


0 引言
西北内陆河流域是中国最干旱的地区,水资源紧缺使其生态系统十分脆弱[1-3],尤其水生态系统更加单调和贫瘠[4-6]。内陆河流域不同时期的浮游植物群落结构特征明晰了水生态系统的综合变化规律,且在时间与空间尺度上的演变过程反映了河流水生态系统健康状况的变迁[7-10]。内陆河流域浮游植物群落特征主要表现为种类少、生物量低且分布很不均衡[11-13]。黑河流域浮游植物的空间分布有着与河流水文分布特征相对应的垂直地带性和水平地带性分异特征,中下游浮游植物群落结构的复杂程度和稳定性均高于上游[14-15]。以往研究表明,流域气候[16]、流速[17]、悬浮物[18]和营养[15,19]的经向变化共同造就了河流浮游植物分布的空间异质性,但鲜见针对黑河浮游植物的长周期分析,在时间和空间尺度上其群落演变特征及影响因素仍不明确。干旱区河流水环境状况与水域面积紧密相关。因此,本文尝试研究干旱区河流水域面积的变化对河流浮游植物群落的沿程变化和历史变化过程的影响,深入反映水文要素与水生态的变化规律,以期为内陆河河流健康提供更为有效的评判方法。
1 数据与方法
1.1 研究区域概况
黑河是中国西北地区第二大内陆河流。发源于祁连山的绵羊岭和讨赖山,流经青海祁连县、甘肃张掖盆地,最后注入内蒙古居延海,地理位置38°—42°N、98°—101°E。干流全长821 km,流域面积14.29万km2[19]。本研究主要选取黑河干流甘肃段,从上至下按照地理条件和气候特征分为3段进行对比研究。以八宝河与野牛沟汇合处至黑河出山口莺落峡为上游,莺落峡至高台县正义峡为中游,正义峡以下至甘肃省界哨马营为下游[20]。黑河上游处于祁连山区,属高寒半干旱气候区,为黑河主要产流区[21]。中游地势平坦、农垦发达,属温带干旱区,是黑河流域水资源主要开发利用区[22]。下游黑河途经戈壁沙漠,为极度干旱区,径流损失严重[23]。黑河水文过程与流域生态环境均受气候和人类活动的影响,分带性变化十分显著[24]。
1.2 数据来源
本文浮游植物调查数据均采用黑河夏季干流甘肃段的调查数据。1981年浮游植物数据主要参考《甘肃渔业资源与区划》[25]及相关渔业资源调查数据,调查时间为6—9月;1997年莺落峡至正义峡数据来自于李鹏等[14]研究,调查时间为5—8月;2008年数据来自甘肃丰源生态生物体系咨询中心完成的小孤山等水电站竣工环保验收调查数据及兰州大学完成的《张掖黑河湿地国家级自然保护区综合科学考察报告》等,调查时间为6—8月。2017年黑河流域浮游植物数据为现场调查数据,调查时间为7月,调查区域覆盖甘肃境内的黑河莺落峡以上(H1—H6)、莺落峡至正义峡(H7—H17)、正义峡至哨马营(H18—H19)3段共19个采样点(图1),包括自然河段、湿地、水库库区、水库减水河段、水库尾水河段等类型。浮游植物样品具体采集与分析方法同文献[26]所述。
图1
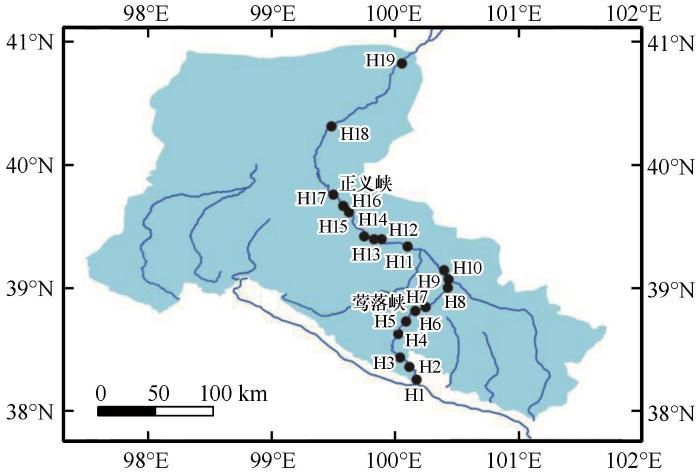
图1
黑河甘肃段浮游植物采样断面分布
Fig.1
Distribution of sampling sites in Gansu section of Heihe River
1.3 数据分析和处理
群落特征参数主要包括各断面浮游植物的物种组成、生物量、密度、物种多样性以及群落优势种。物种多样性采用Shannon-Weaver多样性指数(H)表征。H越大,群落的稳定性和抵抗外界干扰的程度越高。优势种用Manaughton优势度指数(Y)来度量。H和Y计算方法均采取传统计算方法[26]。为表征干旱区水生生物的活动能力和活跃程度,将每公顷水域面积条件下的物种多样性指数与样品总数量的乘积定义为生物活动力指数(M)。
式中:H为Shannon-Weaver多样性指数,N为样品浮游植物总数量(万cells,数值以密度计),Sa为水域面积(hm2)。M越大,浮游植物活动范围越大,活动能力越强。
黑河中游湿地面积数据采用遥感影像解译分析获取,数据源为1984—2015年覆盖全流域的Landsat多光谱影像存档数据集。图像预处理包括图像几何校正、辐射纠正、图像匹配与镶嵌等;图像增强处理包括比值拉伸、直方图均衡化、滤波处理等。针对流域下垫面结构特征,通过阈值分割分离出湿地,湿地主要包括河流、湖泊、河滩、沼泽、水库坑塘等。
2 结果与分析
2.1 物种的演变
黑河浮游植物群落结构有着显著的地理分布特征(表1)。2017年调查中共检出浮游植物8门75属。其中莺落峡以上上游浮游植物为40属,莺落峡至正义峡中游61属,正义峡至哨马营下游37属。全流域绿藻门物种最为丰富多达30属,占40.0%;硅藻门次之达24属,占32.0%。其他依次是蓝藻门8属,裸藻门5属,甲藻门3属,黄藻门、金藻门各2属,隐藻门仅1属占1.3%。从地理分布情况看,除金藻门、隐藻门主要出现在中游水域外,其余6门在全河段均有检出且各自成分比率相对稳定。
表1 黑河干流浮游植物群落结构特征
Table 1
| 种类(属) | 莺落峡以上 | 莺落峡至正义峡 | 正义峡以下 | 黑河流域 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1981年 | 2008年 | 2017年 | 1981年 | 1997年 | 2008年 | 2017年 | 1981年 | 2017年 | 1981年 | 2008年 | 2017年 | ||||
| 绿藻门 | 15 | 17 | 17 | 19 | 15 | 19 | 24 | 17 | 13 | 19 | 22 | 30 | |||
| 硅藻门 | 12 | 16 | 16 | 18 | 14 | 16 | 20 | 15 | 14 | 18 | 20 | 24 | |||
| 蓝藻门 | 3 | 2 | 3 | 5 | 9 | 3 | 6 | 5 | 4 | 5 | 3 | 8 | |||
| 裸藻门 | 3 | 2 | 2 | 4 | 2 | 3 | 5 | 3 | 1 | 4 | 3 | 5 | |||
| 甲藻门 | 3 | 1 | 1 | 2 | 3 | 2 | 2 | 2 | 3 | 3 | 2 | 3 | |||
| 黄藻门 | 1 | 1 | 1 | 1 | 2 | 2 | 1 | 1 | 2 | 1 | 2 | 2 | |||
| 金藻门 | 0 | 0 | 0 | 2 | 4 | 1 | 2 | 0 | 0 | 2 | 1 | 2 | |||
| 隐藻门 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | |||
| 种类总计 | 37 | 39 | 40 | 52 | 49 | 46 | 61 | 43 | 37 | 53 | 53 | 75 | |||
| 平均生物量/(mg·L-1) | 0.217 | 0.262 | 0.274 | 0.290 | — | 0.251 | 0.303 | 0.252 | 0.233 | 0.253 | 0.255 | 0.270 | |||
| 平均密度/(万cells·L-1) | 35.2 | 44.6 | 45.8 | 56.3 | — | 48.2 | 57.9 | 39.7 | 32.6 | 44.1 | 46.8 | 45.4 | |||
| Shannon-Weaver多样性指数(H) | 2.16 | 2.28 | 2.34 | 2.79 | — | 2.48 | 2.87 | 2.52 | 2.21 | — | — | — | |||
黑河浮游植物在不同时期的分布均呈现出与河流水文特征相对应的水平地带性分异特征。黑河浮游植物物种由1981年的53属,经2008年继续维持53属(不包括正义峡以下物种)到2017年增长到75属。主要以绿藻门、硅藻门物种增加幅度较大,但上中下游物种变化各不相同。上游物种从1981年的37属,经2008年的39属,到2017年为40属,呈递增趋势,绿藻门和硅藻门物种有所增加。中游从1981年的52属降低到1997年的49属和2008年的46属,至2017年又增高到61属,呈“V”型变化趋势。期间物种变化波动较大,绿藻门、硅藻门、裸藻门经1997年减少后又增多,蓝藻门、金藻门物种数则相反,经1997年增多后又减少,呈现出动态平衡的特征。中期未出现的隐藻门物种在2017年重新出现,标志着中游水体重新恢复了“活水”的特征。下游浮游植物从1981年的43种至2017年下降为37种,呈下降趋势。
2.2 生物量和密度变化趋势
如表1所示,2017年浮游植物平均生物量0.270 mg·L-1,平均密度45.4万cells·L-1,处于较低水平,不到中国多数河流的1/4。但是生物量的变幅小、稳定性较好,最大值为0.303 mg·L-1,最小值为0.233 mg·L-1;密度最大值57.9万cells·L-1,最小值32.6万cells·L-1。其中硅藻门占生物总量76.8%,绿藻门占生物总量的12.2%,其余各门占生物总量的11.0%。在地理分布上,黑河上游生物量较低平均只有0.274 mg·L-1、平均密度为45.8万cells·L-1;中游最高,平均生物量达到0.303 mg·L-1、平均密度57.9万cells·L;黑河下游平均生物量为最少只有0.233 mg·L-1,平均密度32.6万cells·L-1。
生物量和密度变化过程与物种相似。1981年上游浮游植物平均生物量为0.217 mg·L-1,平均密度量为35.2万cells·L-1,2017年较1981年平均生物量增加了26.3%,平均密度增加了30.1%。中游1981年浮游植物平均生物量为0.290 mg·L-1,平均密度为56.3万cells·L-1,到2008年平均生物量降低13.4%,平均密度降低14.4%;到2017年平均生物量较1981年增加4.3%,平均密度增加2.8%。下游1981年浮游植物平均生物量为0.252 mg·L-1,平均密度为39.7万cells·L-1,到2017年平均生物量降低8.16%,平均密度降低21.8%。从1981—2017年的36年间来看,黑河上游浮游植物生物量和密度均呈显著的增加趋势,中游经历了逐年减小之后快速回升的变化过程,下游呈持续减小趋势。
2.3 生物多样性和优势种
黑河浮游植物Shannon-Weaver多样性指数2.21—2.87,以黑河中游2.87为最高,黑河下游2.21最低(表1)。如表2所示,黑河优势种以硅藻门的羽纹藻属(Pinnularia)、针杆藻属(Synedra)、小环藻属(Cyclotella)及绿藻门的小球藻属(Chlorella)、栅藻属(Scenedesmus)等喜浅水淡水种为主。其中,喜有机质水体的绿藻门的实球藻属(Pandorina)仅为黑河上游优势种。硅藻门的卵形藻属(Cocconeis)、双菱藻属(Surirella)、直链藻属(Melosira)、桥穹藻属(Cymbella),绿藻门的鼓藻属(Cosmarium)、衣藻属(Chlamydomonas),兰藻门的平裂藻属(Merismopedia)、拟鱼腥藻属(Anabaenopsis),金藻门的锥囊藻属(Dinobryon)及黄藻门的膝口藻属(Gonyostomum)仅为中游优势种。蓝藻门微囊藻属(Microcystis)喜高温强光有机质丰富的碱性静水水体,仅存在于黑河下游。
表2 黑河浮游植物优势种及优势度
Table 2
| 门 | 属 | 莺落峡以上 | 莺落峡至正义峡 | 正义峡以下 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1981年 | 2008年 | 2017年 | 1981年 | 1997年 | 2008年 | 2017年 | 1981年 | 2017年 | ||||
| 硅藻门 | 羽纹藻属(Pinnularia) | + | 0.22 | + | + | + | 0.07 | + | 0.11 | |||
| 针杆藻属(Synedra) | + | 0.08 | + | + | + | 0.08 | + | 0.23 | ||||
| 小环藻属(Cyclotella) | + | + | 0.24 | + | + | + | 0.20 | + | ||||
| 等片藻属(Diatoma) | + | 0.10 | + | 0.21 | ||||||||
| 卵形藻属(Cocconeis) | + | 0.03 | ||||||||||
| 双菱藻属(Surirella) | + | 0.02 | ||||||||||
| 直链藻属(Melosira) | + | 0.07 | ||||||||||
| 舟形藻属(Navicula) | + | 0.07 | 0.18 | |||||||||
| 桥穹藻属(Cymbella) | + | + | 0.05 | |||||||||
| 异端藻属(Gomphonema) | + | 0.09 | + | |||||||||
| 菱形藻属(Nitzschia) | + | + | + | 0.05 | + | |||||||
| 布纹藻属(Gyrosigma) | + | 0.07 | + | 0.04 | ||||||||
| 绿藻门 | 小球藻属(Chlorella) | + | + | 0.03 | + | + | + | 0.02 | + | |||
| 栅藻属(Scenedesmus) | + | 0.02 | + | + | 0.04 | + | 0.06 | |||||
| 绿球藻属(Chlorococcum) | + | + | + | + | ||||||||
| 纤维藻属(Ankistrodesmus) | + | + | + | 0.02 | + | 0.04 | ||||||
| 鼓藻属(Cosmarium) | + | 0.03 | ||||||||||
| 盘藻属(Gonium) | + | + | ||||||||||
| 衣藻属(Chlamydomonas) | + | 0.02 | ||||||||||
| 实球藻属(Pandorina) | + | 0.02 | ||||||||||
| 蓝藻门 | 蓝球藻属(Chroococcus) | + | 0.02 | + | ||||||||
| 颤藻属(Oscillatoria) | + | + | 0.03 | + | + | + | 0.02 | + | 0.02 | |||
| 平裂藻属(Merismopedia) | + | + | 0.03 | |||||||||
| 微囊藻属(Microcystis) | 0.03 | |||||||||||
| 拟鱼腥藻属(Anabaenopsis) | + | + | 0.02 | |||||||||
| 裸藻门 | 壳虫藻属(Trachelomonas) | + | 0.02 | |||||||||
| 裸藻属(Englena) | + | + | + | + | 0.02 | + | ||||||
| 金藻门 | 锥囊藻属(Dinobryon) | + | 0.02 | |||||||||
| 甲藻门 | 角藻属(Ceratium) | + | + | 0.03 | + | |||||||
| 黄藻门 | 膝口藻属(Gonyostomum) | + | + | |||||||||
+表示优势种。
生物多样性变化趋势与物种演变趋势一致。1981—2017年,黑河上游浮游植物多样性指数呈增大趋势,生物多样性更加丰富。黑河中游多样性呈“V”型变化趋势,1997、2008年生物多样性下降,盘藻属(Gonium)、蓝球藻属(Chroococcus)、颤藻属(Oscillatoria)、膝口藻属(Gonyostomum)等常见于浅水的水体污染指示种,代替卵形藻属(Cocconeis)、双菱藻属(Surirella)、直链藻属(Melosira),鼓藻属、锥囊藻属成为黑河中游浮游植物优势种。黑河下游生物多样性呈降低趋势,出现了喜高温、强光的微囊藻属(Microcystis)等优势种。
3 讨论
3.1 浮游植物群落结构时空变化过程
黑河浮游植物群落结构在不同时期的分布,均呈现出与河流水文特征相对应的水平地带性分异特征。在空间分布上,1981年的物种数、生物量、密度、生物多样性指数均呈现出中下游高于上游,而中游高于下游的分布特征。2008年结果显示上游除生物量高于中游外,其他参数均低于中游。通过浮游植物生物量和密度计算浮游植物平均体重,2008年上游浮游植物单体平均质量5.9×10-7 mg, 中游为5.2×10-7 mg。2008年中游来水量减少后,水体营养物质积累,浮游植物呈现个体小而密的特点。本次研究中未收集到2008年下游浮游植物数据,但2009—2010年相关研究表明黑河流域上游物种数明显高于下游,浮游植物密度上游<中游<下游,这可能与下游采样点集中在哨马营以下的居延海附近有关[15]。2017年调查结果表明各群落结构参数均为中游最高,其次是上游高于下游。在时间尺度上,黑河浮游植物物种数自1981年到2017年整体呈持续增加趋势,但上中下游物种变化各不相同。上游物种呈递增趋势,中游呈“V”型变化趋势,下游呈下降趋势。生物多样性、生物量和密度指标均体现出与物种相似的变化过程。
黑河浮游植物群落结构变化过程表明,其地理性空间分布特征与全球动植物种丰度从极地到热带地区、从山地到平原显著增加的多样性分布格局基本一致[27],但在干旱区局部河流水系中浮游植物群落对水环境要素的响应却差异巨大。温度、海拔是影响上游浮游植物分布的主要因素;水体氮磷营养盐、温度是影响黑河中游区域浮游植物群落结构分布的主要因子[15,19];下游受盐碱地影响,盐度成为影响浮游植物的主要环境因子[20]。而本文研究结果表明,营养水体的代表种类微囊藻属[28]在2017年成为下游优势种;喜肥水活水的隐藻门和喜低含量有机质、低温水体的金藻门[29]物种主要出现在中游,而上游与下游均未检出;1997、2008年中游喜营养的盘藻属出现、喜活水的隐藻消失。以上变化特征与黑河调水引起的水文特征的变化相一致,以往观点不能解释内陆河浮游植物群落结构的这种演变特征。
3.2 浮游植物群落与黑河水文特征的关联性
黑河莺落峡水文站监测结果代表了黑河上游天然来水状况,从20世纪70年代平均流量44.8 m3·s-1到90年代平均流量50.2 m3·s-1呈现出显著缓慢的增加趋势;2000年之后的17年间来水量快速增加,达到平均59.7 m3·s-1。黑河上游来水量持续增加的同时,黑河上游浮游植物生物多样性也呈现出持续升高的过程。气温普遍升高加大了黑河流域水分的蒸发,加上中游水资源开发利用程度快速提高,以及黑河流域全线调水没有充分考虑中游生态用水,导致地下水位下降,湿地萎缩[30]。正义峡水文站监测结果代表了人类活动对水量变化的影响[31-32]:从20世纪70年代平均流量33.4 m3·s-1到90年代平均24.5 m3·s-1呈现出显著快速的减少趋势[7]。2000年之后的17年间来水量快速回升到35.3 m3·s-1。黑河中游的水量变化形成1970—2000年上游多下游少的形态,2000年以后回升且下游回升快于上游。中游这种水情变化和扰动对水体水生生物产生了深远的影响。自1981年到2008年,黑河中游浮游植物群落结构衰落至最低点,物种数最低减至46属。
3.3 中游天然湿地及水域空间变化
图2
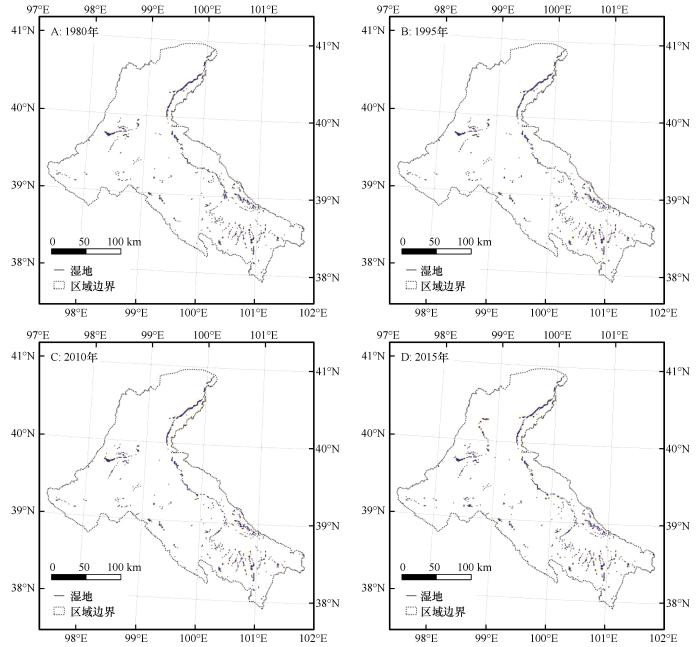
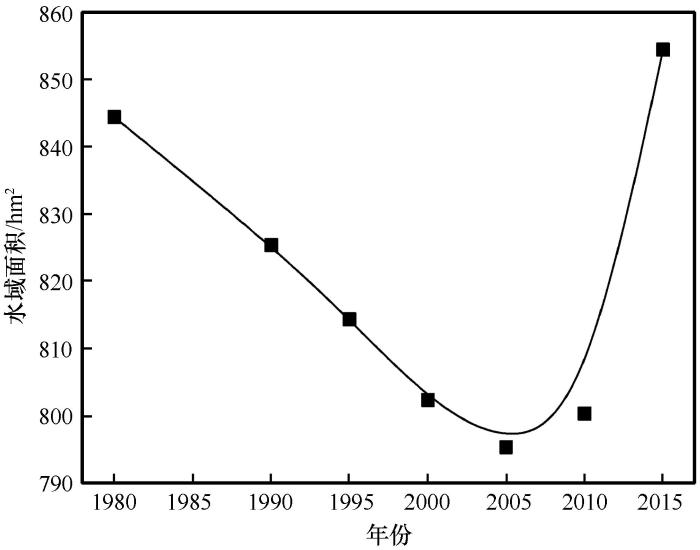
水域是湿地的重要组成部分,水域的变化直接影响水生生物生存与活动的环境动荡。从1980年以来中游水域面积呈现出显著的逐年减小到快速回升的过程(图3)。2000—2010年面积最小,之后快速回升到历史高点水平。以1980年基准面积844.4 hm²计算中游水域面积变化率,10年后的1990年减少了2.2%,20年后的2000年减小了5.0%,30年后的2010年减少了5.2%,35年后的2015年回升到历史最好水平并增加了1.2%。从结果来看仍然是维持相对未变的状况,水域面积变化与黑河流域全线调水、水利工程建设和人工湿地建设等人类活动十分密切[30,34]。相比湿地,水域面积对水文情势变化的响应更加直接、灵敏。
图3
3.4 中游浮游植物群落与水域变化关系
黑河中游浮游植物群落变化趋势与水域面积变化趋势基本一致,均呈现出“V”字型的变化过程。同样以1981年数据为基准计算,物种数和生物多样性指数到2008年分别减小9.8%和7.5%,与水域面积减少幅度很接近;到2017年物种数和生物多样性指数分别增加19.6%和2.9%,水域面积也相应增大了1.2%,物种数变幅大于生物多样性指数和水域面积的变幅(表3)。且隐藻门和金藻门主要出现在湿地保留率高且水域面积广阔的中游,而上游与下游均未检出,特别是在湿地和水域面积均达到历史最小的2000年没有检出隐藻门,这更加印证了其与水域面积的关系密切。
表3 黑河中游水域及浮游植物群落结构变化过程
Table 3
| 年份 | 水域面积及变化率 | 群落结构参数变化率/% | 生物活动力指数 | ||||
|---|---|---|---|---|---|---|---|
| 面积/hm² | 变化率/% | 物种数 | 密度 | Shannon-Weaver指数 | 生物活动力指数/(万cells·hm-2) | 变化率/% | |
| 1981 | 844.4 | — | — | — | — | 0.186 | — |
| 2008 | 800.4 | -5.2 | -9.8 | -14.4 | -7.5 | 0.155 | -16.5 |
| 2017 | 854.4 | 1.2 | 19.6 | 2.8 | 2.9 | 0.194 | 4.3 |
生物活动力指数计算中1981年水域面积采用1980年数据,2008年水域面积采用2010年数据,2017年水域面积采用2015年数据。
生物活动力的变化分析表明2008年生物活动力指数是0.155万cells·hm-²,相对1981年的0.186万cells·hm-²降低了16.5%,变幅高于生物量;2017年则回升到0.194万cells·hm-²,且相对1981年还增加了4.3%。浮游植物所处水域面积越大,活动能力越强。2008年和2017年浮游植物生物活动力指数变化率与水域面积变化率比值分别为3.2、3.6,二者十分接近,因此对干旱区内陆河,可以用水域面积来判断其浮游植物生物活动力、衡量浮游植物群落丰富性。进入21世纪实施黑河全线调水,兴建人工湿地增加水域面积,设立国家级湿地自然保护区等一系列管控和保护措施,提高了浮游植物的活动空间,提高了其获取养分的机会,浮游植物生物活动力指数明显提高,使得浮游植物群落结构快速恢复。因此用水域面积相关的生物活动力指数来衡量水生生物群落结构的活力,对于表征干旱区河流的水生态健康状况具有实质性的意义。
4 结论
以黑河为代表的西北干旱区内陆河流域,水体中的浮游植物群落结构总体处于十分脆弱的状况,生物多样性远低于黄河流域和长江流域的外流河。黑河甘肃段浮游植物最多年份也只有8门75属,其中金藻和隐藻只出现在中游且隐藻只有1属。与生物多样性自上而下沿程持续增加的外流河相比,黑河浮游植物生物多样性的变化形成了上游山区和下游都小于中游区域的特点,这与黑河中游湿地的分布密切相关。在时间尺度上,黑河上游受来水量持续增加影响,浮游植物群落变得持续丰富,水生态状况好转;而下游则呈现出衰落的态势。
黑河中游浮游植物群落结构的演变过程与水域面积的变化呈正相关。1981—2017年间,黑河中游水文过程变化强烈,直接表现为水域面积从1981年开始随着持续减小到2005年以后的快速回升形成“V”变化过程。浮游植物主要依靠适宜的水环境而生存。而水环境在水资源紧缺条件下尤为敏感,从而引发黑河中游浮游植物群落在2008年衰落至最低点后,于2017年又快速恢复到优于1981年的水平;同时,喜贫有机质的金藻仅出现在水域面积广阔的中游,且1997、2008年水域面积最低时中游出现了喜肥水的盘藻,而喜活水的隐藻消失。因此黑河调水和湿地建设对黑河水生态的保护与重建具有重要意义。
生物活动力指数从根本上反映了水资源紧缺的干旱区河流的水生态健康状况。水域面积的增加提高了水生生物的活动空间,提高了其获取养分的机会;同时干旱区河流水环境状况与水域面积紧密相关。因此,与水域面积相关的生物活动力指数更加简明有效地表达了干旱区水生生物群落结构的健康情况。生物活动力指数越高,表明水生生物的活动能力和活跃程度越强,河流生态状况越健康。
参考文献
荒漠河岸林胡杨和柽柳群落小气候特征研究
[J].
黑河流域生态-水文过程集成研究进展
[J].
The role of phytoplankton diversity metrics in shallow lake and river quality assessment
[J].
Global biodiversity,biochemical kinetics,and the energetic-equivalence rule
[J].
Trends and explanatory variables for the major phytoplankton groups of two southwestern Florida estuaries,U.S.A.
[J].
河西内陆河水系的水生生物调查
[J].

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}