祁连山北麓不同海拔土壤化学计量特征
高海宁 , 1 , 2 , 李彩霞 2 , 孙小妹 1 , 张勇 2 , 陈年来 , 1
1.甘肃农业大学 资源与环境学院,甘肃 兰州 730070
2.河西学院 甘肃省河西走廊特色资源利用省级重点实验室,甘肃 张掖 734000
Stoichiometry characteristics of soil at different altitudes in the Qilian Mountains
Gao Haining , 1 , 2 , Li Caixia 2 , Sun Xiaomei 1 , Zhang Yong 2 , Chen Nianlai , 1
1.College of Resource and Environmental Sciences,Gansu Agricultural University,Lanzhou 730070,China
2.Key Laboratory of Hexi Corridor Resources Utilization of Gansu,Hexi University,Zhangye 734000,Gansu,China
通讯作者: 陈年来(E-mail: chennl@163.com )
收稿日期: 2020-09-08
修回日期: 2020-11-24
网络出版日期: 2021-01-29
基金资助:
甘肃省重点研发计划项目 . 1504FKCA003
Received: 2020-09-08
Revised: 2020-11-24
Online: 2021-01-29
摘要
对祁连山中段冰沟流域土壤有机C、N、P含量垂直分布与化学计量特征及其影响因素进行研究。结果表明:冰沟流域土壤有机C、N、P含量在各样点随土层深度而降低,并在表层土中聚集较明显;在0—40 m土层中,土壤有机C、N、P平均含量随海拔升高总体呈现先增高后下降的趋势,在中、高海拔处最高,P含量变异较小。0—40 cm土层中土壤C/N在海拔3 128—2 814 m处高于其他海拔,在3 075 m处C/N最高,土壤C/N、C/P随海拔升高总体呈下降趋势,均在中高海拔处最高,而土壤N/P在海拔3 454 m处最高,在低海拔2 814 m处最低。不同植被类型土壤的C/N、N/P差异性不显著,而土壤SOC/P差异显著,3种植被类型影响土壤C/N和C/P的大小为乔木林>高山灌丛>高山草地,而土壤N/P为灌丛>乔木林>高山草地;植被类型、海拔是影响土壤C、N、P含量和化学计量比的主要因素,其次为盖度、坡向。
关键词:
祁连山 海拔 土壤 化学计量比
Abstract
To elucidate the vertical distribution of soil organic carbon (C), nitrogen (N) and phosphorus (P) concentration in Qilian Mountain northern foot,the eco-stoichiometric characteristics at different elevations and their influencing factors were discussed. The results showed that the soil SOC, N and P content decreased with the increase of soil depth, and the significant aggregation was observed at topsoil; In the 0-40 cm soil layer, the average contents of soil SOC, N and P showed a trend of rising first and then declining with the increase of altitude, and reached the highest at the middle and high altitude area. Notably, the variation of P content is small and relatively stable. The C/N was higher at attitude of 3 128-2 814 m, and especially, the highest at 3 075 m. However, soil C/P and N/P showed a downward trend with the increase of altitude, and they were all the highest at the middle and high altitude area, while soil N/P showed the highest level at 3 454 m and the lowest level at 2 814 m. Meanwhile, there was no significant difference in soil C/N and N/P among different land types but significant difference in C/P. The effects of the three vegetation types on soil C/N and C/P were: arbor forest > alpine shrub > alpine grassland, while N/P was: alpine shrub > arbor forest > alpine grassland. Vegetation type and altitude are the main factors affecting the eco-stoichiometric ratio of soil C, N and P, followed by coverage and slope aspect.
Keywords:
Qilian Mountains altitude soil stoichiometry ratio
本文引用格式
高海宁, 李彩霞, 孙小妹, 张勇, 陈年来. 祁连山北麓不同海拔土壤化学计量特征 . 中国沙漠 [J], 2021, 41(1): 219-227 doi:10.7522/j.issn.1000-694X.2020.00125
Gao Haining, Li Caixia, Sun Xiaomei, Zhang Yong, Chen Nianlai. Stoichiometry characteristics of soil at different altitudes in the Qilian Mountains . Journal of Desert Research
0 引言
土壤是陆地生态系统养分循环的主要载体,是植物生长发育所需养分的主要来源[1 ] ,气候条件,水分状况、凋落物分解都会对土壤养分产生显著影响[2 ] 。土壤有机碳(SOC)直接影响土壤质量[3 ] ,土壤中的N、P是植物生长发育的必须元素,SOC含量与分布特征[4 ] 对区域土壤C、N、P分布具有显著影响[3 ] 。土壤C、N和P的空间分布特征在一定程度上影响着其化学计量比的空间分布特征[5 ] ,从而影响生态系统的群落组成与结构[6 ] 。利用土壤C/N可预测森林土壤硝态氮淋溶程度,土壤N/P可反映植被受干扰程度,土壤元素的含量与分布状况对植物的生长发育有着巨大的影响[7 ] 。
由于生态系统垂直带性的分布差异,不同海拔条件下植被和土壤化学计量特征可以反映植被类型和气候对土壤养分的影响[8 ] ,反映土壤容重和土壤C储量[9 ] ,因此土壤生态化学计量特征的空间差异与变化规律引起许多学者的关注。孙小东等[10 ] 研究中国北方典型风沙区土壤C、N、P化学计量特征随经纬度的分布格局,发现草地和农田土壤有机C、全N、全P化学计量特征随纬度的增加呈逐渐增加趋势,除草地全P含量和N/P外,其余指标均与纬度呈线性关系,随经度的增加呈先减小后增加的趋势,与经度呈二阶多项式分布关系。牛赟等[11 ] 研究结果表明土壤C、N、P化学计量特征与海拔和土壤深度密切相关。由于研究的地理位置、海拔范围及植被状况的不同,海拔梯度土壤生态化学计量特征尚存争议。
祁连山北麓,降水多、蒸发少、气温低、高寒阴湿,是河西走廊生态文明建设和社会经济发展的基础。由于区域地形复杂,水热梯度明显,植被的垂直带性分布影响着生物多样性的分布[12 ] ;因此研究祁连山北麓植被垂直带性分布下的土壤C、N、P化学计量学特征,以阐明祁连山北麓土壤C、N、P循环机制和土壤养分的变化规律势在必行。
本文通过研究祁连山北麓冰沟流域不同海拔(2 814—4 130 m)的土壤C、N、P含量及化学计量比的垂直地带性分布特征,分析海拔等因素对土壤养分含量及化学计量比特征的影响,为祁连山区土壤养分供应状况对植被群落结构和生态功能的影响提供依据。
1 材料与方法
1.1 研究样地
研究区位于祁连县冰沟流域(38°02′18″—38°10′39″N,100°11′24″—100°23′55″E),海拔2 760—4 401 m,年均气温0.5 ℃,年降水量420 mm,平均相对湿度60%,成土岩为石灰岩、砂岩、砾岩,成土母质为残积母质和坡积母质。研究区土壤类型为高山草甸土、高山寒漠土、高山灌丛草甸土、亚高山灌丛草甸土、森林灰褐土。乔木主要有青海云杉(Picea crassifolia )、祁连圆柏(Sabina przewalskii )、山杨(Populus davidiana )等;灌木主要为小叶金露梅(Potentilla parvifolia )、鬼箭锦鸡儿(Caragana jubata )、肋果沙棘(Hippophae neurocarpa )、银露梅(Potentilla grabra )、杯腺柳(Salix cupularis )等。草本有珠芽蓼(Polygonum viviparum )、黄花棘豆(Oxytropis ochrocephala )、线叶嵩草(Kobresia capillifolia )、草地早熟禾(Poa pratensis )等,样地基本概况见表1 。
1.2 研究方法
1.2.1 土壤样品采集
2017年7月,在祁连山北麓冰沟流域,根据植被类型和群落组成,从流域坡底到坡顶(2 814—4 130 m)共设置19个采样点,每个采样点除去表层凋落物层后在剖面分3层(0—10、10—20、20—40 cm)采集土样,每个采样点设置3个重复,共采集171个土壤样品,按样点、海拔和土层深度编号后装入自封袋带回实验室,并捡出每个样品中的石块、植物残体及细根,在室内自然风干后,过0.149 mm筛,用于测定土壤有机C、全N、全P含量。同时用100 cm3 环刀分层取原状土测定土壤容重,用手持GPS仪记录采样点的经纬度和高程。
1.2.2 土壤理化性质测定
土壤有机C、全N、全P、土壤容重(pb ,环刀法)的测定参照《土壤分析技术规范》[13 ] ,土壤C、N、P储量采用下式计算[14 ] 。
S T = ∑ n = 1 n C i × p b × d i 100 (1)
式中:ST 为储量(kg∙m-2 );i 为土层深度(m);Ci 为第i 层土壤有机C(N或P)含量(g∙kg-1 );pb 为第i 层土壤容重(g∙cm-3 );di 为第i 层土壤厚度(cm)。
1.3 数据分析
采用Excel 2010和SPSS 22软件对数据进行处理和统计分析。利用Origin 8.0软件作图,运用单因素方差法(One-way ANOVA)分析不同海拔和深度土壤C、N、P含量及储量化学计量比差异显著性,用最小显著差数法进行多重比较;运用Pearson相关分析对植被类型和土壤C、N、P含量及储量化学计量比之间进行了相关性对比。
2 结果与分析
2.1 土壤C 、N 、P 含量垂直变化特征
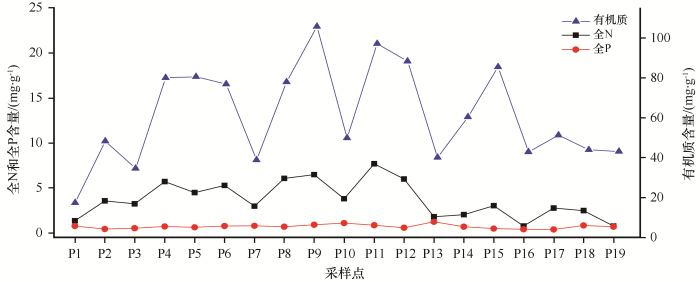
土壤C、N、P含量在各样点变化如表2 所示,各样点随土层深度C、N、P含量均呈下降趋势,0—10 cm土层土壤C、N含量与10—20 cm和20—40 cm土层差异显著(P <0.05),而P含量在0—10 cm土层土壤中较高,但在不同海拔处各层之间差异不显著。0—10 cm土层土壤C含量16.85—163.85 mg∙g-1 ,在海拔3 086、3 379、3 250 m处土壤C含量最高,分别为163.85、145.49、145.81 mg∙g-1 ;N含量11.68—1.37 mg∙g-1 ,在海拔3 250、3 379、3 533 m处最高,分别为11.68、9.10、7.25 mg∙g-1 ;P含量在海拔3 128 m,土层10—20 cm处最高,为1.79 mg∙g-1 。整体而言,土壤C、N和P在表层的聚集现象较为明显,C、N含量随土层变化趋势相近,P含量变化不明显。
由图1 可见,随海拔升高,各样点间0—40 cm深度的土壤C含量变幅较大,为17.37—105.76 mg∙g-1 ,平均值为61.72 mg∙g-1 ,变异系数为39.19%,其中海拔3 379 m和3 250 m与其他海拔差异显著,在海拔3 379 m时最高,其次为3 250 m,海拔4 130 m最低。N含量在3 250 m海拔与其他海拔差异显著(P <0.05),变幅0.77—7.69 mg∙g-1 ,平均3.79 mg∙g-1 ,变异系数53.02%,而P含量变幅0.39—1.25 mg∙g-1 ,平均0.72 mg∙g-1 ,变异系数30.65%,土壤P含量相比C和N在空间变异较小。
图1
图1
土壤C、N、P 含量随海拔梯度变化(0—40 cm)
Fig.1
Soil C, N and P concentration to the altitudes (0-40 cm)
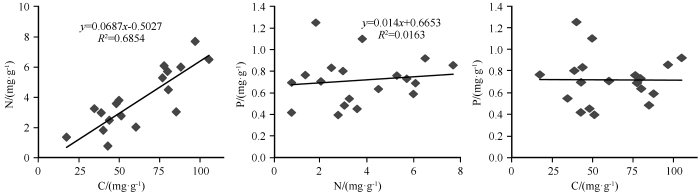
土壤C含量和N含量之间显著相关,决定系数R 2 为0.6854(P <0.05),N和P、C和P之间无相关性(图2 )。综上所述,流域土壤C、N含量随土层变化的尺度基本同步,在中海拔3 379 m和3 250 m最高,而P含量变化滞后于C、N含量。
图2
图2
不同海拔土壤C、N、P相关性
Fig.2
Correlation of soil organic C, N and P at different altitudes
2.2 土壤C 、N 、P 化学计量比垂直变化特征
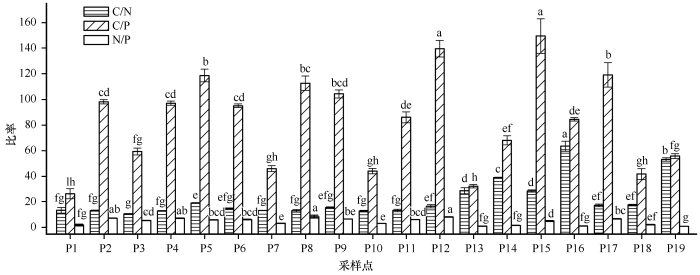
不同海拔土壤C、N、P化学计量比特征结果见图3 ,随着海拔升高C/N呈降低趋势,幅度为10.63—63.38,平均值21.96,变异系数65.31%,C/N在海拔2 814—3 128 m内较高,在各样点(除海拔3 750 m)C/N均高于国内土壤的平均值12.10[15 -18 ] ;C/P在高海拔4 130 m处最低,3 086 m处最高,变幅26.47—149.24,平均82.94,变异系数为42.94%;N/P变幅1.05—8.59,平均4.82,变异系数52.95%。不同海拔C/N、C/P随海拔整体呈现降低趋势,而N/P变化规律不同,在最高和低海拔处最低。
图3
图3
不同海拔土壤C/N、C/P、N/P特征(0—40 cm)
不同小写字母表示样点间差异显著,P <0.05
Fig.3
Characteristics of soil C/N, C/P, N/P at different altitudes (0-40 cm)
2.3 不同植被土壤C 、N 、P 化学计量比特征
不同植被土壤C、N、P化学计量比特征存在一定的差异(表3 )。高山草地、高山灌丛、乔木林随海拔降低土壤C/N增高,在海拔3 128、3 075、2 184 m处C/N分别为28.71、63.38、53.14,而高山草地土壤C/P和N/P呈现先增高后降低的趋势,3种植被类型土壤C/N平均值差异不显著(P >0.05),C/N排序为乔木林>灌丛>高山草地;C/P乔木林和高山灌丛相比较差异不显著(P >0.05),而与高山草地C/P间差异显著(P <0.05),其变化规律与C/N相同;3种植被类型土壤N/P差异不显著(P >0.05),N/P排序为高山灌丛>乔木林>高山草地。由此可见植被的垂直带性分布对土壤C、N、P化学计量比具有显著的影响。
2.4 土壤化学计量比与土壤养分含量、环境因子间的相关性
0—40 cm土壤C含量与C/P、N/P间均呈极显著正相关(P <0.01),与C/N无相关性(表4 );N含量与C/N、C/P、N/P呈极显著相关(P <0.01),其中与C/P、N/P呈正相关,而与C/N呈负相关,P与C/P呈现极显著负相关,与C/N、N/P无显著相关性。盖度与C/N、C/P、N/P之间相关不显著,植被类型与C/N呈极显著正相关(P <0.01),而与C/P呈显著正相关(P <0.05);海拔、坡向与C/N呈极显著正相关(P <0.01),海拔与N/P呈显著负相关(P <0.05),而坡向与N/P呈极显著负相关(P <0.01),从结果分析,植被类型、海拔、坡向对土壤C、N、P化学计量比特征影响差异较大,其中对C/N影响最大,其次为N/P,C/P。从结果来看,土壤C、N、P化学计量特征受多种因素的影响,几种环境因子与C/P相关性较差,说明流域C/P不随环境变化而变化,相对比较稳定。
3 讨论
3.1 土壤C 、N 、P 的垂直变异特征
土壤养分是森林生态系统中植物营养的主要来源,森林群落结构、物种组成及生产力均受土壤养分的影响[19 ] 。土壤养分是海拔梯度主导下地形、气候以及生物因素相互作用的结果,具有显著的空间异质特征[20 ] 。本研究发现,在海拔2 814—4 130 m,乔木林土壤C、N、P含量随土层深度的增加而降低,表层土壤显著高于下层,而P含量相较C、TN含量在土壤剖面上变化不大。在0—40 cm的土层C、N含量的空间分布基本一致,而P含量的变化滞后于C、N含量,C、N和P含量的平均值分别为61.72、3.79 mg∙g-1 ,均高于中国0—10 cm土壤有机C、全N含量的均值24.56、1.88 mg∙g-1 ,而P含量0.72 mg∙g-1 稍低于中国0—10 cm土壤全P含量均值0.78 mg∙g-1[21 ] ,说明该流域土壤C含量高于N,而P含量相对匮乏。0—40 cm土层C、N含量的平均值空间变异系数较大,而P空间变异系数较小,这一结果与吕世丽等[22 ] 的研究结果相似。因为表层土壤受外界环境因素及植被凋落物养分的影响,导致土壤养分在表层密集[22 ] ,土壤C、N除受土壤母质的影响外,还受植物凋落物分解以及植物的吸收利用等影响,而存在着较大的空间变异性[23 ] ;而土壤全P含量主要受成土母质的影响,动植物残体、微生物活动等对全P的影响相对较小[1 ] ,因此,P含量在土层间存在较小的空间变异性,且相对稳定。
海拔的变化将会导致气候特征、植被类型与土壤类型等发生相应改变,从而影响土壤养分的差异[24 ] 。本研究表明,0—40 cm土壤有机C、N平均含量除海拔3 365 m和3 516 m外,在3 204—3 732 m显著高于海拔3 075 m以下和海拔3 750 m以上土壤的C、N含量,而在中高海拔3 379 m和3 250 m土壤C、N平均含量最高,这一结论与Cleveland等[25 ] 的研究结果一致。因为土壤有机质含量与土壤温度和水分有关,适宜的温度有利于有机质的积累,温度过高或过低对土壤有机质的积累均有负效应[8 ,25 ] ;其次,由于各海拔植被类型的不同使得土壤有机C的积累差异较大,N的变化与C的变化基本同步。P含量随海拔升高呈先升高后降低的趋势,在中海拔3 128 m处土壤P含量显著高于其他海拔(P <0.05),可能与该海拔范围内成土母质的分化有关。
3.2 土壤C 、N 、P 化学计量比垂直变化特征
土壤C、N、P生态化学计量比是土壤有机C、N、P各元素总含量的比值,是评价土壤养分限制以及C、N、P饱和诊断和有效的预测性指标[26 ] 。由于地形、气候、土壤质地、植被、动物及土壤微生物等成土因素和人类活动的影响,不同海拔的土壤C、N、P含量有较大的差异,从而使土壤C/N、C/P和N/P存在较大的空间变异性[27 ] 。本研究区域不同海拔0—40 cm土壤C/N差异显著(P <0.05),随海拔升高总体呈下降趋势,在海拔3 128—2 814 m处高于其他海拔,在海拔3 075 m处最高(63.38),研究区土壤C/N的平均值21.96高于全球土壤C/N平均值13.33[2 ] ,表明土壤有机质的矿质化和腐殖质化明显,土壤N含量低。
C/P反映土壤微生物矿化有机物释放P的能力,值越低表明土壤P有效性越高,研究区不同海拔C/P差异显著(P <0.05),随海拔增高总体呈下降趋势,在3 086 m处最高(149.24),在最高海拔4 130 m处最低(26.47)。研究区C/P的平均值82.94稍高于全球森林0—10 cm土壤C/P均值81.90[28] ,表明祁连山北麓土壤有机质矿化释放P的能力弱,土壤P有效性低。
N/P可用作N饱和的诊断指标,也可反映土壤N、P平衡状况,被用于确定养分限制的阈值,能指示植物生长过程中土壤营养成分的供应情况[29 ] 。研究区不同海拔N/P差异显著(P <0.05),在海拔3 204—3 894 m范围内土壤N/P较高,且在3 454 m处最高(8.59),在2 814 m处最低(1.05)。研究区土壤N/P的平均值为4.82,高于中国森林生态系统土壤N/P (3.20)[28] ,说明研究区域土壤P相对缺乏。研究区海拔高、气温低、降雨量大,土壤P元素淋溶强、风化弱,不易累积,土壤P缺乏可能是C/P、N/P高于全球土壤和中国森林生态系统土壤的主要原因。
3.3 植被类型与土壤C/N 、C/P 、N/P 化学计量比的关系
植被类型是影响土壤养分分布的重要因素,不同植被类型的土壤C/N、C/P、N/P存在显著差异[27 ] 。本研究3种植被类型的土壤C/N差异不显著,说明祁连山北麓不同植被类型土壤有机质分解速度基本相近,3种植被类型土壤C/N平均值乔木林28.56>灌丛20.10>高山草地16.54。本研究区乔木林以青海云杉为主,其次为祁连圆柏和山杨,由于其凋落物丰富,凋落物分解速率较快,有效提高了土壤有机C含量,使其C/N提高,而灌丛和高山草地主要为小叶金露梅、银露梅、鬼箭锦鸡儿等,相对于乔木林,凋落物相对少,土壤C/N较低[28] 。乔木林土壤C/P显著高于灌木和草地,变化趋势与C/N相同,即为乔木林105.89>灌丛80.51>高山草地54.00,说明高山草地土壤P的有效性高;土壤N/P可以作为养分限制类型的有效预测指标,祁连山北麓不同植被类型土壤N/P差异不显著,变化趋势为灌丛4.99>乔木林5.06>高山草地4.02,均小于10,说明祁连山北麓不同植被受土壤N限制。
3.4 土壤化学计量比与土壤养分含量、环境因子间的相关性
土壤C与N含量呈极显著正相关,C、N含量与C/P、N/P呈极显著正相关,而N含量与C/N呈极显著负相关,P含量与C/P呈极显著负相关,说明该流域0—40 cm土层土壤养分C、N、P在空间分布上相互耦合[25 ] 。
不同样点土壤C、N、P含量及其化学计量比的差异与海拔、植被类型、盖度和坡向有直接的关系,尤其以海拔和植被类型影响最为显著[29 -30 ] 。研究表明,植被类型与C含量、C/P、N/P呈正相关,因为植被类型不同,每年归还到土壤的凋落物量也不同,且不同植被凋落物C、N、P含量的差异导致土壤C/N、C/P、N/P存在显著的差异。海拔与C/N呈正相关而与N/P、N呈负相关,说明N越低,C/N越高,表明流域整体土壤的C/N和N/P主要受N含量的影响。C/N与植被盖度呈正相关(0.309),但不显著,也说明地上植物是土壤中C素和N素的主要来源。坡向与土壤C/N呈正相关,而与土壤N/P呈负相关。以上分析表明,该区域植被类型、海拔是影响土壤C、N、P含量及其化学计量特征的主要因素。
4 结论
祁连山北麓土壤不同土层有机C、N、P含量随土层深度而降低,主要分布在表层土中;0—40 cm土壤有机C、N含量的平均值均高于全国土壤平均值,而P稍低于全国土壤均值;C、N、P随海拔升高总体呈现先增高后下降的趋势,在中、高海拔处最高;不同海拔梯度土壤C/N,C/P与N/P均值高于全球森林0—10 cm土壤C/P与N/P均值,C/N在海拔3 128—2 814 m处高于其他海拔,在3 075 m处C/N最高,C/P与C/N随海拔总体呈下降趋势。不同植被类型土壤的C/N、N/P差异性不显著,而C/P差异显著,3种植被类型影响土壤C/N和C/P的大小为乔木林>灌丛>高山草地,而N/P为灌丛>乔木林>高山草地,且均小于10,说明祁连山北麓植被受土壤N限制;植被类型、海拔是影响土壤C、N、P化学计量比的主要因素、其次为盖度,坡向影响较小。
参考文献
View Option
[1]
刘超 ,王洋 ,王楠 ,等 陆地生态系统植被氮磷化学计量研究进展
[J].植物生态学报 ,2012 ,36 (11 ):1205 -1216 .
[本文引用: 2]
[2]
邬畏 ,何兴东 ,周启星 生态系统氮磷比化学计量特征研究进展
[J].中国沙漠 ,2010 ,30 (2 ):296 -302 .
[本文引用: 2]
[3]
Sollins P Gregg J W Soil organic matter accumulation in relation to changing soil volume,mass,and structure:concepts and calculations
[J].Geoderma ,2017 ,301 :60 -71 .
[本文引用: 2]
[4]
朱猛 ,刘蔚 ,秦燕燕 ,等 祁连山森林草原带坡面尺度土壤有机碳分布
[J].中国沙漠 ,2016 ,36 (3 ):741 -748 .
[本文引用: 1]
[5]
李新星 ,刘桂民 ,吴小丽 ,等 马衔山不同海拔土壤碳、氮、磷含量及生态化学计量特征
[J].生态学杂志 ,2020 ,39 (3 ):758 -765 .
[本文引用: 1]
[6]
朱湾湾 ,王攀 ,樊瑾 ,等 降水量及N添加对宁夏荒漠草原土壤C∶N∶P生态化学计量特征和植被群落组成的影响
[J].草业学报 ,2019 ,28 (9 ):33 -44 .
[本文引用: 1]
[7]
Fanelli G Lestini M Saulia S Floristic gradients of herbaceous vegetation and P/N ratio in soil in a Mediterranean area
[J]. Plant Ecology ,2007 ,194 (2 ): 231 -242 .
[本文引用: 1]
[8]
Zhang M Zhang X K Liang W J et al Distribution of soil organic carbon fractions along the altitudinal gradient in Changbai Mountain,China
[J].Pedosphere ,2011 ,21 :615 -620 .
[本文引用: 2]
[9]
张梦旭 ,刘蔚 ,朱猛 ,等 黑河上游山地土壤容重分布特征及影响因素
[J].山地学报 ,2019 ,37 (2 ):198 -205 .
[本文引用: 1]
[10]
孙小东 ,宁志英 ,杨红玲 ,等 中国北方典型风沙区土壤碳氮磷化学计量特征
[J].中国沙漠 ,2018 ,38 (6 ):1209 -1218 .
[本文引用: 1]
[11]
牛赟 ,刘贤德 ,赵维俊 ,等 祁连山青海云杉(Picea crassifolia )林浅层土壤碳、氮含量特征及其相互关系
[J].中国沙漠 ,2014 ,34 (2 ):371 -377 .
[本文引用: 1]
[12]
何远政 ,黄文达 ,赵昕 ,等 气候变化对植物多样性的影响研究综述
[J].中国沙漠 ,2021 ,41 (1 ):59 -66 .
[本文引用: 1]
[13]
全国农业技术推广服务中心 土壤分析技术规范 [M].北京 :中国农业出版社 ,2009 .
[本文引用: 1]
[14]
徐芷君 ,刘苑秋 ,方向民 ,等 亚热带2种针叶林土壤碳氮磷储量及化学计量比对混交的响应
[J].水土保持学报 ,2019 ,33 (1 ):165 -170 .
[本文引用: 1]
[16]
张继平 ,张林波 ,王风玉 ,等 井冈山国家级自然保护区森林土壤养分含量的空间变化
[J].土壤 ,2014 ,46 (2 ):262 -268 .
[17]
Wang Y H Zhang L Haimiti Y et al Study on spatial variability of soil nutrients in Ebinur Lake wetlands in China
[J].Journal of Coastal Research ,2015 ,73 (1 ):59 -63 .
[18]
Tian H Q Chen G S Zhang C et al Pattern and variation of C∶N∶P ratios in China’s soils:asynthesis of observational data
[J].Biogeochemistry ,2010 ,98 (3 ):139 -151 .
[本文引用: 1]
[19]
张广帅 ,邓浩俊 ,杜锟 ,等 泥石流频发区山地不同海拔土壤化学计量特征:以云南省小江流域为例
[J].生态学报 ,2016 ,36 (3 ): 675 -687 .
[本文引用: 1]
[20]
朱秋莲 ,邢肖毅 ,张宏 ,等 黄土丘陵沟壑区不同植被区土壤生态化学计量特征
[J].生态学报 ,2013 ,33 (15 ):4674 -4682 .
[本文引用: 1]
[21]
张鹏 ,张涛 ,陈年来 祁连山北麓山体垂直带土壤碳氮分布特征及影响因素
[J].应用生态学报 ,2009 ,20 (3 ):518 -524 .
[本文引用: 1]
[22]
吕世丽 ,李新平 ,李文斌 ,等 牛背梁自然保护区不同海拔高度森林土壤养分特征分析
[J].西北农林科技大学学报(自然科学版) ,2013 ,41 (4 ):161 -177 .
[本文引用: 2]
[23]
Hobbie S E Gough L Foliar and soil nutrients in tundra onglacial landscapes of contrasting ages in northern Alaska
[J].Oecologia ,2016 ,131 (3 ):453 -462 .
[本文引用: 1]
[24]
庞圣江 ,张培 ,贾宏炎 ,等 桂西北不同森林类型土壤生态化学计量特征
[J].中国农学通报 ,2015 ,31 (1 ):17 -23 .
[本文引用: 1]
[25]
Cleveland C C Liptzin D C∶N∶P stoichiometry in soil:is there a“redfield ratio”for the microbial biomass
[J].Biogeochemistry ,2007 ,85 (3 ):235 -252 .
[本文引用: 3]
[26]
Tessier J T Raynal D J Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation
[J].Journal of Applied Ecology ,2003 ,40 (3 ): 523 -534 .
[本文引用: 1]
[27]
Bing H J Wu Y H Zhou J et al Stoichiometric variation of carbon,nitrogen,and phosphorus in soils and its implication for nutrient limitation in alpine ecosystem of Eastern Tibetan Plateau
[J].Journal of Soils & Sediments ,2016 ,16 (2 ):405 -416 .
[本文引用: 2]
[29]
刘秉儒 ,张秀珍 ,胡天华 ,等 贺兰山不同海拔典型植被带土壤微生物多样性
[J].生态学报 ,2013 ,33 (22 ):7211 -7220 .
[本文引用: 2]
[30]
张光德 ,赵传燕 ,戎战磊 ,等 祁连山中部不同植被类型土壤生态化学计量特征研究
[J].兰州大学学报(自然科学版) ,2019 ,55 (4 ):533 -540 .
[本文引用: 1]
陆地生态系统植被氮磷化学计量研究进展
2
2012
... 土壤是陆地生态系统养分循环的主要载体,是植物生长发育所需养分的主要来源[1 ] ,气候条件,水分状况、凋落物分解都会对土壤养分产生显著影响[2 ] .土壤有机碳(SOC)直接影响土壤质量[3 ] ,土壤中的N、P是植物生长发育的必须元素,SOC含量与分布特征[4 ] 对区域土壤C、N、P分布具有显著影响[3 ] .土壤C、N和P的空间分布特征在一定程度上影响着其化学计量比的空间分布特征[5 ] ,从而影响生态系统的群落组成与结构[6 ] .利用土壤C/N可预测森林土壤硝态氮淋溶程度,土壤N/P可反映植被受干扰程度,土壤元素的含量与分布状况对植物的生长发育有着巨大的影响[7 ] . ...
... 土壤养分是森林生态系统中植物营养的主要来源,森林群落结构、物种组成及生产力均受土壤养分的影响[19 ] .土壤养分是海拔梯度主导下地形、气候以及生物因素相互作用的结果,具有显著的空间异质特征[20 ] .本研究发现,在海拔2 814—4 130 m,乔木林土壤C、N、P含量随土层深度的增加而降低,表层土壤显著高于下层,而P含量相较C、TN含量在土壤剖面上变化不大.在0—40 cm的土层C、N含量的空间分布基本一致,而P含量的变化滞后于C、N含量,C、N和P含量的平均值分别为61.72、3.79 mg∙g-1 ,均高于中国0—10 cm土壤有机C、全N含量的均值24.56、1.88 mg∙g-1 ,而P含量0.72 mg∙g-1 稍低于中国0—10 cm土壤全P含量均值0.78 mg∙g-1[21 ] ,说明该流域土壤C含量高于N,而P含量相对匮乏.0—40 cm土层C、N含量的平均值空间变异系数较大,而P空间变异系数较小,这一结果与吕世丽等[22 ] 的研究结果相似.因为表层土壤受外界环境因素及植被凋落物养分的影响,导致土壤养分在表层密集[22 ] ,土壤C、N除受土壤母质的影响外,还受植物凋落物分解以及植物的吸收利用等影响,而存在着较大的空间变异性[23 ] ;而土壤全P含量主要受成土母质的影响,动植物残体、微生物活动等对全P的影响相对较小[1 ] ,因此,P含量在土层间存在较小的空间变异性,且相对稳定. ...
生态系统氮磷比化学计量特征研究进展
2
2010
... 土壤是陆地生态系统养分循环的主要载体,是植物生长发育所需养分的主要来源[1 ] ,气候条件,水分状况、凋落物分解都会对土壤养分产生显著影响[2 ] .土壤有机碳(SOC)直接影响土壤质量[3 ] ,土壤中的N、P是植物生长发育的必须元素,SOC含量与分布特征[4 ] 对区域土壤C、N、P分布具有显著影响[3 ] .土壤C、N和P的空间分布特征在一定程度上影响着其化学计量比的空间分布特征[5 ] ,从而影响生态系统的群落组成与结构[6 ] .利用土壤C/N可预测森林土壤硝态氮淋溶程度,土壤N/P可反映植被受干扰程度,土壤元素的含量与分布状况对植物的生长发育有着巨大的影响[7 ] . ...
... 土壤C、N、P生态化学计量比是土壤有机C、N、P各元素总含量的比值,是评价土壤养分限制以及C、N、P饱和诊断和有效的预测性指标[26 ] .由于地形、气候、土壤质地、植被、动物及土壤微生物等成土因素和人类活动的影响,不同海拔的土壤C、N、P含量有较大的差异,从而使土壤C/N、C/P和N/P存在较大的空间变异性[27 ] .本研究区域不同海拔0—40 cm土壤C/N差异显著(P <0.05),随海拔升高总体呈下降趋势,在海拔3 128—2 814 m处高于其他海拔,在海拔3 075 m处最高(63.38),研究区土壤C/N的平均值21.96高于全球土壤C/N平均值13.33[2 ] ,表明土壤有机质的矿质化和腐殖质化明显,土壤N含量低. ...
Soil organic matter accumulation in relation to changing soil volume,mass,and structure:concepts and calculations
2
2017
... 土壤是陆地生态系统养分循环的主要载体,是植物生长发育所需养分的主要来源[1 ] ,气候条件,水分状况、凋落物分解都会对土壤养分产生显著影响[2 ] .土壤有机碳(SOC)直接影响土壤质量[3 ] ,土壤中的N、P是植物生长发育的必须元素,SOC含量与分布特征[4 ] 对区域土壤C、N、P分布具有显著影响[3 ] .土壤C、N和P的空间分布特征在一定程度上影响着其化学计量比的空间分布特征[5 ] ,从而影响生态系统的群落组成与结构[6 ] .利用土壤C/N可预测森林土壤硝态氮淋溶程度,土壤N/P可反映植被受干扰程度,土壤元素的含量与分布状况对植物的生长发育有着巨大的影响[7 ] . ...
... [3 ].土壤C、N和P的空间分布特征在一定程度上影响着其化学计量比的空间分布特征[5 ] ,从而影响生态系统的群落组成与结构[6 ] .利用土壤C/N可预测森林土壤硝态氮淋溶程度,土壤N/P可反映植被受干扰程度,土壤元素的含量与分布状况对植物的生长发育有着巨大的影响[7 ] . ...
祁连山森林草原带坡面尺度土壤有机碳分布
1
2016
... 土壤是陆地生态系统养分循环的主要载体,是植物生长发育所需养分的主要来源[1 ] ,气候条件,水分状况、凋落物分解都会对土壤养分产生显著影响[2 ] .土壤有机碳(SOC)直接影响土壤质量[3 ] ,土壤中的N、P是植物生长发育的必须元素,SOC含量与分布特征[4 ] 对区域土壤C、N、P分布具有显著影响[3 ] .土壤C、N和P的空间分布特征在一定程度上影响着其化学计量比的空间分布特征[5 ] ,从而影响生态系统的群落组成与结构[6 ] .利用土壤C/N可预测森林土壤硝态氮淋溶程度,土壤N/P可反映植被受干扰程度,土壤元素的含量与分布状况对植物的生长发育有着巨大的影响[7 ] . ...
马衔山不同海拔土壤碳、氮、磷含量及生态化学计量特征
1
2020
... 土壤是陆地生态系统养分循环的主要载体,是植物生长发育所需养分的主要来源[1 ] ,气候条件,水分状况、凋落物分解都会对土壤养分产生显著影响[2 ] .土壤有机碳(SOC)直接影响土壤质量[3 ] ,土壤中的N、P是植物生长发育的必须元素,SOC含量与分布特征[4 ] 对区域土壤C、N、P分布具有显著影响[3 ] .土壤C、N和P的空间分布特征在一定程度上影响着其化学计量比的空间分布特征[5 ] ,从而影响生态系统的群落组成与结构[6 ] .利用土壤C/N可预测森林土壤硝态氮淋溶程度,土壤N/P可反映植被受干扰程度,土壤元素的含量与分布状况对植物的生长发育有着巨大的影响[7 ] . ...
降水量及N添加对宁夏荒漠草原土壤C∶N∶P生态化学计量特征和植被群落组成的影响
1
2019
... 土壤是陆地生态系统养分循环的主要载体,是植物生长发育所需养分的主要来源[1 ] ,气候条件,水分状况、凋落物分解都会对土壤养分产生显著影响[2 ] .土壤有机碳(SOC)直接影响土壤质量[3 ] ,土壤中的N、P是植物生长发育的必须元素,SOC含量与分布特征[4 ] 对区域土壤C、N、P分布具有显著影响[3 ] .土壤C、N和P的空间分布特征在一定程度上影响着其化学计量比的空间分布特征[5 ] ,从而影响生态系统的群落组成与结构[6 ] .利用土壤C/N可预测森林土壤硝态氮淋溶程度,土壤N/P可反映植被受干扰程度,土壤元素的含量与分布状况对植物的生长发育有着巨大的影响[7 ] . ...
Floristic gradients of herbaceous vegetation and P/N ratio in soil in a Mediterranean area
1
2007
... 土壤是陆地生态系统养分循环的主要载体,是植物生长发育所需养分的主要来源[1 ] ,气候条件,水分状况、凋落物分解都会对土壤养分产生显著影响[2 ] .土壤有机碳(SOC)直接影响土壤质量[3 ] ,土壤中的N、P是植物生长发育的必须元素,SOC含量与分布特征[4 ] 对区域土壤C、N、P分布具有显著影响[3 ] .土壤C、N和P的空间分布特征在一定程度上影响着其化学计量比的空间分布特征[5 ] ,从而影响生态系统的群落组成与结构[6 ] .利用土壤C/N可预测森林土壤硝态氮淋溶程度,土壤N/P可反映植被受干扰程度,土壤元素的含量与分布状况对植物的生长发育有着巨大的影响[7 ] . ...
Distribution of soil organic carbon fractions along the altitudinal gradient in Changbai Mountain,China
2
2011
... 由于生态系统垂直带性的分布差异,不同海拔条件下植被和土壤化学计量特征可以反映植被类型和气候对土壤养分的影响[8 ] ,反映土壤容重和土壤C储量[9 ] ,因此土壤生态化学计量特征的空间差异与变化规律引起许多学者的关注.孙小东等[10 ] 研究中国北方典型风沙区土壤C、N、P化学计量特征随经纬度的分布格局,发现草地和农田土壤有机C、全N、全P化学计量特征随纬度的增加呈逐渐增加趋势,除草地全P含量和N/P外,其余指标均与纬度呈线性关系,随经度的增加呈先减小后增加的趋势,与经度呈二阶多项式分布关系.牛赟等[11 ] 研究结果表明土壤C、N、P化学计量特征与海拔和土壤深度密切相关.由于研究的地理位置、海拔范围及植被状况的不同,海拔梯度土壤生态化学计量特征尚存争议. ...
... 海拔的变化将会导致气候特征、植被类型与土壤类型等发生相应改变,从而影响土壤养分的差异[24 ] .本研究表明,0—40 cm土壤有机C、N平均含量除海拔3 365 m和3 516 m外,在3 204—3 732 m显著高于海拔3 075 m以下和海拔3 750 m以上土壤的C、N含量,而在中高海拔3 379 m和3 250 m土壤C、N平均含量最高,这一结论与Cleveland等[25 ] 的研究结果一致.因为土壤有机质含量与土壤温度和水分有关,适宜的温度有利于有机质的积累,温度过高或过低对土壤有机质的积累均有负效应[8 ,25 ] ;其次,由于各海拔植被类型的不同使得土壤有机C的积累差异较大,N的变化与C的变化基本同步.P含量随海拔升高呈先升高后降低的趋势,在中海拔3 128 m处土壤P含量显著高于其他海拔(P <0.05),可能与该海拔范围内成土母质的分化有关. ...
黑河上游山地土壤容重分布特征及影响因素
1
2019
... 由于生态系统垂直带性的分布差异,不同海拔条件下植被和土壤化学计量特征可以反映植被类型和气候对土壤养分的影响[8 ] ,反映土壤容重和土壤C储量[9 ] ,因此土壤生态化学计量特征的空间差异与变化规律引起许多学者的关注.孙小东等[10 ] 研究中国北方典型风沙区土壤C、N、P化学计量特征随经纬度的分布格局,发现草地和农田土壤有机C、全N、全P化学计量特征随纬度的增加呈逐渐增加趋势,除草地全P含量和N/P外,其余指标均与纬度呈线性关系,随经度的增加呈先减小后增加的趋势,与经度呈二阶多项式分布关系.牛赟等[11 ] 研究结果表明土壤C、N、P化学计量特征与海拔和土壤深度密切相关.由于研究的地理位置、海拔范围及植被状况的不同,海拔梯度土壤生态化学计量特征尚存争议. ...
中国北方典型风沙区土壤碳氮磷化学计量特征
1
2018
... 由于生态系统垂直带性的分布差异,不同海拔条件下植被和土壤化学计量特征可以反映植被类型和气候对土壤养分的影响[8 ] ,反映土壤容重和土壤C储量[9 ] ,因此土壤生态化学计量特征的空间差异与变化规律引起许多学者的关注.孙小东等[10 ] 研究中国北方典型风沙区土壤C、N、P化学计量特征随经纬度的分布格局,发现草地和农田土壤有机C、全N、全P化学计量特征随纬度的增加呈逐渐增加趋势,除草地全P含量和N/P外,其余指标均与纬度呈线性关系,随经度的增加呈先减小后增加的趋势,与经度呈二阶多项式分布关系.牛赟等[11 ] 研究结果表明土壤C、N、P化学计量特征与海拔和土壤深度密切相关.由于研究的地理位置、海拔范围及植被状况的不同,海拔梯度土壤生态化学计量特征尚存争议. ...
祁连山青海云杉(Picea crassifolia )林浅层土壤碳、氮含量特征及其相互关系
1
2014
... 由于生态系统垂直带性的分布差异,不同海拔条件下植被和土壤化学计量特征可以反映植被类型和气候对土壤养分的影响[8 ] ,反映土壤容重和土壤C储量[9 ] ,因此土壤生态化学计量特征的空间差异与变化规律引起许多学者的关注.孙小东等[10 ] 研究中国北方典型风沙区土壤C、N、P化学计量特征随经纬度的分布格局,发现草地和农田土壤有机C、全N、全P化学计量特征随纬度的增加呈逐渐增加趋势,除草地全P含量和N/P外,其余指标均与纬度呈线性关系,随经度的增加呈先减小后增加的趋势,与经度呈二阶多项式分布关系.牛赟等[11 ] 研究结果表明土壤C、N、P化学计量特征与海拔和土壤深度密切相关.由于研究的地理位置、海拔范围及植被状况的不同,海拔梯度土壤生态化学计量特征尚存争议. ...
气候变化对植物多样性的影响研究综述
1
2021
... 祁连山北麓,降水多、蒸发少、气温低、高寒阴湿,是河西走廊生态文明建设和社会经济发展的基础.由于区域地形复杂,水热梯度明显,植被的垂直带性分布影响着生物多样性的分布[12 ] ;因此研究祁连山北麓植被垂直带性分布下的土壤C、N、P化学计量学特征,以阐明祁连山北麓土壤C、N、P循环机制和土壤养分的变化规律势在必行. ...
1
2009
... 土壤有机C、全N、全P、土壤容重(pb ,环刀法)的测定参照《土壤分析技术规范》[13 ] ,土壤C、N、P储量采用下式计算[14 ] . ...
亚热带2种针叶林土壤碳氮磷储量及化学计量比对混交的响应
1
2019
... 土壤有机C、全N、全P、土壤容重(pb ,环刀法)的测定参照《土壤分析技术规范》[13 ] ,土壤C、N、P储量采用下式计算[14 ] . ...
1
2000
... 不同海拔土壤C、N、P化学计量比特征结果见图3 ,随着海拔升高C/N呈降低趋势,幅度为10.63—63.38,平均值21.96,变异系数65.31%,C/N在海拔2 814—3 128 m内较高,在各样点(除海拔3 750 m)C/N均高于国内土壤的平均值12.10[15 -18 ] ;C/P在高海拔4 130 m处最低,3 086 m处最高,变幅26.47—149.24,平均82.94,变异系数为42.94%;N/P变幅1.05—8.59,平均4.82,变异系数52.95%.不同海拔C/N、C/P随海拔整体呈现降低趋势,而N/P变化规律不同,在最高和低海拔处最低. ...
井冈山国家级自然保护区森林土壤养分含量的空间变化
2014
Study on spatial variability of soil nutrients in Ebinur Lake wetlands in China
2015
Pattern and variation of C∶N∶P ratios in China’s soils:asynthesis of observational data
1
2010
... 不同海拔土壤C、N、P化学计量比特征结果见图3 ,随着海拔升高C/N呈降低趋势,幅度为10.63—63.38,平均值21.96,变异系数65.31%,C/N在海拔2 814—3 128 m内较高,在各样点(除海拔3 750 m)C/N均高于国内土壤的平均值12.10[15 -18 ] ;C/P在高海拔4 130 m处最低,3 086 m处最高,变幅26.47—149.24,平均82.94,变异系数为42.94%;N/P变幅1.05—8.59,平均4.82,变异系数52.95%.不同海拔C/N、C/P随海拔整体呈现降低趋势,而N/P变化规律不同,在最高和低海拔处最低. ...
泥石流频发区山地不同海拔土壤化学计量特征:以云南省小江流域为例
1
2016
... 土壤养分是森林生态系统中植物营养的主要来源,森林群落结构、物种组成及生产力均受土壤养分的影响[19 ] .土壤养分是海拔梯度主导下地形、气候以及生物因素相互作用的结果,具有显著的空间异质特征[20 ] .本研究发现,在海拔2 814—4 130 m,乔木林土壤C、N、P含量随土层深度的增加而降低,表层土壤显著高于下层,而P含量相较C、TN含量在土壤剖面上变化不大.在0—40 cm的土层C、N含量的空间分布基本一致,而P含量的变化滞后于C、N含量,C、N和P含量的平均值分别为61.72、3.79 mg∙g-1 ,均高于中国0—10 cm土壤有机C、全N含量的均值24.56、1.88 mg∙g-1 ,而P含量0.72 mg∙g-1 稍低于中国0—10 cm土壤全P含量均值0.78 mg∙g-1[21 ] ,说明该流域土壤C含量高于N,而P含量相对匮乏.0—40 cm土层C、N含量的平均值空间变异系数较大,而P空间变异系数较小,这一结果与吕世丽等[22 ] 的研究结果相似.因为表层土壤受外界环境因素及植被凋落物养分的影响,导致土壤养分在表层密集[22 ] ,土壤C、N除受土壤母质的影响外,还受植物凋落物分解以及植物的吸收利用等影响,而存在着较大的空间变异性[23 ] ;而土壤全P含量主要受成土母质的影响,动植物残体、微生物活动等对全P的影响相对较小[1 ] ,因此,P含量在土层间存在较小的空间变异性,且相对稳定. ...
黄土丘陵沟壑区不同植被区土壤生态化学计量特征
1
2013
... 土壤养分是森林生态系统中植物营养的主要来源,森林群落结构、物种组成及生产力均受土壤养分的影响[19 ] .土壤养分是海拔梯度主导下地形、气候以及生物因素相互作用的结果,具有显著的空间异质特征[20 ] .本研究发现,在海拔2 814—4 130 m,乔木林土壤C、N、P含量随土层深度的增加而降低,表层土壤显著高于下层,而P含量相较C、TN含量在土壤剖面上变化不大.在0—40 cm的土层C、N含量的空间分布基本一致,而P含量的变化滞后于C、N含量,C、N和P含量的平均值分别为61.72、3.79 mg∙g-1 ,均高于中国0—10 cm土壤有机C、全N含量的均值24.56、1.88 mg∙g-1 ,而P含量0.72 mg∙g-1 稍低于中国0—10 cm土壤全P含量均值0.78 mg∙g-1[21 ] ,说明该流域土壤C含量高于N,而P含量相对匮乏.0—40 cm土层C、N含量的平均值空间变异系数较大,而P空间变异系数较小,这一结果与吕世丽等[22 ] 的研究结果相似.因为表层土壤受外界环境因素及植被凋落物养分的影响,导致土壤养分在表层密集[22 ] ,土壤C、N除受土壤母质的影响外,还受植物凋落物分解以及植物的吸收利用等影响,而存在着较大的空间变异性[23 ] ;而土壤全P含量主要受成土母质的影响,动植物残体、微生物活动等对全P的影响相对较小[1 ] ,因此,P含量在土层间存在较小的空间变异性,且相对稳定. ...
祁连山北麓山体垂直带土壤碳氮分布特征及影响因素
1
2009
... 土壤养分是森林生态系统中植物营养的主要来源,森林群落结构、物种组成及生产力均受土壤养分的影响[19 ] .土壤养分是海拔梯度主导下地形、气候以及生物因素相互作用的结果,具有显著的空间异质特征[20 ] .本研究发现,在海拔2 814—4 130 m,乔木林土壤C、N、P含量随土层深度的增加而降低,表层土壤显著高于下层,而P含量相较C、TN含量在土壤剖面上变化不大.在0—40 cm的土层C、N含量的空间分布基本一致,而P含量的变化滞后于C、N含量,C、N和P含量的平均值分别为61.72、3.79 mg∙g-1 ,均高于中国0—10 cm土壤有机C、全N含量的均值24.56、1.88 mg∙g-1 ,而P含量0.72 mg∙g-1 稍低于中国0—10 cm土壤全P含量均值0.78 mg∙g-1[21 ] ,说明该流域土壤C含量高于N,而P含量相对匮乏.0—40 cm土层C、N含量的平均值空间变异系数较大,而P空间变异系数较小,这一结果与吕世丽等[22 ] 的研究结果相似.因为表层土壤受外界环境因素及植被凋落物养分的影响,导致土壤养分在表层密集[22 ] ,土壤C、N除受土壤母质的影响外,还受植物凋落物分解以及植物的吸收利用等影响,而存在着较大的空间变异性[23 ] ;而土壤全P含量主要受成土母质的影响,动植物残体、微生物活动等对全P的影响相对较小[1 ] ,因此,P含量在土层间存在较小的空间变异性,且相对稳定. ...
牛背梁自然保护区不同海拔高度森林土壤养分特征分析
2
2013
... 土壤养分是森林生态系统中植物营养的主要来源,森林群落结构、物种组成及生产力均受土壤养分的影响[19 ] .土壤养分是海拔梯度主导下地形、气候以及生物因素相互作用的结果,具有显著的空间异质特征[20 ] .本研究发现,在海拔2 814—4 130 m,乔木林土壤C、N、P含量随土层深度的增加而降低,表层土壤显著高于下层,而P含量相较C、TN含量在土壤剖面上变化不大.在0—40 cm的土层C、N含量的空间分布基本一致,而P含量的变化滞后于C、N含量,C、N和P含量的平均值分别为61.72、3.79 mg∙g-1 ,均高于中国0—10 cm土壤有机C、全N含量的均值24.56、1.88 mg∙g-1 ,而P含量0.72 mg∙g-1 稍低于中国0—10 cm土壤全P含量均值0.78 mg∙g-1[21 ] ,说明该流域土壤C含量高于N,而P含量相对匮乏.0—40 cm土层C、N含量的平均值空间变异系数较大,而P空间变异系数较小,这一结果与吕世丽等[22 ] 的研究结果相似.因为表层土壤受外界环境因素及植被凋落物养分的影响,导致土壤养分在表层密集[22 ] ,土壤C、N除受土壤母质的影响外,还受植物凋落物分解以及植物的吸收利用等影响,而存在着较大的空间变异性[23 ] ;而土壤全P含量主要受成土母质的影响,动植物残体、微生物活动等对全P的影响相对较小[1 ] ,因此,P含量在土层间存在较小的空间变异性,且相对稳定. ...
... [22 ],土壤C、N除受土壤母质的影响外,还受植物凋落物分解以及植物的吸收利用等影响,而存在着较大的空间变异性[23 ] ;而土壤全P含量主要受成土母质的影响,动植物残体、微生物活动等对全P的影响相对较小[1 ] ,因此,P含量在土层间存在较小的空间变异性,且相对稳定. ...
Foliar and soil nutrients in tundra onglacial landscapes of contrasting ages in northern Alaska
1
2016
... 土壤养分是森林生态系统中植物营养的主要来源,森林群落结构、物种组成及生产力均受土壤养分的影响[19 ] .土壤养分是海拔梯度主导下地形、气候以及生物因素相互作用的结果,具有显著的空间异质特征[20 ] .本研究发现,在海拔2 814—4 130 m,乔木林土壤C、N、P含量随土层深度的增加而降低,表层土壤显著高于下层,而P含量相较C、TN含量在土壤剖面上变化不大.在0—40 cm的土层C、N含量的空间分布基本一致,而P含量的变化滞后于C、N含量,C、N和P含量的平均值分别为61.72、3.79 mg∙g-1 ,均高于中国0—10 cm土壤有机C、全N含量的均值24.56、1.88 mg∙g-1 ,而P含量0.72 mg∙g-1 稍低于中国0—10 cm土壤全P含量均值0.78 mg∙g-1[21 ] ,说明该流域土壤C含量高于N,而P含量相对匮乏.0—40 cm土层C、N含量的平均值空间变异系数较大,而P空间变异系数较小,这一结果与吕世丽等[22 ] 的研究结果相似.因为表层土壤受外界环境因素及植被凋落物养分的影响,导致土壤养分在表层密集[22 ] ,土壤C、N除受土壤母质的影响外,还受植物凋落物分解以及植物的吸收利用等影响,而存在着较大的空间变异性[23 ] ;而土壤全P含量主要受成土母质的影响,动植物残体、微生物活动等对全P的影响相对较小[1 ] ,因此,P含量在土层间存在较小的空间变异性,且相对稳定. ...
桂西北不同森林类型土壤生态化学计量特征
1
2015
... 海拔的变化将会导致气候特征、植被类型与土壤类型等发生相应改变,从而影响土壤养分的差异[24 ] .本研究表明,0—40 cm土壤有机C、N平均含量除海拔3 365 m和3 516 m外,在3 204—3 732 m显著高于海拔3 075 m以下和海拔3 750 m以上土壤的C、N含量,而在中高海拔3 379 m和3 250 m土壤C、N平均含量最高,这一结论与Cleveland等[25 ] 的研究结果一致.因为土壤有机质含量与土壤温度和水分有关,适宜的温度有利于有机质的积累,温度过高或过低对土壤有机质的积累均有负效应[8 ,25 ] ;其次,由于各海拔植被类型的不同使得土壤有机C的积累差异较大,N的变化与C的变化基本同步.P含量随海拔升高呈先升高后降低的趋势,在中海拔3 128 m处土壤P含量显著高于其他海拔(P <0.05),可能与该海拔范围内成土母质的分化有关. ...
C∶N∶P stoichiometry in soil:is there a“redfield ratio”for the microbial biomass
3
2007
... 海拔的变化将会导致气候特征、植被类型与土壤类型等发生相应改变,从而影响土壤养分的差异[24 ] .本研究表明,0—40 cm土壤有机C、N平均含量除海拔3 365 m和3 516 m外,在3 204—3 732 m显著高于海拔3 075 m以下和海拔3 750 m以上土壤的C、N含量,而在中高海拔3 379 m和3 250 m土壤C、N平均含量最高,这一结论与Cleveland等[25 ] 的研究结果一致.因为土壤有机质含量与土壤温度和水分有关,适宜的温度有利于有机质的积累,温度过高或过低对土壤有机质的积累均有负效应[8 ,25 ] ;其次,由于各海拔植被类型的不同使得土壤有机C的积累差异较大,N的变化与C的变化基本同步.P含量随海拔升高呈先升高后降低的趋势,在中海拔3 128 m处土壤P含量显著高于其他海拔(P <0.05),可能与该海拔范围内成土母质的分化有关. ...
... ,25 ];其次,由于各海拔植被类型的不同使得土壤有机C的积累差异较大,N的变化与C的变化基本同步.P含量随海拔升高呈先升高后降低的趋势,在中海拔3 128 m处土壤P含量显著高于其他海拔(P <0.05),可能与该海拔范围内成土母质的分化有关. ...
... 土壤C与N含量呈极显著正相关,C、N含量与C/P、N/P呈极显著正相关,而N含量与C/N呈极显著负相关,P含量与C/P呈极显著负相关,说明该流域0—40 cm土层土壤养分C、N、P在空间分布上相互耦合[25 ] . ...
Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation
1
2003
... 土壤C、N、P生态化学计量比是土壤有机C、N、P各元素总含量的比值,是评价土壤养分限制以及C、N、P饱和诊断和有效的预测性指标[26 ] .由于地形、气候、土壤质地、植被、动物及土壤微生物等成土因素和人类活动的影响,不同海拔的土壤C、N、P含量有较大的差异,从而使土壤C/N、C/P和N/P存在较大的空间变异性[27 ] .本研究区域不同海拔0—40 cm土壤C/N差异显著(P <0.05),随海拔升高总体呈下降趋势,在海拔3 128—2 814 m处高于其他海拔,在海拔3 075 m处最高(63.38),研究区土壤C/N的平均值21.96高于全球土壤C/N平均值13.33[2 ] ,表明土壤有机质的矿质化和腐殖质化明显,土壤N含量低. ...
Stoichiometric variation of carbon,nitrogen,and phosphorus in soils and its implication for nutrient limitation in alpine ecosystem of Eastern Tibetan Plateau
2
2016
... 土壤C、N、P生态化学计量比是土壤有机C、N、P各元素总含量的比值,是评价土壤养分限制以及C、N、P饱和诊断和有效的预测性指标[26 ] .由于地形、气候、土壤质地、植被、动物及土壤微生物等成土因素和人类活动的影响,不同海拔的土壤C、N、P含量有较大的差异,从而使土壤C/N、C/P和N/P存在较大的空间变异性[27 ] .本研究区域不同海拔0—40 cm土壤C/N差异显著(P <0.05),随海拔升高总体呈下降趋势,在海拔3 128—2 814 m处高于其他海拔,在海拔3 075 m处最高(63.38),研究区土壤C/N的平均值21.96高于全球土壤C/N平均值13.33[2 ] ,表明土壤有机质的矿质化和腐殖质化明显,土壤N含量低. ...
... 植被类型是影响土壤养分分布的重要因素,不同植被类型的土壤C/N、C/P、N/P存在显著差异[27 ] .本研究3种植被类型的土壤C/N差异不显著,说明祁连山北麓不同植被类型土壤有机质分解速度基本相近,3种植被类型土壤C/N平均值乔木林28.56>灌丛20.10>高山草地16.54.本研究区乔木林以青海云杉为主,其次为祁连圆柏和山杨,由于其凋落物丰富,凋落物分解速率较快,有效提高了土壤有机C含量,使其C/N提高,而灌丛和高山草地主要为小叶金露梅、银露梅、鬼箭锦鸡儿等,相对于乔木林,凋落物相对少,土壤C/N较低[28] .乔木林土壤C/P显著高于灌木和草地,变化趋势与C/N相同,即为乔木林105.89>灌丛80.51>高山草地54.00,说明高山草地土壤P的有效性高;土壤N/P可以作为养分限制类型的有效预测指标,祁连山北麓不同植被类型土壤N/P差异不显著,变化趋势为灌丛4.99>乔木林5.06>高山草地4.02,均小于10,说明祁连山北麓不同植被受土壤N限制. ...
贺兰山不同海拔典型植被带土壤微生物多样性
2
2013
... N/P可用作N饱和的诊断指标,也可反映土壤N、P平衡状况,被用于确定养分限制的阈值,能指示植物生长过程中土壤营养成分的供应情况[29 ] .研究区不同海拔N/P差异显著(P <0.05),在海拔3 204—3 894 m范围内土壤N/P较高,且在3 454 m处最高(8.59),在2 814 m处最低(1.05).研究区土壤N/P的平均值为4.82,高于中国森林生态系统土壤N/P (3.20)[28] ,说明研究区域土壤P相对缺乏.研究区海拔高、气温低、降雨量大,土壤P元素淋溶强、风化弱,不易累积,土壤P缺乏可能是C/P、N/P高于全球土壤和中国森林生态系统土壤的主要原因. ...
... 不同样点土壤C、N、P含量及其化学计量比的差异与海拔、植被类型、盖度和坡向有直接的关系,尤其以海拔和植被类型影响最为显著[29 -30 ] .研究表明,植被类型与C含量、C/P、N/P呈正相关,因为植被类型不同,每年归还到土壤的凋落物量也不同,且不同植被凋落物C、N、P含量的差异导致土壤C/N、C/P、N/P存在显著的差异.海拔与C/N呈正相关而与N/P、N呈负相关,说明N越低,C/N越高,表明流域整体土壤的C/N和N/P主要受N含量的影响.C/N与植被盖度呈正相关(0.309),但不显著,也说明地上植物是土壤中C素和N素的主要来源.坡向与土壤C/N呈正相关,而与土壤N/P呈负相关.以上分析表明,该区域植被类型、海拔是影响土壤C、N、P含量及其化学计量特征的主要因素. ...
祁连山中部不同植被类型土壤生态化学计量特征研究
1
2019
... 不同样点土壤C、N、P含量及其化学计量比的差异与海拔、植被类型、盖度和坡向有直接的关系,尤其以海拔和植被类型影响最为显著[29 -30 ] .研究表明,植被类型与C含量、C/P、N/P呈正相关,因为植被类型不同,每年归还到土壤的凋落物量也不同,且不同植被凋落物C、N、P含量的差异导致土壤C/N、C/P、N/P存在显著的差异.海拔与C/N呈正相关而与N/P、N呈负相关,说明N越低,C/N越高,表明流域整体土壤的C/N和N/P主要受N含量的影响.C/N与植被盖度呈正相关(0.309),但不显著,也说明地上植物是土壤中C素和N素的主要来源.坡向与土壤C/N呈正相关,而与土壤N/P呈负相关.以上分析表明,该区域植被类型、海拔是影响土壤C、N、P含量及其化学计量特征的主要因素. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}