人类活动与气候变化对科尔沁沙质草地植被的影响
1
2008
... 人类活动的加剧显著影响了全球变化,其中降雨格局变化和氮沉降已成为生态学领域的研究热点[1-3].草地生态系统具有一定的敏感性和脆弱性,未来极端降水和极端干旱事件在全球范围内发生的频率将会增加[4-5],这可能会严重地影响草地生态系统的结构与功能[6-7].由于化石燃料的燃烧、含氮化肥的使用等,大气中氮化物含量升高,造成全球大气氮沉降总量增加到原来的3~5倍[8].中国等东南亚地区已成为继北美、欧洲的另一氮沉降集中区[9],并且随着中国工业化的发展,氮沉降量可能还会持续升高[10].氮沉降以湿沉降为主,即氮沉降通常伴随降雨的发生[11],因此两者交互作用可能会对半干旱沙质草地植被特征和叶性状产生复杂的影响.已有研究发现,水氮耦合对植物生长存在一种相互补偿机制[12]. ...
Robust spatially aggregated projections of climate extremes
2013
氮沉降对草地凋落物分解的影响研究进展
1
2017
... 人类活动的加剧显著影响了全球变化,其中降雨格局变化和氮沉降已成为生态学领域的研究热点[1-3].草地生态系统具有一定的敏感性和脆弱性,未来极端降水和极端干旱事件在全球范围内发生的频率将会增加[4-5],这可能会严重地影响草地生态系统的结构与功能[6-7].由于化石燃料的燃烧、含氮化肥的使用等,大气中氮化物含量升高,造成全球大气氮沉降总量增加到原来的3~5倍[8].中国等东南亚地区已成为继北美、欧洲的另一氮沉降集中区[9],并且随着中国工业化的发展,氮沉降量可能还会持续升高[10].氮沉降以湿沉降为主,即氮沉降通常伴随降雨的发生[11],因此两者交互作用可能会对半干旱沙质草地植被特征和叶性状产生复杂的影响.已有研究发现,水氮耦合对植物生长存在一种相互补偿机制[12]. ...
极端降水和极端干旱事件对草原生态系统的影响
1
2014
... 人类活动的加剧显著影响了全球变化,其中降雨格局变化和氮沉降已成为生态学领域的研究热点[1-3].草地生态系统具有一定的敏感性和脆弱性,未来极端降水和极端干旱事件在全球范围内发生的频率将会增加[4-5],这可能会严重地影响草地生态系统的结构与功能[6-7].由于化石燃料的燃烧、含氮化肥的使用等,大气中氮化物含量升高,造成全球大气氮沉降总量增加到原来的3~5倍[8].中国等东南亚地区已成为继北美、欧洲的另一氮沉降集中区[9],并且随着中国工业化的发展,氮沉降量可能还会持续升高[10].氮沉降以湿沉降为主,即氮沉降通常伴随降雨的发生[11],因此两者交互作用可能会对半干旱沙质草地植被特征和叶性状产生复杂的影响.已有研究发现,水氮耦合对植物生长存在一种相互补偿机制[12]. ...
Climate change 2007:impacts, adaptation and vulnerability. contribution of working group ii to the fourth assessment report of the intergovernmental panel on climate change
1
2007
... 人类活动的加剧显著影响了全球变化,其中降雨格局变化和氮沉降已成为生态学领域的研究热点[1-3].草地生态系统具有一定的敏感性和脆弱性,未来极端降水和极端干旱事件在全球范围内发生的频率将会增加[4-5],这可能会严重地影响草地生态系统的结构与功能[6-7].由于化石燃料的燃烧、含氮化肥的使用等,大气中氮化物含量升高,造成全球大气氮沉降总量增加到原来的3~5倍[8].中国等东南亚地区已成为继北美、欧洲的另一氮沉降集中区[9],并且随着中国工业化的发展,氮沉降量可能还会持续升高[10].氮沉降以湿沉降为主,即氮沉降通常伴随降雨的发生[11],因此两者交互作用可能会对半干旱沙质草地植被特征和叶性状产生复杂的影响.已有研究发现,水氮耦合对植物生长存在一种相互补偿机制[12]. ...
科尔沁固定沙地植被特征对降雨变化的响应
3
2014
... 人类活动的加剧显著影响了全球变化,其中降雨格局变化和氮沉降已成为生态学领域的研究热点[1-3].草地生态系统具有一定的敏感性和脆弱性,未来极端降水和极端干旱事件在全球范围内发生的频率将会增加[4-5],这可能会严重地影响草地生态系统的结构与功能[6-7].由于化石燃料的燃烧、含氮化肥的使用等,大气中氮化物含量升高,造成全球大气氮沉降总量增加到原来的3~5倍[8].中国等东南亚地区已成为继北美、欧洲的另一氮沉降集中区[9],并且随着中国工业化的发展,氮沉降量可能还会持续升高[10].氮沉降以湿沉降为主,即氮沉降通常伴随降雨的发生[11],因此两者交互作用可能会对半干旱沙质草地植被特征和叶性状产生复杂的影响.已有研究发现,水氮耦合对植物生长存在一种相互补偿机制[12]. ...
... 降雨是影响植物生长的主要因子,尤其限制干旱半干旱地区植物的生长、发育与繁殖[13].降雨格局的变化会影响植物生长,改变物种间关系,进而影响群落组成和结构,最终影响生态系统的结构与功能[14-15],同时生态系统结构与功能能够在群落尺度对降雨变化做出快速响应[16].在生态学研究中,群落生物量作为重要的数量特征,可衡量植被的动态和生态系统功能的变化[17],是表征草地生态系统恢复进程最直接的指标[18].降雨量对植物群落生产力的影响结果不一致.有学者发现极端降雨和极端干旱对生态系统无显著影响[19].而另一些学者则认为增加降雨会显著提高植物群落生产力[20-22].也有研究指出,降雨量对群落生产力的影响存在一定的范围,当降雨量超出植物生长所适宜的水分范围时后,群落生产力反而会下降[6,15].植物功能性状作为衡量植被与环境之间关系的重要依据[23-24],通过改变微环境条件直接影响植物个体的响应从而间接影响群落的动态变化[25].岳喜元等[26]发现,随着降水减少,典型草原植物群落和优势种羊草的叶面积与叶干物质含量降低,而比叶面积与叶片氮含量增加.田青等[27]发现,降雨量与比叶面积和叶干物质含量间不存在简单的线性关系. ...
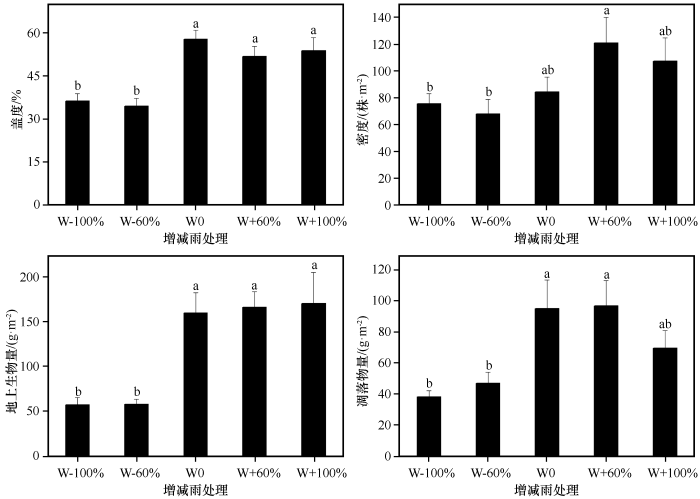
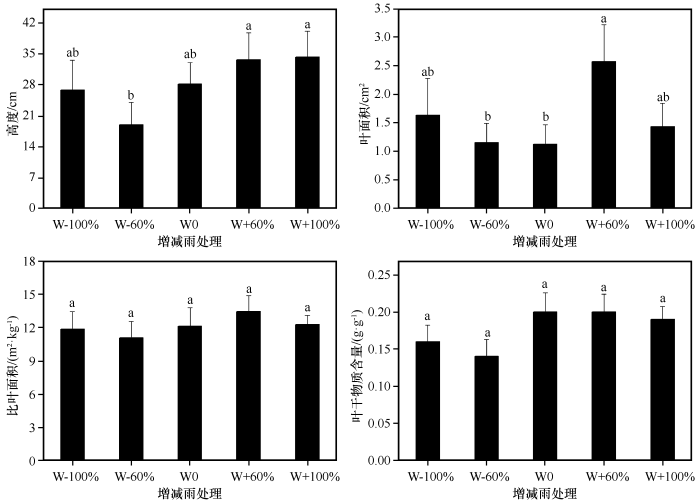
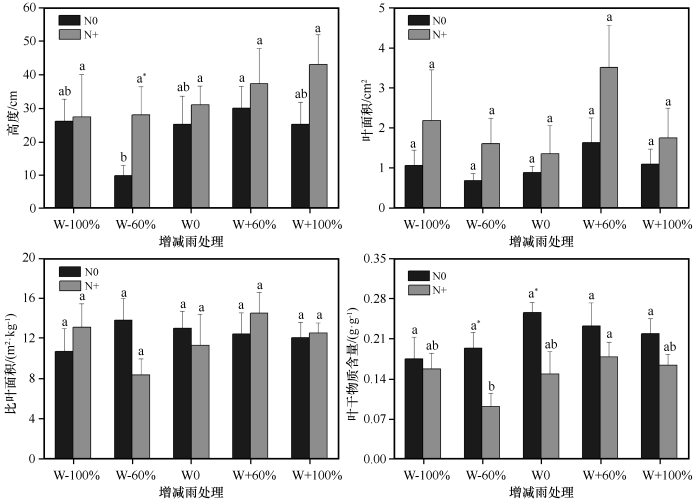
... 降雨是沙地植被形成和发展的主要制约因素[44],降雨量的变化会影响土壤含水量发生变化,通常土壤水分增加会促进植物生长[45].本研究中,减雨(W-60%与W-100%)显著降低植物群落盖度和地上生物量,说明极端干旱不利于沙地植物群落生产力的提高,而增雨(W+60%与W+100%)对其无显著影响,这可能是由于样地中大部分植物为沙生植物,适当增雨会增加群落生产力,但过度增雨可能不利于甚至会抑制沙生植物的生长,使其出现不适应的现象,从而导致增雨对植物群落盖度、地上生物量等影响不明显.增雨60%后群落密度显著高于减雨(W-60%与W-100%),说明增雨60%对群落密度影响更大.张腊梅等[6]发现,相比于增雨60%,增雨30%更有利于植物生长,这与本研究结果有一定的差异,可能是由于试验期间降雨量不同引起的.凋落物是植物将养分返还土壤的一种形式[46],由于一年生植物通常会在较短的时间内快速完成资源的积累和生活史周期[47],因此随着降雨的减少植物会加快衰老的速度,进而产生的凋落物会增多,而本研究发现减雨(W-60%与W-100%)显著降低了群落凋落物量,这可能是由于沙地中原有的现存生物量就少,在降雨减少的情况下,新生的现存生物量更少,进而产生的凋落物也就更少[14].本研究中植物群落高度在减雨60%处理下最低,之后随着降雨的增加而逐渐增加,但相比于增雨60%,增雨100%处理下群落高度几乎没有增加.同样地,叶面积在增雨60%处理下到达最大值,尤其显著高于对照和减雨60%处理,说明相比于增雨100%,增雨60%可以更好地提高植物对光资源的获取能力,表明降雨超过一定范围后植物群落对降雨变化的响应会减弱.比叶面积和叶干物质含量反映植物对资源的利用状况,两者都易受水分的束缚,但叶干物质含量很少受其他性状的影响,如叶片厚度等[48].本研究发现降雨变化对植物群落比叶面积和叶干物质含量均无显著影响,可能是由于降雨对植物的影响具有一定的滞后性[20,49],且试验周期较短. ...
降雨格局的变化对草原生态系统的影响研究进展
1
2010
... 人类活动的加剧显著影响了全球变化,其中降雨格局变化和氮沉降已成为生态学领域的研究热点[1-3].草地生态系统具有一定的敏感性和脆弱性,未来极端降水和极端干旱事件在全球范围内发生的频率将会增加[4-5],这可能会严重地影响草地生态系统的结构与功能[6-7].由于化石燃料的燃烧、含氮化肥的使用等,大气中氮化物含量升高,造成全球大气氮沉降总量增加到原来的3~5倍[8].中国等东南亚地区已成为继北美、欧洲的另一氮沉降集中区[9],并且随着中国工业化的发展,氮沉降量可能还会持续升高[10].氮沉降以湿沉降为主,即氮沉降通常伴随降雨的发生[11],因此两者交互作用可能会对半干旱沙质草地植被特征和叶性状产生复杂的影响.已有研究发现,水氮耦合对植物生长存在一种相互补偿机制[12]. ...
大气氮沉降研究进展
1
2012
... 人类活动的加剧显著影响了全球变化,其中降雨格局变化和氮沉降已成为生态学领域的研究热点[1-3].草地生态系统具有一定的敏感性和脆弱性,未来极端降水和极端干旱事件在全球范围内发生的频率将会增加[4-5],这可能会严重地影响草地生态系统的结构与功能[6-7].由于化石燃料的燃烧、含氮化肥的使用等,大气中氮化物含量升高,造成全球大气氮沉降总量增加到原来的3~5倍[8].中国等东南亚地区已成为继北美、欧洲的另一氮沉降集中区[9],并且随着中国工业化的发展,氮沉降量可能还会持续升高[10].氮沉降以湿沉降为主,即氮沉降通常伴随降雨的发生[11],因此两者交互作用可能会对半干旱沙质草地植被特征和叶性状产生复杂的影响.已有研究发现,水氮耦合对植物生长存在一种相互补偿机制[12]. ...
Contemporary and pre-industrial global reactive nitrogen budgets
1
1999
... 人类活动的加剧显著影响了全球变化,其中降雨格局变化和氮沉降已成为生态学领域的研究热点[1-3].草地生态系统具有一定的敏感性和脆弱性,未来极端降水和极端干旱事件在全球范围内发生的频率将会增加[4-5],这可能会严重地影响草地生态系统的结构与功能[6-7].由于化石燃料的燃烧、含氮化肥的使用等,大气中氮化物含量升高,造成全球大气氮沉降总量增加到原来的3~5倍[8].中国等东南亚地区已成为继北美、欧洲的另一氮沉降集中区[9],并且随着中国工业化的发展,氮沉降量可能还会持续升高[10].氮沉降以湿沉降为主,即氮沉降通常伴随降雨的发生[11],因此两者交互作用可能会对半干旱沙质草地植被特征和叶性状产生复杂的影响.已有研究发现,水氮耦合对植物生长存在一种相互补偿机制[12]. ...
Global nitrogen deposition and carbon sinks
1
2008
... 人类活动的加剧显著影响了全球变化,其中降雨格局变化和氮沉降已成为生态学领域的研究热点[1-3].草地生态系统具有一定的敏感性和脆弱性,未来极端降水和极端干旱事件在全球范围内发生的频率将会增加[4-5],这可能会严重地影响草地生态系统的结构与功能[6-7].由于化石燃料的燃烧、含氮化肥的使用等,大气中氮化物含量升高,造成全球大气氮沉降总量增加到原来的3~5倍[8].中国等东南亚地区已成为继北美、欧洲的另一氮沉降集中区[9],并且随着中国工业化的发展,氮沉降量可能还会持续升高[10].氮沉降以湿沉降为主,即氮沉降通常伴随降雨的发生[11],因此两者交互作用可能会对半干旱沙质草地植被特征和叶性状产生复杂的影响.已有研究发现,水氮耦合对植物生长存在一种相互补偿机制[12]. ...
降雨量与氮沉降及其耦合对羊草主要叶经济性状的影响
1
2016
... 人类活动的加剧显著影响了全球变化,其中降雨格局变化和氮沉降已成为生态学领域的研究热点[1-3].草地生态系统具有一定的敏感性和脆弱性,未来极端降水和极端干旱事件在全球范围内发生的频率将会增加[4-5],这可能会严重地影响草地生态系统的结构与功能[6-7].由于化石燃料的燃烧、含氮化肥的使用等,大气中氮化物含量升高,造成全球大气氮沉降总量增加到原来的3~5倍[8].中国等东南亚地区已成为继北美、欧洲的另一氮沉降集中区[9],并且随着中国工业化的发展,氮沉降量可能还会持续升高[10].氮沉降以湿沉降为主,即氮沉降通常伴随降雨的发生[11],因此两者交互作用可能会对半干旱沙质草地植被特征和叶性状产生复杂的影响.已有研究发现,水氮耦合对植物生长存在一种相互补偿机制[12]. ...
模拟氮沉降和干旱对准噶尔盆地两种一年生荒漠植物生长和光合生理的影响
1
2010
... 人类活动的加剧显著影响了全球变化,其中降雨格局变化和氮沉降已成为生态学领域的研究热点[1-3].草地生态系统具有一定的敏感性和脆弱性,未来极端降水和极端干旱事件在全球范围内发生的频率将会增加[4-5],这可能会严重地影响草地生态系统的结构与功能[6-7].由于化石燃料的燃烧、含氮化肥的使用等,大气中氮化物含量升高,造成全球大气氮沉降总量增加到原来的3~5倍[8].中国等东南亚地区已成为继北美、欧洲的另一氮沉降集中区[9],并且随着中国工业化的发展,氮沉降量可能还会持续升高[10].氮沉降以湿沉降为主,即氮沉降通常伴随降雨的发生[11],因此两者交互作用可能会对半干旱沙质草地植被特征和叶性状产生复杂的影响.已有研究发现,水氮耦合对植物生长存在一种相互补偿机制[12]. ...
天然白刺灌丛对人工模拟增雨的响应
1
2012
... 降雨是影响植物生长的主要因子,尤其限制干旱半干旱地区植物的生长、发育与繁殖[13].降雨格局的变化会影响植物生长,改变物种间关系,进而影响群落组成和结构,最终影响生态系统的结构与功能[14-15],同时生态系统结构与功能能够在群落尺度对降雨变化做出快速响应[16].在生态学研究中,群落生物量作为重要的数量特征,可衡量植被的动态和生态系统功能的变化[17],是表征草地生态系统恢复进程最直接的指标[18].降雨量对植物群落生产力的影响结果不一致.有学者发现极端降雨和极端干旱对生态系统无显著影响[19].而另一些学者则认为增加降雨会显著提高植物群落生产力[20-22].也有研究指出,降雨量对群落生产力的影响存在一定的范围,当降雨量超出植物生长所适宜的水分范围时后,群落生产力反而会下降[6,15].植物功能性状作为衡量植被与环境之间关系的重要依据[23-24],通过改变微环境条件直接影响植物个体的响应从而间接影响群落的动态变化[25].岳喜元等[26]发现,随着降水减少,典型草原植物群落和优势种羊草的叶面积与叶干物质含量降低,而比叶面积与叶片氮含量增加.田青等[27]发现,降雨量与比叶面积和叶干物质含量间不存在简单的线性关系. ...
荒漠草原沙生针茅(Stipa glareosa)群落物种多样性和地上生物量对降雨量的响应
2
2019
... 降雨是影响植物生长的主要因子,尤其限制干旱半干旱地区植物的生长、发育与繁殖[13].降雨格局的变化会影响植物生长,改变物种间关系,进而影响群落组成和结构,最终影响生态系统的结构与功能[14-15],同时生态系统结构与功能能够在群落尺度对降雨变化做出快速响应[16].在生态学研究中,群落生物量作为重要的数量特征,可衡量植被的动态和生态系统功能的变化[17],是表征草地生态系统恢复进程最直接的指标[18].降雨量对植物群落生产力的影响结果不一致.有学者发现极端降雨和极端干旱对生态系统无显著影响[19].而另一些学者则认为增加降雨会显著提高植物群落生产力[20-22].也有研究指出,降雨量对群落生产力的影响存在一定的范围,当降雨量超出植物生长所适宜的水分范围时后,群落生产力反而会下降[6,15].植物功能性状作为衡量植被与环境之间关系的重要依据[23-24],通过改变微环境条件直接影响植物个体的响应从而间接影响群落的动态变化[25].岳喜元等[26]发现,随着降水减少,典型草原植物群落和优势种羊草的叶面积与叶干物质含量降低,而比叶面积与叶片氮含量增加.田青等[27]发现,降雨量与比叶面积和叶干物质含量间不存在简单的线性关系. ...
... 降雨是沙地植被形成和发展的主要制约因素[44],降雨量的变化会影响土壤含水量发生变化,通常土壤水分增加会促进植物生长[45].本研究中,减雨(W-60%与W-100%)显著降低植物群落盖度和地上生物量,说明极端干旱不利于沙地植物群落生产力的提高,而增雨(W+60%与W+100%)对其无显著影响,这可能是由于样地中大部分植物为沙生植物,适当增雨会增加群落生产力,但过度增雨可能不利于甚至会抑制沙生植物的生长,使其出现不适应的现象,从而导致增雨对植物群落盖度、地上生物量等影响不明显.增雨60%后群落密度显著高于减雨(W-60%与W-100%),说明增雨60%对群落密度影响更大.张腊梅等[6]发现,相比于增雨60%,增雨30%更有利于植物生长,这与本研究结果有一定的差异,可能是由于试验期间降雨量不同引起的.凋落物是植物将养分返还土壤的一种形式[46],由于一年生植物通常会在较短的时间内快速完成资源的积累和生活史周期[47],因此随着降雨的减少植物会加快衰老的速度,进而产生的凋落物会增多,而本研究发现减雨(W-60%与W-100%)显著降低了群落凋落物量,这可能是由于沙地中原有的现存生物量就少,在降雨减少的情况下,新生的现存生物量更少,进而产生的凋落物也就更少[14].本研究中植物群落高度在减雨60%处理下最低,之后随着降雨的增加而逐渐增加,但相比于增雨60%,增雨100%处理下群落高度几乎没有增加.同样地,叶面积在增雨60%处理下到达最大值,尤其显著高于对照和减雨60%处理,说明相比于增雨100%,增雨60%可以更好地提高植物对光资源的获取能力,表明降雨超过一定范围后植物群落对降雨变化的响应会减弱.比叶面积和叶干物质含量反映植物对资源的利用状况,两者都易受水分的束缚,但叶干物质含量很少受其他性状的影响,如叶片厚度等[48].本研究发现降雨变化对植物群落比叶面积和叶干物质含量均无显著影响,可能是由于降雨对植物的影响具有一定的滞后性[20,49],且试验周期较短. ...
模拟短期降雨量变化对准噶尔荒漠植物群落的影响
3
2019
... 降雨是影响植物生长的主要因子,尤其限制干旱半干旱地区植物的生长、发育与繁殖[13].降雨格局的变化会影响植物生长,改变物种间关系,进而影响群落组成和结构,最终影响生态系统的结构与功能[14-15],同时生态系统结构与功能能够在群落尺度对降雨变化做出快速响应[16].在生态学研究中,群落生物量作为重要的数量特征,可衡量植被的动态和生态系统功能的变化[17],是表征草地生态系统恢复进程最直接的指标[18].降雨量对植物群落生产力的影响结果不一致.有学者发现极端降雨和极端干旱对生态系统无显著影响[19].而另一些学者则认为增加降雨会显著提高植物群落生产力[20-22].也有研究指出,降雨量对群落生产力的影响存在一定的范围,当降雨量超出植物生长所适宜的水分范围时后,群落生产力反而会下降[6,15].植物功能性状作为衡量植被与环境之间关系的重要依据[23-24],通过改变微环境条件直接影响植物个体的响应从而间接影响群落的动态变化[25].岳喜元等[26]发现,随着降水减少,典型草原植物群落和优势种羊草的叶面积与叶干物质含量降低,而比叶面积与叶片氮含量增加.田青等[27]发现,降雨量与比叶面积和叶干物质含量间不存在简单的线性关系. ...
... ,15].植物功能性状作为衡量植被与环境之间关系的重要依据[23-24],通过改变微环境条件直接影响植物个体的响应从而间接影响群落的动态变化[25].岳喜元等[26]发现,随着降水减少,典型草原植物群落和优势种羊草的叶面积与叶干物质含量降低,而比叶面积与叶片氮含量增加.田青等[27]发现,降雨量与比叶面积和叶干物质含量间不存在简单的线性关系. ...
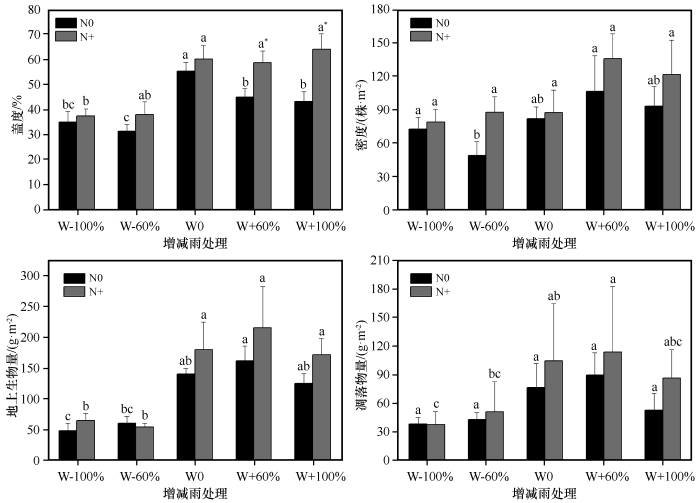
... 水氮之间对植物生长存在相互补偿机制,主要表现为氮添加能缓解干旱对植物造成的影响,但同时干旱可以减弱甚至抑制氮对植物生长的促进作用[56].增雨(W+60%和W+100%)处理下,氮添加使得群落盖度显著增加,群落密度、地上生物量和凋落物量在增雨60%和氮添加交互处理下达到最大值,而增雨100%与氮添加耦合反而使其降低,说明适当增雨更有利于氮素肥力的激发,降雨过多反而会缩短氮素滞留时间,造成氮素流失[15].同一干旱处理(W-60%与W-100%)下,氮添加增加了群落高度和叶面积,尤其减雨60%处理下氮添加显著提高了群落高度,说明氮添加可以在一定程度上缓解干旱对植物造成的影响.同样地,同一增雨处理(W+60%和W+100%)下氮添加也使得高度和叶面积有所增加,这可能是由于增雨更好地激发了氮肥效力,提高了土壤中氮素的可利用性,进而促进植物对氮素的吸收和利用效率.此外,与增雨100%相比,叶面积在增雨60%和氮添加交互处理下最大,说明并不是降雨越多越有利于植物生长,超过一定范围后反而会抑制植物生长.就叶干物质含量而言,在相同降雨处理下,氮添加降低了植物群落叶干物质含量,尤其在W-60%和W0处理下叶干物质含量显著降低,而比叶面积在水氮交互处理下虽有波动变化,但无明显变化规律,这可能是由于不同物种在面对气候变化时所采取的策略不同[52]. ...
长期降雨波动对半干旱灌木群落生物量和土壤水分动态的效应
1
2010
... 降雨是影响植物生长的主要因子,尤其限制干旱半干旱地区植物的生长、发育与繁殖[13].降雨格局的变化会影响植物生长,改变物种间关系,进而影响群落组成和结构,最终影响生态系统的结构与功能[14-15],同时生态系统结构与功能能够在群落尺度对降雨变化做出快速响应[16].在生态学研究中,群落生物量作为重要的数量特征,可衡量植被的动态和生态系统功能的变化[17],是表征草地生态系统恢复进程最直接的指标[18].降雨量对植物群落生产力的影响结果不一致.有学者发现极端降雨和极端干旱对生态系统无显著影响[19].而另一些学者则认为增加降雨会显著提高植物群落生产力[20-22].也有研究指出,降雨量对群落生产力的影响存在一定的范围,当降雨量超出植物生长所适宜的水分范围时后,群落生产力反而会下降[6,15].植物功能性状作为衡量植被与环境之间关系的重要依据[23-24],通过改变微环境条件直接影响植物个体的响应从而间接影响群落的动态变化[25].岳喜元等[26]发现,随着降水减少,典型草原植物群落和优势种羊草的叶面积与叶干物质含量降低,而比叶面积与叶片氮含量增加.田青等[27]发现,降雨量与比叶面积和叶干物质含量间不存在简单的线性关系. ...
半干旱沙地生境变化对植物地上生物量及其碳、氮储量的影响
1
2014
... 降雨是影响植物生长的主要因子,尤其限制干旱半干旱地区植物的生长、发育与繁殖[13].降雨格局的变化会影响植物生长,改变物种间关系,进而影响群落组成和结构,最终影响生态系统的结构与功能[14-15],同时生态系统结构与功能能够在群落尺度对降雨变化做出快速响应[16].在生态学研究中,群落生物量作为重要的数量特征,可衡量植被的动态和生态系统功能的变化[17],是表征草地生态系统恢复进程最直接的指标[18].降雨量对植物群落生产力的影响结果不一致.有学者发现极端降雨和极端干旱对生态系统无显著影响[19].而另一些学者则认为增加降雨会显著提高植物群落生产力[20-22].也有研究指出,降雨量对群落生产力的影响存在一定的范围,当降雨量超出植物生长所适宜的水分范围时后,群落生产力反而会下降[6,15].植物功能性状作为衡量植被与环境之间关系的重要依据[23-24],通过改变微环境条件直接影响植物个体的响应从而间接影响群落的动态变化[25].岳喜元等[26]发现,随着降水减少,典型草原植物群落和优势种羊草的叶面积与叶干物质含量降低,而比叶面积与叶片氮含量增加.田青等[27]发现,降雨量与比叶面积和叶干物质含量间不存在简单的线性关系. ...
短期降雨改变对荒漠草原植物群落特征的影响
1
2018
... 降雨是影响植物生长的主要因子,尤其限制干旱半干旱地区植物的生长、发育与繁殖[13].降雨格局的变化会影响植物生长,改变物种间关系,进而影响群落组成和结构,最终影响生态系统的结构与功能[14-15],同时生态系统结构与功能能够在群落尺度对降雨变化做出快速响应[16].在生态学研究中,群落生物量作为重要的数量特征,可衡量植被的动态和生态系统功能的变化[17],是表征草地生态系统恢复进程最直接的指标[18].降雨量对植物群落生产力的影响结果不一致.有学者发现极端降雨和极端干旱对生态系统无显著影响[19].而另一些学者则认为增加降雨会显著提高植物群落生产力[20-22].也有研究指出,降雨量对群落生产力的影响存在一定的范围,当降雨量超出植物生长所适宜的水分范围时后,群落生产力反而会下降[6,15].植物功能性状作为衡量植被与环境之间关系的重要依据[23-24],通过改变微环境条件直接影响植物个体的响应从而间接影响群落的动态变化[25].岳喜元等[26]发现,随着降水减少,典型草原植物群落和优势种羊草的叶面积与叶干物质含量降低,而比叶面积与叶片氮含量增加.田青等[27]发现,降雨量与比叶面积和叶干物质含量间不存在简单的线性关系. ...
R:S ratios of temperate steppe and the environmental controls in Inner Mongolia
1
2006
... 降雨是影响植物生长的主要因子,尤其限制干旱半干旱地区植物的生长、发育与繁殖[13].降雨格局的变化会影响植物生长,改变物种间关系,进而影响群落组成和结构,最终影响生态系统的结构与功能[14-15],同时生态系统结构与功能能够在群落尺度对降雨变化做出快速响应[16].在生态学研究中,群落生物量作为重要的数量特征,可衡量植被的动态和生态系统功能的变化[17],是表征草地生态系统恢复进程最直接的指标[18].降雨量对植物群落生产力的影响结果不一致.有学者发现极端降雨和极端干旱对生态系统无显著影响[19].而另一些学者则认为增加降雨会显著提高植物群落生产力[20-22].也有研究指出,降雨量对群落生产力的影响存在一定的范围,当降雨量超出植物生长所适宜的水分范围时后,群落生产力反而会下降[6,15].植物功能性状作为衡量植被与环境之间关系的重要依据[23-24],通过改变微环境条件直接影响植物个体的响应从而间接影响群落的动态变化[25].岳喜元等[26]发现,随着降水减少,典型草原植物群落和优势种羊草的叶面积与叶干物质含量降低,而比叶面积与叶片氮含量增加.田青等[27]发现,降雨量与比叶面积和叶干物质含量间不存在简单的线性关系. ...
气候变化对中国草原生态系统影响研究综述
2
2014
... 降雨是影响植物生长的主要因子,尤其限制干旱半干旱地区植物的生长、发育与繁殖[13].降雨格局的变化会影响植物生长,改变物种间关系,进而影响群落组成和结构,最终影响生态系统的结构与功能[14-15],同时生态系统结构与功能能够在群落尺度对降雨变化做出快速响应[16].在生态学研究中,群落生物量作为重要的数量特征,可衡量植被的动态和生态系统功能的变化[17],是表征草地生态系统恢复进程最直接的指标[18].降雨量对植物群落生产力的影响结果不一致.有学者发现极端降雨和极端干旱对生态系统无显著影响[19].而另一些学者则认为增加降雨会显著提高植物群落生产力[20-22].也有研究指出,降雨量对群落生产力的影响存在一定的范围,当降雨量超出植物生长所适宜的水分范围时后,群落生产力反而会下降[6,15].植物功能性状作为衡量植被与环境之间关系的重要依据[23-24],通过改变微环境条件直接影响植物个体的响应从而间接影响群落的动态变化[25].岳喜元等[26]发现,随着降水减少,典型草原植物群落和优势种羊草的叶面积与叶干物质含量降低,而比叶面积与叶片氮含量增加.田青等[27]发现,降雨量与比叶面积和叶干物质含量间不存在简单的线性关系. ...
... 降雨是沙地植被形成和发展的主要制约因素[44],降雨量的变化会影响土壤含水量发生变化,通常土壤水分增加会促进植物生长[45].本研究中,减雨(W-60%与W-100%)显著降低植物群落盖度和地上生物量,说明极端干旱不利于沙地植物群落生产力的提高,而增雨(W+60%与W+100%)对其无显著影响,这可能是由于样地中大部分植物为沙生植物,适当增雨会增加群落生产力,但过度增雨可能不利于甚至会抑制沙生植物的生长,使其出现不适应的现象,从而导致增雨对植物群落盖度、地上生物量等影响不明显.增雨60%后群落密度显著高于减雨(W-60%与W-100%),说明增雨60%对群落密度影响更大.张腊梅等[6]发现,相比于增雨60%,增雨30%更有利于植物生长,这与本研究结果有一定的差异,可能是由于试验期间降雨量不同引起的.凋落物是植物将养分返还土壤的一种形式[46],由于一年生植物通常会在较短的时间内快速完成资源的积累和生活史周期[47],因此随着降雨的减少植物会加快衰老的速度,进而产生的凋落物会增多,而本研究发现减雨(W-60%与W-100%)显著降低了群落凋落物量,这可能是由于沙地中原有的现存生物量就少,在降雨减少的情况下,新生的现存生物量更少,进而产生的凋落物也就更少[14].本研究中植物群落高度在减雨60%处理下最低,之后随着降雨的增加而逐渐增加,但相比于增雨60%,增雨100%处理下群落高度几乎没有增加.同样地,叶面积在增雨60%处理下到达最大值,尤其显著高于对照和减雨60%处理,说明相比于增雨100%,增雨60%可以更好地提高植物对光资源的获取能力,表明降雨超过一定范围后植物群落对降雨变化的响应会减弱.比叶面积和叶干物质含量反映植物对资源的利用状况,两者都易受水分的束缚,但叶干物质含量很少受其他性状的影响,如叶片厚度等[48].本研究发现降雨变化对植物群落比叶面积和叶干物质含量均无显著影响,可能是由于降雨对植物的影响具有一定的滞后性[20,49],且试验周期较短. ...
Precipitation patterns alter growth of temperate vegetation
2005
Primary production and rain use efficiency across a precipitation gradient on the mongolia plateau
1
2008
... 降雨是影响植物生长的主要因子,尤其限制干旱半干旱地区植物的生长、发育与繁殖[13].降雨格局的变化会影响植物生长,改变物种间关系,进而影响群落组成和结构,最终影响生态系统的结构与功能[14-15],同时生态系统结构与功能能够在群落尺度对降雨变化做出快速响应[16].在生态学研究中,群落生物量作为重要的数量特征,可衡量植被的动态和生态系统功能的变化[17],是表征草地生态系统恢复进程最直接的指标[18].降雨量对植物群落生产力的影响结果不一致.有学者发现极端降雨和极端干旱对生态系统无显著影响[19].而另一些学者则认为增加降雨会显著提高植物群落生产力[20-22].也有研究指出,降雨量对群落生产力的影响存在一定的范围,当降雨量超出植物生长所适宜的水分范围时后,群落生产力反而会下降[6,15].植物功能性状作为衡量植被与环境之间关系的重要依据[23-24],通过改变微环境条件直接影响植物个体的响应从而间接影响群落的动态变化[25].岳喜元等[26]发现,随着降水减少,典型草原植物群落和优势种羊草的叶面积与叶干物质含量降低,而比叶面积与叶片氮含量增加.田青等[27]发现,降雨量与比叶面积和叶干物质含量间不存在简单的线性关系. ...
Plant functional traits and environmental filters at a regional scale
1
1998
... 降雨是影响植物生长的主要因子,尤其限制干旱半干旱地区植物的生长、发育与繁殖[13].降雨格局的变化会影响植物生长,改变物种间关系,进而影响群落组成和结构,最终影响生态系统的结构与功能[14-15],同时生态系统结构与功能能够在群落尺度对降雨变化做出快速响应[16].在生态学研究中,群落生物量作为重要的数量特征,可衡量植被的动态和生态系统功能的变化[17],是表征草地生态系统恢复进程最直接的指标[18].降雨量对植物群落生产力的影响结果不一致.有学者发现极端降雨和极端干旱对生态系统无显著影响[19].而另一些学者则认为增加降雨会显著提高植物群落生产力[20-22].也有研究指出,降雨量对群落生产力的影响存在一定的范围,当降雨量超出植物生长所适宜的水分范围时后,群落生产力反而会下降[6,15].植物功能性状作为衡量植被与环境之间关系的重要依据[23-24],通过改变微环境条件直接影响植物个体的响应从而间接影响群落的动态变化[25].岳喜元等[26]发现,随着降水减少,典型草原植物群落和优势种羊草的叶面积与叶干物质含量降低,而比叶面积与叶片氮含量增加.田青等[27]发现,降雨量与比叶面积和叶干物质含量间不存在简单的线性关系. ...
Using plant functional traits to explain community composition across a strong environmental filter in Australian alpine snowpatches
1
2011
... 降雨是影响植物生长的主要因子,尤其限制干旱半干旱地区植物的生长、发育与繁殖[13].降雨格局的变化会影响植物生长,改变物种间关系,进而影响群落组成和结构,最终影响生态系统的结构与功能[14-15],同时生态系统结构与功能能够在群落尺度对降雨变化做出快速响应[16].在生态学研究中,群落生物量作为重要的数量特征,可衡量植被的动态和生态系统功能的变化[17],是表征草地生态系统恢复进程最直接的指标[18].降雨量对植物群落生产力的影响结果不一致.有学者发现极端降雨和极端干旱对生态系统无显著影响[19].而另一些学者则认为增加降雨会显著提高植物群落生产力[20-22].也有研究指出,降雨量对群落生产力的影响存在一定的范围,当降雨量超出植物生长所适宜的水分范围时后,群落生产力反而会下降[6,15].植物功能性状作为衡量植被与环境之间关系的重要依据[23-24],通过改变微环境条件直接影响植物个体的响应从而间接影响群落的动态变化[25].岳喜元等[26]发现,随着降水减少,典型草原植物群落和优势种羊草的叶面积与叶干物质含量降低,而比叶面积与叶片氮含量增加.田青等[27]发现,降雨量与比叶面积和叶干物质含量间不存在简单的线性关系. ...
呼伦贝尔草原3种植物的功能性状对氮磷添加的响应
1
2019
... 降雨是影响植物生长的主要因子,尤其限制干旱半干旱地区植物的生长、发育与繁殖[13].降雨格局的变化会影响植物生长,改变物种间关系,进而影响群落组成和结构,最终影响生态系统的结构与功能[14-15],同时生态系统结构与功能能够在群落尺度对降雨变化做出快速响应[16].在生态学研究中,群落生物量作为重要的数量特征,可衡量植被的动态和生态系统功能的变化[17],是表征草地生态系统恢复进程最直接的指标[18].降雨量对植物群落生产力的影响结果不一致.有学者发现极端降雨和极端干旱对生态系统无显著影响[19].而另一些学者则认为增加降雨会显著提高植物群落生产力[20-22].也有研究指出,降雨量对群落生产力的影响存在一定的范围,当降雨量超出植物生长所适宜的水分范围时后,群落生产力反而会下降[6,15].植物功能性状作为衡量植被与环境之间关系的重要依据[23-24],通过改变微环境条件直接影响植物个体的响应从而间接影响群落的动态变化[25].岳喜元等[26]发现,随着降水减少,典型草原植物群落和优势种羊草的叶面积与叶干物质含量降低,而比叶面积与叶片氮含量增加.田青等[27]发现,降雨量与比叶面积和叶干物质含量间不存在简单的线性关系. ...
降水量和短期极端干旱对典型草原植物群落及优势种羊草(Leymus chinensis)叶性状的影响
1
2018
... 降雨是影响植物生长的主要因子,尤其限制干旱半干旱地区植物的生长、发育与繁殖[13].降雨格局的变化会影响植物生长,改变物种间关系,进而影响群落组成和结构,最终影响生态系统的结构与功能[14-15],同时生态系统结构与功能能够在群落尺度对降雨变化做出快速响应[16].在生态学研究中,群落生物量作为重要的数量特征,可衡量植被的动态和生态系统功能的变化[17],是表征草地生态系统恢复进程最直接的指标[18].降雨量对植物群落生产力的影响结果不一致.有学者发现极端降雨和极端干旱对生态系统无显著影响[19].而另一些学者则认为增加降雨会显著提高植物群落生产力[20-22].也有研究指出,降雨量对群落生产力的影响存在一定的范围,当降雨量超出植物生长所适宜的水分范围时后,群落生产力反而会下降[6,15].植物功能性状作为衡量植被与环境之间关系的重要依据[23-24],通过改变微环境条件直接影响植物个体的响应从而间接影响群落的动态变化[25].岳喜元等[26]发现,随着降水减少,典型草原植物群落和优势种羊草的叶面积与叶干物质含量降低,而比叶面积与叶片氮含量增加.田青等[27]发现,降雨量与比叶面积和叶干物质含量间不存在简单的线性关系. ...
典型草原14种植物叶片性状对模拟降雨变化的响应
1
2008
... 降雨是影响植物生长的主要因子,尤其限制干旱半干旱地区植物的生长、发育与繁殖[13].降雨格局的变化会影响植物生长,改变物种间关系,进而影响群落组成和结构,最终影响生态系统的结构与功能[14-15],同时生态系统结构与功能能够在群落尺度对降雨变化做出快速响应[16].在生态学研究中,群落生物量作为重要的数量特征,可衡量植被的动态和生态系统功能的变化[17],是表征草地生态系统恢复进程最直接的指标[18].降雨量对植物群落生产力的影响结果不一致.有学者发现极端降雨和极端干旱对生态系统无显著影响[19].而另一些学者则认为增加降雨会显著提高植物群落生产力[20-22].也有研究指出,降雨量对群落生产力的影响存在一定的范围,当降雨量超出植物生长所适宜的水分范围时后,群落生产力反而会下降[6,15].植物功能性状作为衡量植被与环境之间关系的重要依据[23-24],通过改变微环境条件直接影响植物个体的响应从而间接影响群落的动态变化[25].岳喜元等[26]发现,随着降水减少,典型草原植物群落和优势种羊草的叶面积与叶干物质含量降低,而比叶面积与叶片氮含量增加.田青等[27]发现,降雨量与比叶面积和叶干物质含量间不存在简单的线性关系. ...
Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China
1
2005
... 氮作为植物生命活动必需的营养元素,对植物生长发育有着至关重要的作用[28].在氮沉降初期,氮素作为营养物质通过增加土壤氮含量从而促进植物的生长,尤其是生长在受氮素限制的生态系统中的植物[29-30].但过度氮沉降导致植物吸收过量的氮素,超过生态系统临界氮容量,打破植物体内固有的养分平衡,从而对植物个体、群落乃至整个生态系统产生影响[29,31].宋彦涛等[32]发现,氮添加提高了羊草地上生物量,比叶面积和叶片厚度无显著变化.杨浩等[33]发现,低氮添加主要影响糙隐子草叶片形态性状,而高氮添加主要影响叶生理性状.李禄军等[34]通过模拟氮沉降发现,氮素增加改变了植物群落物种组成和优势物种,使植物高度、盖度和生物量增加,而物种丰富度和生物多样性减少.Mao等[35]发现,氮添加增加了小叶章(Deyeuxia angustifolia)和狭叶甜茅(Glyceria spiculosa)的高度和地上生物量,但对其比叶面积和叶干物质含量无显著影响. ...
植物响应大气氮沉降研究进展
3
2019
... 氮作为植物生命活动必需的营养元素,对植物生长发育有着至关重要的作用[28].在氮沉降初期,氮素作为营养物质通过增加土壤氮含量从而促进植物的生长,尤其是生长在受氮素限制的生态系统中的植物[29-30].但过度氮沉降导致植物吸收过量的氮素,超过生态系统临界氮容量,打破植物体内固有的养分平衡,从而对植物个体、群落乃至整个生态系统产生影响[29,31].宋彦涛等[32]发现,氮添加提高了羊草地上生物量,比叶面积和叶片厚度无显著变化.杨浩等[33]发现,低氮添加主要影响糙隐子草叶片形态性状,而高氮添加主要影响叶生理性状.李禄军等[34]通过模拟氮沉降发现,氮素增加改变了植物群落物种组成和优势物种,使植物高度、盖度和生物量增加,而物种丰富度和生物多样性减少.Mao等[35]发现,氮添加增加了小叶章(Deyeuxia angustifolia)和狭叶甜茅(Glyceria spiculosa)的高度和地上生物量,但对其比叶面积和叶干物质含量无显著影响. ...
... [29,31].宋彦涛等[32]发现,氮添加提高了羊草地上生物量,比叶面积和叶片厚度无显著变化.杨浩等[33]发现,低氮添加主要影响糙隐子草叶片形态性状,而高氮添加主要影响叶生理性状.李禄军等[34]通过模拟氮沉降发现,氮素增加改变了植物群落物种组成和优势物种,使植物高度、盖度和生物量增加,而物种丰富度和生物多样性减少.Mao等[35]发现,氮添加增加了小叶章(Deyeuxia angustifolia)和狭叶甜茅(Glyceria spiculosa)的高度和地上生物量,但对其比叶面积和叶干物质含量无显著影响. ...
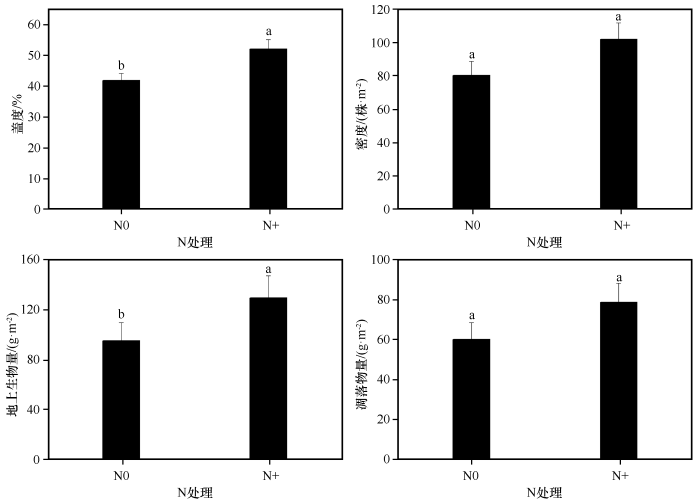
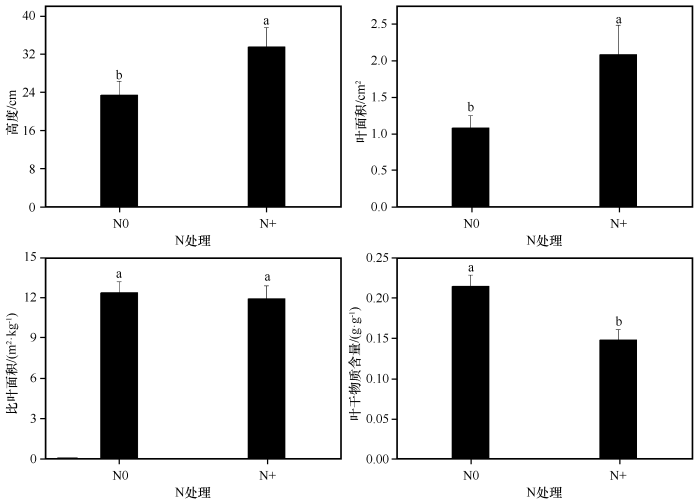
... 在干旱半干旱地区,植物生长不仅受水分限制,较低的氮含量也成为主要限制因子[50].目前,有关氮添加对植物群落生产力影响的研究报道有很多[31-32,51-52],研究结果也不尽相同.本研究中氮添加显著增加了群落盖度和地上生物量,密度也有一定程度的增加,说明短暂的氮添加在一定时期内减缓了该地区氮素对植物生长的限制,促进植物快速生长,增加生物量的累积,与前人研究结果基本一致[34,51].但氮添加也加剧了植物种间或种内竞争,导致一些竞争能力较弱的个体减少甚至死亡,这解释了本研究中群落密度虽有增加但不显著的原因.此外,本研究发现氮添加增加了群落凋落物量,这可能是由于过量的氮添加改变了植物养分再吸收的过程[53],使得器官脱落前植物个体减弱了对养分的再吸收能力[29],同时由于竞争造成个体死亡的增加也为凋落物量的增加做出了一定贡献.植物高度反映植物对光资源竞争能力的强弱[41].研究发现,植物叶片氮含量的高低会影响植物光合作用[54],氮添加可以短暂地消除氮素对沙地生态系统的限制,增加植物可利用氮素水平,使得植物对地下养分的竞争转化为对光的竞争[34],但过量的氮素会打破植物体内的养分平衡,降低植物光合速率[55].本研究中氮添加显著增加了植物群落高度,进一步验证了氮添加可以提高沙地植物群落的光合能力.植物比叶面积越高、叶干物质含量越低,其光捕获能力越强,这些变化有利于光竞争力的提高[48].本研究中氮添加使得群落叶面积显著增加,叶干物质含量显著降低,说明沙地植物群落主要通过叶面积和叶干物质含量的变化来响应氮添加.比叶面积在响应短期氮添加时敏感性较弱,且不同物种对氮添加的响应不同,而群落水平上很容易掩盖这种种间差异,因此比叶面积在群落水平上响应氮添加时没有明显变化. ...
Nitrogen deposition contributes to soil acidification in tropical ecosystems
1
2014
... 氮作为植物生命活动必需的营养元素,对植物生长发育有着至关重要的作用[28].在氮沉降初期,氮素作为营养物质通过增加土壤氮含量从而促进植物的生长,尤其是生长在受氮素限制的生态系统中的植物[29-30].但过度氮沉降导致植物吸收过量的氮素,超过生态系统临界氮容量,打破植物体内固有的养分平衡,从而对植物个体、群落乃至整个生态系统产生影响[29,31].宋彦涛等[32]发现,氮添加提高了羊草地上生物量,比叶面积和叶片厚度无显著变化.杨浩等[33]发现,低氮添加主要影响糙隐子草叶片形态性状,而高氮添加主要影响叶生理性状.李禄军等[34]通过模拟氮沉降发现,氮素增加改变了植物群落物种组成和优势物种,使植物高度、盖度和生物量增加,而物种丰富度和生物多样性减少.Mao等[35]发现,氮添加增加了小叶章(Deyeuxia angustifolia)和狭叶甜茅(Glyceria spiculosa)的高度和地上生物量,但对其比叶面积和叶干物质含量无显著影响. ...
施肥对科尔沁沙质草地群落物种组成和多样性的影响
2
2010
... 氮作为植物生命活动必需的营养元素,对植物生长发育有着至关重要的作用[28].在氮沉降初期,氮素作为营养物质通过增加土壤氮含量从而促进植物的生长,尤其是生长在受氮素限制的生态系统中的植物[29-30].但过度氮沉降导致植物吸收过量的氮素,超过生态系统临界氮容量,打破植物体内固有的养分平衡,从而对植物个体、群落乃至整个生态系统产生影响[29,31].宋彦涛等[32]发现,氮添加提高了羊草地上生物量,比叶面积和叶片厚度无显著变化.杨浩等[33]发现,低氮添加主要影响糙隐子草叶片形态性状,而高氮添加主要影响叶生理性状.李禄军等[34]通过模拟氮沉降发现,氮素增加改变了植物群落物种组成和优势物种,使植物高度、盖度和生物量增加,而物种丰富度和生物多样性减少.Mao等[35]发现,氮添加增加了小叶章(Deyeuxia angustifolia)和狭叶甜茅(Glyceria spiculosa)的高度和地上生物量,但对其比叶面积和叶干物质含量无显著影响. ...
... 在干旱半干旱地区,植物生长不仅受水分限制,较低的氮含量也成为主要限制因子[50].目前,有关氮添加对植物群落生产力影响的研究报道有很多[31-32,51-52],研究结果也不尽相同.本研究中氮添加显著增加了群落盖度和地上生物量,密度也有一定程度的增加,说明短暂的氮添加在一定时期内减缓了该地区氮素对植物生长的限制,促进植物快速生长,增加生物量的累积,与前人研究结果基本一致[34,51].但氮添加也加剧了植物种间或种内竞争,导致一些竞争能力较弱的个体减少甚至死亡,这解释了本研究中群落密度虽有增加但不显著的原因.此外,本研究发现氮添加增加了群落凋落物量,这可能是由于过量的氮添加改变了植物养分再吸收的过程[53],使得器官脱落前植物个体减弱了对养分的再吸收能力[29],同时由于竞争造成个体死亡的增加也为凋落物量的增加做出了一定贡献.植物高度反映植物对光资源竞争能力的强弱[41].研究发现,植物叶片氮含量的高低会影响植物光合作用[54],氮添加可以短暂地消除氮素对沙地生态系统的限制,增加植物可利用氮素水平,使得植物对地下养分的竞争转化为对光的竞争[34],但过量的氮素会打破植物体内的养分平衡,降低植物光合速率[55].本研究中氮添加显著增加了植物群落高度,进一步验证了氮添加可以提高沙地植物群落的光合能力.植物比叶面积越高、叶干物质含量越低,其光捕获能力越强,这些变化有利于光竞争力的提高[48].本研究中氮添加使得群落叶面积显著增加,叶干物质含量显著降低,说明沙地植物群落主要通过叶面积和叶干物质含量的变化来响应氮添加.比叶面积在响应短期氮添加时敏感性较弱,且不同物种对氮添加的响应不同,而群落水平上很容易掩盖这种种间差异,因此比叶面积在群落水平上响应氮添加时没有明显变化. ...
羊草功能性状和地上生物量对氮素添加的响应
2
2016
... 氮作为植物生命活动必需的营养元素,对植物生长发育有着至关重要的作用[28].在氮沉降初期,氮素作为营养物质通过增加土壤氮含量从而促进植物的生长,尤其是生长在受氮素限制的生态系统中的植物[29-30].但过度氮沉降导致植物吸收过量的氮素,超过生态系统临界氮容量,打破植物体内固有的养分平衡,从而对植物个体、群落乃至整个生态系统产生影响[29,31].宋彦涛等[32]发现,氮添加提高了羊草地上生物量,比叶面积和叶片厚度无显著变化.杨浩等[33]发现,低氮添加主要影响糙隐子草叶片形态性状,而高氮添加主要影响叶生理性状.李禄军等[34]通过模拟氮沉降发现,氮素增加改变了植物群落物种组成和优势物种,使植物高度、盖度和生物量增加,而物种丰富度和生物多样性减少.Mao等[35]发现,氮添加增加了小叶章(Deyeuxia angustifolia)和狭叶甜茅(Glyceria spiculosa)的高度和地上生物量,但对其比叶面积和叶干物质含量无显著影响. ...
... 在干旱半干旱地区,植物生长不仅受水分限制,较低的氮含量也成为主要限制因子[50].目前,有关氮添加对植物群落生产力影响的研究报道有很多[31-32,51-52],研究结果也不尽相同.本研究中氮添加显著增加了群落盖度和地上生物量,密度也有一定程度的增加,说明短暂的氮添加在一定时期内减缓了该地区氮素对植物生长的限制,促进植物快速生长,增加生物量的累积,与前人研究结果基本一致[34,51].但氮添加也加剧了植物种间或种内竞争,导致一些竞争能力较弱的个体减少甚至死亡,这解释了本研究中群落密度虽有增加但不显著的原因.此外,本研究发现氮添加增加了群落凋落物量,这可能是由于过量的氮添加改变了植物养分再吸收的过程[53],使得器官脱落前植物个体减弱了对养分的再吸收能力[29],同时由于竞争造成个体死亡的增加也为凋落物量的增加做出了一定贡献.植物高度反映植物对光资源竞争能力的强弱[41].研究发现,植物叶片氮含量的高低会影响植物光合作用[54],氮添加可以短暂地消除氮素对沙地生态系统的限制,增加植物可利用氮素水平,使得植物对地下养分的竞争转化为对光的竞争[34],但过量的氮素会打破植物体内的养分平衡,降低植物光合速率[55].本研究中氮添加显著增加了植物群落高度,进一步验证了氮添加可以提高沙地植物群落的光合能力.植物比叶面积越高、叶干物质含量越低,其光捕获能力越强,这些变化有利于光竞争力的提高[48].本研究中氮添加使得群落叶面积显著增加,叶干物质含量显著降低,说明沙地植物群落主要通过叶面积和叶干物质含量的变化来响应氮添加.比叶面积在响应短期氮添加时敏感性较弱,且不同物种对氮添加的响应不同,而群落水平上很容易掩盖这种种间差异,因此比叶面积在群落水平上响应氮添加时没有明显变化. ...
糙隐子草功能性状对氮添加和干旱的响应
1
2015
... 氮作为植物生命活动必需的营养元素,对植物生长发育有着至关重要的作用[28].在氮沉降初期,氮素作为营养物质通过增加土壤氮含量从而促进植物的生长,尤其是生长在受氮素限制的生态系统中的植物[29-30].但过度氮沉降导致植物吸收过量的氮素,超过生态系统临界氮容量,打破植物体内固有的养分平衡,从而对植物个体、群落乃至整个生态系统产生影响[29,31].宋彦涛等[32]发现,氮添加提高了羊草地上生物量,比叶面积和叶片厚度无显著变化.杨浩等[33]发现,低氮添加主要影响糙隐子草叶片形态性状,而高氮添加主要影响叶生理性状.李禄军等[34]通过模拟氮沉降发现,氮素增加改变了植物群落物种组成和优势物种,使植物高度、盖度和生物量增加,而物种丰富度和生物多样性减少.Mao等[35]发现,氮添加增加了小叶章(Deyeuxia angustifolia)和狭叶甜茅(Glyceria spiculosa)的高度和地上生物量,但对其比叶面积和叶干物质含量无显著影响. ...
氮素添加对科尔沁沙质草地物种多样性和生产力的影响
3
2009
... 氮作为植物生命活动必需的营养元素,对植物生长发育有着至关重要的作用[28].在氮沉降初期,氮素作为营养物质通过增加土壤氮含量从而促进植物的生长,尤其是生长在受氮素限制的生态系统中的植物[29-30].但过度氮沉降导致植物吸收过量的氮素,超过生态系统临界氮容量,打破植物体内固有的养分平衡,从而对植物个体、群落乃至整个生态系统产生影响[29,31].宋彦涛等[32]发现,氮添加提高了羊草地上生物量,比叶面积和叶片厚度无显著变化.杨浩等[33]发现,低氮添加主要影响糙隐子草叶片形态性状,而高氮添加主要影响叶生理性状.李禄军等[34]通过模拟氮沉降发现,氮素增加改变了植物群落物种组成和优势物种,使植物高度、盖度和生物量增加,而物种丰富度和生物多样性减少.Mao等[35]发现,氮添加增加了小叶章(Deyeuxia angustifolia)和狭叶甜茅(Glyceria spiculosa)的高度和地上生物量,但对其比叶面积和叶干物质含量无显著影响. ...
... 在干旱半干旱地区,植物生长不仅受水分限制,较低的氮含量也成为主要限制因子[50].目前,有关氮添加对植物群落生产力影响的研究报道有很多[31-32,51-52],研究结果也不尽相同.本研究中氮添加显著增加了群落盖度和地上生物量,密度也有一定程度的增加,说明短暂的氮添加在一定时期内减缓了该地区氮素对植物生长的限制,促进植物快速生长,增加生物量的累积,与前人研究结果基本一致[34,51].但氮添加也加剧了植物种间或种内竞争,导致一些竞争能力较弱的个体减少甚至死亡,这解释了本研究中群落密度虽有增加但不显著的原因.此外,本研究发现氮添加增加了群落凋落物量,这可能是由于过量的氮添加改变了植物养分再吸收的过程[53],使得器官脱落前植物个体减弱了对养分的再吸收能力[29],同时由于竞争造成个体死亡的增加也为凋落物量的增加做出了一定贡献.植物高度反映植物对光资源竞争能力的强弱[41].研究发现,植物叶片氮含量的高低会影响植物光合作用[54],氮添加可以短暂地消除氮素对沙地生态系统的限制,增加植物可利用氮素水平,使得植物对地下养分的竞争转化为对光的竞争[34],但过量的氮素会打破植物体内的养分平衡,降低植物光合速率[55].本研究中氮添加显著增加了植物群落高度,进一步验证了氮添加可以提高沙地植物群落的光合能力.植物比叶面积越高、叶干物质含量越低,其光捕获能力越强,这些变化有利于光竞争力的提高[48].本研究中氮添加使得群落叶面积显著增加,叶干物质含量显著降低,说明沙地植物群落主要通过叶面积和叶干物质含量的变化来响应氮添加.比叶面积在响应短期氮添加时敏感性较弱,且不同物种对氮添加的响应不同,而群落水平上很容易掩盖这种种间差异,因此比叶面积在群落水平上响应氮添加时没有明显变化. ...
... [34],但过量的氮素会打破植物体内的养分平衡,降低植物光合速率[55].本研究中氮添加显著增加了植物群落高度,进一步验证了氮添加可以提高沙地植物群落的光合能力.植物比叶面积越高、叶干物质含量越低,其光捕获能力越强,这些变化有利于光竞争力的提高[48].本研究中氮添加使得群落叶面积显著增加,叶干物质含量显著降低,说明沙地植物群落主要通过叶面积和叶干物质含量的变化来响应氮添加.比叶面积在响应短期氮添加时敏感性较弱,且不同物种对氮添加的响应不同,而群落水平上很容易掩盖这种种间差异,因此比叶面积在群落水平上响应氮添加时没有明显变化. ...
Effects of nitrogen addition on plant functional traits in freshwater wetland of Sanjiang Plain,Northeast China
1
2014
... 氮作为植物生命活动必需的营养元素,对植物生长发育有着至关重要的作用[28].在氮沉降初期,氮素作为营养物质通过增加土壤氮含量从而促进植物的生长,尤其是生长在受氮素限制的生态系统中的植物[29-30].但过度氮沉降导致植物吸收过量的氮素,超过生态系统临界氮容量,打破植物体内固有的养分平衡,从而对植物个体、群落乃至整个生态系统产生影响[29,31].宋彦涛等[32]发现,氮添加提高了羊草地上生物量,比叶面积和叶片厚度无显著变化.杨浩等[33]发现,低氮添加主要影响糙隐子草叶片形态性状,而高氮添加主要影响叶生理性状.李禄军等[34]通过模拟氮沉降发现,氮素增加改变了植物群落物种组成和优势物种,使植物高度、盖度和生物量增加,而物种丰富度和生物多样性减少.Mao等[35]发现,氮添加增加了小叶章(Deyeuxia angustifolia)和狭叶甜茅(Glyceria spiculosa)的高度和地上生物量,但对其比叶面积和叶干物质含量无显著影响. ...
1
1996
... 全球草地总面积约占地球陆地总面积的1/6,中国草地约占国土面积的41.7%[36].过去几十年,由于人口的不断增加、土地利用不合理以及全球环境的剧烈变化,科尔沁沙地沙漠化进程加快,生态环境不断恶化,已成为中国北方农牧交错带沙漠化最为严重的地区[37].因此,本文以科尔沁沙质草地植物为研究对象,通过野外控制试验探究降雨变化、氮添加及其交互作用对群落植被特征和叶性状的影响,旨在探讨沙地植物群落性状对全球气候变化的响应与适应,掌握沙地植物群落为适应环境所形成的特有生态适应策略,从而为干旱半干旱沙地生态系统退化植被的恢复及其可持续管理提供理论基础. ...
我国北方农牧交错带的草地植被类型、特征及其生态问题
1
2003
... 全球草地总面积约占地球陆地总面积的1/6,中国草地约占国土面积的41.7%[36].过去几十年,由于人口的不断增加、土地利用不合理以及全球环境的剧烈变化,科尔沁沙地沙漠化进程加快,生态环境不断恶化,已成为中国北方农牧交错带沙漠化最为严重的地区[37].因此,本文以科尔沁沙质草地植物为研究对象,通过野外控制试验探究降雨变化、氮添加及其交互作用对群落植被特征和叶性状的影响,旨在探讨沙地植物群落性状对全球气候变化的响应与适应,掌握沙地植物群落为适应环境所形成的特有生态适应策略,从而为干旱半干旱沙地生态系统退化植被的恢复及其可持续管理提供理论基础. ...
近40 a我国北方农牧交错区气候变化及其与土地沙漠化的关系
1
2000
... 研究区(42°55′—42°57′N,120°40′—120°43′E)位于内蒙古通辽市奈曼旗境内,地处科尔沁沙地中南部,属温带大陆性半干旱季风气候,海拔约360 m,年均气温约6.4 ℃,1月最低气温 -13.1 ℃,7月最高气温23.7 ℃,全年≥10 ℃的有效积温大于3 000 ℃,无霜期150 d,年降水量约360 mm,其中70%~80%的降水在6—8月,年蒸发量1 972.8 mm.年均风速3.2~4.1 m·s-1,春冬两季盛行西北风,夏秋两季为西南风[38].该研究区流动沙丘、半固定沙丘、固定沙丘、丘间低地等各种沙丘类型交错分布,土壤类型以风沙土、沼泽土、沙质栗钙土为主.试验区主要植物有狗尾草(Setaria viridis)、白草(Pennisetum centrasiaticum)、砂蓝刺头(Echinops gmelini)、尖头叶藜(Cleistogenes squarrosa)、差巴嘎蒿(Artemisia halodendron)、黄蒿(Artemisia scoparia)、糙隐子草(Cleistogenes squarrosa)等[39]. ...
科尔沁沙地封育过程中植被特征的动态变化
1
2018
... 研究区(42°55′—42°57′N,120°40′—120°43′E)位于内蒙古通辽市奈曼旗境内,地处科尔沁沙地中南部,属温带大陆性半干旱季风气候,海拔约360 m,年均气温约6.4 ℃,1月最低气温 -13.1 ℃,7月最高气温23.7 ℃,全年≥10 ℃的有效积温大于3 000 ℃,无霜期150 d,年降水量约360 mm,其中70%~80%的降水在6—8月,年蒸发量1 972.8 mm.年均风速3.2~4.1 m·s-1,春冬两季盛行西北风,夏秋两季为西南风[38].该研究区流动沙丘、半固定沙丘、固定沙丘、丘间低地等各种沙丘类型交错分布,土壤类型以风沙土、沼泽土、沙质栗钙土为主.试验区主要植物有狗尾草(Setaria viridis)、白草(Pennisetum centrasiaticum)、砂蓝刺头(Echinops gmelini)、尖头叶藜(Cleistogenes squarrosa)、差巴嘎蒿(Artemisia halodendron)、黄蒿(Artemisia scoparia)、糙隐子草(Cleistogenes squarrosa)等[39]. ...
Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis
1
2010
... 本研究依托中国科学院西北生态环境资源研究院奈曼沙漠化研究站,选取水氮交互处理的野外综合试验场地开展试验.为探究降雨量和降雨时间分配对半干旱沙质草地植物生长的影响,根据该区域长期监测的多年平均降水量设置水氮处理,降雨处理包括:对照(W0)、增减雨60%(W-60%与W+60%,5—7月)、增减雨100%(W-100%与W+100%,5—6月);氮添加处理包括不加氮(N0)和添加氮(N+).每个处理6个重复,共60个小区.模拟降雨试验采用搭建遮雨棚的方法进行,其中凹槽遮雨板的材质为高透光聚碳酸酯,透光率达90%.每个试验小区的遮雨棚面积为6 m×6 m,相邻试验小区间设置2 m的缓冲带,生长季结束后拆除遮雨板.为保证空气畅通将遮雨棚四面敞开.模拟氮沉降时使用尿素进行氮添加处理,氮添加量为20 g·m-2·a-1,分两次施,每次10 g·m-2·a-1[40].施肥时间选择5月和7月,氮肥施用量根据小区面积和尿素含氮量(46%)计算.为减小水分和氮素侧向干扰,在每个小区的四周挖1 m深,用包有塑料纸的金属隔板隔开.本研究选取的样地及试验处理位于连续的同一地段,地势平坦,环境条件相对均质,有效地避免了空间异质性的干扰[41].试验期间对试验地进行围封,避免家畜及人为干扰. ...
内蒙古不同草地类型隐子草种群对放牧强度的响应
2
2016
... 本研究依托中国科学院西北生态环境资源研究院奈曼沙漠化研究站,选取水氮交互处理的野外综合试验场地开展试验.为探究降雨量和降雨时间分配对半干旱沙质草地植物生长的影响,根据该区域长期监测的多年平均降水量设置水氮处理,降雨处理包括:对照(W0)、增减雨60%(W-60%与W+60%,5—7月)、增减雨100%(W-100%与W+100%,5—6月);氮添加处理包括不加氮(N0)和添加氮(N+).每个处理6个重复,共60个小区.模拟降雨试验采用搭建遮雨棚的方法进行,其中凹槽遮雨板的材质为高透光聚碳酸酯,透光率达90%.每个试验小区的遮雨棚面积为6 m×6 m,相邻试验小区间设置2 m的缓冲带,生长季结束后拆除遮雨板.为保证空气畅通将遮雨棚四面敞开.模拟氮沉降时使用尿素进行氮添加处理,氮添加量为20 g·m-2·a-1,分两次施,每次10 g·m-2·a-1[40].施肥时间选择5月和7月,氮肥施用量根据小区面积和尿素含氮量(46%)计算.为减小水分和氮素侧向干扰,在每个小区的四周挖1 m深,用包有塑料纸的金属隔板隔开.本研究选取的样地及试验处理位于连续的同一地段,地势平坦,环境条件相对均质,有效地避免了空间异质性的干扰[41].试验期间对试验地进行围封,避免家畜及人为干扰. ...
... 在干旱半干旱地区,植物生长不仅受水分限制,较低的氮含量也成为主要限制因子[50].目前,有关氮添加对植物群落生产力影响的研究报道有很多[31-32,51-52],研究结果也不尽相同.本研究中氮添加显著增加了群落盖度和地上生物量,密度也有一定程度的增加,说明短暂的氮添加在一定时期内减缓了该地区氮素对植物生长的限制,促进植物快速生长,增加生物量的累积,与前人研究结果基本一致[34,51].但氮添加也加剧了植物种间或种内竞争,导致一些竞争能力较弱的个体减少甚至死亡,这解释了本研究中群落密度虽有增加但不显著的原因.此外,本研究发现氮添加增加了群落凋落物量,这可能是由于过量的氮添加改变了植物养分再吸收的过程[53],使得器官脱落前植物个体减弱了对养分的再吸收能力[29],同时由于竞争造成个体死亡的增加也为凋落物量的增加做出了一定贡献.植物高度反映植物对光资源竞争能力的强弱[41].研究发现,植物叶片氮含量的高低会影响植物光合作用[54],氮添加可以短暂地消除氮素对沙地生态系统的限制,增加植物可利用氮素水平,使得植物对地下养分的竞争转化为对光的竞争[34],但过量的氮素会打破植物体内的养分平衡,降低植物光合速率[55].本研究中氮添加显著增加了植物群落高度,进一步验证了氮添加可以提高沙地植物群落的光合能力.植物比叶面积越高、叶干物质含量越低,其光捕获能力越强,这些变化有利于光竞争力的提高[48].本研究中氮添加使得群落叶面积显著增加,叶干物质含量显著降低,说明沙地植物群落主要通过叶面积和叶干物质含量的变化来响应氮添加.比叶面积在响应短期氮添加时敏感性较弱,且不同物种对氮添加的响应不同,而群落水平上很容易掩盖这种种间差异,因此比叶面积在群落水平上响应氮添加时没有明显变化. ...
A handbook of protocols for standardised and easy measurement of plant functional traits worldwide
1
2003
... 于2019年8月生物量高峰期进行调查取样,采样时为了避免边缘效应,限制采样点离边界至少1 m.在每个样地内随机设置3个1 m×1 m的样方,调查样方中植物群落物种组成、高度、盖度和密度,并采用齐地面刈割法分物种收获每种植物,同时收取地上的凋落物,装入信封袋带回实验室,70 ℃下烘干至恒重后称重.每个样方中选择2~4种优势物种,参照Cornelissen等[42]的《植物功能性状标准测定手册》,测定并计算每个样方内优势物种的叶面积(cm2)、比叶面积(m2·kg-1)和叶干物质含量(g·g-1).比叶面积=叶面积/叶干重;叶干物质含量=叶干重/叶饱和鲜重. ...
Biodiversity enhances ecosystem reliability
1
1997
... 群落水平的植物性状值用植物群落功能性状(CWM)表示[43]. ...
Water resources: an overview of world deserts
1
1991
... 降雨是沙地植被形成和发展的主要制约因素[44],降雨量的变化会影响土壤含水量发生变化,通常土壤水分增加会促进植物生长[45].本研究中,减雨(W-60%与W-100%)显著降低植物群落盖度和地上生物量,说明极端干旱不利于沙地植物群落生产力的提高,而增雨(W+60%与W+100%)对其无显著影响,这可能是由于样地中大部分植物为沙生植物,适当增雨会增加群落生产力,但过度增雨可能不利于甚至会抑制沙生植物的生长,使其出现不适应的现象,从而导致增雨对植物群落盖度、地上生物量等影响不明显.增雨60%后群落密度显著高于减雨(W-60%与W-100%),说明增雨60%对群落密度影响更大.张腊梅等[6]发现,相比于增雨60%,增雨30%更有利于植物生长,这与本研究结果有一定的差异,可能是由于试验期间降雨量不同引起的.凋落物是植物将养分返还土壤的一种形式[46],由于一年生植物通常会在较短的时间内快速完成资源的积累和生活史周期[47],因此随着降雨的减少植物会加快衰老的速度,进而产生的凋落物会增多,而本研究发现减雨(W-60%与W-100%)显著降低了群落凋落物量,这可能是由于沙地中原有的现存生物量就少,在降雨减少的情况下,新生的现存生物量更少,进而产生的凋落物也就更少[14].本研究中植物群落高度在减雨60%处理下最低,之后随着降雨的增加而逐渐增加,但相比于增雨60%,增雨100%处理下群落高度几乎没有增加.同样地,叶面积在增雨60%处理下到达最大值,尤其显著高于对照和减雨60%处理,说明相比于增雨100%,增雨60%可以更好地提高植物对光资源的获取能力,表明降雨超过一定范围后植物群落对降雨变化的响应会减弱.比叶面积和叶干物质含量反映植物对资源的利用状况,两者都易受水分的束缚,但叶干物质含量很少受其他性状的影响,如叶片厚度等[48].本研究发现降雨变化对植物群落比叶面积和叶干物质含量均无显著影响,可能是由于降雨对植物的影响具有一定的滞后性[20,49],且试验周期较短. ...
水分和养分对内蒙古荒漠草原植被恢复及碳储量的影响
1
2012
... 降雨是沙地植被形成和发展的主要制约因素[44],降雨量的变化会影响土壤含水量发生变化,通常土壤水分增加会促进植物生长[45].本研究中,减雨(W-60%与W-100%)显著降低植物群落盖度和地上生物量,说明极端干旱不利于沙地植物群落生产力的提高,而增雨(W+60%与W+100%)对其无显著影响,这可能是由于样地中大部分植物为沙生植物,适当增雨会增加群落生产力,但过度增雨可能不利于甚至会抑制沙生植物的生长,使其出现不适应的现象,从而导致增雨对植物群落盖度、地上生物量等影响不明显.增雨60%后群落密度显著高于减雨(W-60%与W-100%),说明增雨60%对群落密度影响更大.张腊梅等[6]发现,相比于增雨60%,增雨30%更有利于植物生长,这与本研究结果有一定的差异,可能是由于试验期间降雨量不同引起的.凋落物是植物将养分返还土壤的一种形式[46],由于一年生植物通常会在较短的时间内快速完成资源的积累和生活史周期[47],因此随着降雨的减少植物会加快衰老的速度,进而产生的凋落物会增多,而本研究发现减雨(W-60%与W-100%)显著降低了群落凋落物量,这可能是由于沙地中原有的现存生物量就少,在降雨减少的情况下,新生的现存生物量更少,进而产生的凋落物也就更少[14].本研究中植物群落高度在减雨60%处理下最低,之后随着降雨的增加而逐渐增加,但相比于增雨60%,增雨100%处理下群落高度几乎没有增加.同样地,叶面积在增雨60%处理下到达最大值,尤其显著高于对照和减雨60%处理,说明相比于增雨100%,增雨60%可以更好地提高植物对光资源的获取能力,表明降雨超过一定范围后植物群落对降雨变化的响应会减弱.比叶面积和叶干物质含量反映植物对资源的利用状况,两者都易受水分的束缚,但叶干物质含量很少受其他性状的影响,如叶片厚度等[48].本研究发现降雨变化对植物群落比叶面积和叶干物质含量均无显著影响,可能是由于降雨对植物的影响具有一定的滞后性[20,49],且试验周期较短. ...
Global pattern of leaf litter nitrogen and phosphorus in woody plants
1
2010
... 降雨是沙地植被形成和发展的主要制约因素[44],降雨量的变化会影响土壤含水量发生变化,通常土壤水分增加会促进植物生长[45].本研究中,减雨(W-60%与W-100%)显著降低植物群落盖度和地上生物量,说明极端干旱不利于沙地植物群落生产力的提高,而增雨(W+60%与W+100%)对其无显著影响,这可能是由于样地中大部分植物为沙生植物,适当增雨会增加群落生产力,但过度增雨可能不利于甚至会抑制沙生植物的生长,使其出现不适应的现象,从而导致增雨对植物群落盖度、地上生物量等影响不明显.增雨60%后群落密度显著高于减雨(W-60%与W-100%),说明增雨60%对群落密度影响更大.张腊梅等[6]发现,相比于增雨60%,增雨30%更有利于植物生长,这与本研究结果有一定的差异,可能是由于试验期间降雨量不同引起的.凋落物是植物将养分返还土壤的一种形式[46],由于一年生植物通常会在较短的时间内快速完成资源的积累和生活史周期[47],因此随着降雨的减少植物会加快衰老的速度,进而产生的凋落物会增多,而本研究发现减雨(W-60%与W-100%)显著降低了群落凋落物量,这可能是由于沙地中原有的现存生物量就少,在降雨减少的情况下,新生的现存生物量更少,进而产生的凋落物也就更少[14].本研究中植物群落高度在减雨60%处理下最低,之后随着降雨的增加而逐渐增加,但相比于增雨60%,增雨100%处理下群落高度几乎没有增加.同样地,叶面积在增雨60%处理下到达最大值,尤其显著高于对照和减雨60%处理,说明相比于增雨100%,增雨60%可以更好地提高植物对光资源的获取能力,表明降雨超过一定范围后植物群落对降雨变化的响应会减弱.比叶面积和叶干物质含量反映植物对资源的利用状况,两者都易受水分的束缚,但叶干物质含量很少受其他性状的影响,如叶片厚度等[48].本研究发现降雨变化对植物群落比叶面积和叶干物质含量均无显著影响,可能是由于降雨对植物的影响具有一定的滞后性[20,49],且试验周期较短. ...
The worldwide leaf economics spectrum
1
2004
... 降雨是沙地植被形成和发展的主要制约因素[44],降雨量的变化会影响土壤含水量发生变化,通常土壤水分增加会促进植物生长[45].本研究中,减雨(W-60%与W-100%)显著降低植物群落盖度和地上生物量,说明极端干旱不利于沙地植物群落生产力的提高,而增雨(W+60%与W+100%)对其无显著影响,这可能是由于样地中大部分植物为沙生植物,适当增雨会增加群落生产力,但过度增雨可能不利于甚至会抑制沙生植物的生长,使其出现不适应的现象,从而导致增雨对植物群落盖度、地上生物量等影响不明显.增雨60%后群落密度显著高于减雨(W-60%与W-100%),说明增雨60%对群落密度影响更大.张腊梅等[6]发现,相比于增雨60%,增雨30%更有利于植物生长,这与本研究结果有一定的差异,可能是由于试验期间降雨量不同引起的.凋落物是植物将养分返还土壤的一种形式[46],由于一年生植物通常会在较短的时间内快速完成资源的积累和生活史周期[47],因此随着降雨的减少植物会加快衰老的速度,进而产生的凋落物会增多,而本研究发现减雨(W-60%与W-100%)显著降低了群落凋落物量,这可能是由于沙地中原有的现存生物量就少,在降雨减少的情况下,新生的现存生物量更少,进而产生的凋落物也就更少[14].本研究中植物群落高度在减雨60%处理下最低,之后随着降雨的增加而逐渐增加,但相比于增雨60%,增雨100%处理下群落高度几乎没有增加.同样地,叶面积在增雨60%处理下到达最大值,尤其显著高于对照和减雨60%处理,说明相比于增雨100%,增雨60%可以更好地提高植物对光资源的获取能力,表明降雨超过一定范围后植物群落对降雨变化的响应会减弱.比叶面积和叶干物质含量反映植物对资源的利用状况,两者都易受水分的束缚,但叶干物质含量很少受其他性状的影响,如叶片厚度等[48].本研究发现降雨变化对植物群落比叶面积和叶干物质含量均无显著影响,可能是由于降雨对植物的影响具有一定的滞后性[20,49],且试验周期较短. ...
Specific leaf area and leaf dry matter content as alternative predictors of plant strategies
2
1999
... 降雨是沙地植被形成和发展的主要制约因素[44],降雨量的变化会影响土壤含水量发生变化,通常土壤水分增加会促进植物生长[45].本研究中,减雨(W-60%与W-100%)显著降低植物群落盖度和地上生物量,说明极端干旱不利于沙地植物群落生产力的提高,而增雨(W+60%与W+100%)对其无显著影响,这可能是由于样地中大部分植物为沙生植物,适当增雨会增加群落生产力,但过度增雨可能不利于甚至会抑制沙生植物的生长,使其出现不适应的现象,从而导致增雨对植物群落盖度、地上生物量等影响不明显.增雨60%后群落密度显著高于减雨(W-60%与W-100%),说明增雨60%对群落密度影响更大.张腊梅等[6]发现,相比于增雨60%,增雨30%更有利于植物生长,这与本研究结果有一定的差异,可能是由于试验期间降雨量不同引起的.凋落物是植物将养分返还土壤的一种形式[46],由于一年生植物通常会在较短的时间内快速完成资源的积累和生活史周期[47],因此随着降雨的减少植物会加快衰老的速度,进而产生的凋落物会增多,而本研究发现减雨(W-60%与W-100%)显著降低了群落凋落物量,这可能是由于沙地中原有的现存生物量就少,在降雨减少的情况下,新生的现存生物量更少,进而产生的凋落物也就更少[14].本研究中植物群落高度在减雨60%处理下最低,之后随着降雨的增加而逐渐增加,但相比于增雨60%,增雨100%处理下群落高度几乎没有增加.同样地,叶面积在增雨60%处理下到达最大值,尤其显著高于对照和减雨60%处理,说明相比于增雨100%,增雨60%可以更好地提高植物对光资源的获取能力,表明降雨超过一定范围后植物群落对降雨变化的响应会减弱.比叶面积和叶干物质含量反映植物对资源的利用状况,两者都易受水分的束缚,但叶干物质含量很少受其他性状的影响,如叶片厚度等[48].本研究发现降雨变化对植物群落比叶面积和叶干物质含量均无显著影响,可能是由于降雨对植物的影响具有一定的滞后性[20,49],且试验周期较短. ...
... 在干旱半干旱地区,植物生长不仅受水分限制,较低的氮含量也成为主要限制因子[50].目前,有关氮添加对植物群落生产力影响的研究报道有很多[31-32,51-52],研究结果也不尽相同.本研究中氮添加显著增加了群落盖度和地上生物量,密度也有一定程度的增加,说明短暂的氮添加在一定时期内减缓了该地区氮素对植物生长的限制,促进植物快速生长,增加生物量的累积,与前人研究结果基本一致[34,51].但氮添加也加剧了植物种间或种内竞争,导致一些竞争能力较弱的个体减少甚至死亡,这解释了本研究中群落密度虽有增加但不显著的原因.此外,本研究发现氮添加增加了群落凋落物量,这可能是由于过量的氮添加改变了植物养分再吸收的过程[53],使得器官脱落前植物个体减弱了对养分的再吸收能力[29],同时由于竞争造成个体死亡的增加也为凋落物量的增加做出了一定贡献.植物高度反映植物对光资源竞争能力的强弱[41].研究发现,植物叶片氮含量的高低会影响植物光合作用[54],氮添加可以短暂地消除氮素对沙地生态系统的限制,增加植物可利用氮素水平,使得植物对地下养分的竞争转化为对光的竞争[34],但过量的氮素会打破植物体内的养分平衡,降低植物光合速率[55].本研究中氮添加显著增加了植物群落高度,进一步验证了氮添加可以提高沙地植物群落的光合能力.植物比叶面积越高、叶干物质含量越低,其光捕获能力越强,这些变化有利于光竞争力的提高[48].本研究中氮添加使得群落叶面积显著增加,叶干物质含量显著降低,说明沙地植物群落主要通过叶面积和叶干物质含量的变化来响应氮添加.比叶面积在响应短期氮添加时敏感性较弱,且不同物种对氮添加的响应不同,而群落水平上很容易掩盖这种种间差异,因此比叶面积在群落水平上响应氮添加时没有明显变化. ...
内蒙古草地生长季植被变化对气候因子的响应
1
2013
... 降雨是沙地植被形成和发展的主要制约因素[44],降雨量的变化会影响土壤含水量发生变化,通常土壤水分增加会促进植物生长[45].本研究中,减雨(W-60%与W-100%)显著降低植物群落盖度和地上生物量,说明极端干旱不利于沙地植物群落生产力的提高,而增雨(W+60%与W+100%)对其无显著影响,这可能是由于样地中大部分植物为沙生植物,适当增雨会增加群落生产力,但过度增雨可能不利于甚至会抑制沙生植物的生长,使其出现不适应的现象,从而导致增雨对植物群落盖度、地上生物量等影响不明显.增雨60%后群落密度显著高于减雨(W-60%与W-100%),说明增雨60%对群落密度影响更大.张腊梅等[6]发现,相比于增雨60%,增雨30%更有利于植物生长,这与本研究结果有一定的差异,可能是由于试验期间降雨量不同引起的.凋落物是植物将养分返还土壤的一种形式[46],由于一年生植物通常会在较短的时间内快速完成资源的积累和生活史周期[47],因此随着降雨的减少植物会加快衰老的速度,进而产生的凋落物会增多,而本研究发现减雨(W-60%与W-100%)显著降低了群落凋落物量,这可能是由于沙地中原有的现存生物量就少,在降雨减少的情况下,新生的现存生物量更少,进而产生的凋落物也就更少[14].本研究中植物群落高度在减雨60%处理下最低,之后随着降雨的增加而逐渐增加,但相比于增雨60%,增雨100%处理下群落高度几乎没有增加.同样地,叶面积在增雨60%处理下到达最大值,尤其显著高于对照和减雨60%处理,说明相比于增雨100%,增雨60%可以更好地提高植物对光资源的获取能力,表明降雨超过一定范围后植物群落对降雨变化的响应会减弱.比叶面积和叶干物质含量反映植物对资源的利用状况,两者都易受水分的束缚,但叶干物质含量很少受其他性状的影响,如叶片厚度等[48].本研究发现降雨变化对植物群落比叶面积和叶干物质含量均无显著影响,可能是由于降雨对植物的影响具有一定的滞后性[20,49],且试验周期较短. ...
Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed
1
2008
... 在干旱半干旱地区,植物生长不仅受水分限制,较低的氮含量也成为主要限制因子[50].目前,有关氮添加对植物群落生产力影响的研究报道有很多[31-32,51-52],研究结果也不尽相同.本研究中氮添加显著增加了群落盖度和地上生物量,密度也有一定程度的增加,说明短暂的氮添加在一定时期内减缓了该地区氮素对植物生长的限制,促进植物快速生长,增加生物量的累积,与前人研究结果基本一致[34,51].但氮添加也加剧了植物种间或种内竞争,导致一些竞争能力较弱的个体减少甚至死亡,这解释了本研究中群落密度虽有增加但不显著的原因.此外,本研究发现氮添加增加了群落凋落物量,这可能是由于过量的氮添加改变了植物养分再吸收的过程[53],使得器官脱落前植物个体减弱了对养分的再吸收能力[29],同时由于竞争造成个体死亡的增加也为凋落物量的增加做出了一定贡献.植物高度反映植物对光资源竞争能力的强弱[41].研究发现,植物叶片氮含量的高低会影响植物光合作用[54],氮添加可以短暂地消除氮素对沙地生态系统的限制,增加植物可利用氮素水平,使得植物对地下养分的竞争转化为对光的竞争[34],但过量的氮素会打破植物体内的养分平衡,降低植物光合速率[55].本研究中氮添加显著增加了植物群落高度,进一步验证了氮添加可以提高沙地植物群落的光合能力.植物比叶面积越高、叶干物质含量越低,其光捕获能力越强,这些变化有利于光竞争力的提高[48].本研究中氮添加使得群落叶面积显著增加,叶干物质含量显著降低,说明沙地植物群落主要通过叶面积和叶干物质含量的变化来响应氮添加.比叶面积在响应短期氮添加时敏感性较弱,且不同物种对氮添加的响应不同,而群落水平上很容易掩盖这种种间差异,因此比叶面积在群落水平上响应氮添加时没有明显变化. ...
Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning:evidence from inner Mongolia Grasslands
2
2010
... 在干旱半干旱地区,植物生长不仅受水分限制,较低的氮含量也成为主要限制因子[50].目前,有关氮添加对植物群落生产力影响的研究报道有很多[31-32,51-52],研究结果也不尽相同.本研究中氮添加显著增加了群落盖度和地上生物量,密度也有一定程度的增加,说明短暂的氮添加在一定时期内减缓了该地区氮素对植物生长的限制,促进植物快速生长,增加生物量的累积,与前人研究结果基本一致[34,51].但氮添加也加剧了植物种间或种内竞争,导致一些竞争能力较弱的个体减少甚至死亡,这解释了本研究中群落密度虽有增加但不显著的原因.此外,本研究发现氮添加增加了群落凋落物量,这可能是由于过量的氮添加改变了植物养分再吸收的过程[53],使得器官脱落前植物个体减弱了对养分的再吸收能力[29],同时由于竞争造成个体死亡的增加也为凋落物量的增加做出了一定贡献.植物高度反映植物对光资源竞争能力的强弱[41].研究发现,植物叶片氮含量的高低会影响植物光合作用[54],氮添加可以短暂地消除氮素对沙地生态系统的限制,增加植物可利用氮素水平,使得植物对地下养分的竞争转化为对光的竞争[34],但过量的氮素会打破植物体内的养分平衡,降低植物光合速率[55].本研究中氮添加显著增加了植物群落高度,进一步验证了氮添加可以提高沙地植物群落的光合能力.植物比叶面积越高、叶干物质含量越低,其光捕获能力越强,这些变化有利于光竞争力的提高[48].本研究中氮添加使得群落叶面积显著增加,叶干物质含量显著降低,说明沙地植物群落主要通过叶面积和叶干物质含量的变化来响应氮添加.比叶面积在响应短期氮添加时敏感性较弱,且不同物种对氮添加的响应不同,而群落水平上很容易掩盖这种种间差异,因此比叶面积在群落水平上响应氮添加时没有明显变化. ...
... ,51].但氮添加也加剧了植物种间或种内竞争,导致一些竞争能力较弱的个体减少甚至死亡,这解释了本研究中群落密度虽有增加但不显著的原因.此外,本研究发现氮添加增加了群落凋落物量,这可能是由于过量的氮添加改变了植物养分再吸收的过程[53],使得器官脱落前植物个体减弱了对养分的再吸收能力[29],同时由于竞争造成个体死亡的增加也为凋落物量的增加做出了一定贡献.植物高度反映植物对光资源竞争能力的强弱[41].研究发现,植物叶片氮含量的高低会影响植物光合作用[54],氮添加可以短暂地消除氮素对沙地生态系统的限制,增加植物可利用氮素水平,使得植物对地下养分的竞争转化为对光的竞争[34],但过量的氮素会打破植物体内的养分平衡,降低植物光合速率[55].本研究中氮添加显著增加了植物群落高度,进一步验证了氮添加可以提高沙地植物群落的光合能力.植物比叶面积越高、叶干物质含量越低,其光捕获能力越强,这些变化有利于光竞争力的提高[48].本研究中氮添加使得群落叶面积显著增加,叶干物质含量显著降低,说明沙地植物群落主要通过叶面积和叶干物质含量的变化来响应氮添加.比叶面积在响应短期氮添加时敏感性较弱,且不同物种对氮添加的响应不同,而群落水平上很容易掩盖这种种间差异,因此比叶面积在群落水平上响应氮添加时没有明显变化. ...
Plant nutrients do not covary with soil nutrients under changing climatic conditions
2
2015
... 在干旱半干旱地区,植物生长不仅受水分限制,较低的氮含量也成为主要限制因子[50].目前,有关氮添加对植物群落生产力影响的研究报道有很多[31-32,51-52],研究结果也不尽相同.本研究中氮添加显著增加了群落盖度和地上生物量,密度也有一定程度的增加,说明短暂的氮添加在一定时期内减缓了该地区氮素对植物生长的限制,促进植物快速生长,增加生物量的累积,与前人研究结果基本一致[34,51].但氮添加也加剧了植物种间或种内竞争,导致一些竞争能力较弱的个体减少甚至死亡,这解释了本研究中群落密度虽有增加但不显著的原因.此外,本研究发现氮添加增加了群落凋落物量,这可能是由于过量的氮添加改变了植物养分再吸收的过程[53],使得器官脱落前植物个体减弱了对养分的再吸收能力[29],同时由于竞争造成个体死亡的增加也为凋落物量的增加做出了一定贡献.植物高度反映植物对光资源竞争能力的强弱[41].研究发现,植物叶片氮含量的高低会影响植物光合作用[54],氮添加可以短暂地消除氮素对沙地生态系统的限制,增加植物可利用氮素水平,使得植物对地下养分的竞争转化为对光的竞争[34],但过量的氮素会打破植物体内的养分平衡,降低植物光合速率[55].本研究中氮添加显著增加了植物群落高度,进一步验证了氮添加可以提高沙地植物群落的光合能力.植物比叶面积越高、叶干物质含量越低,其光捕获能力越强,这些变化有利于光竞争力的提高[48].本研究中氮添加使得群落叶面积显著增加,叶干物质含量显著降低,说明沙地植物群落主要通过叶面积和叶干物质含量的变化来响应氮添加.比叶面积在响应短期氮添加时敏感性较弱,且不同物种对氮添加的响应不同,而群落水平上很容易掩盖这种种间差异,因此比叶面积在群落水平上响应氮添加时没有明显变化. ...
... 水氮之间对植物生长存在相互补偿机制,主要表现为氮添加能缓解干旱对植物造成的影响,但同时干旱可以减弱甚至抑制氮对植物生长的促进作用[56].增雨(W+60%和W+100%)处理下,氮添加使得群落盖度显著增加,群落密度、地上生物量和凋落物量在增雨60%和氮添加交互处理下达到最大值,而增雨100%与氮添加耦合反而使其降低,说明适当增雨更有利于氮素肥力的激发,降雨过多反而会缩短氮素滞留时间,造成氮素流失[15].同一干旱处理(W-60%与W-100%)下,氮添加增加了群落高度和叶面积,尤其减雨60%处理下氮添加显著提高了群落高度,说明氮添加可以在一定程度上缓解干旱对植物造成的影响.同样地,同一增雨处理(W+60%和W+100%)下氮添加也使得高度和叶面积有所增加,这可能是由于增雨更好地激发了氮肥效力,提高了土壤中氮素的可利用性,进而促进植物对氮素的吸收和利用效率.此外,与增雨100%相比,叶面积在增雨60%和氮添加交互处理下最大,说明并不是降雨越多越有利于植物生长,超过一定范围后反而会抑制植物生长.就叶干物质含量而言,在相同降雨处理下,氮添加降低了植物群落叶干物质含量,尤其在W-60%和W0处理下叶干物质含量显著降低,而比叶面积在水氮交互处理下虽有波动变化,但无明显变化规律,这可能是由于不同物种在面对气候变化时所采取的策略不同[52]. ...
5种荒漠短命植物养分再吸收对水氮添加的响应
1
2016
... 在干旱半干旱地区,植物生长不仅受水分限制,较低的氮含量也成为主要限制因子[50].目前,有关氮添加对植物群落生产力影响的研究报道有很多[31-32,51-52],研究结果也不尽相同.本研究中氮添加显著增加了群落盖度和地上生物量,密度也有一定程度的增加,说明短暂的氮添加在一定时期内减缓了该地区氮素对植物生长的限制,促进植物快速生长,增加生物量的累积,与前人研究结果基本一致[34,51].但氮添加也加剧了植物种间或种内竞争,导致一些竞争能力较弱的个体减少甚至死亡,这解释了本研究中群落密度虽有增加但不显著的原因.此外,本研究发现氮添加增加了群落凋落物量,这可能是由于过量的氮添加改变了植物养分再吸收的过程[53],使得器官脱落前植物个体减弱了对养分的再吸收能力[29],同时由于竞争造成个体死亡的增加也为凋落物量的增加做出了一定贡献.植物高度反映植物对光资源竞争能力的强弱[41].研究发现,植物叶片氮含量的高低会影响植物光合作用[54],氮添加可以短暂地消除氮素对沙地生态系统的限制,增加植物可利用氮素水平,使得植物对地下养分的竞争转化为对光的竞争[34],但过量的氮素会打破植物体内的养分平衡,降低植物光合速率[55].本研究中氮添加显著增加了植物群落高度,进一步验证了氮添加可以提高沙地植物群落的光合能力.植物比叶面积越高、叶干物质含量越低,其光捕获能力越强,这些变化有利于光竞争力的提高[48].本研究中氮添加使得群落叶面积显著增加,叶干物质含量显著降低,说明沙地植物群落主要通过叶面积和叶干物质含量的变化来响应氮添加.比叶面积在响应短期氮添加时敏感性较弱,且不同物种对氮添加的响应不同,而群落水平上很容易掩盖这种种间差异,因此比叶面积在群落水平上响应氮添加时没有明显变化. ...
荒漠草原植物群落光合速率对水氮添加的响应
1
2019
... 在干旱半干旱地区,植物生长不仅受水分限制,较低的氮含量也成为主要限制因子[50].目前,有关氮添加对植物群落生产力影响的研究报道有很多[31-32,51-52],研究结果也不尽相同.本研究中氮添加显著增加了群落盖度和地上生物量,密度也有一定程度的增加,说明短暂的氮添加在一定时期内减缓了该地区氮素对植物生长的限制,促进植物快速生长,增加生物量的累积,与前人研究结果基本一致[34,51].但氮添加也加剧了植物种间或种内竞争,导致一些竞争能力较弱的个体减少甚至死亡,这解释了本研究中群落密度虽有增加但不显著的原因.此外,本研究发现氮添加增加了群落凋落物量,这可能是由于过量的氮添加改变了植物养分再吸收的过程[53],使得器官脱落前植物个体减弱了对养分的再吸收能力[29],同时由于竞争造成个体死亡的增加也为凋落物量的增加做出了一定贡献.植物高度反映植物对光资源竞争能力的强弱[41].研究发现,植物叶片氮含量的高低会影响植物光合作用[54],氮添加可以短暂地消除氮素对沙地生态系统的限制,增加植物可利用氮素水平,使得植物对地下养分的竞争转化为对光的竞争[34],但过量的氮素会打破植物体内的养分平衡,降低植物光合速率[55].本研究中氮添加显著增加了植物群落高度,进一步验证了氮添加可以提高沙地植物群落的光合能力.植物比叶面积越高、叶干物质含量越低,其光捕获能力越强,这些变化有利于光竞争力的提高[48].本研究中氮添加使得群落叶面积显著增加,叶干物质含量显著降低,说明沙地植物群落主要通过叶面积和叶干物质含量的变化来响应氮添加.比叶面积在响应短期氮添加时敏感性较弱,且不同物种对氮添加的响应不同,而群落水平上很容易掩盖这种种间差异,因此比叶面积在群落水平上响应氮添加时没有明显变化. ...
氮沉降对植物生长的影响研究进展
1
2017
... 在干旱半干旱地区,植物生长不仅受水分限制,较低的氮含量也成为主要限制因子[50].目前,有关氮添加对植物群落生产力影响的研究报道有很多[31-32,51-52],研究结果也不尽相同.本研究中氮添加显著增加了群落盖度和地上生物量,密度也有一定程度的增加,说明短暂的氮添加在一定时期内减缓了该地区氮素对植物生长的限制,促进植物快速生长,增加生物量的累积,与前人研究结果基本一致[34,51].但氮添加也加剧了植物种间或种内竞争,导致一些竞争能力较弱的个体减少甚至死亡,这解释了本研究中群落密度虽有增加但不显著的原因.此外,本研究发现氮添加增加了群落凋落物量,这可能是由于过量的氮添加改变了植物养分再吸收的过程[53],使得器官脱落前植物个体减弱了对养分的再吸收能力[29],同时由于竞争造成个体死亡的增加也为凋落物量的增加做出了一定贡献.植物高度反映植物对光资源竞争能力的强弱[41].研究发现,植物叶片氮含量的高低会影响植物光合作用[54],氮添加可以短暂地消除氮素对沙地生态系统的限制,增加植物可利用氮素水平,使得植物对地下养分的竞争转化为对光的竞争[34],但过量的氮素会打破植物体内的养分平衡,降低植物光合速率[55].本研究中氮添加显著增加了植物群落高度,进一步验证了氮添加可以提高沙地植物群落的光合能力.植物比叶面积越高、叶干物质含量越低,其光捕获能力越强,这些变化有利于光竞争力的提高[48].本研究中氮添加使得群落叶面积显著增加,叶干物质含量显著降低,说明沙地植物群落主要通过叶面积和叶干物质含量的变化来响应氮添加.比叶面积在响应短期氮添加时敏感性较弱,且不同物种对氮添加的响应不同,而群落水平上很容易掩盖这种种间差异,因此比叶面积在群落水平上响应氮添加时没有明显变化. ...
骆驼刺幼苗生长和功能性状对不同水氮添加的响应
1
2016
... 水氮之间对植物生长存在相互补偿机制,主要表现为氮添加能缓解干旱对植物造成的影响,但同时干旱可以减弱甚至抑制氮对植物生长的促进作用[56].增雨(W+60%和W+100%)处理下,氮添加使得群落盖度显著增加,群落密度、地上生物量和凋落物量在增雨60%和氮添加交互处理下达到最大值,而增雨100%与氮添加耦合反而使其降低,说明适当增雨更有利于氮素肥力的激发,降雨过多反而会缩短氮素滞留时间,造成氮素流失[15].同一干旱处理(W-60%与W-100%)下,氮添加增加了群落高度和叶面积,尤其减雨60%处理下氮添加显著提高了群落高度,说明氮添加可以在一定程度上缓解干旱对植物造成的影响.同样地,同一增雨处理(W+60%和W+100%)下氮添加也使得高度和叶面积有所增加,这可能是由于增雨更好地激发了氮肥效力,提高了土壤中氮素的可利用性,进而促进植物对氮素的吸收和利用效率.此外,与增雨100%相比,叶面积在增雨60%和氮添加交互处理下最大,说明并不是降雨越多越有利于植物生长,超过一定范围后反而会抑制植物生长.就叶干物质含量而言,在相同降雨处理下,氮添加降低了植物群落叶干物质含量,尤其在W-60%和W0处理下叶干物质含量显著降低,而比叶面积在水氮交互处理下虽有波动变化,但无明显变化规律,这可能是由于不同物种在面对气候变化时所采取的策略不同[52]. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}