C4 plant species and geographical distribution in relation to climate in the desert vegetation of China
1
2011
... C4植物水分利用效率高于C3植物,CAM(Crassulacean Acid Metabolism,景天酸代谢)植物具有最高的水分利用效率.在中国荒漠地区,CAM植物稀少,C4植物,特别是C4木本植物在荒漠生态系统中具有重要地位和作用.中国荒漠地区有C4木本植物45种(包括半木本植物),占中国荒漠植物总种数的6%,集中在藜科和蓼科,分别为19种和26种,主要分布在贺兰山以西的西北干旱荒漠区,其丰度与干旱紧密相关[1]. ...
Comparison of δ13C values and gas exchange of assimilating shoots of desert plants Haloxylon ammodendron and Calligonum mongolicum with other plants
4
2004
... 以前,人们熟知C4作物玉米、高粱、甘蔗,认为C4植物只限于草本,木本中没有C4植物.2004年Su等[2]从花环结构、δ13C值、CO2补偿点、光饱和点和最大光合速率等方面全面系统地证明了广泛分布于中国荒漠地区的木本植物梭梭和沙拐枣为C4植物.C4植物CO2同化的最初产物不是C3植物的三碳化合物3-磷酸甘油酸,而是四碳化合物草酰乙酸,故此得名. ...
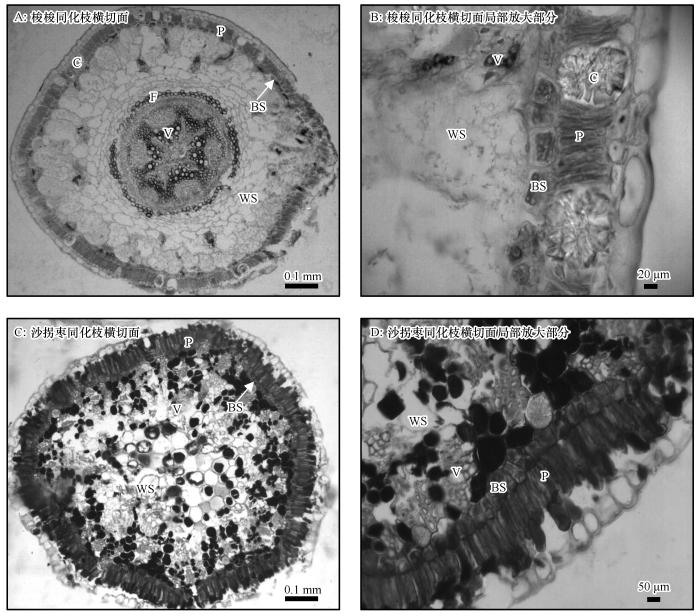
... 第一次到沙漠里,见到在相同的环境条件下,有的植物生长旺盛,植株高大;有的长势衰弱,个体矮小;为什么差异会这么大呢?后来了解到,高大灌木和类似小乔木的是梭梭,大量枯枝宿存与活枝一起构成植冠、地表根系发达的是沙拐枣.这2种植物为什么如此特殊,这激起了我的浓厚兴趣,从机理上认识它们的想法油然而生,于是将其与同域的C3植物柠条(Caragana korshinskii)和花棒(Hedysarum scoparium)进行了对比研究,得出荒漠植物梭梭和沙拐枣为典型的C4木本植物,栅栏细胞、维管束鞘细胞和维管束组成了花环结构[2,4](图1),也就是在C4草本植物叶片中发现的由叶肉细胞和维管束鞘细胞围绕维管束以同心圆形式排列组成的类似花环的结构.栅栏细胞在叶片中称为叶肉细胞,也叫光合细胞,在同化枝中形状细长排列紧密似栅栏状,故此得名.维管束鞘细胞也叫花环细胞.梭梭和沙拐枣同化枝的维管束,有小维管束和大维管束,小维管束散布在维管束鞘细胞内侧和贮水组织内,大维管束分布在同化枝中央.典型荒漠C4木本植物与C4草本植物花环结构的区别在于:一是光合器官不同,C4木本植物在同化枝,C4草本植物在叶片;二是C4木本植物的花环细胞围绕在小维管束和大维管束及贮水组织周围,而C4草本植物围绕在小维管束周围[5],但有的荒漠C4草本植物花环细胞圈内也有贮水细胞;三是C4木本植物花环结构中有许多异形细胞,如含晶细胞和黏液细胞,而C4草本植物没有或很少.这2种荒漠木本植物花环结构的揭示,使原先观测出的一些光合生理特性和生态适应特征从光合器官结构上得以诠释,改变了人们的传统认知,梭梭和沙拐枣做为典型的C4木本植物在防沙治沙和荒漠植被恢复中被广泛应用,“蚂蚁森林”项目的首选树木就是梭梭. ...
... [
2]
P:栅栏细胞;BS:维管束鞘细胞;WS:贮水组织;V:维管组织;F:纤维细胞;C:含晶细胞 ...
... P:栅栏细胞;BS:维管束鞘细胞;WS:贮水组织;V:维管组织;F:纤维细胞;C:含晶细胞
Kranz anatomy of assimilating shoots of desert plants <i>Haloxylon ammodendron</i> and <i>Calligonum mongolicum</i><sup>[<xref ref-type="bibr" rid="R2">2</xref>]</sup>Fig.1![]()
当时通过稳定碳同位素技术和光合作用生理特性观测研究得出,梭梭和沙拐枣具有C4光合特性,于是广泛查阅文献资料发现,澳大利亚的Winter[6]在20世纪80年代初就对原苏联和以色列等干旱地区分布的藜科和蓼科中的C4植物进行了研究,主要通过测定植物叶片的稳定碳同位素比率来判定是否具有C4光合途径,材料来源于干燥的植物标本和一些植物园,他认为梭梭属的梭梭、黑梭梭(Haloxylon aphyllum)、白梭梭(H. persicum)和沙拐枣属的乔木沙拐枣(Calligonum arborescens)、头状沙拐枣(C. caput-medusae)、白皮沙拐枣(C. leucocladum)等具有C4光合特征,但没有提供具有花环结构的叶片解剖结构图.Gamaley等[7]报道沙拐枣具有花环结构,沙拐枣属种类具有类似的花环结构,像猪毛菜属类型(salsoloid type).关于木本植物中有C4植物的报道,最早于1975年,Pearcy等[8]采用稳定碳同位素技术,在美国夏威夷的热带雨林中发现有C4乔木,为大戟科大戟属植物,进一步分析证明夏威夷群岛上分布的大戟属植物,从匍匐小灌木、灌木到乔木都是C4植物,它们生长在雨林和沼泽环境[8-9],有什么样的光合生理和生态适应结构,并没有展示. ...
1
2014
... 在干旱缺水和高温炎热的极端环境下,梭梭和沙拐枣为什么比其他荒漠植物生长旺盛、生物量大.21世纪初,我从参加共和国农业史上“黄淮海战役”[3]的黄河下游故道沙荒地改造利用的山东禹城片区,转战到“西部大开发”的甘肃临泽荒漠绿洲区,当时中国科学院搞知识创新工程,我主动应聘从事生态与农业研究,承担了中国科学院知识创新工程项目,即中国科学院重大项目——西部行动计划项目“黑河流域水-生态-经济系统综合管理试验示范”中游课题的一个专题“绿洲农业系统节水型种植结构调整与管理技术试验示范”,利用在黄淮海平原工作的经历和积累开展绿洲节水农业研究.以任务带学科试图认识荒漠植物的逆境适应性,从事荒漠植物生理生态基础研究. ...
荒漠植物梭梭和沙拐枣的花环结构及C4光合特征
2
2005
... 第一次到沙漠里,见到在相同的环境条件下,有的植物生长旺盛,植株高大;有的长势衰弱,个体矮小;为什么差异会这么大呢?后来了解到,高大灌木和类似小乔木的是梭梭,大量枯枝宿存与活枝一起构成植冠、地表根系发达的是沙拐枣.这2种植物为什么如此特殊,这激起了我的浓厚兴趣,从机理上认识它们的想法油然而生,于是将其与同域的C3植物柠条(Caragana korshinskii)和花棒(Hedysarum scoparium)进行了对比研究,得出荒漠植物梭梭和沙拐枣为典型的C4木本植物,栅栏细胞、维管束鞘细胞和维管束组成了花环结构[2,4](图1),也就是在C4草本植物叶片中发现的由叶肉细胞和维管束鞘细胞围绕维管束以同心圆形式排列组成的类似花环的结构.栅栏细胞在叶片中称为叶肉细胞,也叫光合细胞,在同化枝中形状细长排列紧密似栅栏状,故此得名.维管束鞘细胞也叫花环细胞.梭梭和沙拐枣同化枝的维管束,有小维管束和大维管束,小维管束散布在维管束鞘细胞内侧和贮水组织内,大维管束分布在同化枝中央.典型荒漠C4木本植物与C4草本植物花环结构的区别在于:一是光合器官不同,C4木本植物在同化枝,C4草本植物在叶片;二是C4木本植物的花环细胞围绕在小维管束和大维管束及贮水组织周围,而C4草本植物围绕在小维管束周围[5],但有的荒漠C4草本植物花环细胞圈内也有贮水细胞;三是C4木本植物花环结构中有许多异形细胞,如含晶细胞和黏液细胞,而C4草本植物没有或很少.这2种荒漠木本植物花环结构的揭示,使原先观测出的一些光合生理特性和生态适应特征从光合器官结构上得以诠释,改变了人们的传统认知,梭梭和沙拐枣做为典型的C4木本植物在防沙治沙和荒漠植被恢复中被广泛应用,“蚂蚁森林”项目的首选树木就是梭梭. ...
... 在查阅资料和回答一些疑难问题时,后来又找到当时在我单位做中国科学院“百人计划”的安黎哲教授,多次向他请教、讨论,他推荐了一些关键参考资料,帮助我对梭梭和沙拐枣的花环结构进行了详细解读和判定,进一步提高了对C4木本植物的认识[4].梭梭和沙拐枣花环结构的不同点是,梭梭具有较大的维管束,即维管组织,位于同化枝中央髓周围;还有一些小维管束,散布于贮水组织和近微管束鞘细胞处.沙拐枣同化枝大部分为小维管束.梭梭在栅栏细胞之间和贮水组织中具有大量含晶细胞,而沙拐枣栅栏细胞许多富含黏液,同时许多贮水细胞也富含黏液物(图1).梭梭和白梭梭在外形上较难区分,通过解剖结构的对比发现,梭梭的同化枝中部中央维管束外围有一圈生活的纤维细胞,白梭梭同化枝中部中央维管束外围有两大束生活的纤维细胞[9]. ...
2
2016
... 第一次到沙漠里,见到在相同的环境条件下,有的植物生长旺盛,植株高大;有的长势衰弱,个体矮小;为什么差异会这么大呢?后来了解到,高大灌木和类似小乔木的是梭梭,大量枯枝宿存与活枝一起构成植冠、地表根系发达的是沙拐枣.这2种植物为什么如此特殊,这激起了我的浓厚兴趣,从机理上认识它们的想法油然而生,于是将其与同域的C3植物柠条(Caragana korshinskii)和花棒(Hedysarum scoparium)进行了对比研究,得出荒漠植物梭梭和沙拐枣为典型的C4木本植物,栅栏细胞、维管束鞘细胞和维管束组成了花环结构[2,4](图1),也就是在C4草本植物叶片中发现的由叶肉细胞和维管束鞘细胞围绕维管束以同心圆形式排列组成的类似花环的结构.栅栏细胞在叶片中称为叶肉细胞,也叫光合细胞,在同化枝中形状细长排列紧密似栅栏状,故此得名.维管束鞘细胞也叫花环细胞.梭梭和沙拐枣同化枝的维管束,有小维管束和大维管束,小维管束散布在维管束鞘细胞内侧和贮水组织内,大维管束分布在同化枝中央.典型荒漠C4木本植物与C4草本植物花环结构的区别在于:一是光合器官不同,C4木本植物在同化枝,C4草本植物在叶片;二是C4木本植物的花环细胞围绕在小维管束和大维管束及贮水组织周围,而C4草本植物围绕在小维管束周围[5],但有的荒漠C4草本植物花环细胞圈内也有贮水细胞;三是C4木本植物花环结构中有许多异形细胞,如含晶细胞和黏液细胞,而C4草本植物没有或很少.这2种荒漠木本植物花环结构的揭示,使原先观测出的一些光合生理特性和生态适应特征从光合器官结构上得以诠释,改变了人们的传统认知,梭梭和沙拐枣做为典型的C4木本植物在防沙治沙和荒漠植被恢复中被广泛应用,“蚂蚁森林”项目的首选树木就是梭梭. ...
... 那时我具有的科研条件和知识储备使我难以单独完成解剖结构展示这一艰巨工作,我就找到西北师范大学的马瑞君教授,让她帮忙做个切片图,她做出的同化枝横切面图效果很好,彩色图片层次分明、很清晰,但当时我还是不能很好地判读,我的博士生导师刘新民研究员就推荐我请教兰州大学的王亚馥教授,跟她交流讨论掌握了很多细胞学知识,受到了很大启发.在这一过程中,查阅了许多中国学者做的关于梭梭、白梭梭和沙拐枣同化枝的解剖结构图.20世纪80年代初,与Winter同一年,中国李正理等[15]描述,白梭梭栅栏组织细胞再往里面则为一些近乎方形的细胞所组成的细胞层.赵翠仙等[16]对腾格里沙漠主要旱生植物抗旱性结构的研究表明,沙拐枣和梭梭的解剖构造别具一格,2—3层排列整齐的栅栏组织包围着贮水组织和输导组织,在栅栏层下有一层由砖形黏液细胞组成的内皮层.刘家琼[17]描述,在梭梭同化枝解剖结构中,栅栏细胞往里为一圈长方形细胞,该细胞富含树胶物质.后来,陆续有梭梭等荒漠植物解剖结构的文章发表,罗秀英等[18]对白梭梭解剖结构描述为,栅栏组织内为一层近方形的含树胶细胞,排列整齐紧密.黄振英等[19]观察到梭梭栅栏层为1—2层,梭梭中除正常维管束外,还具有小维管组织.侯彩霞等[20]在研究超旱生植物的解剖结构时,用电子显微镜观察发现,梭梭的胶质细胞中充满叶绿体,叶绿体基粒不发达,这正是维管束鞘细胞的特征,C4植物只有在维管束鞘细胞内形成淀粉,在叶肉细胞中没有淀粉[5].他们未能确定是维管束鞘细胞,沿用罗秀英等[18]的定义,将这层细胞称之为胶质细胞或第二层光合细胞.以上这些研究都不同程度观察到了梭梭、白梭梭和沙拐枣同化枝解剖结构的特殊性,并对这一细胞进行了描述.但由于缺乏其他资料佐证,未能明确这一层细胞就是维管束鞘细胞.以上文献资料的解剖结构图都不同程度能看到维管束鞘细胞和小维管束,但没有认识到它们和栅栏细胞一起组成了花环结构,是C4植物结构. ...
C4 plant of high biomass in arid regions of Asia-occurrence of C4 photosynthesis in Chenopodiaceae and Polygonaceae from the Middle East and USSR
2
1981
... 当时通过稳定碳同位素技术和光合作用生理特性观测研究得出,梭梭和沙拐枣具有C4光合特性,于是广泛查阅文献资料发现,澳大利亚的Winter[6]在20世纪80年代初就对原苏联和以色列等干旱地区分布的藜科和蓼科中的C4植物进行了研究,主要通过测定植物叶片的稳定碳同位素比率来判定是否具有C4光合途径,材料来源于干燥的植物标本和一些植物园,他认为梭梭属的梭梭、黑梭梭(Haloxylon aphyllum)、白梭梭(H. persicum)和沙拐枣属的乔木沙拐枣(Calligonum arborescens)、头状沙拐枣(C. caput-medusae)、白皮沙拐枣(C. leucocladum)等具有C4光合特征,但没有提供具有花环结构的叶片解剖结构图.Gamaley等[7]报道沙拐枣具有花环结构,沙拐枣属种类具有类似的花环结构,像猪毛菜属类型(salsoloid type).关于木本植物中有C4植物的报道,最早于1975年,Pearcy等[8]采用稳定碳同位素技术,在美国夏威夷的热带雨林中发现有C4乔木,为大戟科大戟属植物,进一步分析证明夏威夷群岛上分布的大戟属植物,从匍匐小灌木、灌木到乔木都是C4植物,它们生长在雨林和沼泽环境[8-9],有什么样的光合生理和生态适应结构,并没有展示. ...
... 中国学者1993年发表了C4光合作用植物名录[10],收录了梭梭、黑梭梭、白梭梭、乔木沙拐枣、头状沙拐枣和白皮沙拐枣等木本植物为C4植物,参考文献均为Winter[6]发表在《Oecologia》上的文章,随后研究了中国C4植物地理分布与气候环境的关系[11].中国内蒙古地区的C4植物名录中,列出梭梭、阿拉善沙拐枣(Calligonum alashanicum)和沙拐枣为C4植物[12],参考依据都是用稳定碳同位素技术得出的梭梭属和沙拐枣属的这些木本植物为C4植物,没有气体交换特征数据,也没有花环结构图片,这些木本植物的花环结构是什么样的,它和草本植物有什么不同,看不出来.公维昌等[13]在比较多枝柽柳(Tamarix ramosissima)和梭梭光合器官解剖结构时,在讨论中提到“Lyshede发现梭梭同化枝存在Kranz结构,是C4植物”.但是,查了一下Lyshede[14]的原文,研究利用嫩枝进行光合作用的Spartocytisus filipes、Spartocytisus supranubius和Genista aetnensis共3种豆科植物的同化枝解剖结构,并没有包含藜科的梭梭,引言和讨论中也没有提到梭梭. ...
The structure of plants of Trans-Altai Gobi
1
1988
... 当时通过稳定碳同位素技术和光合作用生理特性观测研究得出,梭梭和沙拐枣具有C4光合特性,于是广泛查阅文献资料发现,澳大利亚的Winter[6]在20世纪80年代初就对原苏联和以色列等干旱地区分布的藜科和蓼科中的C4植物进行了研究,主要通过测定植物叶片的稳定碳同位素比率来判定是否具有C4光合途径,材料来源于干燥的植物标本和一些植物园,他认为梭梭属的梭梭、黑梭梭(Haloxylon aphyllum)、白梭梭(H. persicum)和沙拐枣属的乔木沙拐枣(Calligonum arborescens)、头状沙拐枣(C. caput-medusae)、白皮沙拐枣(C. leucocladum)等具有C4光合特征,但没有提供具有花环结构的叶片解剖结构图.Gamaley等[7]报道沙拐枣具有花环结构,沙拐枣属种类具有类似的花环结构,像猪毛菜属类型(salsoloid type).关于木本植物中有C4植物的报道,最早于1975年,Pearcy等[8]采用稳定碳同位素技术,在美国夏威夷的热带雨林中发现有C4乔木,为大戟科大戟属植物,进一步分析证明夏威夷群岛上分布的大戟属植物,从匍匐小灌木、灌木到乔木都是C4植物,它们生长在雨林和沼泽环境[8-9],有什么样的光合生理和生态适应结构,并没有展示. ...
C4 photosynthesis in tree form Euphorbia species from Hawaiian rainforest sites
2
1975
... 当时通过稳定碳同位素技术和光合作用生理特性观测研究得出,梭梭和沙拐枣具有C4光合特性,于是广泛查阅文献资料发现,澳大利亚的Winter[6]在20世纪80年代初就对原苏联和以色列等干旱地区分布的藜科和蓼科中的C4植物进行了研究,主要通过测定植物叶片的稳定碳同位素比率来判定是否具有C4光合途径,材料来源于干燥的植物标本和一些植物园,他认为梭梭属的梭梭、黑梭梭(Haloxylon aphyllum)、白梭梭(H. persicum)和沙拐枣属的乔木沙拐枣(Calligonum arborescens)、头状沙拐枣(C. caput-medusae)、白皮沙拐枣(C. leucocladum)等具有C4光合特征,但没有提供具有花环结构的叶片解剖结构图.Gamaley等[7]报道沙拐枣具有花环结构,沙拐枣属种类具有类似的花环结构,像猪毛菜属类型(salsoloid type).关于木本植物中有C4植物的报道,最早于1975年,Pearcy等[8]采用稳定碳同位素技术,在美国夏威夷的热带雨林中发现有C4乔木,为大戟科大戟属植物,进一步分析证明夏威夷群岛上分布的大戟属植物,从匍匐小灌木、灌木到乔木都是C4植物,它们生长在雨林和沼泽环境[8-9],有什么样的光合生理和生态适应结构,并没有展示. ...
... [8-9],有什么样的光合生理和生态适应结构,并没有展示. ...
5
2019
... 当时通过稳定碳同位素技术和光合作用生理特性观测研究得出,梭梭和沙拐枣具有C4光合特性,于是广泛查阅文献资料发现,澳大利亚的Winter[6]在20世纪80年代初就对原苏联和以色列等干旱地区分布的藜科和蓼科中的C4植物进行了研究,主要通过测定植物叶片的稳定碳同位素比率来判定是否具有C4光合途径,材料来源于干燥的植物标本和一些植物园,他认为梭梭属的梭梭、黑梭梭(Haloxylon aphyllum)、白梭梭(H. persicum)和沙拐枣属的乔木沙拐枣(Calligonum arborescens)、头状沙拐枣(C. caput-medusae)、白皮沙拐枣(C. leucocladum)等具有C4光合特征,但没有提供具有花环结构的叶片解剖结构图.Gamaley等[7]报道沙拐枣具有花环结构,沙拐枣属种类具有类似的花环结构,像猪毛菜属类型(salsoloid type).关于木本植物中有C4植物的报道,最早于1975年,Pearcy等[8]采用稳定碳同位素技术,在美国夏威夷的热带雨林中发现有C4乔木,为大戟科大戟属植物,进一步分析证明夏威夷群岛上分布的大戟属植物,从匍匐小灌木、灌木到乔木都是C4植物,它们生长在雨林和沼泽环境[8-9],有什么样的光合生理和生态适应结构,并没有展示. ...
... 在查阅资料和回答一些疑难问题时,后来又找到当时在我单位做中国科学院“百人计划”的安黎哲教授,多次向他请教、讨论,他推荐了一些关键参考资料,帮助我对梭梭和沙拐枣的花环结构进行了详细解读和判定,进一步提高了对C4木本植物的认识[4].梭梭和沙拐枣花环结构的不同点是,梭梭具有较大的维管束,即维管组织,位于同化枝中央髓周围;还有一些小维管束,散布于贮水组织和近微管束鞘细胞处.沙拐枣同化枝大部分为小维管束.梭梭在栅栏细胞之间和贮水组织中具有大量含晶细胞,而沙拐枣栅栏细胞许多富含黏液,同时许多贮水细胞也富含黏液物(图1).梭梭和白梭梭在外形上较难区分,通过解剖结构的对比发现,梭梭的同化枝中部中央维管束外围有一圈生活的纤维细胞,白梭梭同化枝中部中央维管束外围有两大束生活的纤维细胞[9]. ...
... 梭梭同化枝的含晶细胞,随着枝条生长其数量也不断增多[18].后来继续研究表明,梭梭同化枝顶部与中部相比较,含晶细胞明显少.基部切片同化枝节间部分,可以看出同化枝两边对生的鳞片状退化叶,退化叶的基部并合,形成一个短鞘围绕着节间基部,退化叶中也含有晶体[9]. ...
... 梭梭含晶细胞中的物质,具有通过提高渗透压来提高植物的保水性与吸水力的作用[17].沙拐枣黏液细胞具有保水能力,从而为其周围的细胞提供一个较湿润的小环境.沙拐枣也含有少量的含晶细胞,同化枝顶部的晶体较大,中部和基部要稍小一些[9].植物的任何组织中都可观察到晶体,无论是在哪个组织中发现的晶体,大部分通常聚积在特化细胞——含晶细胞的液泡中[23].进一步结合能谱分析,并综合晶体的酸碱溶解性及硝酸银组化分析判定,梭梭、白梭梭和沙拐枣同化枝中所含晶体的主要成分为草酸钙(CaC2O4·nH2O)[24],草酸钙功能涉及钙调节和离子平衡,是一种钙调节机制[25].比较不同土壤水分条件下盆栽梭梭同化枝横截面解剖结构图时发现,在土壤湿润条件下,梭梭同化枝中含晶细胞的尺寸要比干旱条件下的大一些,并且晶体充满了含晶细胞腔,说明含晶细胞内晶体大小与水分条件存在一定联系[9].植物组织中的草酸钙主要以两种水合状态存在,即一水草酸钙(CaC2O4·H2O)和多水草酸钙(CaC2O4·(2+x)H2O)[26]. ...
... [9].植物组织中的草酸钙主要以两种水合状态存在,即一水草酸钙(CaC2O4·H2O)和多水草酸钙(CaC2O4·(2+x)H2O)[26]. ...
C4光合作用植物名录
1
1993
... 中国学者1993年发表了C4光合作用植物名录[10],收录了梭梭、黑梭梭、白梭梭、乔木沙拐枣、头状沙拐枣和白皮沙拐枣等木本植物为C4植物,参考文献均为Winter[6]发表在《Oecologia》上的文章,随后研究了中国C4植物地理分布与气候环境的关系[11].中国内蒙古地区的C4植物名录中,列出梭梭、阿拉善沙拐枣(Calligonum alashanicum)和沙拐枣为C4植物[12],参考依据都是用稳定碳同位素技术得出的梭梭属和沙拐枣属的这些木本植物为C4植物,没有气体交换特征数据,也没有花环结构图片,这些木本植物的花环结构是什么样的,它和草本植物有什么不同,看不出来.公维昌等[13]在比较多枝柽柳(Tamarix ramosissima)和梭梭光合器官解剖结构时,在讨论中提到“Lyshede发现梭梭同化枝存在Kranz结构,是C4植物”.但是,查了一下Lyshede[14]的原文,研究利用嫩枝进行光合作用的Spartocytisus filipes、Spartocytisus supranubius和Genista aetnensis共3种豆科植物的同化枝解剖结构,并没有包含藜科的梭梭,引言和讨论中也没有提到梭梭. ...
中国C4植物的地理分布与生态学研究:I.中国C4植物及其与气候环境的关系
1
1997
... 中国学者1993年发表了C4光合作用植物名录[10],收录了梭梭、黑梭梭、白梭梭、乔木沙拐枣、头状沙拐枣和白皮沙拐枣等木本植物为C4植物,参考文献均为Winter[6]发表在《Oecologia》上的文章,随后研究了中国C4植物地理分布与气候环境的关系[11].中国内蒙古地区的C4植物名录中,列出梭梭、阿拉善沙拐枣(Calligonum alashanicum)和沙拐枣为C4植物[12],参考依据都是用稳定碳同位素技术得出的梭梭属和沙拐枣属的这些木本植物为C4植物,没有气体交换特征数据,也没有花环结构图片,这些木本植物的花环结构是什么样的,它和草本植物有什么不同,看不出来.公维昌等[13]在比较多枝柽柳(Tamarix ramosissima)和梭梭光合器官解剖结构时,在讨论中提到“Lyshede发现梭梭同化枝存在Kranz结构,是C4植物”.但是,查了一下Lyshede[14]的原文,研究利用嫩枝进行光合作用的Spartocytisus filipes、Spartocytisus supranubius和Genista aetnensis共3种豆科植物的同化枝解剖结构,并没有包含藜科的梭梭,引言和讨论中也没有提到梭梭. ...
内蒙古地区的C4植物名录
1
2001
... 中国学者1993年发表了C4光合作用植物名录[10],收录了梭梭、黑梭梭、白梭梭、乔木沙拐枣、头状沙拐枣和白皮沙拐枣等木本植物为C4植物,参考文献均为Winter[6]发表在《Oecologia》上的文章,随后研究了中国C4植物地理分布与气候环境的关系[11].中国内蒙古地区的C4植物名录中,列出梭梭、阿拉善沙拐枣(Calligonum alashanicum)和沙拐枣为C4植物[12],参考依据都是用稳定碳同位素技术得出的梭梭属和沙拐枣属的这些木本植物为C4植物,没有气体交换特征数据,也没有花环结构图片,这些木本植物的花环结构是什么样的,它和草本植物有什么不同,看不出来.公维昌等[13]在比较多枝柽柳(Tamarix ramosissima)和梭梭光合器官解剖结构时,在讨论中提到“Lyshede发现梭梭同化枝存在Kranz结构,是C4植物”.但是,查了一下Lyshede[14]的原文,研究利用嫩枝进行光合作用的Spartocytisus filipes、Spartocytisus supranubius和Genista aetnensis共3种豆科植物的同化枝解剖结构,并没有包含藜科的梭梭,引言和讨论中也没有提到梭梭. ...
多枝柽柳与梭梭光合器官形态解剖结构的生态适应性
1
2011
... 中国学者1993年发表了C4光合作用植物名录[10],收录了梭梭、黑梭梭、白梭梭、乔木沙拐枣、头状沙拐枣和白皮沙拐枣等木本植物为C4植物,参考文献均为Winter[6]发表在《Oecologia》上的文章,随后研究了中国C4植物地理分布与气候环境的关系[11].中国内蒙古地区的C4植物名录中,列出梭梭、阿拉善沙拐枣(Calligonum alashanicum)和沙拐枣为C4植物[12],参考依据都是用稳定碳同位素技术得出的梭梭属和沙拐枣属的这些木本植物为C4植物,没有气体交换特征数据,也没有花环结构图片,这些木本植物的花环结构是什么样的,它和草本植物有什么不同,看不出来.公维昌等[13]在比较多枝柽柳(Tamarix ramosissima)和梭梭光合器官解剖结构时,在讨论中提到“Lyshede发现梭梭同化枝存在Kranz结构,是C4植物”.但是,查了一下Lyshede[14]的原文,研究利用嫩枝进行光合作用的Spartocytisus filipes、Spartocytisus supranubius和Genista aetnensis共3种豆科植物的同化枝解剖结构,并没有包含藜科的梭梭,引言和讨论中也没有提到梭梭. ...
Xeromorphic features of three stem assimilating in relation to their ecology
1
1979
... 中国学者1993年发表了C4光合作用植物名录[10],收录了梭梭、黑梭梭、白梭梭、乔木沙拐枣、头状沙拐枣和白皮沙拐枣等木本植物为C4植物,参考文献均为Winter[6]发表在《Oecologia》上的文章,随后研究了中国C4植物地理分布与气候环境的关系[11].中国内蒙古地区的C4植物名录中,列出梭梭、阿拉善沙拐枣(Calligonum alashanicum)和沙拐枣为C4植物[12],参考依据都是用稳定碳同位素技术得出的梭梭属和沙拐枣属的这些木本植物为C4植物,没有气体交换特征数据,也没有花环结构图片,这些木本植物的花环结构是什么样的,它和草本植物有什么不同,看不出来.公维昌等[13]在比较多枝柽柳(Tamarix ramosissima)和梭梭光合器官解剖结构时,在讨论中提到“Lyshede发现梭梭同化枝存在Kranz结构,是C4植物”.但是,查了一下Lyshede[14]的原文,研究利用嫩枝进行光合作用的Spartocytisus filipes、Spartocytisus supranubius和Genista aetnensis共3种豆科植物的同化枝解剖结构,并没有包含藜科的梭梭,引言和讨论中也没有提到梭梭. ...
我国甘肃九种旱生植物同化枝的解剖观察
1
1981
... 那时我具有的科研条件和知识储备使我难以单独完成解剖结构展示这一艰巨工作,我就找到西北师范大学的马瑞君教授,让她帮忙做个切片图,她做出的同化枝横切面图效果很好,彩色图片层次分明、很清晰,但当时我还是不能很好地判读,我的博士生导师刘新民研究员就推荐我请教兰州大学的王亚馥教授,跟她交流讨论掌握了很多细胞学知识,受到了很大启发.在这一过程中,查阅了许多中国学者做的关于梭梭、白梭梭和沙拐枣同化枝的解剖结构图.20世纪80年代初,与Winter同一年,中国李正理等[15]描述,白梭梭栅栏组织细胞再往里面则为一些近乎方形的细胞所组成的细胞层.赵翠仙等[16]对腾格里沙漠主要旱生植物抗旱性结构的研究表明,沙拐枣和梭梭的解剖构造别具一格,2—3层排列整齐的栅栏组织包围着贮水组织和输导组织,在栅栏层下有一层由砖形黏液细胞组成的内皮层.刘家琼[17]描述,在梭梭同化枝解剖结构中,栅栏细胞往里为一圈长方形细胞,该细胞富含树胶物质.后来,陆续有梭梭等荒漠植物解剖结构的文章发表,罗秀英等[18]对白梭梭解剖结构描述为,栅栏组织内为一层近方形的含树胶细胞,排列整齐紧密.黄振英等[19]观察到梭梭栅栏层为1—2层,梭梭中除正常维管束外,还具有小维管组织.侯彩霞等[20]在研究超旱生植物的解剖结构时,用电子显微镜观察发现,梭梭的胶质细胞中充满叶绿体,叶绿体基粒不发达,这正是维管束鞘细胞的特征,C4植物只有在维管束鞘细胞内形成淀粉,在叶肉细胞中没有淀粉[5].他们未能确定是维管束鞘细胞,沿用罗秀英等[18]的定义,将这层细胞称之为胶质细胞或第二层光合细胞.以上这些研究都不同程度观察到了梭梭、白梭梭和沙拐枣同化枝解剖结构的特殊性,并对这一细胞进行了描述.但由于缺乏其他资料佐证,未能明确这一层细胞就是维管束鞘细胞.以上文献资料的解剖结构图都不同程度能看到维管束鞘细胞和小维管束,但没有认识到它们和栅栏细胞一起组成了花环结构,是C4植物结构. ...
腾格里沙漠主要旱生植物旱性结构的初步研究
1
1981
... 那时我具有的科研条件和知识储备使我难以单独完成解剖结构展示这一艰巨工作,我就找到西北师范大学的马瑞君教授,让她帮忙做个切片图,她做出的同化枝横切面图效果很好,彩色图片层次分明、很清晰,但当时我还是不能很好地判读,我的博士生导师刘新民研究员就推荐我请教兰州大学的王亚馥教授,跟她交流讨论掌握了很多细胞学知识,受到了很大启发.在这一过程中,查阅了许多中国学者做的关于梭梭、白梭梭和沙拐枣同化枝的解剖结构图.20世纪80年代初,与Winter同一年,中国李正理等[15]描述,白梭梭栅栏组织细胞再往里面则为一些近乎方形的细胞所组成的细胞层.赵翠仙等[16]对腾格里沙漠主要旱生植物抗旱性结构的研究表明,沙拐枣和梭梭的解剖构造别具一格,2—3层排列整齐的栅栏组织包围着贮水组织和输导组织,在栅栏层下有一层由砖形黏液细胞组成的内皮层.刘家琼[17]描述,在梭梭同化枝解剖结构中,栅栏细胞往里为一圈长方形细胞,该细胞富含树胶物质.后来,陆续有梭梭等荒漠植物解剖结构的文章发表,罗秀英等[18]对白梭梭解剖结构描述为,栅栏组织内为一层近方形的含树胶细胞,排列整齐紧密.黄振英等[19]观察到梭梭栅栏层为1—2层,梭梭中除正常维管束外,还具有小维管组织.侯彩霞等[20]在研究超旱生植物的解剖结构时,用电子显微镜观察发现,梭梭的胶质细胞中充满叶绿体,叶绿体基粒不发达,这正是维管束鞘细胞的特征,C4植物只有在维管束鞘细胞内形成淀粉,在叶肉细胞中没有淀粉[5].他们未能确定是维管束鞘细胞,沿用罗秀英等[18]的定义,将这层细胞称之为胶质细胞或第二层光合细胞.以上这些研究都不同程度观察到了梭梭、白梭梭和沙拐枣同化枝解剖结构的特殊性,并对这一细胞进行了描述.但由于缺乏其他资料佐证,未能明确这一层细胞就是维管束鞘细胞.以上文献资料的解剖结构图都不同程度能看到维管束鞘细胞和小维管束,但没有认识到它们和栅栏细胞一起组成了花环结构,是C4植物结构. ...
我国荒漠不同生态类型植物的旱生结构
2
1982
... 那时我具有的科研条件和知识储备使我难以单独完成解剖结构展示这一艰巨工作,我就找到西北师范大学的马瑞君教授,让她帮忙做个切片图,她做出的同化枝横切面图效果很好,彩色图片层次分明、很清晰,但当时我还是不能很好地判读,我的博士生导师刘新民研究员就推荐我请教兰州大学的王亚馥教授,跟她交流讨论掌握了很多细胞学知识,受到了很大启发.在这一过程中,查阅了许多中国学者做的关于梭梭、白梭梭和沙拐枣同化枝的解剖结构图.20世纪80年代初,与Winter同一年,中国李正理等[15]描述,白梭梭栅栏组织细胞再往里面则为一些近乎方形的细胞所组成的细胞层.赵翠仙等[16]对腾格里沙漠主要旱生植物抗旱性结构的研究表明,沙拐枣和梭梭的解剖构造别具一格,2—3层排列整齐的栅栏组织包围着贮水组织和输导组织,在栅栏层下有一层由砖形黏液细胞组成的内皮层.刘家琼[17]描述,在梭梭同化枝解剖结构中,栅栏细胞往里为一圈长方形细胞,该细胞富含树胶物质.后来,陆续有梭梭等荒漠植物解剖结构的文章发表,罗秀英等[18]对白梭梭解剖结构描述为,栅栏组织内为一层近方形的含树胶细胞,排列整齐紧密.黄振英等[19]观察到梭梭栅栏层为1—2层,梭梭中除正常维管束外,还具有小维管组织.侯彩霞等[20]在研究超旱生植物的解剖结构时,用电子显微镜观察发现,梭梭的胶质细胞中充满叶绿体,叶绿体基粒不发达,这正是维管束鞘细胞的特征,C4植物只有在维管束鞘细胞内形成淀粉,在叶肉细胞中没有淀粉[5].他们未能确定是维管束鞘细胞,沿用罗秀英等[18]的定义,将这层细胞称之为胶质细胞或第二层光合细胞.以上这些研究都不同程度观察到了梭梭、白梭梭和沙拐枣同化枝解剖结构的特殊性,并对这一细胞进行了描述.但由于缺乏其他资料佐证,未能明确这一层细胞就是维管束鞘细胞.以上文献资料的解剖结构图都不同程度能看到维管束鞘细胞和小维管束,但没有认识到它们和栅栏细胞一起组成了花环结构,是C4植物结构. ...
... 梭梭含晶细胞中的物质,具有通过提高渗透压来提高植物的保水性与吸水力的作用[17].沙拐枣黏液细胞具有保水能力,从而为其周围的细胞提供一个较湿润的小环境.沙拐枣也含有少量的含晶细胞,同化枝顶部的晶体较大,中部和基部要稍小一些[9].植物的任何组织中都可观察到晶体,无论是在哪个组织中发现的晶体,大部分通常聚积在特化细胞——含晶细胞的液泡中[23].进一步结合能谱分析,并综合晶体的酸碱溶解性及硝酸银组化分析判定,梭梭、白梭梭和沙拐枣同化枝中所含晶体的主要成分为草酸钙(CaC2O4·nH2O)[24],草酸钙功能涉及钙调节和离子平衡,是一种钙调节机制[25].比较不同土壤水分条件下盆栽梭梭同化枝横截面解剖结构图时发现,在土壤湿润条件下,梭梭同化枝中含晶细胞的尺寸要比干旱条件下的大一些,并且晶体充满了含晶细胞腔,说明含晶细胞内晶体大小与水分条件存在一定联系[9].植物组织中的草酸钙主要以两种水合状态存在,即一水草酸钙(CaC2O4·H2O)和多水草酸钙(CaC2O4·(2+x)H2O)[26]. ...
新疆几种旱生植物叶(同化枝)的解剖结构观察
3
1986
... 那时我具有的科研条件和知识储备使我难以单独完成解剖结构展示这一艰巨工作,我就找到西北师范大学的马瑞君教授,让她帮忙做个切片图,她做出的同化枝横切面图效果很好,彩色图片层次分明、很清晰,但当时我还是不能很好地判读,我的博士生导师刘新民研究员就推荐我请教兰州大学的王亚馥教授,跟她交流讨论掌握了很多细胞学知识,受到了很大启发.在这一过程中,查阅了许多中国学者做的关于梭梭、白梭梭和沙拐枣同化枝的解剖结构图.20世纪80年代初,与Winter同一年,中国李正理等[15]描述,白梭梭栅栏组织细胞再往里面则为一些近乎方形的细胞所组成的细胞层.赵翠仙等[16]对腾格里沙漠主要旱生植物抗旱性结构的研究表明,沙拐枣和梭梭的解剖构造别具一格,2—3层排列整齐的栅栏组织包围着贮水组织和输导组织,在栅栏层下有一层由砖形黏液细胞组成的内皮层.刘家琼[17]描述,在梭梭同化枝解剖结构中,栅栏细胞往里为一圈长方形细胞,该细胞富含树胶物质.后来,陆续有梭梭等荒漠植物解剖结构的文章发表,罗秀英等[18]对白梭梭解剖结构描述为,栅栏组织内为一层近方形的含树胶细胞,排列整齐紧密.黄振英等[19]观察到梭梭栅栏层为1—2层,梭梭中除正常维管束外,还具有小维管组织.侯彩霞等[20]在研究超旱生植物的解剖结构时,用电子显微镜观察发现,梭梭的胶质细胞中充满叶绿体,叶绿体基粒不发达,这正是维管束鞘细胞的特征,C4植物只有在维管束鞘细胞内形成淀粉,在叶肉细胞中没有淀粉[5].他们未能确定是维管束鞘细胞,沿用罗秀英等[18]的定义,将这层细胞称之为胶质细胞或第二层光合细胞.以上这些研究都不同程度观察到了梭梭、白梭梭和沙拐枣同化枝解剖结构的特殊性,并对这一细胞进行了描述.但由于缺乏其他资料佐证,未能明确这一层细胞就是维管束鞘细胞.以上文献资料的解剖结构图都不同程度能看到维管束鞘细胞和小维管束,但没有认识到它们和栅栏细胞一起组成了花环结构,是C4植物结构. ...
... [18]的定义,将这层细胞称之为胶质细胞或第二层光合细胞.以上这些研究都不同程度观察到了梭梭、白梭梭和沙拐枣同化枝解剖结构的特殊性,并对这一细胞进行了描述.但由于缺乏其他资料佐证,未能明确这一层细胞就是维管束鞘细胞.以上文献资料的解剖结构图都不同程度能看到维管束鞘细胞和小维管束,但没有认识到它们和栅栏细胞一起组成了花环结构,是C4植物结构. ...
... 梭梭同化枝的含晶细胞,随着枝条生长其数量也不断增多[18].后来继续研究表明,梭梭同化枝顶部与中部相比较,含晶细胞明显少.基部切片同化枝节间部分,可以看出同化枝两边对生的鳞片状退化叶,退化叶的基部并合,形成一个短鞘围绕着节间基部,退化叶中也含有晶体[9]. ...
30种新疆沙生植物的结构及其对沙漠环境的适应
1
1997
... 那时我具有的科研条件和知识储备使我难以单独完成解剖结构展示这一艰巨工作,我就找到西北师范大学的马瑞君教授,让她帮忙做个切片图,她做出的同化枝横切面图效果很好,彩色图片层次分明、很清晰,但当时我还是不能很好地判读,我的博士生导师刘新民研究员就推荐我请教兰州大学的王亚馥教授,跟她交流讨论掌握了很多细胞学知识,受到了很大启发.在这一过程中,查阅了许多中国学者做的关于梭梭、白梭梭和沙拐枣同化枝的解剖结构图.20世纪80年代初,与Winter同一年,中国李正理等[15]描述,白梭梭栅栏组织细胞再往里面则为一些近乎方形的细胞所组成的细胞层.赵翠仙等[16]对腾格里沙漠主要旱生植物抗旱性结构的研究表明,沙拐枣和梭梭的解剖构造别具一格,2—3层排列整齐的栅栏组织包围着贮水组织和输导组织,在栅栏层下有一层由砖形黏液细胞组成的内皮层.刘家琼[17]描述,在梭梭同化枝解剖结构中,栅栏细胞往里为一圈长方形细胞,该细胞富含树胶物质.后来,陆续有梭梭等荒漠植物解剖结构的文章发表,罗秀英等[18]对白梭梭解剖结构描述为,栅栏组织内为一层近方形的含树胶细胞,排列整齐紧密.黄振英等[19]观察到梭梭栅栏层为1—2层,梭梭中除正常维管束外,还具有小维管组织.侯彩霞等[20]在研究超旱生植物的解剖结构时,用电子显微镜观察发现,梭梭的胶质细胞中充满叶绿体,叶绿体基粒不发达,这正是维管束鞘细胞的特征,C4植物只有在维管束鞘细胞内形成淀粉,在叶肉细胞中没有淀粉[5].他们未能确定是维管束鞘细胞,沿用罗秀英等[18]的定义,将这层细胞称之为胶质细胞或第二层光合细胞.以上这些研究都不同程度观察到了梭梭、白梭梭和沙拐枣同化枝解剖结构的特殊性,并对这一细胞进行了描述.但由于缺乏其他资料佐证,未能明确这一层细胞就是维管束鞘细胞.以上文献资料的解剖结构图都不同程度能看到维管束鞘细胞和小维管束,但没有认识到它们和栅栏细胞一起组成了花环结构,是C4植物结构. ...
水分胁迫下超旱生植物梭梭的结构变化
1
1997
... 那时我具有的科研条件和知识储备使我难以单独完成解剖结构展示这一艰巨工作,我就找到西北师范大学的马瑞君教授,让她帮忙做个切片图,她做出的同化枝横切面图效果很好,彩色图片层次分明、很清晰,但当时我还是不能很好地判读,我的博士生导师刘新民研究员就推荐我请教兰州大学的王亚馥教授,跟她交流讨论掌握了很多细胞学知识,受到了很大启发.在这一过程中,查阅了许多中国学者做的关于梭梭、白梭梭和沙拐枣同化枝的解剖结构图.20世纪80年代初,与Winter同一年,中国李正理等[15]描述,白梭梭栅栏组织细胞再往里面则为一些近乎方形的细胞所组成的细胞层.赵翠仙等[16]对腾格里沙漠主要旱生植物抗旱性结构的研究表明,沙拐枣和梭梭的解剖构造别具一格,2—3层排列整齐的栅栏组织包围着贮水组织和输导组织,在栅栏层下有一层由砖形黏液细胞组成的内皮层.刘家琼[17]描述,在梭梭同化枝解剖结构中,栅栏细胞往里为一圈长方形细胞,该细胞富含树胶物质.后来,陆续有梭梭等荒漠植物解剖结构的文章发表,罗秀英等[18]对白梭梭解剖结构描述为,栅栏组织内为一层近方形的含树胶细胞,排列整齐紧密.黄振英等[19]观察到梭梭栅栏层为1—2层,梭梭中除正常维管束外,还具有小维管组织.侯彩霞等[20]在研究超旱生植物的解剖结构时,用电子显微镜观察发现,梭梭的胶质细胞中充满叶绿体,叶绿体基粒不发达,这正是维管束鞘细胞的特征,C4植物只有在维管束鞘细胞内形成淀粉,在叶肉细胞中没有淀粉[5].他们未能确定是维管束鞘细胞,沿用罗秀英等[18]的定义,将这层细胞称之为胶质细胞或第二层光合细胞.以上这些研究都不同程度观察到了梭梭、白梭梭和沙拐枣同化枝解剖结构的特殊性,并对这一细胞进行了描述.但由于缺乏其他资料佐证,未能明确这一层细胞就是维管束鞘细胞.以上文献资料的解剖结构图都不同程度能看到维管束鞘细胞和小维管束,但没有认识到它们和栅栏细胞一起组成了花环结构,是C4植物结构. ...
Kranz anatomy is not essential for terrestrial C4 plant photosynthesis
2
2001
... 当时在我单位做中国科学院“百人计划”的张立新研究员,在我深入研究木本C4植物方面提供了帮助和关键思路,他推荐让我阅读国际顶级刊物《Nature》杂志上的一篇文章,俄罗斯科学院的Voznesenskaya等[21]通过对C3植物盘果碱蓬 (Suaeda heterophylla) 和C4植物落叶松状猪毛菜 (Salsola laricina)的比较研究,揭示了并非所有的C4植物都具有花环结构,发现了在单一光合细胞内进行C4光合作用的植物异子蓬 (Borszczowia aralocaspica),它同属于藜科,有C4植物光合特征,但并没有花环结构,这种植物的C4光合作用,是在绿色组织叶肉细胞质中,通过光合酶在空间上的分隔、两种类型叶绿体的分离和其他细胞器在不同的位置来完成的[21].在低的大气CO2浓度下C4光合作用比C3光合作用更有利,藻青菌类、藻类和一些植物对光合作用CO2限制的重要适应就是进化形成了CO2浓缩机制[22]. ...
... [21].在低的大气CO2浓度下C4光合作用比C3光合作用更有利,藻青菌类、藻类和一些植物对光合作用CO2限制的重要适应就是进化形成了CO2浓缩机制[22]. ...
1
2000
... 当时在我单位做中国科学院“百人计划”的张立新研究员,在我深入研究木本C4植物方面提供了帮助和关键思路,他推荐让我阅读国际顶级刊物《Nature》杂志上的一篇文章,俄罗斯科学院的Voznesenskaya等[21]通过对C3植物盘果碱蓬 (Suaeda heterophylla) 和C4植物落叶松状猪毛菜 (Salsola laricina)的比较研究,揭示了并非所有的C4植物都具有花环结构,发现了在单一光合细胞内进行C4光合作用的植物异子蓬 (Borszczowia aralocaspica),它同属于藜科,有C4植物光合特征,但并没有花环结构,这种植物的C4光合作用,是在绿色组织叶肉细胞质中,通过光合酶在空间上的分隔、两种类型叶绿体的分离和其他细胞器在不同的位置来完成的[21].在低的大气CO2浓度下C4光合作用比C3光合作用更有利,藻青菌类、藻类和一些植物对光合作用CO2限制的重要适应就是进化形成了CO2浓缩机制[22]. ...
植物含晶细胞的结构与功能
1
2006
... 梭梭含晶细胞中的物质,具有通过提高渗透压来提高植物的保水性与吸水力的作用[17].沙拐枣黏液细胞具有保水能力,从而为其周围的细胞提供一个较湿润的小环境.沙拐枣也含有少量的含晶细胞,同化枝顶部的晶体较大,中部和基部要稍小一些[9].植物的任何组织中都可观察到晶体,无论是在哪个组织中发现的晶体,大部分通常聚积在特化细胞——含晶细胞的液泡中[23].进一步结合能谱分析,并综合晶体的酸碱溶解性及硝酸银组化分析判定,梭梭、白梭梭和沙拐枣同化枝中所含晶体的主要成分为草酸钙(CaC2O4·nH2O)[24],草酸钙功能涉及钙调节和离子平衡,是一种钙调节机制[25].比较不同土壤水分条件下盆栽梭梭同化枝横截面解剖结构图时发现,在土壤湿润条件下,梭梭同化枝中含晶细胞的尺寸要比干旱条件下的大一些,并且晶体充满了含晶细胞腔,说明含晶细胞内晶体大小与水分条件存在一定联系[9].植物组织中的草酸钙主要以两种水合状态存在,即一水草酸钙(CaC2O4·H2O)和多水草酸钙(CaC2O4·(2+x)H2O)[26]. ...
五种C4荒漠植物光合器官中含晶细胞的比较分析
1
2008
... 梭梭含晶细胞中的物质,具有通过提高渗透压来提高植物的保水性与吸水力的作用[17].沙拐枣黏液细胞具有保水能力,从而为其周围的细胞提供一个较湿润的小环境.沙拐枣也含有少量的含晶细胞,同化枝顶部的晶体较大,中部和基部要稍小一些[9].植物的任何组织中都可观察到晶体,无论是在哪个组织中发现的晶体,大部分通常聚积在特化细胞——含晶细胞的液泡中[23].进一步结合能谱分析,并综合晶体的酸碱溶解性及硝酸银组化分析判定,梭梭、白梭梭和沙拐枣同化枝中所含晶体的主要成分为草酸钙(CaC2O4·nH2O)[24],草酸钙功能涉及钙调节和离子平衡,是一种钙调节机制[25].比较不同土壤水分条件下盆栽梭梭同化枝横截面解剖结构图时发现,在土壤湿润条件下,梭梭同化枝中含晶细胞的尺寸要比干旱条件下的大一些,并且晶体充满了含晶细胞腔,说明含晶细胞内晶体大小与水分条件存在一定联系[9].植物组织中的草酸钙主要以两种水合状态存在,即一水草酸钙(CaC2O4·H2O)和多水草酸钙(CaC2O4·(2+x)H2O)[26]. ...
The role of druse and raphide calcium oxalate crystals in tissue calcium regulation in Pistia Stratiotes leaves
1
2002
... 梭梭含晶细胞中的物质,具有通过提高渗透压来提高植物的保水性与吸水力的作用[17].沙拐枣黏液细胞具有保水能力,从而为其周围的细胞提供一个较湿润的小环境.沙拐枣也含有少量的含晶细胞,同化枝顶部的晶体较大,中部和基部要稍小一些[9].植物的任何组织中都可观察到晶体,无论是在哪个组织中发现的晶体,大部分通常聚积在特化细胞——含晶细胞的液泡中[23].进一步结合能谱分析,并综合晶体的酸碱溶解性及硝酸银组化分析判定,梭梭、白梭梭和沙拐枣同化枝中所含晶体的主要成分为草酸钙(CaC2O4·nH2O)[24],草酸钙功能涉及钙调节和离子平衡,是一种钙调节机制[25].比较不同土壤水分条件下盆栽梭梭同化枝横截面解剖结构图时发现,在土壤湿润条件下,梭梭同化枝中含晶细胞的尺寸要比干旱条件下的大一些,并且晶体充满了含晶细胞腔,说明含晶细胞内晶体大小与水分条件存在一定联系[9].植物组织中的草酸钙主要以两种水合状态存在,即一水草酸钙(CaC2O4·H2O)和多水草酸钙(CaC2O4·(2+x)H2O)[26]. ...
Crystallography of the two hydrates of crystalline calcium oxalate in plants
1
1981
... 梭梭含晶细胞中的物质,具有通过提高渗透压来提高植物的保水性与吸水力的作用[17].沙拐枣黏液细胞具有保水能力,从而为其周围的细胞提供一个较湿润的小环境.沙拐枣也含有少量的含晶细胞,同化枝顶部的晶体较大,中部和基部要稍小一些[9].植物的任何组织中都可观察到晶体,无论是在哪个组织中发现的晶体,大部分通常聚积在特化细胞——含晶细胞的液泡中[23].进一步结合能谱分析,并综合晶体的酸碱溶解性及硝酸银组化分析判定,梭梭、白梭梭和沙拐枣同化枝中所含晶体的主要成分为草酸钙(CaC2O4·nH2O)[24],草酸钙功能涉及钙调节和离子平衡,是一种钙调节机制[25].比较不同土壤水分条件下盆栽梭梭同化枝横截面解剖结构图时发现,在土壤湿润条件下,梭梭同化枝中含晶细胞的尺寸要比干旱条件下的大一些,并且晶体充满了含晶细胞腔,说明含晶细胞内晶体大小与水分条件存在一定联系[9].植物组织中的草酸钙主要以两种水合状态存在,即一水草酸钙(CaC2O4·H2O)和多水草酸钙(CaC2O4·(2+x)H2O)[26]. ...
C4 photosynthesis,atmospheric CO2,and climate
1
1997
... 植物光合途径除由遗传特性决定外,适应极端环境的进化改变也是一个很重要方面[27].环境条件可以引起C3、C4光合途径间相互转化,光合途径的改变是植物适应逆境的根本变化.Pyankov等[28]对黑梭梭和白梭梭的研究表明,这2种植物的同化枝完全具有C4光合作用特征,而种子萌发后的幼小子叶则通过C3途径进行光合作用,它们通过光合器官的改变及光合途径从C3转变为C4来适应严酷的环境.因为梭梭属植物种子缺乏胚乳,自萌发后约1个月长的时间靠子叶维持生长.可见,梭梭属植物CO2固定途径及光合器官的变化,是它们在极端荒漠条件下得以生存、生长和繁殖的重要因素. ...
Features of photosynthesis in Haloxylon species of Chenopodiaceae that are dominant plants in central Asian deserts
1
1999
... 植物光合途径除由遗传特性决定外,适应极端环境的进化改变也是一个很重要方面[27].环境条件可以引起C3、C4光合途径间相互转化,光合途径的改变是植物适应逆境的根本变化.Pyankov等[28]对黑梭梭和白梭梭的研究表明,这2种植物的同化枝完全具有C4光合作用特征,而种子萌发后的幼小子叶则通过C3途径进行光合作用,它们通过光合器官的改变及光合途径从C3转变为C4来适应严酷的环境.因为梭梭属植物种子缺乏胚乳,自萌发后约1个月长的时间靠子叶维持生长.可见,梭梭属植物CO2固定途径及光合器官的变化,是它们在极端荒漠条件下得以生存、生长和繁殖的重要因素. ...
.C4荒漠植物猪毛菜与木本猪毛菜的叶片解剖结构及光合生理特征
2
2009
... 我们也比较了同属C4草本和木本植物的环境适应差异,同一生境混生群落中,C4草本猪毛菜(Salsola collina)光合速率和蒸腾速率都明显高于C4木本猪毛菜(S. arbuscula),但木本猪毛菜的水分利用效率高于猪毛菜,特别是在中午高温强光下差别更大,木本猪毛菜可高出猪毛菜40%以上[29].比较C4木本猪毛菜和草本猪毛菜解剖结构得出,木本猪毛菜叶片具有更厚的角质层,表皮下有1层下皮细胞,其栅栏组织细胞较长,排列更紧密[29]. ...
... [29]. ...
Photosynthetic regulation of C4 desert plants Haloxylon ammodendron under drought stress
2
2007
... 平时发现,即使很少的降雨,只要空气湿度增加,梭梭树冠就明显变绿,新稍长势旺盛.对于这一现象,我们设计了试验,对成龄梭梭在典型干旱环境、雨后近地层空气湿润和人工树下挖坑浇水补充土壤水分等3种水分条件下的气体交换和叶绿素荧光特征进行观测比较,得出在空气湿润或土壤湿度提高条件下,梭梭在干旱环境下出现的光抑制现象消失[30].充分证明水分胁迫导致C4荒漠植物梭梭出现光抑制,提高空气湿度或者土壤湿度,都能避免光抑制,提高光能利用率[30].空气湿度增加并没有改善根系分布层土壤水分状况,但有利于荒漠植物生长.荒漠地区有效降水量的提法对于荒漠植物而言还需斟酌. ...
... [30].空气湿度增加并没有改善根系分布层土壤水分状况,但有利于荒漠植物生长.荒漠地区有效降水量的提法对于荒漠植物而言还需斟酌. ...
Canopy and leaf gas exchange of Haloxylon ammodendron under different soil moisture regimes
1
2010
... 土壤水分阈值和适宜含水量是荒漠植被恢复和保育措施实施的重要基础参数.土壤水分控制试验观测得出,梭梭在土壤含水量为田间持水量的50%时光合能力最强,升高或降低土壤水分,其光合能力都下降[31].作物生长不受水分限制的土壤含水量为田间持水量的70%—80%[32].可见,荒漠C4木本植物生长发育的适宜土壤含水量要明显低于绿洲作物.C3和C4荒漠植物生理需水量和总耗水量随土壤湿度增加而增加,随空气湿度减小而增加;减少土壤含水量,使植物处于水分胁迫状态,生理需水量基本不变,但土壤蒸发量和总耗水量显著减少[32]. ...
Partitioning evapotranspiration of desert plants under different water regimes in the inland Heihe River Basin,Northwestern China
2
2016
... 土壤水分阈值和适宜含水量是荒漠植被恢复和保育措施实施的重要基础参数.土壤水分控制试验观测得出,梭梭在土壤含水量为田间持水量的50%时光合能力最强,升高或降低土壤水分,其光合能力都下降[31].作物生长不受水分限制的土壤含水量为田间持水量的70%—80%[32].可见,荒漠C4木本植物生长发育的适宜土壤含水量要明显低于绿洲作物.C3和C4荒漠植物生理需水量和总耗水量随土壤湿度增加而增加,随空气湿度减小而增加;减少土壤含水量,使植物处于水分胁迫状态,生理需水量基本不变,但土壤蒸发量和总耗水量显著减少[32]. ...
... [32]. ...
梭梭根系的水分再分配特征对其生理和形态的影响
1
2017
... 利用2年生梭梭幼苗,通过分根生长装置设置不同环境条件研究得出,梭梭具有水分再分配作用,在夜间弱蒸腾条件下,根系吸收下层湿润土壤中的水分并释放到上层干燥土壤中,以缓解干旱环境下植物的水分亏缺[33].梭梭幼苗根系分布浅,只能利用降水带来的有限的浅层土壤水,而成年梭梭能最大限度地利用深层土壤水分,减少表层土壤干旱对其产生的不利影响[34]. ...
根深决定不同个体大小梭梭对夏季干旱生理响应的差异
1
2021
... 利用2年生梭梭幼苗,通过分根生长装置设置不同环境条件研究得出,梭梭具有水分再分配作用,在夜间弱蒸腾条件下,根系吸收下层湿润土壤中的水分并释放到上层干燥土壤中,以缓解干旱环境下植物的水分亏缺[33].梭梭幼苗根系分布浅,只能利用降水带来的有限的浅层土壤水,而成年梭梭能最大限度地利用深层土壤水分,减少表层土壤干旱对其产生的不利影响[34]. ...
Carbon assimilation characteristics of plants in oasis-desert ecotone and their response to CO2 enrichment
1
2004
... 大气CO2浓度升高,气候变暖背景下,不同光合途径的荒漠植物适应性如何,这对维护绿洲稳定至关重要.在河西走廊中部临泽荒漠绿洲过渡带,通过加富CO2浓度观测表明,C4植物梭梭和沙拐枣能适应高CO2浓度环境,而C3植物柠条和花棒则难以适应,会逐渐衰退以至死亡[35].花棒虽然蒸腾耗水少,但因在环境严重缺水时光合效率低而影响其对荒漠环境的适应能力[36].从CO2加富试验得出结论10年后调查,临泽荒漠绿洲过渡带的沙拐枣种群属于增长型[37]. ...
几种荒漠植物对干旱过程的生理适应性比较
1
2008
... 大气CO2浓度升高,气候变暖背景下,不同光合途径的荒漠植物适应性如何,这对维护绿洲稳定至关重要.在河西走廊中部临泽荒漠绿洲过渡带,通过加富CO2浓度观测表明,C4植物梭梭和沙拐枣能适应高CO2浓度环境,而C3植物柠条和花棒则难以适应,会逐渐衰退以至死亡[35].花棒虽然蒸腾耗水少,但因在环境严重缺水时光合效率低而影响其对荒漠环境的适应能力[36].从CO2加富试验得出结论10年后调查,临泽荒漠绿洲过渡带的沙拐枣种群属于增长型[37]. ...
荒漠绿洲过渡带沙拐枣种群结构及动态特征
1
2014
... 大气CO2浓度升高,气候变暖背景下,不同光合途径的荒漠植物适应性如何,这对维护绿洲稳定至关重要.在河西走廊中部临泽荒漠绿洲过渡带,通过加富CO2浓度观测表明,C4植物梭梭和沙拐枣能适应高CO2浓度环境,而C3植物柠条和花棒则难以适应,会逐渐衰退以至死亡[35].花棒虽然蒸腾耗水少,但因在环境严重缺水时光合效率低而影响其对荒漠环境的适应能力[36].从CO2加富试验得出结论10年后调查,临泽荒漠绿洲过渡带的沙拐枣种群属于增长型[37]. ...
Associated growth of C3 and C4 desert plants helps the C3 species at the cost of the C4 species
3
2012
... 在荒漠植物分布格局与环境关系的调查中发现,在同一戈壁生境下,C3荒漠植物红砂(Reaumuria soongorica)和C4荒漠植物珍珠(Salsola passerina)有的单生.有的联生,和珍珠联生的红砂长势要好于单生.通过对比观测研究,揭示了红砂和珍珠联生有利于红砂固有光合特性的表达和生存,但抑制珍珠生长[38],二者采取不同的生存策略[39],总体表现为联生降低蒸腾速率,有利于群落水分利用效率的提高[38].C3和C4草本植物混生较多,但C3和C4木本植物(包括半木本植物)混生,特别是联生现象少见,C3木本植物红砂和C4半木本植物珍珠联生表明,C3和C4荒漠植物联生C4植物付出代价促进C3植物生长,这是C4荒漠植物的一个重要特性[38]. ...
... [38].C3和C4草本植物混生较多,但C3和C4木本植物(包括半木本植物)混生,特别是联生现象少见,C3木本植物红砂和C4半木本植物珍珠联生表明,C3和C4荒漠植物联生C4植物付出代价促进C3植物生长,这是C4荒漠植物的一个重要特性[38]. ...
... [38]. ...
干旱程度对C3植物红砂和C4植物珍珠光合生理参数的影响
1
2012
... 在荒漠植物分布格局与环境关系的调查中发现,在同一戈壁生境下,C3荒漠植物红砂(Reaumuria soongorica)和C4荒漠植物珍珠(Salsola passerina)有的单生.有的联生,和珍珠联生的红砂长势要好于单生.通过对比观测研究,揭示了红砂和珍珠联生有利于红砂固有光合特性的表达和生存,但抑制珍珠生长[38],二者采取不同的生存策略[39],总体表现为联生降低蒸腾速率,有利于群落水分利用效率的提高[38].C3和C4草本植物混生较多,但C3和C4木本植物(包括半木本植物)混生,特别是联生现象少见,C3木本植物红砂和C4半木本植物珍珠联生表明,C3和C4荒漠植物联生C4植物付出代价促进C3植物生长,这是C4荒漠植物的一个重要特性[38]. ...
干旱区荒漠无机固碳能力及土壤碳同化途径
14
2018
... 土壤固碳被认为是一种潜在的解决气候变化的方法,但人们一直注重土壤的有机碳储存,对土壤无机固碳还没有引起足够的重视.干旱区荒漠的无机固碳,也叫非生物固碳,观点不一,至今没有引起广泛重视.荒漠地区降水稀少、蒸发量大、气候干燥,植被贫乏、环境荒凉,根据荒漠景观和颗粒组成,可将荒漠划分为三大类,即砾质荒漠(戈壁)、沙质荒漠(沙漠)和壤质荒漠[40],中国西北干旱区河西走廊中部黑河流域荒漠区0—30 cm表土层戈壁、沙漠和壤质荒漠平均土壤无机碳(SIC)密度为2.8 kg·m-2,占该层总碳的81%;相应西南高寒地区沼泽、湿草甸和干草甸平均SIC密度为1.5 kg·m-2,占该层总碳的7%;西北荒漠区和西南高寒区1 m土层SIC密度占总碳密度分别为85%和6%.可以看出,西北干旱荒漠区土壤以无机固碳为主,西南高寒区以有机固碳为主[40]. ...
... [40]. ...
... 土壤碳库包括土壤有机碳(SOC)和土壤无机碳,干旱荒漠有限的生产力、较低的土壤含水量以及特殊的土壤理化性质,如较高的土壤pH值,使得土壤碳储量表现为SOC储量相对贫乏,而SIC储量比较丰富[40].在这些干旱荒漠区,SIC的储量是SOC的2—10倍[41],且SIC的累积速率通常高于SOC[42].新疆焉耆盆地的绿洲农田SIC储量是SOC的5倍多[42],干旱土地SIC密度和储量远大于SOC.干旱荒漠区土壤中的溶解性无机碳并非来自土壤本身的碳酸盐,而是来自土壤空气或大气CO2[43]. ...
... 尽管如此,质疑声并没有平息,争论依然很激烈,2017年Schlesinger[57]通过考证非生物CO2吸收的可能机制(空气CO2分压差、碳酸盐溶解、过滤通过包气带的土壤水溶液检查淋溶),认为每一种机制都不足以解释涡度相关系统的观测结果,吸收的CO2存在哪儿,怀疑荒漠生态系统测定结果的准确性[40, 57].可见,更多的科学问题还需要深入研究. ...
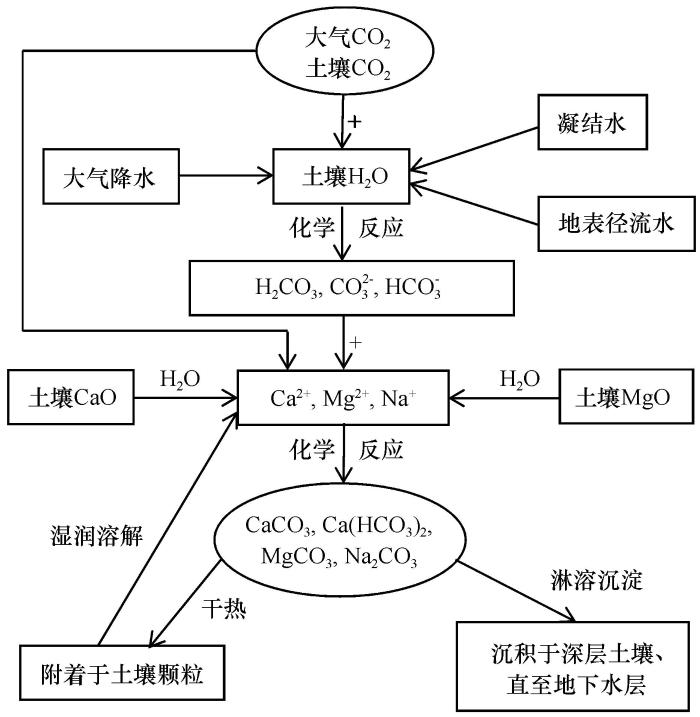
... 针对土壤对CO2的吸收和无机固定,2018年苏培玺等[40]提出了土壤碳同化(soil carbon assimilation)概念,认为土壤碳同化和植物碳同化是自然界CO2固定的两条根本途径.定义土壤碳同化是土壤将大气CO2或土壤CO2通过化学反应生成无机碳酸盐固定的过程,区别于植物通过光合作用同化碳的有机固碳能力, 将CO2通过土壤中化学反应固定成无机碳酸盐碳的能力称为无机固碳能力[40].荒漠生态系统固碳是植物碳同化与土壤碳同化,即有机和无机固碳过程的结合,中国内陆黑河流域荒漠土壤无机碳密度和碳储量是有机碳的5倍[40].苏培玺等[40]将土壤碳同化途径划分为3个阶段,一是CO2与H2O反应阶段,二是CO2或弱碳酸与土壤溶液阳离子反应阶段,三是生成溶解碳酸盐与沉淀碳酸盐附着于土壤颗粒和向下沉积阶段(图2).通过这3个紧密相连,随自身土壤环境和大气环境条件而变化的过程,大气和土壤CO2固定于土壤中,随土壤水分入渗进入地下水层. ...
... [40].荒漠生态系统固碳是植物碳同化与土壤碳同化,即有机和无机固碳过程的结合,中国内陆黑河流域荒漠土壤无机碳密度和碳储量是有机碳的5倍[40].苏培玺等[40]将土壤碳同化途径划分为3个阶段,一是CO2与H2O反应阶段,二是CO2或弱碳酸与土壤溶液阳离子反应阶段,三是生成溶解碳酸盐与沉淀碳酸盐附着于土壤颗粒和向下沉积阶段(图2).通过这3个紧密相连,随自身土壤环境和大气环境条件而变化的过程,大气和土壤CO2固定于土壤中,随土壤水分入渗进入地下水层. ...
... [40].苏培玺等[40]将土壤碳同化途径划分为3个阶段,一是CO2与H2O反应阶段,二是CO2或弱碳酸与土壤溶液阳离子反应阶段,三是生成溶解碳酸盐与沉淀碳酸盐附着于土壤颗粒和向下沉积阶段(图2).通过这3个紧密相连,随自身土壤环境和大气环境条件而变化的过程,大气和土壤CO2固定于土壤中,随土壤水分入渗进入地下水层. ...
... [40]将土壤碳同化途径划分为3个阶段,一是CO2与H2O反应阶段,二是CO2或弱碳酸与土壤溶液阳离子反应阶段,三是生成溶解碳酸盐与沉淀碳酸盐附着于土壤颗粒和向下沉积阶段(图2).通过这3个紧密相连,随自身土壤环境和大气环境条件而变化的过程,大气和土壤CO2固定于土壤中,随土壤水分入渗进入地下水层. ...
... [
40]
The pathway of soil carbon assimilation<sup>[<xref ref-type="bibr" rid="R40">40</xref>]</sup>Fig.2![]()
随后,2019年以色列Carmi等[59]在研究半干旱区森林土壤非饱和带无机碳时得出,土壤中的溶解碳酸盐中的碳来自于大气CO2,认为土壤吸收大气CO2过程是客观存在的,土壤水入渗速率与大气CO2作为无机碳的长期固存速率紧密相关.2021年意大利Laudicina等[60]在半干旱的地中海环境中研究了石膏(CaSO4·2H2O)中的Ca2+溶解,以及土壤无机碳酸盐的形成过程,区分原生和次生碳酸盐,认为石膏中的Ca2+是土壤无机碳净积累的驱动因素,有助于固定大气CO2.同一年,以色列Sagi等[61]提出了“干旱土地碳汇集(dryland carbon influx, DCI)”框架,解释了调节非生物土壤CO2吸收的因素和过程,回答了在何种条件下会产生夜间CO2流入,表明土壤CO2汇集在全世界旱地普遍存在,这是由于土壤和气候条件促进了非生物CO2交换和抑制了生物呼吸. ...
... [
40]
Fig.2![]()
随后,2019年以色列Carmi等[59]在研究半干旱区森林土壤非饱和带无机碳时得出,土壤中的溶解碳酸盐中的碳来自于大气CO2,认为土壤吸收大气CO2过程是客观存在的,土壤水入渗速率与大气CO2作为无机碳的长期固存速率紧密相关.2021年意大利Laudicina等[60]在半干旱的地中海环境中研究了石膏(CaSO4·2H2O)中的Ca2+溶解,以及土壤无机碳酸盐的形成过程,区分原生和次生碳酸盐,认为石膏中的Ca2+是土壤无机碳净积累的驱动因素,有助于固定大气CO2.同一年,以色列Sagi等[61]提出了“干旱土地碳汇集(dryland carbon influx, DCI)”框架,解释了调节非生物土壤CO2吸收的因素和过程,回答了在何种条件下会产生夜间CO2流入,表明土壤CO2汇集在全世界旱地普遍存在,这是由于土壤和气候条件促进了非生物CO2交换和抑制了生物呼吸. ...
... 荒漠土壤本身沙粒含量高,沙粒的化学成分主要为SiO2,还含有钙、镁、铝、铁等氧化物,二者反应形成的硅酸盐分解可产生Ca2+、Mg2+、Al3+、Fe2+等阳离子[40,62],或硅酸盐与CO2直接反应生成沉淀碳酸盐,化学反应式: ...
... 此外,荒漠系统外山区来水带有Ca2+、Mg2+、K+、Na+等阳离子,提高了土壤阳离子含量[63];这些阳离子具有一定的表聚性,为同化CO2气体生成碳酸盐奠定了丰富的物质基础[40],如同植物碳同化的叶绿素一样重要.土壤碳同化能力随着土壤有机碳含量、含盐量、水分、粉粒和黏粒含量的增加而提高[40],荒漠变绿洲,绿洲农业的丰产栽培管理都有利于土壤无机固碳能力的提高,这是因为农田土壤有机碳分解释放的CO2和施肥、灌溉以及地下水额外带来的Ca2+和Mg2+阳离子,为农田无机碳的形成提供了充足的碳源和钙镁源[42]. ...
... [40],荒漠变绿洲,绿洲农业的丰产栽培管理都有利于土壤无机固碳能力的提高,这是因为农田土壤有机碳分解释放的CO2和施肥、灌溉以及地下水额外带来的Ca2+和Mg2+阳离子,为农田无机碳的形成提供了充足的碳源和钙镁源[42]. ...
... 北京师范大学王秀君团队研究表明,在干旱区农田,SIC存储与SOC存储在1 m土层中呈显著正相关[64].我们团队通过对干旱区戈壁、沙漠和壤质荒漠1 m深范围内SIC和SOC密度的对比研究也得出相同的结论,即荒漠SIC储量与SOC储量呈显著正相关[40].SIC具有较长的固碳地质滞留时间,但最近的研究表明,SIC并不像以前认为的那样稳定,可以被某些微生物利用并转化为土壤中的有机物.北京林业大学张宇清团队研究发现,某些微生物采用化学自养和异养途径,即暗微生物途径,可将干旱土壤中的SIC转化为SOC[65].当然,这种转化还存在很大的未知需要探索. ...
Soil inorganic carbon stock under different soil types and land uses on the Loess Plateau region of China
1
2014
... 土壤碳库包括土壤有机碳(SOC)和土壤无机碳,干旱荒漠有限的生产力、较低的土壤含水量以及特殊的土壤理化性质,如较高的土壤pH值,使得土壤碳储量表现为SOC储量相对贫乏,而SIC储量比较丰富[40].在这些干旱荒漠区,SIC的储量是SOC的2—10倍[41],且SIC的累积速率通常高于SOC[42].新疆焉耆盆地的绿洲农田SIC储量是SOC的5倍多[42],干旱土地SIC密度和储量远大于SOC.干旱荒漠区土壤中的溶解性无机碳并非来自土壤本身的碳酸盐,而是来自土壤空气或大气CO2[43]. ...
Carbon accumulation in arid croplands of northwest China:pedogenic carbonate exceeding organic carbon
3
2015
... 土壤碳库包括土壤有机碳(SOC)和土壤无机碳,干旱荒漠有限的生产力、较低的土壤含水量以及特殊的土壤理化性质,如较高的土壤pH值,使得土壤碳储量表现为SOC储量相对贫乏,而SIC储量比较丰富[40].在这些干旱荒漠区,SIC的储量是SOC的2—10倍[41],且SIC的累积速率通常高于SOC[42].新疆焉耆盆地的绿洲农田SIC储量是SOC的5倍多[42],干旱土地SIC密度和储量远大于SOC.干旱荒漠区土壤中的溶解性无机碳并非来自土壤本身的碳酸盐,而是来自土壤空气或大气CO2[43]. ...
... [42],干旱土地SIC密度和储量远大于SOC.干旱荒漠区土壤中的溶解性无机碳并非来自土壤本身的碳酸盐,而是来自土壤空气或大气CO2[43]. ...
... 此外,荒漠系统外山区来水带有Ca2+、Mg2+、K+、Na+等阳离子,提高了土壤阳离子含量[63];这些阳离子具有一定的表聚性,为同化CO2气体生成碳酸盐奠定了丰富的物质基础[40],如同植物碳同化的叶绿素一样重要.土壤碳同化能力随着土壤有机碳含量、含盐量、水分、粉粒和黏粒含量的增加而提高[40],荒漠变绿洲,绿洲农业的丰产栽培管理都有利于土壤无机固碳能力的提高,这是因为农田土壤有机碳分解释放的CO2和施肥、灌溉以及地下水额外带来的Ca2+和Mg2+阳离子,为农田无机碳的形成提供了充足的碳源和钙镁源[42]. ...
重新被“激活”的土壤无机碳研究
2
2016
... 土壤碳库包括土壤有机碳(SOC)和土壤无机碳,干旱荒漠有限的生产力、较低的土壤含水量以及特殊的土壤理化性质,如较高的土壤pH值,使得土壤碳储量表现为SOC储量相对贫乏,而SIC储量比较丰富[40].在这些干旱荒漠区,SIC的储量是SOC的2—10倍[41],且SIC的累积速率通常高于SOC[42].新疆焉耆盆地的绿洲农田SIC储量是SOC的5倍多[42],干旱土地SIC密度和储量远大于SOC.干旱荒漠区土壤中的溶解性无机碳并非来自土壤本身的碳酸盐,而是来自土壤空气或大气CO2[43]. ...
... 中国科学院新疆生地所李彦等[43]将这一现象称为“无机呼吸”、“负呼吸”等,查阅资料发现南极寒漠地区也有这种现象[50].2008年,Stone[51]在美国《Science》杂志撰文评述指出:2005年,李彦在中国西部古尔班通古特沙漠的碳通量测量时发现CO2负通量现象,起初他认为设备出了故障,后来排除了仪器因素,在发现夜间CO2吸收现象时,他的团队排除了稀疏的植被造成CO2固定的可能性,并得出一个令人惊讶的结论,即古尔班通古特沙漠的碱性土壤以无机形式吸收了大量的CO2.21世纪初10年中国和其他国家观测到的这一负通量现象,引起了关注这一问题学者的广泛热议,掀起了土壤是否有无机碳汇能力的讨论. ...
Diurnal,seasonal and annual variation in the net ecosystem CO2 exchange of a desert shrub community (Sarcocaulescent) in Baja California,Mexico
1
2005
... 2013—2017年,我负责完成了国家重点基础研究发展计划(973)课题“碳循环-气候变化互馈作用及地球系统敏感性”,与项目负责人清华大学林光辉教授和我们课题组中国科学院植物研究所郭庆华研究员等,就碳循环领域的不确定性问题进行广泛交流,启发良多.我们利用该课题经费对在墨西哥[44]、美国[45]和中国[46]的干旱荒漠区观测到的CO2负通量现象进行不同区域观测资料的对比分析,美国内华达的莫哈韦沙漠和中国新疆的古尔班通古特沙漠,年降水量都不足200 mm,属于典型的干旱荒漠区,都观测到了荒漠生态系统吸收的碳超过100 g C·m-2·a-1[45-46].美国Hamerlynck等[47]确认荒漠区土壤表面夜间有CO2负通量时土壤亚表层中CO2分压低于大气,土壤从大气中直接吸收CO2.中国Liu等[48]用13C示踪研究显示,大气CO2可以直接进入沙漠土壤,并成为土壤颗粒中的一部分.Rey[49]分析了各种可能导致CO2吸收异常的非生物机制,认为沙漠中碳吸收可能与温度日变化或天气模式导致的土壤孔隙压力有关. ...
Large annual net ecosystem CO2 uptake of a Mojave Desert ecosystem
2
2008
... 2013—2017年,我负责完成了国家重点基础研究发展计划(973)课题“碳循环-气候变化互馈作用及地球系统敏感性”,与项目负责人清华大学林光辉教授和我们课题组中国科学院植物研究所郭庆华研究员等,就碳循环领域的不确定性问题进行广泛交流,启发良多.我们利用该课题经费对在墨西哥[44]、美国[45]和中国[46]的干旱荒漠区观测到的CO2负通量现象进行不同区域观测资料的对比分析,美国内华达的莫哈韦沙漠和中国新疆的古尔班通古特沙漠,年降水量都不足200 mm,属于典型的干旱荒漠区,都观测到了荒漠生态系统吸收的碳超过100 g C·m-2·a-1[45-46].美国Hamerlynck等[47]确认荒漠区土壤表面夜间有CO2负通量时土壤亚表层中CO2分压低于大气,土壤从大气中直接吸收CO2.中国Liu等[48]用13C示踪研究显示,大气CO2可以直接进入沙漠土壤,并成为土壤颗粒中的一部分.Rey[49]分析了各种可能导致CO2吸收异常的非生物机制,认为沙漠中碳吸收可能与温度日变化或天气模式导致的土壤孔隙压力有关. ...
... -1[45-46].美国Hamerlynck等[47]确认荒漠区土壤表面夜间有CO2负通量时土壤亚表层中CO2分压低于大气,土壤从大气中直接吸收CO2.中国Liu等[48]用13C示踪研究显示,大气CO2可以直接进入沙漠土壤,并成为土壤颗粒中的一部分.Rey[49]分析了各种可能导致CO2吸收异常的非生物机制,认为沙漠中碳吸收可能与温度日变化或天气模式导致的土壤孔隙压力有关. ...
CO2 absorption by alkaline soils and its implication to the global carbon cycle
3
2009
... 2013—2017年,我负责完成了国家重点基础研究发展计划(973)课题“碳循环-气候变化互馈作用及地球系统敏感性”,与项目负责人清华大学林光辉教授和我们课题组中国科学院植物研究所郭庆华研究员等,就碳循环领域的不确定性问题进行广泛交流,启发良多.我们利用该课题经费对在墨西哥[44]、美国[45]和中国[46]的干旱荒漠区观测到的CO2负通量现象进行不同区域观测资料的对比分析,美国内华达的莫哈韦沙漠和中国新疆的古尔班通古特沙漠,年降水量都不足200 mm,属于典型的干旱荒漠区,都观测到了荒漠生态系统吸收的碳超过100 g C·m-2·a-1[45-46].美国Hamerlynck等[47]确认荒漠区土壤表面夜间有CO2负通量时土壤亚表层中CO2分压低于大气,土壤从大气中直接吸收CO2.中国Liu等[48]用13C示踪研究显示,大气CO2可以直接进入沙漠土壤,并成为土壤颗粒中的一部分.Rey[49]分析了各种可能导致CO2吸收异常的非生物机制,认为沙漠中碳吸收可能与温度日变化或天气模式导致的土壤孔隙压力有关. ...
... -46].美国Hamerlynck等[47]确认荒漠区土壤表面夜间有CO2负通量时土壤亚表层中CO2分压低于大气,土壤从大气中直接吸收CO2.中国Liu等[48]用13C示踪研究显示,大气CO2可以直接进入沙漠土壤,并成为土壤颗粒中的一部分.Rey[49]分析了各种可能导致CO2吸收异常的非生物机制,认为沙漠中碳吸收可能与温度日变化或天气模式导致的土壤孔隙压力有关. ...
... 李彦团队利用箱式法在荒漠盐碱土上观测到了进入土壤的CO2通量,得出中国西北地区盐碱土对SIC的吸收可高达622 g·m-2·a-1,普遍高于沙漠生态系统的碳积累速率[46].土壤无机碳库研究表明,黄土高原土壤SIC在1 m土层的平均密度和储量是SOC库的两倍多[54],中国西北干旱区土壤SIC库是SOC库的2—5倍,约占全国土壤无机碳库的60%[55]. ...
Nocturnal soil CO2 uptake and its relationship to subsurface soil and ecosystem carbon fluxes in a Chihuahuan Desert shrubland
1
2013
... 2013—2017年,我负责完成了国家重点基础研究发展计划(973)课题“碳循环-气候变化互馈作用及地球系统敏感性”,与项目负责人清华大学林光辉教授和我们课题组中国科学院植物研究所郭庆华研究员等,就碳循环领域的不确定性问题进行广泛交流,启发良多.我们利用该课题经费对在墨西哥[44]、美国[45]和中国[46]的干旱荒漠区观测到的CO2负通量现象进行不同区域观测资料的对比分析,美国内华达的莫哈韦沙漠和中国新疆的古尔班通古特沙漠,年降水量都不足200 mm,属于典型的干旱荒漠区,都观测到了荒漠生态系统吸收的碳超过100 g C·m-2·a-1[45-46].美国Hamerlynck等[47]确认荒漠区土壤表面夜间有CO2负通量时土壤亚表层中CO2分压低于大气,土壤从大气中直接吸收CO2.中国Liu等[48]用13C示踪研究显示,大气CO2可以直接进入沙漠土壤,并成为土壤颗粒中的一部分.Rey[49]分析了各种可能导致CO2吸收异常的非生物机制,认为沙漠中碳吸收可能与温度日变化或天气模式导致的土壤孔隙压力有关. ...
Abiotic CO2 uptake from the atmosphere by semiarid desert soil and its partitioning into soil phases
1
2015
... 2013—2017年,我负责完成了国家重点基础研究发展计划(973)课题“碳循环-气候变化互馈作用及地球系统敏感性”,与项目负责人清华大学林光辉教授和我们课题组中国科学院植物研究所郭庆华研究员等,就碳循环领域的不确定性问题进行广泛交流,启发良多.我们利用该课题经费对在墨西哥[44]、美国[45]和中国[46]的干旱荒漠区观测到的CO2负通量现象进行不同区域观测资料的对比分析,美国内华达的莫哈韦沙漠和中国新疆的古尔班通古特沙漠,年降水量都不足200 mm,属于典型的干旱荒漠区,都观测到了荒漠生态系统吸收的碳超过100 g C·m-2·a-1[45-46].美国Hamerlynck等[47]确认荒漠区土壤表面夜间有CO2负通量时土壤亚表层中CO2分压低于大气,土壤从大气中直接吸收CO2.中国Liu等[48]用13C示踪研究显示,大气CO2可以直接进入沙漠土壤,并成为土壤颗粒中的一部分.Rey[49]分析了各种可能导致CO2吸收异常的非生物机制,认为沙漠中碳吸收可能与温度日变化或天气模式导致的土壤孔隙压力有关. ...
Mind the gap:non-biological processes contributing to soil CO2 efflux
1
2015
... 2013—2017年,我负责完成了国家重点基础研究发展计划(973)课题“碳循环-气候变化互馈作用及地球系统敏感性”,与项目负责人清华大学林光辉教授和我们课题组中国科学院植物研究所郭庆华研究员等,就碳循环领域的不确定性问题进行广泛交流,启发良多.我们利用该课题经费对在墨西哥[44]、美国[45]和中国[46]的干旱荒漠区观测到的CO2负通量现象进行不同区域观测资料的对比分析,美国内华达的莫哈韦沙漠和中国新疆的古尔班通古特沙漠,年降水量都不足200 mm,属于典型的干旱荒漠区,都观测到了荒漠生态系统吸收的碳超过100 g C·m-2·a-1[45-46].美国Hamerlynck等[47]确认荒漠区土壤表面夜间有CO2负通量时土壤亚表层中CO2分压低于大气,土壤从大气中直接吸收CO2.中国Liu等[48]用13C示踪研究显示,大气CO2可以直接进入沙漠土壤,并成为土壤颗粒中的一部分.Rey[49]分析了各种可能导致CO2吸收异常的非生物机制,认为沙漠中碳吸收可能与温度日变化或天气模式导致的土壤孔隙压力有关. ...
Abiotic processes dominate CO2 fluxes in Antarctic soils
1
2012
... 中国科学院新疆生地所李彦等[43]将这一现象称为“无机呼吸”、“负呼吸”等,查阅资料发现南极寒漠地区也有这种现象[50].2008年,Stone[51]在美国《Science》杂志撰文评述指出:2005年,李彦在中国西部古尔班通古特沙漠的碳通量测量时发现CO2负通量现象,起初他认为设备出了故障,后来排除了仪器因素,在发现夜间CO2吸收现象时,他的团队排除了稀疏的植被造成CO2固定的可能性,并得出一个令人惊讶的结论,即古尔班通古特沙漠的碱性土壤以无机形式吸收了大量的CO2.21世纪初10年中国和其他国家观测到的这一负通量现象,引起了关注这一问题学者的广泛热议,掀起了土壤是否有无机碳汇能力的讨论. ...
Have desert researchers discovered a hidden loop in the carbon cycle?
2
2008
... 中国科学院新疆生地所李彦等[43]将这一现象称为“无机呼吸”、“负呼吸”等,查阅资料发现南极寒漠地区也有这种现象[50].2008年,Stone[51]在美国《Science》杂志撰文评述指出:2005年,李彦在中国西部古尔班通古特沙漠的碳通量测量时发现CO2负通量现象,起初他认为设备出了故障,后来排除了仪器因素,在发现夜间CO2吸收现象时,他的团队排除了稀疏的植被造成CO2固定的可能性,并得出一个令人惊讶的结论,即古尔班通古特沙漠的碱性土壤以无机形式吸收了大量的CO2.21世纪初10年中国和其他国家观测到的这一负通量现象,引起了关注这一问题学者的广泛热议,掀起了土壤是否有无机碳汇能力的讨论. ...
... 在中国,每年干旱区土壤吸收大气中的碳达1.5 Tg·a-1,1 m深度SIC总存储估计在53.3—77.9 Pg[56].以美国内华达的莫哈韦沙漠的观测结果计算的全球荒漠和半干旱土壤能吸收5.2 Pg·a-1[51],这里面有对研究区域的不同理解,但荒漠和广义干旱土壤吸收无机碳的事实不断涌现. ...
1
1994
... 全球碳循环包括碳固定和碳释放2个过程,在全球碳平衡研究和估算时发现,有近20%的CO2排放去向不明,这就是全球气候变化与碳循环领域熟知的CO2失汇,即“碳黑洞”问题[52-53].干旱区盐碱土吸收CO2被认为部分揭示了碳黑洞现象[53],干旱区盐碱土在全球分布广泛,中国盐碱土从干旱区到半湿润区、从内陆到南方沿海地区均有分布,滨海湿地土壤盐渍化也很严重.中国长江以北地区土壤多数为中性和碱性,长江以南在碳酸钙母岩发育土壤和沿海地区海冲母岩情况下,土壤呈中性或微碱性,土壤中含有Na+、Ca2+、Mg2+等阳离子. ...
干旱区盐碱土的秘密:碳黑洞释义
3
2014
... 全球碳循环包括碳固定和碳释放2个过程,在全球碳平衡研究和估算时发现,有近20%的CO2排放去向不明,这就是全球气候变化与碳循环领域熟知的CO2失汇,即“碳黑洞”问题[52-53].干旱区盐碱土吸收CO2被认为部分揭示了碳黑洞现象[53],干旱区盐碱土在全球分布广泛,中国盐碱土从干旱区到半湿润区、从内陆到南方沿海地区均有分布,滨海湿地土壤盐渍化也很严重.中国长江以北地区土壤多数为中性和碱性,长江以南在碳酸钙母岩发育土壤和沿海地区海冲母岩情况下,土壤呈中性或微碱性,土壤中含有Na+、Ca2+、Mg2+等阳离子. ...
... [53],干旱区盐碱土在全球分布广泛,中国盐碱土从干旱区到半湿润区、从内陆到南方沿海地区均有分布,滨海湿地土壤盐渍化也很严重.中国长江以北地区土壤多数为中性和碱性,长江以南在碳酸钙母岩发育土壤和沿海地区海冲母岩情况下,土壤呈中性或微碱性,土壤中含有Na+、Ca2+、Mg2+等阳离子. ...
... 海洋碳汇除了高、低等植物的有机固碳外,另一个很重要的方面就是海水溶解CO2,CO2与H2O反应的产物或者CO2直接与海水中的阳离子发生反应,生成溶解或沉淀碳酸盐.土壤碳同化类似于海洋的无机碳汇,只是介质截然不同,影响因素更多,过程更复杂.平时溶解在雨水或灌溉水中的大气CO2同样可形成溶解或沉淀碳酸盐,最终外流河输送到海洋,内陆河输送到尾闾湖,漫长下渗固结最终沉积下来,通过不同的成岩过程,形成石灰岩、白云石和碳质页岩等[53]. ...
Effect of environmental factors on regional soil organic carbon stocks across the Loess Plateau region,China
1
2011
... 李彦团队利用箱式法在荒漠盐碱土上观测到了进入土壤的CO2通量,得出中国西北地区盐碱土对SIC的吸收可高达622 g·m-2·a-1,普遍高于沙漠生态系统的碳积累速率[46].土壤无机碳库研究表明,黄土高原土壤SIC在1 m土层的平均密度和储量是SOC库的两倍多[54],中国西北干旱区土壤SIC库是SOC库的2—5倍,约占全国土壤无机碳库的60%[55]. ...
Assessment of soil organic and carbonate carbon storage in China
1
2007
... 李彦团队利用箱式法在荒漠盐碱土上观测到了进入土壤的CO2通量,得出中国西北地区盐碱土对SIC的吸收可高达622 g·m-2·a-1,普遍高于沙漠生态系统的碳积累速率[46].土壤无机碳库研究表明,黄土高原土壤SIC在1 m土层的平均密度和储量是SOC库的两倍多[54],中国西北干旱区土壤SIC库是SOC库的2—5倍,约占全国土壤无机碳库的60%[55]. ...
Soil inorganic carbon storage pattern in China
1
2008
... 在中国,每年干旱区土壤吸收大气中的碳达1.5 Tg·a-1,1 m深度SIC总存储估计在53.3—77.9 Pg[56].以美国内华达的莫哈韦沙漠的观测结果计算的全球荒漠和半干旱土壤能吸收5.2 Pg·a-1[51],这里面有对研究区域的不同理解,但荒漠和广义干旱土壤吸收无机碳的事实不断涌现. ...
An evaluation of abiotic carbon sinks in deserts
2
2017
... 尽管如此,质疑声并没有平息,争论依然很激烈,2017年Schlesinger[57]通过考证非生物CO2吸收的可能机制(空气CO2分压差、碳酸盐溶解、过滤通过包气带的土壤水溶液检查淋溶),认为每一种机制都不足以解释涡度相关系统的观测结果,吸收的CO2存在哪儿,怀疑荒漠生态系统测定结果的准确性[40, 57].可见,更多的科学问题还需要深入研究. ...
... , 57].可见,更多的科学问题还需要深入研究. ...
Hidden carbon sink beneath desert
1
2015
... Li等[58]进一步研究证实了荒漠盐碱土以无机方式大量吸收CO2,并回答了吸收的CO2存在于荒漠地下咸水层,干旱区绿洲的洗盐过程是重要的碳汇过程,不是简单的无机碳迁移,荒漠地下咸水形成一个类似于海洋的碳汇.盐碱地改良洗盐过程强化了盐碱地碳汇能力,洗盐过程同时洗去了土壤中的可溶性无机碳酸盐和沉积无机碳酸盐,进入地下咸水层.在荒漠绿洲冬灌过程中,通过改变土壤胶体表面吸附性能,使更多的碳酸盐随水下渗沉积于深层土壤,直至母岩层,最终形成碳质岩石,成为地质结构的一部分. ...
Sequestration of atmospheric carbon dioxide as inorganic carbon in the unsaturated zone under semi-arid forests
1
2019
... 随后,2019年以色列Carmi等[59]在研究半干旱区森林土壤非饱和带无机碳时得出,土壤中的溶解碳酸盐中的碳来自于大气CO2,认为土壤吸收大气CO2过程是客观存在的,土壤水入渗速率与大气CO2作为无机碳的长期固存速率紧密相关.2021年意大利Laudicina等[60]在半干旱的地中海环境中研究了石膏(CaSO4·2H2O)中的Ca2+溶解,以及土壤无机碳酸盐的形成过程,区分原生和次生碳酸盐,认为石膏中的Ca2+是土壤无机碳净积累的驱动因素,有助于固定大气CO2.同一年,以色列Sagi等[61]提出了“干旱土地碳汇集(dryland carbon influx, DCI)”框架,解释了调节非生物土壤CO2吸收的因素和过程,回答了在何种条件下会产生夜间CO2流入,表明土壤CO2汇集在全世界旱地普遍存在,这是由于土壤和气候条件促进了非生物CO2交换和抑制了生物呼吸. ...
Relief and calcium from gypsum as key factors for net inorganic carbon accumulation in soils of a semiarid Mediterranean environment
1
2021
... 随后,2019年以色列Carmi等[59]在研究半干旱区森林土壤非饱和带无机碳时得出,土壤中的溶解碳酸盐中的碳来自于大气CO2,认为土壤吸收大气CO2过程是客观存在的,土壤水入渗速率与大气CO2作为无机碳的长期固存速率紧密相关.2021年意大利Laudicina等[60]在半干旱的地中海环境中研究了石膏(CaSO4·2H2O)中的Ca2+溶解,以及土壤无机碳酸盐的形成过程,区分原生和次生碳酸盐,认为石膏中的Ca2+是土壤无机碳净积累的驱动因素,有助于固定大气CO2.同一年,以色列Sagi等[61]提出了“干旱土地碳汇集(dryland carbon influx, DCI)”框架,解释了调节非生物土壤CO2吸收的因素和过程,回答了在何种条件下会产生夜间CO2流入,表明土壤CO2汇集在全世界旱地普遍存在,这是由于土壤和气候条件促进了非生物CO2交换和抑制了生物呼吸. ...
Soil CO2 influx in drylands:a conceptual framework and empirical examination
1
2021
... 随后,2019年以色列Carmi等[59]在研究半干旱区森林土壤非饱和带无机碳时得出,土壤中的溶解碳酸盐中的碳来自于大气CO2,认为土壤吸收大气CO2过程是客观存在的,土壤水入渗速率与大气CO2作为无机碳的长期固存速率紧密相关.2021年意大利Laudicina等[60]在半干旱的地中海环境中研究了石膏(CaSO4·2H2O)中的Ca2+溶解,以及土壤无机碳酸盐的形成过程,区分原生和次生碳酸盐,认为石膏中的Ca2+是土壤无机碳净积累的驱动因素,有助于固定大气CO2.同一年,以色列Sagi等[61]提出了“干旱土地碳汇集(dryland carbon influx, DCI)”框架,解释了调节非生物土壤CO2吸收的因素和过程,回答了在何种条件下会产生夜间CO2流入,表明土壤CO2汇集在全世界旱地普遍存在,这是由于土壤和气候条件促进了非生物CO2交换和抑制了生物呼吸. ...
Pertilization enhancing carbon sequestration as carbonate in arid cropland:assessments of long-term experiments in northern China
1
2014
... 荒漠土壤本身沙粒含量高,沙粒的化学成分主要为SiO2,还含有钙、镁、铝、铁等氧化物,二者反应形成的硅酸盐分解可产生Ca2+、Mg2+、Al3+、Fe2+等阳离子[40,62],或硅酸盐与CO2直接反应生成沉淀碳酸盐,化学反应式: ...
Carbon sequestration in dryland ecosystems
1
2004
... 此外,荒漠系统外山区来水带有Ca2+、Mg2+、K+、Na+等阳离子,提高了土壤阳离子含量[63];这些阳离子具有一定的表聚性,为同化CO2气体生成碳酸盐奠定了丰富的物质基础[40],如同植物碳同化的叶绿素一样重要.土壤碳同化能力随着土壤有机碳含量、含盐量、水分、粉粒和黏粒含量的增加而提高[40],荒漠变绿洲,绿洲农业的丰产栽培管理都有利于土壤无机固碳能力的提高,这是因为农田土壤有机碳分解释放的CO2和施肥、灌溉以及地下水额外带来的Ca2+和Mg2+阳离子,为农田无机碳的形成提供了充足的碳源和钙镁源[42]. ...
Relationship between soil inorganic carbon and organic carbon in the wheat-maize cropland of the North China Plain
1
2017
... 北京师范大学王秀君团队研究表明,在干旱区农田,SIC存储与SOC存储在1 m土层中呈显著正相关[64].我们团队通过对干旱区戈壁、沙漠和壤质荒漠1 m深范围内SIC和SOC密度的对比研究也得出相同的结论,即荒漠SIC储量与SOC储量呈显著正相关[40].SIC具有较长的固碳地质滞留时间,但最近的研究表明,SIC并不像以前认为的那样稳定,可以被某些微生物利用并转化为土壤中的有机物.北京林业大学张宇清团队研究发现,某些微生物采用化学自养和异养途径,即暗微生物途径,可将干旱土壤中的SIC转化为SOC[65].当然,这种转化还存在很大的未知需要探索. ...
Soil microbes transform inorganic carbon into organic carbon by dark fixation pathways in desert soil
1
2021
... 北京师范大学王秀君团队研究表明,在干旱区农田,SIC存储与SOC存储在1 m土层中呈显著正相关[64].我们团队通过对干旱区戈壁、沙漠和壤质荒漠1 m深范围内SIC和SOC密度的对比研究也得出相同的结论,即荒漠SIC储量与SOC储量呈显著正相关[40].SIC具有较长的固碳地质滞留时间,但最近的研究表明,SIC并不像以前认为的那样稳定,可以被某些微生物利用并转化为土壤中的有机物.北京林业大学张宇清团队研究发现,某些微生物采用化学自养和异养途径,即暗微生物途径,可将干旱土壤中的SIC转化为SOC[65].当然,这种转化还存在很大的未知需要探索. ...
Decoupling of soil nutrient cycles as a function of aridity in global drylands
1
2013
... 干旱区(干旱、半干旱和半湿润易旱地区)面积占全球陆地面积的40%以上[66-67],中国广义干旱区面积约占国土面积的50%,干旱区分布着大面积的荒漠和盐碱土.在未来气候变暖条件下, 干旱区降水有增加的趋势[68],相应地土壤无机固碳能力将增加,荒漠生态系统碳汇能力将增强.干旱区灌溉农业的发展, 有利于干旱区生产功能和生态功能的协同提高和发展. ...
Groundwater-dependent ecosystems:recent insights from satellite and field-based studies
1
2015
... 干旱区(干旱、半干旱和半湿润易旱地区)面积占全球陆地面积的40%以上[66-67],中国广义干旱区面积约占国土面积的50%,干旱区分布着大面积的荒漠和盐碱土.在未来气候变暖条件下, 干旱区降水有增加的趋势[68],相应地土壤无机固碳能力将增加,荒漠生态系统碳汇能力将增强.干旱区灌溉农业的发展, 有利于干旱区生产功能和生态功能的协同提高和发展. ...
1
2003
... 干旱区(干旱、半干旱和半湿润易旱地区)面积占全球陆地面积的40%以上[66-67],中国广义干旱区面积约占国土面积的50%,干旱区分布着大面积的荒漠和盐碱土.在未来气候变暖条件下, 干旱区降水有增加的趋势[68],相应地土壤无机固碳能力将增加,荒漠生态系统碳汇能力将增强.干旱区灌溉农业的发展, 有利于干旱区生产功能和生态功能的协同提高和发展. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}