中亚干旱荒漠区植被碳储量估算
2
2013
... 荒漠生态系统是陆地生态系统的重要子系统,虽然生物量及碳储量较森林和草原低,但面积大、分布广,固碳潜力不容忽视[1].古尔班通古特沙漠作为中国第二大固定和半固定沙漠,具有典型的大陆性荒漠气候,降雨季节分布比较均匀,冬季还有大量积雪[2-5].在早春时期,短命植物与类短命植物以及少量一年生和多年生草本植物利用较为丰富的水分快速生长,形成了类似于荒漠草原的独特景观[1,5-6].这些不同功能型的草本植物贡献了绝大部分的生产力和多样性,在荒漠生态系统地表稳定的维持、荒漠植物群落物种构成、能量流动、养分循环等方面起着重要作用[3,7-8].以短命植物为例,生产力占草本群落产量的60%以上,在它完成生活史后,还会在6个月内把凋落物中35%的氮和60%的磷再释放到土壤中,为下阶段生长的灌木和草本植物提供养分,被认为是沙漠环境的养分库[9-10].除此之外,古尔班通古特沙漠中还包括许多单种的科和属,虽然特有成分较少,但优势现象明显,例如牻牛儿苗科、车前科和莎草科等,它们在沙漠植被中具有重要作用[11]. ...
... [1,5-6].这些不同功能型的草本植物贡献了绝大部分的生产力和多样性,在荒漠生态系统地表稳定的维持、荒漠植物群落物种构成、能量流动、养分循环等方面起着重要作用[3,7-8].以短命植物为例,生产力占草本群落产量的60%以上,在它完成生活史后,还会在6个月内把凋落物中35%的氮和60%的磷再释放到土壤中,为下阶段生长的灌木和草本植物提供养分,被认为是沙漠环境的养分库[9-10].除此之外,古尔班通古特沙漠中还包括许多单种的科和属,虽然特有成分较少,但优势现象明显,例如牻牛儿苗科、车前科和莎草科等,它们在沙漠植被中具有重要作用[11]. ...
Distribution of ephemeral plants and their significance in dune stabilization in Gurbantunggut Desert
2
2003
... 荒漠生态系统是陆地生态系统的重要子系统,虽然生物量及碳储量较森林和草原低,但面积大、分布广,固碳潜力不容忽视[1].古尔班通古特沙漠作为中国第二大固定和半固定沙漠,具有典型的大陆性荒漠气候,降雨季节分布比较均匀,冬季还有大量积雪[2-5].在早春时期,短命植物与类短命植物以及少量一年生和多年生草本植物利用较为丰富的水分快速生长,形成了类似于荒漠草原的独特景观[1,5-6].这些不同功能型的草本植物贡献了绝大部分的生产力和多样性,在荒漠生态系统地表稳定的维持、荒漠植物群落物种构成、能量流动、养分循环等方面起着重要作用[3,7-8].以短命植物为例,生产力占草本群落产量的60%以上,在它完成生活史后,还会在6个月内把凋落物中35%的氮和60%的磷再释放到土壤中,为下阶段生长的灌木和草本植物提供养分,被认为是沙漠环境的养分库[9-10].除此之外,古尔班通古特沙漠中还包括许多单种的科和属,虽然特有成分较少,但优势现象明显,例如牻牛儿苗科、车前科和莎草科等,它们在沙漠植被中具有重要作用[11]. ...
... 研究区位于新疆北部的古尔班通古特沙漠(44°11′—46°50′N、84°31′—91°20′E,面积4.88万km2).该沙漠年均蒸发量>2 000 mm,而年均降水量普遍不超过150 mm,沙漠腹地仅有70—100 mm,集中于冬春季;年均气温6—10 ℃,极端最高气温为40 ℃以上,≥10 ℃的年积温3 000—3 500 ℃,空气相对湿度平均50%—60%,5—8月通常小于45%[2].梭梭(Haloxylon ammodendron)和白梭梭(Haloxylon persicum)是准噶尔荒漠植物群落的主要灌木建群种.该地区具有丰富的短命和类短命植物,是早春荒漠生态系统的重要组成部分[5-6].此外,该沙漠还广泛分布着生物土壤结皮,包括藻结皮、地衣结皮和苔藓结皮3种主要类型,也是维持荒漠地表稳定的重要生物因子[29]. ...
短命植物分布与沙垄表层土壤水分的关系:以古尔班通古特沙漠为例
1
2004
... 荒漠生态系统是陆地生态系统的重要子系统,虽然生物量及碳储量较森林和草原低,但面积大、分布广,固碳潜力不容忽视[1].古尔班通古特沙漠作为中国第二大固定和半固定沙漠,具有典型的大陆性荒漠气候,降雨季节分布比较均匀,冬季还有大量积雪[2-5].在早春时期,短命植物与类短命植物以及少量一年生和多年生草本植物利用较为丰富的水分快速生长,形成了类似于荒漠草原的独特景观[1,5-6].这些不同功能型的草本植物贡献了绝大部分的生产力和多样性,在荒漠生态系统地表稳定的维持、荒漠植物群落物种构成、能量流动、养分循环等方面起着重要作用[3,7-8].以短命植物为例,生产力占草本群落产量的60%以上,在它完成生活史后,还会在6个月内把凋落物中35%的氮和60%的磷再释放到土壤中,为下阶段生长的灌木和草本植物提供养分,被认为是沙漠环境的养分库[9-10].除此之外,古尔班通古特沙漠中还包括许多单种的科和属,虽然特有成分较少,但优势现象明显,例如牻牛儿苗科、车前科和莎草科等,它们在沙漠植被中具有重要作用[11]. ...
新疆准噶尔荒漠短命植物群落特征及其水热适应性
1
2010
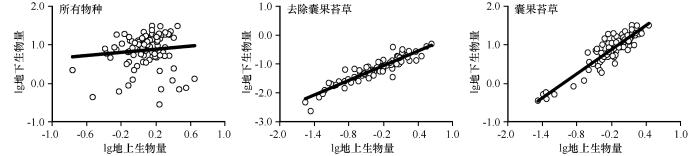
... 目前关于草本植物群落/层片地上和地下生物量分配机制在异速生长假说和最优分配假说之间仍有争议.考虑到两种理论在研究生物量分配中的局限性,许多研究者将异速生长法和最优分配假说结合来进行了应用[42-44].在群落水平上,有诸多研究均证明了草本植物地上生物量与地下生物量之间的等速生长关系[19,28,45-46].从本研究结果来看,地上与地下生物量之间没有明显的相关生长关系,与马安娜等[47]对温带草原、山地草甸、温性草甸草原地上与地下生物量之间关系的研究结果类似.当去除类短命植物囊果苔草后,虽然草本层片地上生物量大于地下部分,但地上与地下生物量分配却符合等速生长理论,该结果与其他相关研究结果一致[41,46].有研究表明,不同草本植物群落生物量具有不同的分配规律,其原因可能是群落物种组成和生长类型的影响[48].群落内不同物种间因对环境的适应性不同或者是系统分类地位和遗传发育的差异[4],导致不同生活型的草本植物对群落生产力的贡献率不同,其中优势物种的特性在很大程度上决定了群落的生物量.因此,在研究区内,物种间的遗传差异使得各物种采取不同的生存策略,不同类型的植物为了适应环境具有不同的生物量分配策略,在资源有限的情况下,选择将更多生物量分配给遭受胁迫的器官,符合最优分配假说[26].但本研究在去除囊果苔草对整个层片影响的情况下,层片地上与地下生物量分配关系符合异速生长理论,可见最优分配假说与异速分配假说两者并不矛盾,在一定条件下可以结合起来[42],共同解释该草本植物层片生物量的不同分配模式. ...
新疆北部早春短命植物区系纲要
3
1994
... 荒漠生态系统是陆地生态系统的重要子系统,虽然生物量及碳储量较森林和草原低,但面积大、分布广,固碳潜力不容忽视[1].古尔班通古特沙漠作为中国第二大固定和半固定沙漠,具有典型的大陆性荒漠气候,降雨季节分布比较均匀,冬季还有大量积雪[2-5].在早春时期,短命植物与类短命植物以及少量一年生和多年生草本植物利用较为丰富的水分快速生长,形成了类似于荒漠草原的独特景观[1,5-6].这些不同功能型的草本植物贡献了绝大部分的生产力和多样性,在荒漠生态系统地表稳定的维持、荒漠植物群落物种构成、能量流动、养分循环等方面起着重要作用[3,7-8].以短命植物为例,生产力占草本群落产量的60%以上,在它完成生活史后,还会在6个月内把凋落物中35%的氮和60%的磷再释放到土壤中,为下阶段生长的灌木和草本植物提供养分,被认为是沙漠环境的养分库[9-10].除此之外,古尔班通古特沙漠中还包括许多单种的科和属,虽然特有成分较少,但优势现象明显,例如牻牛儿苗科、车前科和莎草科等,它们在沙漠植被中具有重要作用[11]. ...
... ,5-6].这些不同功能型的草本植物贡献了绝大部分的生产力和多样性,在荒漠生态系统地表稳定的维持、荒漠植物群落物种构成、能量流动、养分循环等方面起着重要作用[3,7-8].以短命植物为例,生产力占草本群落产量的60%以上,在它完成生活史后,还会在6个月内把凋落物中35%的氮和60%的磷再释放到土壤中,为下阶段生长的灌木和草本植物提供养分,被认为是沙漠环境的养分库[9-10].除此之外,古尔班通古特沙漠中还包括许多单种的科和属,虽然特有成分较少,但优势现象明显,例如牻牛儿苗科、车前科和莎草科等,它们在沙漠植被中具有重要作用[11]. ...
... 研究区位于新疆北部的古尔班通古特沙漠(44°11′—46°50′N、84°31′—91°20′E,面积4.88万km2).该沙漠年均蒸发量>2 000 mm,而年均降水量普遍不超过150 mm,沙漠腹地仅有70—100 mm,集中于冬春季;年均气温6—10 ℃,极端最高气温为40 ℃以上,≥10 ℃的年积温3 000—3 500 ℃,空气相对湿度平均50%—60%,5—8月通常小于45%[2].梭梭(Haloxylon ammodendron)和白梭梭(Haloxylon persicum)是准噶尔荒漠植物群落的主要灌木建群种.该地区具有丰富的短命和类短命植物,是早春荒漠生态系统的重要组成部分[5-6].此外,该沙漠还广泛分布着生物土壤结皮,包括藻结皮、地衣结皮和苔藓结皮3种主要类型,也是维持荒漠地表稳定的重要生物因子[29]. ...
论古尔班通古特沙漠植物多样性的一般特点
2
2002
... 荒漠生态系统是陆地生态系统的重要子系统,虽然生物量及碳储量较森林和草原低,但面积大、分布广,固碳潜力不容忽视[1].古尔班通古特沙漠作为中国第二大固定和半固定沙漠,具有典型的大陆性荒漠气候,降雨季节分布比较均匀,冬季还有大量积雪[2-5].在早春时期,短命植物与类短命植物以及少量一年生和多年生草本植物利用较为丰富的水分快速生长,形成了类似于荒漠草原的独特景观[1,5-6].这些不同功能型的草本植物贡献了绝大部分的生产力和多样性,在荒漠生态系统地表稳定的维持、荒漠植物群落物种构成、能量流动、养分循环等方面起着重要作用[3,7-8].以短命植物为例,生产力占草本群落产量的60%以上,在它完成生活史后,还会在6个月内把凋落物中35%的氮和60%的磷再释放到土壤中,为下阶段生长的灌木和草本植物提供养分,被认为是沙漠环境的养分库[9-10].除此之外,古尔班通古特沙漠中还包括许多单种的科和属,虽然特有成分较少,但优势现象明显,例如牻牛儿苗科、车前科和莎草科等,它们在沙漠植被中具有重要作用[11]. ...
... 研究区位于新疆北部的古尔班通古特沙漠(44°11′—46°50′N、84°31′—91°20′E,面积4.88万km2).该沙漠年均蒸发量>2 000 mm,而年均降水量普遍不超过150 mm,沙漠腹地仅有70—100 mm,集中于冬春季;年均气温6—10 ℃,极端最高气温为40 ℃以上,≥10 ℃的年积温3 000—3 500 ℃,空气相对湿度平均50%—60%,5—8月通常小于45%[2].梭梭(Haloxylon ammodendron)和白梭梭(Haloxylon persicum)是准噶尔荒漠植物群落的主要灌木建群种.该地区具有丰富的短命和类短命植物,是早春荒漠生态系统的重要组成部分[5-6].此外,该沙漠还广泛分布着生物土壤结皮,包括藻结皮、地衣结皮和苔藓结皮3种主要类型,也是维持荒漠地表稳定的重要生物因子[29]. ...
The ecological significance of the Herbaceous Layer in temperate forest ecosystems
1
2007
... 荒漠生态系统是陆地生态系统的重要子系统,虽然生物量及碳储量较森林和草原低,但面积大、分布广,固碳潜力不容忽视[1].古尔班通古特沙漠作为中国第二大固定和半固定沙漠,具有典型的大陆性荒漠气候,降雨季节分布比较均匀,冬季还有大量积雪[2-5].在早春时期,短命植物与类短命植物以及少量一年生和多年生草本植物利用较为丰富的水分快速生长,形成了类似于荒漠草原的独特景观[1,5-6].这些不同功能型的草本植物贡献了绝大部分的生产力和多样性,在荒漠生态系统地表稳定的维持、荒漠植物群落物种构成、能量流动、养分循环等方面起着重要作用[3,7-8].以短命植物为例,生产力占草本群落产量的60%以上,在它完成生活史后,还会在6个月内把凋落物中35%的氮和60%的磷再释放到土壤中,为下阶段生长的灌木和草本植物提供养分,被认为是沙漠环境的养分库[9-10].除此之外,古尔班通古特沙漠中还包括许多单种的科和属,虽然特有成分较少,但优势现象明显,例如牻牛儿苗科、车前科和莎草科等,它们在沙漠植被中具有重要作用[11]. ...
3种荒漠植物群落物种组成与丰富度的季节变化及地上生物量特征
1
2011
... 荒漠生态系统是陆地生态系统的重要子系统,虽然生物量及碳储量较森林和草原低,但面积大、分布广,固碳潜力不容忽视[1].古尔班通古特沙漠作为中国第二大固定和半固定沙漠,具有典型的大陆性荒漠气候,降雨季节分布比较均匀,冬季还有大量积雪[2-5].在早春时期,短命植物与类短命植物以及少量一年生和多年生草本植物利用较为丰富的水分快速生长,形成了类似于荒漠草原的独特景观[1,5-6].这些不同功能型的草本植物贡献了绝大部分的生产力和多样性,在荒漠生态系统地表稳定的维持、荒漠植物群落物种构成、能量流动、养分循环等方面起着重要作用[3,7-8].以短命植物为例,生产力占草本群落产量的60%以上,在它完成生活史后,还会在6个月内把凋落物中35%的氮和60%的磷再释放到土壤中,为下阶段生长的灌木和草本植物提供养分,被认为是沙漠环境的养分库[9-10].除此之外,古尔班通古特沙漠中还包括许多单种的科和属,虽然特有成分较少,但优势现象明显,例如牻牛儿苗科、车前科和莎草科等,它们在沙漠植被中具有重要作用[11]. ...
新疆莫索湾地区短命植物的初步研究
1
1985
... 荒漠生态系统是陆地生态系统的重要子系统,虽然生物量及碳储量较森林和草原低,但面积大、分布广,固碳潜力不容忽视[1].古尔班通古特沙漠作为中国第二大固定和半固定沙漠,具有典型的大陆性荒漠气候,降雨季节分布比较均匀,冬季还有大量积雪[2-5].在早春时期,短命植物与类短命植物以及少量一年生和多年生草本植物利用较为丰富的水分快速生长,形成了类似于荒漠草原的独特景观[1,5-6].这些不同功能型的草本植物贡献了绝大部分的生产力和多样性,在荒漠生态系统地表稳定的维持、荒漠植物群落物种构成、能量流动、养分循环等方面起着重要作用[3,7-8].以短命植物为例,生产力占草本群落产量的60%以上,在它完成生活史后,还会在6个月内把凋落物中35%的氮和60%的磷再释放到土壤中,为下阶段生长的灌木和草本植物提供养分,被认为是沙漠环境的养分库[9-10].除此之外,古尔班通古特沙漠中还包括许多单种的科和属,虽然特有成分较少,但优势现象明显,例如牻牛儿苗科、车前科和莎草科等,它们在沙漠植被中具有重要作用[11]. ...
The role of spring ephemerals and soil microbes in soil nutrient retention in a temperate desert
1
2016
... 荒漠生态系统是陆地生态系统的重要子系统,虽然生物量及碳储量较森林和草原低,但面积大、分布广,固碳潜力不容忽视[1].古尔班通古特沙漠作为中国第二大固定和半固定沙漠,具有典型的大陆性荒漠气候,降雨季节分布比较均匀,冬季还有大量积雪[2-5].在早春时期,短命植物与类短命植物以及少量一年生和多年生草本植物利用较为丰富的水分快速生长,形成了类似于荒漠草原的独特景观[1,5-6].这些不同功能型的草本植物贡献了绝大部分的生产力和多样性,在荒漠生态系统地表稳定的维持、荒漠植物群落物种构成、能量流动、养分循环等方面起着重要作用[3,7-8].以短命植物为例,生产力占草本群落产量的60%以上,在它完成生活史后,还会在6个月内把凋落物中35%的氮和60%的磷再释放到土壤中,为下阶段生长的灌木和草本植物提供养分,被认为是沙漠环境的养分库[9-10].除此之外,古尔班通古特沙漠中还包括许多单种的科和属,虽然特有成分较少,但优势现象明显,例如牻牛儿苗科、车前科和莎草科等,它们在沙漠植被中具有重要作用[11]. ...
古尔班通古特沙漠南部植物多样性及群落分类
1
2012
... 荒漠生态系统是陆地生态系统的重要子系统,虽然生物量及碳储量较森林和草原低,但面积大、分布广,固碳潜力不容忽视[1].古尔班通古特沙漠作为中国第二大固定和半固定沙漠,具有典型的大陆性荒漠气候,降雨季节分布比较均匀,冬季还有大量积雪[2-5].在早春时期,短命植物与类短命植物以及少量一年生和多年生草本植物利用较为丰富的水分快速生长,形成了类似于荒漠草原的独特景观[1,5-6].这些不同功能型的草本植物贡献了绝大部分的生产力和多样性,在荒漠生态系统地表稳定的维持、荒漠植物群落物种构成、能量流动、养分循环等方面起着重要作用[3,7-8].以短命植物为例,生产力占草本群落产量的60%以上,在它完成生活史后,还会在6个月内把凋落物中35%的氮和60%的磷再释放到土壤中,为下阶段生长的灌木和草本植物提供养分,被认为是沙漠环境的养分库[9-10].除此之外,古尔班通古特沙漠中还包括许多单种的科和属,虽然特有成分较少,但优势现象明显,例如牻牛儿苗科、车前科和莎草科等,它们在沙漠植被中具有重要作用[11]. ...
科尔沁沙地70种草本植物个体和构件生物量比较研究
1
2009
... 目前关于荒漠区植被地上和地下生物量分配特征的研究有很多,但是由于环境条件、时间、物种等因素的不同,分别得出了不同的地上和地下生物量分配比率.在个体水平上,李雪华等[12]通过对沙漠多种草本植物生物量的研究发现,生活史短的植物投入到地上构件的生物量较多,而多年生植物则会向地下部分投入较多的能量.陶冶等[13]分别对不同的类短命植物生物量分配特征进行了研究,发现不同科、不同属及同属不同种之间的生物量大小及器官分配比例均没有一致规律.对古尔班通古特沙漠不同地区的3种不同生活型草本植物的研究表明,类短命植物和一年生草本植物地上与地下生物量间呈现出异速生长格局,短命植物地上和地下的生物量分配为等速生长关系[14-16].综上可知,不同生命形式的草本植物在生长发育过程中,可能会采取不同的策略来调节地上和地下生物量的分配以适应变化的环境条件[17]. ...
准噶尔荒漠6种类短命植物生物量分配与异速生长关系
3
2014
... 目前关于荒漠区植被地上和地下生物量分配特征的研究有很多,但是由于环境条件、时间、物种等因素的不同,分别得出了不同的地上和地下生物量分配比率.在个体水平上,李雪华等[12]通过对沙漠多种草本植物生物量的研究发现,生活史短的植物投入到地上构件的生物量较多,而多年生植物则会向地下部分投入较多的能量.陶冶等[13]分别对不同的类短命植物生物量分配特征进行了研究,发现不同科、不同属及同属不同种之间的生物量大小及器官分配比例均没有一致规律.对古尔班通古特沙漠不同地区的3种不同生活型草本植物的研究表明,类短命植物和一年生草本植物地上与地下生物量间呈现出异速生长格局,短命植物地上和地下的生物量分配为等速生长关系[14-16].综上可知,不同生命形式的草本植物在生长发育过程中,可能会采取不同的策略来调节地上和地下生物量的分配以适应变化的环境条件[17]. ...
... 生物量是研究生态系统碳循环的基础,可以反映生态系统生产力的大小,不仅受到物种多样性、密度和均匀度的影响,还与物种本身的生物学特性和外部环境资源有关[23].生物量的分配是指植物将光合产物用于地上和地下不同器官中的比例,是植物对获取资源和利用资源的权衡,可以反映植物对不同的生长环境和选择压力的适应[24-26],会受到物种特征、个体发育和植物生存环境的强烈影响.关于生物量分配有两种流行的假设:最优分配和异速分配.最优分配理论指出植物为适应环境变化,会将生物量分配给获得最有限资源的器官,即通过调整体内生物量的分配以最大限度地获取光、营养和水等有限的资源[27].异速生长理论表明,在自然条件下种内和种间成熟个体的生物量分配速率相同[20,27-28],也有研究得出生物量呈异速分配模式[13].有研究者提出上述两种假说是相辅相成的,在解释植物生物量分配方面可能是互补的而非独立的[26],但这还需要大量研究加以验证. ...
... 采用全株挖掘法获取样方内全部植物地上和地下生物量.植物根系挖掘的深度因物种而异[13,15-16],根据以往经验及文献报道,样方内短命植物、一年生植物及类短命植物囊果苔草根系一般分布于0.3 m以内,但为保证根系能全部获取,我们将取样深度定为0.5 m,并根据野外实际情况灵活变动.在挖出的土块中,直接收获粗壮主根,断裂的细根利用1—2 mm土筛筛选后挑出.仔细除去附着在目标植物根系上的土壤及其他植物根系,同时去除处于半分解状态的死根,并对植株进行分类标记保存.回到实验室,将每株植物冲洗干净,将地上和地下部分剪开,同样方内的同种植物同器官样品包装在一起,所有样品置于70 ℃烘箱内烘干至恒重. ...
Ecological biomass allocation strategies in plant species with different life forms in a cold desert,China
2
2019
... 目前关于荒漠区植被地上和地下生物量分配特征的研究有很多,但是由于环境条件、时间、物种等因素的不同,分别得出了不同的地上和地下生物量分配比率.在个体水平上,李雪华等[12]通过对沙漠多种草本植物生物量的研究发现,生活史短的植物投入到地上构件的生物量较多,而多年生植物则会向地下部分投入较多的能量.陶冶等[13]分别对不同的类短命植物生物量分配特征进行了研究,发现不同科、不同属及同属不同种之间的生物量大小及器官分配比例均没有一致规律.对古尔班通古特沙漠不同地区的3种不同生活型草本植物的研究表明,类短命植物和一年生草本植物地上与地下生物量间呈现出异速生长格局,短命植物地上和地下的生物量分配为等速生长关系[14-16].综上可知,不同生命形式的草本植物在生长发育过程中,可能会采取不同的策略来调节地上和地下生物量的分配以适应变化的环境条件[17]. ...
... 物种多样性是指生态系统中物种的丰富度,是评价一个地区生物资源丰富程度的重要指标,对维持生态系统的稳定性具有重要意义[14].物种多样性和生产力因受尺度、干扰、群落历史等因素的影响而呈现出正相关、负相关、U型、单峰型和不相关等多种表达形式[49].尽管目前已有许多研究表明干旱、半干旱区域草本植物群落的生物量与物种丰富度之间为单峰关系[50-52].但因沙漠生态系统生产力通常受到水分、养分有效性的限制,并且在空间和时间上具有高度可变性,地理位置、生物地理历史以及极端温度等因素也导致沙漠中的物种多样性差异很大,所以干旱生态系统中生产力和物种多样性之间还没有明确的关系[51].有研究认为有利的气候条件如高的降雨量,会增加物种丰富度和地下生物量,增大群落对极端气候的抵抗力[53].还有研究得出干旱胁迫虽然使得生物量显著减少,但并未改变群落物种组成和物种多样性,因为沙漠植物可塑性较高,会通过增加优势物种的生殖器官的生物量,来维持群落的物种丰富度和稳定性[54]. ...
古尔班通古特沙漠南缘沙垄表面草本植物分布特征及生物量分配研究
1
2015
... 采用全株挖掘法获取样方内全部植物地上和地下生物量.植物根系挖掘的深度因物种而异[13,15-16],根据以往经验及文献报道,样方内短命植物、一年生植物及类短命植物囊果苔草根系一般分布于0.3 m以内,但为保证根系能全部获取,我们将取样深度定为0.5 m,并根据野外实际情况灵活变动.在挖出的土块中,直接收获粗壮主根,断裂的细根利用1—2 mm土筛筛选后挑出.仔细除去附着在目标植物根系上的土壤及其他植物根系,同时去除处于半分解状态的死根,并对植株进行分类标记保存.回到实验室,将每株植物冲洗干净,将地上和地下部分剪开,同样方内的同种植物同器官样品包装在一起,所有样品置于70 ℃烘箱内烘干至恒重. ...
古尔班通古特沙漠6种荒漠草本植物的生物量分配与相关生长关系
3
2016
... 目前关于荒漠区植被地上和地下生物量分配特征的研究有很多,但是由于环境条件、时间、物种等因素的不同,分别得出了不同的地上和地下生物量分配比率.在个体水平上,李雪华等[12]通过对沙漠多种草本植物生物量的研究发现,生活史短的植物投入到地上构件的生物量较多,而多年生植物则会向地下部分投入较多的能量.陶冶等[13]分别对不同的类短命植物生物量分配特征进行了研究,发现不同科、不同属及同属不同种之间的生物量大小及器官分配比例均没有一致规律.对古尔班通古特沙漠不同地区的3种不同生活型草本植物的研究表明,类短命植物和一年生草本植物地上与地下生物量间呈现出异速生长格局,短命植物地上和地下的生物量分配为等速生长关系[14-16].综上可知,不同生命形式的草本植物在生长发育过程中,可能会采取不同的策略来调节地上和地下生物量的分配以适应变化的环境条件[17]. ...
... 采用全株挖掘法获取样方内全部植物地上和地下生物量.植物根系挖掘的深度因物种而异[13,15-16],根据以往经验及文献报道,样方内短命植物、一年生植物及类短命植物囊果苔草根系一般分布于0.3 m以内,但为保证根系能全部获取,我们将取样深度定为0.5 m,并根据野外实际情况灵活变动.在挖出的土块中,直接收获粗壮主根,断裂的细根利用1—2 mm土筛筛选后挑出.仔细除去附着在目标植物根系上的土壤及其他植物根系,同时去除处于半分解状态的死根,并对植株进行分类标记保存.回到实验室,将每株植物冲洗干净,将地上和地下部分剪开,同样方内的同种植物同器官样品包装在一起,所有样品置于70 ℃烘箱内烘干至恒重. ...
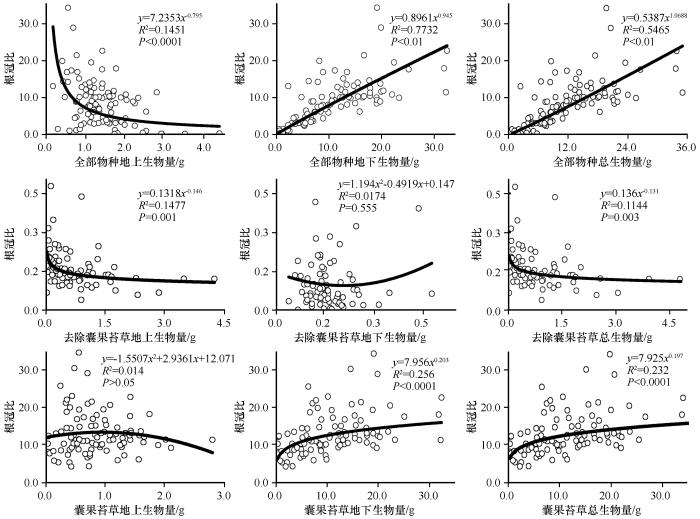
... 生物量的分配是指植物生长发育过程中将光合产物用于地上、地下不同器官中的比例,体现了植物对资源的权衡和环境的适应[39].研究区草本植物层片地上和地下生物量变化范围均小于冀西北和黄土丘陵不同植被带的草本植物群落,但由于类短命植物的存在,使得根冠比的变化范围大于后两者[40-41].对于去除囊果苔草的草本层片来说,根冠比与地下生物量之间没有相关关系,随地上生物量和总生物量的增加而降低但变化范围较窄,表明去除囊果苔草的草本层片对繁殖分配和营养分配的权衡主要发生在地上部分.以层片中短命植物为例,为了在短时间内资源较为丰富的条件下快速完成生活史,选择将较多的能量投入到地上部分,以保证植物的有性繁殖和种群扩散[16].其中层片水平的地下生物量和总生物量与根冠比之间的关系与层片中囊果苔草的变化趋势相似,说明草本群落/层片水平的地下生物量受囊果苔草地下生物量的影响,使得整个草本层片的生物量分配倾向于根系部分.草本植物层片R/S随地上生物量的增大显著降低,随地下生物量和总生物量的增加显著增大,而且变化范围都较大,证明了群落/层片根冠比具有生物量大小依赖性. ...
古尔班通古特沙漠南缘3种生活型草本植物生物量分配及相关生长关系
1
2017
... 目前关于荒漠区植被地上和地下生物量分配特征的研究有很多,但是由于环境条件、时间、物种等因素的不同,分别得出了不同的地上和地下生物量分配比率.在个体水平上,李雪华等[12]通过对沙漠多种草本植物生物量的研究发现,生活史短的植物投入到地上构件的生物量较多,而多年生植物则会向地下部分投入较多的能量.陶冶等[13]分别对不同的类短命植物生物量分配特征进行了研究,发现不同科、不同属及同属不同种之间的生物量大小及器官分配比例均没有一致规律.对古尔班通古特沙漠不同地区的3种不同生活型草本植物的研究表明,类短命植物和一年生草本植物地上与地下生物量间呈现出异速生长格局,短命植物地上和地下的生物量分配为等速生长关系[14-16].综上可知,不同生命形式的草本植物在生长发育过程中,可能会采取不同的策略来调节地上和地下生物量的分配以适应变化的环境条件[17]. ...
Patterns of above- and belowground biomass allocation in China's grasslands:evidence from individual-level observations
1
2010
... 同样,在群落水平上,地上、地下生物量相关生长关系的研究也有很大发展.有学者发现中国草地生物量分配不符合等速生长假说[18],而Yang等[19]通过对青藏高原多个地点的调查研究发现,地上与地下生物量符合等速分配格局.程栋梁[20]研究了中国西北地区5个不同自然湿度梯度的植物生物量分配关系,发现干旱虽然会影响异速生长常数,但地上和地下生物量之间仍存在相关生长关系.王敏等[21]对黑河中游的地上和地下生物量进行了调查,发现无论是个体水平还是群落水平,地上与地下生物量均为等速生长关系.陶冶[22]对准噶尔荒漠典型植物群落生物的分配关系进行了研究,结果表明大部分草本植物的地上与地下生物量之间为等速生长关系,并且地上和地下生物量间的相关生长关系随生长期的变化而发生变化,在成熟期趋于等速生长.不同物种和群落间的地上与地下生物量生长关系存在差异,虽然诸多研究证明了草本植物群落地上与地下生物量之间具有等速生长关系,但是这种等速生长关系是否符合所有群落类型,特别是干旱荒漠区的草本植物,还没有一致结论. ...
Above- and belowground biomass allocation in Tibetan grasslands
2
2009
... 同样,在群落水平上,地上、地下生物量相关生长关系的研究也有很大发展.有学者发现中国草地生物量分配不符合等速生长假说[18],而Yang等[19]通过对青藏高原多个地点的调查研究发现,地上与地下生物量符合等速分配格局.程栋梁[20]研究了中国西北地区5个不同自然湿度梯度的植物生物量分配关系,发现干旱虽然会影响异速生长常数,但地上和地下生物量之间仍存在相关生长关系.王敏等[21]对黑河中游的地上和地下生物量进行了调查,发现无论是个体水平还是群落水平,地上与地下生物量均为等速生长关系.陶冶[22]对准噶尔荒漠典型植物群落生物的分配关系进行了研究,结果表明大部分草本植物的地上与地下生物量之间为等速生长关系,并且地上和地下生物量间的相关生长关系随生长期的变化而发生变化,在成熟期趋于等速生长.不同物种和群落间的地上与地下生物量生长关系存在差异,虽然诸多研究证明了草本植物群落地上与地下生物量之间具有等速生长关系,但是这种等速生长关系是否符合所有群落类型,特别是干旱荒漠区的草本植物,还没有一致结论. ...
... 目前关于草本植物群落/层片地上和地下生物量分配机制在异速生长假说和最优分配假说之间仍有争议.考虑到两种理论在研究生物量分配中的局限性,许多研究者将异速生长法和最优分配假说结合来进行了应用[42-44].在群落水平上,有诸多研究均证明了草本植物地上生物量与地下生物量之间的等速生长关系[19,28,45-46].从本研究结果来看,地上与地下生物量之间没有明显的相关生长关系,与马安娜等[47]对温带草原、山地草甸、温性草甸草原地上与地下生物量之间关系的研究结果类似.当去除类短命植物囊果苔草后,虽然草本层片地上生物量大于地下部分,但地上与地下生物量分配却符合等速生长理论,该结果与其他相关研究结果一致[41,46].有研究表明,不同草本植物群落生物量具有不同的分配规律,其原因可能是群落物种组成和生长类型的影响[48].群落内不同物种间因对环境的适应性不同或者是系统分类地位和遗传发育的差异[4],导致不同生活型的草本植物对群落生产力的贡献率不同,其中优势物种的特性在很大程度上决定了群落的生物量.因此,在研究区内,物种间的遗传差异使得各物种采取不同的生存策略,不同类型的植物为了适应环境具有不同的生物量分配策略,在资源有限的情况下,选择将更多生物量分配给遭受胁迫的器官,符合最优分配假说[26].但本研究在去除囊果苔草对整个层片影响的情况下,层片地上与地下生物量分配关系符合异速生长理论,可见最优分配假说与异速分配假说两者并不矛盾,在一定条件下可以结合起来[42],共同解释该草本植物层片生物量的不同分配模式. ...
植物生物量分配模式与生长速率的相关规律研究
2
2007
... 同样,在群落水平上,地上、地下生物量相关生长关系的研究也有很大发展.有学者发现中国草地生物量分配不符合等速生长假说[18],而Yang等[19]通过对青藏高原多个地点的调查研究发现,地上与地下生物量符合等速分配格局.程栋梁[20]研究了中国西北地区5个不同自然湿度梯度的植物生物量分配关系,发现干旱虽然会影响异速生长常数,但地上和地下生物量之间仍存在相关生长关系.王敏等[21]对黑河中游的地上和地下生物量进行了调查,发现无论是个体水平还是群落水平,地上与地下生物量均为等速生长关系.陶冶[22]对准噶尔荒漠典型植物群落生物的分配关系进行了研究,结果表明大部分草本植物的地上与地下生物量之间为等速生长关系,并且地上和地下生物量间的相关生长关系随生长期的变化而发生变化,在成熟期趋于等速生长.不同物种和群落间的地上与地下生物量生长关系存在差异,虽然诸多研究证明了草本植物群落地上与地下生物量之间具有等速生长关系,但是这种等速生长关系是否符合所有群落类型,特别是干旱荒漠区的草本植物,还没有一致结论. ...
... 生物量是研究生态系统碳循环的基础,可以反映生态系统生产力的大小,不仅受到物种多样性、密度和均匀度的影响,还与物种本身的生物学特性和外部环境资源有关[23].生物量的分配是指植物将光合产物用于地上和地下不同器官中的比例,是植物对获取资源和利用资源的权衡,可以反映植物对不同的生长环境和选择压力的适应[24-26],会受到物种特征、个体发育和植物生存环境的强烈影响.关于生物量分配有两种流行的假设:最优分配和异速分配.最优分配理论指出植物为适应环境变化,会将生物量分配给获得最有限资源的器官,即通过调整体内生物量的分配以最大限度地获取光、营养和水等有限的资源[27].异速生长理论表明,在自然条件下种内和种间成熟个体的生物量分配速率相同[20,27-28],也有研究得出生物量呈异速分配模式[13].有研究者提出上述两种假说是相辅相成的,在解释植物生物量分配方面可能是互补的而非独立的[26],但这还需要大量研究加以验证. ...
黑河中游荒漠草地地上和地下生物量的分配格局
1
2014
... 同样,在群落水平上,地上、地下生物量相关生长关系的研究也有很大发展.有学者发现中国草地生物量分配不符合等速生长假说[18],而Yang等[19]通过对青藏高原多个地点的调查研究发现,地上与地下生物量符合等速分配格局.程栋梁[20]研究了中国西北地区5个不同自然湿度梯度的植物生物量分配关系,发现干旱虽然会影响异速生长常数,但地上和地下生物量之间仍存在相关生长关系.王敏等[21]对黑河中游的地上和地下生物量进行了调查,发现无论是个体水平还是群落水平,地上与地下生物量均为等速生长关系.陶冶[22]对准噶尔荒漠典型植物群落生物的分配关系进行了研究,结果表明大部分草本植物的地上与地下生物量之间为等速生长关系,并且地上和地下生物量间的相关生长关系随生长期的变化而发生变化,在成熟期趋于等速生长.不同物种和群落间的地上与地下生物量生长关系存在差异,虽然诸多研究证明了草本植物群落地上与地下生物量之间具有等速生长关系,但是这种等速生长关系是否符合所有群落类型,特别是干旱荒漠区的草本植物,还没有一致结论. ...
准噶尔荒漠典型植物群落生物量的分配、估测与空间分布
1
2014
... 同样,在群落水平上,地上、地下生物量相关生长关系的研究也有很大发展.有学者发现中国草地生物量分配不符合等速生长假说[18],而Yang等[19]通过对青藏高原多个地点的调查研究发现,地上与地下生物量符合等速分配格局.程栋梁[20]研究了中国西北地区5个不同自然湿度梯度的植物生物量分配关系,发现干旱虽然会影响异速生长常数,但地上和地下生物量之间仍存在相关生长关系.王敏等[21]对黑河中游的地上和地下生物量进行了调查,发现无论是个体水平还是群落水平,地上与地下生物量均为等速生长关系.陶冶[22]对准噶尔荒漠典型植物群落生物的分配关系进行了研究,结果表明大部分草本植物的地上与地下生物量之间为等速生长关系,并且地上和地下生物量间的相关生长关系随生长期的变化而发生变化,在成熟期趋于等速生长.不同物种和群落间的地上与地下生物量生长关系存在差异,虽然诸多研究证明了草本植物群落地上与地下生物量之间具有等速生长关系,但是这种等速生长关系是否符合所有群落类型,特别是干旱荒漠区的草本植物,还没有一致结论. ...
青藏高原高寒草甸植物群落物种组成和生物量沿环境梯度的变化
1
2007
... 生物量是研究生态系统碳循环的基础,可以反映生态系统生产力的大小,不仅受到物种多样性、密度和均匀度的影响,还与物种本身的生物学特性和外部环境资源有关[23].生物量的分配是指植物将光合产物用于地上和地下不同器官中的比例,是植物对获取资源和利用资源的权衡,可以反映植物对不同的生长环境和选择压力的适应[24-26],会受到物种特征、个体发育和植物生存环境的强烈影响.关于生物量分配有两种流行的假设:最优分配和异速分配.最优分配理论指出植物为适应环境变化,会将生物量分配给获得最有限资源的器官,即通过调整体内生物量的分配以最大限度地获取光、营养和水等有限的资源[27].异速生长理论表明,在自然条件下种内和种间成熟个体的生物量分配速率相同[20,27-28],也有研究得出生物量呈异速分配模式[13].有研究者提出上述两种假说是相辅相成的,在解释植物生物量分配方面可能是互补的而非独立的[26],但这还需要大量研究加以验证. ...
Above-and below-ground biomass relationships across 1534 forested communities
1
2007
... 生物量是研究生态系统碳循环的基础,可以反映生态系统生产力的大小,不仅受到物种多样性、密度和均匀度的影响,还与物种本身的生物学特性和外部环境资源有关[23].生物量的分配是指植物将光合产物用于地上和地下不同器官中的比例,是植物对获取资源和利用资源的权衡,可以反映植物对不同的生长环境和选择压力的适应[24-26],会受到物种特征、个体发育和植物生存环境的强烈影响.关于生物量分配有两种流行的假设:最优分配和异速分配.最优分配理论指出植物为适应环境变化,会将生物量分配给获得最有限资源的器官,即通过调整体内生物量的分配以最大限度地获取光、营养和水等有限的资源[27].异速生长理论表明,在自然条件下种内和种间成熟个体的生物量分配速率相同[20,27-28],也有研究得出生物量呈异速分配模式[13].有研究者提出上述两种假说是相辅相成的,在解释植物生物量分配方面可能是互补的而非独立的[26],但这还需要大量研究加以验证. ...
Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation
0
2007
The role of biomass allocation in the growth response of plants to different levels of light,CO2,nutrients and water: a quantitative review
4
2000
... 生物量是研究生态系统碳循环的基础,可以反映生态系统生产力的大小,不仅受到物种多样性、密度和均匀度的影响,还与物种本身的生物学特性和外部环境资源有关[23].生物量的分配是指植物将光合产物用于地上和地下不同器官中的比例,是植物对获取资源和利用资源的权衡,可以反映植物对不同的生长环境和选择压力的适应[24-26],会受到物种特征、个体发育和植物生存环境的强烈影响.关于生物量分配有两种流行的假设:最优分配和异速分配.最优分配理论指出植物为适应环境变化,会将生物量分配给获得最有限资源的器官,即通过调整体内生物量的分配以最大限度地获取光、营养和水等有限的资源[27].异速生长理论表明,在自然条件下种内和种间成熟个体的生物量分配速率相同[20,27-28],也有研究得出生物量呈异速分配模式[13].有研究者提出上述两种假说是相辅相成的,在解释植物生物量分配方面可能是互补的而非独立的[26],但这还需要大量研究加以验证. ...
... [26],但这还需要大量研究加以验证. ...
... 目前关于草本植物群落/层片地上和地下生物量分配机制在异速生长假说和最优分配假说之间仍有争议.考虑到两种理论在研究生物量分配中的局限性,许多研究者将异速生长法和最优分配假说结合来进行了应用[42-44].在群落水平上,有诸多研究均证明了草本植物地上生物量与地下生物量之间的等速生长关系[19,28,45-46].从本研究结果来看,地上与地下生物量之间没有明显的相关生长关系,与马安娜等[47]对温带草原、山地草甸、温性草甸草原地上与地下生物量之间关系的研究结果类似.当去除类短命植物囊果苔草后,虽然草本层片地上生物量大于地下部分,但地上与地下生物量分配却符合等速生长理论,该结果与其他相关研究结果一致[41,46].有研究表明,不同草本植物群落生物量具有不同的分配规律,其原因可能是群落物种组成和生长类型的影响[48].群落内不同物种间因对环境的适应性不同或者是系统分类地位和遗传发育的差异[4],导致不同生活型的草本植物对群落生产力的贡献率不同,其中优势物种的特性在很大程度上决定了群落的生物量.因此,在研究区内,物种间的遗传差异使得各物种采取不同的生存策略,不同类型的植物为了适应环境具有不同的生物量分配策略,在资源有限的情况下,选择将更多生物量分配给遭受胁迫的器官,符合最优分配假说[26].但本研究在去除囊果苔草对整个层片影响的情况下,层片地上与地下生物量分配关系符合异速生长理论,可见最优分配假说与异速分配假说两者并不矛盾,在一定条件下可以结合起来[42],共同解释该草本植物层片生物量的不同分配模式. ...
... 密度制约效应是物种共存的重要调控机制[56].随着密度的增加,邻近植株间可获得的资源减少,继而引起对光资源、水分与矿质营养甚至是生存空间的竞争,相对应的植物也会采取不同的生物量分配策略来响应不同的种群密度[38].根冠比做为研究地上-地下生物量分配的有用工具,常用来表征植物光合产物在地上、地下生物量分配格局的变化[57],Poorter等[26]曾指出生物量分配受物种类型、个体发育和栖息地环境的影响. ...
Global Allocation rules for patterns of biomass partitioning in seed plants
2
2002
... 生物量是研究生态系统碳循环的基础,可以反映生态系统生产力的大小,不仅受到物种多样性、密度和均匀度的影响,还与物种本身的生物学特性和外部环境资源有关[23].生物量的分配是指植物将光合产物用于地上和地下不同器官中的比例,是植物对获取资源和利用资源的权衡,可以反映植物对不同的生长环境和选择压力的适应[24-26],会受到物种特征、个体发育和植物生存环境的强烈影响.关于生物量分配有两种流行的假设:最优分配和异速分配.最优分配理论指出植物为适应环境变化,会将生物量分配给获得最有限资源的器官,即通过调整体内生物量的分配以最大限度地获取光、营养和水等有限的资源[27].异速生长理论表明,在自然条件下种内和种间成熟个体的生物量分配速率相同[20,27-28],也有研究得出生物量呈异速分配模式[13].有研究者提出上述两种假说是相辅相成的,在解释植物生物量分配方面可能是互补的而非独立的[26],但这还需要大量研究加以验证. ...
... ,27-28],也有研究得出生物量呈异速分配模式[13].有研究者提出上述两种假说是相辅相成的,在解释植物生物量分配方面可能是互补的而非独立的[26],但这还需要大量研究加以验证. ...
A general model for the structure and allometry of plant vascular systems
2
1999
... 生物量是研究生态系统碳循环的基础,可以反映生态系统生产力的大小,不仅受到物种多样性、密度和均匀度的影响,还与物种本身的生物学特性和外部环境资源有关[23].生物量的分配是指植物将光合产物用于地上和地下不同器官中的比例,是植物对获取资源和利用资源的权衡,可以反映植物对不同的生长环境和选择压力的适应[24-26],会受到物种特征、个体发育和植物生存环境的强烈影响.关于生物量分配有两种流行的假设:最优分配和异速分配.最优分配理论指出植物为适应环境变化,会将生物量分配给获得最有限资源的器官,即通过调整体内生物量的分配以最大限度地获取光、营养和水等有限的资源[27].异速生长理论表明,在自然条件下种内和种间成熟个体的生物量分配速率相同[20,27-28],也有研究得出生物量呈异速分配模式[13].有研究者提出上述两种假说是相辅相成的,在解释植物生物量分配方面可能是互补的而非独立的[26],但这还需要大量研究加以验证. ...
... 目前关于草本植物群落/层片地上和地下生物量分配机制在异速生长假说和最优分配假说之间仍有争议.考虑到两种理论在研究生物量分配中的局限性,许多研究者将异速生长法和最优分配假说结合来进行了应用[42-44].在群落水平上,有诸多研究均证明了草本植物地上生物量与地下生物量之间的等速生长关系[19,28,45-46].从本研究结果来看,地上与地下生物量之间没有明显的相关生长关系,与马安娜等[47]对温带草原、山地草甸、温性草甸草原地上与地下生物量之间关系的研究结果类似.当去除类短命植物囊果苔草后,虽然草本层片地上生物量大于地下部分,但地上与地下生物量分配却符合等速生长理论,该结果与其他相关研究结果一致[41,46].有研究表明,不同草本植物群落生物量具有不同的分配规律,其原因可能是群落物种组成和生长类型的影响[48].群落内不同物种间因对环境的适应性不同或者是系统分类地位和遗传发育的差异[4],导致不同生活型的草本植物对群落生产力的贡献率不同,其中优势物种的特性在很大程度上决定了群落的生物量.因此,在研究区内,物种间的遗传差异使得各物种采取不同的生存策略,不同类型的植物为了适应环境具有不同的生物量分配策略,在资源有限的情况下,选择将更多生物量分配给遭受胁迫的器官,符合最优分配假说[26].但本研究在去除囊果苔草对整个层片影响的情况下,层片地上与地下生物量分配关系符合异速生长理论,可见最优分配假说与异速分配假说两者并不矛盾,在一定条件下可以结合起来[42],共同解释该草本植物层片生物量的不同分配模式. ...
1
2008
... 研究区位于新疆北部的古尔班通古特沙漠(44°11′—46°50′N、84°31′—91°20′E,面积4.88万km2).该沙漠年均蒸发量>2 000 mm,而年均降水量普遍不超过150 mm,沙漠腹地仅有70—100 mm,集中于冬春季;年均气温6—10 ℃,极端最高气温为40 ℃以上,≥10 ℃的年积温3 000—3 500 ℃,空气相对湿度平均50%—60%,5—8月通常小于45%[2].梭梭(Haloxylon ammodendron)和白梭梭(Haloxylon persicum)是准噶尔荒漠植物群落的主要灌木建群种.该地区具有丰富的短命和类短命植物,是早春荒漠生态系统的重要组成部分[5-6].此外,该沙漠还广泛分布着生物土壤结皮,包括藻结皮、地衣结皮和苔藓结皮3种主要类型,也是维持荒漠地表稳定的重要生物因子[29]. ...
幂指数异速生长机制模型综述
1
2008
... 式中:Y是某种生物学特征或者功能;β是标准化常数;X指个体大小;α是异速生长指数[30-31].α=1为等速关系,即因变量和自变量呈均匀或等比例变化;α≠1为异速关系,即因变量和自变量间为不均匀或不等比例变化.确定异速生长参数时,将幂函数转换为: ...
A general model for the origin of allometric scaling laws in biology
1
1997
... 式中:Y是某种生物学特征或者功能;β是标准化常数;X指个体大小;α是异速生长指数[30-31].α=1为等速关系,即因变量和自变量呈均匀或等比例变化;α≠1为异速关系,即因变量和自变量间为不均匀或不等比例变化.确定异速生长参数时,将幂函数转换为: ...
Invariant scaling relationships for interspecific plant biomass production rates and body size
1
2001
... 利用降主轴回归(reduced major axis regression,RMA)方法计算回归模型的指数、95%置信区间(95% CI)和决定系数(R2)[32-33].对数化之后的α即为幂函数对数化之后线性回归的斜率,lgβ为线性回归的截距.异速生长指数(α)和截距(lgβ)的计算、等速生长检验及α的方差分析均使用SMATR软件完成[34]. ...
Plant allometry:Is there a grand unifying theory?
1
2004
... 利用降主轴回归(reduced major axis regression,RMA)方法计算回归模型的指数、95%置信区间(95% CI)和决定系数(R2)[32-33].对数化之后的α即为幂函数对数化之后线性回归的斜率,lgβ为线性回归的截距.异速生长指数(α)和截距(lgβ)的计算、等速生长检验及α的方差分析均使用SMATR软件完成[34]. ...
1
Version 1
... 利用降主轴回归(reduced major axis regression,RMA)方法计算回归模型的指数、95%置信区间(95% CI)和决定系数(R2)[32-33].对数化之后的α即为幂函数对数化之后线性回归的斜率,lgβ为线性回归的截距.异速生长指数(α)和截距(lgβ)的计算、等速生长检验及α的方差分析均使用SMATR软件完成[34]. ...
The relationship between productivity and species richness
2
1999
... 式中:B是生物量;b0、b1、b2是回归常数[35]. ...
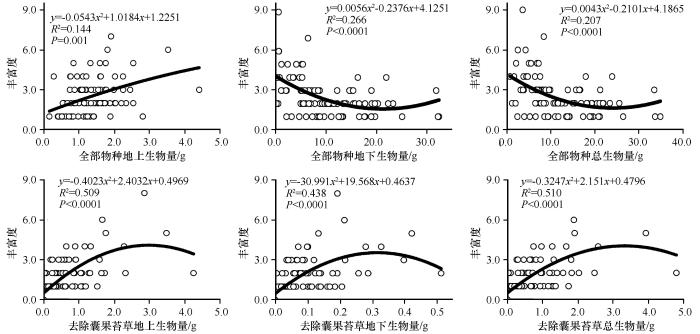
... 目前许多实验研究都支持植物物种多样性受生产力的影响,也有实验表明物种的数量和种类反过来也会影响生产力[35],但关于这个观点有以下两种争论,一是生产力的增加是由物种丰富度本身增加造成的,二是不同功能组或特定物种的加入导致生产力发生变化.多年生囊果苔草作为层片内的优势物种,对群落结构和功能以及稳定性的维持有重要的作用.因其具有发达的地下根系,在整个层片地下生物量占比很大,竞争能力较强,可以更有效地利用资源而创造更高的生产力[55],所以可以认为囊果苔草的加入与否,在很大程度影响了草本植物层片生产力于物种丰富度之间的关系,在某种程度上支持物种多样性影响群落生产力的观点. ...
Self-thinning in overcrowded pure stands under cultivated and natural conditions
1
1963
... 经典的密度制约理论常用关系式为[36-37]: ...
Plant energetics and population density-Reply
1
1999
... 经典的密度制约理论常用关系式为[36-37]: ...
密度制约决定的植物生物量分配格局
3
2011
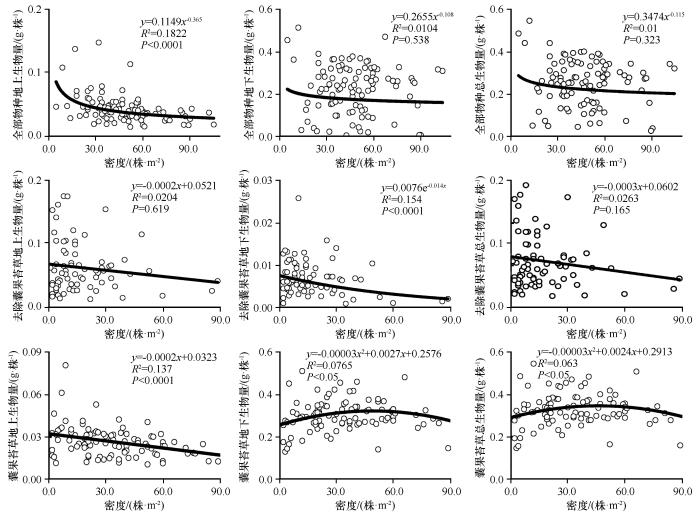
... 式中:Y代表每个样方平均个体地上、地下或总生物量;N为种群密度(单位面积个体数);C为常数项;α为比例指数或者异速指数,反映的是生物特征的密度依赖效应,也称为密度调控指数.也可以选择线性函数(y=a+bx)、指数函数(y=aebx)、二项式函数(y=ax2+bx+c)等相关程度最高的函数来表达生物量与密度之间的相关关系[38].除了个体生物量外,根冠比也被引入作为一个变量,探究植物密度变化是否影响层片水平植物地上与地下生物量的分配特征. ...
... 密度制约效应是物种共存的重要调控机制[56].随着密度的增加,邻近植株间可获得的资源减少,继而引起对光资源、水分与矿质营养甚至是生存空间的竞争,相对应的植物也会采取不同的生物量分配策略来响应不同的种群密度[38].根冠比做为研究地上-地下生物量分配的有用工具,常用来表征植物光合产物在地上、地下生物量分配格局的变化[57],Poorter等[26]曾指出生物量分配受物种类型、个体发育和栖息地环境的影响. ...
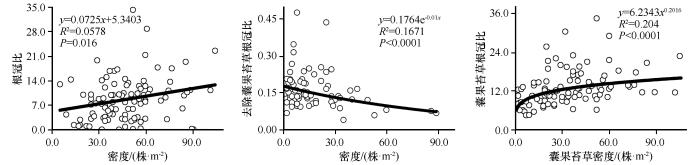
... 从本研究结果来看,去除群落内优势物种囊果苔草后,地上生物量不随密度变化,总生物量在不同密度下基本维持平衡,地下生物量随密度的增加而减小,根冠比与密度之间呈负相关关系.这说明随着密度的增加,群落内生活史短的植物之间存在对水分和养分的竞争作用,为了自身的营养和繁殖生长,以减小地下生物量的分配方式来维持对地上繁殖器官养分的供应[58].不论是整个草本植物层片还是囊果苔草,地上生物量都随密度的增加而减小,根冠比都随密度的增加而增大.优势物种囊果苔草的地上生物量和总生物量与密度之间呈现出先增加后减小的趋势,说明当囊果苔草的密度增加到一定值时,彼此根系之间也存在一定的密度制约作用,而整个层片水平的地下生物量和总生物量均不随密度的变化而变化,说明生存空间的扩大减弱了苔草彼此根系之间对空间的竞争作用.但类短命植物囊果苔草为维持自身庞大根系的正常生长和繁殖,在与群落内其他植株竞争养分过程中可能处于优势地位,会抑制群落中根相对浅或者少的植株生长,并且有研究表明这种竞争的不对称性会随着植物的生长而放大[59].虽然从层片水平来看,地下生物量不随密度的变化而变化,但地下仍存在植物对资源的竞争作用,会导致群落内的个体以牺牲地上器官的生物量分配保证种群的生存繁衍,维持群落的稳定[38].从两种不同的根冠比与密度关系可以看出,植物群落种类的不同导致彼此之间竞争强度存在差异,该不对称竞争对于有限环境资源条件下的物种共存非常重要.研究表明,密度是种群内植株个体生长、资源竞争和分配的重要影响因素[56],在环境胁迫下植物间的相互作用(不对称竞争)会随着密度的变化而发生变化[60]. ...
Allocation,plasticity and allometry
1
2004
... 生物量的分配是指植物生长发育过程中将光合产物用于地上、地下不同器官中的比例,体现了植物对资源的权衡和环境的适应[39].研究区草本植物层片地上和地下生物量变化范围均小于冀西北和黄土丘陵不同植被带的草本植物群落,但由于类短命植物的存在,使得根冠比的变化范围大于后两者[40-41].对于去除囊果苔草的草本层片来说,根冠比与地下生物量之间没有相关关系,随地上生物量和总生物量的增加而降低但变化范围较窄,表明去除囊果苔草的草本层片对繁殖分配和营养分配的权衡主要发生在地上部分.以层片中短命植物为例,为了在短时间内资源较为丰富的条件下快速完成生活史,选择将较多的能量投入到地上部分,以保证植物的有性繁殖和种群扩散[16].其中层片水平的地下生物量和总生物量与根冠比之间的关系与层片中囊果苔草的变化趋势相似,说明草本群落/层片水平的地下生物量受囊果苔草地下生物量的影响,使得整个草本层片的生物量分配倾向于根系部分.草本植物层片R/S随地上生物量的增大显著降低,随地下生物量和总生物量的增加显著增大,而且变化范围都较大,证明了群落/层片根冠比具有生物量大小依赖性. ...
冀西北中低山区灌草群落生物量分配及其与土壤因子的关系
1
2020
... 生物量的分配是指植物生长发育过程中将光合产物用于地上、地下不同器官中的比例,体现了植物对资源的权衡和环境的适应[39].研究区草本植物层片地上和地下生物量变化范围均小于冀西北和黄土丘陵不同植被带的草本植物群落,但由于类短命植物的存在,使得根冠比的变化范围大于后两者[40-41].对于去除囊果苔草的草本层片来说,根冠比与地下生物量之间没有相关关系,随地上生物量和总生物量的增加而降低但变化范围较窄,表明去除囊果苔草的草本层片对繁殖分配和营养分配的权衡主要发生在地上部分.以层片中短命植物为例,为了在短时间内资源较为丰富的条件下快速完成生活史,选择将较多的能量投入到地上部分,以保证植物的有性繁殖和种群扩散[16].其中层片水平的地下生物量和总生物量与根冠比之间的关系与层片中囊果苔草的变化趋势相似,说明草本群落/层片水平的地下生物量受囊果苔草地下生物量的影响,使得整个草本层片的生物量分配倾向于根系部分.草本植物层片R/S随地上生物量的增大显著降低,随地下生物量和总生物量的增加显著增大,而且变化范围都较大,证明了群落/层片根冠比具有生物量大小依赖性. ...
黄土丘陵区草本群落生物量空间分布格局及其影响因素
2
2018
... 生物量的分配是指植物生长发育过程中将光合产物用于地上、地下不同器官中的比例,体现了植物对资源的权衡和环境的适应[39].研究区草本植物层片地上和地下生物量变化范围均小于冀西北和黄土丘陵不同植被带的草本植物群落,但由于类短命植物的存在,使得根冠比的变化范围大于后两者[40-41].对于去除囊果苔草的草本层片来说,根冠比与地下生物量之间没有相关关系,随地上生物量和总生物量的增加而降低但变化范围较窄,表明去除囊果苔草的草本层片对繁殖分配和营养分配的权衡主要发生在地上部分.以层片中短命植物为例,为了在短时间内资源较为丰富的条件下快速完成生活史,选择将较多的能量投入到地上部分,以保证植物的有性繁殖和种群扩散[16].其中层片水平的地下生物量和总生物量与根冠比之间的关系与层片中囊果苔草的变化趋势相似,说明草本群落/层片水平的地下生物量受囊果苔草地下生物量的影响,使得整个草本层片的生物量分配倾向于根系部分.草本植物层片R/S随地上生物量的增大显著降低,随地下生物量和总生物量的增加显著增大,而且变化范围都较大,证明了群落/层片根冠比具有生物量大小依赖性. ...
... 目前关于草本植物群落/层片地上和地下生物量分配机制在异速生长假说和最优分配假说之间仍有争议.考虑到两种理论在研究生物量分配中的局限性,许多研究者将异速生长法和最优分配假说结合来进行了应用[42-44].在群落水平上,有诸多研究均证明了草本植物地上生物量与地下生物量之间的等速生长关系[19,28,45-46].从本研究结果来看,地上与地下生物量之间没有明显的相关生长关系,与马安娜等[47]对温带草原、山地草甸、温性草甸草原地上与地下生物量之间关系的研究结果类似.当去除类短命植物囊果苔草后,虽然草本层片地上生物量大于地下部分,但地上与地下生物量分配却符合等速生长理论,该结果与其他相关研究结果一致[41,46].有研究表明,不同草本植物群落生物量具有不同的分配规律,其原因可能是群落物种组成和生长类型的影响[48].群落内不同物种间因对环境的适应性不同或者是系统分类地位和遗传发育的差异[4],导致不同生活型的草本植物对群落生产力的贡献率不同,其中优势物种的特性在很大程度上决定了群落的生物量.因此,在研究区内,物种间的遗传差异使得各物种采取不同的生存策略,不同类型的植物为了适应环境具有不同的生物量分配策略,在资源有限的情况下,选择将更多生物量分配给遭受胁迫的器官,符合最优分配假说[26].但本研究在去除囊果苔草对整个层片影响的情况下,层片地上与地下生物量分配关系符合异速生长理论,可见最优分配假说与异速分配假说两者并不矛盾,在一定条件下可以结合起来[42],共同解释该草本植物层片生物量的不同分配模式. ...
Pitfalls and possibilities in the analysis of biomass allocation patterns in plants
2
2012
... 目前关于草本植物群落/层片地上和地下生物量分配机制在异速生长假说和最优分配假说之间仍有争议.考虑到两种理论在研究生物量分配中的局限性,许多研究者将异速生长法和最优分配假说结合来进行了应用[42-44].在群落水平上,有诸多研究均证明了草本植物地上生物量与地下生物量之间的等速生长关系[19,28,45-46].从本研究结果来看,地上与地下生物量之间没有明显的相关生长关系,与马安娜等[47]对温带草原、山地草甸、温性草甸草原地上与地下生物量之间关系的研究结果类似.当去除类短命植物囊果苔草后,虽然草本层片地上生物量大于地下部分,但地上与地下生物量分配却符合等速生长理论,该结果与其他相关研究结果一致[41,46].有研究表明,不同草本植物群落生物量具有不同的分配规律,其原因可能是群落物种组成和生长类型的影响[48].群落内不同物种间因对环境的适应性不同或者是系统分类地位和遗传发育的差异[4],导致不同生活型的草本植物对群落生产力的贡献率不同,其中优势物种的特性在很大程度上决定了群落的生物量.因此,在研究区内,物种间的遗传差异使得各物种采取不同的生存策略,不同类型的植物为了适应环境具有不同的生物量分配策略,在资源有限的情况下,选择将更多生物量分配给遭受胁迫的器官,符合最优分配假说[26].但本研究在去除囊果苔草对整个层片影响的情况下,层片地上与地下生物量分配关系符合异速生长理论,可见最优分配假说与异速分配假说两者并不矛盾,在一定条件下可以结合起来[42],共同解释该草本植物层片生物量的不同分配模式. ...
... [42],共同解释该草本植物层片生物量的不同分配模式. ...
甘肃南部7种高寒杜鹃生物量分配的异速生长关系
0
2020
川西北高寒草甸优势植物生物量分配对策
1
2018
... 目前关于草本植物群落/层片地上和地下生物量分配机制在异速生长假说和最优分配假说之间仍有争议.考虑到两种理论在研究生物量分配中的局限性,许多研究者将异速生长法和最优分配假说结合来进行了应用[42-44].在群落水平上,有诸多研究均证明了草本植物地上生物量与地下生物量之间的等速生长关系[19,28,45-46].从本研究结果来看,地上与地下生物量之间没有明显的相关生长关系,与马安娜等[47]对温带草原、山地草甸、温性草甸草原地上与地下生物量之间关系的研究结果类似.当去除类短命植物囊果苔草后,虽然草本层片地上生物量大于地下部分,但地上与地下生物量分配却符合等速生长理论,该结果与其他相关研究结果一致[41,46].有研究表明,不同草本植物群落生物量具有不同的分配规律,其原因可能是群落物种组成和生长类型的影响[48].群落内不同物种间因对环境的适应性不同或者是系统分类地位和遗传发育的差异[4],导致不同生活型的草本植物对群落生产力的贡献率不同,其中优势物种的特性在很大程度上决定了群落的生物量.因此,在研究区内,物种间的遗传差异使得各物种采取不同的生存策略,不同类型的植物为了适应环境具有不同的生物量分配策略,在资源有限的情况下,选择将更多生物量分配给遭受胁迫的器官,符合最优分配假说[26].但本研究在去除囊果苔草对整个层片影响的情况下,层片地上与地下生物量分配关系符合异速生长理论,可见最优分配假说与异速分配假说两者并不矛盾,在一定条件下可以结合起来[42],共同解释该草本植物层片生物量的不同分配模式. ...
中国北方典型草地物种丰富度与生产力的关系
1
2006
... 目前关于草本植物群落/层片地上和地下生物量分配机制在异速生长假说和最优分配假说之间仍有争议.考虑到两种理论在研究生物量分配中的局限性,许多研究者将异速生长法和最优分配假说结合来进行了应用[42-44].在群落水平上,有诸多研究均证明了草本植物地上生物量与地下生物量之间的等速生长关系[19,28,45-46].从本研究结果来看,地上与地下生物量之间没有明显的相关生长关系,与马安娜等[47]对温带草原、山地草甸、温性草甸草原地上与地下生物量之间关系的研究结果类似.当去除类短命植物囊果苔草后,虽然草本层片地上生物量大于地下部分,但地上与地下生物量分配却符合等速生长理论,该结果与其他相关研究结果一致[41,46].有研究表明,不同草本植物群落生物量具有不同的分配规律,其原因可能是群落物种组成和生长类型的影响[48].群落内不同物种间因对环境的适应性不同或者是系统分类地位和遗传发育的差异[4],导致不同生活型的草本植物对群落生产力的贡献率不同,其中优势物种的特性在很大程度上决定了群落的生物量.因此,在研究区内,物种间的遗传差异使得各物种采取不同的生存策略,不同类型的植物为了适应环境具有不同的生物量分配策略,在资源有限的情况下,选择将更多生物量分配给遭受胁迫的器官,符合最优分配假说[26].但本研究在去除囊果苔草对整个层片影响的情况下,层片地上与地下生物量分配关系符合异速生长理论,可见最优分配假说与异速分配假说两者并不矛盾,在一定条件下可以结合起来[42],共同解释该草本植物层片生物量的不同分配模式. ...
Biomass and nutrient allocation strategies in a desert ecosystem in the Hexi Corridor,northwest China
2
2017
... 目前关于草本植物群落/层片地上和地下生物量分配机制在异速生长假说和最优分配假说之间仍有争议.考虑到两种理论在研究生物量分配中的局限性,许多研究者将异速生长法和最优分配假说结合来进行了应用[42-44].在群落水平上,有诸多研究均证明了草本植物地上生物量与地下生物量之间的等速生长关系[19,28,45-46].从本研究结果来看,地上与地下生物量之间没有明显的相关生长关系,与马安娜等[47]对温带草原、山地草甸、温性草甸草原地上与地下生物量之间关系的研究结果类似.当去除类短命植物囊果苔草后,虽然草本层片地上生物量大于地下部分,但地上与地下生物量分配却符合等速生长理论,该结果与其他相关研究结果一致[41,46].有研究表明,不同草本植物群落生物量具有不同的分配规律,其原因可能是群落物种组成和生长类型的影响[48].群落内不同物种间因对环境的适应性不同或者是系统分类地位和遗传发育的差异[4],导致不同生活型的草本植物对群落生产力的贡献率不同,其中优势物种的特性在很大程度上决定了群落的生物量.因此,在研究区内,物种间的遗传差异使得各物种采取不同的生存策略,不同类型的植物为了适应环境具有不同的生物量分配策略,在资源有限的情况下,选择将更多生物量分配给遭受胁迫的器官,符合最优分配假说[26].但本研究在去除囊果苔草对整个层片影响的情况下,层片地上与地下生物量分配关系符合异速生长理论,可见最优分配假说与异速分配假说两者并不矛盾,在一定条件下可以结合起来[42],共同解释该草本植物层片生物量的不同分配模式. ...
... ,46].有研究表明,不同草本植物群落生物量具有不同的分配规律,其原因可能是群落物种组成和生长类型的影响[48].群落内不同物种间因对环境的适应性不同或者是系统分类地位和遗传发育的差异[4],导致不同生活型的草本植物对群落生产力的贡献率不同,其中优势物种的特性在很大程度上决定了群落的生物量.因此,在研究区内,物种间的遗传差异使得各物种采取不同的生存策略,不同类型的植物为了适应环境具有不同的生物量分配策略,在资源有限的情况下,选择将更多生物量分配给遭受胁迫的器官,符合最优分配假说[26].但本研究在去除囊果苔草对整个层片影响的情况下,层片地上与地下生物量分配关系符合异速生长理论,可见最优分配假说与异速分配假说两者并不矛盾,在一定条件下可以结合起来[42],共同解释该草本植物层片生物量的不同分配模式. ...
中国草地植被地上和地下生物量的关系分析
1
2014
... 目前关于草本植物群落/层片地上和地下生物量分配机制在异速生长假说和最优分配假说之间仍有争议.考虑到两种理论在研究生物量分配中的局限性,许多研究者将异速生长法和最优分配假说结合来进行了应用[42-44].在群落水平上,有诸多研究均证明了草本植物地上生物量与地下生物量之间的等速生长关系[19,28,45-46].从本研究结果来看,地上与地下生物量之间没有明显的相关生长关系,与马安娜等[47]对温带草原、山地草甸、温性草甸草原地上与地下生物量之间关系的研究结果类似.当去除类短命植物囊果苔草后,虽然草本层片地上生物量大于地下部分,但地上与地下生物量分配却符合等速生长理论,该结果与其他相关研究结果一致[41,46].有研究表明,不同草本植物群落生物量具有不同的分配规律,其原因可能是群落物种组成和生长类型的影响[48].群落内不同物种间因对环境的适应性不同或者是系统分类地位和遗传发育的差异[4],导致不同生活型的草本植物对群落生产力的贡献率不同,其中优势物种的特性在很大程度上决定了群落的生物量.因此,在研究区内,物种间的遗传差异使得各物种采取不同的生存策略,不同类型的植物为了适应环境具有不同的生物量分配策略,在资源有限的情况下,选择将更多生物量分配给遭受胁迫的器官,符合最优分配假说[26].但本研究在去除囊果苔草对整个层片影响的情况下,层片地上与地下生物量分配关系符合异速生长理论,可见最优分配假说与异速分配假说两者并不矛盾,在一定条件下可以结合起来[42],共同解释该草本植物层片生物量的不同分配模式. ...
Ecosystem stability and compensatory effects in the Inner Mongolia grassland
1
2004
... 目前关于草本植物群落/层片地上和地下生物量分配机制在异速生长假说和最优分配假说之间仍有争议.考虑到两种理论在研究生物量分配中的局限性,许多研究者将异速生长法和最优分配假说结合来进行了应用[42-44].在群落水平上,有诸多研究均证明了草本植物地上生物量与地下生物量之间的等速生长关系[19,28,45-46].从本研究结果来看,地上与地下生物量之间没有明显的相关生长关系,与马安娜等[47]对温带草原、山地草甸、温性草甸草原地上与地下生物量之间关系的研究结果类似.当去除类短命植物囊果苔草后,虽然草本层片地上生物量大于地下部分,但地上与地下生物量分配却符合等速生长理论,该结果与其他相关研究结果一致[41,46].有研究表明,不同草本植物群落生物量具有不同的分配规律,其原因可能是群落物种组成和生长类型的影响[48].群落内不同物种间因对环境的适应性不同或者是系统分类地位和遗传发育的差异[4],导致不同生活型的草本植物对群落生产力的贡献率不同,其中优势物种的特性在很大程度上决定了群落的生物量.因此,在研究区内,物种间的遗传差异使得各物种采取不同的生存策略,不同类型的植物为了适应环境具有不同的生物量分配策略,在资源有限的情况下,选择将更多生物量分配给遭受胁迫的器官,符合最优分配假说[26].但本研究在去除囊果苔草对整个层片影响的情况下,层片地上与地下生物量分配关系符合异速生长理论,可见最优分配假说与异速分配假说两者并不矛盾,在一定条件下可以结合起来[42],共同解释该草本植物层片生物量的不同分配模式. ...
What is the observed relationship between species richness and productivity?
1
2001
... 物种多样性是指生态系统中物种的丰富度,是评价一个地区生物资源丰富程度的重要指标,对维持生态系统的稳定性具有重要意义[14].物种多样性和生产力因受尺度、干扰、群落历史等因素的影响而呈现出正相关、负相关、U型、单峰型和不相关等多种表达形式[49].尽管目前已有许多研究表明干旱、半干旱区域草本植物群落的生物量与物种丰富度之间为单峰关系[50-52].但因沙漠生态系统生产力通常受到水分、养分有效性的限制,并且在空间和时间上具有高度可变性,地理位置、生物地理历史以及极端温度等因素也导致沙漠中的物种多样性差异很大,所以干旱生态系统中生产力和物种多样性之间还没有明确的关系[51].有研究认为有利的气候条件如高的降雨量,会增加物种丰富度和地下生物量,增大群落对极端气候的抵抗力[53].还有研究得出干旱胁迫虽然使得生物量显著减少,但并未改变群落物种组成和物种多样性,因为沙漠植物可塑性较高,会通过增加优势物种的生殖器官的生物量,来维持群落的物种丰富度和稳定性[54]. ...
科尔沁沙质草地物种多样性与生产力的关系
1
2007
... 物种多样性是指生态系统中物种的丰富度,是评价一个地区生物资源丰富程度的重要指标,对维持生态系统的稳定性具有重要意义[14].物种多样性和生产力因受尺度、干扰、群落历史等因素的影响而呈现出正相关、负相关、U型、单峰型和不相关等多种表达形式[49].尽管目前已有许多研究表明干旱、半干旱区域草本植物群落的生物量与物种丰富度之间为单峰关系[50-52].但因沙漠生态系统生产力通常受到水分、养分有效性的限制,并且在空间和时间上具有高度可变性,地理位置、生物地理历史以及极端温度等因素也导致沙漠中的物种多样性差异很大,所以干旱生态系统中生产力和物种多样性之间还没有明确的关系[51].有研究认为有利的气候条件如高的降雨量,会增加物种丰富度和地下生物量,增大群落对极端气候的抵抗力[53].还有研究得出干旱胁迫虽然使得生物量显著减少,但并未改变群落物种组成和物种多样性,因为沙漠植物可塑性较高,会通过增加优势物种的生殖器官的生物量,来维持群落的物种丰富度和稳定性[54]. ...
Species richness and biomass:dissection of the hump-shaped relationships
2
1998
... 物种多样性是指生态系统中物种的丰富度,是评价一个地区生物资源丰富程度的重要指标,对维持生态系统的稳定性具有重要意义[14].物种多样性和生产力因受尺度、干扰、群落历史等因素的影响而呈现出正相关、负相关、U型、单峰型和不相关等多种表达形式[49].尽管目前已有许多研究表明干旱、半干旱区域草本植物群落的生物量与物种丰富度之间为单峰关系[50-52].但因沙漠生态系统生产力通常受到水分、养分有效性的限制,并且在空间和时间上具有高度可变性,地理位置、生物地理历史以及极端温度等因素也导致沙漠中的物种多样性差异很大,所以干旱生态系统中生产力和物种多样性之间还没有明确的关系[51].有研究认为有利的气候条件如高的降雨量,会增加物种丰富度和地下生物量,增大群落对极端气候的抵抗力[53].还有研究得出干旱胁迫虽然使得生物量显著减少,但并未改变群落物种组成和物种多样性,因为沙漠植物可塑性较高,会通过增加优势物种的生殖器官的生物量,来维持群落的物种丰富度和稳定性[54]. ...
... 研究区草本植物层片地下生物量、总生物量与丰富度之间为倒U型关系,地上生物量与丰富度之间为负二次函数关系,且只显示出丰富度随地上生物量的增加而增加的阶段,未达到单峰曲线的最大值,可能的原因是该草本植物层片的生产力范围较小.有学者认为小尺度上存在的线性正相关、负相关以及不相关是较大尺度上观察到的单峰型关系的组成部分,即单峰型空间关系是正、负线性关系以及不相关累积的结果[51].本研究在去除群落内囊果苔草的条件下,草本植物层片的生物量与物种丰富度之间符合单峰关系,即群落/层片丰富度和多样性在中等生物量时达到顶峰,可根据竞争排除假说来解释. ...
Humped relationship between herbaceous species richness and biomass reveals a potential for increasing productivity in a temperate desert in Central Asia
1
2020
... 物种多样性是指生态系统中物种的丰富度,是评价一个地区生物资源丰富程度的重要指标,对维持生态系统的稳定性具有重要意义[14].物种多样性和生产力因受尺度、干扰、群落历史等因素的影响而呈现出正相关、负相关、U型、单峰型和不相关等多种表达形式[49].尽管目前已有许多研究表明干旱、半干旱区域草本植物群落的生物量与物种丰富度之间为单峰关系[50-52].但因沙漠生态系统生产力通常受到水分、养分有效性的限制,并且在空间和时间上具有高度可变性,地理位置、生物地理历史以及极端温度等因素也导致沙漠中的物种多样性差异很大,所以干旱生态系统中生产力和物种多样性之间还没有明确的关系[51].有研究认为有利的气候条件如高的降雨量,会增加物种丰富度和地下生物量,增大群落对极端气候的抵抗力[53].还有研究得出干旱胁迫虽然使得生物量显著减少,但并未改变群落物种组成和物种多样性,因为沙漠植物可塑性较高,会通过增加优势物种的生殖器官的生物量,来维持群落的物种丰富度和稳定性[54]. ...
Plant diversity enhances productivity and soil carbon storage
1
2018
... 物种多样性是指生态系统中物种的丰富度,是评价一个地区生物资源丰富程度的重要指标,对维持生态系统的稳定性具有重要意义[14].物种多样性和生产力因受尺度、干扰、群落历史等因素的影响而呈现出正相关、负相关、U型、单峰型和不相关等多种表达形式[49].尽管目前已有许多研究表明干旱、半干旱区域草本植物群落的生物量与物种丰富度之间为单峰关系[50-52].但因沙漠生态系统生产力通常受到水分、养分有效性的限制,并且在空间和时间上具有高度可变性,地理位置、生物地理历史以及极端温度等因素也导致沙漠中的物种多样性差异很大,所以干旱生态系统中生产力和物种多样性之间还没有明确的关系[51].有研究认为有利的气候条件如高的降雨量,会增加物种丰富度和地下生物量,增大群落对极端气候的抵抗力[53].还有研究得出干旱胁迫虽然使得生物量显著减少,但并未改变群落物种组成和物种多样性,因为沙漠植物可塑性较高,会通过增加优势物种的生殖器官的生物量,来维持群落的物种丰富度和稳定性[54]. ...
Extreme drought affects the productivity,but not the composition,of a desert plant community in Central Asia differentially across microtopographies
1
2020
... 物种多样性是指生态系统中物种的丰富度,是评价一个地区生物资源丰富程度的重要指标,对维持生态系统的稳定性具有重要意义[14].物种多样性和生产力因受尺度、干扰、群落历史等因素的影响而呈现出正相关、负相关、U型、单峰型和不相关等多种表达形式[49].尽管目前已有许多研究表明干旱、半干旱区域草本植物群落的生物量与物种丰富度之间为单峰关系[50-52].但因沙漠生态系统生产力通常受到水分、养分有效性的限制,并且在空间和时间上具有高度可变性,地理位置、生物地理历史以及极端温度等因素也导致沙漠中的物种多样性差异很大,所以干旱生态系统中生产力和物种多样性之间还没有明确的关系[51].有研究认为有利的气候条件如高的降雨量,会增加物种丰富度和地下生物量,增大群落对极端气候的抵抗力[53].还有研究得出干旱胁迫虽然使得生物量显著减少,但并未改变群落物种组成和物种多样性,因为沙漠植物可塑性较高,会通过增加优势物种的生殖器官的生物量,来维持群落的物种丰富度和稳定性[54]. ...
Plant diversity and ecosystem?productivity:theoretical considerations
1
1997
... 目前许多实验研究都支持植物物种多样性受生产力的影响,也有实验表明物种的数量和种类反过来也会影响生产力[35],但关于这个观点有以下两种争论,一是生产力的增加是由物种丰富度本身增加造成的,二是不同功能组或特定物种的加入导致生产力发生变化.多年生囊果苔草作为层片内的优势物种,对群落结构和功能以及稳定性的维持有重要的作用.因其具有发达的地下根系,在整个层片地下生物量占比很大,竞争能力较强,可以更有效地利用资源而创造更高的生产力[55],所以可以认为囊果苔草的加入与否,在很大程度影响了草本植物层片生产力于物种丰富度之间的关系,在某种程度上支持物种多样性影响群落生产力的观点. ...
Plant diversity in tropical forests: a review of mechanisms of species coexistence
2
2002
... 密度制约效应是物种共存的重要调控机制[56].随着密度的增加,邻近植株间可获得的资源减少,继而引起对光资源、水分与矿质营养甚至是生存空间的竞争,相对应的植物也会采取不同的生物量分配策略来响应不同的种群密度[38].根冠比做为研究地上-地下生物量分配的有用工具,常用来表征植物光合产物在地上、地下生物量分配格局的变化[57],Poorter等[26]曾指出生物量分配受物种类型、个体发育和栖息地环境的影响. ...
... 从本研究结果来看,去除群落内优势物种囊果苔草后,地上生物量不随密度变化,总生物量在不同密度下基本维持平衡,地下生物量随密度的增加而减小,根冠比与密度之间呈负相关关系.这说明随着密度的增加,群落内生活史短的植物之间存在对水分和养分的竞争作用,为了自身的营养和繁殖生长,以减小地下生物量的分配方式来维持对地上繁殖器官养分的供应[58].不论是整个草本植物层片还是囊果苔草,地上生物量都随密度的增加而减小,根冠比都随密度的增加而增大.优势物种囊果苔草的地上生物量和总生物量与密度之间呈现出先增加后减小的趋势,说明当囊果苔草的密度增加到一定值时,彼此根系之间也存在一定的密度制约作用,而整个层片水平的地下生物量和总生物量均不随密度的变化而变化,说明生存空间的扩大减弱了苔草彼此根系之间对空间的竞争作用.但类短命植物囊果苔草为维持自身庞大根系的正常生长和繁殖,在与群落内其他植株竞争养分过程中可能处于优势地位,会抑制群落中根相对浅或者少的植株生长,并且有研究表明这种竞争的不对称性会随着植物的生长而放大[59].虽然从层片水平来看,地下生物量不随密度的变化而变化,但地下仍存在植物对资源的竞争作用,会导致群落内的个体以牺牲地上器官的生物量分配保证种群的生存繁衍,维持群落的稳定[38].从两种不同的根冠比与密度关系可以看出,植物群落种类的不同导致彼此之间竞争强度存在差异,该不对称竞争对于有限环境资源条件下的物种共存非常重要.研究表明,密度是种群内植株个体生长、资源竞争和分配的重要影响因素[56],在环境胁迫下植物间的相互作用(不对称竞争)会随着密度的变化而发生变化[60]. ...
Critical analysis of root:shoot ratios in terrestrial biomes
1
2010
... 密度制约效应是物种共存的重要调控机制[56].随着密度的增加,邻近植株间可获得的资源减少,继而引起对光资源、水分与矿质营养甚至是生存空间的竞争,相对应的植物也会采取不同的生物量分配策略来响应不同的种群密度[38].根冠比做为研究地上-地下生物量分配的有用工具,常用来表征植物光合产物在地上、地下生物量分配格局的变化[57],Poorter等[26]曾指出生物量分配受物种类型、个体发育和栖息地环境的影响. ...
准噶尔荒漠早春短命植物的光合特性及生物量分配特点
1
2007
... 从本研究结果来看,去除群落内优势物种囊果苔草后,地上生物量不随密度变化,总生物量在不同密度下基本维持平衡,地下生物量随密度的增加而减小,根冠比与密度之间呈负相关关系.这说明随着密度的增加,群落内生活史短的植物之间存在对水分和养分的竞争作用,为了自身的营养和繁殖生长,以减小地下生物量的分配方式来维持对地上繁殖器官养分的供应[58].不论是整个草本植物层片还是囊果苔草,地上生物量都随密度的增加而减小,根冠比都随密度的增加而增大.优势物种囊果苔草的地上生物量和总生物量与密度之间呈现出先增加后减小的趋势,说明当囊果苔草的密度增加到一定值时,彼此根系之间也存在一定的密度制约作用,而整个层片水平的地下生物量和总生物量均不随密度的变化而变化,说明生存空间的扩大减弱了苔草彼此根系之间对空间的竞争作用.但类短命植物囊果苔草为维持自身庞大根系的正常生长和繁殖,在与群落内其他植株竞争养分过程中可能处于优势地位,会抑制群落中根相对浅或者少的植株生长,并且有研究表明这种竞争的不对称性会随着植物的生长而放大[59].虽然从层片水平来看,地下生物量不随密度的变化而变化,但地下仍存在植物对资源的竞争作用,会导致群落内的个体以牺牲地上器官的生物量分配保证种群的生存繁衍,维持群落的稳定[38].从两种不同的根冠比与密度关系可以看出,植物群落种类的不同导致彼此之间竞争强度存在差异,该不对称竞争对于有限环境资源条件下的物种共存非常重要.研究表明,密度是种群内植株个体生长、资源竞争和分配的重要影响因素[56],在环境胁迫下植物间的相互作用(不对称竞争)会随着密度的变化而发生变化[60]. ...
Empirical corn yield loss estimation from common lambsquarters (Chenopodium album) and giant foxtail (Setaria faberi) in mixed communities
1
2003
... 从本研究结果来看,去除群落内优势物种囊果苔草后,地上生物量不随密度变化,总生物量在不同密度下基本维持平衡,地下生物量随密度的增加而减小,根冠比与密度之间呈负相关关系.这说明随着密度的增加,群落内生活史短的植物之间存在对水分和养分的竞争作用,为了自身的营养和繁殖生长,以减小地下生物量的分配方式来维持对地上繁殖器官养分的供应[58].不论是整个草本植物层片还是囊果苔草,地上生物量都随密度的增加而减小,根冠比都随密度的增加而增大.优势物种囊果苔草的地上生物量和总生物量与密度之间呈现出先增加后减小的趋势,说明当囊果苔草的密度增加到一定值时,彼此根系之间也存在一定的密度制约作用,而整个层片水平的地下生物量和总生物量均不随密度的变化而变化,说明生存空间的扩大减弱了苔草彼此根系之间对空间的竞争作用.但类短命植物囊果苔草为维持自身庞大根系的正常生长和繁殖,在与群落内其他植株竞争养分过程中可能处于优势地位,会抑制群落中根相对浅或者少的植株生长,并且有研究表明这种竞争的不对称性会随着植物的生长而放大[59].虽然从层片水平来看,地下生物量不随密度的变化而变化,但地下仍存在植物对资源的竞争作用,会导致群落内的个体以牺牲地上器官的生物量分配保证种群的生存繁衍,维持群落的稳定[38].从两种不同的根冠比与密度关系可以看出,植物群落种类的不同导致彼此之间竞争强度存在差异,该不对称竞争对于有限环境资源条件下的物种共存非常重要.研究表明,密度是种群内植株个体生长、资源竞争和分配的重要影响因素[56],在环境胁迫下植物间的相互作用(不对称竞争)会随着密度的变化而发生变化[60]. ...
Density-dependence tips the change of plant-plant interactions under environmental stress
1
2020
... 从本研究结果来看,去除群落内优势物种囊果苔草后,地上生物量不随密度变化,总生物量在不同密度下基本维持平衡,地下生物量随密度的增加而减小,根冠比与密度之间呈负相关关系.这说明随着密度的增加,群落内生活史短的植物之间存在对水分和养分的竞争作用,为了自身的营养和繁殖生长,以减小地下生物量的分配方式来维持对地上繁殖器官养分的供应[58].不论是整个草本植物层片还是囊果苔草,地上生物量都随密度的增加而减小,根冠比都随密度的增加而增大.优势物种囊果苔草的地上生物量和总生物量与密度之间呈现出先增加后减小的趋势,说明当囊果苔草的密度增加到一定值时,彼此根系之间也存在一定的密度制约作用,而整个层片水平的地下生物量和总生物量均不随密度的变化而变化,说明生存空间的扩大减弱了苔草彼此根系之间对空间的竞争作用.但类短命植物囊果苔草为维持自身庞大根系的正常生长和繁殖,在与群落内其他植株竞争养分过程中可能处于优势地位,会抑制群落中根相对浅或者少的植株生长,并且有研究表明这种竞争的不对称性会随着植物的生长而放大[59].虽然从层片水平来看,地下生物量不随密度的变化而变化,但地下仍存在植物对资源的竞争作用,会导致群落内的个体以牺牲地上器官的生物量分配保证种群的生存繁衍,维持群落的稳定[38].从两种不同的根冠比与密度关系可以看出,植物群落种类的不同导致彼此之间竞争强度存在差异,该不对称竞争对于有限环境资源条件下的物种共存非常重要.研究表明,密度是种群内植株个体生长、资源竞争和分配的重要影响因素[56],在环境胁迫下植物间的相互作用(不对称竞争)会随着密度的变化而发生变化[60]. ...
Conspecific negative density dependence of trees varies with plant functional traits and environmental conditions across scales in a 60-ha tropical rainforest dynamics plot
1
2021
... 无论是温带森林还是热带森林,都存在负密度制约效应,并且密度制约效应在不同物种之间存在差异[61-62].杨元武等[63]曾提出密度调控指数不是恒定的,干旱与盐分胁迫、土壤养分等环境因素都会通过影响植物间的相互作用,进而影响密度调控规律.邓建明[64]研究表明,随着干旱胁迫强度的加大,地上部的α的绝对值将变小,而地下部α接近 -4/3,即植物地上和地下器官的α对环境有不同的响应.本研究得出的群落地上生物量与密度之间的调控指数为-0.365,不仅大于湿润梯度较大的浙江省天目山的密度调控指数-1.65—-1.3[65],还大于理论预测的-3/2、-4/3.主要原因是森林群落不受水资源限制的,其自疏效应主要是由冠层对光的竞争引起的[66],而干旱荒漠区植被大都是开放的冠层,引起密度调节的主要驱动因素是根[64,67]. ...
Density dependence is not very prevalent in a heterogeneous subtropical forest
1
2012
... 无论是温带森林还是热带森林,都存在负密度制约效应,并且密度制约效应在不同物种之间存在差异[61-62].杨元武等[63]曾提出密度调控指数不是恒定的,干旱与盐分胁迫、土壤养分等环境因素都会通过影响植物间的相互作用,进而影响密度调控规律.邓建明[64]研究表明,随着干旱胁迫强度的加大,地上部的α的绝对值将变小,而地下部α接近 -4/3,即植物地上和地下器官的α对环境有不同的响应.本研究得出的群落地上生物量与密度之间的调控指数为-0.365,不仅大于湿润梯度较大的浙江省天目山的密度调控指数-1.65—-1.3[65],还大于理论预测的-3/2、-4/3.主要原因是森林群落不受水资源限制的,其自疏效应主要是由冠层对光的竞争引起的[66],而干旱荒漠区植被大都是开放的冠层,引起密度调节的主要驱动因素是根[64,67]. ...
植物密度调控及其对环境变化响应的研究进展
1
2011
... 无论是温带森林还是热带森林,都存在负密度制约效应,并且密度制约效应在不同物种之间存在差异[61-62].杨元武等[63]曾提出密度调控指数不是恒定的,干旱与盐分胁迫、土壤养分等环境因素都会通过影响植物间的相互作用,进而影响密度调控规律.邓建明[64]研究表明,随着干旱胁迫强度的加大,地上部的α的绝对值将变小,而地下部α接近 -4/3,即植物地上和地下器官的α对环境有不同的响应.本研究得出的群落地上生物量与密度之间的调控指数为-0.365,不仅大于湿润梯度较大的浙江省天目山的密度调控指数-1.65—-1.3[65],还大于理论预测的-3/2、-4/3.主要原因是森林群落不受水资源限制的,其自疏效应主要是由冠层对光的竞争引起的[66],而干旱荒漠区植被大都是开放的冠层,引起密度调节的主要驱动因素是根[64,67]. ...
水分梯度条件下植物群体调控的异速比例变化规律研究
2
2007
... 无论是温带森林还是热带森林,都存在负密度制约效应,并且密度制约效应在不同物种之间存在差异[61-62].杨元武等[63]曾提出密度调控指数不是恒定的,干旱与盐分胁迫、土壤养分等环境因素都会通过影响植物间的相互作用,进而影响密度调控规律.邓建明[64]研究表明,随着干旱胁迫强度的加大,地上部的α的绝对值将变小,而地下部α接近 -4/3,即植物地上和地下器官的α对环境有不同的响应.本研究得出的群落地上生物量与密度之间的调控指数为-0.365,不仅大于湿润梯度较大的浙江省天目山的密度调控指数-1.65—-1.3[65],还大于理论预测的-3/2、-4/3.主要原因是森林群落不受水资源限制的,其自疏效应主要是由冠层对光的竞争引起的[66],而干旱荒漠区植被大都是开放的冠层,引起密度调节的主要驱动因素是根[64,67]. ...
... [64,67]. ...
Plant height-crown radius and canopy coverage-density relationships determine above-ground biomass-density relationship in stressful environments
1
2009
... 无论是温带森林还是热带森林,都存在负密度制约效应,并且密度制约效应在不同物种之间存在差异[61-62].杨元武等[63]曾提出密度调控指数不是恒定的,干旱与盐分胁迫、土壤养分等环境因素都会通过影响植物间的相互作用,进而影响密度调控规律.邓建明[64]研究表明,随着干旱胁迫强度的加大,地上部的α的绝对值将变小,而地下部α接近 -4/3,即植物地上和地下器官的α对环境有不同的响应.本研究得出的群落地上生物量与密度之间的调控指数为-0.365,不仅大于湿润梯度较大的浙江省天目山的密度调控指数-1.65—-1.3[65],还大于理论预测的-3/2、-4/3.主要原因是森林群落不受水资源限制的,其自疏效应主要是由冠层对光的竞争引起的[66],而干旱荒漠区植被大都是开放的冠层,引起密度调节的主要驱动因素是根[64,67]. ...
植物光合产物分配及其影响因子研究进展
1
2010
... 无论是温带森林还是热带森林,都存在负密度制约效应,并且密度制约效应在不同物种之间存在差异[61-62].杨元武等[63]曾提出密度调控指数不是恒定的,干旱与盐分胁迫、土壤养分等环境因素都会通过影响植物间的相互作用,进而影响密度调控规律.邓建明[64]研究表明,随着干旱胁迫强度的加大,地上部的α的绝对值将变小,而地下部α接近 -4/3,即植物地上和地下器官的α对环境有不同的响应.本研究得出的群落地上生物量与密度之间的调控指数为-0.365,不仅大于湿润梯度较大的浙江省天目山的密度调控指数-1.65—-1.3[65],还大于理论预测的-3/2、-4/3.主要原因是森林群落不受水资源限制的,其自疏效应主要是由冠层对光的竞争引起的[66],而干旱荒漠区植被大都是开放的冠层,引起密度调节的主要驱动因素是根[64,67]. ...
植物地下和地上密度调控的差异随湿润度梯度变化的定量规律研究
1
2011
... 无论是温带森林还是热带森林,都存在负密度制约效应,并且密度制约效应在不同物种之间存在差异[61-62].杨元武等[63]曾提出密度调控指数不是恒定的,干旱与盐分胁迫、土壤养分等环境因素都会通过影响植物间的相互作用,进而影响密度调控规律.邓建明[64]研究表明,随着干旱胁迫强度的加大,地上部的α的绝对值将变小,而地下部α接近 -4/3,即植物地上和地下器官的α对环境有不同的响应.本研究得出的群落地上生物量与密度之间的调控指数为-0.365,不仅大于湿润梯度较大的浙江省天目山的密度调控指数-1.65—-1.3[65],还大于理论预测的-3/2、-4/3.主要原因是森林群落不受水资源限制的,其自疏效应主要是由冠层对光的竞争引起的[66],而干旱荒漠区植被大都是开放的冠层,引起密度调节的主要驱动因素是根[64,67]. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}