荒漠土壤微生物群落结构特征研究进展
2
2018
... 荒漠生态系统是干旱、半干旱区以荒漠植物为主的生态系统,主要由耐旱和超旱生的乔木、灌木以及草本植物组成,约占全球陆地面积的三分之一[1].荒漠生态系统通常受水分和养分资源限制,植被稀疏,表现出明显的斑块状格局,是陆地生态系统中最为脆弱的系统之一[2].荒漠中除天然植被外,人工植被作为重要生态修复措施对荒漠生态系统稳定性的维持同样至关重要.然而在极端恶劣的环境中,荒漠却拥有大量独特的微生物资源,它们作为荒漠生态系统重要组成部分,参与了重要的生态过程,对稳定生态系统、完善生态系统功能有重要意义[1]. ...
... [1]. ...
Ecological stoichiometry:from sea to lake to land
2
2000
... 荒漠生态系统是干旱、半干旱区以荒漠植物为主的生态系统,主要由耐旱和超旱生的乔木、灌木以及草本植物组成,约占全球陆地面积的三分之一[1].荒漠生态系统通常受水分和养分资源限制,植被稀疏,表现出明显的斑块状格局,是陆地生态系统中最为脆弱的系统之一[2].荒漠中除天然植被外,人工植被作为重要生态修复措施对荒漠生态系统稳定性的维持同样至关重要.然而在极端恶劣的环境中,荒漠却拥有大量独特的微生物资源,它们作为荒漠生态系统重要组成部分,参与了重要的生态过程,对稳定生态系统、完善生态系统功能有重要意义[1]. ...
... 生态化学计量学是通过研究元素比率来揭示生态过程和作用的学科,为研究生态系统内各种化学元素平衡关系和能量流动以及生态系统结构与功能提供了一种重要方法[2].其主要研究内容是组成生物有机体元素含量及其比值的变化,其中对碳(C)、氮(N)、磷(P)含量及计量比的分析是研究的重点[3].由于生态系统内部的C、N、P在植物、凋落物与土壤之间相互转换,生物有机体的生态化学计量特征不仅受自身的结构限制和代谢需求的影响,同时受环境因素影响,往往表现出一定的环境可塑性[4].因此,研究C、N、P的计量关系,分析比较生态系统不同组分中C∶N∶P的变化,能够有效地反映有机体的行为特性及与生态系统间的相互作用.这对揭示元素生物地球化学循环规律、生产力形成机制以及生态系统稳定性维持机制等具有重要意义[5-6].对水分和养分同时限制的荒漠生态系统而言,开展植物、凋落物、土壤、土壤微生物和土壤酶的生态化学计量研究有望为判断其限制性元素、土壤营养的供给能力、养分的有效性及C、N、P对环境变化的适应和反馈调节能力提供重要科学依据. ...
毛乌素沙地典型植物叶片生态化学计量特征
2
2016
... 生态化学计量学是通过研究元素比率来揭示生态过程和作用的学科,为研究生态系统内各种化学元素平衡关系和能量流动以及生态系统结构与功能提供了一种重要方法[2].其主要研究内容是组成生物有机体元素含量及其比值的变化,其中对碳(C)、氮(N)、磷(P)含量及计量比的分析是研究的重点[3].由于生态系统内部的C、N、P在植物、凋落物与土壤之间相互转换,生物有机体的生态化学计量特征不仅受自身的结构限制和代谢需求的影响,同时受环境因素影响,往往表现出一定的环境可塑性[4].因此,研究C、N、P的计量关系,分析比较生态系统不同组分中C∶N∶P的变化,能够有效地反映有机体的行为特性及与生态系统间的相互作用.这对揭示元素生物地球化学循环规律、生产力形成机制以及生态系统稳定性维持机制等具有重要意义[5-6].对水分和养分同时限制的荒漠生态系统而言,开展植物、凋落物、土壤、土壤微生物和土壤酶的生态化学计量研究有望为判断其限制性元素、土壤营养的供给能力、养分的有效性及C、N、P对环境变化的适应和反馈调节能力提供重要科学依据. ...
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
The C∶N∶P stoichiometry of organisms and ecosystems in a changing world:a review and perspectives
1
2012
... 生态化学计量学是通过研究元素比率来揭示生态过程和作用的学科,为研究生态系统内各种化学元素平衡关系和能量流动以及生态系统结构与功能提供了一种重要方法[2].其主要研究内容是组成生物有机体元素含量及其比值的变化,其中对碳(C)、氮(N)、磷(P)含量及计量比的分析是研究的重点[3].由于生态系统内部的C、N、P在植物、凋落物与土壤之间相互转换,生物有机体的生态化学计量特征不仅受自身的结构限制和代谢需求的影响,同时受环境因素影响,往往表现出一定的环境可塑性[4].因此,研究C、N、P的计量关系,分析比较生态系统不同组分中C∶N∶P的变化,能够有效地反映有机体的行为特性及与生态系统间的相互作用.这对揭示元素生物地球化学循环规律、生产力形成机制以及生态系统稳定性维持机制等具有重要意义[5-6].对水分和养分同时限制的荒漠生态系统而言,开展植物、凋落物、土壤、土壤微生物和土壤酶的生态化学计量研究有望为判断其限制性元素、土壤营养的供给能力、养分的有效性及C、N、P对环境变化的适应和反馈调节能力提供重要科学依据. ...
Terrestrial phosphorus limitation:mechanisms,implications,and nitrogen-phosphorus interactions
1
2010
... 生态化学计量学是通过研究元素比率来揭示生态过程和作用的学科,为研究生态系统内各种化学元素平衡关系和能量流动以及生态系统结构与功能提供了一种重要方法[2].其主要研究内容是组成生物有机体元素含量及其比值的变化,其中对碳(C)、氮(N)、磷(P)含量及计量比的分析是研究的重点[3].由于生态系统内部的C、N、P在植物、凋落物与土壤之间相互转换,生物有机体的生态化学计量特征不仅受自身的结构限制和代谢需求的影响,同时受环境因素影响,往往表现出一定的环境可塑性[4].因此,研究C、N、P的计量关系,分析比较生态系统不同组分中C∶N∶P的变化,能够有效地反映有机体的行为特性及与生态系统间的相互作用.这对揭示元素生物地球化学循环规律、生产力形成机制以及生态系统稳定性维持机制等具有重要意义[5-6].对水分和养分同时限制的荒漠生态系统而言,开展植物、凋落物、土壤、土壤微生物和土壤酶的生态化学计量研究有望为判断其限制性元素、土壤营养的供给能力、养分的有效性及C、N、P对环境变化的适应和反馈调节能力提供重要科学依据. ...
Microbial stoichiometry overrides biomass as a regulator of soil carbon and nitrogen cycling
2
2016
... 生态化学计量学是通过研究元素比率来揭示生态过程和作用的学科,为研究生态系统内各种化学元素平衡关系和能量流动以及生态系统结构与功能提供了一种重要方法[2].其主要研究内容是组成生物有机体元素含量及其比值的变化,其中对碳(C)、氮(N)、磷(P)含量及计量比的分析是研究的重点[3].由于生态系统内部的C、N、P在植物、凋落物与土壤之间相互转换,生物有机体的生态化学计量特征不仅受自身的结构限制和代谢需求的影响,同时受环境因素影响,往往表现出一定的环境可塑性[4].因此,研究C、N、P的计量关系,分析比较生态系统不同组分中C∶N∶P的变化,能够有效地反映有机体的行为特性及与生态系统间的相互作用.这对揭示元素生物地球化学循环规律、生产力形成机制以及生态系统稳定性维持机制等具有重要意义[5-6].对水分和养分同时限制的荒漠生态系统而言,开展植物、凋落物、土壤、土壤微生物和土壤酶的生态化学计量研究有望为判断其限制性元素、土壤营养的供给能力、养分的有效性及C、N、P对环境变化的适应和反馈调节能力提供重要科学依据. ...
... 研究发现,全球森林生态系统植物叶片C∶N∶P为1 212∶28∶1,而当植物叶片凋亡后,养分的再吸收过程使得枯叶的C∶N∶P变为3 007∶45∶1[60];植物凋亡后根的C∶N∶P平均值为4 184∶43∶1[61];土壤C∶N∶P最低为287∶17∶1[62];土壤微生物量C∶N∶P平均值为42∶6∶1,并随土壤深度增加显著减小[26];Sinsabaugh等[57]发现土壤酶中C∶N∶P接近于1∶1∶1.相比之下,从植物叶片、凋落物到土壤再到土壤微生物及酶的计量特征C、N元素整体处于减少趋势.Xu等[37]研究发现,在区域尺度上,随着生物区系不同,C∶N∶P存在很大变异性,如天然湿地中的C∶N∶P为131∶14∶1,而沙漠中的C∶N∶P则为31∶4∶1.面对底物和微生物化学计量间的巨大差异,微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例和元素利用效率等机制来响应和调节.目前为止,关于植物-土壤-微生物-酶C、N、P关系的研究多聚焦于探讨土壤微生物与土壤、植被间的变化及其影响因素.研究表明,随着纬度、年均温、年降水量的变化,微生物量N∶P与植物组织的N∶P呈现出一致的变化趋势,即低纬度地区更受P的限制[63].这表明在高纬度地区微生物和植物一样由于生长季短而需要更高的生长速率,进而需要高的N、P含量和低的N∶P维持[6].Li等[64]分析了中国亚热带地区土壤数据后认为,微生物量C∶N和C∶P会随土壤C∶N和C∶P的增加而增加;周正虎等[38]却发现中国森林土壤微生物量C∶N随着土壤C∶N的增加而减小,与Cleveland等[65]对全球森林的数据分析结果一致.近期,吕星宇等[35]在腾格里沙漠开展的人工固沙植被研究表明,固沙植被演替过程中,微生物功能群数量C∶N∶P的变化主要受到草本、灌木和土壤C∶N∶P变化的影响.受灌木C∶N∶P增加及草本C∶N∶P减小的影响,微生物功能群数量C∶N∶P随之增加;对土壤酶C∶N∶P影响最大的因子分别是微生物功能群数量C∶N∶P和凋落物C∶N∶P,土壤酶C∶N∶P随着微生物功能群数量C∶N∶P增加而增加. ...
Dynamics of soil microbial C∶N∶P stoichiometry and its driving mechanisms following natural vegetation restoration after farmland abandonment
1
2019
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
N∶P ratios in terrestrial plants:variation and functional significance
1
2004
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
Soil and plant components ecological stoichiometry in four steppe communities in the Loess Plateau of China
2
2016
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
... 目前,绝大多数有关微生物C∶N∶P的研究多聚焦于MBC∶MBN∶MBP及其与土壤C∶N∶P之间的关系.全球尺度上的MBC∶MBN∶MBP平均值为42∶6∶1[37],不同的植被类型间差异明显[9].如沙漠土壤中MBC∶MBN∶MBP仅为31∶4∶1,而在天然湿地中则高达131∶14∶1[37].土壤MBC、MBN、MBP与土壤C、N、P呈显著正相关,MBC随土壤C∶N的增加而减小,而MBN却随土壤C∶N的增加而增加[38].张义凡等[39]以宁夏盐池荒漠草原的柠条、沙蒿、短花针茅典型群落为研究对象,分析了不同生境(冠下、丛间)和不同土层间MBC、MBN的变化特征,发现3种群落土壤微生物量变化差异较大,且MBC和MBN的含量均表现为冠下>丛间,并随土层深度的增加而降低,土壤C∶N与MBC∶MBN显著正相关.这表明土壤MBC、MBN具有一定的生物学指示特性,可以作为评价土壤质量的生物学指标. ...
Temporal variation in sediment C,N,and P stoichiometry in a plateau lake during sediment burial
1
2019
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
Stoichiometric N∶P flexibility and mycorrhizal symbiosis favour plant resistance against drought
1
2017
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
Drought effect on plant nitrogen and phosphorus:a meta-analysis
1
2014
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
Leaf element concentrations of terrestrial plants across China are influenced by taxonomy and the environment
1
2012
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
Global-scale patterns of nutrient resorption associated with latitude,temperature and precipitation
1
2009
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
Nutrient levels within leaves,stems,and roots of the xeric species Reaumuria soongoricain relation to geographical,climatic,and soil conditions
1
2015
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China
1
2005
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
Scaling of C∶N∶P stoichiometry in forests worldwide:applications of terrestrial redfield-type ratios
1
2004
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
Linkages of plant and soil C∶N∶P stoichiometry and their relationships to forest growth in subtropical plantations
1
2015
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
The role of plants in the effects of global change on nutrient availability and stoichiometry in the plant-soil system
1
2012
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
Effects of drought on nutrient uptake and assimilation in vegetable crops
1
2012
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
古尔班通古特沙漠4种草本植物叶片与土壤的化学计量特征
1
2015
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
植物化学计量学:一个方兴未艾的生态学研究方向
1
2018
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
Linking microbial C∶N∶P stoichiometry to microbial community and abiotic factors along a 3500-km grassland transect on the Tibetan Plateau
1
2016
... C、N、P是植物生长所必须的元素,植物对N、P的吸收和对C的同化作用及凋落物的分解等驱动着生物地球化学循环[7].植物C、N、P的生态化学计量组成关系影响许多植物的生理和生态系统过程,C∶N∶P也可以反映植物潜在的生化分配和生活史对策[8].因此,研究荒漠生态系统中植物和凋落物C∶N∶P特征与生态因子的关系、对气候变化的响应与适应能力,能够有效分析土壤养分供应潜力与评估陆地生态系统生产力.对这些问题的探讨有助于丰富中国荒漠生态系统植物生态化学计量学领域的研究.陆地生态系统中的植物通过光合作用和生物固氮从大气中固定C和N并从土壤中汲取P,同时以凋落物、根系及分泌物的形式将其归还于土壤.目前,有关植物C∶N∶P的研究已经在全球、区域、生态系统、群落及植物个体尺度上开展了较为深入的研究[9-10].众多研究表明,植物C∶N∶P与气候、物种、土壤水分、土壤养分等因子密切相关[11].在全球尺度上,干旱胁迫通常减少植物N、P含量,但增加植物N∶P,同时土壤速效氮的增加和土壤P的减少可能导致植物N∶P增加[12].陆地生态系统植物叶片的N、P含量与纬度正相关,但不同的生活型及物种的C∶N∶P差异显著[13].植物凋落物C∶N∶P平均值约为3 000∶46∶1[14],并随生物区系、生态系统以及植物生活型而变化,如荒漠草本植物的N∶P明显低于灌木[15].在区域尺度上,Han等[16]发现中国境内753种陆生植物叶片N∶P与纬度的相关性不显著,这可能与土壤本底元素的异质性较大有关.McGroddy等[17]发现森林生态系统中55种植物的凋落物C∶N∶P为3 007∶45∶1,叶片C∶N∶P为1 212∶28∶1,凋落物和叶片C∶N∶P存在较大的变化,叶片N∶P随纬度升高显著降低.对长白山森林不同演替阶段的典型群落优势种植物元素含量研究发现,随演替阶段的进展植物叶片N含量明显增加,而P含量减少,演替前期群落倾向于受N限制,后期受N、P共同限制,P限制更为强烈[18].在水分短缺的干旱半干旱区域,植物经过长期的适应进化,形成了独特的生态化学计量特征.荒漠中不同植物对环境的适应策略和养分利用状况体现在C、N、P化学计量特征上[3].大量研究表明干旱通过减少植物对N、P的吸收、转运和再分配来抑制植物生长[19].而叶片C含量高的植物具有较强的抗旱能力,如毛乌素沙地典型植物叶片的平均C含量较高,通过增强抵御能力来适应沙地水分缺乏、土壤养分贫瘠的不利环境[20].而从植物个体角度来看,古尔班通古特沙漠4种草本植物叶片N、P含量高于阿拉善荒漠植物[21].可见,由于植被结构和组成上的差异,植被C∶N∶P特征在不同的生态系统之间变异很大[22-23]. ...
Plant litter quality affects the accumulation rate,composition,and stability of mineral-associated soil organic matter
2
2018
... 土壤作为最活跃的生命层,是植物生长繁育的基础,具有养分转化的生态功能,土壤也是陆地生态系统中最为复杂多变的环境因子[24].土地利用方式的改变、人为因素的扰动以及冰期、气候地区间的差异使得土壤中的养分循环具有一定的空间异质性[25].此外,植物枯落物的凋落、土壤中水分的流动、植物与大气之间的相互作用使得养分在陆地生态系统中不断地进行再分配[24].元素在土壤中的渗透和扩散速率也缓慢得多,这使得陆生动植物对土壤的反馈作用更多地被限制在表层土壤中[26].可见,研究表层土壤中C∶N∶P的变化是荒漠生态系统C∶N∶P研究的基础. ...
... [24].元素在土壤中的渗透和扩散速率也缓慢得多,这使得陆生动植物对土壤的反馈作用更多地被限制在表层土壤中[26].可见,研究表层土壤中C∶N∶P的变化是荒漠生态系统C∶N∶P研究的基础. ...
土壤与植物生态化学计量学研究进展
1
2016
... 土壤作为最活跃的生命层,是植物生长繁育的基础,具有养分转化的生态功能,土壤也是陆地生态系统中最为复杂多变的环境因子[24].土地利用方式的改变、人为因素的扰动以及冰期、气候地区间的差异使得土壤中的养分循环具有一定的空间异质性[25].此外,植物枯落物的凋落、土壤中水分的流动、植物与大气之间的相互作用使得养分在陆地生态系统中不断地进行再分配[24].元素在土壤中的渗透和扩散速率也缓慢得多,这使得陆生动植物对土壤的反馈作用更多地被限制在表层土壤中[26].可见,研究表层土壤中C∶N∶P的变化是荒漠生态系统C∶N∶P研究的基础. ...
Pattern and variation of C∶N∶P ratios in China's soils:a synthesis of observational data
3
2010
... 土壤作为最活跃的生命层,是植物生长繁育的基础,具有养分转化的生态功能,土壤也是陆地生态系统中最为复杂多变的环境因子[24].土地利用方式的改变、人为因素的扰动以及冰期、气候地区间的差异使得土壤中的养分循环具有一定的空间异质性[25].此外,植物枯落物的凋落、土壤中水分的流动、植物与大气之间的相互作用使得养分在陆地生态系统中不断地进行再分配[24].元素在土壤中的渗透和扩散速率也缓慢得多,这使得陆生动植物对土壤的反馈作用更多地被限制在表层土壤中[26].可见,研究表层土壤中C∶N∶P的变化是荒漠生态系统C∶N∶P研究的基础. ...
... 植物根系和凋落物是土壤有机碳(SOC)周转的主要物质来源和驱动力[27].土壤中N素来源主要有生物固氮及氮沉降两种途径,其中,凋落物中的有机氮经矿化作用形成土壤有效N是最重要的途径.而P在生态系统中的生物地球化学循环是沉积型循环,主要源于含P的矿物岩石的缓慢风化作用[28].土壤中的C∶N∶P比值是评价土壤质量的重要指标.Aponte等[29]发现在全球范围内,虽然土壤中C、N、P含量的变化范围很大,但它们三者之间具有显著的正相关关系,而C∶N∶P也被限制在一个很窄的范围内,就全球平均值而言,土壤中的C∶N∶P恒定,即186∶13∶1.中国2 384个完整土壤剖面的C∶N、C∶P及N∶P原子比低于全球表层土壤,说明尽管不同土层C、N、P相互制约,但其生态化学计量学会随环境的改变而变化[26].而在荒漠地区,土壤有机质含量低并且分解缓慢影响到C∶N∶P.陶冶等[30]分析了准噶尔荒漠79个样点浅层(0—10 cm)土壤化学计量特征,发现与全国及全球平均水平相比,准噶尔荒漠土壤C、N、P含量、C∶P及N∶P均较低而C∶N较高,养分元素缺乏程度表现为N>C>P;而古尔班通古特沙漠土壤C、N、P养分元素含量及其化学计量比均远低于全球及全国平均水平,C∶N和P含量具有较高的稳定性[31-32].有研究发现土壤化学计量特征在不同群落类型间、不同生物土壤结皮(Biological Soil Crusts,BSC)类型间均有一定差异,植物个体对土壤C、N、P含量及比值也有显著影响[33]. ...
... 研究发现,全球森林生态系统植物叶片C∶N∶P为1 212∶28∶1,而当植物叶片凋亡后,养分的再吸收过程使得枯叶的C∶N∶P变为3 007∶45∶1[60];植物凋亡后根的C∶N∶P平均值为4 184∶43∶1[61];土壤C∶N∶P最低为287∶17∶1[62];土壤微生物量C∶N∶P平均值为42∶6∶1,并随土壤深度增加显著减小[26];Sinsabaugh等[57]发现土壤酶中C∶N∶P接近于1∶1∶1.相比之下,从植物叶片、凋落物到土壤再到土壤微生物及酶的计量特征C、N元素整体处于减少趋势.Xu等[37]研究发现,在区域尺度上,随着生物区系不同,C∶N∶P存在很大变异性,如天然湿地中的C∶N∶P为131∶14∶1,而沙漠中的C∶N∶P则为31∶4∶1.面对底物和微生物化学计量间的巨大差异,微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例和元素利用效率等机制来响应和调节.目前为止,关于植物-土壤-微生物-酶C、N、P关系的研究多聚焦于探讨土壤微生物与土壤、植被间的变化及其影响因素.研究表明,随着纬度、年均温、年降水量的变化,微生物量N∶P与植物组织的N∶P呈现出一致的变化趋势,即低纬度地区更受P的限制[63].这表明在高纬度地区微生物和植物一样由于生长季短而需要更高的生长速率,进而需要高的N、P含量和低的N∶P维持[6].Li等[64]分析了中国亚热带地区土壤数据后认为,微生物量C∶N和C∶P会随土壤C∶N和C∶P的增加而增加;周正虎等[38]却发现中国森林土壤微生物量C∶N随着土壤C∶N的增加而减小,与Cleveland等[65]对全球森林的数据分析结果一致.近期,吕星宇等[35]在腾格里沙漠开展的人工固沙植被研究表明,固沙植被演替过程中,微生物功能群数量C∶N∶P的变化主要受到草本、灌木和土壤C∶N∶P变化的影响.受灌木C∶N∶P增加及草本C∶N∶P减小的影响,微生物功能群数量C∶N∶P随之增加;对土壤酶C∶N∶P影响最大的因子分别是微生物功能群数量C∶N∶P和凋落物C∶N∶P,土壤酶C∶N∶P随着微生物功能群数量C∶N∶P增加而增加. ...
Mycorrhizal fungi as mediators of soil organic matter dynamics
1
2019
... 植物根系和凋落物是土壤有机碳(SOC)周转的主要物质来源和驱动力[27].土壤中N素来源主要有生物固氮及氮沉降两种途径,其中,凋落物中的有机氮经矿化作用形成土壤有效N是最重要的途径.而P在生态系统中的生物地球化学循环是沉积型循环,主要源于含P的矿物岩石的缓慢风化作用[28].土壤中的C∶N∶P比值是评价土壤质量的重要指标.Aponte等[29]发现在全球范围内,虽然土壤中C、N、P含量的变化范围很大,但它们三者之间具有显著的正相关关系,而C∶N∶P也被限制在一个很窄的范围内,就全球平均值而言,土壤中的C∶N∶P恒定,即186∶13∶1.中国2 384个完整土壤剖面的C∶N、C∶P及N∶P原子比低于全球表层土壤,说明尽管不同土层C、N、P相互制约,但其生态化学计量学会随环境的改变而变化[26].而在荒漠地区,土壤有机质含量低并且分解缓慢影响到C∶N∶P.陶冶等[30]分析了准噶尔荒漠79个样点浅层(0—10 cm)土壤化学计量特征,发现与全国及全球平均水平相比,准噶尔荒漠土壤C、N、P含量、C∶P及N∶P均较低而C∶N较高,养分元素缺乏程度表现为N>C>P;而古尔班通古特沙漠土壤C、N、P养分元素含量及其化学计量比均远低于全球及全国平均水平,C∶N和P含量具有较高的稳定性[31-32].有研究发现土壤化学计量特征在不同群落类型间、不同生物土壤结皮(Biological Soil Crusts,BSC)类型间均有一定差异,植物个体对土壤C、N、P含量及比值也有显著影响[33]. ...
陆地生态系统磷素循环及其影响因素
1
2005
... 植物根系和凋落物是土壤有机碳(SOC)周转的主要物质来源和驱动力[27].土壤中N素来源主要有生物固氮及氮沉降两种途径,其中,凋落物中的有机氮经矿化作用形成土壤有效N是最重要的途径.而P在生态系统中的生物地球化学循环是沉积型循环,主要源于含P的矿物岩石的缓慢风化作用[28].土壤中的C∶N∶P比值是评价土壤质量的重要指标.Aponte等[29]发现在全球范围内,虽然土壤中C、N、P含量的变化范围很大,但它们三者之间具有显著的正相关关系,而C∶N∶P也被限制在一个很窄的范围内,就全球平均值而言,土壤中的C∶N∶P恒定,即186∶13∶1.中国2 384个完整土壤剖面的C∶N、C∶P及N∶P原子比低于全球表层土壤,说明尽管不同土层C、N、P相互制约,但其生态化学计量学会随环境的改变而变化[26].而在荒漠地区,土壤有机质含量低并且分解缓慢影响到C∶N∶P.陶冶等[30]分析了准噶尔荒漠79个样点浅层(0—10 cm)土壤化学计量特征,发现与全国及全球平均水平相比,准噶尔荒漠土壤C、N、P含量、C∶P及N∶P均较低而C∶N较高,养分元素缺乏程度表现为N>C>P;而古尔班通古特沙漠土壤C、N、P养分元素含量及其化学计量比均远低于全球及全国平均水平,C∶N和P含量具有较高的稳定性[31-32].有研究发现土壤化学计量特征在不同群落类型间、不同生物土壤结皮(Biological Soil Crusts,BSC)类型间均有一定差异,植物个体对土壤C、N、P含量及比值也有显著影响[33]. ...
Microbial N and P in soils of Mediterranean oak forests:influence of season,canopy cover and soil depth
1
2010
... 植物根系和凋落物是土壤有机碳(SOC)周转的主要物质来源和驱动力[27].土壤中N素来源主要有生物固氮及氮沉降两种途径,其中,凋落物中的有机氮经矿化作用形成土壤有效N是最重要的途径.而P在生态系统中的生物地球化学循环是沉积型循环,主要源于含P的矿物岩石的缓慢风化作用[28].土壤中的C∶N∶P比值是评价土壤质量的重要指标.Aponte等[29]发现在全球范围内,虽然土壤中C、N、P含量的变化范围很大,但它们三者之间具有显著的正相关关系,而C∶N∶P也被限制在一个很窄的范围内,就全球平均值而言,土壤中的C∶N∶P恒定,即186∶13∶1.中国2 384个完整土壤剖面的C∶N、C∶P及N∶P原子比低于全球表层土壤,说明尽管不同土层C、N、P相互制约,但其生态化学计量学会随环境的改变而变化[26].而在荒漠地区,土壤有机质含量低并且分解缓慢影响到C∶N∶P.陶冶等[30]分析了准噶尔荒漠79个样点浅层(0—10 cm)土壤化学计量特征,发现与全国及全球平均水平相比,准噶尔荒漠土壤C、N、P含量、C∶P及N∶P均较低而C∶N较高,养分元素缺乏程度表现为N>C>P;而古尔班通古特沙漠土壤C、N、P养分元素含量及其化学计量比均远低于全球及全国平均水平,C∶N和P含量具有较高的稳定性[31-32].有研究发现土壤化学计量特征在不同群落类型间、不同生物土壤结皮(Biological Soil Crusts,BSC)类型间均有一定差异,植物个体对土壤C、N、P含量及比值也有显著影响[33]. ...
准噶尔荒漠区域尺度浅层土壤化学计量特征及其空间分布格局
1
2016
... 植物根系和凋落物是土壤有机碳(SOC)周转的主要物质来源和驱动力[27].土壤中N素来源主要有生物固氮及氮沉降两种途径,其中,凋落物中的有机氮经矿化作用形成土壤有效N是最重要的途径.而P在生态系统中的生物地球化学循环是沉积型循环,主要源于含P的矿物岩石的缓慢风化作用[28].土壤中的C∶N∶P比值是评价土壤质量的重要指标.Aponte等[29]发现在全球范围内,虽然土壤中C、N、P含量的变化范围很大,但它们三者之间具有显著的正相关关系,而C∶N∶P也被限制在一个很窄的范围内,就全球平均值而言,土壤中的C∶N∶P恒定,即186∶13∶1.中国2 384个完整土壤剖面的C∶N、C∶P及N∶P原子比低于全球表层土壤,说明尽管不同土层C、N、P相互制约,但其生态化学计量学会随环境的改变而变化[26].而在荒漠地区,土壤有机质含量低并且分解缓慢影响到C∶N∶P.陶冶等[30]分析了准噶尔荒漠79个样点浅层(0—10 cm)土壤化学计量特征,发现与全国及全球平均水平相比,准噶尔荒漠土壤C、N、P含量、C∶P及N∶P均较低而C∶N较高,养分元素缺乏程度表现为N>C>P;而古尔班通古特沙漠土壤C、N、P养分元素含量及其化学计量比均远低于全球及全国平均水平,C∶N和P含量具有较高的稳定性[31-32].有研究发现土壤化学计量特征在不同群落类型间、不同生物土壤结皮(Biological Soil Crusts,BSC)类型间均有一定差异,植物个体对土壤C、N、P含量及比值也有显著影响[33]. ...
古尔班通古特沙漠典型灌木群落土壤化学计量特征及其影响因素
1
2017
... 植物根系和凋落物是土壤有机碳(SOC)周转的主要物质来源和驱动力[27].土壤中N素来源主要有生物固氮及氮沉降两种途径,其中,凋落物中的有机氮经矿化作用形成土壤有效N是最重要的途径.而P在生态系统中的生物地球化学循环是沉积型循环,主要源于含P的矿物岩石的缓慢风化作用[28].土壤中的C∶N∶P比值是评价土壤质量的重要指标.Aponte等[29]发现在全球范围内,虽然土壤中C、N、P含量的变化范围很大,但它们三者之间具有显著的正相关关系,而C∶N∶P也被限制在一个很窄的范围内,就全球平均值而言,土壤中的C∶N∶P恒定,即186∶13∶1.中国2 384个完整土壤剖面的C∶N、C∶P及N∶P原子比低于全球表层土壤,说明尽管不同土层C、N、P相互制约,但其生态化学计量学会随环境的改变而变化[26].而在荒漠地区,土壤有机质含量低并且分解缓慢影响到C∶N∶P.陶冶等[30]分析了准噶尔荒漠79个样点浅层(0—10 cm)土壤化学计量特征,发现与全国及全球平均水平相比,准噶尔荒漠土壤C、N、P含量、C∶P及N∶P均较低而C∶N较高,养分元素缺乏程度表现为N>C>P;而古尔班通古特沙漠土壤C、N、P养分元素含量及其化学计量比均远低于全球及全国平均水平,C∶N和P含量具有较高的稳定性[31-32].有研究发现土壤化学计量特征在不同群落类型间、不同生物土壤结皮(Biological Soil Crusts,BSC)类型间均有一定差异,植物个体对土壤C、N、P含量及比值也有显著影响[33]. ...
古尔班通古特沙漠5种植物凋落物分解特征
1
2020
... 植物根系和凋落物是土壤有机碳(SOC)周转的主要物质来源和驱动力[27].土壤中N素来源主要有生物固氮及氮沉降两种途径,其中,凋落物中的有机氮经矿化作用形成土壤有效N是最重要的途径.而P在生态系统中的生物地球化学循环是沉积型循环,主要源于含P的矿物岩石的缓慢风化作用[28].土壤中的C∶N∶P比值是评价土壤质量的重要指标.Aponte等[29]发现在全球范围内,虽然土壤中C、N、P含量的变化范围很大,但它们三者之间具有显著的正相关关系,而C∶N∶P也被限制在一个很窄的范围内,就全球平均值而言,土壤中的C∶N∶P恒定,即186∶13∶1.中国2 384个完整土壤剖面的C∶N、C∶P及N∶P原子比低于全球表层土壤,说明尽管不同土层C、N、P相互制约,但其生态化学计量学会随环境的改变而变化[26].而在荒漠地区,土壤有机质含量低并且分解缓慢影响到C∶N∶P.陶冶等[30]分析了准噶尔荒漠79个样点浅层(0—10 cm)土壤化学计量特征,发现与全国及全球平均水平相比,准噶尔荒漠土壤C、N、P含量、C∶P及N∶P均较低而C∶N较高,养分元素缺乏程度表现为N>C>P;而古尔班通古特沙漠土壤C、N、P养分元素含量及其化学计量比均远低于全球及全国平均水平,C∶N和P含量具有较高的稳定性[31-32].有研究发现土壤化学计量特征在不同群落类型间、不同生物土壤结皮(Biological Soil Crusts,BSC)类型间均有一定差异,植物个体对土壤C、N、P含量及比值也有显著影响[33]. ...
黄土丘陵区生物结皮土壤生态化学计量学特征及其对干扰的响应
1
2019
... 植物根系和凋落物是土壤有机碳(SOC)周转的主要物质来源和驱动力[27].土壤中N素来源主要有生物固氮及氮沉降两种途径,其中,凋落物中的有机氮经矿化作用形成土壤有效N是最重要的途径.而P在生态系统中的生物地球化学循环是沉积型循环,主要源于含P的矿物岩石的缓慢风化作用[28].土壤中的C∶N∶P比值是评价土壤质量的重要指标.Aponte等[29]发现在全球范围内,虽然土壤中C、N、P含量的变化范围很大,但它们三者之间具有显著的正相关关系,而C∶N∶P也被限制在一个很窄的范围内,就全球平均值而言,土壤中的C∶N∶P恒定,即186∶13∶1.中国2 384个完整土壤剖面的C∶N、C∶P及N∶P原子比低于全球表层土壤,说明尽管不同土层C、N、P相互制约,但其生态化学计量学会随环境的改变而变化[26].而在荒漠地区,土壤有机质含量低并且分解缓慢影响到C∶N∶P.陶冶等[30]分析了准噶尔荒漠79个样点浅层(0—10 cm)土壤化学计量特征,发现与全国及全球平均水平相比,准噶尔荒漠土壤C、N、P含量、C∶P及N∶P均较低而C∶N较高,养分元素缺乏程度表现为N>C>P;而古尔班通古特沙漠土壤C、N、P养分元素含量及其化学计量比均远低于全球及全国平均水平,C∶N和P含量具有较高的稳定性[31-32].有研究发现土壤化学计量特征在不同群落类型间、不同生物土壤结皮(Biological Soil Crusts,BSC)类型间均有一定差异,植物个体对土壤C、N、P含量及比值也有显著影响[33]. ...
Interactions among functional groups in the cycling of,carbon,nitrogen and phosphorus in the rhizosphere of three successional species of tropical woody trees
1
2005
... 土壤微生物是植物生长过程中养分循环的重要调控者,其种类、数量及空间分布直接或间接反映了土壤生境的特征及其转化规律,因此成为表征陆地生态系统土壤质量变化、指示土壤发育状况和植被演替的重要指标[34].土壤微生物量是活的土壤有机质部分,是土壤养分特别是N和P等的“源”和“库”[35].土壤微生物量通过固持和矿化作用控制土壤生态系统C、N、P的流量.因而研究土壤微生物量碳(MBC)、氮(MBN)、磷(MBP)的生态化学计量特征对理解土壤生态系统的限制性营养元素具有重要意义[36]. ...
固沙植被区土壤质地与土壤微生物数量的关系
4
2019
... 土壤微生物是植物生长过程中养分循环的重要调控者,其种类、数量及空间分布直接或间接反映了土壤生境的特征及其转化规律,因此成为表征陆地生态系统土壤质量变化、指示土壤发育状况和植被演替的重要指标[34].土壤微生物量是活的土壤有机质部分,是土壤养分特别是N和P等的“源”和“库”[35].土壤微生物量通过固持和矿化作用控制土壤生态系统C、N、P的流量.因而研究土壤微生物量碳(MBC)、氮(MBN)、磷(MBP)的生态化学计量特征对理解土壤生态系统的限制性营养元素具有重要意义[36]. ...
... 土壤微生物根据功能可以划分为不同的类群,依据在C、N、P循环过程中的作用,土壤微生物可划分为纤维素分解菌、氨化细菌、硝化细菌、溶磷菌等生理类群[40].不同功能群在养分循环中生物学功能各异[41].土壤中纤维素降解微生物功能群不仅是土壤碳素循环的主要驱动者,也是维持土壤生物活性、植物生长碳素和能源的重要供给来源[42];氨化和硝化微生物功能群在土壤氮素循环过程中发挥重要作用[42];溶磷微生物功能群可使土壤中难以被植物吸收利用的磷转变成可被利用的磷化物[43].因此,对土壤微生物以功能为单位进行研究,可以更好地掌握生物地球化学循环及生态系统的演替规律,特别是对退化生态系统的恢复及提高其生物学功能均有重要的理论和实践意义[42].同样,环境变化可直接或间接地影响微生物群落结构和多样性,进而对整个生态系统产生影响.大量研究表明,土壤类型、土壤理化性质(水分、温度、pH和C、N、P元素含量等)[44-45]、植被类型及物种多样性等是影响土壤微生物功能群的重要因素[39],而在干旱区土壤微生物对这些因素的变化更加敏感.如张静茹等[46]论述了降雨对干旱半干旱地区土壤微生物数量的直接和间接影响;Liu等[47]在中国北方温带半干旱草原进行增温增雨试验,发现土壤微生物对增温的效应比增雨更强;此外,植被恢复年限、植物种、BSC、季节变化等均被证实能显著影响干旱沙区土壤微生物数量[35,48].近期,Wang等[49]发现随着中国北方干旱沙区优势植物物种变化,N含量是决定土壤细菌群落的关键因素.从研究方法上看,现阶段关于微生物群落结构的研究多采用传统培养法、高通量基因组测序法(如TFLP和DGGE)、磷脂脂肪酸法(PLFA)进行微生物分类和多样性的研究[36],或应用Biology-GN技术和基因芯片技术研究微生物功能群的组成与丰度[50].然而,由于土壤中可培养的微生物仅占很小的一部分、微生物底物来源非常繁杂、磷脂脂肪酸的复杂性和非专一性等原因,使用单一方法很难获取微生物功能群的真实情况.从研究内容上来看,近年来围绕固沙植被演替过程中土壤微生物数量、分类和多样性开展了一些研究[51],发现干旱沙区微生物功能基因丰度随植被重建年限增加而增大,尤其是C、N循环基因丰度的增加促进了微生物的代谢潜力[48]. ...
... 研究发现,全球森林生态系统植物叶片C∶N∶P为1 212∶28∶1,而当植物叶片凋亡后,养分的再吸收过程使得枯叶的C∶N∶P变为3 007∶45∶1[60];植物凋亡后根的C∶N∶P平均值为4 184∶43∶1[61];土壤C∶N∶P最低为287∶17∶1[62];土壤微生物量C∶N∶P平均值为42∶6∶1,并随土壤深度增加显著减小[26];Sinsabaugh等[57]发现土壤酶中C∶N∶P接近于1∶1∶1.相比之下,从植物叶片、凋落物到土壤再到土壤微生物及酶的计量特征C、N元素整体处于减少趋势.Xu等[37]研究发现,在区域尺度上,随着生物区系不同,C∶N∶P存在很大变异性,如天然湿地中的C∶N∶P为131∶14∶1,而沙漠中的C∶N∶P则为31∶4∶1.面对底物和微生物化学计量间的巨大差异,微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例和元素利用效率等机制来响应和调节.目前为止,关于植物-土壤-微生物-酶C、N、P关系的研究多聚焦于探讨土壤微生物与土壤、植被间的变化及其影响因素.研究表明,随着纬度、年均温、年降水量的变化,微生物量N∶P与植物组织的N∶P呈现出一致的变化趋势,即低纬度地区更受P的限制[63].这表明在高纬度地区微生物和植物一样由于生长季短而需要更高的生长速率,进而需要高的N、P含量和低的N∶P维持[6].Li等[64]分析了中国亚热带地区土壤数据后认为,微生物量C∶N和C∶P会随土壤C∶N和C∶P的增加而增加;周正虎等[38]却发现中国森林土壤微生物量C∶N随着土壤C∶N的增加而减小,与Cleveland等[65]对全球森林的数据分析结果一致.近期,吕星宇等[35]在腾格里沙漠开展的人工固沙植被研究表明,固沙植被演替过程中,微生物功能群数量C∶N∶P的变化主要受到草本、灌木和土壤C∶N∶P变化的影响.受灌木C∶N∶P增加及草本C∶N∶P减小的影响,微生物功能群数量C∶N∶P随之增加;对土壤酶C∶N∶P影响最大的因子分别是微生物功能群数量C∶N∶P和凋落物C∶N∶P,土壤酶C∶N∶P随着微生物功能群数量C∶N∶P增加而增加. ...
... 植被重建是干旱半干旱荒漠区防止土地沙化、减少风沙危害、促进生态恢复的重要途径[63].中国利用植物固沙已有60多年的历史,国家先后在风沙危害区启动了“三北”防护林建设、退耕还林还草和京津风沙源治理等以人工植被建设作为主要修复措施的一批重大生态建设工程.60余年来,中国沙区植被建设取得了举世瞩目的成就,有效遏制了沙漠化的发展,促进了区域生境恢复[66].人工固沙植被是荒漠生态系统重要的组成部分,是土地荒漠化防治和水土流失治理的主要调控者[66].这些先后建立的人工植被为我们进行荒漠生态系统恢复过程中化学计量学研究奠定了基础.以沙坡头为例,人工固沙植被建立后,原有的流沙演变成一个复杂的人工-天然复合的荒漠生态系统.土壤性质的改善和成土作用的加剧,不但使得流沙逐渐向地带性土壤演变,而且促使生物地球化学循环逐渐集中在浅表层[35].此外,随着土壤肥力的提升,植被也逐渐向天然植物群落演变,植物多样性增加的同时,鸟类、动物及昆虫也明显增多[64].开展人工固沙植被生态系统C∶N∶P化学计量的研究,是理解固沙植被系统C、N、P循环过程及植被、微生物和土壤三者之间互馈关系的核心和关键,也是进行科学防沙治沙和管理固沙植被的指导依据.然而,针对沙区人工植被建立后土壤微生物功能群及其化学计量学的研究目前相对较少,需加强人工固沙植被生态系统演替进程中C∶N∶P化学计量的研究. ...
Embracing the unknown:disentangling the complexities of the soil microbiome
5
2017
... 土壤微生物是植物生长过程中养分循环的重要调控者,其种类、数量及空间分布直接或间接反映了土壤生境的特征及其转化规律,因此成为表征陆地生态系统土壤质量变化、指示土壤发育状况和植被演替的重要指标[34].土壤微生物量是活的土壤有机质部分,是土壤养分特别是N和P等的“源”和“库”[35].土壤微生物量通过固持和矿化作用控制土壤生态系统C、N、P的流量.因而研究土壤微生物量碳(MBC)、氮(MBN)、磷(MBP)的生态化学计量特征对理解土壤生态系统的限制性营养元素具有重要意义[36]. ...
... 土壤微生物根据功能可以划分为不同的类群,依据在C、N、P循环过程中的作用,土壤微生物可划分为纤维素分解菌、氨化细菌、硝化细菌、溶磷菌等生理类群[40].不同功能群在养分循环中生物学功能各异[41].土壤中纤维素降解微生物功能群不仅是土壤碳素循环的主要驱动者,也是维持土壤生物活性、植物生长碳素和能源的重要供给来源[42];氨化和硝化微生物功能群在土壤氮素循环过程中发挥重要作用[42];溶磷微生物功能群可使土壤中难以被植物吸收利用的磷转变成可被利用的磷化物[43].因此,对土壤微生物以功能为单位进行研究,可以更好地掌握生物地球化学循环及生态系统的演替规律,特别是对退化生态系统的恢复及提高其生物学功能均有重要的理论和实践意义[42].同样,环境变化可直接或间接地影响微生物群落结构和多样性,进而对整个生态系统产生影响.大量研究表明,土壤类型、土壤理化性质(水分、温度、pH和C、N、P元素含量等)[44-45]、植被类型及物种多样性等是影响土壤微生物功能群的重要因素[39],而在干旱区土壤微生物对这些因素的变化更加敏感.如张静茹等[46]论述了降雨对干旱半干旱地区土壤微生物数量的直接和间接影响;Liu等[47]在中国北方温带半干旱草原进行增温增雨试验,发现土壤微生物对增温的效应比增雨更强;此外,植被恢复年限、植物种、BSC、季节变化等均被证实能显著影响干旱沙区土壤微生物数量[35,48].近期,Wang等[49]发现随着中国北方干旱沙区优势植物物种变化,N含量是决定土壤细菌群落的关键因素.从研究方法上看,现阶段关于微生物群落结构的研究多采用传统培养法、高通量基因组测序法(如TFLP和DGGE)、磷脂脂肪酸法(PLFA)进行微生物分类和多样性的研究[36],或应用Biology-GN技术和基因芯片技术研究微生物功能群的组成与丰度[50].然而,由于土壤中可培养的微生物仅占很小的一部分、微生物底物来源非常繁杂、磷脂脂肪酸的复杂性和非专一性等原因,使用单一方法很难获取微生物功能群的真实情况.从研究内容上来看,近年来围绕固沙植被演替过程中土壤微生物数量、分类和多样性开展了一些研究[51],发现干旱沙区微生物功能基因丰度随植被重建年限增加而增大,尤其是C、N循环基因丰度的增加促进了微生物的代谢潜力[48]. ...
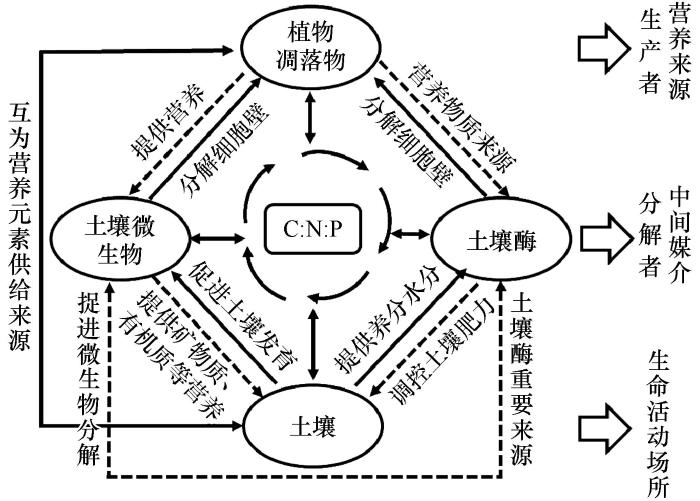
... 生态系统中C、N、P等元素的生物地球化学循环过程将植被、土壤、土壤微生物和酶有机地联系在一起[41].植物是生产者,植物通过其凋落物和根系及分泌物为土壤微生物和土壤酶提供营养,促进土壤微生物和酶的多样性.而土壤微生物则是主要分解者,可以分解植物及凋落物的残体,对环境起着天然的“过滤”和“净化”作用.同时,土壤微生物能够分泌土壤酶[36],大多数土壤酶是微生物对环境响应表达的产物,能分解植物和微生物的细胞壁,降解大分子,并最终产生可溶性底物供微生物同化.土壤为生物地球化学循环和能量流动过程提供场所,是生态系统稳定性维持的基础.植物从土壤中吸收生长所需的营养物质,再以凋落物的形式将养分元素返还给土壤.植物-凋落物-土壤系统外部的元素能够通过特定微生物和酶进入土壤,植物或凋落物在分解的过程中还受到微生物和酶的共同作用.同时,土壤有机质和有机残体在土壤酶的作用下被分解为不同的产物,为微生物和植物的生长发育提供了物质和能量基础.因此,微生物和酶作为中间媒介,土壤作为载体,共同调控C、N、P在生态系统中的循环,进而影响植物-凋落物-土壤生态化学计量特征的变化(图1). ...
... 此外,建立微生物群落结构与功能具体的联系是土壤微生物学面临的一大挑战.许多微生物过程并不是单一途径的代谢产物,而是可以被分类的综合代谢途径的产物.如土壤中几乎所有的异养细菌活性类群都可能促进有机碳分解代谢或氮矿化的速率[36].甚至就植物纤维素分解代谢而言,可能需要多个不同类型的代谢过程共同完成[68].所以将微生物以功能群的形式进行划分,研究不同微生物功能群C∶N∶P化学计量也将是未来研究的重点. ...
... 随着现代分子技术的发展,荒漠中潜在的微生物资源也被充分挖掘.如何利用丰富的基因组、宏基因组和标记基因数据来提高我们对土壤微生物群功能的理解,也是未来研究中需要攻克的难题.基于DNA测序方法,可广泛用于量化微生物群落的相对丰度.然而,土壤中微生物功能基因(如宏基因组测序)通常只提供某一类微生物的相对丰度或基因百分比,而不是它们的绝对丰度.土壤过程更可能与微生物功能群、基因或基因产物的绝对数量有关,而不是与它们在群落中的相对比例有关[36].推断微生物功能群、基因或基因产物的绝对丰度需要不同研究方法的结合使用.如将传统培养法、BIOLOG-GN法及基因芯片测序等方法结合使用,探讨如何采用多种方法进行土壤微生物功能群C∶N∶P的探索性研究目前还处于空白状态,相信在未来研究中也将被予以重视. ...
A global analysis of soil microbial biomass carbon,nitrogen and phosphorus in terrestrial ecosystems
3
2013
... 目前,绝大多数有关微生物C∶N∶P的研究多聚焦于MBC∶MBN∶MBP及其与土壤C∶N∶P之间的关系.全球尺度上的MBC∶MBN∶MBP平均值为42∶6∶1[37],不同的植被类型间差异明显[9].如沙漠土壤中MBC∶MBN∶MBP仅为31∶4∶1,而在天然湿地中则高达131∶14∶1[37].土壤MBC、MBN、MBP与土壤C、N、P呈显著正相关,MBC随土壤C∶N的增加而减小,而MBN却随土壤C∶N的增加而增加[38].张义凡等[39]以宁夏盐池荒漠草原的柠条、沙蒿、短花针茅典型群落为研究对象,分析了不同生境(冠下、丛间)和不同土层间MBC、MBN的变化特征,发现3种群落土壤微生物量变化差异较大,且MBC和MBN的含量均表现为冠下>丛间,并随土层深度的增加而降低,土壤C∶N与MBC∶MBN显著正相关.这表明土壤MBC、MBN具有一定的生物学指示特性,可以作为评价土壤质量的生物学指标. ...
... [37].土壤MBC、MBN、MBP与土壤C、N、P呈显著正相关,MBC随土壤C∶N的增加而减小,而MBN却随土壤C∶N的增加而增加[38].张义凡等[39]以宁夏盐池荒漠草原的柠条、沙蒿、短花针茅典型群落为研究对象,分析了不同生境(冠下、丛间)和不同土层间MBC、MBN的变化特征,发现3种群落土壤微生物量变化差异较大,且MBC和MBN的含量均表现为冠下>丛间,并随土层深度的增加而降低,土壤C∶N与MBC∶MBN显著正相关.这表明土壤MBC、MBN具有一定的生物学指示特性,可以作为评价土壤质量的生物学指标. ...
... 研究发现,全球森林生态系统植物叶片C∶N∶P为1 212∶28∶1,而当植物叶片凋亡后,养分的再吸收过程使得枯叶的C∶N∶P变为3 007∶45∶1[60];植物凋亡后根的C∶N∶P平均值为4 184∶43∶1[61];土壤C∶N∶P最低为287∶17∶1[62];土壤微生物量C∶N∶P平均值为42∶6∶1,并随土壤深度增加显著减小[26];Sinsabaugh等[57]发现土壤酶中C∶N∶P接近于1∶1∶1.相比之下,从植物叶片、凋落物到土壤再到土壤微生物及酶的计量特征C、N元素整体处于减少趋势.Xu等[37]研究发现,在区域尺度上,随着生物区系不同,C∶N∶P存在很大变异性,如天然湿地中的C∶N∶P为131∶14∶1,而沙漠中的C∶N∶P则为31∶4∶1.面对底物和微生物化学计量间的巨大差异,微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例和元素利用效率等机制来响应和调节.目前为止,关于植物-土壤-微生物-酶C、N、P关系的研究多聚焦于探讨土壤微生物与土壤、植被间的变化及其影响因素.研究表明,随着纬度、年均温、年降水量的变化,微生物量N∶P与植物组织的N∶P呈现出一致的变化趋势,即低纬度地区更受P的限制[63].这表明在高纬度地区微生物和植物一样由于生长季短而需要更高的生长速率,进而需要高的N、P含量和低的N∶P维持[6].Li等[64]分析了中国亚热带地区土壤数据后认为,微生物量C∶N和C∶P会随土壤C∶N和C∶P的增加而增加;周正虎等[38]却发现中国森林土壤微生物量C∶N随着土壤C∶N的增加而减小,与Cleveland等[65]对全球森林的数据分析结果一致.近期,吕星宇等[35]在腾格里沙漠开展的人工固沙植被研究表明,固沙植被演替过程中,微生物功能群数量C∶N∶P的变化主要受到草本、灌木和土壤C∶N∶P变化的影响.受灌木C∶N∶P增加及草本C∶N∶P减小的影响,微生物功能群数量C∶N∶P随之增加;对土壤酶C∶N∶P影响最大的因子分别是微生物功能群数量C∶N∶P和凋落物C∶N∶P,土壤酶C∶N∶P随着微生物功能群数量C∶N∶P增加而增加. ...
土地利用变化对东北温带幼龄林土壤碳氮磷含量及其化学计量特征的影响
2
2015
... 目前,绝大多数有关微生物C∶N∶P的研究多聚焦于MBC∶MBN∶MBP及其与土壤C∶N∶P之间的关系.全球尺度上的MBC∶MBN∶MBP平均值为42∶6∶1[37],不同的植被类型间差异明显[9].如沙漠土壤中MBC∶MBN∶MBP仅为31∶4∶1,而在天然湿地中则高达131∶14∶1[37].土壤MBC、MBN、MBP与土壤C、N、P呈显著正相关,MBC随土壤C∶N的增加而减小,而MBN却随土壤C∶N的增加而增加[38].张义凡等[39]以宁夏盐池荒漠草原的柠条、沙蒿、短花针茅典型群落为研究对象,分析了不同生境(冠下、丛间)和不同土层间MBC、MBN的变化特征,发现3种群落土壤微生物量变化差异较大,且MBC和MBN的含量均表现为冠下>丛间,并随土层深度的增加而降低,土壤C∶N与MBC∶MBN显著正相关.这表明土壤MBC、MBN具有一定的生物学指示特性,可以作为评价土壤质量的生物学指标. ...
... 研究发现,全球森林生态系统植物叶片C∶N∶P为1 212∶28∶1,而当植物叶片凋亡后,养分的再吸收过程使得枯叶的C∶N∶P变为3 007∶45∶1[60];植物凋亡后根的C∶N∶P平均值为4 184∶43∶1[61];土壤C∶N∶P最低为287∶17∶1[62];土壤微生物量C∶N∶P平均值为42∶6∶1,并随土壤深度增加显著减小[26];Sinsabaugh等[57]发现土壤酶中C∶N∶P接近于1∶1∶1.相比之下,从植物叶片、凋落物到土壤再到土壤微生物及酶的计量特征C、N元素整体处于减少趋势.Xu等[37]研究发现,在区域尺度上,随着生物区系不同,C∶N∶P存在很大变异性,如天然湿地中的C∶N∶P为131∶14∶1,而沙漠中的C∶N∶P则为31∶4∶1.面对底物和微生物化学计量间的巨大差异,微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例和元素利用效率等机制来响应和调节.目前为止,关于植物-土壤-微生物-酶C、N、P关系的研究多聚焦于探讨土壤微生物与土壤、植被间的变化及其影响因素.研究表明,随着纬度、年均温、年降水量的变化,微生物量N∶P与植物组织的N∶P呈现出一致的变化趋势,即低纬度地区更受P的限制[63].这表明在高纬度地区微生物和植物一样由于生长季短而需要更高的生长速率,进而需要高的N、P含量和低的N∶P维持[6].Li等[64]分析了中国亚热带地区土壤数据后认为,微生物量C∶N和C∶P会随土壤C∶N和C∶P的增加而增加;周正虎等[38]却发现中国森林土壤微生物量C∶N随着土壤C∶N的增加而减小,与Cleveland等[65]对全球森林的数据分析结果一致.近期,吕星宇等[35]在腾格里沙漠开展的人工固沙植被研究表明,固沙植被演替过程中,微生物功能群数量C∶N∶P的变化主要受到草本、灌木和土壤C∶N∶P变化的影响.受灌木C∶N∶P增加及草本C∶N∶P减小的影响,微生物功能群数量C∶N∶P随之增加;对土壤酶C∶N∶P影响最大的因子分别是微生物功能群数量C∶N∶P和凋落物C∶N∶P,土壤酶C∶N∶P随着微生物功能群数量C∶N∶P增加而增加. ...
荒漠草原3种典型群落类型的土壤微生物量碳氮研究
2
2017
... 目前,绝大多数有关微生物C∶N∶P的研究多聚焦于MBC∶MBN∶MBP及其与土壤C∶N∶P之间的关系.全球尺度上的MBC∶MBN∶MBP平均值为42∶6∶1[37],不同的植被类型间差异明显[9].如沙漠土壤中MBC∶MBN∶MBP仅为31∶4∶1,而在天然湿地中则高达131∶14∶1[37].土壤MBC、MBN、MBP与土壤C、N、P呈显著正相关,MBC随土壤C∶N的增加而减小,而MBN却随土壤C∶N的增加而增加[38].张义凡等[39]以宁夏盐池荒漠草原的柠条、沙蒿、短花针茅典型群落为研究对象,分析了不同生境(冠下、丛间)和不同土层间MBC、MBN的变化特征,发现3种群落土壤微生物量变化差异较大,且MBC和MBN的含量均表现为冠下>丛间,并随土层深度的增加而降低,土壤C∶N与MBC∶MBN显著正相关.这表明土壤MBC、MBN具有一定的生物学指示特性,可以作为评价土壤质量的生物学指标. ...
... 土壤微生物根据功能可以划分为不同的类群,依据在C、N、P循环过程中的作用,土壤微生物可划分为纤维素分解菌、氨化细菌、硝化细菌、溶磷菌等生理类群[40].不同功能群在养分循环中生物学功能各异[41].土壤中纤维素降解微生物功能群不仅是土壤碳素循环的主要驱动者,也是维持土壤生物活性、植物生长碳素和能源的重要供给来源[42];氨化和硝化微生物功能群在土壤氮素循环过程中发挥重要作用[42];溶磷微生物功能群可使土壤中难以被植物吸收利用的磷转变成可被利用的磷化物[43].因此,对土壤微生物以功能为单位进行研究,可以更好地掌握生物地球化学循环及生态系统的演替规律,特别是对退化生态系统的恢复及提高其生物学功能均有重要的理论和实践意义[42].同样,环境变化可直接或间接地影响微生物群落结构和多样性,进而对整个生态系统产生影响.大量研究表明,土壤类型、土壤理化性质(水分、温度、pH和C、N、P元素含量等)[44-45]、植被类型及物种多样性等是影响土壤微生物功能群的重要因素[39],而在干旱区土壤微生物对这些因素的变化更加敏感.如张静茹等[46]论述了降雨对干旱半干旱地区土壤微生物数量的直接和间接影响;Liu等[47]在中国北方温带半干旱草原进行增温增雨试验,发现土壤微生物对增温的效应比增雨更强;此外,植被恢复年限、植物种、BSC、季节变化等均被证实能显著影响干旱沙区土壤微生物数量[35,48].近期,Wang等[49]发现随着中国北方干旱沙区优势植物物种变化,N含量是决定土壤细菌群落的关键因素.从研究方法上看,现阶段关于微生物群落结构的研究多采用传统培养法、高通量基因组测序法(如TFLP和DGGE)、磷脂脂肪酸法(PLFA)进行微生物分类和多样性的研究[36],或应用Biology-GN技术和基因芯片技术研究微生物功能群的组成与丰度[50].然而,由于土壤中可培养的微生物仅占很小的一部分、微生物底物来源非常繁杂、磷脂脂肪酸的复杂性和非专一性等原因,使用单一方法很难获取微生物功能群的真实情况.从研究内容上来看,近年来围绕固沙植被演替过程中土壤微生物数量、分类和多样性开展了一些研究[51],发现干旱沙区微生物功能基因丰度随植被重建年限增加而增大,尤其是C、N循环基因丰度的增加促进了微生物的代谢潜力[48]. ...
东祁连山亚高山草地土壤微生物功能群数量动态及其与土壤环境关系
1
2007
... 土壤微生物根据功能可以划分为不同的类群,依据在C、N、P循环过程中的作用,土壤微生物可划分为纤维素分解菌、氨化细菌、硝化细菌、溶磷菌等生理类群[40].不同功能群在养分循环中生物学功能各异[41].土壤中纤维素降解微生物功能群不仅是土壤碳素循环的主要驱动者,也是维持土壤生物活性、植物生长碳素和能源的重要供给来源[42];氨化和硝化微生物功能群在土壤氮素循环过程中发挥重要作用[42];溶磷微生物功能群可使土壤中难以被植物吸收利用的磷转变成可被利用的磷化物[43].因此,对土壤微生物以功能为单位进行研究,可以更好地掌握生物地球化学循环及生态系统的演替规律,特别是对退化生态系统的恢复及提高其生物学功能均有重要的理论和实践意义[42].同样,环境变化可直接或间接地影响微生物群落结构和多样性,进而对整个生态系统产生影响.大量研究表明,土壤类型、土壤理化性质(水分、温度、pH和C、N、P元素含量等)[44-45]、植被类型及物种多样性等是影响土壤微生物功能群的重要因素[39],而在干旱区土壤微生物对这些因素的变化更加敏感.如张静茹等[46]论述了降雨对干旱半干旱地区土壤微生物数量的直接和间接影响;Liu等[47]在中国北方温带半干旱草原进行增温增雨试验,发现土壤微生物对增温的效应比增雨更强;此外,植被恢复年限、植物种、BSC、季节变化等均被证实能显著影响干旱沙区土壤微生物数量[35,48].近期,Wang等[49]发现随着中国北方干旱沙区优势植物物种变化,N含量是决定土壤细菌群落的关键因素.从研究方法上看,现阶段关于微生物群落结构的研究多采用传统培养法、高通量基因组测序法(如TFLP和DGGE)、磷脂脂肪酸法(PLFA)进行微生物分类和多样性的研究[36],或应用Biology-GN技术和基因芯片技术研究微生物功能群的组成与丰度[50].然而,由于土壤中可培养的微生物仅占很小的一部分、微生物底物来源非常繁杂、磷脂脂肪酸的复杂性和非专一性等原因,使用单一方法很难获取微生物功能群的真实情况.从研究内容上来看,近年来围绕固沙植被演替过程中土壤微生物数量、分类和多样性开展了一些研究[51],发现干旱沙区微生物功能基因丰度随植被重建年限增加而增大,尤其是C、N循环基因丰度的增加促进了微生物的代谢潜力[48]. ...
土壤-微生物-植物系统中矿物风化与元素循环
2
2014
... 土壤微生物根据功能可以划分为不同的类群,依据在C、N、P循环过程中的作用,土壤微生物可划分为纤维素分解菌、氨化细菌、硝化细菌、溶磷菌等生理类群[40].不同功能群在养分循环中生物学功能各异[41].土壤中纤维素降解微生物功能群不仅是土壤碳素循环的主要驱动者,也是维持土壤生物活性、植物生长碳素和能源的重要供给来源[42];氨化和硝化微生物功能群在土壤氮素循环过程中发挥重要作用[42];溶磷微生物功能群可使土壤中难以被植物吸收利用的磷转变成可被利用的磷化物[43].因此,对土壤微生物以功能为单位进行研究,可以更好地掌握生物地球化学循环及生态系统的演替规律,特别是对退化生态系统的恢复及提高其生物学功能均有重要的理论和实践意义[42].同样,环境变化可直接或间接地影响微生物群落结构和多样性,进而对整个生态系统产生影响.大量研究表明,土壤类型、土壤理化性质(水分、温度、pH和C、N、P元素含量等)[44-45]、植被类型及物种多样性等是影响土壤微生物功能群的重要因素[39],而在干旱区土壤微生物对这些因素的变化更加敏感.如张静茹等[46]论述了降雨对干旱半干旱地区土壤微生物数量的直接和间接影响;Liu等[47]在中国北方温带半干旱草原进行增温增雨试验,发现土壤微生物对增温的效应比增雨更强;此外,植被恢复年限、植物种、BSC、季节变化等均被证实能显著影响干旱沙区土壤微生物数量[35,48].近期,Wang等[49]发现随着中国北方干旱沙区优势植物物种变化,N含量是决定土壤细菌群落的关键因素.从研究方法上看,现阶段关于微生物群落结构的研究多采用传统培养法、高通量基因组测序法(如TFLP和DGGE)、磷脂脂肪酸法(PLFA)进行微生物分类和多样性的研究[36],或应用Biology-GN技术和基因芯片技术研究微生物功能群的组成与丰度[50].然而,由于土壤中可培养的微生物仅占很小的一部分、微生物底物来源非常繁杂、磷脂脂肪酸的复杂性和非专一性等原因,使用单一方法很难获取微生物功能群的真实情况.从研究内容上来看,近年来围绕固沙植被演替过程中土壤微生物数量、分类和多样性开展了一些研究[51],发现干旱沙区微生物功能基因丰度随植被重建年限增加而增大,尤其是C、N循环基因丰度的增加促进了微生物的代谢潜力[48]. ...
... 生态系统中C、N、P等元素的生物地球化学循环过程将植被、土壤、土壤微生物和酶有机地联系在一起[41].植物是生产者,植物通过其凋落物和根系及分泌物为土壤微生物和土壤酶提供营养,促进土壤微生物和酶的多样性.而土壤微生物则是主要分解者,可以分解植物及凋落物的残体,对环境起着天然的“过滤”和“净化”作用.同时,土壤微生物能够分泌土壤酶[36],大多数土壤酶是微生物对环境响应表达的产物,能分解植物和微生物的细胞壁,降解大分子,并最终产生可溶性底物供微生物同化.土壤为生物地球化学循环和能量流动过程提供场所,是生态系统稳定性维持的基础.植物从土壤中吸收生长所需的营养物质,再以凋落物的形式将养分元素返还给土壤.植物-凋落物-土壤系统外部的元素能够通过特定微生物和酶进入土壤,植物或凋落物在分解的过程中还受到微生物和酶的共同作用.同时,土壤有机质和有机残体在土壤酶的作用下被分解为不同的产物,为微生物和植物的生长发育提供了物质和能量基础.因此,微生物和酶作为中间媒介,土壤作为载体,共同调控C、N、P在生态系统中的循环,进而影响植物-凋落物-土壤生态化学计量特征的变化(图1). ...
土壤微生物功能群及其研究进展
3
2008
... 土壤微生物根据功能可以划分为不同的类群,依据在C、N、P循环过程中的作用,土壤微生物可划分为纤维素分解菌、氨化细菌、硝化细菌、溶磷菌等生理类群[40].不同功能群在养分循环中生物学功能各异[41].土壤中纤维素降解微生物功能群不仅是土壤碳素循环的主要驱动者,也是维持土壤生物活性、植物生长碳素和能源的重要供给来源[42];氨化和硝化微生物功能群在土壤氮素循环过程中发挥重要作用[42];溶磷微生物功能群可使土壤中难以被植物吸收利用的磷转变成可被利用的磷化物[43].因此,对土壤微生物以功能为单位进行研究,可以更好地掌握生物地球化学循环及生态系统的演替规律,特别是对退化生态系统的恢复及提高其生物学功能均有重要的理论和实践意义[42].同样,环境变化可直接或间接地影响微生物群落结构和多样性,进而对整个生态系统产生影响.大量研究表明,土壤类型、土壤理化性质(水分、温度、pH和C、N、P元素含量等)[44-45]、植被类型及物种多样性等是影响土壤微生物功能群的重要因素[39],而在干旱区土壤微生物对这些因素的变化更加敏感.如张静茹等[46]论述了降雨对干旱半干旱地区土壤微生物数量的直接和间接影响;Liu等[47]在中国北方温带半干旱草原进行增温增雨试验,发现土壤微生物对增温的效应比增雨更强;此外,植被恢复年限、植物种、BSC、季节变化等均被证实能显著影响干旱沙区土壤微生物数量[35,48].近期,Wang等[49]发现随着中国北方干旱沙区优势植物物种变化,N含量是决定土壤细菌群落的关键因素.从研究方法上看,现阶段关于微生物群落结构的研究多采用传统培养法、高通量基因组测序法(如TFLP和DGGE)、磷脂脂肪酸法(PLFA)进行微生物分类和多样性的研究[36],或应用Biology-GN技术和基因芯片技术研究微生物功能群的组成与丰度[50].然而,由于土壤中可培养的微生物仅占很小的一部分、微生物底物来源非常繁杂、磷脂脂肪酸的复杂性和非专一性等原因,使用单一方法很难获取微生物功能群的真实情况.从研究内容上来看,近年来围绕固沙植被演替过程中土壤微生物数量、分类和多样性开展了一些研究[51],发现干旱沙区微生物功能基因丰度随植被重建年限增加而增大,尤其是C、N循环基因丰度的增加促进了微生物的代谢潜力[48]. ...
... [42];溶磷微生物功能群可使土壤中难以被植物吸收利用的磷转变成可被利用的磷化物[43].因此,对土壤微生物以功能为单位进行研究,可以更好地掌握生物地球化学循环及生态系统的演替规律,特别是对退化生态系统的恢复及提高其生物学功能均有重要的理论和实践意义[42].同样,环境变化可直接或间接地影响微生物群落结构和多样性,进而对整个生态系统产生影响.大量研究表明,土壤类型、土壤理化性质(水分、温度、pH和C、N、P元素含量等)[44-45]、植被类型及物种多样性等是影响土壤微生物功能群的重要因素[39],而在干旱区土壤微生物对这些因素的变化更加敏感.如张静茹等[46]论述了降雨对干旱半干旱地区土壤微生物数量的直接和间接影响;Liu等[47]在中国北方温带半干旱草原进行增温增雨试验,发现土壤微生物对增温的效应比增雨更强;此外,植被恢复年限、植物种、BSC、季节变化等均被证实能显著影响干旱沙区土壤微生物数量[35,48].近期,Wang等[49]发现随着中国北方干旱沙区优势植物物种变化,N含量是决定土壤细菌群落的关键因素.从研究方法上看,现阶段关于微生物群落结构的研究多采用传统培养法、高通量基因组测序法(如TFLP和DGGE)、磷脂脂肪酸法(PLFA)进行微生物分类和多样性的研究[36],或应用Biology-GN技术和基因芯片技术研究微生物功能群的组成与丰度[50].然而,由于土壤中可培养的微生物仅占很小的一部分、微生物底物来源非常繁杂、磷脂脂肪酸的复杂性和非专一性等原因,使用单一方法很难获取微生物功能群的真实情况.从研究内容上来看,近年来围绕固沙植被演替过程中土壤微生物数量、分类和多样性开展了一些研究[51],发现干旱沙区微生物功能基因丰度随植被重建年限增加而增大,尤其是C、N循环基因丰度的增加促进了微生物的代谢潜力[48]. ...
... [42].同样,环境变化可直接或间接地影响微生物群落结构和多样性,进而对整个生态系统产生影响.大量研究表明,土壤类型、土壤理化性质(水分、温度、pH和C、N、P元素含量等)[44-45]、植被类型及物种多样性等是影响土壤微生物功能群的重要因素[39],而在干旱区土壤微生物对这些因素的变化更加敏感.如张静茹等[46]论述了降雨对干旱半干旱地区土壤微生物数量的直接和间接影响;Liu等[47]在中国北方温带半干旱草原进行增温增雨试验,发现土壤微生物对增温的效应比增雨更强;此外,植被恢复年限、植物种、BSC、季节变化等均被证实能显著影响干旱沙区土壤微生物数量[35,48].近期,Wang等[49]发现随着中国北方干旱沙区优势植物物种变化,N含量是决定土壤细菌群落的关键因素.从研究方法上看,现阶段关于微生物群落结构的研究多采用传统培养法、高通量基因组测序法(如TFLP和DGGE)、磷脂脂肪酸法(PLFA)进行微生物分类和多样性的研究[36],或应用Biology-GN技术和基因芯片技术研究微生物功能群的组成与丰度[50].然而,由于土壤中可培养的微生物仅占很小的一部分、微生物底物来源非常繁杂、磷脂脂肪酸的复杂性和非专一性等原因,使用单一方法很难获取微生物功能群的真实情况.从研究内容上来看,近年来围绕固沙植被演替过程中土壤微生物数量、分类和多样性开展了一些研究[51],发现干旱沙区微生物功能基因丰度随植被重建年限增加而增大,尤其是C、N循环基因丰度的增加促进了微生物的代谢潜力[48]. ...
Seasonal changes in soil phosphorus and associated microbial properties under adjacent grassland and forest in New Zealand
1
2003
... 土壤微生物根据功能可以划分为不同的类群,依据在C、N、P循环过程中的作用,土壤微生物可划分为纤维素分解菌、氨化细菌、硝化细菌、溶磷菌等生理类群[40].不同功能群在养分循环中生物学功能各异[41].土壤中纤维素降解微生物功能群不仅是土壤碳素循环的主要驱动者,也是维持土壤生物活性、植物生长碳素和能源的重要供给来源[42];氨化和硝化微生物功能群在土壤氮素循环过程中发挥重要作用[42];溶磷微生物功能群可使土壤中难以被植物吸收利用的磷转变成可被利用的磷化物[43].因此,对土壤微生物以功能为单位进行研究,可以更好地掌握生物地球化学循环及生态系统的演替规律,特别是对退化生态系统的恢复及提高其生物学功能均有重要的理论和实践意义[42].同样,环境变化可直接或间接地影响微生物群落结构和多样性,进而对整个生态系统产生影响.大量研究表明,土壤类型、土壤理化性质(水分、温度、pH和C、N、P元素含量等)[44-45]、植被类型及物种多样性等是影响土壤微生物功能群的重要因素[39],而在干旱区土壤微生物对这些因素的变化更加敏感.如张静茹等[46]论述了降雨对干旱半干旱地区土壤微生物数量的直接和间接影响;Liu等[47]在中国北方温带半干旱草原进行增温增雨试验,发现土壤微生物对增温的效应比增雨更强;此外,植被恢复年限、植物种、BSC、季节变化等均被证实能显著影响干旱沙区土壤微生物数量[35,48].近期,Wang等[49]发现随着中国北方干旱沙区优势植物物种变化,N含量是决定土壤细菌群落的关键因素.从研究方法上看,现阶段关于微生物群落结构的研究多采用传统培养法、高通量基因组测序法(如TFLP和DGGE)、磷脂脂肪酸法(PLFA)进行微生物分类和多样性的研究[36],或应用Biology-GN技术和基因芯片技术研究微生物功能群的组成与丰度[50].然而,由于土壤中可培养的微生物仅占很小的一部分、微生物底物来源非常繁杂、磷脂脂肪酸的复杂性和非专一性等原因,使用单一方法很难获取微生物功能群的真实情况.从研究内容上来看,近年来围绕固沙植被演替过程中土壤微生物数量、分类和多样性开展了一些研究[51],发现干旱沙区微生物功能基因丰度随植被重建年限增加而增大,尤其是C、N循环基因丰度的增加促进了微生物的代谢潜力[48]. ...
High throughput sequencing analysis of biogeographical distribution of bacterial communities in the black soils of northeast China
1
2014
... 土壤微生物根据功能可以划分为不同的类群,依据在C、N、P循环过程中的作用,土壤微生物可划分为纤维素分解菌、氨化细菌、硝化细菌、溶磷菌等生理类群[40].不同功能群在养分循环中生物学功能各异[41].土壤中纤维素降解微生物功能群不仅是土壤碳素循环的主要驱动者,也是维持土壤生物活性、植物生长碳素和能源的重要供给来源[42];氨化和硝化微生物功能群在土壤氮素循环过程中发挥重要作用[42];溶磷微生物功能群可使土壤中难以被植物吸收利用的磷转变成可被利用的磷化物[43].因此,对土壤微生物以功能为单位进行研究,可以更好地掌握生物地球化学循环及生态系统的演替规律,特别是对退化生态系统的恢复及提高其生物学功能均有重要的理论和实践意义[42].同样,环境变化可直接或间接地影响微生物群落结构和多样性,进而对整个生态系统产生影响.大量研究表明,土壤类型、土壤理化性质(水分、温度、pH和C、N、P元素含量等)[44-45]、植被类型及物种多样性等是影响土壤微生物功能群的重要因素[39],而在干旱区土壤微生物对这些因素的变化更加敏感.如张静茹等[46]论述了降雨对干旱半干旱地区土壤微生物数量的直接和间接影响;Liu等[47]在中国北方温带半干旱草原进行增温增雨试验,发现土壤微生物对增温的效应比增雨更强;此外,植被恢复年限、植物种、BSC、季节变化等均被证实能显著影响干旱沙区土壤微生物数量[35,48].近期,Wang等[49]发现随着中国北方干旱沙区优势植物物种变化,N含量是决定土壤细菌群落的关键因素.从研究方法上看,现阶段关于微生物群落结构的研究多采用传统培养法、高通量基因组测序法(如TFLP和DGGE)、磷脂脂肪酸法(PLFA)进行微生物分类和多样性的研究[36],或应用Biology-GN技术和基因芯片技术研究微生物功能群的组成与丰度[50].然而,由于土壤中可培养的微生物仅占很小的一部分、微生物底物来源非常繁杂、磷脂脂肪酸的复杂性和非专一性等原因,使用单一方法很难获取微生物功能群的真实情况.从研究内容上来看,近年来围绕固沙植被演替过程中土壤微生物数量、分类和多样性开展了一些研究[51],发现干旱沙区微生物功能基因丰度随植被重建年限增加而增大,尤其是C、N循环基因丰度的增加促进了微生物的代谢潜力[48]. ...
Scale-dependent effects of climate and geographic distance on bacterial diversity patterns across northern China's grasslands
1
2018
... 土壤微生物根据功能可以划分为不同的类群,依据在C、N、P循环过程中的作用,土壤微生物可划分为纤维素分解菌、氨化细菌、硝化细菌、溶磷菌等生理类群[40].不同功能群在养分循环中生物学功能各异[41].土壤中纤维素降解微生物功能群不仅是土壤碳素循环的主要驱动者,也是维持土壤生物活性、植物生长碳素和能源的重要供给来源[42];氨化和硝化微生物功能群在土壤氮素循环过程中发挥重要作用[42];溶磷微生物功能群可使土壤中难以被植物吸收利用的磷转变成可被利用的磷化物[43].因此,对土壤微生物以功能为单位进行研究,可以更好地掌握生物地球化学循环及生态系统的演替规律,特别是对退化生态系统的恢复及提高其生物学功能均有重要的理论和实践意义[42].同样,环境变化可直接或间接地影响微生物群落结构和多样性,进而对整个生态系统产生影响.大量研究表明,土壤类型、土壤理化性质(水分、温度、pH和C、N、P元素含量等)[44-45]、植被类型及物种多样性等是影响土壤微生物功能群的重要因素[39],而在干旱区土壤微生物对这些因素的变化更加敏感.如张静茹等[46]论述了降雨对干旱半干旱地区土壤微生物数量的直接和间接影响;Liu等[47]在中国北方温带半干旱草原进行增温增雨试验,发现土壤微生物对增温的效应比增雨更强;此外,植被恢复年限、植物种、BSC、季节变化等均被证实能显著影响干旱沙区土壤微生物数量[35,48].近期,Wang等[49]发现随着中国北方干旱沙区优势植物物种变化,N含量是决定土壤细菌群落的关键因素.从研究方法上看,现阶段关于微生物群落结构的研究多采用传统培养法、高通量基因组测序法(如TFLP和DGGE)、磷脂脂肪酸法(PLFA)进行微生物分类和多样性的研究[36],或应用Biology-GN技术和基因芯片技术研究微生物功能群的组成与丰度[50].然而,由于土壤中可培养的微生物仅占很小的一部分、微生物底物来源非常繁杂、磷脂脂肪酸的复杂性和非专一性等原因,使用单一方法很难获取微生物功能群的真实情况.从研究内容上来看,近年来围绕固沙植被演替过程中土壤微生物数量、分类和多样性开展了一些研究[51],发现干旱沙区微生物功能基因丰度随植被重建年限增加而增大,尤其是C、N循环基因丰度的增加促进了微生物的代谢潜力[48]. ...
降雨对干旱半干旱地区土壤微生物影响研究进展
1
2014
... 土壤微生物根据功能可以划分为不同的类群,依据在C、N、P循环过程中的作用,土壤微生物可划分为纤维素分解菌、氨化细菌、硝化细菌、溶磷菌等生理类群[40].不同功能群在养分循环中生物学功能各异[41].土壤中纤维素降解微生物功能群不仅是土壤碳素循环的主要驱动者,也是维持土壤生物活性、植物生长碳素和能源的重要供给来源[42];氨化和硝化微生物功能群在土壤氮素循环过程中发挥重要作用[42];溶磷微生物功能群可使土壤中难以被植物吸收利用的磷转变成可被利用的磷化物[43].因此,对土壤微生物以功能为单位进行研究,可以更好地掌握生物地球化学循环及生态系统的演替规律,特别是对退化生态系统的恢复及提高其生物学功能均有重要的理论和实践意义[42].同样,环境变化可直接或间接地影响微生物群落结构和多样性,进而对整个生态系统产生影响.大量研究表明,土壤类型、土壤理化性质(水分、温度、pH和C、N、P元素含量等)[44-45]、植被类型及物种多样性等是影响土壤微生物功能群的重要因素[39],而在干旱区土壤微生物对这些因素的变化更加敏感.如张静茹等[46]论述了降雨对干旱半干旱地区土壤微生物数量的直接和间接影响;Liu等[47]在中国北方温带半干旱草原进行增温增雨试验,发现土壤微生物对增温的效应比增雨更强;此外,植被恢复年限、植物种、BSC、季节变化等均被证实能显著影响干旱沙区土壤微生物数量[35,48].近期,Wang等[49]发现随着中国北方干旱沙区优势植物物种变化,N含量是决定土壤细菌群落的关键因素.从研究方法上看,现阶段关于微生物群落结构的研究多采用传统培养法、高通量基因组测序法(如TFLP和DGGE)、磷脂脂肪酸法(PLFA)进行微生物分类和多样性的研究[36],或应用Biology-GN技术和基因芯片技术研究微生物功能群的组成与丰度[50].然而,由于土壤中可培养的微生物仅占很小的一部分、微生物底物来源非常繁杂、磷脂脂肪酸的复杂性和非专一性等原因,使用单一方法很难获取微生物功能群的真实情况.从研究内容上来看,近年来围绕固沙植被演替过程中土壤微生物数量、分类和多样性开展了一些研究[51],发现干旱沙区微生物功能基因丰度随植被重建年限增加而增大,尤其是C、N循环基因丰度的增加促进了微生物的代谢潜力[48]. ...
Predominant role of water in regulating soil and microbial respiration and their responses to climate change in a semiarid grassland
1
2010
... 土壤微生物根据功能可以划分为不同的类群,依据在C、N、P循环过程中的作用,土壤微生物可划分为纤维素分解菌、氨化细菌、硝化细菌、溶磷菌等生理类群[40].不同功能群在养分循环中生物学功能各异[41].土壤中纤维素降解微生物功能群不仅是土壤碳素循环的主要驱动者,也是维持土壤生物活性、植物生长碳素和能源的重要供给来源[42];氨化和硝化微生物功能群在土壤氮素循环过程中发挥重要作用[42];溶磷微生物功能群可使土壤中难以被植物吸收利用的磷转变成可被利用的磷化物[43].因此,对土壤微生物以功能为单位进行研究,可以更好地掌握生物地球化学循环及生态系统的演替规律,特别是对退化生态系统的恢复及提高其生物学功能均有重要的理论和实践意义[42].同样,环境变化可直接或间接地影响微生物群落结构和多样性,进而对整个生态系统产生影响.大量研究表明,土壤类型、土壤理化性质(水分、温度、pH和C、N、P元素含量等)[44-45]、植被类型及物种多样性等是影响土壤微生物功能群的重要因素[39],而在干旱区土壤微生物对这些因素的变化更加敏感.如张静茹等[46]论述了降雨对干旱半干旱地区土壤微生物数量的直接和间接影响;Liu等[47]在中国北方温带半干旱草原进行增温增雨试验,发现土壤微生物对增温的效应比增雨更强;此外,植被恢复年限、植物种、BSC、季节变化等均被证实能显著影响干旱沙区土壤微生物数量[35,48].近期,Wang等[49]发现随着中国北方干旱沙区优势植物物种变化,N含量是决定土壤细菌群落的关键因素.从研究方法上看,现阶段关于微生物群落结构的研究多采用传统培养法、高通量基因组测序法(如TFLP和DGGE)、磷脂脂肪酸法(PLFA)进行微生物分类和多样性的研究[36],或应用Biology-GN技术和基因芯片技术研究微生物功能群的组成与丰度[50].然而,由于土壤中可培养的微生物仅占很小的一部分、微生物底物来源非常繁杂、磷脂脂肪酸的复杂性和非专一性等原因,使用单一方法很难获取微生物功能群的真实情况.从研究内容上来看,近年来围绕固沙植被演替过程中土壤微生物数量、分类和多样性开展了一些研究[51],发现干旱沙区微生物功能基因丰度随植被重建年限增加而增大,尤其是C、N循环基因丰度的增加促进了微生物的代谢潜力[48]. ...
Changes in functional gene structure and metabolic potential of the microbial community in biological soil crusts along a revegetation chronosequence in the Tengger Desert
2
2018
... 土壤微生物根据功能可以划分为不同的类群,依据在C、N、P循环过程中的作用,土壤微生物可划分为纤维素分解菌、氨化细菌、硝化细菌、溶磷菌等生理类群[40].不同功能群在养分循环中生物学功能各异[41].土壤中纤维素降解微生物功能群不仅是土壤碳素循环的主要驱动者,也是维持土壤生物活性、植物生长碳素和能源的重要供给来源[42];氨化和硝化微生物功能群在土壤氮素循环过程中发挥重要作用[42];溶磷微生物功能群可使土壤中难以被植物吸收利用的磷转变成可被利用的磷化物[43].因此,对土壤微生物以功能为单位进行研究,可以更好地掌握生物地球化学循环及生态系统的演替规律,特别是对退化生态系统的恢复及提高其生物学功能均有重要的理论和实践意义[42].同样,环境变化可直接或间接地影响微生物群落结构和多样性,进而对整个生态系统产生影响.大量研究表明,土壤类型、土壤理化性质(水分、温度、pH和C、N、P元素含量等)[44-45]、植被类型及物种多样性等是影响土壤微生物功能群的重要因素[39],而在干旱区土壤微生物对这些因素的变化更加敏感.如张静茹等[46]论述了降雨对干旱半干旱地区土壤微生物数量的直接和间接影响;Liu等[47]在中国北方温带半干旱草原进行增温增雨试验,发现土壤微生物对增温的效应比增雨更强;此外,植被恢复年限、植物种、BSC、季节变化等均被证实能显著影响干旱沙区土壤微生物数量[35,48].近期,Wang等[49]发现随着中国北方干旱沙区优势植物物种变化,N含量是决定土壤细菌群落的关键因素.从研究方法上看,现阶段关于微生物群落结构的研究多采用传统培养法、高通量基因组测序法(如TFLP和DGGE)、磷脂脂肪酸法(PLFA)进行微生物分类和多样性的研究[36],或应用Biology-GN技术和基因芯片技术研究微生物功能群的组成与丰度[50].然而,由于土壤中可培养的微生物仅占很小的一部分、微生物底物来源非常繁杂、磷脂脂肪酸的复杂性和非专一性等原因,使用单一方法很难获取微生物功能群的真实情况.从研究内容上来看,近年来围绕固沙植被演替过程中土壤微生物数量、分类和多样性开展了一些研究[51],发现干旱沙区微生物功能基因丰度随植被重建年限增加而增大,尤其是C、N循环基因丰度的增加促进了微生物的代谢潜力[48]. ...
... [48]. ...
Dominant plant species shape soil bacterial community in semiarid sandy land of northern China
1
2018
... 土壤微生物根据功能可以划分为不同的类群,依据在C、N、P循环过程中的作用,土壤微生物可划分为纤维素分解菌、氨化细菌、硝化细菌、溶磷菌等生理类群[40].不同功能群在养分循环中生物学功能各异[41].土壤中纤维素降解微生物功能群不仅是土壤碳素循环的主要驱动者,也是维持土壤生物活性、植物生长碳素和能源的重要供给来源[42];氨化和硝化微生物功能群在土壤氮素循环过程中发挥重要作用[42];溶磷微生物功能群可使土壤中难以被植物吸收利用的磷转变成可被利用的磷化物[43].因此,对土壤微生物以功能为单位进行研究,可以更好地掌握生物地球化学循环及生态系统的演替规律,特别是对退化生态系统的恢复及提高其生物学功能均有重要的理论和实践意义[42].同样,环境变化可直接或间接地影响微生物群落结构和多样性,进而对整个生态系统产生影响.大量研究表明,土壤类型、土壤理化性质(水分、温度、pH和C、N、P元素含量等)[44-45]、植被类型及物种多样性等是影响土壤微生物功能群的重要因素[39],而在干旱区土壤微生物对这些因素的变化更加敏感.如张静茹等[46]论述了降雨对干旱半干旱地区土壤微生物数量的直接和间接影响;Liu等[47]在中国北方温带半干旱草原进行增温增雨试验,发现土壤微生物对增温的效应比增雨更强;此外,植被恢复年限、植物种、BSC、季节变化等均被证实能显著影响干旱沙区土壤微生物数量[35,48].近期,Wang等[49]发现随着中国北方干旱沙区优势植物物种变化,N含量是决定土壤细菌群落的关键因素.从研究方法上看,现阶段关于微生物群落结构的研究多采用传统培养法、高通量基因组测序法(如TFLP和DGGE)、磷脂脂肪酸法(PLFA)进行微生物分类和多样性的研究[36],或应用Biology-GN技术和基因芯片技术研究微生物功能群的组成与丰度[50].然而,由于土壤中可培养的微生物仅占很小的一部分、微生物底物来源非常繁杂、磷脂脂肪酸的复杂性和非专一性等原因,使用单一方法很难获取微生物功能群的真实情况.从研究内容上来看,近年来围绕固沙植被演替过程中土壤微生物数量、分类和多样性开展了一些研究[51],发现干旱沙区微生物功能基因丰度随植被重建年限增加而增大,尤其是C、N循环基因丰度的增加促进了微生物的代谢潜力[48]. ...
The microbe-mediated mechanisms affecting topsoil carbon stock in Tibetan grasslands
1
2015
... 土壤微生物根据功能可以划分为不同的类群,依据在C、N、P循环过程中的作用,土壤微生物可划分为纤维素分解菌、氨化细菌、硝化细菌、溶磷菌等生理类群[40].不同功能群在养分循环中生物学功能各异[41].土壤中纤维素降解微生物功能群不仅是土壤碳素循环的主要驱动者,也是维持土壤生物活性、植物生长碳素和能源的重要供给来源[42];氨化和硝化微生物功能群在土壤氮素循环过程中发挥重要作用[42];溶磷微生物功能群可使土壤中难以被植物吸收利用的磷转变成可被利用的磷化物[43].因此,对土壤微生物以功能为单位进行研究,可以更好地掌握生物地球化学循环及生态系统的演替规律,特别是对退化生态系统的恢复及提高其生物学功能均有重要的理论和实践意义[42].同样,环境变化可直接或间接地影响微生物群落结构和多样性,进而对整个生态系统产生影响.大量研究表明,土壤类型、土壤理化性质(水分、温度、pH和C、N、P元素含量等)[44-45]、植被类型及物种多样性等是影响土壤微生物功能群的重要因素[39],而在干旱区土壤微生物对这些因素的变化更加敏感.如张静茹等[46]论述了降雨对干旱半干旱地区土壤微生物数量的直接和间接影响;Liu等[47]在中国北方温带半干旱草原进行增温增雨试验,发现土壤微生物对增温的效应比增雨更强;此外,植被恢复年限、植物种、BSC、季节变化等均被证实能显著影响干旱沙区土壤微生物数量[35,48].近期,Wang等[49]发现随着中国北方干旱沙区优势植物物种变化,N含量是决定土壤细菌群落的关键因素.从研究方法上看,现阶段关于微生物群落结构的研究多采用传统培养法、高通量基因组测序法(如TFLP和DGGE)、磷脂脂肪酸法(PLFA)进行微生物分类和多样性的研究[36],或应用Biology-GN技术和基因芯片技术研究微生物功能群的组成与丰度[50].然而,由于土壤中可培养的微生物仅占很小的一部分、微生物底物来源非常繁杂、磷脂脂肪酸的复杂性和非专一性等原因,使用单一方法很难获取微生物功能群的真实情况.从研究内容上来看,近年来围绕固沙植被演替过程中土壤微生物数量、分类和多样性开展了一些研究[51],发现干旱沙区微生物功能基因丰度随植被重建年限增加而增大,尤其是C、N循环基因丰度的增加促进了微生物的代谢潜力[48]. ...
Fungal-bacterial diversity and microbiome complexity predict ecosystem functioning
1
2019
... 土壤微生物根据功能可以划分为不同的类群,依据在C、N、P循环过程中的作用,土壤微生物可划分为纤维素分解菌、氨化细菌、硝化细菌、溶磷菌等生理类群[40].不同功能群在养分循环中生物学功能各异[41].土壤中纤维素降解微生物功能群不仅是土壤碳素循环的主要驱动者,也是维持土壤生物活性、植物生长碳素和能源的重要供给来源[42];氨化和硝化微生物功能群在土壤氮素循环过程中发挥重要作用[42];溶磷微生物功能群可使土壤中难以被植物吸收利用的磷转变成可被利用的磷化物[43].因此,对土壤微生物以功能为单位进行研究,可以更好地掌握生物地球化学循环及生态系统的演替规律,特别是对退化生态系统的恢复及提高其生物学功能均有重要的理论和实践意义[42].同样,环境变化可直接或间接地影响微生物群落结构和多样性,进而对整个生态系统产生影响.大量研究表明,土壤类型、土壤理化性质(水分、温度、pH和C、N、P元素含量等)[44-45]、植被类型及物种多样性等是影响土壤微生物功能群的重要因素[39],而在干旱区土壤微生物对这些因素的变化更加敏感.如张静茹等[46]论述了降雨对干旱半干旱地区土壤微生物数量的直接和间接影响;Liu等[47]在中国北方温带半干旱草原进行增温增雨试验,发现土壤微生物对增温的效应比增雨更强;此外,植被恢复年限、植物种、BSC、季节变化等均被证实能显著影响干旱沙区土壤微生物数量[35,48].近期,Wang等[49]发现随着中国北方干旱沙区优势植物物种变化,N含量是决定土壤细菌群落的关键因素.从研究方法上看,现阶段关于微生物群落结构的研究多采用传统培养法、高通量基因组测序法(如TFLP和DGGE)、磷脂脂肪酸法(PLFA)进行微生物分类和多样性的研究[36],或应用Biology-GN技术和基因芯片技术研究微生物功能群的组成与丰度[50].然而,由于土壤中可培养的微生物仅占很小的一部分、微生物底物来源非常繁杂、磷脂脂肪酸的复杂性和非专一性等原因,使用单一方法很难获取微生物功能群的真实情况.从研究内容上来看,近年来围绕固沙植被演替过程中土壤微生物数量、分类和多样性开展了一些研究[51],发现干旱沙区微生物功能基因丰度随植被重建年限增加而增大,尤其是C、N循环基因丰度的增加促进了微生物的代谢潜力[48]. ...
微生物对分解底物碳氮磷化学计量的响应和调节机制
1
2016
... 土壤微生物除了通过调整或改变自身的C∶N∶P功能群结构来响应底物的变异,还可通过调整胞外酶的比例来适应底物的变化[52-53].大多数土壤酶是微生物对环境响应表达的产物,是一种数量极微而作用极大的土壤组成部分,它能分解植物和微生物的细胞壁,解聚大分子,并最终产生可溶性底物供微生物同化.土壤中酶的种类十分繁杂且功能各异.从代谢功能来看,与土壤有机质转化有关的土壤酶主要有:纤维素酶,能够催化土壤中纤维素的水解;木聚糖酶,能够水解植物残体中的半纤维素;蔗糖酶,主要对水溶性蔗糖进行水解并产生果糖和葡萄糖.人们常用木质素分解酶、纤维素分解酶、多糖分解酶来指示土壤碳的转换[54].与有机氮转化相关的酶主要包括:几丁质酶,目前研究主要集中于β-乙酰氨基葡萄糖苷酶;蛋白酶,主要分解肽类、氨基酸和蛋白质;肽酶,能够从肽链的末端水解形成氨基酸,其主要研究对象是氨基肽酶;脲酶,能够水解尿素,同时生成CO2和水.另外,研究者常使用生物固氮酶、硝化酶和氨同化酶来指示土壤N的循环过程[55].土壤中有机磷的转化主要依靠磷酸酶来完成,它水解磷酸脂类形成磷酸及其他产物[56]. ...
Microbial community dynamics alleviate stoichiometric constraints during litter decay
1
2014
... 土壤微生物除了通过调整或改变自身的C∶N∶P功能群结构来响应底物的变异,还可通过调整胞外酶的比例来适应底物的变化[52-53].大多数土壤酶是微生物对环境响应表达的产物,是一种数量极微而作用极大的土壤组成部分,它能分解植物和微生物的细胞壁,解聚大分子,并最终产生可溶性底物供微生物同化.土壤中酶的种类十分繁杂且功能各异.从代谢功能来看,与土壤有机质转化有关的土壤酶主要有:纤维素酶,能够催化土壤中纤维素的水解;木聚糖酶,能够水解植物残体中的半纤维素;蔗糖酶,主要对水溶性蔗糖进行水解并产生果糖和葡萄糖.人们常用木质素分解酶、纤维素分解酶、多糖分解酶来指示土壤碳的转换[54].与有机氮转化相关的酶主要包括:几丁质酶,目前研究主要集中于β-乙酰氨基葡萄糖苷酶;蛋白酶,主要分解肽类、氨基酸和蛋白质;肽酶,能够从肽链的末端水解形成氨基酸,其主要研究对象是氨基肽酶;脲酶,能够水解尿素,同时生成CO2和水.另外,研究者常使用生物固氮酶、硝化酶和氨同化酶来指示土壤N的循环过程[55].土壤中有机磷的转化主要依靠磷酸酶来完成,它水解磷酸脂类形成磷酸及其他产物[56]. ...
Soil enzyme activity and stoichiometry in forest ecosystems along the North-South Transect in eastern China
1
2017
... 土壤微生物除了通过调整或改变自身的C∶N∶P功能群结构来响应底物的变异,还可通过调整胞外酶的比例来适应底物的变化[52-53].大多数土壤酶是微生物对环境响应表达的产物,是一种数量极微而作用极大的土壤组成部分,它能分解植物和微生物的细胞壁,解聚大分子,并最终产生可溶性底物供微生物同化.土壤中酶的种类十分繁杂且功能各异.从代谢功能来看,与土壤有机质转化有关的土壤酶主要有:纤维素酶,能够催化土壤中纤维素的水解;木聚糖酶,能够水解植物残体中的半纤维素;蔗糖酶,主要对水溶性蔗糖进行水解并产生果糖和葡萄糖.人们常用木质素分解酶、纤维素分解酶、多糖分解酶来指示土壤碳的转换[54].与有机氮转化相关的酶主要包括:几丁质酶,目前研究主要集中于β-乙酰氨基葡萄糖苷酶;蛋白酶,主要分解肽类、氨基酸和蛋白质;肽酶,能够从肽链的末端水解形成氨基酸,其主要研究对象是氨基肽酶;脲酶,能够水解尿素,同时生成CO2和水.另外,研究者常使用生物固氮酶、硝化酶和氨同化酶来指示土壤N的循环过程[55].土壤中有机磷的转化主要依靠磷酸酶来完成,它水解磷酸脂类形成磷酸及其他产物[56]. ...
荒漠草原不同种植年限人工柠条林土壤碳氮磷化学计量特征
1
2015
... 土壤微生物除了通过调整或改变自身的C∶N∶P功能群结构来响应底物的变异,还可通过调整胞外酶的比例来适应底物的变化[52-53].大多数土壤酶是微生物对环境响应表达的产物,是一种数量极微而作用极大的土壤组成部分,它能分解植物和微生物的细胞壁,解聚大分子,并最终产生可溶性底物供微生物同化.土壤中酶的种类十分繁杂且功能各异.从代谢功能来看,与土壤有机质转化有关的土壤酶主要有:纤维素酶,能够催化土壤中纤维素的水解;木聚糖酶,能够水解植物残体中的半纤维素;蔗糖酶,主要对水溶性蔗糖进行水解并产生果糖和葡萄糖.人们常用木质素分解酶、纤维素分解酶、多糖分解酶来指示土壤碳的转换[54].与有机氮转化相关的酶主要包括:几丁质酶,目前研究主要集中于β-乙酰氨基葡萄糖苷酶;蛋白酶,主要分解肽类、氨基酸和蛋白质;肽酶,能够从肽链的末端水解形成氨基酸,其主要研究对象是氨基肽酶;脲酶,能够水解尿素,同时生成CO2和水.另外,研究者常使用生物固氮酶、硝化酶和氨同化酶来指示土壤N的循环过程[55].土壤中有机磷的转化主要依靠磷酸酶来完成,它水解磷酸脂类形成磷酸及其他产物[56]. ...
我国土壤水解酶活性研究进展
1
2012
... 土壤微生物除了通过调整或改变自身的C∶N∶P功能群结构来响应底物的变异,还可通过调整胞外酶的比例来适应底物的变化[52-53].大多数土壤酶是微生物对环境响应表达的产物,是一种数量极微而作用极大的土壤组成部分,它能分解植物和微生物的细胞壁,解聚大分子,并最终产生可溶性底物供微生物同化.土壤中酶的种类十分繁杂且功能各异.从代谢功能来看,与土壤有机质转化有关的土壤酶主要有:纤维素酶,能够催化土壤中纤维素的水解;木聚糖酶,能够水解植物残体中的半纤维素;蔗糖酶,主要对水溶性蔗糖进行水解并产生果糖和葡萄糖.人们常用木质素分解酶、纤维素分解酶、多糖分解酶来指示土壤碳的转换[54].与有机氮转化相关的酶主要包括:几丁质酶,目前研究主要集中于β-乙酰氨基葡萄糖苷酶;蛋白酶,主要分解肽类、氨基酸和蛋白质;肽酶,能够从肽链的末端水解形成氨基酸,其主要研究对象是氨基肽酶;脲酶,能够水解尿素,同时生成CO2和水.另外,研究者常使用生物固氮酶、硝化酶和氨同化酶来指示土壤N的循环过程[55].土壤中有机磷的转化主要依靠磷酸酶来完成,它水解磷酸脂类形成磷酸及其他产物[56]. ...
Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment
4
2010
... 通常情况下,β-1,4葡萄糖苷酶(BG)、β-1,4乙酰胺糖苷酶(NAG)、亮氨酸氨基肽酶(LAP)和酸性(碱性)磷酸酶(AP)常用来对微生物代谢、生物地球化学循环过程及微生物养分需求进行评价.BG与土壤转化酶一样能够用来表征土壤中的C循环,可降解纤维素,水解来自纤维素二糖的葡萄糖;NAG可降解几丁质和肽聚糖,水解来自于壳二糖的氨基葡萄糖;LAP可水解蛋白质,水解亮氨酸和来自多肽氮末端的疏水氨基酸,是与NAG一样共同参与土壤N循环的重要物质;AP可水解来自磷酸多糖和磷脂中的磷酸盐.近年来,以Sinsabaugh为代表的酶学家认为这4种酶可以用来反映土壤中C、N、P的转化利用状况,提出了“生态酶化学计量”的概念,即用BG∶(NAG+LAP)∶AP来反映微生物生物量C∶N∶P与底物C∶N∶P以及与微生物的同化和生长效率之间的联系[57-58].一般来讲,植被组成、土壤养分、微生物数量都会影响土壤酶活性,如沙区人工植被土壤酶活性与土壤有机C、N、P、MBC、MBN、微生物数量显著正相关[59].Sinsabaugh等[57]采用Meta分析发现BG、NAG和AP活性随着土壤有机质的增加而逐渐提高,在总体上BG∶(NAG+LAP)∶AP接近于1∶1∶1. ...
... [57]采用Meta分析发现BG、NAG和AP活性随着土壤有机质的增加而逐渐提高,在总体上BG∶(NAG+LAP)∶AP接近于1∶1∶1. ...
... 研究发现,全球森林生态系统植物叶片C∶N∶P为1 212∶28∶1,而当植物叶片凋亡后,养分的再吸收过程使得枯叶的C∶N∶P变为3 007∶45∶1[60];植物凋亡后根的C∶N∶P平均值为4 184∶43∶1[61];土壤C∶N∶P最低为287∶17∶1[62];土壤微生物量C∶N∶P平均值为42∶6∶1,并随土壤深度增加显著减小[26];Sinsabaugh等[57]发现土壤酶中C∶N∶P接近于1∶1∶1.相比之下,从植物叶片、凋落物到土壤再到土壤微生物及酶的计量特征C、N元素整体处于减少趋势.Xu等[37]研究发现,在区域尺度上,随着生物区系不同,C∶N∶P存在很大变异性,如天然湿地中的C∶N∶P为131∶14∶1,而沙漠中的C∶N∶P则为31∶4∶1.面对底物和微生物化学计量间的巨大差异,微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例和元素利用效率等机制来响应和调节.目前为止,关于植物-土壤-微生物-酶C、N、P关系的研究多聚焦于探讨土壤微生物与土壤、植被间的变化及其影响因素.研究表明,随着纬度、年均温、年降水量的变化,微生物量N∶P与植物组织的N∶P呈现出一致的变化趋势,即低纬度地区更受P的限制[63].这表明在高纬度地区微生物和植物一样由于生长季短而需要更高的生长速率,进而需要高的N、P含量和低的N∶P维持[6].Li等[64]分析了中国亚热带地区土壤数据后认为,微生物量C∶N和C∶P会随土壤C∶N和C∶P的增加而增加;周正虎等[38]却发现中国森林土壤微生物量C∶N随着土壤C∶N的增加而减小,与Cleveland等[65]对全球森林的数据分析结果一致.近期,吕星宇等[35]在腾格里沙漠开展的人工固沙植被研究表明,固沙植被演替过程中,微生物功能群数量C∶N∶P的变化主要受到草本、灌木和土壤C∶N∶P变化的影响.受灌木C∶N∶P增加及草本C∶N∶P减小的影响,微生物功能群数量C∶N∶P随之增加;对土壤酶C∶N∶P影响最大的因子分别是微生物功能群数量C∶N∶P和凋落物C∶N∶P,土壤酶C∶N∶P随着微生物功能群数量C∶N∶P增加而增加. ...
... 一般来讲,植被组成、土壤养分、微生物数量都会影响土壤酶活性[57].由于土壤酶C∶N∶P的研究才刚刚起步,尤其针对荒漠生态系统植被演替过程中土壤酶的生态化学计量的研究还远远不够.那么,以BG、NAG、LAP和AP作为C、N、P的特征酶在荒漠生态系统是否具有代表性还需要进一步的验证,其化学计量特征在固沙植被演替过程中有什么规律仍有待探索.其次,对荒漠人工植被演替过程中土壤酶C∶N∶P特征时空变化,可通过底物添加控制试验,评估土壤酶活性对不同植物凋落物添加的敏感性,建立凋落物属性同土壤、土壤微生物功能群和酶的C∶N∶P之间的关系,结合凋落物分解速率揭示底物影响土壤酶C∶N∶P的机理. ...
Ecoenzymatic stoichiometry of recalcitrant organic matter decomposition:the growth rate hypothesis in reverse
1
2011
... 通常情况下,β-1,4葡萄糖苷酶(BG)、β-1,4乙酰胺糖苷酶(NAG)、亮氨酸氨基肽酶(LAP)和酸性(碱性)磷酸酶(AP)常用来对微生物代谢、生物地球化学循环过程及微生物养分需求进行评价.BG与土壤转化酶一样能够用来表征土壤中的C循环,可降解纤维素,水解来自纤维素二糖的葡萄糖;NAG可降解几丁质和肽聚糖,水解来自于壳二糖的氨基葡萄糖;LAP可水解蛋白质,水解亮氨酸和来自多肽氮末端的疏水氨基酸,是与NAG一样共同参与土壤N循环的重要物质;AP可水解来自磷酸多糖和磷脂中的磷酸盐.近年来,以Sinsabaugh为代表的酶学家认为这4种酶可以用来反映土壤中C、N、P的转化利用状况,提出了“生态酶化学计量”的概念,即用BG∶(NAG+LAP)∶AP来反映微生物生物量C∶N∶P与底物C∶N∶P以及与微生物的同化和生长效率之间的联系[57-58].一般来讲,植被组成、土壤养分、微生物数量都会影响土壤酶活性,如沙区人工植被土壤酶活性与土壤有机C、N、P、MBC、MBN、微生物数量显著正相关[59].Sinsabaugh等[57]采用Meta分析发现BG、NAG和AP活性随着土壤有机质的增加而逐渐提高,在总体上BG∶(NAG+LAP)∶AP接近于1∶1∶1. ...
科尔沁沙地土壤酶活性与植被的关系
1
2006
... 通常情况下,β-1,4葡萄糖苷酶(BG)、β-1,4乙酰胺糖苷酶(NAG)、亮氨酸氨基肽酶(LAP)和酸性(碱性)磷酸酶(AP)常用来对微生物代谢、生物地球化学循环过程及微生物养分需求进行评价.BG与土壤转化酶一样能够用来表征土壤中的C循环,可降解纤维素,水解来自纤维素二糖的葡萄糖;NAG可降解几丁质和肽聚糖,水解来自于壳二糖的氨基葡萄糖;LAP可水解蛋白质,水解亮氨酸和来自多肽氮末端的疏水氨基酸,是与NAG一样共同参与土壤N循环的重要物质;AP可水解来自磷酸多糖和磷脂中的磷酸盐.近年来,以Sinsabaugh为代表的酶学家认为这4种酶可以用来反映土壤中C、N、P的转化利用状况,提出了“生态酶化学计量”的概念,即用BG∶(NAG+LAP)∶AP来反映微生物生物量C∶N∶P与底物C∶N∶P以及与微生物的同化和生长效率之间的联系[57-58].一般来讲,植被组成、土壤养分、微生物数量都会影响土壤酶活性,如沙区人工植被土壤酶活性与土壤有机C、N、P、MBC、MBN、微生物数量显著正相关[59].Sinsabaugh等[57]采用Meta分析发现BG、NAG和AP活性随着土壤有机质的增加而逐渐提高,在总体上BG∶(NAG+LAP)∶AP接近于1∶1∶1. ...
Correction:relationship between the relative limitation and resorption efficiency of nitrogen vs phosphorus in woody plants
1
2014
... 研究发现,全球森林生态系统植物叶片C∶N∶P为1 212∶28∶1,而当植物叶片凋亡后,养分的再吸收过程使得枯叶的C∶N∶P变为3 007∶45∶1[60];植物凋亡后根的C∶N∶P平均值为4 184∶43∶1[61];土壤C∶N∶P最低为287∶17∶1[62];土壤微生物量C∶N∶P平均值为42∶6∶1,并随土壤深度增加显著减小[26];Sinsabaugh等[57]发现土壤酶中C∶N∶P接近于1∶1∶1.相比之下,从植物叶片、凋落物到土壤再到土壤微生物及酶的计量特征C、N元素整体处于减少趋势.Xu等[37]研究发现,在区域尺度上,随着生物区系不同,C∶N∶P存在很大变异性,如天然湿地中的C∶N∶P为131∶14∶1,而沙漠中的C∶N∶P则为31∶4∶1.面对底物和微生物化学计量间的巨大差异,微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例和元素利用效率等机制来响应和调节.目前为止,关于植物-土壤-微生物-酶C、N、P关系的研究多聚焦于探讨土壤微生物与土壤、植被间的变化及其影响因素.研究表明,随着纬度、年均温、年降水量的变化,微生物量N∶P与植物组织的N∶P呈现出一致的变化趋势,即低纬度地区更受P的限制[63].这表明在高纬度地区微生物和植物一样由于生长季短而需要更高的生长速率,进而需要高的N、P含量和低的N∶P维持[6].Li等[64]分析了中国亚热带地区土壤数据后认为,微生物量C∶N和C∶P会随土壤C∶N和C∶P的增加而增加;周正虎等[38]却发现中国森林土壤微生物量C∶N随着土壤C∶N的增加而减小,与Cleveland等[65]对全球森林的数据分析结果一致.近期,吕星宇等[35]在腾格里沙漠开展的人工固沙植被研究表明,固沙植被演替过程中,微生物功能群数量C∶N∶P的变化主要受到草本、灌木和土壤C∶N∶P变化的影响.受灌木C∶N∶P增加及草本C∶N∶P减小的影响,微生物功能群数量C∶N∶P随之增加;对土壤酶C∶N∶P影响最大的因子分别是微生物功能群数量C∶N∶P和凋落物C∶N∶P,土壤酶C∶N∶P随着微生物功能群数量C∶N∶P增加而增加. ...
Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus
1
2011
... 研究发现,全球森林生态系统植物叶片C∶N∶P为1 212∶28∶1,而当植物叶片凋亡后,养分的再吸收过程使得枯叶的C∶N∶P变为3 007∶45∶1[60];植物凋亡后根的C∶N∶P平均值为4 184∶43∶1[61];土壤C∶N∶P最低为287∶17∶1[62];土壤微生物量C∶N∶P平均值为42∶6∶1,并随土壤深度增加显著减小[26];Sinsabaugh等[57]发现土壤酶中C∶N∶P接近于1∶1∶1.相比之下,从植物叶片、凋落物到土壤再到土壤微生物及酶的计量特征C、N元素整体处于减少趋势.Xu等[37]研究发现,在区域尺度上,随着生物区系不同,C∶N∶P存在很大变异性,如天然湿地中的C∶N∶P为131∶14∶1,而沙漠中的C∶N∶P则为31∶4∶1.面对底物和微生物化学计量间的巨大差异,微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例和元素利用效率等机制来响应和调节.目前为止,关于植物-土壤-微生物-酶C、N、P关系的研究多聚焦于探讨土壤微生物与土壤、植被间的变化及其影响因素.研究表明,随着纬度、年均温、年降水量的变化,微生物量N∶P与植物组织的N∶P呈现出一致的变化趋势,即低纬度地区更受P的限制[63].这表明在高纬度地区微生物和植物一样由于生长季短而需要更高的生长速率,进而需要高的N、P含量和低的N∶P维持[6].Li等[64]分析了中国亚热带地区土壤数据后认为,微生物量C∶N和C∶P会随土壤C∶N和C∶P的增加而增加;周正虎等[38]却发现中国森林土壤微生物量C∶N随着土壤C∶N的增加而减小,与Cleveland等[65]对全球森林的数据分析结果一致.近期,吕星宇等[35]在腾格里沙漠开展的人工固沙植被研究表明,固沙植被演替过程中,微生物功能群数量C∶N∶P的变化主要受到草本、灌木和土壤C∶N∶P变化的影响.受灌木C∶N∶P增加及草本C∶N∶P减小的影响,微生物功能群数量C∶N∶P随之增加;对土壤酶C∶N∶P影响最大的因子分别是微生物功能群数量C∶N∶P和凋落物C∶N∶P,土壤酶C∶N∶P随着微生物功能群数量C∶N∶P增加而增加. ...
Differential responses of soil hydrolytic and oxidative enzyme activities to the natural forest conversion
1
2020
... 研究发现,全球森林生态系统植物叶片C∶N∶P为1 212∶28∶1,而当植物叶片凋亡后,养分的再吸收过程使得枯叶的C∶N∶P变为3 007∶45∶1[60];植物凋亡后根的C∶N∶P平均值为4 184∶43∶1[61];土壤C∶N∶P最低为287∶17∶1[62];土壤微生物量C∶N∶P平均值为42∶6∶1,并随土壤深度增加显著减小[26];Sinsabaugh等[57]发现土壤酶中C∶N∶P接近于1∶1∶1.相比之下,从植物叶片、凋落物到土壤再到土壤微生物及酶的计量特征C、N元素整体处于减少趋势.Xu等[37]研究发现,在区域尺度上,随着生物区系不同,C∶N∶P存在很大变异性,如天然湿地中的C∶N∶P为131∶14∶1,而沙漠中的C∶N∶P则为31∶4∶1.面对底物和微生物化学计量间的巨大差异,微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例和元素利用效率等机制来响应和调节.目前为止,关于植物-土壤-微生物-酶C、N、P关系的研究多聚焦于探讨土壤微生物与土壤、植被间的变化及其影响因素.研究表明,随着纬度、年均温、年降水量的变化,微生物量N∶P与植物组织的N∶P呈现出一致的变化趋势,即低纬度地区更受P的限制[63].这表明在高纬度地区微生物和植物一样由于生长季短而需要更高的生长速率,进而需要高的N、P含量和低的N∶P维持[6].Li等[64]分析了中国亚热带地区土壤数据后认为,微生物量C∶N和C∶P会随土壤C∶N和C∶P的增加而增加;周正虎等[38]却发现中国森林土壤微生物量C∶N随着土壤C∶N的增加而减小,与Cleveland等[65]对全球森林的数据分析结果一致.近期,吕星宇等[35]在腾格里沙漠开展的人工固沙植被研究表明,固沙植被演替过程中,微生物功能群数量C∶N∶P的变化主要受到草本、灌木和土壤C∶N∶P变化的影响.受灌木C∶N∶P增加及草本C∶N∶P减小的影响,微生物功能群数量C∶N∶P随之增加;对土壤酶C∶N∶P影响最大的因子分别是微生物功能群数量C∶N∶P和凋落物C∶N∶P,土壤酶C∶N∶P随着微生物功能群数量C∶N∶P增加而增加. ...
Ecological restoration and recovery in the wind-blown sand hazard areas of northern China:relationship between soil water and carrying capacity for vegetation in the Tengger Desert
3
2014
... 研究发现,全球森林生态系统植物叶片C∶N∶P为1 212∶28∶1,而当植物叶片凋亡后,养分的再吸收过程使得枯叶的C∶N∶P变为3 007∶45∶1[60];植物凋亡后根的C∶N∶P平均值为4 184∶43∶1[61];土壤C∶N∶P最低为287∶17∶1[62];土壤微生物量C∶N∶P平均值为42∶6∶1,并随土壤深度增加显著减小[26];Sinsabaugh等[57]发现土壤酶中C∶N∶P接近于1∶1∶1.相比之下,从植物叶片、凋落物到土壤再到土壤微生物及酶的计量特征C、N元素整体处于减少趋势.Xu等[37]研究发现,在区域尺度上,随着生物区系不同,C∶N∶P存在很大变异性,如天然湿地中的C∶N∶P为131∶14∶1,而沙漠中的C∶N∶P则为31∶4∶1.面对底物和微生物化学计量间的巨大差异,微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例和元素利用效率等机制来响应和调节.目前为止,关于植物-土壤-微生物-酶C、N、P关系的研究多聚焦于探讨土壤微生物与土壤、植被间的变化及其影响因素.研究表明,随着纬度、年均温、年降水量的变化,微生物量N∶P与植物组织的N∶P呈现出一致的变化趋势,即低纬度地区更受P的限制[63].这表明在高纬度地区微生物和植物一样由于生长季短而需要更高的生长速率,进而需要高的N、P含量和低的N∶P维持[6].Li等[64]分析了中国亚热带地区土壤数据后认为,微生物量C∶N和C∶P会随土壤C∶N和C∶P的增加而增加;周正虎等[38]却发现中国森林土壤微生物量C∶N随着土壤C∶N的增加而减小,与Cleveland等[65]对全球森林的数据分析结果一致.近期,吕星宇等[35]在腾格里沙漠开展的人工固沙植被研究表明,固沙植被演替过程中,微生物功能群数量C∶N∶P的变化主要受到草本、灌木和土壤C∶N∶P变化的影响.受灌木C∶N∶P增加及草本C∶N∶P减小的影响,微生物功能群数量C∶N∶P随之增加;对土壤酶C∶N∶P影响最大的因子分别是微生物功能群数量C∶N∶P和凋落物C∶N∶P,土壤酶C∶N∶P随着微生物功能群数量C∶N∶P增加而增加. ...
... 植被重建是干旱半干旱荒漠区防止土地沙化、减少风沙危害、促进生态恢复的重要途径[63].中国利用植物固沙已有60多年的历史,国家先后在风沙危害区启动了“三北”防护林建设、退耕还林还草和京津风沙源治理等以人工植被建设作为主要修复措施的一批重大生态建设工程.60余年来,中国沙区植被建设取得了举世瞩目的成就,有效遏制了沙漠化的发展,促进了区域生境恢复[66].人工固沙植被是荒漠生态系统重要的组成部分,是土地荒漠化防治和水土流失治理的主要调控者[66].这些先后建立的人工植被为我们进行荒漠生态系统恢复过程中化学计量学研究奠定了基础.以沙坡头为例,人工固沙植被建立后,原有的流沙演变成一个复杂的人工-天然复合的荒漠生态系统.土壤性质的改善和成土作用的加剧,不但使得流沙逐渐向地带性土壤演变,而且促使生物地球化学循环逐渐集中在浅表层[35].此外,随着土壤肥力的提升,植被也逐渐向天然植物群落演变,植物多样性增加的同时,鸟类、动物及昆虫也明显增多[64].开展人工固沙植被生态系统C∶N∶P化学计量的研究,是理解固沙植被系统C、N、P循环过程及植被、微生物和土壤三者之间互馈关系的核心和关键,也是进行科学防沙治沙和管理固沙植被的指导依据.然而,针对沙区人工植被建立后土壤微生物功能群及其化学计量学的研究目前相对较少,需加强人工固沙植被生态系统演替进程中C∶N∶P化学计量的研究. ...
... BSC是由隐花植物(如蓝藻、地衣、苔藓)和土壤微生物及其他相关生物体通过分泌物、菌丝体和假根等与土壤表层颗粒胶结而成的复合体,是荒漠地表景观的重要组成[66].大多数荒漠地区受到非生物因子(如水分匮乏)的调控和胁迫,使得地表维管束植物群落呈斑块状分布,而这种分布格局为BSC在荒漠地区的繁殖和覆盖提供了合适的空间和适宜的生态位,使BSC广泛分布于荒漠地区[63].BSC对于荒漠景观过程,土壤生态、水文、生物过程,地球化学循环及生态修复过程起着重要作用[67].由于目前还没有针对BSC或隐花植物C∶N∶P计量的研究,缺乏BSC对土壤C、N、P响应的生态化学计量认识.因此,开展BSC或隐花植物C∶N∶P化学计量特征的研究,有助于揭示荒漠生态系统植被演替过程中C、N、P的变化规律. ...
The effects of sand stabilization and revegetation on cryptogam species diversity and soil fertility in the Tengger Desert,Northern China
2
2003
... 研究发现,全球森林生态系统植物叶片C∶N∶P为1 212∶28∶1,而当植物叶片凋亡后,养分的再吸收过程使得枯叶的C∶N∶P变为3 007∶45∶1[60];植物凋亡后根的C∶N∶P平均值为4 184∶43∶1[61];土壤C∶N∶P最低为287∶17∶1[62];土壤微生物量C∶N∶P平均值为42∶6∶1,并随土壤深度增加显著减小[26];Sinsabaugh等[57]发现土壤酶中C∶N∶P接近于1∶1∶1.相比之下,从植物叶片、凋落物到土壤再到土壤微生物及酶的计量特征C、N元素整体处于减少趋势.Xu等[37]研究发现,在区域尺度上,随着生物区系不同,C∶N∶P存在很大变异性,如天然湿地中的C∶N∶P为131∶14∶1,而沙漠中的C∶N∶P则为31∶4∶1.面对底物和微生物化学计量间的巨大差异,微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例和元素利用效率等机制来响应和调节.目前为止,关于植物-土壤-微生物-酶C、N、P关系的研究多聚焦于探讨土壤微生物与土壤、植被间的变化及其影响因素.研究表明,随着纬度、年均温、年降水量的变化,微生物量N∶P与植物组织的N∶P呈现出一致的变化趋势,即低纬度地区更受P的限制[63].这表明在高纬度地区微生物和植物一样由于生长季短而需要更高的生长速率,进而需要高的N、P含量和低的N∶P维持[6].Li等[64]分析了中国亚热带地区土壤数据后认为,微生物量C∶N和C∶P会随土壤C∶N和C∶P的增加而增加;周正虎等[38]却发现中国森林土壤微生物量C∶N随着土壤C∶N的增加而减小,与Cleveland等[65]对全球森林的数据分析结果一致.近期,吕星宇等[35]在腾格里沙漠开展的人工固沙植被研究表明,固沙植被演替过程中,微生物功能群数量C∶N∶P的变化主要受到草本、灌木和土壤C∶N∶P变化的影响.受灌木C∶N∶P增加及草本C∶N∶P减小的影响,微生物功能群数量C∶N∶P随之增加;对土壤酶C∶N∶P影响最大的因子分别是微生物功能群数量C∶N∶P和凋落物C∶N∶P,土壤酶C∶N∶P随着微生物功能群数量C∶N∶P增加而增加. ...
... 植被重建是干旱半干旱荒漠区防止土地沙化、减少风沙危害、促进生态恢复的重要途径[63].中国利用植物固沙已有60多年的历史,国家先后在风沙危害区启动了“三北”防护林建设、退耕还林还草和京津风沙源治理等以人工植被建设作为主要修复措施的一批重大生态建设工程.60余年来,中国沙区植被建设取得了举世瞩目的成就,有效遏制了沙漠化的发展,促进了区域生境恢复[66].人工固沙植被是荒漠生态系统重要的组成部分,是土地荒漠化防治和水土流失治理的主要调控者[66].这些先后建立的人工植被为我们进行荒漠生态系统恢复过程中化学计量学研究奠定了基础.以沙坡头为例,人工固沙植被建立后,原有的流沙演变成一个复杂的人工-天然复合的荒漠生态系统.土壤性质的改善和成土作用的加剧,不但使得流沙逐渐向地带性土壤演变,而且促使生物地球化学循环逐渐集中在浅表层[35].此外,随着土壤肥力的提升,植被也逐渐向天然植物群落演变,植物多样性增加的同时,鸟类、动物及昆虫也明显增多[64].开展人工固沙植被生态系统C∶N∶P化学计量的研究,是理解固沙植被系统C、N、P循环过程及植被、微生物和土壤三者之间互馈关系的核心和关键,也是进行科学防沙治沙和管理固沙植被的指导依据.然而,针对沙区人工植被建立后土壤微生物功能群及其化学计量学的研究目前相对较少,需加强人工固沙植被生态系统演替进程中C∶N∶P化学计量的研究. ...
C∶N∶P stoichiometry in soil:is there a "Redfield ratio" for the microbial biomass?
1
2007
... 研究发现,全球森林生态系统植物叶片C∶N∶P为1 212∶28∶1,而当植物叶片凋亡后,养分的再吸收过程使得枯叶的C∶N∶P变为3 007∶45∶1[60];植物凋亡后根的C∶N∶P平均值为4 184∶43∶1[61];土壤C∶N∶P最低为287∶17∶1[62];土壤微生物量C∶N∶P平均值为42∶6∶1,并随土壤深度增加显著减小[26];Sinsabaugh等[57]发现土壤酶中C∶N∶P接近于1∶1∶1.相比之下,从植物叶片、凋落物到土壤再到土壤微生物及酶的计量特征C、N元素整体处于减少趋势.Xu等[37]研究发现,在区域尺度上,随着生物区系不同,C∶N∶P存在很大变异性,如天然湿地中的C∶N∶P为131∶14∶1,而沙漠中的C∶N∶P则为31∶4∶1.面对底物和微生物化学计量间的巨大差异,微生物主要通过调整自身化学计量特征、群落结构、胞外酶比例和元素利用效率等机制来响应和调节.目前为止,关于植物-土壤-微生物-酶C、N、P关系的研究多聚焦于探讨土壤微生物与土壤、植被间的变化及其影响因素.研究表明,随着纬度、年均温、年降水量的变化,微生物量N∶P与植物组织的N∶P呈现出一致的变化趋势,即低纬度地区更受P的限制[63].这表明在高纬度地区微生物和植物一样由于生长季短而需要更高的生长速率,进而需要高的N、P含量和低的N∶P维持[6].Li等[64]分析了中国亚热带地区土壤数据后认为,微生物量C∶N和C∶P会随土壤C∶N和C∶P的增加而增加;周正虎等[38]却发现中国森林土壤微生物量C∶N随着土壤C∶N的增加而减小,与Cleveland等[65]对全球森林的数据分析结果一致.近期,吕星宇等[35]在腾格里沙漠开展的人工固沙植被研究表明,固沙植被演替过程中,微生物功能群数量C∶N∶P的变化主要受到草本、灌木和土壤C∶N∶P变化的影响.受灌木C∶N∶P增加及草本C∶N∶P减小的影响,微生物功能群数量C∶N∶P随之增加;对土壤酶C∶N∶P影响最大的因子分别是微生物功能群数量C∶N∶P和凋落物C∶N∶P,土壤酶C∶N∶P随着微生物功能群数量C∶N∶P增加而增加. ...
我国沙区人工植被系统生态-水文过程和互馈机理研究评述
3
2013
... 植被重建是干旱半干旱荒漠区防止土地沙化、减少风沙危害、促进生态恢复的重要途径[63].中国利用植物固沙已有60多年的历史,国家先后在风沙危害区启动了“三北”防护林建设、退耕还林还草和京津风沙源治理等以人工植被建设作为主要修复措施的一批重大生态建设工程.60余年来,中国沙区植被建设取得了举世瞩目的成就,有效遏制了沙漠化的发展,促进了区域生境恢复[66].人工固沙植被是荒漠生态系统重要的组成部分,是土地荒漠化防治和水土流失治理的主要调控者[66].这些先后建立的人工植被为我们进行荒漠生态系统恢复过程中化学计量学研究奠定了基础.以沙坡头为例,人工固沙植被建立后,原有的流沙演变成一个复杂的人工-天然复合的荒漠生态系统.土壤性质的改善和成土作用的加剧,不但使得流沙逐渐向地带性土壤演变,而且促使生物地球化学循环逐渐集中在浅表层[35].此外,随着土壤肥力的提升,植被也逐渐向天然植物群落演变,植物多样性增加的同时,鸟类、动物及昆虫也明显增多[64].开展人工固沙植被生态系统C∶N∶P化学计量的研究,是理解固沙植被系统C、N、P循环过程及植被、微生物和土壤三者之间互馈关系的核心和关键,也是进行科学防沙治沙和管理固沙植被的指导依据.然而,针对沙区人工植被建立后土壤微生物功能群及其化学计量学的研究目前相对较少,需加强人工固沙植被生态系统演替进程中C∶N∶P化学计量的研究. ...
... [66].这些先后建立的人工植被为我们进行荒漠生态系统恢复过程中化学计量学研究奠定了基础.以沙坡头为例,人工固沙植被建立后,原有的流沙演变成一个复杂的人工-天然复合的荒漠生态系统.土壤性质的改善和成土作用的加剧,不但使得流沙逐渐向地带性土壤演变,而且促使生物地球化学循环逐渐集中在浅表层[35].此外,随着土壤肥力的提升,植被也逐渐向天然植物群落演变,植物多样性增加的同时,鸟类、动物及昆虫也明显增多[64].开展人工固沙植被生态系统C∶N∶P化学计量的研究,是理解固沙植被系统C、N、P循环过程及植被、微生物和土壤三者之间互馈关系的核心和关键,也是进行科学防沙治沙和管理固沙植被的指导依据.然而,针对沙区人工植被建立后土壤微生物功能群及其化学计量学的研究目前相对较少,需加强人工固沙植被生态系统演替进程中C∶N∶P化学计量的研究. ...
... BSC是由隐花植物(如蓝藻、地衣、苔藓)和土壤微生物及其他相关生物体通过分泌物、菌丝体和假根等与土壤表层颗粒胶结而成的复合体,是荒漠地表景观的重要组成[66].大多数荒漠地区受到非生物因子(如水分匮乏)的调控和胁迫,使得地表维管束植物群落呈斑块状分布,而这种分布格局为BSC在荒漠地区的繁殖和覆盖提供了合适的空间和适宜的生态位,使BSC广泛分布于荒漠地区[63].BSC对于荒漠景观过程,土壤生态、水文、生物过程,地球化学循环及生态修复过程起着重要作用[67].由于目前还没有针对BSC或隐花植物C∶N∶P计量的研究,缺乏BSC对土壤C、N、P响应的生态化学计量认识.因此,开展BSC或隐花植物C∶N∶P化学计量特征的研究,有助于揭示荒漠生态系统植被演替过程中C、N、P的变化规律. ...
沙坡头生态水文学研究进展及水量平衡自动模拟监测系统
1
2018
... BSC是由隐花植物(如蓝藻、地衣、苔藓)和土壤微生物及其他相关生物体通过分泌物、菌丝体和假根等与土壤表层颗粒胶结而成的复合体,是荒漠地表景观的重要组成[66].大多数荒漠地区受到非生物因子(如水分匮乏)的调控和胁迫,使得地表维管束植物群落呈斑块状分布,而这种分布格局为BSC在荒漠地区的繁殖和覆盖提供了合适的空间和适宜的生态位,使BSC广泛分布于荒漠地区[63].BSC对于荒漠景观过程,土壤生态、水文、生物过程,地球化学循环及生态修复过程起着重要作用[67].由于目前还没有针对BSC或隐花植物C∶N∶P计量的研究,缺乏BSC对土壤C、N、P响应的生态化学计量认识.因此,开展BSC或隐花植物C∶N∶P化学计量特征的研究,有助于揭示荒漠生态系统植被演替过程中C、N、P的变化规律. ...
Dispersing misconceptions and identifying opportunities for the use of 'omics' in soil microbial ecology
1
2015
... 此外,建立微生物群落结构与功能具体的联系是土壤微生物学面临的一大挑战.许多微生物过程并不是单一途径的代谢产物,而是可以被分类的综合代谢途径的产物.如土壤中几乎所有的异养细菌活性类群都可能促进有机碳分解代谢或氮矿化的速率[36].甚至就植物纤维素分解代谢而言,可能需要多个不同类型的代谢过程共同完成[68].所以将微生物以功能群的形式进行划分,研究不同微生物功能群C∶N∶P化学计量也将是未来研究的重点. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}