荒漠绿洲湿地水分来源及植物水分利用策略
赵颖 , 1 , 2 , 刘冰 , 1 , 赵文智 1 , 温紫娟 3 , 王宵 1 , 2
1.中国科学院西北生态环境资源研究院 临泽内陆河流域研究站/中国科学院内陆河流域生态水文重点实验室,甘肃 兰州 730000
2.中国科学院大学,北京 100049
3.兰州大学,甘肃 兰州 730000
Water sources and strategies of plant water use in desert oasis wetland of China
Zhao Ying , 1 , 2 , Liu Bing , 1 , Zhao Wenzhi 1 , Wen Zijuan 3 , Wang Xiao 1 , 2
1.Linze Inland River Basin Research Station / Key Laboratory of Ecohydrology of Inland River Basin,Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
2.University of Chinese Academy of Sciences,Beijing 100049,China
3.Lanzhou University,Lanzhou 730000,China
通讯作者: 刘冰(E-mail: liubing@lzb.ac.cn )
收稿日期: 2021-09-04
修回日期: 2021-12-20
基金资助:
中国科学院A类战略性先导科技专项课题 . XDA23060304 国家自然科学基金项目 . 41771038 . 42071048
Received: 2021-09-04
Revised: 2021-12-20
摘要
水分是干旱区不同景观界面间的水分循环过程与水力联系的主体,维持着干旱区湿地生态系统的结构与功能。为量化水分来源及其对植物水分的贡献率,以河岸灌木湿地和草地盐沼湿地为研究对象,通过测定降水、径流、地下水、土壤水和植物水中δ D、δ 18 O组成,利用多源线性混合模型分析水分来源对荒漠植物水分利用的贡献率。结果表明:(1) 黑河流域荒漠绿洲湿地年均降水量104.6 mm,约占蒸散量(604.47 mm)的17.03%,具有明显的季节性分布特征。地下水位与土壤含水量的波动取决于河流距离,离河道较近的河岸灌木湿地地下水深度及土壤含水量随季节波动较小,而离河道较远的草地盐沼湿地则变化很大。(2) 当地大气降水线δ D=6.33δ 18 O+4.04 (R 2 =0.931),斜率和截距均略小于全球大气降水线则符合黑河流域湿地整体降水少而蒸散量大的特点。黑河径流δ D和δ 18 O均值分别为-43.80‰±12.09‰和-8.65‰±23.33‰,地下水为-50.98‰±13.18‰和-9.74‰±25.49‰,土壤水为 -42.07‰±6.89‰和-7.22‰±2.49‰,植物水为-51.84‰±14.46‰和-8.50‰±24.13‰。(3) 地表蒸发是荒漠绿洲湿地土壤氢、氧同位素富集的主要原因。地下水和河水分别是草地盐沼湿地与河岸灌木湿地的主要水分来源,贡献率分别约为61%和50%,表明湿地植物相比于干旱区脉冲式降水更依赖较为稳定的水源。(4) 植物根系深度和毛细根分布是决定荒漠绿洲湿地植物水分利用策略的重要因素。
关键词:
荒漠绿洲湿地 水分来源 氢氧同位素 贡献率 水分利用策略
Abstract
Water is the backbone of hydraulic connection and water cycle process between different landscape in arid area, which further maintains the structure and function of wetland ecosystem in arid area. In order to quantify water source and its contribution to plant water, this paper takes riparian shrub wetland and salt-marsh grassland wetland as research objects, by measuring the δ D and δ 18 O composition of precipitation, runoff, groundwater, soil water and plant water, to analyze the contribution rate of water sources to water use of desert plants with the linear mixed model. The results showed that: (1) annual rainfall average 104.6 mm, accounting for about 17.03% of the evapotranspiration (604.47 mm) in desert oasis wetlands in which has obvious seasonal distribution characteristics. The fluctuation of groundwater and soil water content depends on the distance from the river. The depth of groundwater and soil water content of riparian shrub wetlands near the river vary little with the seasons, while vary greatly in salt marsh grass wetlands far away from the river channel. (2) The precipitation line of local atmospheric is δ D=6.33δ 18 O+4.04 (R 2 =0.931), and the slope and intercept are slightly smaller than the precipitation line of global atmospheric, which is consistent with the characteristics of less precipitation and large evapotranspiration in the wetlands of the Heihe River Basin. The mean values of δ D and δ 18 O of Heihe River groundwater, runoff, soil waterand and plant water were -50.98‰±13.18‰ and -9.74‰±25.49‰, -43.80‰±12.09‰ and -8.65‰±23.33‰, -42.07‰±6.89‰ and -7.22‰±2.49‰, -51.84‰±14.46‰ and -8.50‰±24.13‰, respectively. (3) Surface evaporation is the main cause of soil hydrogen and oxygen isotope enrichment in desert oasis wetland. Runoff and groundwater were the main water sources of desert oasis wetlands,and the contribution rate of runoff to plant water use was about 50% in riparian shrub wetland, while the contribution rate of groundwater was about 61% in salt marsh grass wetland. However, precipitation infiltration contributed little to the water use of riparian shrub wetlands and salt marsh grass wetlands, with values of 4% and 13%, respectively. (4) Depth of root and distribution of capillary root were important determinants of plant water use strategy in desert oasis wetland.
Keywords:
desert oasis wetland water sources hydrogen and oxygen isotopes contribution rate water use strategy
本文引用格式
赵颖, 刘冰, 赵文智, 温紫娟, 王宵. 荒漠绿洲湿地水分来源及植物水分利用策略 . 中国沙漠 [J], 2022, 42(4): 151-162 doi:10.7522/j.issn.1000-694X.2021.00157
Zhao Ying, Liu Bing, Zhao Wenzhi, Wen Zijuan, Wang Xiao. Water sources and strategies of plant water use in desert oasis wetland of China . Journal of Desert Research
0 引言
在干旱区,湿地作为荒漠绿洲核心地理单元,不仅在涵养水源、净化水质、蓄洪抗旱、调节气候和维护生物多样性等方面发挥着重要作用[1 ] ,而且对河岸林锐减、绿洲萎缩、草场退化和荒漠化等具有抑制功能,在维持区域生态平衡与保障内陆河流域可持续发展方面具有生态屏障作用[2 ] 。气候变化通过对气温、降水以及干旱洪涝极端水文事件发生频率和强度的影响,改变着湿地蒸散发、径流、水位、水文周期等关键水文过程[3 ] ,调控着湿地水分来源、补给途径及其植物水分利用策略,对荒漠绿洲湿地植物区系组成、种群格局和稳定性产生深远影响[4 ] 。与此同时,农田开垦、灌溉引水等人类活动通过改变植被覆盖和径流过程间接或直接影响着湿地水文过程,加剧了水资源短缺和地下水枯竭,使荒漠绿洲湿地出现面积萎缩、植被退化、多样性减少、盐碱化加剧等生态环境问题[5 ] 。所以,人类活动与气候变化共同作用是荒漠绿洲湿地环境恶化的主要驱动力,定量研究荒漠绿洲湿地水分来源、补给途径及其植物水分利用策略,是解决荒漠绿洲湿地环境恶化问题的前提条件,将为干旱区湿地环境治理和生态安全提供理论依据与决策支持。
干旱区降水稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[4 -5 ] 。通常,受植被类型、水分状况、河流径流以及地下水位的影响,植物水分来源与利用方式的辨析与识别具有高度的变异性和复杂性[6 ] 。研究表明,除极少数盐生植物外,陆地植物根系吸收水分在木质部运输过程中一般不发生同位素分馏,因此植物茎木质部提取的氢氧稳定同位素比率可反映植物的水分来源[7 ] ,为荒漠绿洲湿地水分来源的辨析提供了水文学依据。目前,国内学者基于稳定同位素技术在荒漠、河岸、海岸等系统中乔灌草等不同生活型植物水分来源和利用策略研究中开展了广泛应用,包括干旱半干旱区[8 -10 ] 、川西亚高山区[11 ] 、青海湖流域[12 ] 和黑河流域[13 -14 ] 等,为探明湿地水分来源、补给途径及其植物水分利用策略提供了思路与技术。然而,基于稳定同位素技术研究干旱区湿地水分来源、补给途径及其植物水分利用策略的研究迄今为止仍未见报道。
在黑河流域,荒漠绿洲湿地主要分布在河流源头、绿洲、河滩和内陆湖滨等季节性或常年性积水地段,是河西走廊甚至西北地区重要的水源涵养地,对黑河流域生态安全及社会经济发展发挥重要作用[14 ] 。因此,以黑河流域中游河岸灌木湿地与草地盐沼湿地为研究对象,基于稳定同位素技术研究降水、径流、地下水、土壤水和植物水的同位素组成,探究不同水分来源对荒漠植物水分利用的贡献率,将为解释干旱区湿地植物水分利用机理及适应策略提供科学依据和理论基础。
1 材料和方法
1.1 研究区概况
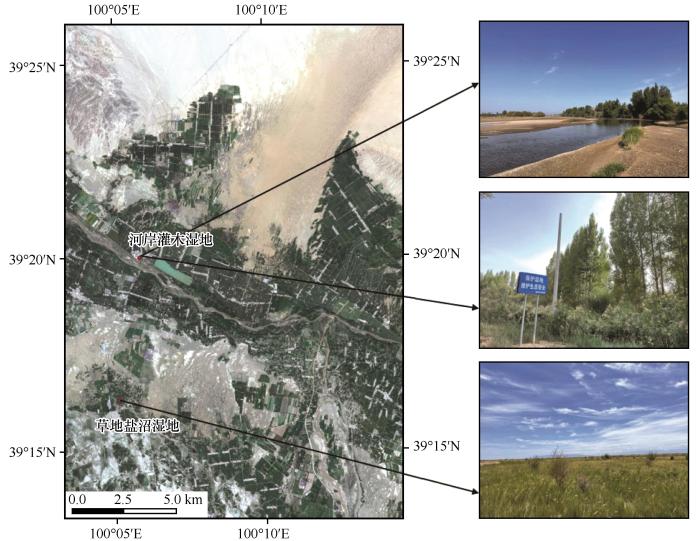
研究区位于黑河中游张掖国家湿地保护区临泽县境内,为典型的荒漠绿洲湿地(图1 )。该区属温带大陆性干旱气候,夏季干燥炎热,冬季寒冷。年均气温7.6 ℃,最高达39.1 ℃, 最低为-27.3 ℃;年均降水量仅117 mm,集中于6—9月,约占年降水量的65%[14 ] 。研究区湿地面积2.10×105 hm2 ,其中自然湿地面积为1.99×105 hm2 ,占湿地面积的94.81%;人工湿地面积1.09×104 hm2 ,占湿地面积的5.19%[14 ] 。湿地类型包括永久性河流、季节性河流、洪泛平原湿地、永久性淡水湖、季节性淡水湖、草本沼泽、高山湿地、灌丛湿地、内陆盐沼、池塘、灌溉渠系、稻田、蓄水区及盐田14个类型[15 ] 。土壤以盐渍化草甸土和盐土为主,水分来源主要依靠河流径流、浅层地下水和农田灌溉渗漏支撑。植物区系受中亚和蒙古植物区系的影响,湿地植被以多年生草本植物为主,伴生乔灌木树种主要有梭梭(Haloxylon ammodendron )、沙枣(Elaeagnus angustifolia )、柽柳(Tamarix chinensis )等,伴生草本植物有小粒苔草(Carex karoi )、小花灯芯草(Juncus articulatus )、芦苇(Phragmites australis )、盐爪爪(Kalidium foliatum )、赖草(Leymus secalinus )和苦豆子(Sophora alopecuroides )等[16 ] 。
图1
图1
荒漠绿洲湿地研究区概况图
Fig.1
Location map of study area in desert oasis wetland
1.2 试验设计
1.2.1 气象测量
在试验期,通过波文比能量平衡法(BREB)测量土壤与大气交界处的水热通量。传感器架两层架设,分别为距地面2 m和3 m。观测项目包括降水量、风向、风速、气温、地温、相对湿度、净辐射、水汽压和地热流等。风速和风向利用三维声波风速计(ATI Electronics Inc.,Boulder,美国)监测,水汽压用红外线气体分析仪(CS105,Vaisala,Helsinki,芬兰),净辐射用辐射传感器(LI190SB,Li-Cor Inc.,Lincoln,NE,美国),土壤热通量用土壤热通量板(model HFT-3,Radiation Energy Balance Systems,美国),降水量用倾斗式雨量计(model TE525,metric;Texas Electronics, Dallas, TX,美国),气温和相对湿度分别用温度传感器(107-L,Campbell Scientific Ltd.,Edmonton,Alberta,加拿大)与湿度传感器(Decagon Devices, Pullman,WA,美国),数据频率为10 Hz,CR1000数据记录仪(Campbell Scientific Inc.,Logan,UT,美国)每5 min记录1次,30 min存储平均数据,其中降水和风速数据每10 min存储均值。
1.2.2 水文测量
在荒漠绿洲湿地试验场,选取100 m×100 m的固定试验区。在试验区内沿垂直于河道方向布设直径5 cm的PVC地下水观测井,所有观测井均安装水位传感器HOBO U24获取地下水位动态变化。利用ECH2 O-TE土壤水热盐观测系统连续监测试验区土壤水分、温度和电导率,其埋深依次为20、40、60、80、100 cm。采用烘干法每10 d对土壤水分数据进行校正。
1.2.3 生物量、含水量及同位素测量
在试验期间,采集降水、植物茎、土壤水、地表径流、地下水等样品。降水水样在气象站安装的存储式雨量计中收集。在生长期,选择树木年龄超过两年且发育状况基本一致的荒漠湿地植物柽柳、沙枣各10株,每月中旬将枝条剪成直径0.5 cm、长5 cm左右,迅速将外皮和韧皮部去掉,保留木质部。所取样品迅速装入玻璃瓶内并盖好瓶塞,每瓶装3—5段。在荒漠绿洲湿地试验场,利用土钻挖取土壤用于提取土壤水和测定土壤含水量,挖取深度直至饱和含水层。在2015—2017年干旱期(2015年6月19日)、极干旱期(2016年3月12—13日)、极湿润期(2017年11月28—29日)和湿润期(2017年1月8日)对不同深度土壤进行取样(0—10、10—20、20—40、40—60 cm,此后每20 cm一层采集至浅层地下水)。同时,利用烘干法校正仪器观测的土壤含水量数据。地下水样取自地下水观测井。试验区内6口观测井均匀分布。河水取自样地附近的地表径流和河流径流。水样采集后装在4 mL玻璃样品瓶中用Parafilm封口膜密封,防止水分蒸发,5个重复。植物茎和土壤样品在实验室冷冻保存,降水、地下水、地表径流等水样冷藏保存。
同位素样品在中国科学院内陆河流域生态水文重点实验室进行测定分析。先用低温真空抽蒸馏提法抽提植物茎秆样品和土壤样品中的水分, 抽提出的水分与降水、河水、地下水等水样分别放在玻璃样品瓶中密封,并低温(2 ℃)保存,然后采用液态水同位素分析仪(DLT-100,Los Gatos Research Inc.,Mountain View,美国)测量不同样品中的δ 18 O值。
δ 18 O = R s a R s t - 1 × 1000 ‰ (1)
式中:R sa 是样品中元素的重轻同位素丰度之比(18 O/16 O);R st 是国际通用标准物稳定同位素丰度之比(H、O稳定同位素采用v-SMOW)。
1.3 分析方法
1.3.1 波文比能量平衡法
波文比能量平衡法是在水热交换的基础上估算实际蒸散量,并且计算感热与潜热之间的对流分量。所以,地表能量平衡方程可以表达为:
R n - G - H - λ E T = 0 (2)
式中:R n 为净辐射(W·m-2 );G 为土壤地热流(W·m-2 );H 为感热通量(W·m-2 );λ -1 );ET 为实际蒸散量(mm)。
波文比是能量分配指标,为感热通量与潜热通量之比。计算波文比、潜热通量以及感热通量的表达式分别为:
β = H λ E T (3)
λ E T = R n - G 1 + β (4)
H = β ( R n - G ) 1 + β (5)
在实际计算中,波文比是用冠层之上两个高度之间的温度和水汽压梯度来计算的:
β = γ ∆ T ∆ e (6)
式中:γ -1 );∆T 和∆e 分别为2 m和3 m的温度差(℃)和水汽压差(kPa)[3 ] 。
E T = 1 λ R n - G 1 + γ ∆ T / ∆ e (7)
1.3.2 分析方法
利用多元线性混合模型(IsoSource)上下限方法计算土壤水、河水、地下水、降水等不同潜在水源对湿地植物的贡献率。考虑到干旱环境下部分植物在吸水过程中氢同位素会发生分馏,而氧同位素不会分馏,采用δ 18 O计算不同水源对植物的水分贡献率更为准确[7 ,17 ] 。多元线性混合模型的计算方法如下:
f 1 δ 18 O 1 + f 2 δ 18 O 2 + f 3 δ 18 O 3 + … + f n δ 18 O n = δ 18 O p (8)
f 1 + f 2 + … + f n = 1 (9)
式中:δ 18 O p δ 18 O1 、δ 18 O2 、…、δ 18 O n n 种水源中氧同位素值;f 1 、 f 2 、 …、 fn 分别为不同水源对植物水的贡献比例。
采用SPSS18.0统计软件进行数据统计与分析,采用Origin软件进行图表的绘制。
2 结果与分析
2.1 水文情况特征
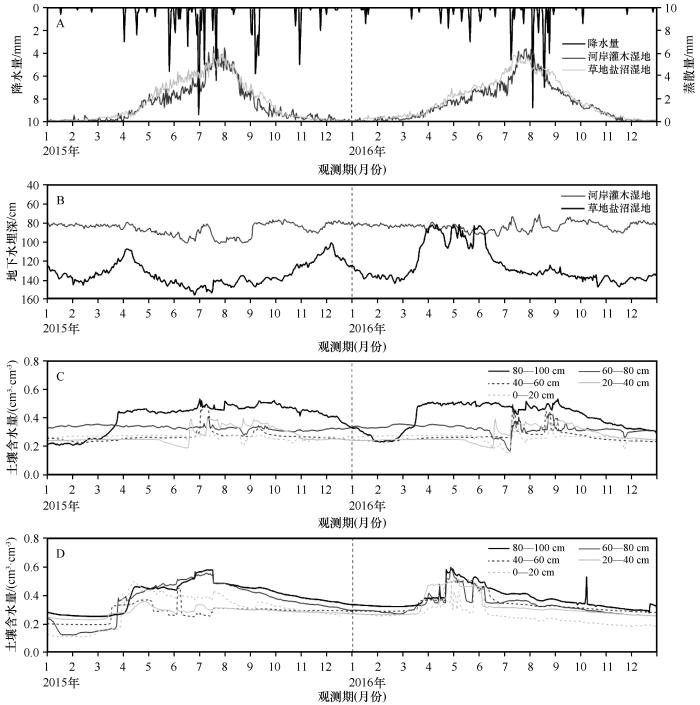
在研究期(2015—2017年)该地区降水有明显的季节差异,降水年内整体呈正态分布趋势,2015年降水明显高于2016年(图2 )。试验区地处荒漠干旱区,降水事件主要发生在生长季(6—8月),非生长季降水极少,且降水量≤5 mm的事件频率较高,小降水事件较多;≥10 mm的事件少,持续时间长且降水量大的事件较少发生,具有明显的季节性分布特征。2015年降水103.2 mm,生长季(6—8月)降水占年降水的69.8%;2016年降水65.4 mm,生长季降水占75.5%。根据卡方检验,生长季占年降水比例的差异不显著(χ 2 =0.66,P =0.42)。降水分布在年际间也略有差异,2015年7月降水量最高,为33 mm;而2016年8月降水量最高,为34.8 mm。
图2
图2
河岸灌木湿地和草地盐沼湿地的蒸散发及2015、2016年降水量(A),地下水埋深(B)和土壤含水量(C和D分别代表灌木和草地生境在不同土壤深度下的土壤含水量)
Fig.2
Evapotranspiration and precipitation of shrub wetland and grassland in 2015 and 2016 (A), groundwater depth (B), C and D represent soil water content of shrub wetland and grassland
与降水数据相似,河岸灌木湿地与草地盐沼湿地蒸散量整体也呈正态分布,年均蒸散量为604.47 mm。在生长季,蒸散量呈单峰变化,而在冻融期土壤冻结,蒸散量接近于0(图2 ),但不同湿地的蒸散发特征具有比较明显差异。在1—3月,河岸灌木湿地与草地盐沼湿地的蒸散量均在0—1 mm;自4月起,蒸散量开始快速增加,草地盐沼湿地增长尤为迅猛,5—7月均高于河岸灌木湿地蒸散量,最高可达6.01 mm·d-1 。7月下旬至8月初灌木湿地蒸散量达到峰值,最高可达6.61 mm·d-1 。自9月后,温度降低导致水汽压降低的影响,湿地蒸散发均急剧下降,直至12月趋于0 mm。
河岸灌木湿地与草地盐沼湿地的地下水波动程度截然不同(图2 )。在离河道距离较近的灌木湿地,地下水深度随季节波动较小,地下水全年埋深70—110 cm,年均85.8±9 mm;而在草地盐沼湿地地下水深度波动较大,为80—160 cm,年均埋深为130.1±12.2 mm。在草地盐沼湿地中,地下水位呈现先升后减再升的趋势,从3、4月开始上升,夏季后逐渐下降。
除80—100 cm土层的含水量波动较大外,河岸灌木湿地含水量较为稳定(图2 ),原因是河水侧渗长期补充地下水位,并且维持水位在80 cm左右波动。受农田灌溉退水的影响,草地盐沼湿地土壤含水量波动较大且具有季节性变化,不同深度的土层含水量变化趋势相似,即在春季开始升高并在夏季后逐步下降。
2.2 降水稳定同位素特征
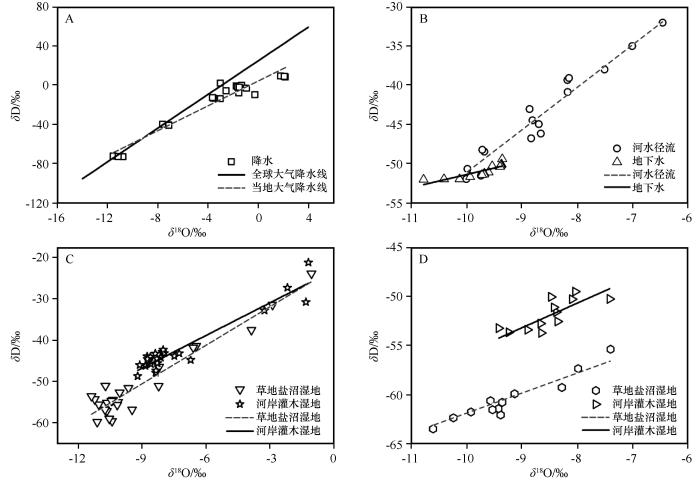
黑河中游,降水量极少且分布不均,年降水量的90%发生在生长期。在研究区,降水中δ D和δ 18 O存在较大波动,具有明显的季节变化规律。δ D值 -73.21‰—9.34‰,平均-18.21‰±26.92‰,变异系数-147.8%;δ 18 O值-11.54‰—2.17‰,平均-3.52‰±4.14‰,变异系数-117.67%(表1 )。生长季前期与生长季后期δ 18 O值较高,生长旺盛期δ 18 O处于较低水平。当地大气降水线(δ D=6.33δ 18 O+4.04,R 2 =0.931)斜率和截距均略小于全球大气降水线,但地表径流与降水的截距和斜率较为接近,而地下水与降水和径流的拟合曲线差异较大(图3 ,表2 )。降水δ D和δ 18 O波动范围最大,其最大值极显著地高于径流、地下水和植物水(表1 )。同时,径流和地下水与植物水的均值差异较大。
图3
图3
全球大气降水线和当地大气降水线(A)、径流和地下水(B)、土壤水(C)和植物木质部水(D)与δ D与δ 18 O的回归线性关系
Fig.3
Linear relationship between global and local precipitation lines (A), runoff and groundwater (B), soil water (C) and plant xylem water (D) and δ D and δ 18 O
2.3 黑河径流、地下水、土壤水和植物水同位素特征
试验期间,黑河径流δ D和δ 18 O值分别为-50.71‰—-32.00‰和-9.98‰—-6.45‰,均值分别为-43.80‰±12.09‰和-8.65‰±23.33‰(表1 ),变异系数分别为 -27.61%和-26.97%,中等变异,拟合方程为δ D=5.44δ 18 O+3.23(R 2 =0.91,表2 )。地下水δ D和δ 18 O值分别为 -52.06‰—-49.44‰和-10.78‰—-9.35‰,均值分别为 -50.98‰±13.18‰和-9.74‰±25.49‰,变异系数分别为-25.87%和-26.17%,中等变异,拟合方程为δ D=1.66δ 18 O-34.80 (R 2 =0.70,图3 )。地下水与径流同位素部分重叠且差异较大,表明部分地下水来自于河水。
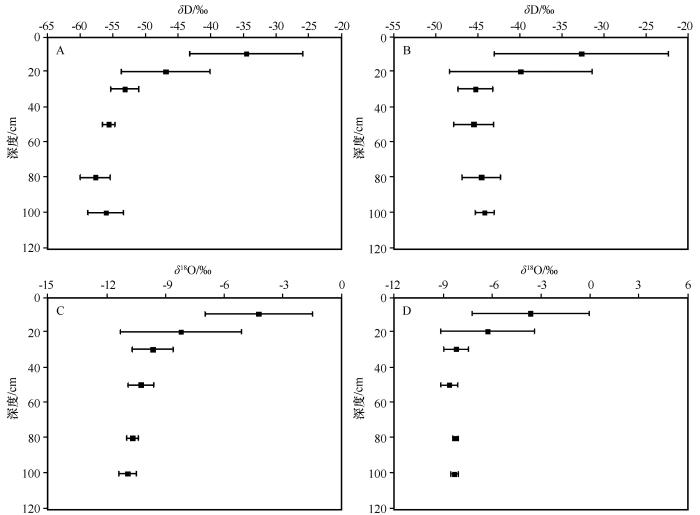
在荒漠绿洲湿地,河岸灌木湿地土壤水δ D与δ 18 O值略高,分别为-48.74‰—-28.11‰和-9.22‰—-1.18‰,均值分别为-42.07‰±6.89‰和-7.22‰±2.49‰(表1 ),变异系数分别为25.63%和39.17%,拟合曲线为δ D=2.64δ 18 O-23.02(R 2 =0.90,表2 );然而,草地盐沼湿地的δ D与δ 18 O值跨度较大,分别为 -59.90‰—-23.81‰和-11.38‰—-1.04‰,均值分别为-50.72‰±13.57‰和-9.00‰±3.30‰(表1 ),变异系数分别为-26.76%和-36.72%,拟合曲线为δ D=3.12δ 18 O-22.64(R 2 =0.92,表2 )。土壤水分布于降水线右下方,表明土壤水δ D与δ 18 O经历了不同程度的二次蒸发分馏效应使得δ D与δ 18 O发生富集作用(图3 )。在垂直方向上,土壤水稳定同位素值存在明显差异(P <0.05)。随土层增加,土壤水δ D和δ 18 O值有依次降低的变化趋势(图4 )。表层土壤水δ D和δ 18 O值与降水有少量重合,表明表层土壤受降水的补给。0—20 cm土层δ D和δ 18 O值变化最大,向下变幅逐渐减少,表明强烈蒸发是表层土壤水δ 18 O富集的主要原因。土层30 cm以下,土壤水δ 18 O和δ D值变化较小,土壤水δ D和δ 18 O值接近河水与地下水,表明降水对土壤水的贡献很小。同时,当深度大于30 cm之后,河岸灌木湿地地下水的δ 18 O和δ D均小于黑河径流,表明地下水的贡献较小。
图4
图4
草地盐沼湿地(A,C)和河岸灌木湿地(B,D)生境不同深度土壤水的δ D(A,B)和δ 18 O(C,D)含量
Fig.4
δ D (A,B) and δ 18 O (C,D) values of grassland wetland (A,C) and shrub wetland (B,D) at different depths
在河岸灌木湿地与草地盐沼湿地,植物水同位素存在显著差异(P <0.05),这表明两种植物生长所需的水分来源不同(图3 )。河岸灌木林植物水δ D和δ 18 O值分别为-53.72‰—-49.49‰和-9.42‰—-7.41‰,均值分别为-51.84‰±14.46‰和-8.50‰±24.13‰(表1 ),变异系数分别为-27.89%和-28.39%,拟合曲线为δ D=2.52δ 18 O-30.54 (R 2 =0.61,表2 );草地盐沼湿地植物水的δ D与δ 18 O值跨度较大,分别为-63.56‰—-55.41‰和-10.61‰—-7.40‰,均值分别为-60.55‰±16.94‰和-9.24‰±27.13‰(表1 ),变异系数分别为-27.97%和-29.36%,拟合曲线为δ D=2.04δ 18 O-41.52(R 2 =0.71,表2 )。
2.4 土壤水及植物水与各水源影响因子的关系
基于同位素示踪信息对土壤水及植物水与不同水分来源的相关关系进行分析(表3 )。荒漠绿洲湿地不同降水事件对土壤水和植物水δ D的贡献率有差异,但未达到显著水平。土壤水及土壤水与地下水的交互作用显著地影响着河岸灌木湿地、草地盐沼湿地土壤水和植物水δ D(P <0.001),在草地盐沼湿地,地下水对土壤水与植物水δ D的影响存在显著性差异(P <0.01)。此外,降水、地下水和土壤水的交互作用也显著地影响着盐沼草地湿地土壤水和植物水δ D (P <0.001)。
2.5 水分来源对植物水分利用的贡献
多元线性混合模型表明河岸灌木湿地与草地盐沼湿地水分来源存在显著差异(P <0.05,图5 )。在河岸灌木湿地,水分主要来源于河水,约占50%;其次是地下水,占37%,而降水仅占13%。然而,离河较远的草地盐沼湿地水分主要来源于地下水,达67%;其次是河水径流,约占29%,而降水只占到水分来源的4%。
图5
图5
河岸灌木湿地与盐沼草地湿地各潜在水源对植物水分利用的贡献率
Fig.5
Contribution rate of water resources of shrub wetland and grassland wetland
2.6 不同土层根系生物量
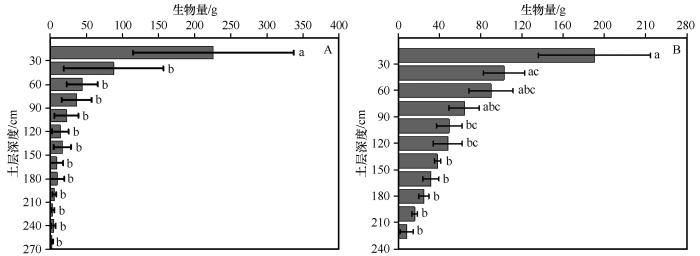
在荒漠绿洲湿地,河岸灌木湿地优势种柽柳和草地盐沼湿地优势种沙枣根系生物量在不同土层深度具有显著差异性(图6 )。柽柳浅层根系发达,细根主要分布在土层0—60 cm (57.69%),0—20 cm细根生物量225.4±125.6 g,约占土层内总细根生物量的28.67%,其次20—40 cm和40—60 cm土层则分别占15.47%和13.55%;在60—80、80—100、100—120、120—140、140—160、160—180、180—200、200—220 cm各层细根生物量占总细根生物量的百分比分别为9.64%、7.46%、7.24%、5.76%、4.80%、3.77%、2.42%、1.23%。沙枣细根主要分布在0—40 cm (64.83%),0—20 cm细根生物量186.5±65.5 g,约占总细根生物量的46.66%,20—40 cm占18.17%;随土层加深,沙枣细根逐渐减少,在40—60、60—80、80—100、100—120、120—140、140—160、160—180、180—200、200—220、220—240、240—260 cm各层细根生物量占总细根生物量的百分比分别为9.19%、7.61%、4.68%、2.93%、3.48%、1.82%、2.05%、1.20%、0.75%、0.99%、0.48%。
图6
图6
河岸灌木湿地优势种柽柳(A)和草地盐沼湿地优势种沙枣(B)根系生物量分布图
Fig.6
Root biomass of dominant species at different depths in shrub wetland (A) and grassland wetland (B)
3 讨论
3.1 湿地水分来源
水分来源及其植物水分利用策略是决定湿地植物区系组成、种群分布格局的关键因子[4 -5 ] ,导致荒漠绿洲湿地植被演替往往朝着不确定性和复杂性方向发展。在干旱区,降水量稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[5 ] 。因此,确定荒漠绿洲湿地水分来源以及植物水分利用方式是干旱区受损湿地植被保护与恢复急需解决的关键问题[15 -16 ] 。在干旱区,小降水(<5 mm)是降水事件的主体,而大降水事件的发生相对较少。本地大气降水线斜率和截距均小于全球大气降水线,而河水、地下水、土壤水都位于大气降水线附近,说明降水对各水源均有补给作用[17 ] ,但并不显著。黑河水主要来源于山区冰雪融水,对地下水补给量大,所以δ 18 O值位于大气降水线左上方;干旱区蒸发强烈,造成土壤水δ 18 O的富集,所以位于大气降水线下方[17 -18 ] 。在干旱区,水分输出量远大于降水量,小降水事件的雨滴在降落过程中受云下二次蒸发的影响而发生强烈的同位素动力分馏效应,导致降水中δ D和δ 18 O存在较大波动[19 ] 。然而,土壤水分入渗几乎不影响同位素分馏。因此,地表蒸发是导致荒漠绿洲湿地土壤氢、氧同位素富集的主要原因[20 -21 ] 。
在荒漠绿洲湿地,通常受植被类型、植物生活型、离河距离、地下水埋深以及地貌特征的影响,水分来源和植物水分利用方式表现出显著差异性[22 ] 。研究区处于非季风区,属典型的大陆性干旱气候,在降水量小、极端降水频率大及蒸发强烈等因素影响下,降水不能作为荒漠绿洲湿地植物生长的稳定水源[23 -24 ] ,这与我们研究的结论基本一致。通常,干旱地区植物水分利用方式普遍符合两层水分利用模式假设,即深根系木本植物如乔木或灌木利用稳定的深层水源(地下水、深层土壤水),而浅根系多年生或一年生草本植物则依靠表层不稳定的水源(表层土壤水、降水)[25 ] 。研究表明,盐沼草地湿地浅层土壤的δ D、δ 18 O均高于河岸灌木湿地,表明地下水是盐沼草本湿地水分蒸散的主要来源,而受河流补给较少;此外河水是河岸灌木湿地的主要水分来源,说明在荒漠绿洲湿地水分来源对植物利用的贡献差异关键是由离河道距离所决定的。
3.2 土壤剖面水分补给特征
通常,土壤水δ 18 O和δ D值主要受蒸发、灌溉、地下水、降水和土壤质地等因素的影响。在干旱区,降水主要以脉动的方式输入,从而引发受降水和蒸发作用控制的表层土壤水稳定同位素组成呈脉动式变化[26 -28 ] 。在干旱区,小降水事件只能引起表层土壤水分变化,而大降水事件虽然发生频率较低,但能够有效补给深层土壤水分[29 ] ,降水量越大,降水向深层土壤渗透越快[27 ,30 ] 。降水入渗后与土壤原有水分混合构成新的土壤水,其稳定同位素值随着时间的推移不断发生着变化[31 ] 。降水事件发生后土壤水δ 18 O值的变化与降水的δ 18 O值和降雨量的大小密切相关[32 ] ,如8 mm降水入渗后荒漠绿洲湿地δ 18 O值的变化幅度相对较小,而24.8 mm降水能显著改变土壤水的δ 18 O值,即降水量与土壤质地共同影响着降水入渗量和入渗深度[29 ] 。在荒漠绿洲湿地,受蒸发作用影响重同位素富集[33 ] ,导致浅层土壤水的δ D和δ 18 O值较大,甚至在20 cm深度左右土壤水都出现了δ 18 O最大值,而在20 cm深度以下土壤水δ 18 O随深度增加而逐渐减小。在接近饱和地下水位之前,河岸灌木湿地与草地盐沼湿地土壤水的δ 18 O值都接近地下水的δ 18 O值,表明土壤剖面在上界面受蒸发作用而存在强烈的同位素分馏,而下界面则接受地下水的补给。
与深层土壤相比,0—30 cm浅层土壤水的δ D和δ 18 O值变化更复杂[34 ] 。受强烈蒸发的影响,0—10 cm表层土壤水δ D和δ 18 O富集,且随着土层深度的增加,蒸发作用逐渐减弱,δ D和δ 18 O趋于贫化,即浅层土壤水的δ D和δ 18 O值较大,深层土壤水的δ D和δ 18 O值较小。然而,荒漠绿洲湿地土壤水δ D和δ 18 O的最小值均出现在30 cm土层附近,这是由于该土层的黏粒含量最高且土壤持水性好[35 -36 ] ,土壤水氢氧同位素不易被蒸发而产生富集,故其稳定同位素值小[37 ] 。然而,随着灌溉和降水事件的发生,土壤δ D和δ 18 O值逐渐减小。而大于30 cm的深层土壤水主要受地下水稳定补给的影响,其氢氧稳定同位素值比较平稳。因此,土壤水δ D和δ 18 O值的时空变化表现了土壤蒸发、灌溉水及降水入渗、地下水补给等因素对土壤水的影响。
3.3 湿地植物水分利用策略
植物水分利用方式与植物种类型、根系分布、水分条件和生长阶段等有关,但有研究表明不同量级的降水对不同生境优势种的植物水贡献率也有影响[38 ] ,小降水事件对表层(0—20 cm)根系丰富树种植物水的贡献率较高,而较大降水对根系分布较深的植物水的贡献率更高[31 ] ,不同量级降水对各优势树种植物水的贡献率表明深根植物和浅根植物具有不同的水分利用策略[39 ] 。根系主要通过根尖的根毛区来吸收水分,植物根系深度和毛细根分布是决定植物吸收水源的重要因素,所以植物细根分布对植物水分利用策略有很大影响[40 ] 。本研究发现,荒漠绿洲湿地河岸灌木湿地优势种柽柳浅层根系发达,细根主要分布在40 cm深度内的浅层土壤,随土层加深,柽柳细根逐渐减少。草地盐沼湿地优势种沙枣虽浅层侧根发育较少但主根发达[33 ] 。褚建民[34 ] 的研究也表明沙枣是深根植物,能够直接利用浅层地下水,与本研究结果基本相符。
潜在水源对植物水分利用策略至关重要[41 ] 。在荒漠绿洲湿地,不同降水事件对土壤水和植物水δ D的贡献率虽有差异,但未达到显著水平(表3 )。同时,土壤水及土壤水与地下水的交互作用显著地影响着河岸灌木湿地、盐沼草地湿地土壤水和植物水δ D(P <0.001),说明土壤水是维持荒漠绿洲湿地植物生长的决定性因素。在草地盐沼湿地中,地下水对土壤水与植物水δ D的影响存在显著性差异(P <0.01),表明地下水是离河较远的草地湿地土壤与植物水最主要水分来源。此外,降水、地下水和土壤水的交互作用也显著地影响着盐沼草地湿地土壤水和植物水δ D (P <0.001)。由此可见,在河岸灌木湿地、盐沼草地,深根植物和浅根植物具有不同的水分利用策略[42 ] ,地下水对荒漠绿洲湿地植物水的贡献率随着距离河道距离而减小。
4 结论
水分来源及其植物水分利用策略是决定湿地植物区系组成、种群分布格局的关键因子,已成为干旱区受损湿地植被保护与恢复急需解决的关键问题。因此,基于稳定同位素技术研究降水、径流、地下水、土壤水和植物水的同位素组成,探究不同水分来源对荒漠绿洲湿地植物水分利用的贡献率。
黑河流域当地大气降水线为δ D=6.33δ 18 O+4.04 (R 2 =0.931),当地大气降水线斜率和截距均小于全球大气降水线,而河水、地下水、土壤水都位于大气降水线附近,说明降水对各水源均有补给作用,但并不显著。
地表蒸发是导致荒漠绿洲湿地土壤氢、氧同位素富集的主要原因。地下水和河水是盐沼草本湿地与河岸灌木湿地的主要水分来源,说明在水分来源对植物水分利用的贡献差异是由河道距离所决定的。在干旱区,受降水和蒸发作用控制的表层土壤水稳定同位素组成呈脉动式变化,土壤剖面在上界面受蒸发作用而存在强烈的同位素分馏,而下界面则接受地下水的补给。
参考文献
View Option
[1]
章光新 ,武瑶 ,吴燕锋 ,等 湿地生态水文学研究综述
[J].水科学进展 ,2018 ,29 (5 ):737 -749 .
[本文引用: 1]
[2]
梅荣 干旱区绿洲湿地生态系统研究的意义和重点
[J].内蒙古科技与经济 ,2007 ,139 (9 ):115 -116 .
[本文引用: 1]
[3]
刘冰 ,常学向 ,李守波 荒漠区土壤水分对降水脉动响应
[J].中国沙漠 ,2011 ,31 (3 ):716 -723 .
[本文引用: 2]
[4]
Liu B Zhao W Z Wen Z J et al Response of water and energy exchange to the environmental variable in a desert-oasis wetland of Northwest China
[J].Hydrological Processes ,2014 ,28 (25 ):6098 -6112 .
[本文引用: 3]
[5]
Liu B Zhao W Z Wen Z J et al Hydrochemical characteristics jointly determine the transport and cycling of soil carbon,nitrogen,and phosphorus in an arid Chinese wetland
[J].Journal of Geophysical Research: Biogeosciences ,2020 ,125 (7 ):1 -21 .
[本文引用: 4]
[6]
张飞 气候和人类活动影响的湿地植被高光谱研究现状与展望
[J].新疆大学学报(自然科学版) ,2019 ,36 (1 ):25 -33 ,48 .
[本文引用: 1]
[7]
许秀丽 ,李云良 ,谭志强 ,等 鄱阳湖湿地典型中生植物水分利用来源的同位素示踪
[J].湖泊科学 ,2020 ,32 (6 ):1749 -1760 .
[本文引用: 2]
[8]
Patrick Z E David G W Hydrogen isotope fractionation during water uptake by woody xerophytes
[J].Plant and Soil ,2007 ,291 (1/2 ):93 -107 .
[本文引用: 1]
[9]
李宁 ,周海 ,任珩 ,等 不同地下水位处梭梭(Haloxylon ammodendron )水分来源特征
[J].中国沙漠 ,2021 ,41 (4 ):79 -86 .
[10]
周海 河西走廊绿洲边缘典型荒漠灌木根系分布特征及其水分利用策略
[D].北京 :中国科学院大学 ,2016 .
[本文引用: 1]
[11]
徐庆 ,刘世荣 ,安树青 ,等 川西亚高山暗针叶林降水分配过程中氧稳定同位素特征
[J].植物生态学报 ,2006 (1 ):83 -89 .
[本文引用: 1]
[12]
赵国琴 ,李小雁 ,吴华武 ,等 青海湖流域具鳞水柏枝植物水分利用氢同位素示踪研究
[J].植物生态学报 ,2013 ,37 (12 ):1091 -1100 .
[本文引用: 1]
[13]
赵良菊 ,肖洪浪 ,程国栋 ,等 黑河下游河岸林植物水分来源初步研究
[J].地球学报 ,2008 ,29 (6 ):709 -718 .
[本文引用: 1]
[14]
庄艳丽 ,赵文智 临泽荒漠绿洲湿地植物生态系列及其物种多样性研究
[J].湿地科学 ,2014 ,12 (4 ):477 -484 .
[本文引用: 4]
[15]
张锋 ,李自珍 ,惠苍 中国湿地物种多样性与生境面积关系及其生态学机理的模拟研究
[J].西北植物学报 ,2004 (3 ):392 -396 .
[本文引用: 2]
[16]
李小燕 黑河流域湿地典型植物群落的物种多样性
[J].草业科学 ,2012 ,29 (9 ):1335 -1339 .
[本文引用: 2]
[17]
Donald L P Jillian W G Source partitioning using stable isotopes: coping with too many sources
[J].Oecologia ,2003 ,136 (2 ):261 -269 .
[本文引用: 3]
[18]
Li D F Gao G Y Lü Y H et al Multi-scale variability of soil carbon and nitrogen in the middle reaches of the Heihe River basin, northwestern China
[J].Catena ,2016 ,137 :328 -339 .
[本文引用: 1]
[19]
孟阳阳 ,刘冰 ,刘婵 荒漠绿洲湿地土壤水热盐动态过程及其影响机制
[J].中国沙漠 ,2019 ,39 (1 ):149 -160 .
[本文引用: 1]
[20]
Zhao L J Xiao H L Zhou M X et al Factors controlling spatial and seasonal distributions of precipitation δ 18 O in China
[J].Hydrological Processes ,2012 ,26 (1 ):143 -152 .
[本文引用: 1]
[21]
孟玉川 ,刘国东 长江流域降水稳定同位素的云下二次蒸发效应
[J].水科学进展 ,2010 ,21 (3 ):327 -334 .
[本文引用: 1]
[22]
李海涛 白皮沙拐枣(Calligonum leucocladum )根系与环境关系的初步研究
[J].新疆农业大学学报 ,1996 ,19 (1 ):56-61,72 -72 .
[本文引用: 1]
[23]
陈亚宁 ,李卫红 ,陈亚鹏 ,等 新疆塔里木河下游断流河道输水与生态恢复
[J].生态学报 ,2007 (2 ):538 -545 .
[本文引用: 1]
[24]
王希义 ,徐海量 ,潘存德 ,等 塔里木河下游优势草本植物与地下水埋深的关系
[J].中国沙漠 ,2016 ,36 (1 ):216 -224 .
[本文引用: 1]
[25]
罗维成 ,曾凡江 ,刘波 ,等 疏叶骆驼刺母株与子株间的水分整合
[J].植物生态学报 ,2013 ,37 (2 ):164 -172 .
[本文引用: 1]
[26]
周海 ,郑新军 ,唐立松 ,等 准噶尔盆地东南缘多枝柽柳、白刺和红砂水分来源的异同
[J].植物生态学报 ,2013 ,37 (7 ):665 -673 .
[本文引用: 1]
[27]
马迎宾 ,张蓓蓓 ,徐庆 ,等 绍兴淡水湿地人工林优势树种水分利用策略
[J].林业科学 ,2019 ,55 (12 ):140 -150 .
[本文引用: 1]
[28]
巩国丽 ,陈辉 ,段德玉 利用稳定氢氧同位素定量区分白刺水分来源的方法比较
[J].生态学报 ,2011 ,31 (24 ):7533 -7541 .
[本文引用: 1]
[29]
Ohte N Koba K Yoshikawa K et al Water utilization of natural and planted trees in the semiarid desert of Inner Mongolia,China
[J].Ecological Applications ,2003 ,13 (2 ):337 -351 .
[本文引用: 2]
[30]
Saha A K Sternberg L D S O Ross M S et al Water source utilization and foliar nutrient status differs between upland and flooded plant communities in wetland tree islands
[J].Wetlands Ecology & Management ,2010 ,18 (3 ):343 -355 .
[本文引用: 1]
[31]
赵文智 ,吉喜斌 ,刘鹄 蒸散发观测研究进展及绿洲蒸散研究展望
[J].干旱区研究 ,2011 ,28 (3 ):463 -470 .
[本文引用: 2]
[32]
林光辉 稳定同位素生态学 [M].北京 :高等教育出版社 ,2013 :18 -19 .
[本文引用: 1]
[33]
朱建佳 ,陈辉 ,邢星 ,等 柴达木盆地荒漠植物水分来源定量研究:以格尔木样区为例
[J].地理研究 ,2015 ,34 (2 ):285 -292 .
[本文引用: 2]
[34]
褚建民 干旱区植物的水分选择性利用研究
[D].北京 :中国林业科学研究院 ,2007 .
[本文引用: 2]
[35]
刘树宝 ,陈亚宁 ,李卫红 ,等 黑河下游不同林龄胡杨水分来源的D、δ 18 O同位素示踪
[J].干旱区地理 ,2014 ,37 (5 ):988 -995 .
[本文引用: 1]
[36]
孟阳阳 ,刘冰 ,刘婵 水盐梯度下湿地柽柳(Tamarix ramosissima )光合响应特征和水分利用效率
[J].中国沙漠 ,2018 ,38 (3 ):568 -577 .
[本文引用: 1]
[37]
李亚飞 ,于静洁 ,陆凯 ,等 额济纳三角洲胡杨和多枝柽柳水分来源解析
[J].植物生态学报 ,2017 ,41 (5 ):519 -528 .
[本文引用: 1]
[38]
林欢 ,许秀丽 ,张奇 鄱阳湖典型洲滩湿地水分补排关系
[J].湖泊科学 ,2017 ,29 (1 ):160 -175 .
[本文引用: 1]
[39]
尹力 ,赵良菊 ,阮云峰 ,等 黑河下游典型生态系统水分补给源及优势植物水分来源研究
[J].冰川冻土 ,2012 ,34 (6 ):1478 -1486 .
[本文引用: 1]
[40]
Zhang X Xiao Y Wan H et al Using stable hydrogen and oxygen isotopes to study water movement in soil-plant-atmosphere continuum at Poyang Lake wetland,China
[J].Wetlands Ecology and Management ,2017 ,25 (2 ):1 -14 .
[本文引用: 1]
[41]
段德玉 ,欧阳华 稳定氢氧同位素在定量区分植物水分利用来源中的应用
[J].生态环境 ,2007 (2 ):655 -660 .
[本文引用: 1]
[42]
余绍文 ,孙自永 ,周爱国 ,等 用D、δ 18 O同位素确定黑河中游戈壁地区植物水分来源
[J].中国沙漠 ,2012 ,32 (3 ):717 -723 .
[本文引用: 1]
湿地生态水文学研究综述
1
2018
... 在干旱区,湿地作为荒漠绿洲核心地理单元,不仅在涵养水源、净化水质、蓄洪抗旱、调节气候和维护生物多样性等方面发挥着重要作用[1 ] ,而且对河岸林锐减、绿洲萎缩、草场退化和荒漠化等具有抑制功能,在维持区域生态平衡与保障内陆河流域可持续发展方面具有生态屏障作用[2 ] .气候变化通过对气温、降水以及干旱洪涝极端水文事件发生频率和强度的影响,改变着湿地蒸散发、径流、水位、水文周期等关键水文过程[3 ] ,调控着湿地水分来源、补给途径及其植物水分利用策略,对荒漠绿洲湿地植物区系组成、种群格局和稳定性产生深远影响[4 ] .与此同时,农田开垦、灌溉引水等人类活动通过改变植被覆盖和径流过程间接或直接影响着湿地水文过程,加剧了水资源短缺和地下水枯竭,使荒漠绿洲湿地出现面积萎缩、植被退化、多样性减少、盐碱化加剧等生态环境问题[5 ] .所以,人类活动与气候变化共同作用是荒漠绿洲湿地环境恶化的主要驱动力,定量研究荒漠绿洲湿地水分来源、补给途径及其植物水分利用策略,是解决荒漠绿洲湿地环境恶化问题的前提条件,将为干旱区湿地环境治理和生态安全提供理论依据与决策支持. ...
干旱区绿洲湿地生态系统研究的意义和重点
1
2007
... 在干旱区,湿地作为荒漠绿洲核心地理单元,不仅在涵养水源、净化水质、蓄洪抗旱、调节气候和维护生物多样性等方面发挥着重要作用[1 ] ,而且对河岸林锐减、绿洲萎缩、草场退化和荒漠化等具有抑制功能,在维持区域生态平衡与保障内陆河流域可持续发展方面具有生态屏障作用[2 ] .气候变化通过对气温、降水以及干旱洪涝极端水文事件发生频率和强度的影响,改变着湿地蒸散发、径流、水位、水文周期等关键水文过程[3 ] ,调控着湿地水分来源、补给途径及其植物水分利用策略,对荒漠绿洲湿地植物区系组成、种群格局和稳定性产生深远影响[4 ] .与此同时,农田开垦、灌溉引水等人类活动通过改变植被覆盖和径流过程间接或直接影响着湿地水文过程,加剧了水资源短缺和地下水枯竭,使荒漠绿洲湿地出现面积萎缩、植被退化、多样性减少、盐碱化加剧等生态环境问题[5 ] .所以,人类活动与气候变化共同作用是荒漠绿洲湿地环境恶化的主要驱动力,定量研究荒漠绿洲湿地水分来源、补给途径及其植物水分利用策略,是解决荒漠绿洲湿地环境恶化问题的前提条件,将为干旱区湿地环境治理和生态安全提供理论依据与决策支持. ...
荒漠区土壤水分对降水脉动响应
2
2011
... 在干旱区,湿地作为荒漠绿洲核心地理单元,不仅在涵养水源、净化水质、蓄洪抗旱、调节气候和维护生物多样性等方面发挥着重要作用[1 ] ,而且对河岸林锐减、绿洲萎缩、草场退化和荒漠化等具有抑制功能,在维持区域生态平衡与保障内陆河流域可持续发展方面具有生态屏障作用[2 ] .气候变化通过对气温、降水以及干旱洪涝极端水文事件发生频率和强度的影响,改变着湿地蒸散发、径流、水位、水文周期等关键水文过程[3 ] ,调控着湿地水分来源、补给途径及其植物水分利用策略,对荒漠绿洲湿地植物区系组成、种群格局和稳定性产生深远影响[4 ] .与此同时,农田开垦、灌溉引水等人类活动通过改变植被覆盖和径流过程间接或直接影响着湿地水文过程,加剧了水资源短缺和地下水枯竭,使荒漠绿洲湿地出现面积萎缩、植被退化、多样性减少、盐碱化加剧等生态环境问题[5 ] .所以,人类活动与气候变化共同作用是荒漠绿洲湿地环境恶化的主要驱动力,定量研究荒漠绿洲湿地水分来源、补给途径及其植物水分利用策略,是解决荒漠绿洲湿地环境恶化问题的前提条件,将为干旱区湿地环境治理和生态安全提供理论依据与决策支持. ...
... 式中:γ -1 );∆T 和∆e 分别为2 m和3 m的温度差(℃)和水汽压差(kPa)[3 ] . ...
Response of water and energy exchange to the environmental variable in a desert-oasis wetland of Northwest China
3
2014
... 在干旱区,湿地作为荒漠绿洲核心地理单元,不仅在涵养水源、净化水质、蓄洪抗旱、调节气候和维护生物多样性等方面发挥着重要作用[1 ] ,而且对河岸林锐减、绿洲萎缩、草场退化和荒漠化等具有抑制功能,在维持区域生态平衡与保障内陆河流域可持续发展方面具有生态屏障作用[2 ] .气候变化通过对气温、降水以及干旱洪涝极端水文事件发生频率和强度的影响,改变着湿地蒸散发、径流、水位、水文周期等关键水文过程[3 ] ,调控着湿地水分来源、补给途径及其植物水分利用策略,对荒漠绿洲湿地植物区系组成、种群格局和稳定性产生深远影响[4 ] .与此同时,农田开垦、灌溉引水等人类活动通过改变植被覆盖和径流过程间接或直接影响着湿地水文过程,加剧了水资源短缺和地下水枯竭,使荒漠绿洲湿地出现面积萎缩、植被退化、多样性减少、盐碱化加剧等生态环境问题[5 ] .所以,人类活动与气候变化共同作用是荒漠绿洲湿地环境恶化的主要驱动力,定量研究荒漠绿洲湿地水分来源、补给途径及其植物水分利用策略,是解决荒漠绿洲湿地环境恶化问题的前提条件,将为干旱区湿地环境治理和生态安全提供理论依据与决策支持. ...
... 干旱区降水稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[4 -5 ] .通常,受植被类型、水分状况、河流径流以及地下水位的影响,植物水分来源与利用方式的辨析与识别具有高度的变异性和复杂性[6 ] .研究表明,除极少数盐生植物外,陆地植物根系吸收水分在木质部运输过程中一般不发生同位素分馏,因此植物茎木质部提取的氢氧稳定同位素比率可反映植物的水分来源[7 ] ,为荒漠绿洲湿地水分来源的辨析提供了水文学依据.目前,国内学者基于稳定同位素技术在荒漠、河岸、海岸等系统中乔灌草等不同生活型植物水分来源和利用策略研究中开展了广泛应用,包括干旱半干旱区[8 -10 ] 、川西亚高山区[11 ] 、青海湖流域[12 ] 和黑河流域[13 -14 ] 等,为探明湿地水分来源、补给途径及其植物水分利用策略提供了思路与技术.然而,基于稳定同位素技术研究干旱区湿地水分来源、补给途径及其植物水分利用策略的研究迄今为止仍未见报道. ...
... 水分来源及其植物水分利用策略是决定湿地植物区系组成、种群分布格局的关键因子[4 -5 ] ,导致荒漠绿洲湿地植被演替往往朝着不确定性和复杂性方向发展.在干旱区,降水量稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[5 ] .因此,确定荒漠绿洲湿地水分来源以及植物水分利用方式是干旱区受损湿地植被保护与恢复急需解决的关键问题[15 -16 ] .在干旱区,小降水(<5 mm)是降水事件的主体,而大降水事件的发生相对较少.本地大气降水线斜率和截距均小于全球大气降水线,而河水、地下水、土壤水都位于大气降水线附近,说明降水对各水源均有补给作用[17 ] ,但并不显著.黑河水主要来源于山区冰雪融水,对地下水补给量大,所以δ 18 O值位于大气降水线左上方;干旱区蒸发强烈,造成土壤水δ 18 O的富集,所以位于大气降水线下方[17 -18 ] .在干旱区,水分输出量远大于降水量,小降水事件的雨滴在降落过程中受云下二次蒸发的影响而发生强烈的同位素动力分馏效应,导致降水中δ D和δ 18 O存在较大波动[19 ] .然而,土壤水分入渗几乎不影响同位素分馏.因此,地表蒸发是导致荒漠绿洲湿地土壤氢、氧同位素富集的主要原因[20 -21 ] . ...
Hydrochemical characteristics jointly determine the transport and cycling of soil carbon,nitrogen,and phosphorus in an arid Chinese wetland
4
2020
... 在干旱区,湿地作为荒漠绿洲核心地理单元,不仅在涵养水源、净化水质、蓄洪抗旱、调节气候和维护生物多样性等方面发挥着重要作用[1 ] ,而且对河岸林锐减、绿洲萎缩、草场退化和荒漠化等具有抑制功能,在维持区域生态平衡与保障内陆河流域可持续发展方面具有生态屏障作用[2 ] .气候变化通过对气温、降水以及干旱洪涝极端水文事件发生频率和强度的影响,改变着湿地蒸散发、径流、水位、水文周期等关键水文过程[3 ] ,调控着湿地水分来源、补给途径及其植物水分利用策略,对荒漠绿洲湿地植物区系组成、种群格局和稳定性产生深远影响[4 ] .与此同时,农田开垦、灌溉引水等人类活动通过改变植被覆盖和径流过程间接或直接影响着湿地水文过程,加剧了水资源短缺和地下水枯竭,使荒漠绿洲湿地出现面积萎缩、植被退化、多样性减少、盐碱化加剧等生态环境问题[5 ] .所以,人类活动与气候变化共同作用是荒漠绿洲湿地环境恶化的主要驱动力,定量研究荒漠绿洲湿地水分来源、补给途径及其植物水分利用策略,是解决荒漠绿洲湿地环境恶化问题的前提条件,将为干旱区湿地环境治理和生态安全提供理论依据与决策支持. ...
... 干旱区降水稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[4 -5 ] .通常,受植被类型、水分状况、河流径流以及地下水位的影响,植物水分来源与利用方式的辨析与识别具有高度的变异性和复杂性[6 ] .研究表明,除极少数盐生植物外,陆地植物根系吸收水分在木质部运输过程中一般不发生同位素分馏,因此植物茎木质部提取的氢氧稳定同位素比率可反映植物的水分来源[7 ] ,为荒漠绿洲湿地水分来源的辨析提供了水文学依据.目前,国内学者基于稳定同位素技术在荒漠、河岸、海岸等系统中乔灌草等不同生活型植物水分来源和利用策略研究中开展了广泛应用,包括干旱半干旱区[8 -10 ] 、川西亚高山区[11 ] 、青海湖流域[12 ] 和黑河流域[13 -14 ] 等,为探明湿地水分来源、补给途径及其植物水分利用策略提供了思路与技术.然而,基于稳定同位素技术研究干旱区湿地水分来源、补给途径及其植物水分利用策略的研究迄今为止仍未见报道. ...
... 水分来源及其植物水分利用策略是决定湿地植物区系组成、种群分布格局的关键因子[4 -5 ] ,导致荒漠绿洲湿地植被演替往往朝着不确定性和复杂性方向发展.在干旱区,降水量稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[5 ] .因此,确定荒漠绿洲湿地水分来源以及植物水分利用方式是干旱区受损湿地植被保护与恢复急需解决的关键问题[15 -16 ] .在干旱区,小降水(<5 mm)是降水事件的主体,而大降水事件的发生相对较少.本地大气降水线斜率和截距均小于全球大气降水线,而河水、地下水、土壤水都位于大气降水线附近,说明降水对各水源均有补给作用[17 ] ,但并不显著.黑河水主要来源于山区冰雪融水,对地下水补给量大,所以δ 18 O值位于大气降水线左上方;干旱区蒸发强烈,造成土壤水δ 18 O的富集,所以位于大气降水线下方[17 -18 ] .在干旱区,水分输出量远大于降水量,小降水事件的雨滴在降落过程中受云下二次蒸发的影响而发生强烈的同位素动力分馏效应,导致降水中δ D和δ 18 O存在较大波动[19 ] .然而,土壤水分入渗几乎不影响同位素分馏.因此,地表蒸发是导致荒漠绿洲湿地土壤氢、氧同位素富集的主要原因[20 -21 ] . ...
... [5 ].因此,确定荒漠绿洲湿地水分来源以及植物水分利用方式是干旱区受损湿地植被保护与恢复急需解决的关键问题[15 -16 ] .在干旱区,小降水(<5 mm)是降水事件的主体,而大降水事件的发生相对较少.本地大气降水线斜率和截距均小于全球大气降水线,而河水、地下水、土壤水都位于大气降水线附近,说明降水对各水源均有补给作用[17 ] ,但并不显著.黑河水主要来源于山区冰雪融水,对地下水补给量大,所以δ 18 O值位于大气降水线左上方;干旱区蒸发强烈,造成土壤水δ 18 O的富集,所以位于大气降水线下方[17 -18 ] .在干旱区,水分输出量远大于降水量,小降水事件的雨滴在降落过程中受云下二次蒸发的影响而发生强烈的同位素动力分馏效应,导致降水中δ D和δ 18 O存在较大波动[19 ] .然而,土壤水分入渗几乎不影响同位素分馏.因此,地表蒸发是导致荒漠绿洲湿地土壤氢、氧同位素富集的主要原因[20 -21 ] . ...
气候和人类活动影响的湿地植被高光谱研究现状与展望
1
2019
... 干旱区降水稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[4 -5 ] .通常,受植被类型、水分状况、河流径流以及地下水位的影响,植物水分来源与利用方式的辨析与识别具有高度的变异性和复杂性[6 ] .研究表明,除极少数盐生植物外,陆地植物根系吸收水分在木质部运输过程中一般不发生同位素分馏,因此植物茎木质部提取的氢氧稳定同位素比率可反映植物的水分来源[7 ] ,为荒漠绿洲湿地水分来源的辨析提供了水文学依据.目前,国内学者基于稳定同位素技术在荒漠、河岸、海岸等系统中乔灌草等不同生活型植物水分来源和利用策略研究中开展了广泛应用,包括干旱半干旱区[8 -10 ] 、川西亚高山区[11 ] 、青海湖流域[12 ] 和黑河流域[13 -14 ] 等,为探明湿地水分来源、补给途径及其植物水分利用策略提供了思路与技术.然而,基于稳定同位素技术研究干旱区湿地水分来源、补给途径及其植物水分利用策略的研究迄今为止仍未见报道. ...
鄱阳湖湿地典型中生植物水分利用来源的同位素示踪
2
2020
... 干旱区降水稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[4 -5 ] .通常,受植被类型、水分状况、河流径流以及地下水位的影响,植物水分来源与利用方式的辨析与识别具有高度的变异性和复杂性[6 ] .研究表明,除极少数盐生植物外,陆地植物根系吸收水分在木质部运输过程中一般不发生同位素分馏,因此植物茎木质部提取的氢氧稳定同位素比率可反映植物的水分来源[7 ] ,为荒漠绿洲湿地水分来源的辨析提供了水文学依据.目前,国内学者基于稳定同位素技术在荒漠、河岸、海岸等系统中乔灌草等不同生活型植物水分来源和利用策略研究中开展了广泛应用,包括干旱半干旱区[8 -10 ] 、川西亚高山区[11 ] 、青海湖流域[12 ] 和黑河流域[13 -14 ] 等,为探明湿地水分来源、补给途径及其植物水分利用策略提供了思路与技术.然而,基于稳定同位素技术研究干旱区湿地水分来源、补给途径及其植物水分利用策略的研究迄今为止仍未见报道. ...
... 利用多元线性混合模型(IsoSource)上下限方法计算土壤水、河水、地下水、降水等不同潜在水源对湿地植物的贡献率.考虑到干旱环境下部分植物在吸水过程中氢同位素会发生分馏,而氧同位素不会分馏,采用δ 18 O计算不同水源对植物的水分贡献率更为准确[7 ,17 ] .多元线性混合模型的计算方法如下: ...
Hydrogen isotope fractionation during water uptake by woody xerophytes
1
2007
... 干旱区降水稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[4 -5 ] .通常,受植被类型、水分状况、河流径流以及地下水位的影响,植物水分来源与利用方式的辨析与识别具有高度的变异性和复杂性[6 ] .研究表明,除极少数盐生植物外,陆地植物根系吸收水分在木质部运输过程中一般不发生同位素分馏,因此植物茎木质部提取的氢氧稳定同位素比率可反映植物的水分来源[7 ] ,为荒漠绿洲湿地水分来源的辨析提供了水文学依据.目前,国内学者基于稳定同位素技术在荒漠、河岸、海岸等系统中乔灌草等不同生活型植物水分来源和利用策略研究中开展了广泛应用,包括干旱半干旱区[8 -10 ] 、川西亚高山区[11 ] 、青海湖流域[12 ] 和黑河流域[13 -14 ] 等,为探明湿地水分来源、补给途径及其植物水分利用策略提供了思路与技术.然而,基于稳定同位素技术研究干旱区湿地水分来源、补给途径及其植物水分利用策略的研究迄今为止仍未见报道. ...
不同地下水位处梭梭(Haloxylon ammodendron )水分来源特征
0
2021
河西走廊绿洲边缘典型荒漠灌木根系分布特征及其水分利用策略
1
2016
... 干旱区降水稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[4 -5 ] .通常,受植被类型、水分状况、河流径流以及地下水位的影响,植物水分来源与利用方式的辨析与识别具有高度的变异性和复杂性[6 ] .研究表明,除极少数盐生植物外,陆地植物根系吸收水分在木质部运输过程中一般不发生同位素分馏,因此植物茎木质部提取的氢氧稳定同位素比率可反映植物的水分来源[7 ] ,为荒漠绿洲湿地水分来源的辨析提供了水文学依据.目前,国内学者基于稳定同位素技术在荒漠、河岸、海岸等系统中乔灌草等不同生活型植物水分来源和利用策略研究中开展了广泛应用,包括干旱半干旱区[8 -10 ] 、川西亚高山区[11 ] 、青海湖流域[12 ] 和黑河流域[13 -14 ] 等,为探明湿地水分来源、补给途径及其植物水分利用策略提供了思路与技术.然而,基于稳定同位素技术研究干旱区湿地水分来源、补给途径及其植物水分利用策略的研究迄今为止仍未见报道. ...
川西亚高山暗针叶林降水分配过程中氧稳定同位素特征
1
2006
... 干旱区降水稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[4 -5 ] .通常,受植被类型、水分状况、河流径流以及地下水位的影响,植物水分来源与利用方式的辨析与识别具有高度的变异性和复杂性[6 ] .研究表明,除极少数盐生植物外,陆地植物根系吸收水分在木质部运输过程中一般不发生同位素分馏,因此植物茎木质部提取的氢氧稳定同位素比率可反映植物的水分来源[7 ] ,为荒漠绿洲湿地水分来源的辨析提供了水文学依据.目前,国内学者基于稳定同位素技术在荒漠、河岸、海岸等系统中乔灌草等不同生活型植物水分来源和利用策略研究中开展了广泛应用,包括干旱半干旱区[8 -10 ] 、川西亚高山区[11 ] 、青海湖流域[12 ] 和黑河流域[13 -14 ] 等,为探明湿地水分来源、补给途径及其植物水分利用策略提供了思路与技术.然而,基于稳定同位素技术研究干旱区湿地水分来源、补给途径及其植物水分利用策略的研究迄今为止仍未见报道. ...
青海湖流域具鳞水柏枝植物水分利用氢同位素示踪研究
1
2013
... 干旱区降水稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[4 -5 ] .通常,受植被类型、水分状况、河流径流以及地下水位的影响,植物水分来源与利用方式的辨析与识别具有高度的变异性和复杂性[6 ] .研究表明,除极少数盐生植物外,陆地植物根系吸收水分在木质部运输过程中一般不发生同位素分馏,因此植物茎木质部提取的氢氧稳定同位素比率可反映植物的水分来源[7 ] ,为荒漠绿洲湿地水分来源的辨析提供了水文学依据.目前,国内学者基于稳定同位素技术在荒漠、河岸、海岸等系统中乔灌草等不同生活型植物水分来源和利用策略研究中开展了广泛应用,包括干旱半干旱区[8 -10 ] 、川西亚高山区[11 ] 、青海湖流域[12 ] 和黑河流域[13 -14 ] 等,为探明湿地水分来源、补给途径及其植物水分利用策略提供了思路与技术.然而,基于稳定同位素技术研究干旱区湿地水分来源、补给途径及其植物水分利用策略的研究迄今为止仍未见报道. ...
黑河下游河岸林植物水分来源初步研究
1
2008
... 干旱区降水稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[4 -5 ] .通常,受植被类型、水分状况、河流径流以及地下水位的影响,植物水分来源与利用方式的辨析与识别具有高度的变异性和复杂性[6 ] .研究表明,除极少数盐生植物外,陆地植物根系吸收水分在木质部运输过程中一般不发生同位素分馏,因此植物茎木质部提取的氢氧稳定同位素比率可反映植物的水分来源[7 ] ,为荒漠绿洲湿地水分来源的辨析提供了水文学依据.目前,国内学者基于稳定同位素技术在荒漠、河岸、海岸等系统中乔灌草等不同生活型植物水分来源和利用策略研究中开展了广泛应用,包括干旱半干旱区[8 -10 ] 、川西亚高山区[11 ] 、青海湖流域[12 ] 和黑河流域[13 -14 ] 等,为探明湿地水分来源、补给途径及其植物水分利用策略提供了思路与技术.然而,基于稳定同位素技术研究干旱区湿地水分来源、补给途径及其植物水分利用策略的研究迄今为止仍未见报道. ...
临泽荒漠绿洲湿地植物生态系列及其物种多样性研究
4
2014
... 干旱区降水稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[4 -5 ] .通常,受植被类型、水分状况、河流径流以及地下水位的影响,植物水分来源与利用方式的辨析与识别具有高度的变异性和复杂性[6 ] .研究表明,除极少数盐生植物外,陆地植物根系吸收水分在木质部运输过程中一般不发生同位素分馏,因此植物茎木质部提取的氢氧稳定同位素比率可反映植物的水分来源[7 ] ,为荒漠绿洲湿地水分来源的辨析提供了水文学依据.目前,国内学者基于稳定同位素技术在荒漠、河岸、海岸等系统中乔灌草等不同生活型植物水分来源和利用策略研究中开展了广泛应用,包括干旱半干旱区[8 -10 ] 、川西亚高山区[11 ] 、青海湖流域[12 ] 和黑河流域[13 -14 ] 等,为探明湿地水分来源、补给途径及其植物水分利用策略提供了思路与技术.然而,基于稳定同位素技术研究干旱区湿地水分来源、补给途径及其植物水分利用策略的研究迄今为止仍未见报道. ...
... 在黑河流域,荒漠绿洲湿地主要分布在河流源头、绿洲、河滩和内陆湖滨等季节性或常年性积水地段,是河西走廊甚至西北地区重要的水源涵养地,对黑河流域生态安全及社会经济发展发挥重要作用[14 ] .因此,以黑河流域中游河岸灌木湿地与草地盐沼湿地为研究对象,基于稳定同位素技术研究降水、径流、地下水、土壤水和植物水的同位素组成,探究不同水分来源对荒漠植物水分利用的贡献率,将为解释干旱区湿地植物水分利用机理及适应策略提供科学依据和理论基础. ...
... 研究区位于黑河中游张掖国家湿地保护区临泽县境内,为典型的荒漠绿洲湿地(图1 ).该区属温带大陆性干旱气候,夏季干燥炎热,冬季寒冷.年均气温7.6 ℃,最高达39.1 ℃, 最低为-27.3 ℃;年均降水量仅117 mm,集中于6—9月,约占年降水量的65%[14 ] .研究区湿地面积2.10×105 hm2 ,其中自然湿地面积为1.99×105 hm2 ,占湿地面积的94.81%;人工湿地面积1.09×104 hm2 ,占湿地面积的5.19%[14 ] .湿地类型包括永久性河流、季节性河流、洪泛平原湿地、永久性淡水湖、季节性淡水湖、草本沼泽、高山湿地、灌丛湿地、内陆盐沼、池塘、灌溉渠系、稻田、蓄水区及盐田14个类型[15 ] .土壤以盐渍化草甸土和盐土为主,水分来源主要依靠河流径流、浅层地下水和农田灌溉渗漏支撑.植物区系受中亚和蒙古植物区系的影响,湿地植被以多年生草本植物为主,伴生乔灌木树种主要有梭梭(Haloxylon ammodendron )、沙枣(Elaeagnus angustifolia )、柽柳(Tamarix chinensis )等,伴生草本植物有小粒苔草(Carex karoi )、小花灯芯草(Juncus articulatus )、芦苇(Phragmites australis )、盐爪爪(Kalidium foliatum )、赖草(Leymus secalinus )和苦豆子(Sophora alopecuroides )等[16 ] . ...
... [14 ].湿地类型包括永久性河流、季节性河流、洪泛平原湿地、永久性淡水湖、季节性淡水湖、草本沼泽、高山湿地、灌丛湿地、内陆盐沼、池塘、灌溉渠系、稻田、蓄水区及盐田14个类型[15 ] .土壤以盐渍化草甸土和盐土为主,水分来源主要依靠河流径流、浅层地下水和农田灌溉渗漏支撑.植物区系受中亚和蒙古植物区系的影响,湿地植被以多年生草本植物为主,伴生乔灌木树种主要有梭梭(Haloxylon ammodendron )、沙枣(Elaeagnus angustifolia )、柽柳(Tamarix chinensis )等,伴生草本植物有小粒苔草(Carex karoi )、小花灯芯草(Juncus articulatus )、芦苇(Phragmites australis )、盐爪爪(Kalidium foliatum )、赖草(Leymus secalinus )和苦豆子(Sophora alopecuroides )等[16 ] . ...
中国湿地物种多样性与生境面积关系及其生态学机理的模拟研究
2
2004
... 研究区位于黑河中游张掖国家湿地保护区临泽县境内,为典型的荒漠绿洲湿地(图1 ).该区属温带大陆性干旱气候,夏季干燥炎热,冬季寒冷.年均气温7.6 ℃,最高达39.1 ℃, 最低为-27.3 ℃;年均降水量仅117 mm,集中于6—9月,约占年降水量的65%[14 ] .研究区湿地面积2.10×105 hm2 ,其中自然湿地面积为1.99×105 hm2 ,占湿地面积的94.81%;人工湿地面积1.09×104 hm2 ,占湿地面积的5.19%[14 ] .湿地类型包括永久性河流、季节性河流、洪泛平原湿地、永久性淡水湖、季节性淡水湖、草本沼泽、高山湿地、灌丛湿地、内陆盐沼、池塘、灌溉渠系、稻田、蓄水区及盐田14个类型[15 ] .土壤以盐渍化草甸土和盐土为主,水分来源主要依靠河流径流、浅层地下水和农田灌溉渗漏支撑.植物区系受中亚和蒙古植物区系的影响,湿地植被以多年生草本植物为主,伴生乔灌木树种主要有梭梭(Haloxylon ammodendron )、沙枣(Elaeagnus angustifolia )、柽柳(Tamarix chinensis )等,伴生草本植物有小粒苔草(Carex karoi )、小花灯芯草(Juncus articulatus )、芦苇(Phragmites australis )、盐爪爪(Kalidium foliatum )、赖草(Leymus secalinus )和苦豆子(Sophora alopecuroides )等[16 ] . ...
... 水分来源及其植物水分利用策略是决定湿地植物区系组成、种群分布格局的关键因子[4 -5 ] ,导致荒漠绿洲湿地植被演替往往朝着不确定性和复杂性方向发展.在干旱区,降水量稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[5 ] .因此,确定荒漠绿洲湿地水分来源以及植物水分利用方式是干旱区受损湿地植被保护与恢复急需解决的关键问题[15 -16 ] .在干旱区,小降水(<5 mm)是降水事件的主体,而大降水事件的发生相对较少.本地大气降水线斜率和截距均小于全球大气降水线,而河水、地下水、土壤水都位于大气降水线附近,说明降水对各水源均有补给作用[17 ] ,但并不显著.黑河水主要来源于山区冰雪融水,对地下水补给量大,所以δ 18 O值位于大气降水线左上方;干旱区蒸发强烈,造成土壤水δ 18 O的富集,所以位于大气降水线下方[17 -18 ] .在干旱区,水分输出量远大于降水量,小降水事件的雨滴在降落过程中受云下二次蒸发的影响而发生强烈的同位素动力分馏效应,导致降水中δ D和δ 18 O存在较大波动[19 ] .然而,土壤水分入渗几乎不影响同位素分馏.因此,地表蒸发是导致荒漠绿洲湿地土壤氢、氧同位素富集的主要原因[20 -21 ] . ...
黑河流域湿地典型植物群落的物种多样性
2
2012
... 研究区位于黑河中游张掖国家湿地保护区临泽县境内,为典型的荒漠绿洲湿地(图1 ).该区属温带大陆性干旱气候,夏季干燥炎热,冬季寒冷.年均气温7.6 ℃,最高达39.1 ℃, 最低为-27.3 ℃;年均降水量仅117 mm,集中于6—9月,约占年降水量的65%[14 ] .研究区湿地面积2.10×105 hm2 ,其中自然湿地面积为1.99×105 hm2 ,占湿地面积的94.81%;人工湿地面积1.09×104 hm2 ,占湿地面积的5.19%[14 ] .湿地类型包括永久性河流、季节性河流、洪泛平原湿地、永久性淡水湖、季节性淡水湖、草本沼泽、高山湿地、灌丛湿地、内陆盐沼、池塘、灌溉渠系、稻田、蓄水区及盐田14个类型[15 ] .土壤以盐渍化草甸土和盐土为主,水分来源主要依靠河流径流、浅层地下水和农田灌溉渗漏支撑.植物区系受中亚和蒙古植物区系的影响,湿地植被以多年生草本植物为主,伴生乔灌木树种主要有梭梭(Haloxylon ammodendron )、沙枣(Elaeagnus angustifolia )、柽柳(Tamarix chinensis )等,伴生草本植物有小粒苔草(Carex karoi )、小花灯芯草(Juncus articulatus )、芦苇(Phragmites australis )、盐爪爪(Kalidium foliatum )、赖草(Leymus secalinus )和苦豆子(Sophora alopecuroides )等[16 ] . ...
... 水分来源及其植物水分利用策略是决定湿地植物区系组成、种群分布格局的关键因子[4 -5 ] ,导致荒漠绿洲湿地植被演替往往朝着不确定性和复杂性方向发展.在干旱区,降水量稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[5 ] .因此,确定荒漠绿洲湿地水分来源以及植物水分利用方式是干旱区受损湿地植被保护与恢复急需解决的关键问题[15 -16 ] .在干旱区,小降水(<5 mm)是降水事件的主体,而大降水事件的发生相对较少.本地大气降水线斜率和截距均小于全球大气降水线,而河水、地下水、土壤水都位于大气降水线附近,说明降水对各水源均有补给作用[17 ] ,但并不显著.黑河水主要来源于山区冰雪融水,对地下水补给量大,所以δ 18 O值位于大气降水线左上方;干旱区蒸发强烈,造成土壤水δ 18 O的富集,所以位于大气降水线下方[17 -18 ] .在干旱区,水分输出量远大于降水量,小降水事件的雨滴在降落过程中受云下二次蒸发的影响而发生强烈的同位素动力分馏效应,导致降水中δ D和δ 18 O存在较大波动[19 ] .然而,土壤水分入渗几乎不影响同位素分馏.因此,地表蒸发是导致荒漠绿洲湿地土壤氢、氧同位素富集的主要原因[20 -21 ] . ...
Source partitioning using stable isotopes: coping with too many sources
3
2003
... 利用多元线性混合模型(IsoSource)上下限方法计算土壤水、河水、地下水、降水等不同潜在水源对湿地植物的贡献率.考虑到干旱环境下部分植物在吸水过程中氢同位素会发生分馏,而氧同位素不会分馏,采用δ 18 O计算不同水源对植物的水分贡献率更为准确[7 ,17 ] .多元线性混合模型的计算方法如下: ...
... 水分来源及其植物水分利用策略是决定湿地植物区系组成、种群分布格局的关键因子[4 -5 ] ,导致荒漠绿洲湿地植被演替往往朝着不确定性和复杂性方向发展.在干旱区,降水量稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[5 ] .因此,确定荒漠绿洲湿地水分来源以及植物水分利用方式是干旱区受损湿地植被保护与恢复急需解决的关键问题[15 -16 ] .在干旱区,小降水(<5 mm)是降水事件的主体,而大降水事件的发生相对较少.本地大气降水线斜率和截距均小于全球大气降水线,而河水、地下水、土壤水都位于大气降水线附近,说明降水对各水源均有补给作用[17 ] ,但并不显著.黑河水主要来源于山区冰雪融水,对地下水补给量大,所以δ 18 O值位于大气降水线左上方;干旱区蒸发强烈,造成土壤水δ 18 O的富集,所以位于大气降水线下方[17 -18 ] .在干旱区,水分输出量远大于降水量,小降水事件的雨滴在降落过程中受云下二次蒸发的影响而发生强烈的同位素动力分馏效应,导致降水中δ D和δ 18 O存在较大波动[19 ] .然而,土壤水分入渗几乎不影响同位素分馏.因此,地表蒸发是导致荒漠绿洲湿地土壤氢、氧同位素富集的主要原因[20 -21 ] . ...
... [17 -18 ].在干旱区,水分输出量远大于降水量,小降水事件的雨滴在降落过程中受云下二次蒸发的影响而发生强烈的同位素动力分馏效应,导致降水中δ D和δ 18 O存在较大波动[19 ] .然而,土壤水分入渗几乎不影响同位素分馏.因此,地表蒸发是导致荒漠绿洲湿地土壤氢、氧同位素富集的主要原因[20 -21 ] . ...
Multi-scale variability of soil carbon and nitrogen in the middle reaches of the Heihe River basin, northwestern China
1
2016
... 水分来源及其植物水分利用策略是决定湿地植物区系组成、种群分布格局的关键因子[4 -5 ] ,导致荒漠绿洲湿地植被演替往往朝着不确定性和复杂性方向发展.在干旱区,降水量稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[5 ] .因此,确定荒漠绿洲湿地水分来源以及植物水分利用方式是干旱区受损湿地植被保护与恢复急需解决的关键问题[15 -16 ] .在干旱区,小降水(<5 mm)是降水事件的主体,而大降水事件的发生相对较少.本地大气降水线斜率和截距均小于全球大气降水线,而河水、地下水、土壤水都位于大气降水线附近,说明降水对各水源均有补给作用[17 ] ,但并不显著.黑河水主要来源于山区冰雪融水,对地下水补给量大,所以δ 18 O值位于大气降水线左上方;干旱区蒸发强烈,造成土壤水δ 18 O的富集,所以位于大气降水线下方[17 -18 ] .在干旱区,水分输出量远大于降水量,小降水事件的雨滴在降落过程中受云下二次蒸发的影响而发生强烈的同位素动力分馏效应,导致降水中δ D和δ 18 O存在较大波动[19 ] .然而,土壤水分入渗几乎不影响同位素分馏.因此,地表蒸发是导致荒漠绿洲湿地土壤氢、氧同位素富集的主要原因[20 -21 ] . ...
荒漠绿洲湿地土壤水热盐动态过程及其影响机制
1
2019
... 水分来源及其植物水分利用策略是决定湿地植物区系组成、种群分布格局的关键因子[4 -5 ] ,导致荒漠绿洲湿地植被演替往往朝着不确定性和复杂性方向发展.在干旱区,降水量稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[5 ] .因此,确定荒漠绿洲湿地水分来源以及植物水分利用方式是干旱区受损湿地植被保护与恢复急需解决的关键问题[15 -16 ] .在干旱区,小降水(<5 mm)是降水事件的主体,而大降水事件的发生相对较少.本地大气降水线斜率和截距均小于全球大气降水线,而河水、地下水、土壤水都位于大气降水线附近,说明降水对各水源均有补给作用[17 ] ,但并不显著.黑河水主要来源于山区冰雪融水,对地下水补给量大,所以δ 18 O值位于大气降水线左上方;干旱区蒸发强烈,造成土壤水δ 18 O的富集,所以位于大气降水线下方[17 -18 ] .在干旱区,水分输出量远大于降水量,小降水事件的雨滴在降落过程中受云下二次蒸发的影响而发生强烈的同位素动力分馏效应,导致降水中δ D和δ 18 O存在较大波动[19 ] .然而,土壤水分入渗几乎不影响同位素分馏.因此,地表蒸发是导致荒漠绿洲湿地土壤氢、氧同位素富集的主要原因[20 -21 ] . ...
Factors controlling spatial and seasonal distributions of precipitation δ 18 O in China
1
2012
... 水分来源及其植物水分利用策略是决定湿地植物区系组成、种群分布格局的关键因子[4 -5 ] ,导致荒漠绿洲湿地植被演替往往朝着不确定性和复杂性方向发展.在干旱区,降水量稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[5 ] .因此,确定荒漠绿洲湿地水分来源以及植物水分利用方式是干旱区受损湿地植被保护与恢复急需解决的关键问题[15 -16 ] .在干旱区,小降水(<5 mm)是降水事件的主体,而大降水事件的发生相对较少.本地大气降水线斜率和截距均小于全球大气降水线,而河水、地下水、土壤水都位于大气降水线附近,说明降水对各水源均有补给作用[17 ] ,但并不显著.黑河水主要来源于山区冰雪融水,对地下水补给量大,所以δ 18 O值位于大气降水线左上方;干旱区蒸发强烈,造成土壤水δ 18 O的富集,所以位于大气降水线下方[17 -18 ] .在干旱区,水分输出量远大于降水量,小降水事件的雨滴在降落过程中受云下二次蒸发的影响而发生强烈的同位素动力分馏效应,导致降水中δ D和δ 18 O存在较大波动[19 ] .然而,土壤水分入渗几乎不影响同位素分馏.因此,地表蒸发是导致荒漠绿洲湿地土壤氢、氧同位素富集的主要原因[20 -21 ] . ...
长江流域降水稳定同位素的云下二次蒸发效应
1
2010
... 水分来源及其植物水分利用策略是决定湿地植物区系组成、种群分布格局的关键因子[4 -5 ] ,导致荒漠绿洲湿地植被演替往往朝着不确定性和复杂性方向发展.在干旱区,降水量稀少,河流径流、浅层地下水及农田退水成为荒漠绿洲湿地主要水分来源[5 ] .因此,确定荒漠绿洲湿地水分来源以及植物水分利用方式是干旱区受损湿地植被保护与恢复急需解决的关键问题[15 -16 ] .在干旱区,小降水(<5 mm)是降水事件的主体,而大降水事件的发生相对较少.本地大气降水线斜率和截距均小于全球大气降水线,而河水、地下水、土壤水都位于大气降水线附近,说明降水对各水源均有补给作用[17 ] ,但并不显著.黑河水主要来源于山区冰雪融水,对地下水补给量大,所以δ 18 O值位于大气降水线左上方;干旱区蒸发强烈,造成土壤水δ 18 O的富集,所以位于大气降水线下方[17 -18 ] .在干旱区,水分输出量远大于降水量,小降水事件的雨滴在降落过程中受云下二次蒸发的影响而发生强烈的同位素动力分馏效应,导致降水中δ D和δ 18 O存在较大波动[19 ] .然而,土壤水分入渗几乎不影响同位素分馏.因此,地表蒸发是导致荒漠绿洲湿地土壤氢、氧同位素富集的主要原因[20 -21 ] . ...
白皮沙拐枣(Calligonum leucocladum )根系与环境关系的初步研究
1
1996
... 在荒漠绿洲湿地,通常受植被类型、植物生活型、离河距离、地下水埋深以及地貌特征的影响,水分来源和植物水分利用方式表现出显著差异性[22 ] .研究区处于非季风区,属典型的大陆性干旱气候,在降水量小、极端降水频率大及蒸发强烈等因素影响下,降水不能作为荒漠绿洲湿地植物生长的稳定水源[23 -24 ] ,这与我们研究的结论基本一致.通常,干旱地区植物水分利用方式普遍符合两层水分利用模式假设,即深根系木本植物如乔木或灌木利用稳定的深层水源(地下水、深层土壤水),而浅根系多年生或一年生草本植物则依靠表层不稳定的水源(表层土壤水、降水)[25 ] .研究表明,盐沼草地湿地浅层土壤的δ D、δ 18 O均高于河岸灌木湿地,表明地下水是盐沼草本湿地水分蒸散的主要来源,而受河流补给较少;此外河水是河岸灌木湿地的主要水分来源,说明在荒漠绿洲湿地水分来源对植物利用的贡献差异关键是由离河道距离所决定的. ...
新疆塔里木河下游断流河道输水与生态恢复
1
2007
... 在荒漠绿洲湿地,通常受植被类型、植物生活型、离河距离、地下水埋深以及地貌特征的影响,水分来源和植物水分利用方式表现出显著差异性[22 ] .研究区处于非季风区,属典型的大陆性干旱气候,在降水量小、极端降水频率大及蒸发强烈等因素影响下,降水不能作为荒漠绿洲湿地植物生长的稳定水源[23 -24 ] ,这与我们研究的结论基本一致.通常,干旱地区植物水分利用方式普遍符合两层水分利用模式假设,即深根系木本植物如乔木或灌木利用稳定的深层水源(地下水、深层土壤水),而浅根系多年生或一年生草本植物则依靠表层不稳定的水源(表层土壤水、降水)[25 ] .研究表明,盐沼草地湿地浅层土壤的δ D、δ 18 O均高于河岸灌木湿地,表明地下水是盐沼草本湿地水分蒸散的主要来源,而受河流补给较少;此外河水是河岸灌木湿地的主要水分来源,说明在荒漠绿洲湿地水分来源对植物利用的贡献差异关键是由离河道距离所决定的. ...
塔里木河下游优势草本植物与地下水埋深的关系
1
2016
... 在荒漠绿洲湿地,通常受植被类型、植物生活型、离河距离、地下水埋深以及地貌特征的影响,水分来源和植物水分利用方式表现出显著差异性[22 ] .研究区处于非季风区,属典型的大陆性干旱气候,在降水量小、极端降水频率大及蒸发强烈等因素影响下,降水不能作为荒漠绿洲湿地植物生长的稳定水源[23 -24 ] ,这与我们研究的结论基本一致.通常,干旱地区植物水分利用方式普遍符合两层水分利用模式假设,即深根系木本植物如乔木或灌木利用稳定的深层水源(地下水、深层土壤水),而浅根系多年生或一年生草本植物则依靠表层不稳定的水源(表层土壤水、降水)[25 ] .研究表明,盐沼草地湿地浅层土壤的δ D、δ 18 O均高于河岸灌木湿地,表明地下水是盐沼草本湿地水分蒸散的主要来源,而受河流补给较少;此外河水是河岸灌木湿地的主要水分来源,说明在荒漠绿洲湿地水分来源对植物利用的贡献差异关键是由离河道距离所决定的. ...
疏叶骆驼刺母株与子株间的水分整合
1
2013
... 在荒漠绿洲湿地,通常受植被类型、植物生活型、离河距离、地下水埋深以及地貌特征的影响,水分来源和植物水分利用方式表现出显著差异性[22 ] .研究区处于非季风区,属典型的大陆性干旱气候,在降水量小、极端降水频率大及蒸发强烈等因素影响下,降水不能作为荒漠绿洲湿地植物生长的稳定水源[23 -24 ] ,这与我们研究的结论基本一致.通常,干旱地区植物水分利用方式普遍符合两层水分利用模式假设,即深根系木本植物如乔木或灌木利用稳定的深层水源(地下水、深层土壤水),而浅根系多年生或一年生草本植物则依靠表层不稳定的水源(表层土壤水、降水)[25 ] .研究表明,盐沼草地湿地浅层土壤的δ D、δ 18 O均高于河岸灌木湿地,表明地下水是盐沼草本湿地水分蒸散的主要来源,而受河流补给较少;此外河水是河岸灌木湿地的主要水分来源,说明在荒漠绿洲湿地水分来源对植物利用的贡献差异关键是由离河道距离所决定的. ...
准噶尔盆地东南缘多枝柽柳、白刺和红砂水分来源的异同
1
2013
... 通常,土壤水δ 18 O和δ D值主要受蒸发、灌溉、地下水、降水和土壤质地等因素的影响.在干旱区,降水主要以脉动的方式输入,从而引发受降水和蒸发作用控制的表层土壤水稳定同位素组成呈脉动式变化[26 -28 ] .在干旱区,小降水事件只能引起表层土壤水分变化,而大降水事件虽然发生频率较低,但能够有效补给深层土壤水分[29 ] ,降水量越大,降水向深层土壤渗透越快[27 ,30 ] .降水入渗后与土壤原有水分混合构成新的土壤水,其稳定同位素值随着时间的推移不断发生着变化[31 ] .降水事件发生后土壤水δ 18 O值的变化与降水的δ 18 O值和降雨量的大小密切相关[32 ] ,如8 mm降水入渗后荒漠绿洲湿地δ 18 O值的变化幅度相对较小,而24.8 mm降水能显著改变土壤水的δ 18 O值,即降水量与土壤质地共同影响着降水入渗量和入渗深度[29 ] .在荒漠绿洲湿地,受蒸发作用影响重同位素富集[33 ] ,导致浅层土壤水的δ D和δ 18 O值较大,甚至在20 cm深度左右土壤水都出现了δ 18 O最大值,而在20 cm深度以下土壤水δ 18 O随深度增加而逐渐减小.在接近饱和地下水位之前,河岸灌木湿地与草地盐沼湿地土壤水的δ 18 O值都接近地下水的δ 18 O值,表明土壤剖面在上界面受蒸发作用而存在强烈的同位素分馏,而下界面则接受地下水的补给. ...
绍兴淡水湿地人工林优势树种水分利用策略
1
2019
... 通常,土壤水δ 18 O和δ D值主要受蒸发、灌溉、地下水、降水和土壤质地等因素的影响.在干旱区,降水主要以脉动的方式输入,从而引发受降水和蒸发作用控制的表层土壤水稳定同位素组成呈脉动式变化[26 -28 ] .在干旱区,小降水事件只能引起表层土壤水分变化,而大降水事件虽然发生频率较低,但能够有效补给深层土壤水分[29 ] ,降水量越大,降水向深层土壤渗透越快[27 ,30 ] .降水入渗后与土壤原有水分混合构成新的土壤水,其稳定同位素值随着时间的推移不断发生着变化[31 ] .降水事件发生后土壤水δ 18 O值的变化与降水的δ 18 O值和降雨量的大小密切相关[32 ] ,如8 mm降水入渗后荒漠绿洲湿地δ 18 O值的变化幅度相对较小,而24.8 mm降水能显著改变土壤水的δ 18 O值,即降水量与土壤质地共同影响着降水入渗量和入渗深度[29 ] .在荒漠绿洲湿地,受蒸发作用影响重同位素富集[33 ] ,导致浅层土壤水的δ D和δ 18 O值较大,甚至在20 cm深度左右土壤水都出现了δ 18 O最大值,而在20 cm深度以下土壤水δ 18 O随深度增加而逐渐减小.在接近饱和地下水位之前,河岸灌木湿地与草地盐沼湿地土壤水的δ 18 O值都接近地下水的δ 18 O值,表明土壤剖面在上界面受蒸发作用而存在强烈的同位素分馏,而下界面则接受地下水的补给. ...
利用稳定氢氧同位素定量区分白刺水分来源的方法比较
1
2011
... 通常,土壤水δ 18 O和δ D值主要受蒸发、灌溉、地下水、降水和土壤质地等因素的影响.在干旱区,降水主要以脉动的方式输入,从而引发受降水和蒸发作用控制的表层土壤水稳定同位素组成呈脉动式变化[26 -28 ] .在干旱区,小降水事件只能引起表层土壤水分变化,而大降水事件虽然发生频率较低,但能够有效补给深层土壤水分[29 ] ,降水量越大,降水向深层土壤渗透越快[27 ,30 ] .降水入渗后与土壤原有水分混合构成新的土壤水,其稳定同位素值随着时间的推移不断发生着变化[31 ] .降水事件发生后土壤水δ 18 O值的变化与降水的δ 18 O值和降雨量的大小密切相关[32 ] ,如8 mm降水入渗后荒漠绿洲湿地δ 18 O值的变化幅度相对较小,而24.8 mm降水能显著改变土壤水的δ 18 O值,即降水量与土壤质地共同影响着降水入渗量和入渗深度[29 ] .在荒漠绿洲湿地,受蒸发作用影响重同位素富集[33 ] ,导致浅层土壤水的δ D和δ 18 O值较大,甚至在20 cm深度左右土壤水都出现了δ 18 O最大值,而在20 cm深度以下土壤水δ 18 O随深度增加而逐渐减小.在接近饱和地下水位之前,河岸灌木湿地与草地盐沼湿地土壤水的δ 18 O值都接近地下水的δ 18 O值,表明土壤剖面在上界面受蒸发作用而存在强烈的同位素分馏,而下界面则接受地下水的补给. ...
Water utilization of natural and planted trees in the semiarid desert of Inner Mongolia,China
2
2003
... 通常,土壤水δ 18 O和δ D值主要受蒸发、灌溉、地下水、降水和土壤质地等因素的影响.在干旱区,降水主要以脉动的方式输入,从而引发受降水和蒸发作用控制的表层土壤水稳定同位素组成呈脉动式变化[26 -28 ] .在干旱区,小降水事件只能引起表层土壤水分变化,而大降水事件虽然发生频率较低,但能够有效补给深层土壤水分[29 ] ,降水量越大,降水向深层土壤渗透越快[27 ,30 ] .降水入渗后与土壤原有水分混合构成新的土壤水,其稳定同位素值随着时间的推移不断发生着变化[31 ] .降水事件发生后土壤水δ 18 O值的变化与降水的δ 18 O值和降雨量的大小密切相关[32 ] ,如8 mm降水入渗后荒漠绿洲湿地δ 18 O值的变化幅度相对较小,而24.8 mm降水能显著改变土壤水的δ 18 O值,即降水量与土壤质地共同影响着降水入渗量和入渗深度[29 ] .在荒漠绿洲湿地,受蒸发作用影响重同位素富集[33 ] ,导致浅层土壤水的δ D和δ 18 O值较大,甚至在20 cm深度左右土壤水都出现了δ 18 O最大值,而在20 cm深度以下土壤水δ 18 O随深度增加而逐渐减小.在接近饱和地下水位之前,河岸灌木湿地与草地盐沼湿地土壤水的δ 18 O值都接近地下水的δ 18 O值,表明土壤剖面在上界面受蒸发作用而存在强烈的同位素分馏,而下界面则接受地下水的补给. ...
... [29 ].在荒漠绿洲湿地,受蒸发作用影响重同位素富集[33 ] ,导致浅层土壤水的δ D和δ 18 O值较大,甚至在20 cm深度左右土壤水都出现了δ 18 O最大值,而在20 cm深度以下土壤水δ 18 O随深度增加而逐渐减小.在接近饱和地下水位之前,河岸灌木湿地与草地盐沼湿地土壤水的δ 18 O值都接近地下水的δ 18 O值,表明土壤剖面在上界面受蒸发作用而存在强烈的同位素分馏,而下界面则接受地下水的补给. ...
Water source utilization and foliar nutrient status differs between upland and flooded plant communities in wetland tree islands
1
2010
... 通常,土壤水δ 18 O和δ D值主要受蒸发、灌溉、地下水、降水和土壤质地等因素的影响.在干旱区,降水主要以脉动的方式输入,从而引发受降水和蒸发作用控制的表层土壤水稳定同位素组成呈脉动式变化[26 -28 ] .在干旱区,小降水事件只能引起表层土壤水分变化,而大降水事件虽然发生频率较低,但能够有效补给深层土壤水分[29 ] ,降水量越大,降水向深层土壤渗透越快[27 ,30 ] .降水入渗后与土壤原有水分混合构成新的土壤水,其稳定同位素值随着时间的推移不断发生着变化[31 ] .降水事件发生后土壤水δ 18 O值的变化与降水的δ 18 O值和降雨量的大小密切相关[32 ] ,如8 mm降水入渗后荒漠绿洲湿地δ 18 O值的变化幅度相对较小,而24.8 mm降水能显著改变土壤水的δ 18 O值,即降水量与土壤质地共同影响着降水入渗量和入渗深度[29 ] .在荒漠绿洲湿地,受蒸发作用影响重同位素富集[33 ] ,导致浅层土壤水的δ D和δ 18 O值较大,甚至在20 cm深度左右土壤水都出现了δ 18 O最大值,而在20 cm深度以下土壤水δ 18 O随深度增加而逐渐减小.在接近饱和地下水位之前,河岸灌木湿地与草地盐沼湿地土壤水的δ 18 O值都接近地下水的δ 18 O值,表明土壤剖面在上界面受蒸发作用而存在强烈的同位素分馏,而下界面则接受地下水的补给. ...
蒸散发观测研究进展及绿洲蒸散研究展望
2
2011
... 通常,土壤水δ 18 O和δ D值主要受蒸发、灌溉、地下水、降水和土壤质地等因素的影响.在干旱区,降水主要以脉动的方式输入,从而引发受降水和蒸发作用控制的表层土壤水稳定同位素组成呈脉动式变化[26 -28 ] .在干旱区,小降水事件只能引起表层土壤水分变化,而大降水事件虽然发生频率较低,但能够有效补给深层土壤水分[29 ] ,降水量越大,降水向深层土壤渗透越快[27 ,30 ] .降水入渗后与土壤原有水分混合构成新的土壤水,其稳定同位素值随着时间的推移不断发生着变化[31 ] .降水事件发生后土壤水δ 18 O值的变化与降水的δ 18 O值和降雨量的大小密切相关[32 ] ,如8 mm降水入渗后荒漠绿洲湿地δ 18 O值的变化幅度相对较小,而24.8 mm降水能显著改变土壤水的δ 18 O值,即降水量与土壤质地共同影响着降水入渗量和入渗深度[29 ] .在荒漠绿洲湿地,受蒸发作用影响重同位素富集[33 ] ,导致浅层土壤水的δ D和δ 18 O值较大,甚至在20 cm深度左右土壤水都出现了δ 18 O最大值,而在20 cm深度以下土壤水δ 18 O随深度增加而逐渐减小.在接近饱和地下水位之前,河岸灌木湿地与草地盐沼湿地土壤水的δ 18 O值都接近地下水的δ 18 O值,表明土壤剖面在上界面受蒸发作用而存在强烈的同位素分馏,而下界面则接受地下水的补给. ...
... 植物水分利用方式与植物种类型、根系分布、水分条件和生长阶段等有关,但有研究表明不同量级的降水对不同生境优势种的植物水贡献率也有影响[38 ] ,小降水事件对表层(0—20 cm)根系丰富树种植物水的贡献率较高,而较大降水对根系分布较深的植物水的贡献率更高[31 ] ,不同量级降水对各优势树种植物水的贡献率表明深根植物和浅根植物具有不同的水分利用策略[39 ] .根系主要通过根尖的根毛区来吸收水分,植物根系深度和毛细根分布是决定植物吸收水源的重要因素,所以植物细根分布对植物水分利用策略有很大影响[40 ] .本研究发现,荒漠绿洲湿地河岸灌木湿地优势种柽柳浅层根系发达,细根主要分布在40 cm深度内的浅层土壤,随土层加深,柽柳细根逐渐减少.草地盐沼湿地优势种沙枣虽浅层侧根发育较少但主根发达[33 ] .褚建民[34 ] 的研究也表明沙枣是深根植物,能够直接利用浅层地下水,与本研究结果基本相符. ...
1
2013
... 通常,土壤水δ 18 O和δ D值主要受蒸发、灌溉、地下水、降水和土壤质地等因素的影响.在干旱区,降水主要以脉动的方式输入,从而引发受降水和蒸发作用控制的表层土壤水稳定同位素组成呈脉动式变化[26 -28 ] .在干旱区,小降水事件只能引起表层土壤水分变化,而大降水事件虽然发生频率较低,但能够有效补给深层土壤水分[29 ] ,降水量越大,降水向深层土壤渗透越快[27 ,30 ] .降水入渗后与土壤原有水分混合构成新的土壤水,其稳定同位素值随着时间的推移不断发生着变化[31 ] .降水事件发生后土壤水δ 18 O值的变化与降水的δ 18 O值和降雨量的大小密切相关[32 ] ,如8 mm降水入渗后荒漠绿洲湿地δ 18 O值的变化幅度相对较小,而24.8 mm降水能显著改变土壤水的δ 18 O值,即降水量与土壤质地共同影响着降水入渗量和入渗深度[29 ] .在荒漠绿洲湿地,受蒸发作用影响重同位素富集[33 ] ,导致浅层土壤水的δ D和δ 18 O值较大,甚至在20 cm深度左右土壤水都出现了δ 18 O最大值,而在20 cm深度以下土壤水δ 18 O随深度增加而逐渐减小.在接近饱和地下水位之前,河岸灌木湿地与草地盐沼湿地土壤水的δ 18 O值都接近地下水的δ 18 O值,表明土壤剖面在上界面受蒸发作用而存在强烈的同位素分馏,而下界面则接受地下水的补给. ...
柴达木盆地荒漠植物水分来源定量研究:以格尔木样区为例
2
2015
... 通常,土壤水δ 18 O和δ D值主要受蒸发、灌溉、地下水、降水和土壤质地等因素的影响.在干旱区,降水主要以脉动的方式输入,从而引发受降水和蒸发作用控制的表层土壤水稳定同位素组成呈脉动式变化[26 -28 ] .在干旱区,小降水事件只能引起表层土壤水分变化,而大降水事件虽然发生频率较低,但能够有效补给深层土壤水分[29 ] ,降水量越大,降水向深层土壤渗透越快[27 ,30 ] .降水入渗后与土壤原有水分混合构成新的土壤水,其稳定同位素值随着时间的推移不断发生着变化[31 ] .降水事件发生后土壤水δ 18 O值的变化与降水的δ 18 O值和降雨量的大小密切相关[32 ] ,如8 mm降水入渗后荒漠绿洲湿地δ 18 O值的变化幅度相对较小,而24.8 mm降水能显著改变土壤水的δ 18 O值,即降水量与土壤质地共同影响着降水入渗量和入渗深度[29 ] .在荒漠绿洲湿地,受蒸发作用影响重同位素富集[33 ] ,导致浅层土壤水的δ D和δ 18 O值较大,甚至在20 cm深度左右土壤水都出现了δ 18 O最大值,而在20 cm深度以下土壤水δ 18 O随深度增加而逐渐减小.在接近饱和地下水位之前,河岸灌木湿地与草地盐沼湿地土壤水的δ 18 O值都接近地下水的δ 18 O值,表明土壤剖面在上界面受蒸发作用而存在强烈的同位素分馏,而下界面则接受地下水的补给. ...
... 植物水分利用方式与植物种类型、根系分布、水分条件和生长阶段等有关,但有研究表明不同量级的降水对不同生境优势种的植物水贡献率也有影响[38 ] ,小降水事件对表层(0—20 cm)根系丰富树种植物水的贡献率较高,而较大降水对根系分布较深的植物水的贡献率更高[31 ] ,不同量级降水对各优势树种植物水的贡献率表明深根植物和浅根植物具有不同的水分利用策略[39 ] .根系主要通过根尖的根毛区来吸收水分,植物根系深度和毛细根分布是决定植物吸收水源的重要因素,所以植物细根分布对植物水分利用策略有很大影响[40 ] .本研究发现,荒漠绿洲湿地河岸灌木湿地优势种柽柳浅层根系发达,细根主要分布在40 cm深度内的浅层土壤,随土层加深,柽柳细根逐渐减少.草地盐沼湿地优势种沙枣虽浅层侧根发育较少但主根发达[33 ] .褚建民[34 ] 的研究也表明沙枣是深根植物,能够直接利用浅层地下水,与本研究结果基本相符. ...
干旱区植物的水分选择性利用研究
2
2007
... 与深层土壤相比,0—30 cm浅层土壤水的δ D和δ 18 O值变化更复杂[34 ] .受强烈蒸发的影响,0—10 cm表层土壤水δ D和δ 18 O富集,且随着土层深度的增加,蒸发作用逐渐减弱,δ D和δ 18 O趋于贫化,即浅层土壤水的δ D和δ 18 O值较大,深层土壤水的δ D和δ 18 O值较小.然而,荒漠绿洲湿地土壤水δ D和δ 18 O的最小值均出现在30 cm土层附近,这是由于该土层的黏粒含量最高且土壤持水性好[35 -36 ] ,土壤水氢氧同位素不易被蒸发而产生富集,故其稳定同位素值小[37 ] .然而,随着灌溉和降水事件的发生,土壤δ D和δ 18 O值逐渐减小.而大于30 cm的深层土壤水主要受地下水稳定补给的影响,其氢氧稳定同位素值比较平稳.因此,土壤水δ D和δ 18 O值的时空变化表现了土壤蒸发、灌溉水及降水入渗、地下水补给等因素对土壤水的影响. ...
... 植物水分利用方式与植物种类型、根系分布、水分条件和生长阶段等有关,但有研究表明不同量级的降水对不同生境优势种的植物水贡献率也有影响[38 ] ,小降水事件对表层(0—20 cm)根系丰富树种植物水的贡献率较高,而较大降水对根系分布较深的植物水的贡献率更高[31 ] ,不同量级降水对各优势树种植物水的贡献率表明深根植物和浅根植物具有不同的水分利用策略[39 ] .根系主要通过根尖的根毛区来吸收水分,植物根系深度和毛细根分布是决定植物吸收水源的重要因素,所以植物细根分布对植物水分利用策略有很大影响[40 ] .本研究发现,荒漠绿洲湿地河岸灌木湿地优势种柽柳浅层根系发达,细根主要分布在40 cm深度内的浅层土壤,随土层加深,柽柳细根逐渐减少.草地盐沼湿地优势种沙枣虽浅层侧根发育较少但主根发达[33 ] .褚建民[34 ] 的研究也表明沙枣是深根植物,能够直接利用浅层地下水,与本研究结果基本相符. ...
黑河下游不同林龄胡杨水分来源的D、δ 18 O同位素示踪
1
2014
... 与深层土壤相比,0—30 cm浅层土壤水的δ D和δ 18 O值变化更复杂[34 ] .受强烈蒸发的影响,0—10 cm表层土壤水δ D和δ 18 O富集,且随着土层深度的增加,蒸发作用逐渐减弱,δ D和δ 18 O趋于贫化,即浅层土壤水的δ D和δ 18 O值较大,深层土壤水的δ D和δ 18 O值较小.然而,荒漠绿洲湿地土壤水δ D和δ 18 O的最小值均出现在30 cm土层附近,这是由于该土层的黏粒含量最高且土壤持水性好[35 -36 ] ,土壤水氢氧同位素不易被蒸发而产生富集,故其稳定同位素值小[37 ] .然而,随着灌溉和降水事件的发生,土壤δ D和δ 18 O值逐渐减小.而大于30 cm的深层土壤水主要受地下水稳定补给的影响,其氢氧稳定同位素值比较平稳.因此,土壤水δ D和δ 18 O值的时空变化表现了土壤蒸发、灌溉水及降水入渗、地下水补给等因素对土壤水的影响. ...
水盐梯度下湿地柽柳(Tamarix ramosissima )光合响应特征和水分利用效率
1
2018
... 与深层土壤相比,0—30 cm浅层土壤水的δ D和δ 18 O值变化更复杂[34 ] .受强烈蒸发的影响,0—10 cm表层土壤水δ D和δ 18 O富集,且随着土层深度的增加,蒸发作用逐渐减弱,δ D和δ 18 O趋于贫化,即浅层土壤水的δ D和δ 18 O值较大,深层土壤水的δ D和δ 18 O值较小.然而,荒漠绿洲湿地土壤水δ D和δ 18 O的最小值均出现在30 cm土层附近,这是由于该土层的黏粒含量最高且土壤持水性好[35 -36 ] ,土壤水氢氧同位素不易被蒸发而产生富集,故其稳定同位素值小[37 ] .然而,随着灌溉和降水事件的发生,土壤δ D和δ 18 O值逐渐减小.而大于30 cm的深层土壤水主要受地下水稳定补给的影响,其氢氧稳定同位素值比较平稳.因此,土壤水δ D和δ 18 O值的时空变化表现了土壤蒸发、灌溉水及降水入渗、地下水补给等因素对土壤水的影响. ...
额济纳三角洲胡杨和多枝柽柳水分来源解析
1
2017
... 与深层土壤相比,0—30 cm浅层土壤水的δ D和δ 18 O值变化更复杂[34 ] .受强烈蒸发的影响,0—10 cm表层土壤水δ D和δ 18 O富集,且随着土层深度的增加,蒸发作用逐渐减弱,δ D和δ 18 O趋于贫化,即浅层土壤水的δ D和δ 18 O值较大,深层土壤水的δ D和δ 18 O值较小.然而,荒漠绿洲湿地土壤水δ D和δ 18 O的最小值均出现在30 cm土层附近,这是由于该土层的黏粒含量最高且土壤持水性好[35 -36 ] ,土壤水氢氧同位素不易被蒸发而产生富集,故其稳定同位素值小[37 ] .然而,随着灌溉和降水事件的发生,土壤δ D和δ 18 O值逐渐减小.而大于30 cm的深层土壤水主要受地下水稳定补给的影响,其氢氧稳定同位素值比较平稳.因此,土壤水δ D和δ 18 O值的时空变化表现了土壤蒸发、灌溉水及降水入渗、地下水补给等因素对土壤水的影响. ...
鄱阳湖典型洲滩湿地水分补排关系
1
2017
... 植物水分利用方式与植物种类型、根系分布、水分条件和生长阶段等有关,但有研究表明不同量级的降水对不同生境优势种的植物水贡献率也有影响[38 ] ,小降水事件对表层(0—20 cm)根系丰富树种植物水的贡献率较高,而较大降水对根系分布较深的植物水的贡献率更高[31 ] ,不同量级降水对各优势树种植物水的贡献率表明深根植物和浅根植物具有不同的水分利用策略[39 ] .根系主要通过根尖的根毛区来吸收水分,植物根系深度和毛细根分布是决定植物吸收水源的重要因素,所以植物细根分布对植物水分利用策略有很大影响[40 ] .本研究发现,荒漠绿洲湿地河岸灌木湿地优势种柽柳浅层根系发达,细根主要分布在40 cm深度内的浅层土壤,随土层加深,柽柳细根逐渐减少.草地盐沼湿地优势种沙枣虽浅层侧根发育较少但主根发达[33 ] .褚建民[34 ] 的研究也表明沙枣是深根植物,能够直接利用浅层地下水,与本研究结果基本相符. ...
黑河下游典型生态系统水分补给源及优势植物水分来源研究
1
2012
... 植物水分利用方式与植物种类型、根系分布、水分条件和生长阶段等有关,但有研究表明不同量级的降水对不同生境优势种的植物水贡献率也有影响[38 ] ,小降水事件对表层(0—20 cm)根系丰富树种植物水的贡献率较高,而较大降水对根系分布较深的植物水的贡献率更高[31 ] ,不同量级降水对各优势树种植物水的贡献率表明深根植物和浅根植物具有不同的水分利用策略[39 ] .根系主要通过根尖的根毛区来吸收水分,植物根系深度和毛细根分布是决定植物吸收水源的重要因素,所以植物细根分布对植物水分利用策略有很大影响[40 ] .本研究发现,荒漠绿洲湿地河岸灌木湿地优势种柽柳浅层根系发达,细根主要分布在40 cm深度内的浅层土壤,随土层加深,柽柳细根逐渐减少.草地盐沼湿地优势种沙枣虽浅层侧根发育较少但主根发达[33 ] .褚建民[34 ] 的研究也表明沙枣是深根植物,能够直接利用浅层地下水,与本研究结果基本相符. ...
Using stable hydrogen and oxygen isotopes to study water movement in soil-plant-atmosphere continuum at Poyang Lake wetland,China
1
2017
... 植物水分利用方式与植物种类型、根系分布、水分条件和生长阶段等有关,但有研究表明不同量级的降水对不同生境优势种的植物水贡献率也有影响[38 ] ,小降水事件对表层(0—20 cm)根系丰富树种植物水的贡献率较高,而较大降水对根系分布较深的植物水的贡献率更高[31 ] ,不同量级降水对各优势树种植物水的贡献率表明深根植物和浅根植物具有不同的水分利用策略[39 ] .根系主要通过根尖的根毛区来吸收水分,植物根系深度和毛细根分布是决定植物吸收水源的重要因素,所以植物细根分布对植物水分利用策略有很大影响[40 ] .本研究发现,荒漠绿洲湿地河岸灌木湿地优势种柽柳浅层根系发达,细根主要分布在40 cm深度内的浅层土壤,随土层加深,柽柳细根逐渐减少.草地盐沼湿地优势种沙枣虽浅层侧根发育较少但主根发达[33 ] .褚建民[34 ] 的研究也表明沙枣是深根植物,能够直接利用浅层地下水,与本研究结果基本相符. ...
稳定氢氧同位素在定量区分植物水分利用来源中的应用
1
2007
... 潜在水源对植物水分利用策略至关重要[41 ] .在荒漠绿洲湿地,不同降水事件对土壤水和植物水δ D的贡献率虽有差异,但未达到显著水平(表3 ).同时,土壤水及土壤水与地下水的交互作用显著地影响着河岸灌木湿地、盐沼草地湿地土壤水和植物水δ D(P <0.001),说明土壤水是维持荒漠绿洲湿地植物生长的决定性因素.在草地盐沼湿地中,地下水对土壤水与植物水δ D的影响存在显著性差异(P <0.01),表明地下水是离河较远的草地湿地土壤与植物水最主要水分来源.此外,降水、地下水和土壤水的交互作用也显著地影响着盐沼草地湿地土壤水和植物水δ D (P <0.001).由此可见,在河岸灌木湿地、盐沼草地,深根植物和浅根植物具有不同的水分利用策略[42 ] ,地下水对荒漠绿洲湿地植物水的贡献率随着距离河道距离而减小. ...
用D、δ 18 O同位素确定黑河中游戈壁地区植物水分来源
1
2012
... 潜在水源对植物水分利用策略至关重要[41 ] .在荒漠绿洲湿地,不同降水事件对土壤水和植物水δ D的贡献率虽有差异,但未达到显著水平(表3 ).同时,土壤水及土壤水与地下水的交互作用显著地影响着河岸灌木湿地、盐沼草地湿地土壤水和植物水δ D(P <0.001),说明土壤水是维持荒漠绿洲湿地植物生长的决定性因素.在草地盐沼湿地中,地下水对土壤水与植物水δ D的影响存在显著性差异(P <0.01),表明地下水是离河较远的草地湿地土壤与植物水最主要水分来源.此外,降水、地下水和土壤水的交互作用也显著地影响着盐沼草地湿地土壤水和植物水δ D (P <0.001).由此可见,在河岸灌木湿地、盐沼草地,深根植物和浅根植物具有不同的水分利用策略[42 ] ,地下水对荒漠绿洲湿地植物水的贡献率随着距离河道距离而减小. ...





 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}