


0 引言
化石燃料燃烧、氮肥施用、畜禽养殖等人为活动使陆地生态系统氮输入不断增加[1]。1980—2010年,中国总的氮沉降量平均以每年0.4 kg·hm-2的速率持续增加,并且随着经济、农业等的进一步发展,氮沉降量可能还会继续升高[2-3]。氮是植物生长发育必不可缺的营养元素[4],氮素水平直接影响植物的生长发育和新陈代谢[5]。在全球气候变化的背景下,全球降水格局也有着显著变化[6]。水分是干旱、半干旱区草原植物生长、发育最重要的限制因素[7]。降水的改变意味着荒漠生态系统中的植物也将在生长发育、形态特征、生理机能等方面发生改变[8]。氮素和水分是植物生长的重要环境限制因子,尤其是在受养分和水分限制的荒漠生态系统[9]。因此,研究氮沉降和降水变化对荒漠植物的生理特性影响,对揭示全球气候变化背景下荒漠植物响应和适应机制具有重要的意义。氮素和水分之间存在显著的耦合效应,二者对植物的综合效应往往是通过植物本身复杂的生理过程而起作用的[10]。李静静等[11]研究得出,在同等干旱胁迫下,适当增施氮素可提高草地早熟禾(Poa pratensis)叶片中可溶性蛋白含量,而丙二醛(MDA)含量则随着施氮量的增加呈现出先降低后增加的变化趋势,但在相同氮素水平下,过氧化氢酶(CAT)活性则随着水分的增加而增强,MDA含量则随着水分胁迫的加重而增加,超氧化物歧化酶(SOD)、过氧化物酶(POD)活性也均随着水分的降低而呈现出先升高后降低的变化趋势。李香云等[12]研究荒漠草原植物群落光合速率对水氮添加的响应,发现氮添加和增加降水的交互效应显著提高了群落的光合速率和优势种植物高度。因此开展氮沉降和降水变化对典型荒漠植物的影响研究,对揭示全球气候变化背景下荒漠植物响应和适应机制具有重要的意义。
红砂(Reaumuria soongorica)为柽柳科植物,广泛分布在中国干旱沙漠地区,是荒漠地区中的主要植物[13],具有耐瘠薄、适应性广、集沙能力强等特点,对荒漠地区的生态保护具有重要作用[14]。珍珠猪毛菜(Salsola passerina)为藜科植物,具有一系列的抗旱特征[15]。自然分布群落中,有时红砂和珍珠猪毛菜各自单独生长,有时它们紧密生长在一起[16]。近年来,有关降水对红砂和珍珠猪毛菜的光合荧光影响的研究较多[17-19],而关于在不同生长方式下两种荒漠植物对氮沉降和降水变化的生理特性响应的研究相对较少,那么气候变化对红砂和珍珠猪毛菜生长方式是否会产生影响?这种影响通过生理变化是如何来反映的?为此,本研究以2种典型荒漠植物红砂和珍珠猪毛菜为研究材料,通过开展人工模拟氮素沉降和降水处理试验,探究2种植物3种生长方式下的生理响应,以期进一步了解不同竞争模式下荒漠植物应对环境变化的适应能力和适应机制,为深入探讨中国干旱荒漠区植物群落的更新与建设提供科学依据。
1 材料与方法
1.1 材料与处理
试验于2019年4月至2019年9月在甘肃农业大学试验基地的人工遮雨棚内进行。以前期培育好的同一批民勤种源1年实生苗为试验材料。2019年4月初从苗圃选取生长一致的幼苗移栽到上口径18 cm、底口径10 cm、高25.5 cm的花盆内培育,每盆栽种2株。生长方式分为单生红砂(HH:2株红砂为1盆)、单生珍珠猪毛菜(ZZ:2株珍珠猪毛菜为1盆)和混生红砂-珍珠猪毛菜(HZ:红砂和珍珠猪毛菜各1株为1盆)。盆内实验用土均取自民勤种源地红砂-珍珠猪毛菜灌木林下0—20 cm去杂后混合均匀的均质土壤。2019年5月1日选取长势一致(红砂和珍珠猪毛菜株高10 cm左右)的幼苗进行施氮和降水控制试验。
根据Jia等[20]的研究结果,中国的西北湿沉降速率0—1.0 g·m-2·a-1,且近10年来局部地区沉降速度达4.0 g·m-2·a-1,大气氮沉降中的氮元素主要是以NHx-N和NOy-N的形式存在,但在河西地区以NHx-N 为主。鉴于此,施氮分为N0(对照)、N1(中氮4 g·m-2·a-1)、N2(高氮8 g·m-2·a-1)3种处理,由(NH4)2SO4提供,每月初将氮肥溶于100 mL水中施入土壤,对照施入等量的水。依据民勤1961—2008年气象资料显示,该区多年平均降水量为116.7 mm,降水量较高的年份为154.2 mm,比多年平均水平高约30%,降水量最低为81.5 mm,比多年平均水平低30%[21]。因而试验期间降水设置为:降水减少30%(W-)、对照(W)和降水增加30%(W+)3个梯度。以两种植物生长旺盛期(5—9月)每月的降水量为基准对照,换算为各处理每月的总灌水量,分10次施入(每3天1次),月平均降雨量及各水分处理下每次灌水量列于表1。处理期间HH、ZZ和HZ的氮沉降和降水交互处理各9个处理(N0W-、N0W、N0W+、N1W-、N1W、N1W+、N2W-、N2W、N2W+),每个处理取6盆,共计162盆。
表1 1961—2008年5—9月平均降水量及每次灌水量
Table 1
| 处理 时间 | 每月平均 降水量 /mm | 每次灌水量/mL | ||
|---|---|---|---|---|
| W-(降水 减少30%) | W (对照) | W+(降水 增加30%) | ||
| 5月 | 11.2 | 13.05 | 18.64 | 24.23 |
| 6月 | 15.0 | 17.47 | 24.96 | 32.45 |
| 7月 | 23.0 | 26.80 | 38.28 | 49.76 |
| 8月 | 28.1 | 32.73 | 46.76 | 60.79 |
| 9月 | 18.2 | 21.20 | 30.29 | 39.38 |
1.2 指标测定与方法
1.3 数据处理
采用Excel 2010软件对数据进行统计和制表,用统计分析软件SPSS 22进行三因素方差分析检验氮素、降水、生长方式及其交互对红砂和珍珠猪毛菜幼苗的影响,利用单因素方差分析检验同一降雨和氮素不同生长方式下各指标的差异,不同水平间的多重比较用Duncan法。
2 结果与分析
2.1 氮沉降和降水变化对红砂和珍珠猪毛菜叶片MDA的影响
氮沉降、降水量及种植方式对红砂叶片MDA含量影响极显著(P<0.01,表2),氮和生长方式交互影响极显著(P<0.01),而其他交互作用下,对红砂影响均不显著;除水和生长方式交互对珍珠猪毛菜的MDA含量影响不显著,其他对珍珠猪毛菜影响均显著(P<0.05)或极显著(P<0.01)。施氮和增水交互均使不同生长方式下红砂和珍珠猪毛菜叶片MDA含量降低,且在降水增加30%和施氮量8 g·m-2·a-1时最小(图1)。与对照相比,单生红砂、混生红砂、单生珍珠猪毛菜、混生珍珠猪毛菜的MAD含量最大分别降低9.36%、15.13%、14.36%、18.77%。同一氮沉降和降水处理下,红砂和珍珠猪毛菜混生使红砂的MDA含量显著增高(P<0.05),珍珠猪毛菜叶片MDA含量显著降低(P<0.05),N0W-、N0W、N0W+、N1W-、N1W、N1W+、N2W-、N2W、N2W+处理下,混生红砂MAD含量比单生红砂分别增加12.36%、13.14%、11.56%、6.98%、7.77%、8.32%、5.24%、3.24%、5.94%,混生珍珠猪毛菜比单生珍珠猪毛菜分别减少18.19%、21.13%、24.28%、23.68%、24.24%、26.39%、25.16%、26.14%、25.19%。
表2 氮沉降、降水量、生长方式及其交互作用对红砂和珍珠猪毛菜丙二醛(MDA)含量和抗氧化酶活性影响的方差分析
Table 2
| 植物种 | 因素 | 丙二醛MDA | 过氧化氢酶CAT | 过氧化物酶POD | 超氧化物歧化酶SOD |
|---|---|---|---|---|---|
| 红砂 | 氮 | 113.877** | 2 977.441** | 627.185** | 59 320.919** |
| 水 | 310.148** | 8 779.43** | 1067.717** | 16 734.038** | |
| 生长方式 | 458.608** | 1519.006** | 1 651.706** | 25 127.108** | |
| 氮×水 | 2.556 | 387.83** | 13.831** | 499.512** | |
| 氮×生长方式 | 35.004** | 201.458** | 40.865** | 1 892.336** | |
| 水×生长方式 | 0.239 | 91.21** | 21.243** | 819.311** | |
| 氮×水×生长方式 | 1.206 | 40.52** | 24.863** | 218.649** | |
| 珍珠猪毛菜 | 氮 | 199.311** | 5 089.873** | 1 314.613** | 70 938.048** |
| 水 | 165.466** | 6 739.326** | 4 576.053** | 15 140.846** | |
| 生长方式 | 3 330.418** | 14 659.648** | 1 039.276** | 9 521.426** | |
| 氮×水 | 3.117* | 157.069** | 4.986** | 1 929.279** | |
| 氮×生长方式 | 3.856* | 1 230.202** | 11.71** | 1 200.972** | |
| 水×生长方式 | 0.405 | 428.483** | 88.045** | 173.447** | |
| 氮×水×生长方式 | 3.308* | 166.856** | 12.914** | 495.889** |
表中数值为F检验值;*表示在0.05水平上显著相关,**表示在0.01水平上极显著相关。
图1
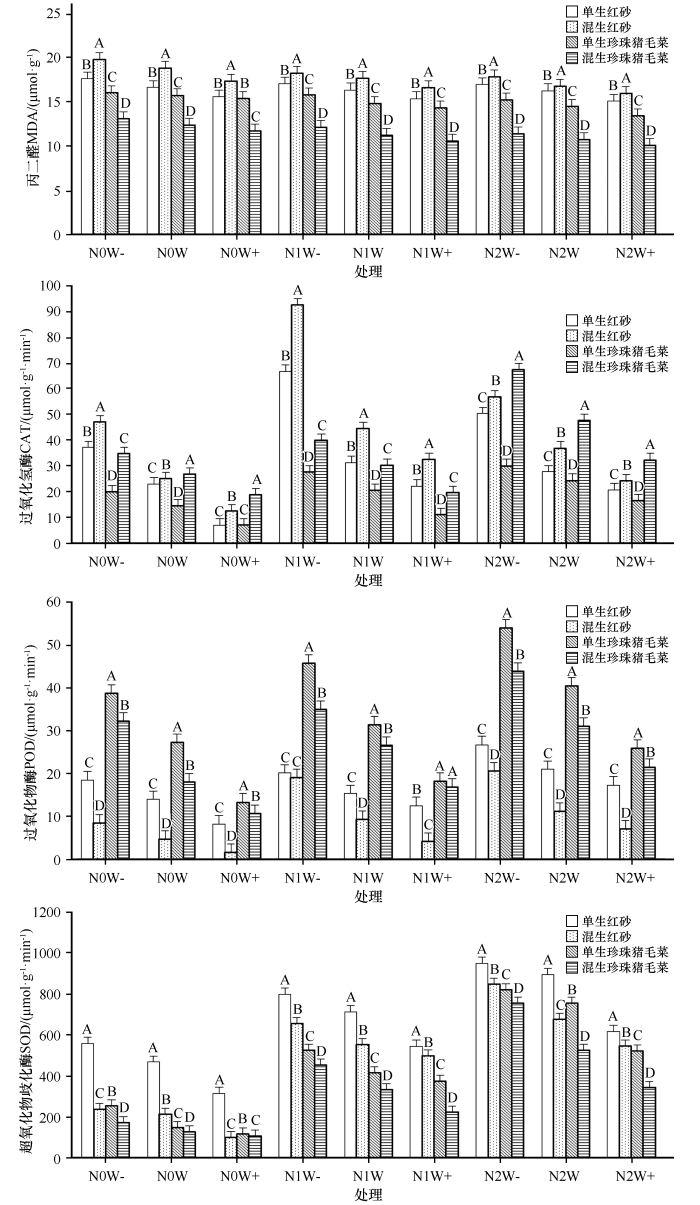
图1
氮沉降、降水量、生长方式及其交互作用对红砂和珍珠猪毛菜丙二醛含量和抗氧化酶活性的影响
不同大写字母表示同一施氮和降水水平下不同生长方式差异显著(P<0.05)
Fig.1
Effects of nitrogen deposition, precipitation, growth mode and their interaction on malondialdehyde content and antioxidant enzyme activity in Reaumuria soongorica and Salsola passerina
2.2 氮沉降和降水变化对红砂和珍珠猪毛菜叶片抗氧化酶活性的影响
施氮、降水和种植方式及其交互对红砂和珍珠猪毛菜叶片CAT、POD、SOD活性影响均极显著(P<0.01,表2)。
单生红砂和混生红砂的CAT活性均随施氮量的增加呈先升后降的变化趋势(N1>N2>N0),随降水量的增加而降低(图1)。与N0W(对照)相比,单生红砂(MDA)N0W-、N1W-、N1W、N2W-、N2W下分别增加61.55%、1.90倍、36.36%、1.19倍、20.27%,N0W+、N1W+、N2W+下分别降低69.40%、3.45%、10.10%;混生红砂N0W-、N1W-、N1W、N1W+、N2W-、N2W下分别增加87.66%、2.70倍、77.61%、29.96%、1.27倍、47.34%,N0W+、N2W+下分别降低50.60%、3.86%。单生珍珠猪毛菜和混生珍珠猪毛菜的CAT活性随氮沉降增加而上升,随降水增加而下降,与N0W(对照)相比,除N0W+和N1W+处理下CAT活性有所降低,其他处理下活性均升高,单生珍珠猪毛菜和混生珍珠猪毛菜N0W-、N1W-、N1W、N2W-、N2W、N2W+下分别增加39.10%、91.84%、42.79%、1.10倍、69.58%、13.98%和30.12%、49.28%、13.38%、1.52倍、78.37%、20.97%,N0W+、N1W+下分别降低50.29%、23.58%和30.26%、26.57%。
不同生长方式下红砂和珍珠猪毛菜的POD、SOD活性均随施氮量增加而升高,随降水增加而降低。单生红砂和混生红砂的POD活性在N0W-、N1W-、N1W、N2W-、N2W、N2W+处理下分别增加32.79%、44.32%、9.69%、92.16%、50.61%、23.67%和80.29%、3.13倍、1.00倍、3.46倍、1.42倍、50.12%,N0W+、N1W+处理下分别降低41.42%、10.57%、和66.78%、11.44%;单生珍珠猪毛菜的POD活性在N0W-、N1W-、N1W、N2W-、N2W处理下分别增加42.58%、68.69%、15.54%、98.68%、46.81%,N0W+、N1W+ 、N2W+处理下分别降低51.25%、33.29%、4.52%;混生珍珠猪毛菜的POD活性在N0W-、N1W-、N1W、N2W-、N2W、N2W+处理下分别增加78.60%、93.99%、46.82%、1.44倍、71.89%、18.93%,N0W+、N1W+处理下分别降低41.23%、6.98%。与对照相比,不同生长方式下红砂和珍珠猪毛菜的SOD活性除在N0W+处理下低于对照,其余处理均高于对照。单生红砂和混生红砂的SOD活性在N0W-、N1W-、N1W、N1W+、N2W-、N2W、N2W+处理下分别增加19.69%、71.11%、52.66%、16.56%、1.03倍、91.85%、32.27%和11.79%、2.10倍、1.60倍、1.35倍、3.00倍、2.18倍、1.58倍,N0W+处理下降低32.91%和53.25%;单生珍珠猪毛菜和混生珍珠猪毛菜的SOD活性在N0W-、N1W-、N1W、N1W+、N2W-、N2W、N2W+处理下分别增加74.14%、2.61倍、1.85倍、1.57倍、4.65倍、4.20倍、2.59倍和36.01%、2.60倍、1.63倍、76.12%、4.99倍、3.15倍、1.73倍,N0W+处理下降低20.37%和15.72%。红砂和珍珠猪毛菜混生显著(P<0.05)提高了红砂和珍珠猪毛菜的CAT活性,而使POD、SOD活性均显著(P<0.05)降低。 N0W-、N0W、N0W+、N1W-、N1W、N1W+、N2W-、N2W、N2W+处理下,混生红砂CAT活性比单生红砂分别增加26.45%、8.86%、75.73%、38.74%、41.78%、46.52%、12.93%、33.36%、16.41%;混生珍珠猪毛菜CAT活性比单生珍珠猪毛菜分别增加74.39%、86.42%、1.62倍、45.06%、48.02%、79.12%、1.24倍、96.08%、97.84%;混生红砂POD、SOD活性比单生红砂分别降低55.09%、66.99%、81.28%、5.53%、39.78%、67.31%、23.41%、46.93%、59.93%和57.60%、54.60%、68.37%、17.88%、22.60%、8.53%、10.63%、24.68%、11.46%;混生珍珠猪毛菜POD、SOD活性比单生珍珠猪毛菜分别降低16.84%、33.61%、19.96%、23.65%、15.64%、7.43%、18.50%、23.31%、17.30%和32.23%、13.22%、8.16%、13.45%、19.91%、40.58%、7.87%、30.70%、34.13%。
2.3 氮沉降和降水变化对红砂和珍珠猪毛菜叶片渗透调节物质的影响
表3 氮沉降、降水量、生长方式及其交互作用对红砂和珍珠渗透调节物质含量影响的方差分析
Table 3
| 植物种 | 因素 | 可溶性蛋白SP | 脯氨酸Pro |
|---|---|---|---|
| 红砂 | 氮 | 1479.236** | 777.134** |
| 水 | 542.641** | 1315.021** | |
| 生长方式 | 110.267** | 1928.662** | |
| 氮×水 | 6.164** | 27.319** | |
| 氮×生长方式 | 1.068 | 59.441** | |
| 水×生长方式 | 5.308* | 120.339** | |
| 氮×水×生长方式 | 1.877 | 62.764** | |
| 珍珠猪毛菜 | 氮 | 4145.49** | 1843.819** |
| 水 | 1579.516** | 464.782** | |
| 生长方式 | 1614.298** | 10740.754** | |
| 氮×水 | 110.396** | 7.208** | |
| 氮×生长方式 | 50.814** | 1391.517** | |
| 水×生长方式 | 40.809** | 260.556** | |
| 氮×水×生长方式 | 18.191** | 6.376** |
表中数值为F检验值;*表示在0.05水平上显著相关,**表示在0.01水平上极显著相关。
图2
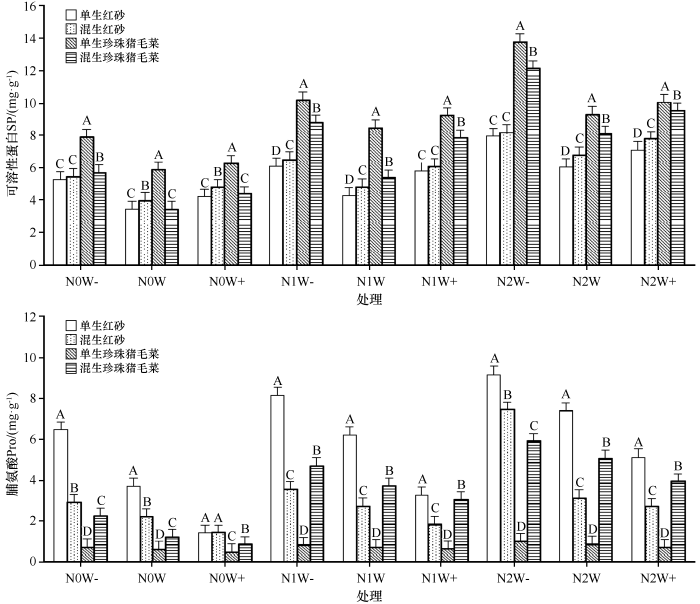
图2
氮沉降、降水量、生长方式及其交互作用对红砂和珍珠猪毛菜渗透调节物质(SP、Pro)含量的影响(不同字母表明差异显著)
Fig.2
Effects of nitrogen deposition, precipitation, growth mode and their interaction on the contents of osmotic adjustment substances (SP, Pro) in Reaumuria soongorica and Salsola passerina(Different letters indicate significant differences)
与N0W(对照)相比,N0W-、N0W+、N1W-、N1W、N1W+、N2W-、N2W、N2W+处理下单生红砂和混生红砂的SP含量分别增加52.63%、22.36%、76.90%、24.45%、68.45%、1.30倍、75.28%、1.06倍和37.91%、21.38%、64.78%、22.02%、53.61%、1.06倍、72.14%、96.41%,单生珍珠猪毛菜和混生珍珠猪毛菜的SP含量分别增加34.18%、6.80%、73.70%、44.26%、56.73%、1.35倍、58.13%、71.29%和65.56%、27.07%、1.55倍、56.62%、1.28倍、2.53倍、1.35倍、1.77倍。
不同生长方式下,红砂的Pro含量除N0W+和N1W+低于对照外,其余处理均高于对照,而珍珠猪毛菜除N0W处理低于对照,其他处理均高于对照。与对照相比,在N0W-、N1W-、N1W、N2W-、N2W、N2W+处理下单生红砂和混生红砂的Pro含量分别增加74.91%、1.21倍、67.88%、1.48倍、1.01倍、38.56%和32.11%、60.84%、23.84%、2.38倍、41.65%、23.35%,N0W+、N1W+处理下分别减少61.50%、11.65%和35.77%、17.31%;在N0W-、N1W-、N1W、N1W+、N2W-、N2W、N2W+处理下单生珍珠猪毛菜和混生珍珠猪毛菜的Pro含量分别增加18.57%、34.51%、16.33%、2.86%、64.34%、43.18%、16.90%和86.31%、2.89倍、2.08倍、1.51倍、3.89倍、3.19倍、2.25倍,N0W+处理下减少24.12%和30.30%。
红砂的SP含量在混生下显著(P<0.05)高于单生,而Pro则相反;珍珠猪毛菜的SP含量单生下显著(P<0.05)高于混生,Pro含量则混生显著(P<0.05)高于单生。N0W-、N0W、N0W+、N1W-、N1W、N1W+、N2W-、N2W、N2W+处理下,混生红砂SP含量比单生红砂分别增加3.28%、14.31%、13.39%、6.48%、12.08%、4.23%、2.55%、12.26%,而Pro含量分别减少55.06%、40.50%、0.74%、56.68%、56.11%、44.31%、18.92%、57.98%、47.03%;混生珍珠猪毛菜的SP比单生珍珠猪毛菜分别减少27.85%、41.53%、30.42%、14.01%、36.51%、14.84%、12.10%、13.15%、5.60%,而Pro含量分别增加2.15倍、1.00倍、84.18%、4.81倍、4.31倍、3.89倍、4.96倍、4.87倍、4.58倍。
3 讨论
氮沉降和降水变化会改变养分供应状况,进而影响植物的生理特性。增施氮肥可显著提高保护酶的活性,降低MDA含量,从而减少细胞质膜的过氧化损伤,而随降水增加,抗氧化酶活性及MDA含量均有所降低[25-27]。也有研究表明,适当增施氮肥能提高干旱胁迫下植物叶片内源保护酶活性,对细胞膜的稳定性有促进作用,明显减缓了植物叶片细胞膜系统受损伤的程度[28],但高氮下植物受胁迫加强,反而不利于植物生长[29]。本研究结果显示,增施氮肥使不同生长方式下红砂和珍珠猪毛菜的抗氧化酶活性均提高,而降水增加又使其活性降低,氮和水交互处理下,除施氮与降水增加交互处理下低于对照外,其他处理均高于对照。这说明在降水减少和正常降水下增施氮肥均能显著提高红砂和珍珠猪毛菜的抗氧化能力,在水分充足条件下,即使增施氮肥,抗氧化酶仍保持较低活性水平。氮水交互可显著降低不同生长方式下红砂和珍珠猪毛菜的MDA含量,且在高氮和降水增加交互下达最小,说明水氮耦合下,对两种植物的MDA影响更显著。在氮和水充足环境条件下,红砂和珍珠猪毛菜不受胁迫或受很小的胁迫强度,所以MDA含量处于较低水平。因为高氮和降水增加处理下,MDA含量较低,所以抗氧化酶活性保持较低水平依然能保证植物正常生长。植物体内渗透调节物质Pro含量的变化与植物的抗逆性相关[30],SP含量的增加也有利于提高植物的抗胁迫能力与适应性[31]。施氮量增加可提高土沉香(Aquilaria sinensis)幼苗Pro含量[32]和辣椒(Capsicum annuum)叶片SP含量[33],降雨量的改变会引起樟子松(Pinus sylvestris var. mongolica)针叶Pro和SP含量的升高[34]。本研究中,红砂和珍珠猪毛菜叶片SP含量均在高氮与降水减少交互处理和高氮与降水增加处理下含量较高。姬江莉等[27]研究得出,水分减少时,红砂通过渗透调节物质的积累来增强抗旱性,而降雨增加时这些物质的含量也有所增加,可能是因为在得到水分补偿时,红砂开始进行旺盛的生理生化反应与代谢活动从而促使其体内这些物质的含量升高。所以,本研究中降水减少下增施氮肥,显著增加SP含量,从而提高红砂和珍珠猪毛菜的抗逆性,而水氮充足条件下,植物合成蛋白能力加强,满足旺盛的生长代谢需求。而Pro含量对水氮变化响应十分敏感,在降水减少和高氮交互下增加最大,从而说明增施氮肥能提高干旱条件下植物的渗透调节物质,提高其抗旱性。
红砂和珍珠猪毛菜混生时两者表现为互惠关系,促进了根系对土壤养分和水分的吸收能力,且混生群落适应极端环境的能力要高于单生群落,这种混生群落对维持生态系统稳定和高效利用降水资源有着极其重要的作用[35]。本试验中,红砂和珍珠猪毛菜混生,均使二者CAT活性显著提高,而POD、SOD活性反而降低。混生红砂的MDA含量显著高于单生红砂,而混生珍珠猪毛菜叶片MDA含量低于单生珍珠猪毛菜,说明混生红砂的POD、SOD活性降低,降低了活性氧的清除能力,而混生珍珠猪毛菜因CAT活性的增加能补偿由于POD、SOD活性降低而引起活性氧的清除能力降低,从而达到高效清除能力,也可能是红砂在清除其体内活性氧自由基时以POD、SOD为主,CAT为辅,而珍珠猪毛菜以CAT为主。红砂和珍珠猪毛菜混生时,对两者的渗透调节物质的影响差异很大,红砂是以提高SP含量来增强其渗透调节能力,而珍珠猪毛菜则是增加Pro含量。植物种间互利的强度或重要性与环境密切相关[36]。氮添加会造成土壤酸化,打破土壤中的元素平衡,致使对土壤环境变化敏感的植物竞争力降低[37]。本研究中,增施氮肥和降水增加交互能降低红砂混生下MDA增加的幅度,而混生珍珠猪毛菜的MDA减少幅度增大;水氮处理也能更好地维持混生红砂和珍珠猪毛菜的抗氧化和渗透调节能力。
4 结论
氮沉降和降水交互作用下,对红砂和珍珠猪毛菜的生理特性有不同程度的影响,总体上表现为抗氧化酶活性升高和渗透调节物质增加,MDA含量降低,从而增强其抗逆境能力,说明在未来氮沉降和降水变化下有利于红砂和珍珠猪毛菜的生存。而水氮处理下能更好地维持混生红砂-珍珠猪毛菜的抗氧化和渗透调节能力,从而说明混生红砂-珍珠猪毛菜能适应未来环境变化,发挥竞争优势,稳定群落地位,具有成为主要生长方式趋势。
参考文献
Transformation of the nitrogen cycle:recent trends,questions,and potential solutions
[J].
Enhanced nitrogen deposition over China
[J].
Reactive nitrogen and the world:200 years of change
[J].
Photosynthetic responses of Mojave Desert shrubs to free air CO2 enrichment are greatest during wet years
[J].
The water adaptability of Jatrophacurcas is modulated by soil nitrogen availability
[J].
红砂(Reaumuria soongorica)忍耐极度干旱的保护机制: 叶片脱落和茎中蔗糖累积
[J].
Associated growth of C3 and C4 desert plants helps the C3 species at the cost of the C4 species
[J].
荒漠环境梯度下联生红砂(Reaumuria soongorica)、珍珠(Salsola passerina)荧光特性
[J].
红砂(Reaumuria soongorica)、珍珠(Salsola passerine)蒸腾耗水规律的尺度整合
[J].
Spatial and decadal variations in inorganic nitrogen wet deposition in China induced by human activity
[J].
Membrane lipid peroxidation,nitrogen fixation andleghemoglobin content in soybean root nodules
[J].
氮素营养调控小麦旗叶衰老和光合功能衰退的生理机制
[J].
Differential response of salt stress on Brassica juncea:photosynthetic performance,pigment,proline,D1 and antioxidant enzymes
[J].

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}