白刺( Nitraria tangutorum
李小乐 , 1 , 党晓宏 , 1 , 2 , 翟波 1 , 魏亚娟 1 , 迟旭 1 , 吴惠敏 1
1.内蒙古农业大学 沙漠治理学院,内蒙古 呼和浩特 010018
2.内蒙古杭锦荒漠生态系统国家定位观测研究站,内蒙古 鄂尔多斯 017400
Adventitious root architecture and growth characteristics of Nitraria tangutorum shrub
Li Xiaole , 1 , Dang Xiaohong , 1 , 2 , Zhai Bo 1 , Wei Yajuan 1 , Chi Xu 1 , Wu Huimin 1
1.College of Desert Control Science and Engineering,Inner Mongolia Agricultural University,Hohhot 010018,China
2.Hangjin National Positioning Observation and Research Station of Desert Ecosystem,Ordos 017400,Inner Mongolia,China
通讯作者: 党晓宏(E-mail: dangxiaohong1986@126.com )
收稿日期: 2021-10-21
修回日期: 2022-02-20
基金资助:
国家自然科学基金项目 . 41967009 内蒙古自治区研究生科研创新项目 . S20210211Z
Received: 2021-10-21
Revised: 2022-02-20
摘要
植物根系构型和生物量分配模式能有效反映植物对干旱贫瘠环境的适应策略。以西鄂尔多斯国家级自然保护区内的唐古特白刺(Nitraria tangutorum )灌丛为研究对象,采用传统挖掘法对灌丛不定根系构型特征和生物量分配模式进行野外实地调查。结果表明:(1)不同生长阶段白刺灌丛不定根系均以垂直分布的深根型为主。(2)不定根系构型均为人字形,雏形阶段根系拓扑指数T1 、修正拓扑参数qa 和qb 分别为0.98、0.95、0.52,发育阶段为0.87、0.69、0.46,成熟阶段为0.85、0.63、0.63。雏形阶段灌丛分支率较发育和成熟阶段分别小8.96%和30.05%。随白刺灌丛不断发育,各级根径逐渐增大,灌丛分支逐渐复杂,资源利用范围逐渐扩大。相较于灌丛不定根系,生物量分配重心始终倾斜于灌丛地上部分。(3)成熟阶段灌丛不定根系与地上部分的根冠比较雏形阶段和发育阶段分别降低32.75%和0.83%,其中不定根系粗根(三级根)占比减少,细根(二级根和一级根)占比提高。白刺灌丛在生长过程中通过提高不定根系分支率、增加灌丛地上部分和不定根系细根的生物量等方式适应胁迫环境。
关键词:
白刺(Nitraria tangutorum ) 不定根系 根系构型 生物量分配 相对生长关系
Abstract
Root architecture and biomass allocation patterns can effectively reflect the adaptive strategies of plants to arid and infertile environments. Abstract: In this study, Nitraria tangutorum shrub in the West Ordos National Nature Reserve was used as the research object, and the characteristics of the root architecture and biomass allocation pattern of the shrub were investigated by using the traditional excavation method. The results showed as follows :(1) the main adventitious roots in Nitraria tangutorum shrub at different growth stages were vertical deep-root type. (2) The root topological index TI and the modified topological parameters qa and qb were 0.98, 0.95 and 0.52, respectively. The developmental stages were 0.87, 0.69, 0.46. The mature stage is 0.85, 0.63, 0.63. The branching rate of shrub in embryonic stage decreased by 8.96% and 30.05% compared with that in developing and mature stage, respectively. With the development of Nitraria tangutorum shrub, the root diameter of different levels increased gradually, the shrub branches became more complex, and the utilization range of resources expanded gradually. However, compared with the adventitious roots, the biomass distribution center was always inclined to the aboveground part of the shrub. (3) Compared with the embryonic stage and development stage, the ratio of coarse root (tertiary root) and fine root (secondary root and primary root) of the adventitious roots in the mature stage decreased by 32.75% and 0.83%, respectively. In the process of growth, Nitraria tangutorum shrub adapted to the stress environment by increasing the branching rate of its adventitious roots and the biomass of its aboveground and fine roots.
Keywords:
Nitraria tangutorum adventitious roots root architecture biomass allocation allometric relationships
本文引用格式
李小乐, 党晓宏, 翟波, 魏亚娟, 迟旭, 吴惠敏. 白刺( Nitraria tangutorum . 中国沙漠 [J], 2022, 42(4): 172-180 doi:10.7522/j.issn.1000-694X.2022.00021
Li Xiaole, Dang Xiaohong, Zhai Bo, Wei Yajuan, Chi Xu, Wu Huimin. Adventitious root architecture and growth characteristics of Nitraria tangutorum shrub . Journal of Desert Research
0 引言
根系是植物与外界环境进行物质交换的重要器官[1 ] ,根系在土壤中的分布和排列方式被称为根系构型[2 ] ,构型特征随土壤环境变化会发生动态调整,这是植物对异质性土壤资源的反馈机制和适应策略[3 ] 。生物量是衡量个体在生态系统中功能与地位的重要指标[4 ] ,植物的根系生物量所占比例反映了植物为适应环境以求对光、水、养分等资源最大吸收利用所做出的适应[5 ] 。处于不同生长阶段的植物会表现出不同的根系构型和生物量分配模式以适应环境[6 -8 ] 。因此,探究植物的根系构型和生物量分配模式在胁迫环境中的适应机制,对于干旱区的荒漠化防治及植物多样性保护具有重要意义。
西鄂尔多斯国家自然保护区位于鄂尔多斯高原西缘,独特的气候条件和地形地貌使这里成为了许多古老孑遗植物的“避难所”[9 ] 。白刺(Nitraria tangutorum )作为研究区主要建群种,因其耐旱耐瘠薄的特性为研究区生态修复发挥着重要作用。白刺灌丛易形成沙堆,沙堆沉积物包含了大量环境信息,对于描述区域土壤退化和风蚀情况有重要意义[10 ] 。对沙堆调查后发现其内部长有大量不定根,并且不同生长阶段的不定根构型和生物量分配表现出显著差异。前人研究发现沙堆的形成和发育与不定根生长有着密切联系[11 ] ,但目前缺少不定根系构型以及生物量分配模式的研究,不定根随灌丛生长和沙堆发育会做出怎样适应策略也不明晰。
鉴于此,本试验对不同生长阶段白刺沙堆内的不定根系构型特征及生物量分配模式开展研究,以期揭示白刺灌丛在适应胁迫环境过程中对不定根系生态策略调整及其与沙堆形态发育的互馈机制,为荒漠植物的保护与恢复提供理论依据及数据支持。
1 材料与方法
1.1 研究区概况
西鄂尔多斯国家级自然保护区(39°13′—40°10′N,106°44′—107°43′E)属暖温带大陆性气候区,年均气温7.8—8.1 ℃,年降水量162—172 mm,集中在6—8月,年潜在蒸发量2 470—3 481 mm。研究区以偏西北风为主,年均风速3.2 m·s-1 ,最大风速可达24.2 m·s-1 。土壤类型主要为棕钙土、灰漠土,具有土层浅薄、土质粗糙、养分贫瘠、腐殖质含量低等特点[12 ] 。研究区是强旱生灌丛为建群种的荒漠化草原,优势种为白刺、沙冬青(Ammopiptanthus mongolicus )、霸王(Zygophyllum xanthoxylon )、红砂(Reaumuria soongarica )、半日花(Helianthemum songaricum ),伴生植物有绵刺(Potaninia mongolica )、珍珠猪毛菜(Salsola passerina )等,并伴有一定数量的草本植物。植物所需水分多来自天然降水,地下水埋深大于15 m[13 ] 。
1.2 研究方法
野外试验时间为2021年8月中旬,在研究区设置一个100 m×100 m的样方,样方内以白刺灌丛为主,并少量分布植被盖度10%—30%的其他小型沙生植物,沙生植物分布于白刺灌丛沙堆四周,未对沙堆内部的不定根生长产生影响(图1 )。采用“空间代替时间”的方法,结合相关文献与样地实际情况[14 ] ,选择研究区内具有代表性的白刺灌丛为研究对象。将白刺灌丛分为雏形、发育和成熟3个生长阶段,每阶段各3丛,共9丛。
图1
图1
白刺灌丛及其不定根
Fig.1
Nitraria tangutorum shrub and its adventitious roots
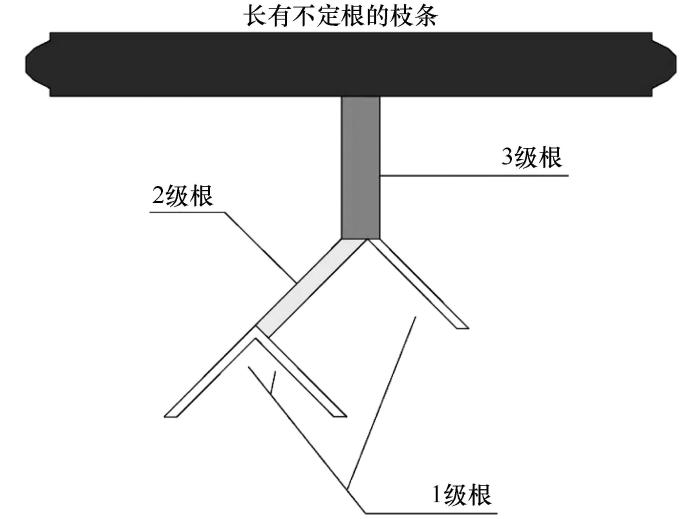
采用Strahler法对不定根系进行分级[15 -16 ] ,由外及内确定根序,最外根系为1级根,两个1级根系相遇为2级根,以此类推(图2 )。若不同级根系交汇取较高级者。
图2
图2
不定根系分级方法示意图
Fig.2
Schematic diagram of adventitious root classification method
用小铲挖掘灌丛沙堆,挖出不定根后用直尺记录不定根生长所处沙埋深度,用量角器测量3级根与地平线的夹角以表示不定根系的空间分布特征(夹角β <30°为水平根,60°>β >30°为倾斜根,β >60°为垂直根)。每株灌丛选择8个3级不定根及其下生长所有次生不定根,用游标卡尺测量各级根径,记录各级根系数量及连接数,结果取均值以减小误差。采集1/4灌丛的全部不定根系和灌丛枝系,拿回实验室风干用于后期生物量测定。
1.2.1 灌丛沙堆形态特征测定
用卷尺对灌丛长轴(L g )、短轴(W g )、株高(H g )及沙堆的长轴(L d )、短轴(W d )、高度(H d )进行测量(表1 )。灌丛冠幅(C )计算公式如下:
C = ( L g + W g ) / 2 (1)
由于灌丛沙堆呈椭圆形。因此,其底面积(S d )及沙堆体积(V d )[17 ] 计算公式如下:
S d = 1 / 4 × π × L d × W d (2)
V d = 1 / 6 × π × L d × W d × H d (3)
1.2.2 不定根分支率计算
根系总分支率(OBRr ):根据Strahler分支等级[15 -16 ] 计算每个等级(i )根的数量Ni ,分别以根级i 和lgNi 为横、纵坐标作图,回归直线的斜率绝对值的逆对数即为根系总分支率。逐级分支率(SBR (i :i +1))为i 级分支数与前一级的分支数之比,即:
S B R ( i : i + 1 ) = N i / ( N i + 1 ) (4)
1.2.3 不定根根径比计算
根径比(RBD (i :i +1))为i 级根和前一级根的比率,即:
R B D ( i : i + 1 ) = B D i / B D i + 1 (5)
式中:BDi 和BDi+ 1 分别为i 级分支和i +1级分支的直径。
1.2.4 拓扑参数计算
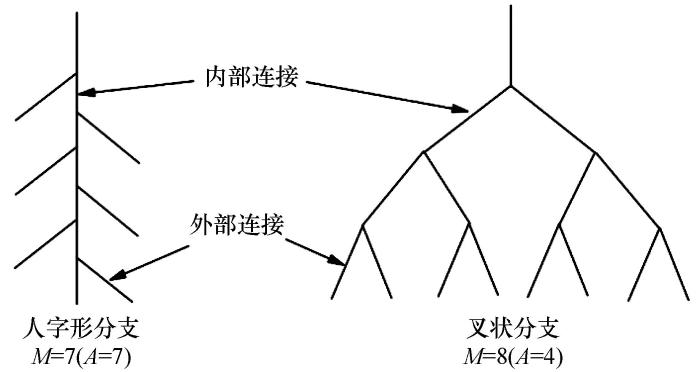
Fitter[18 ] 和Bouma等[19 ] 提出根系分支存在人字形分支和叉状分支两种极端模式(图3 ),并提出拓扑指数表达式:
T I = l g A / l g M (6)
式中:TI 为拓扑指数;A 为最长根系通道内部连接总数;M 为所有外部根系连接总数。TI 越接近1表明A 和M 值相近,根系分支少,形状接近人字形分支结构;TI 越接近0.5表明根系结构越接近叉状分支。Oppelt等[20 ] 对拓扑指数构型进行了进一步研究,并提出新的拓扑参数。其表达式为:
q a = a - 1 - l b v 0 v 0 - 1 - l b v 0 (7)
q b = b - 1 - l b v 0 ( v 0 + 1 ) / 2 - 1 / v 0 - l b v 0 (8)
式中:qa 和qb 分别表示a 和b 的修正值;a 与Fitter模型中的A 相等;b 为平均拓扑长度,b=Pe/v0 ,Pe 为基部带根部末端所有连接总数;lbv 0 = lnv 0 / ln2 ,其中v 0 等同Fitter中的M 。人字形分支为qa =qb =1,叉状形分支为qa =qb =0。
图3
图3
根系拓扑结构示意图
Fig.3
Schematic diagram of root topological structure
1.2.5 不定根根冠比计算
将风干后的灌丛放置烘箱于105 ℃下杀青30 min,在80 ℃下烘至恒重,用电子天平(精度0.001 g)称重后乘4倍表示生物量。不定根根冠比(RSR )计算如式(9)。
R S R = B G B / A G B (9)
式中:AGB 为灌丛地上部分生物量;BGB 为灌丛不定根系生物量。
1.2.6 异速生长机制模型
利用异速生长机制模型对白刺灌丛各生长阶段AGB 与BGB 进行相关生长分析[21 ] 。
Y = β X α (10)
l g Y = l g β + α l g X (11)
式中:Y 为BGB ;X 为AGB ;β 为常数;α 为相关生长指数。当α =1时,BGB 和AGB 为等速生长;当α ≠1时,为异速生长[22 ] 。判断α 是否等于1,利用α 的95%置信区间是否包含1来确定。
1.3 数据处理及分析
运用Microsoft Excel 2010对不定根系构形参数进行计算并构建生物量异速生长机制模型,采用DPS数据处理系统对数据进行单因素方差分析(One-way ANOVA),显著性水平设为0.05。运用Origin 2018对处理后的数据进行绘图。
2 结果与分析
2.1 不同生长阶段的白刺灌丛不定根系构型特征
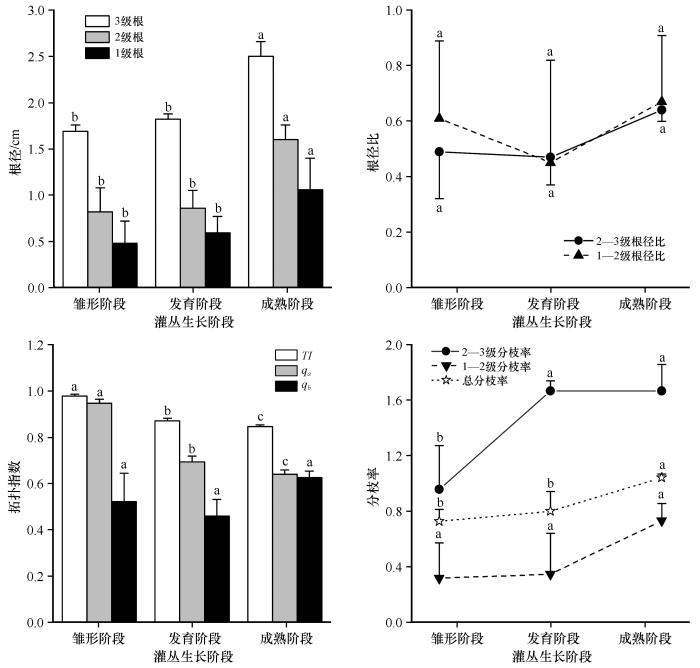
白刺灌丛不定根根径与灌丛形态显著正相关(P <0.05,图4 ),成熟阶段灌丛的1级、2级和3级不定根根径分别较发育阶段、雏形阶段灌丛增加79.66%和120.83%、86.05%和95.12%、37.36%和47.93%。不同生长阶段的白刺灌丛各级不定根根径比为0.47—0.75,且未表现出显著差异性。除qb 无显著差异外,白刺灌丛其他拓扑指数参数均随灌丛发育显著减小,雏形阶段灌丛qa =0.95,qb =0.52,TI =0.98;发育阶段灌丛qa =0.69,qb =0.46,TI =0.87;成熟阶段灌丛qa =0.63,qb =0.63,TI =0.85。不同发育阶段白刺灌丛的不定根系构型均属于人字形分支,但随灌丛发育,拓扑结构逐渐复杂,拥有更多次级分支。不定根总分支率随灌丛生长而逐渐增大,雏形阶段灌丛显著小于发育阶段和成熟阶段灌丛,分别减小8.96%和30.05%。雏形阶段灌丛2—3级不定根分支率显著小于发育阶段和成熟阶段灌丛,分别减小42.50%和42.50%,而不同生长阶段的灌丛1—2级不定根分支率大小各异,但未表现出显著差异。
图4
图4
白刺灌丛不定根系构型特征
不同字母表示差异显著,P <0.05
Fig.4
Adventitious root architecture characteristics of Nitraria tangutorum shrub
不定根系埋深7.42—10.57 cm,且埋深与灌丛的生长阶段无关(表2 )。不同生长阶段的白刺灌丛不定根系均以垂直根(52.40%)为主,倾斜根其次(36.11%),水平根最少(11.49%)。
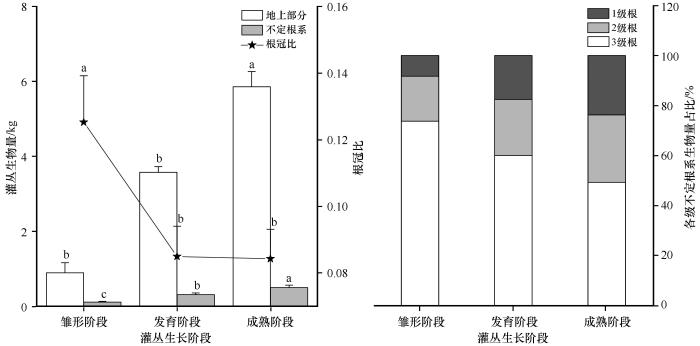
2.2 不同生长阶段的白刺灌丛生物量分配模式
白刺灌丛地上部分和不定根系生物量均随生长阶段的变化逐渐增加,且各生长阶段间存在显著差异(P <0.05,图5 )。灌丛的生长发育可不断改善土壤微环境,成熟阶段灌丛地上部分生物量分别较处于相对胁迫环境的发育阶段、雏形阶段灌丛增加63.83%和554.73%,不定根生物量分别较发育阶段、雏形阶段灌丛增加62.26%和350.13%。白刺灌丛1级根、2级根、3级根所占比例分别为8.29%、18.04%、74.67%(雏形阶段)、17.59%、22.38%、60.02%(发育阶段)、13.73%、27.03%、59.25%(成熟阶段)。在西鄂尔多斯地区胁迫环境条件下的不定根根冠比随生长阶段的不断变化而逐级递减,其中雏形阶段灌丛根冠比较发育阶段和成熟阶段灌丛分别高45.28%和48.69%。
图5
图5
白刺灌丛不定根生物量分配特征
不同字母表示差异显著,P <0.05
Fig.5
Biomass allocation characteristics of adventitious roots in Nitraria tangutorum shrub
2.3 不同生长阶段白刺灌丛地上部分和不定根系生物量相对生长关系
不同生长阶段白刺灌丛的地上部分和不定根系的生长关系均表现为异速生长(表3 )。从相关生长指数的均值和95%置信区间可以看出,不同生长阶段的白刺灌丛相关生长指数均小于1,说明白刺灌丛地上部分生长速度始终大于地下不定根生长速度。从不同生长阶段来看,随着灌丛生长发育,不定根系生长速率不断提高,雏形阶段灌丛相关生长指数较发育阶段和成熟阶段分别低280.00%和190.00%。这可能与不定根系在沙堆内部独特的生长环境有关,灌丛枝系被沙埋后逐渐长出不定根系,而随着灌丛地上部分的不断生长,枝系拦截风沙流能力不断提高,沙堆形态逐渐增大,因此被埋枝系不断增多,致使不定根系增长速率增加,而不定根生长可固定沙堆促使其继续增大[23 ] 。
3 讨论
3.1 不同生长阶段白刺灌丛根系构型特征
根系构型特征随不同的水分、养分条件产生变化,具有一定可塑性[24 ] 。根系构型差异影响根系对土壤水分的获取能力以及碳的消耗[25 ] 与分配[26 ] 。灌丛的根径与其寿命正相关,但较大的根径会消耗更多的碳,而干旱胁迫环境下灌丛根系的高死亡率会导致大量碳损失,不利于灌丛的生存[27 ] 。因此,生长阶段初期的雏形阶段灌丛为适应干旱环境,选择降低根径以减少根系死亡所带来的碳消耗。而成熟阶段灌丛的根径显著大于雏形阶段和发育阶段灌丛也反映了随着灌丛的不断生长其抗干旱胁迫能力不断增强,有能力适应恶劣环境后选择寿命更长的较粗不定根。1—2级根径比小于2—3级,说明了随根径级数增加根径逐渐变小,但不同生长阶段的白刺灌丛不定根系根径比均未表现出显著差异。各级根系分支前后的一致性反映了根系的自相似性,因此可通过已测数据对部分构型指标进行估算,从而获得相关数据。这是对植物构型研究及分析的有效方法,不仅降低了工作难度,并且减少了对植物破坏[7 ] 。
植物会随着环境的不断变化对其根系构型做出调整,以期达到对资源的最大利用[28 ] 。前人研究发现,植物的根系分支率与环境干旱胁迫程度负相关[6 ] 。在等量碳投入条件下,虽然人字形分支分布范围更小,不利于养分占有,但分支结构简单,对资源吸收利用能力更强,更适应胁迫环境[7 ,18 ] 。本研究中随灌丛生长发育其拓扑指数参数显著降低,成熟阶段灌丛的分支率显著高于发育阶段和雏形阶段灌丛,不同生长阶段白刺灌丛不定根系构型均表现为人字形,该结论与单立山等[29 ] 对地下根系的研究结论不同。造成该结论差异的原因可能与不同地区资源差异有关,也可能是由于不定根系和地下根系自身特性差异有关,有待进一步研究。本试验的结果表明了研究区的白刺灌丛随生长发育抗干旱胁迫能力不断增强,成熟阶段灌丛不同于发育阶段和雏形阶段灌丛的将更多精力放在对环境适应上,而是有余力通过增加不定根系分支率来扩张其对资源的吸收利用范围以满足地上部分生长需求,这表现了白刺灌丛不同生长阶段为环境适应而做出的动态调整。白刺灌丛的不定根系中未发现有4级根的出现,该结果的产生与土壤容重有关。不定根系随灌丛发育多向深层土壤生长,而深层土壤较高的紧实度对根系生长形成阻力,不利于根系的扩张,因此影响了不定根生长与分支能力[30 ] 。
根系的形态受遗传和外界环境两方面因素影响,为适应不同环境下的水分、养分条件,会在空间分布上表现出一定的自适应性[31 ] 。本试验发现,白刺灌丛的不定根系的空间分布以垂直根为主,倾斜根其次,水平根最少,表现出深根型特征。这一现象主要受研究区气候条件影响。干旱少雨、日照充足、土壤水分蒸发量大,正是研究区这些恶劣的气候特征导致表层土壤含水率极低。因此白刺灌丛为获得更多水分,不定根更多地向含水率更高的深层土壤生长[29 ] 。这种空间分布特征也表现了白刺灌丛为适宜干旱环境而做出的重要生存策略。也有研究表明倾斜根可以拥有最好的固土能力[32 ] ,因此,灌丛沙堆内部大量倾斜不定根系对沙堆土壤有很强的稳固作用。
3.2 不同生长阶段白刺灌丛及各级不定根的生物量分配模式
植物为了适应周围生态环境,通过调节不同器官生物量分配来达到对资源利用的最大化,而灌丛的不同生长阶段也会因为其自身生长特性表现出不同的分配方式[8 ] 。本研究中,不同生长阶段白刺灌丛的根冠比随植物的生长发育而不断降低,不定根系的生长速率也表现为持续下降,生长关系表现为异速生长。该结论与肖遥等[8 ] 的研究结论相似。不同时期的植物相关生长是动态变化的,且变化趋势也不同。由于西鄂尔多斯地区干旱少雨、土壤贫瘠,特殊的气候条件致使这里植物地下部分生物量占比更高[33 ] ,而雏形阶段白刺灌丛拥有最高的根冠比与研究区的气候因素有很大关系。处于生长初期的雏形阶段灌丛为适应干旱贫瘠的环境,在生存决策上选择在根径更细、分支结构更简单等方面调整以适应环境,生物量分配上同样集中于根系部分,通过增加根系生长速率,以此扩大对土壤水分、养分等资源的利用范围,进而提高对恶劣生存环境的适应能力。而发育阶段和成熟阶段灌丛在不断生长发育的过程中通过根系分泌、枝系拦截等方式改变了土壤微环境[34 ] ,逐渐适应研究区干旱贫瘠的环境后,灌丛为获得更高的光照资源在生态策略上选择降低根系部分生长速率,将更多生物量分配于地上部分,进而提高争夺空间和光照面积的能力,以满足自身生长、繁殖需要[35 ] 。
灌丛根系是固定植物以及吸收利用荒漠环境中养分、水分的重要器官,同时也是地上部分正常生长发育的基本保障[36 ] 。研究灌丛根系生物量可以了解灌丛的生产潜力,揭示灌丛对该区域自然资源的利用能力与效率。不同径级根系的功能不同[37 ] ,粗根用于支撑固定植物和贮存营养物质[36 ] ,细根负责养分与水分的吸收和运输[38 ] 。因此,除根冠比和相对生长关系外,各级根系生物量占比也可反映植物对环境的适应策略。由于灌丛的根径大小随生长阶段变化也表现明显差异,因此本文未采用绝对根径划分灌丛的粗根与细根,而是依据灌丛的径级将其区分为粗根与细根,其中3级根代表粗根,1级根、2级根代表细根。本试验结果显示,灌丛生长发育过程中分支率不断增加,灌丛细根不断生长,其生物量占比不断提高。雏形阶段灌丛为更好地固定灌丛沙堆以及保存营养适应贫瘠土壤,不定根系多为粗根,而发育后期灌丛逐渐稳定,因此将更多生物量分配到细根以求提高资源吸收利用范围及效率。这表明灌丛随生长发育在生态策略上会做出调整。
4 结论
成熟阶段白刺灌丛的1级、2级和3级不定根根径分别较发育阶段、雏形阶段灌丛增加79.66%和120.83%、86.05%和95.12%、37.36%和47.93%。不定根系以垂直分布为主,为深根型特征,有利于利用深层水分。
不同生长阶段白刺灌丛不定根系构型均为人字形,且随生长发育分支逐渐增多。雏形阶段灌丛qa =0.95,qb =0.52,TI =0.98;发育阶段灌丛qa =0.69,qb =0.46,TI =0.87;成熟阶段灌丛qa =0.63,qb =0.63,TI =0.85。不定根总分支率随灌丛生长而逐渐增大,雏形阶段灌丛显著小于发育阶段和成熟阶段灌丛,分别小8.96%和30.05%。白刺逐渐增加分支结构有利于扩大对资源的吸收利用范围。
不同生长阶段的白刺灌丛生物量分配表现为根冠比和根系生长速率随灌丛发育而逐渐降低,粗根占比减少,细根占比提高。生长初期的灌丛将更多生物量放到不定根系及粗根来固定灌丛以及储存营养来适应胁迫环境,而生长后期的灌丛降低根系部分生长速度,将更多生物量分配到地上部分以提高光能资源的利用。
参考文献
View Option
[1]
Dannowski M Block A Fractal geometry and root system structures of heterogeneous plant communities
[J].Plant and Soil ,2005 ,272 (1/2 ):61 -76 .
[本文引用: 1]
[2]
Lynch J Root architecture and plant productivity
[J].Plant Physiology ,1995 ,109 (1 ):7 -13 .
[本文引用: 1]
[3]
Savinov I A Architectural analysis of representatives of the Celastrales Order:structure and rhythm of shoot development in connection with adaptations of species to different environmental conditions
[J].Contemporary Problems of Ecology ,2020 ,13 (3 ):300 -308 .
[本文引用: 1]
[4]
Odum E P Barrett G W Fundamentals of Ecology [M].Philadelphia,USA :Saunders ,1971 .
[本文引用: 1]
[5]
孙儒泳 ,李庆芬 ,牛翠娟 ,等 基础生态学 [M].北京 :高等教育出版社 ,2002 .
[本文引用: 1]
[6]
单立山 ,李毅 ,董秋莲 ,等 红砂根系构型对干旱的生态适应
[J].中国沙漠 ,2012 ,32 (5 ):1283 -1290 .
[本文引用: 2]
[7]
杨小林 ,张希明 ,李义玲 ,等 塔克拉玛干沙漠腹地3种植物根系构型及其生境适应策略
[J].植物生态学报 ,2008 ,32 (6 ):1268 -1276 .
[本文引用: 2]
[8]
肖遥 ,陶冶 ,张元明 古尔班通古特沙漠4种荒漠草本植物不同生长期的生物量分配与叶片化学计量特征
[J].植物生态学报 ,2014 ,38 (9 ):929 -940 .
[本文引用: 3]
[9]
徐道龙 西鄂尔多斯荒漠6种珍稀植物菌根及共生微生物多样性研究
[D].呼和浩特 :内蒙古大学 ,2021 .
[本文引用: 1]
[10]
Li J Zhao Y Liu H et al Sandy desertification cycles in the southwestern Mu Us Desert in China over the past 80 years recorded based on nebkha sediments
[J].Aeolian Research ,2016 ,20 :100 -107 .
[本文引用: 1]
[11]
常兆丰 ,张进虎 ,石学刚 ,等 沙漠植物分层侧影与积沙成丘的关系
[J].生态学报 ,2017 ,37 (21 ):7351 -7358 .
[本文引用: 1]
[12]
党晓宏 ,蒙仲举 ,高永 ,等 西鄂尔多斯地区5种荒漠灌丛光合固碳能力研究
[J].干旱区资源与环境 ,2017 ,31 (11 ):128 -135 .
[本文引用: 1]
[13]
张萍 ,康经理 ,袁瑱 ,等 两类植物型沙丘上植物群落的异同及其对沙丘形态的响应
[J].生态学报 ,2017 ,37 (23 ):7920 -7927 .
[本文引用: 1]
[14]
靳虎甲 ,马全林 ,何明珠 ,等 石羊河下游白刺灌丛演替过程中群落结构及数量特征
[J].生态学报 ,2013 ,33 (7 ):2248 -2259 .
[本文引用: 1]
[15]
Steingraeber D A Waller D M Non-stationarity of tree branching patterns and bifurcation ratios
[J].Proceedings of the Royal Society of London.Series B ,Biological Sciences,1986 ,228 (1251 ):187 -194 .
[本文引用: 2]
[16]
Berntson G M The characterization of topology:a comparison of four topological indices for rooted binary trees
[J].Journal of Theoretical Biology ,1995 ,177 (3 ):271 -281 .
[本文引用: 2]
[17]
高广磊 ,丁国栋 ,赵媛媛 ,等 生物结皮发育对毛乌素沙地土壤粒度特征的影响
[J].农业机械学报 ,2014 ,45 (1 ):115 -120 .
[本文引用: 1]
[18]
Fitter A H An architectural approach to the comparative ecology of plant root systems
[J].New Phytologist ,1987 ,106 (1 ):61 -77 .
[本文引用: 2]
[19]
Bouma T J Nielsen K L Van Hal J et al Root system topology and diameter distribution of species from habitats differing in inundation frequency
[J].Functional Ecology ,2001 ,15 (3 ):360 -369 .
[本文引用: 1]
[20]
Oppelt A L Kurth W Godbold D L Topology,scaling relations and Leonardo's rule in root systems from African tree species
[J].Tree Physiology ,2001 ,21 (2/3 ):117 -128 .
[本文引用: 1]
[21]
Niklas K J Modelling below-and above-ground biomass for non-woody and woody plants
[J].Annals of Botany ,2005 ,95 (2 ):315 -321 .
[本文引用: 1]
[22]
Nquist B J Niklas K J Global allocation rules for patterns of biomass partitioning in seed plants
[J].Science ,2002 ,295 (5559 ):1517 -1520 .
[本文引用: 1]
[23]
张扬 ,李程远 ,韩少杰 ,等 典型黑土区主要树种根系构型特征及其对固土能力的影响
[J].应用生态学报 ,2021 ,32 (5 ):1726 -1734 .
[本文引用: 1]
[24]
马献发 ,宋凤斌 ,张继舟 根系对土壤环境胁迫响应的研究进展
[J].中国农学通报 ,2011 ,27 (5 ):44 -48 .
[本文引用: 1]
[25]
Angela H Graziella B Claude D et al Plant root growth,architecture and function
[J].Plant and Soil ,2009 ,321 (1/2 ):153 -187 .
[本文引用: 1]
[26]
吴静 ,盛茂银 ,肖海龙 ,等 西南喀斯特石漠化环境适生植物细根构型及其与细根和根际土壤养分计量特征的相关性
[J].生态学报 ,2022 ,42 (2 ):677 -687 .
[本文引用: 1]
[27]
李尝君 ,郭京衡 ,曾凡江 ,等 多枝柽柳(Tamarix ramosissima )根、冠构型的年龄差异及其适应意义
[J].中国沙漠 ,2015 ,35 (2 ):365 -372 .
[本文引用: 1]
[28]
班芷桦 ,王琼 喜旱莲子草和接骨草竞争对模拟增温的响应
[J].植物生态学报 ,2015 ,39 (1 ):43 -51 .
[本文引用: 1]
[29]
单立山 ,李毅 ,任伟 ,等 河西走廊中部两种荒漠植物根系构型特征
[J].应用生态学报 ,2013 ,24 (1 ):25 -31 .
[本文引用: 2]
[30]
木巴热克·阿尤普 ,陈亚宁 ,李卫红 ,等 极端干旱环境下的胡杨细根分布与土壤特征
[J].中国沙漠 ,2011 ,31 (6 ):1449 -1458 .
[本文引用: 1]
[31]
Meyer K M Ward D Wiegand K et al Multi-proxy evidence for competition between savanna woody species
[J].Perspectives in Plant Ecology,Evolution and Systematics ,2007 ,10 (1 ):63 -72 .
[本文引用: 1]
[32]
李珍玉 ,欧阳淼 ,肖宏彬 ,等 基于根系构型的调控提高植物边坡根系固土能力
[J].岩土力学 ,2021 ,(12 ):1 -11 .
[本文引用: 1]
[33]
党晓宏 ,高永 ,蒙仲举 ,等 西鄂尔多斯地区5种荒漠优势灌丛生物量分配格局及预测模型
[J].中国沙漠 ,2017 ,37 (1 ):100 -108 .
[本文引用: 1]
[34]
杜建会 ,严平 ,董玉祥 干旱地区灌丛沙堆研究现状与展望
[J].地理学报 ,2010 ,65 (3 ):339 -350 .
[本文引用: 1]
[35]
孙越 ,何怀江 ,李良 ,等 阔叶红松林下6种早夏草本不同生长期生物量分配及模型构建
[J].生态学报 ,2017 ,37 (19 ):6523 -6533 .
[本文引用: 1]
[36]
仇瑶 ,常顺利 ,张毓涛 ,等 天山林区六种灌木生物量的建模及其器官分配的适应性
[J].生态学报 ,2015 ,35 (23 ):7842 -7851 .
[本文引用: 2]
[37]
刘瑞 ,靳虎甲 ,马全林 ,等 甘肃景电灌区不同栽植年限枸杞生物量分配特征
[J].生态学杂志 ,2012 ,31 (10 ):2493 -2500 .
[本文引用: 1]
[38]
王向荣 ,王政权 ,韩有志 ,等 水曲柳和落叶松不同根序之间细根直径的变异研究
[J].植物生态学报 ,2005 ,29 (6 ):5 -11 .
[本文引用: 1]
Fractal geometry and root system structures of heterogeneous plant communities
1
2005
... 根系是植物与外界环境进行物质交换的重要器官[1 ] ,根系在土壤中的分布和排列方式被称为根系构型[2 ] ,构型特征随土壤环境变化会发生动态调整,这是植物对异质性土壤资源的反馈机制和适应策略[3 ] .生物量是衡量个体在生态系统中功能与地位的重要指标[4 ] ,植物的根系生物量所占比例反映了植物为适应环境以求对光、水、养分等资源最大吸收利用所做出的适应[5 ] .处于不同生长阶段的植物会表现出不同的根系构型和生物量分配模式以适应环境[6 -8 ] .因此,探究植物的根系构型和生物量分配模式在胁迫环境中的适应机制,对于干旱区的荒漠化防治及植物多样性保护具有重要意义. ...
Root architecture and plant productivity
1
1995
... 根系是植物与外界环境进行物质交换的重要器官[1 ] ,根系在土壤中的分布和排列方式被称为根系构型[2 ] ,构型特征随土壤环境变化会发生动态调整,这是植物对异质性土壤资源的反馈机制和适应策略[3 ] .生物量是衡量个体在生态系统中功能与地位的重要指标[4 ] ,植物的根系生物量所占比例反映了植物为适应环境以求对光、水、养分等资源最大吸收利用所做出的适应[5 ] .处于不同生长阶段的植物会表现出不同的根系构型和生物量分配模式以适应环境[6 -8 ] .因此,探究植物的根系构型和生物量分配模式在胁迫环境中的适应机制,对于干旱区的荒漠化防治及植物多样性保护具有重要意义. ...
Architectural analysis of representatives of the Celastrales Order:structure and rhythm of shoot development in connection with adaptations of species to different environmental conditions
1
2020
... 根系是植物与外界环境进行物质交换的重要器官[1 ] ,根系在土壤中的分布和排列方式被称为根系构型[2 ] ,构型特征随土壤环境变化会发生动态调整,这是植物对异质性土壤资源的反馈机制和适应策略[3 ] .生物量是衡量个体在生态系统中功能与地位的重要指标[4 ] ,植物的根系生物量所占比例反映了植物为适应环境以求对光、水、养分等资源最大吸收利用所做出的适应[5 ] .处于不同生长阶段的植物会表现出不同的根系构型和生物量分配模式以适应环境[6 -8 ] .因此,探究植物的根系构型和生物量分配模式在胁迫环境中的适应机制,对于干旱区的荒漠化防治及植物多样性保护具有重要意义. ...
1
1971
... 根系是植物与外界环境进行物质交换的重要器官[1 ] ,根系在土壤中的分布和排列方式被称为根系构型[2 ] ,构型特征随土壤环境变化会发生动态调整,这是植物对异质性土壤资源的反馈机制和适应策略[3 ] .生物量是衡量个体在生态系统中功能与地位的重要指标[4 ] ,植物的根系生物量所占比例反映了植物为适应环境以求对光、水、养分等资源最大吸收利用所做出的适应[5 ] .处于不同生长阶段的植物会表现出不同的根系构型和生物量分配模式以适应环境[6 -8 ] .因此,探究植物的根系构型和生物量分配模式在胁迫环境中的适应机制,对于干旱区的荒漠化防治及植物多样性保护具有重要意义. ...
1
2002
... 根系是植物与外界环境进行物质交换的重要器官[1 ] ,根系在土壤中的分布和排列方式被称为根系构型[2 ] ,构型特征随土壤环境变化会发生动态调整,这是植物对异质性土壤资源的反馈机制和适应策略[3 ] .生物量是衡量个体在生态系统中功能与地位的重要指标[4 ] ,植物的根系生物量所占比例反映了植物为适应环境以求对光、水、养分等资源最大吸收利用所做出的适应[5 ] .处于不同生长阶段的植物会表现出不同的根系构型和生物量分配模式以适应环境[6 -8 ] .因此,探究植物的根系构型和生物量分配模式在胁迫环境中的适应机制,对于干旱区的荒漠化防治及植物多样性保护具有重要意义. ...
红砂根系构型对干旱的生态适应
2
2012
... 根系是植物与外界环境进行物质交换的重要器官[1 ] ,根系在土壤中的分布和排列方式被称为根系构型[2 ] ,构型特征随土壤环境变化会发生动态调整,这是植物对异质性土壤资源的反馈机制和适应策略[3 ] .生物量是衡量个体在生态系统中功能与地位的重要指标[4 ] ,植物的根系生物量所占比例反映了植物为适应环境以求对光、水、养分等资源最大吸收利用所做出的适应[5 ] .处于不同生长阶段的植物会表现出不同的根系构型和生物量分配模式以适应环境[6 -8 ] .因此,探究植物的根系构型和生物量分配模式在胁迫环境中的适应机制,对于干旱区的荒漠化防治及植物多样性保护具有重要意义. ...
... 植物会随着环境的不断变化对其根系构型做出调整,以期达到对资源的最大利用[28 ] .前人研究发现,植物的根系分支率与环境干旱胁迫程度负相关[6 ] .在等量碳投入条件下,虽然人字形分支分布范围更小,不利于养分占有,但分支结构简单,对资源吸收利用能力更强,更适应胁迫环境[7 ,18 ] .本研究中随灌丛生长发育其拓扑指数参数显著降低,成熟阶段灌丛的分支率显著高于发育阶段和雏形阶段灌丛,不同生长阶段白刺灌丛不定根系构型均表现为人字形,该结论与单立山等[29 ] 对地下根系的研究结论不同.造成该结论差异的原因可能与不同地区资源差异有关,也可能是由于不定根系和地下根系自身特性差异有关,有待进一步研究.本试验的结果表明了研究区的白刺灌丛随生长发育抗干旱胁迫能力不断增强,成熟阶段灌丛不同于发育阶段和雏形阶段灌丛的将更多精力放在对环境适应上,而是有余力通过增加不定根系分支率来扩张其对资源的吸收利用范围以满足地上部分生长需求,这表现了白刺灌丛不同生长阶段为环境适应而做出的动态调整.白刺灌丛的不定根系中未发现有4级根的出现,该结果的产生与土壤容重有关.不定根系随灌丛发育多向深层土壤生长,而深层土壤较高的紧实度对根系生长形成阻力,不利于根系的扩张,因此影响了不定根生长与分支能力[30 ] . ...
塔克拉玛干沙漠腹地3种植物根系构型及其生境适应策略
2
2008
... 根系构型特征随不同的水分、养分条件产生变化,具有一定可塑性[24 ] .根系构型差异影响根系对土壤水分的获取能力以及碳的消耗[25 ] 与分配[26 ] .灌丛的根径与其寿命正相关,但较大的根径会消耗更多的碳,而干旱胁迫环境下灌丛根系的高死亡率会导致大量碳损失,不利于灌丛的生存[27 ] .因此,生长阶段初期的雏形阶段灌丛为适应干旱环境,选择降低根径以减少根系死亡所带来的碳消耗.而成熟阶段灌丛的根径显著大于雏形阶段和发育阶段灌丛也反映了随着灌丛的不断生长其抗干旱胁迫能力不断增强,有能力适应恶劣环境后选择寿命更长的较粗不定根.1—2级根径比小于2—3级,说明了随根径级数增加根径逐渐变小,但不同生长阶段的白刺灌丛不定根系根径比均未表现出显著差异.各级根系分支前后的一致性反映了根系的自相似性,因此可通过已测数据对部分构型指标进行估算,从而获得相关数据.这是对植物构型研究及分析的有效方法,不仅降低了工作难度,并且减少了对植物破坏[7 ] . ...
... 植物会随着环境的不断变化对其根系构型做出调整,以期达到对资源的最大利用[28 ] .前人研究发现,植物的根系分支率与环境干旱胁迫程度负相关[6 ] .在等量碳投入条件下,虽然人字形分支分布范围更小,不利于养分占有,但分支结构简单,对资源吸收利用能力更强,更适应胁迫环境[7 ,18 ] .本研究中随灌丛生长发育其拓扑指数参数显著降低,成熟阶段灌丛的分支率显著高于发育阶段和雏形阶段灌丛,不同生长阶段白刺灌丛不定根系构型均表现为人字形,该结论与单立山等[29 ] 对地下根系的研究结论不同.造成该结论差异的原因可能与不同地区资源差异有关,也可能是由于不定根系和地下根系自身特性差异有关,有待进一步研究.本试验的结果表明了研究区的白刺灌丛随生长发育抗干旱胁迫能力不断增强,成熟阶段灌丛不同于发育阶段和雏形阶段灌丛的将更多精力放在对环境适应上,而是有余力通过增加不定根系分支率来扩张其对资源的吸收利用范围以满足地上部分生长需求,这表现了白刺灌丛不同生长阶段为环境适应而做出的动态调整.白刺灌丛的不定根系中未发现有4级根的出现,该结果的产生与土壤容重有关.不定根系随灌丛发育多向深层土壤生长,而深层土壤较高的紧实度对根系生长形成阻力,不利于根系的扩张,因此影响了不定根生长与分支能力[30 ] . ...
古尔班通古特沙漠4种荒漠草本植物不同生长期的生物量分配与叶片化学计量特征
3
2014
... 根系是植物与外界环境进行物质交换的重要器官[1 ] ,根系在土壤中的分布和排列方式被称为根系构型[2 ] ,构型特征随土壤环境变化会发生动态调整,这是植物对异质性土壤资源的反馈机制和适应策略[3 ] .生物量是衡量个体在生态系统中功能与地位的重要指标[4 ] ,植物的根系生物量所占比例反映了植物为适应环境以求对光、水、养分等资源最大吸收利用所做出的适应[5 ] .处于不同生长阶段的植物会表现出不同的根系构型和生物量分配模式以适应环境[6 -8 ] .因此,探究植物的根系构型和生物量分配模式在胁迫环境中的适应机制,对于干旱区的荒漠化防治及植物多样性保护具有重要意义. ...
... 植物为了适应周围生态环境,通过调节不同器官生物量分配来达到对资源利用的最大化,而灌丛的不同生长阶段也会因为其自身生长特性表现出不同的分配方式[8 ] .本研究中,不同生长阶段白刺灌丛的根冠比随植物的生长发育而不断降低,不定根系的生长速率也表现为持续下降,生长关系表现为异速生长.该结论与肖遥等[8 ] 的研究结论相似.不同时期的植物相关生长是动态变化的,且变化趋势也不同.由于西鄂尔多斯地区干旱少雨、土壤贫瘠,特殊的气候条件致使这里植物地下部分生物量占比更高[33 ] ,而雏形阶段白刺灌丛拥有最高的根冠比与研究区的气候因素有很大关系.处于生长初期的雏形阶段灌丛为适应干旱贫瘠的环境,在生存决策上选择在根径更细、分支结构更简单等方面调整以适应环境,生物量分配上同样集中于根系部分,通过增加根系生长速率,以此扩大对土壤水分、养分等资源的利用范围,进而提高对恶劣生存环境的适应能力.而发育阶段和成熟阶段灌丛在不断生长发育的过程中通过根系分泌、枝系拦截等方式改变了土壤微环境[34 ] ,逐渐适应研究区干旱贫瘠的环境后,灌丛为获得更高的光照资源在生态策略上选择降低根系部分生长速率,将更多生物量分配于地上部分,进而提高争夺空间和光照面积的能力,以满足自身生长、繁殖需要[35 ] . ...
... [8 ]的研究结论相似.不同时期的植物相关生长是动态变化的,且变化趋势也不同.由于西鄂尔多斯地区干旱少雨、土壤贫瘠,特殊的气候条件致使这里植物地下部分生物量占比更高[33 ] ,而雏形阶段白刺灌丛拥有最高的根冠比与研究区的气候因素有很大关系.处于生长初期的雏形阶段灌丛为适应干旱贫瘠的环境,在生存决策上选择在根径更细、分支结构更简单等方面调整以适应环境,生物量分配上同样集中于根系部分,通过增加根系生长速率,以此扩大对土壤水分、养分等资源的利用范围,进而提高对恶劣生存环境的适应能力.而发育阶段和成熟阶段灌丛在不断生长发育的过程中通过根系分泌、枝系拦截等方式改变了土壤微环境[34 ] ,逐渐适应研究区干旱贫瘠的环境后,灌丛为获得更高的光照资源在生态策略上选择降低根系部分生长速率,将更多生物量分配于地上部分,进而提高争夺空间和光照面积的能力,以满足自身生长、繁殖需要[35 ] . ...
西鄂尔多斯荒漠6种珍稀植物菌根及共生微生物多样性研究
1
2021
... 西鄂尔多斯国家自然保护区位于鄂尔多斯高原西缘,独特的气候条件和地形地貌使这里成为了许多古老孑遗植物的“避难所”[9 ] .白刺(Nitraria tangutorum )作为研究区主要建群种,因其耐旱耐瘠薄的特性为研究区生态修复发挥着重要作用.白刺灌丛易形成沙堆,沙堆沉积物包含了大量环境信息,对于描述区域土壤退化和风蚀情况有重要意义[10 ] .对沙堆调查后发现其内部长有大量不定根,并且不同生长阶段的不定根构型和生物量分配表现出显著差异.前人研究发现沙堆的形成和发育与不定根生长有着密切联系[11 ] ,但目前缺少不定根系构型以及生物量分配模式的研究,不定根随灌丛生长和沙堆发育会做出怎样适应策略也不明晰. ...
Sandy desertification cycles in the southwestern Mu Us Desert in China over the past 80 years recorded based on nebkha sediments
1
2016
... 西鄂尔多斯国家自然保护区位于鄂尔多斯高原西缘,独特的气候条件和地形地貌使这里成为了许多古老孑遗植物的“避难所”[9 ] .白刺(Nitraria tangutorum )作为研究区主要建群种,因其耐旱耐瘠薄的特性为研究区生态修复发挥着重要作用.白刺灌丛易形成沙堆,沙堆沉积物包含了大量环境信息,对于描述区域土壤退化和风蚀情况有重要意义[10 ] .对沙堆调查后发现其内部长有大量不定根,并且不同生长阶段的不定根构型和生物量分配表现出显著差异.前人研究发现沙堆的形成和发育与不定根生长有着密切联系[11 ] ,但目前缺少不定根系构型以及生物量分配模式的研究,不定根随灌丛生长和沙堆发育会做出怎样适应策略也不明晰. ...
沙漠植物分层侧影与积沙成丘的关系
1
2017
... 西鄂尔多斯国家自然保护区位于鄂尔多斯高原西缘,独特的气候条件和地形地貌使这里成为了许多古老孑遗植物的“避难所”[9 ] .白刺(Nitraria tangutorum )作为研究区主要建群种,因其耐旱耐瘠薄的特性为研究区生态修复发挥着重要作用.白刺灌丛易形成沙堆,沙堆沉积物包含了大量环境信息,对于描述区域土壤退化和风蚀情况有重要意义[10 ] .对沙堆调查后发现其内部长有大量不定根,并且不同生长阶段的不定根构型和生物量分配表现出显著差异.前人研究发现沙堆的形成和发育与不定根生长有着密切联系[11 ] ,但目前缺少不定根系构型以及生物量分配模式的研究,不定根随灌丛生长和沙堆发育会做出怎样适应策略也不明晰. ...
西鄂尔多斯地区5种荒漠灌丛光合固碳能力研究
1
2017
... 西鄂尔多斯国家级自然保护区(39°13′—40°10′N,106°44′—107°43′E)属暖温带大陆性气候区,年均气温7.8—8.1 ℃,年降水量162—172 mm,集中在6—8月,年潜在蒸发量2 470—3 481 mm.研究区以偏西北风为主,年均风速3.2 m·s-1 ,最大风速可达24.2 m·s-1 .土壤类型主要为棕钙土、灰漠土,具有土层浅薄、土质粗糙、养分贫瘠、腐殖质含量低等特点[12 ] .研究区是强旱生灌丛为建群种的荒漠化草原,优势种为白刺、沙冬青(Ammopiptanthus mongolicus )、霸王(Zygophyllum xanthoxylon )、红砂(Reaumuria soongarica )、半日花(Helianthemum songaricum ),伴生植物有绵刺(Potaninia mongolica )、珍珠猪毛菜(Salsola passerina )等,并伴有一定数量的草本植物.植物所需水分多来自天然降水,地下水埋深大于15 m[13 ] . ...
两类植物型沙丘上植物群落的异同及其对沙丘形态的响应
1
2017
... 西鄂尔多斯国家级自然保护区(39°13′—40°10′N,106°44′—107°43′E)属暖温带大陆性气候区,年均气温7.8—8.1 ℃,年降水量162—172 mm,集中在6—8月,年潜在蒸发量2 470—3 481 mm.研究区以偏西北风为主,年均风速3.2 m·s-1 ,最大风速可达24.2 m·s-1 .土壤类型主要为棕钙土、灰漠土,具有土层浅薄、土质粗糙、养分贫瘠、腐殖质含量低等特点[12 ] .研究区是强旱生灌丛为建群种的荒漠化草原,优势种为白刺、沙冬青(Ammopiptanthus mongolicus )、霸王(Zygophyllum xanthoxylon )、红砂(Reaumuria soongarica )、半日花(Helianthemum songaricum ),伴生植物有绵刺(Potaninia mongolica )、珍珠猪毛菜(Salsola passerina )等,并伴有一定数量的草本植物.植物所需水分多来自天然降水,地下水埋深大于15 m[13 ] . ...
石羊河下游白刺灌丛演替过程中群落结构及数量特征
1
2013
... 野外试验时间为2021年8月中旬,在研究区设置一个100 m×100 m的样方,样方内以白刺灌丛为主,并少量分布植被盖度10%—30%的其他小型沙生植物,沙生植物分布于白刺灌丛沙堆四周,未对沙堆内部的不定根生长产生影响(图1 ).采用“空间代替时间”的方法,结合相关文献与样地实际情况[14 ] ,选择研究区内具有代表性的白刺灌丛为研究对象.将白刺灌丛分为雏形、发育和成熟3个生长阶段,每阶段各3丛,共9丛. ...
Non-stationarity of tree branching patterns and bifurcation ratios
2
1986
... 采用Strahler法对不定根系进行分级[15 -16 ] ,由外及内确定根序,最外根系为1级根,两个1级根系相遇为2级根,以此类推(图2 ).若不同级根系交汇取较高级者. ...
... 根系总分支率(OBRr ):根据Strahler分支等级[15 -16 ] 计算每个等级(i )根的数量Ni ,分别以根级i 和lgNi 为横、纵坐标作图,回归直线的斜率绝对值的逆对数即为根系总分支率.逐级分支率(SBR (i :i +1))为i 级分支数与前一级的分支数之比,即: ...
The characterization of topology:a comparison of four topological indices for rooted binary trees
2
1995
... 采用Strahler法对不定根系进行分级[15 -16 ] ,由外及内确定根序,最外根系为1级根,两个1级根系相遇为2级根,以此类推(图2 ).若不同级根系交汇取较高级者. ...
... 根系总分支率(OBRr ):根据Strahler分支等级[15 -16 ] 计算每个等级(i )根的数量Ni ,分别以根级i 和lgNi 为横、纵坐标作图,回归直线的斜率绝对值的逆对数即为根系总分支率.逐级分支率(SBR (i :i +1))为i 级分支数与前一级的分支数之比,即: ...
生物结皮发育对毛乌素沙地土壤粒度特征的影响
1
2014
... 由于灌丛沙堆呈椭圆形.因此,其底面积(S d )及沙堆体积(V d )[17 ] 计算公式如下: ...
An architectural approach to the comparative ecology of plant root systems
2
1987
... Fitter[18 ] 和Bouma等[19 ] 提出根系分支存在人字形分支和叉状分支两种极端模式(图3 ),并提出拓扑指数表达式: ...
... 植物会随着环境的不断变化对其根系构型做出调整,以期达到对资源的最大利用[28 ] .前人研究发现,植物的根系分支率与环境干旱胁迫程度负相关[6 ] .在等量碳投入条件下,虽然人字形分支分布范围更小,不利于养分占有,但分支结构简单,对资源吸收利用能力更强,更适应胁迫环境[7 ,18 ] .本研究中随灌丛生长发育其拓扑指数参数显著降低,成熟阶段灌丛的分支率显著高于发育阶段和雏形阶段灌丛,不同生长阶段白刺灌丛不定根系构型均表现为人字形,该结论与单立山等[29 ] 对地下根系的研究结论不同.造成该结论差异的原因可能与不同地区资源差异有关,也可能是由于不定根系和地下根系自身特性差异有关,有待进一步研究.本试验的结果表明了研究区的白刺灌丛随生长发育抗干旱胁迫能力不断增强,成熟阶段灌丛不同于发育阶段和雏形阶段灌丛的将更多精力放在对环境适应上,而是有余力通过增加不定根系分支率来扩张其对资源的吸收利用范围以满足地上部分生长需求,这表现了白刺灌丛不同生长阶段为环境适应而做出的动态调整.白刺灌丛的不定根系中未发现有4级根的出现,该结果的产生与土壤容重有关.不定根系随灌丛发育多向深层土壤生长,而深层土壤较高的紧实度对根系生长形成阻力,不利于根系的扩张,因此影响了不定根生长与分支能力[30 ] . ...
Root system topology and diameter distribution of species from habitats differing in inundation frequency
1
2001
... Fitter[18 ] 和Bouma等[19 ] 提出根系分支存在人字形分支和叉状分支两种极端模式(图3 ),并提出拓扑指数表达式: ...
Topology,scaling relations and Leonardo's rule in root systems from African tree species
1
2001
... 式中:TI 为拓扑指数;A 为最长根系通道内部连接总数;M 为所有外部根系连接总数.TI 越接近1表明A 和M 值相近,根系分支少,形状接近人字形分支结构;TI 越接近0.5表明根系结构越接近叉状分支.Oppelt等[20 ] 对拓扑指数构型进行了进一步研究,并提出新的拓扑参数.其表达式为: ...
Modelling below-and above-ground biomass for non-woody and woody plants
1
2005
... 利用异速生长机制模型对白刺灌丛各生长阶段AGB 与BGB 进行相关生长分析[21 ] . ...
Global allocation rules for patterns of biomass partitioning in seed plants
1
2002
... 式中:Y 为BGB ;X 为AGB ;β 为常数;α 为相关生长指数.当α =1时,BGB 和AGB 为等速生长;当α ≠1时,为异速生长[22 ] .判断α 是否等于1,利用α 的95%置信区间是否包含1来确定. ...
典型黑土区主要树种根系构型特征及其对固土能力的影响
1
2021
... 不同生长阶段白刺灌丛的地上部分和不定根系的生长关系均表现为异速生长(表3 ).从相关生长指数的均值和95%置信区间可以看出,不同生长阶段的白刺灌丛相关生长指数均小于1,说明白刺灌丛地上部分生长速度始终大于地下不定根生长速度.从不同生长阶段来看,随着灌丛生长发育,不定根系生长速率不断提高,雏形阶段灌丛相关生长指数较发育阶段和成熟阶段分别低280.00%和190.00%.这可能与不定根系在沙堆内部独特的生长环境有关,灌丛枝系被沙埋后逐渐长出不定根系,而随着灌丛地上部分的不断生长,枝系拦截风沙流能力不断提高,沙堆形态逐渐增大,因此被埋枝系不断增多,致使不定根系增长速率增加,而不定根生长可固定沙堆促使其继续增大[23 ] . ...
根系对土壤环境胁迫响应的研究进展
1
2011
... 根系构型特征随不同的水分、养分条件产生变化,具有一定可塑性[24 ] .根系构型差异影响根系对土壤水分的获取能力以及碳的消耗[25 ] 与分配[26 ] .灌丛的根径与其寿命正相关,但较大的根径会消耗更多的碳,而干旱胁迫环境下灌丛根系的高死亡率会导致大量碳损失,不利于灌丛的生存[27 ] .因此,生长阶段初期的雏形阶段灌丛为适应干旱环境,选择降低根径以减少根系死亡所带来的碳消耗.而成熟阶段灌丛的根径显著大于雏形阶段和发育阶段灌丛也反映了随着灌丛的不断生长其抗干旱胁迫能力不断增强,有能力适应恶劣环境后选择寿命更长的较粗不定根.1—2级根径比小于2—3级,说明了随根径级数增加根径逐渐变小,但不同生长阶段的白刺灌丛不定根系根径比均未表现出显著差异.各级根系分支前后的一致性反映了根系的自相似性,因此可通过已测数据对部分构型指标进行估算,从而获得相关数据.这是对植物构型研究及分析的有效方法,不仅降低了工作难度,并且减少了对植物破坏[7 ] . ...
Plant root growth,architecture and function
1
2009
... 根系构型特征随不同的水分、养分条件产生变化,具有一定可塑性[24 ] .根系构型差异影响根系对土壤水分的获取能力以及碳的消耗[25 ] 与分配[26 ] .灌丛的根径与其寿命正相关,但较大的根径会消耗更多的碳,而干旱胁迫环境下灌丛根系的高死亡率会导致大量碳损失,不利于灌丛的生存[27 ] .因此,生长阶段初期的雏形阶段灌丛为适应干旱环境,选择降低根径以减少根系死亡所带来的碳消耗.而成熟阶段灌丛的根径显著大于雏形阶段和发育阶段灌丛也反映了随着灌丛的不断生长其抗干旱胁迫能力不断增强,有能力适应恶劣环境后选择寿命更长的较粗不定根.1—2级根径比小于2—3级,说明了随根径级数增加根径逐渐变小,但不同生长阶段的白刺灌丛不定根系根径比均未表现出显著差异.各级根系分支前后的一致性反映了根系的自相似性,因此可通过已测数据对部分构型指标进行估算,从而获得相关数据.这是对植物构型研究及分析的有效方法,不仅降低了工作难度,并且减少了对植物破坏[7 ] . ...
西南喀斯特石漠化环境适生植物细根构型及其与细根和根际土壤养分计量特征的相关性
1
2022
... 根系构型特征随不同的水分、养分条件产生变化,具有一定可塑性[24 ] .根系构型差异影响根系对土壤水分的获取能力以及碳的消耗[25 ] 与分配[26 ] .灌丛的根径与其寿命正相关,但较大的根径会消耗更多的碳,而干旱胁迫环境下灌丛根系的高死亡率会导致大量碳损失,不利于灌丛的生存[27 ] .因此,生长阶段初期的雏形阶段灌丛为适应干旱环境,选择降低根径以减少根系死亡所带来的碳消耗.而成熟阶段灌丛的根径显著大于雏形阶段和发育阶段灌丛也反映了随着灌丛的不断生长其抗干旱胁迫能力不断增强,有能力适应恶劣环境后选择寿命更长的较粗不定根.1—2级根径比小于2—3级,说明了随根径级数增加根径逐渐变小,但不同生长阶段的白刺灌丛不定根系根径比均未表现出显著差异.各级根系分支前后的一致性反映了根系的自相似性,因此可通过已测数据对部分构型指标进行估算,从而获得相关数据.这是对植物构型研究及分析的有效方法,不仅降低了工作难度,并且减少了对植物破坏[7 ] . ...
多枝柽柳(Tamarix ramosissima )根、冠构型的年龄差异及其适应意义
1
2015
... 根系构型特征随不同的水分、养分条件产生变化,具有一定可塑性[24 ] .根系构型差异影响根系对土壤水分的获取能力以及碳的消耗[25 ] 与分配[26 ] .灌丛的根径与其寿命正相关,但较大的根径会消耗更多的碳,而干旱胁迫环境下灌丛根系的高死亡率会导致大量碳损失,不利于灌丛的生存[27 ] .因此,生长阶段初期的雏形阶段灌丛为适应干旱环境,选择降低根径以减少根系死亡所带来的碳消耗.而成熟阶段灌丛的根径显著大于雏形阶段和发育阶段灌丛也反映了随着灌丛的不断生长其抗干旱胁迫能力不断增强,有能力适应恶劣环境后选择寿命更长的较粗不定根.1—2级根径比小于2—3级,说明了随根径级数增加根径逐渐变小,但不同生长阶段的白刺灌丛不定根系根径比均未表现出显著差异.各级根系分支前后的一致性反映了根系的自相似性,因此可通过已测数据对部分构型指标进行估算,从而获得相关数据.这是对植物构型研究及分析的有效方法,不仅降低了工作难度,并且减少了对植物破坏[7 ] . ...
喜旱莲子草和接骨草竞争对模拟增温的响应
1
2015
... 植物会随着环境的不断变化对其根系构型做出调整,以期达到对资源的最大利用[28 ] .前人研究发现,植物的根系分支率与环境干旱胁迫程度负相关[6 ] .在等量碳投入条件下,虽然人字形分支分布范围更小,不利于养分占有,但分支结构简单,对资源吸收利用能力更强,更适应胁迫环境[7 ,18 ] .本研究中随灌丛生长发育其拓扑指数参数显著降低,成熟阶段灌丛的分支率显著高于发育阶段和雏形阶段灌丛,不同生长阶段白刺灌丛不定根系构型均表现为人字形,该结论与单立山等[29 ] 对地下根系的研究结论不同.造成该结论差异的原因可能与不同地区资源差异有关,也可能是由于不定根系和地下根系自身特性差异有关,有待进一步研究.本试验的结果表明了研究区的白刺灌丛随生长发育抗干旱胁迫能力不断增强,成熟阶段灌丛不同于发育阶段和雏形阶段灌丛的将更多精力放在对环境适应上,而是有余力通过增加不定根系分支率来扩张其对资源的吸收利用范围以满足地上部分生长需求,这表现了白刺灌丛不同生长阶段为环境适应而做出的动态调整.白刺灌丛的不定根系中未发现有4级根的出现,该结果的产生与土壤容重有关.不定根系随灌丛发育多向深层土壤生长,而深层土壤较高的紧实度对根系生长形成阻力,不利于根系的扩张,因此影响了不定根生长与分支能力[30 ] . ...
河西走廊中部两种荒漠植物根系构型特征
2
2013
... 植物会随着环境的不断变化对其根系构型做出调整,以期达到对资源的最大利用[28 ] .前人研究发现,植物的根系分支率与环境干旱胁迫程度负相关[6 ] .在等量碳投入条件下,虽然人字形分支分布范围更小,不利于养分占有,但分支结构简单,对资源吸收利用能力更强,更适应胁迫环境[7 ,18 ] .本研究中随灌丛生长发育其拓扑指数参数显著降低,成熟阶段灌丛的分支率显著高于发育阶段和雏形阶段灌丛,不同生长阶段白刺灌丛不定根系构型均表现为人字形,该结论与单立山等[29 ] 对地下根系的研究结论不同.造成该结论差异的原因可能与不同地区资源差异有关,也可能是由于不定根系和地下根系自身特性差异有关,有待进一步研究.本试验的结果表明了研究区的白刺灌丛随生长发育抗干旱胁迫能力不断增强,成熟阶段灌丛不同于发育阶段和雏形阶段灌丛的将更多精力放在对环境适应上,而是有余力通过增加不定根系分支率来扩张其对资源的吸收利用范围以满足地上部分生长需求,这表现了白刺灌丛不同生长阶段为环境适应而做出的动态调整.白刺灌丛的不定根系中未发现有4级根的出现,该结果的产生与土壤容重有关.不定根系随灌丛发育多向深层土壤生长,而深层土壤较高的紧实度对根系生长形成阻力,不利于根系的扩张,因此影响了不定根生长与分支能力[30 ] . ...
... 根系的形态受遗传和外界环境两方面因素影响,为适应不同环境下的水分、养分条件,会在空间分布上表现出一定的自适应性[31 ] .本试验发现,白刺灌丛的不定根系的空间分布以垂直根为主,倾斜根其次,水平根最少,表现出深根型特征.这一现象主要受研究区气候条件影响.干旱少雨、日照充足、土壤水分蒸发量大,正是研究区这些恶劣的气候特征导致表层土壤含水率极低.因此白刺灌丛为获得更多水分,不定根更多地向含水率更高的深层土壤生长[29 ] .这种空间分布特征也表现了白刺灌丛为适宜干旱环境而做出的重要生存策略.也有研究表明倾斜根可以拥有最好的固土能力[32 ] ,因此,灌丛沙堆内部大量倾斜不定根系对沙堆土壤有很强的稳固作用. ...
极端干旱环境下的胡杨细根分布与土壤特征
1
2011
... 植物会随着环境的不断变化对其根系构型做出调整,以期达到对资源的最大利用[28 ] .前人研究发现,植物的根系分支率与环境干旱胁迫程度负相关[6 ] .在等量碳投入条件下,虽然人字形分支分布范围更小,不利于养分占有,但分支结构简单,对资源吸收利用能力更强,更适应胁迫环境[7 ,18 ] .本研究中随灌丛生长发育其拓扑指数参数显著降低,成熟阶段灌丛的分支率显著高于发育阶段和雏形阶段灌丛,不同生长阶段白刺灌丛不定根系构型均表现为人字形,该结论与单立山等[29 ] 对地下根系的研究结论不同.造成该结论差异的原因可能与不同地区资源差异有关,也可能是由于不定根系和地下根系自身特性差异有关,有待进一步研究.本试验的结果表明了研究区的白刺灌丛随生长发育抗干旱胁迫能力不断增强,成熟阶段灌丛不同于发育阶段和雏形阶段灌丛的将更多精力放在对环境适应上,而是有余力通过增加不定根系分支率来扩张其对资源的吸收利用范围以满足地上部分生长需求,这表现了白刺灌丛不同生长阶段为环境适应而做出的动态调整.白刺灌丛的不定根系中未发现有4级根的出现,该结果的产生与土壤容重有关.不定根系随灌丛发育多向深层土壤生长,而深层土壤较高的紧实度对根系生长形成阻力,不利于根系的扩张,因此影响了不定根生长与分支能力[30 ] . ...
Multi-proxy evidence for competition between savanna woody species
1
2007
... 根系的形态受遗传和外界环境两方面因素影响,为适应不同环境下的水分、养分条件,会在空间分布上表现出一定的自适应性[31 ] .本试验发现,白刺灌丛的不定根系的空间分布以垂直根为主,倾斜根其次,水平根最少,表现出深根型特征.这一现象主要受研究区气候条件影响.干旱少雨、日照充足、土壤水分蒸发量大,正是研究区这些恶劣的气候特征导致表层土壤含水率极低.因此白刺灌丛为获得更多水分,不定根更多地向含水率更高的深层土壤生长[29 ] .这种空间分布特征也表现了白刺灌丛为适宜干旱环境而做出的重要生存策略.也有研究表明倾斜根可以拥有最好的固土能力[32 ] ,因此,灌丛沙堆内部大量倾斜不定根系对沙堆土壤有很强的稳固作用. ...
基于根系构型的调控提高植物边坡根系固土能力
1
2021
... 根系的形态受遗传和外界环境两方面因素影响,为适应不同环境下的水分、养分条件,会在空间分布上表现出一定的自适应性[31 ] .本试验发现,白刺灌丛的不定根系的空间分布以垂直根为主,倾斜根其次,水平根最少,表现出深根型特征.这一现象主要受研究区气候条件影响.干旱少雨、日照充足、土壤水分蒸发量大,正是研究区这些恶劣的气候特征导致表层土壤含水率极低.因此白刺灌丛为获得更多水分,不定根更多地向含水率更高的深层土壤生长[29 ] .这种空间分布特征也表现了白刺灌丛为适宜干旱环境而做出的重要生存策略.也有研究表明倾斜根可以拥有最好的固土能力[32 ] ,因此,灌丛沙堆内部大量倾斜不定根系对沙堆土壤有很强的稳固作用. ...
西鄂尔多斯地区5种荒漠优势灌丛生物量分配格局及预测模型
1
2017
... 植物为了适应周围生态环境,通过调节不同器官生物量分配来达到对资源利用的最大化,而灌丛的不同生长阶段也会因为其自身生长特性表现出不同的分配方式[8 ] .本研究中,不同生长阶段白刺灌丛的根冠比随植物的生长发育而不断降低,不定根系的生长速率也表现为持续下降,生长关系表现为异速生长.该结论与肖遥等[8 ] 的研究结论相似.不同时期的植物相关生长是动态变化的,且变化趋势也不同.由于西鄂尔多斯地区干旱少雨、土壤贫瘠,特殊的气候条件致使这里植物地下部分生物量占比更高[33 ] ,而雏形阶段白刺灌丛拥有最高的根冠比与研究区的气候因素有很大关系.处于生长初期的雏形阶段灌丛为适应干旱贫瘠的环境,在生存决策上选择在根径更细、分支结构更简单等方面调整以适应环境,生物量分配上同样集中于根系部分,通过增加根系生长速率,以此扩大对土壤水分、养分等资源的利用范围,进而提高对恶劣生存环境的适应能力.而发育阶段和成熟阶段灌丛在不断生长发育的过程中通过根系分泌、枝系拦截等方式改变了土壤微环境[34 ] ,逐渐适应研究区干旱贫瘠的环境后,灌丛为获得更高的光照资源在生态策略上选择降低根系部分生长速率,将更多生物量分配于地上部分,进而提高争夺空间和光照面积的能力,以满足自身生长、繁殖需要[35 ] . ...
干旱地区灌丛沙堆研究现状与展望
1
2010
... 植物为了适应周围生态环境,通过调节不同器官生物量分配来达到对资源利用的最大化,而灌丛的不同生长阶段也会因为其自身生长特性表现出不同的分配方式[8 ] .本研究中,不同生长阶段白刺灌丛的根冠比随植物的生长发育而不断降低,不定根系的生长速率也表现为持续下降,生长关系表现为异速生长.该结论与肖遥等[8 ] 的研究结论相似.不同时期的植物相关生长是动态变化的,且变化趋势也不同.由于西鄂尔多斯地区干旱少雨、土壤贫瘠,特殊的气候条件致使这里植物地下部分生物量占比更高[33 ] ,而雏形阶段白刺灌丛拥有最高的根冠比与研究区的气候因素有很大关系.处于生长初期的雏形阶段灌丛为适应干旱贫瘠的环境,在生存决策上选择在根径更细、分支结构更简单等方面调整以适应环境,生物量分配上同样集中于根系部分,通过增加根系生长速率,以此扩大对土壤水分、养分等资源的利用范围,进而提高对恶劣生存环境的适应能力.而发育阶段和成熟阶段灌丛在不断生长发育的过程中通过根系分泌、枝系拦截等方式改变了土壤微环境[34 ] ,逐渐适应研究区干旱贫瘠的环境后,灌丛为获得更高的光照资源在生态策略上选择降低根系部分生长速率,将更多生物量分配于地上部分,进而提高争夺空间和光照面积的能力,以满足自身生长、繁殖需要[35 ] . ...
阔叶红松林下6种早夏草本不同生长期生物量分配及模型构建
1
2017
... 植物为了适应周围生态环境,通过调节不同器官生物量分配来达到对资源利用的最大化,而灌丛的不同生长阶段也会因为其自身生长特性表现出不同的分配方式[8 ] .本研究中,不同生长阶段白刺灌丛的根冠比随植物的生长发育而不断降低,不定根系的生长速率也表现为持续下降,生长关系表现为异速生长.该结论与肖遥等[8 ] 的研究结论相似.不同时期的植物相关生长是动态变化的,且变化趋势也不同.由于西鄂尔多斯地区干旱少雨、土壤贫瘠,特殊的气候条件致使这里植物地下部分生物量占比更高[33 ] ,而雏形阶段白刺灌丛拥有最高的根冠比与研究区的气候因素有很大关系.处于生长初期的雏形阶段灌丛为适应干旱贫瘠的环境,在生存决策上选择在根径更细、分支结构更简单等方面调整以适应环境,生物量分配上同样集中于根系部分,通过增加根系生长速率,以此扩大对土壤水分、养分等资源的利用范围,进而提高对恶劣生存环境的适应能力.而发育阶段和成熟阶段灌丛在不断生长发育的过程中通过根系分泌、枝系拦截等方式改变了土壤微环境[34 ] ,逐渐适应研究区干旱贫瘠的环境后,灌丛为获得更高的光照资源在生态策略上选择降低根系部分生长速率,将更多生物量分配于地上部分,进而提高争夺空间和光照面积的能力,以满足自身生长、繁殖需要[35 ] . ...
天山林区六种灌木生物量的建模及其器官分配的适应性
2
2015
... 灌丛根系是固定植物以及吸收利用荒漠环境中养分、水分的重要器官,同时也是地上部分正常生长发育的基本保障[36 ] .研究灌丛根系生物量可以了解灌丛的生产潜力,揭示灌丛对该区域自然资源的利用能力与效率.不同径级根系的功能不同[37 ] ,粗根用于支撑固定植物和贮存营养物质[36 ] ,细根负责养分与水分的吸收和运输[38 ] .因此,除根冠比和相对生长关系外,各级根系生物量占比也可反映植物对环境的适应策略.由于灌丛的根径大小随生长阶段变化也表现明显差异,因此本文未采用绝对根径划分灌丛的粗根与细根,而是依据灌丛的径级将其区分为粗根与细根,其中3级根代表粗根,1级根、2级根代表细根.本试验结果显示,灌丛生长发育过程中分支率不断增加,灌丛细根不断生长,其生物量占比不断提高.雏形阶段灌丛为更好地固定灌丛沙堆以及保存营养适应贫瘠土壤,不定根系多为粗根,而发育后期灌丛逐渐稳定,因此将更多生物量分配到细根以求提高资源吸收利用范围及效率.这表明灌丛随生长发育在生态策略上会做出调整. ...
... [36 ],细根负责养分与水分的吸收和运输[38 ] .因此,除根冠比和相对生长关系外,各级根系生物量占比也可反映植物对环境的适应策略.由于灌丛的根径大小随生长阶段变化也表现明显差异,因此本文未采用绝对根径划分灌丛的粗根与细根,而是依据灌丛的径级将其区分为粗根与细根,其中3级根代表粗根,1级根、2级根代表细根.本试验结果显示,灌丛生长发育过程中分支率不断增加,灌丛细根不断生长,其生物量占比不断提高.雏形阶段灌丛为更好地固定灌丛沙堆以及保存营养适应贫瘠土壤,不定根系多为粗根,而发育后期灌丛逐渐稳定,因此将更多生物量分配到细根以求提高资源吸收利用范围及效率.这表明灌丛随生长发育在生态策略上会做出调整. ...
甘肃景电灌区不同栽植年限枸杞生物量分配特征
1
2012
... 灌丛根系是固定植物以及吸收利用荒漠环境中养分、水分的重要器官,同时也是地上部分正常生长发育的基本保障[36 ] .研究灌丛根系生物量可以了解灌丛的生产潜力,揭示灌丛对该区域自然资源的利用能力与效率.不同径级根系的功能不同[37 ] ,粗根用于支撑固定植物和贮存营养物质[36 ] ,细根负责养分与水分的吸收和运输[38 ] .因此,除根冠比和相对生长关系外,各级根系生物量占比也可反映植物对环境的适应策略.由于灌丛的根径大小随生长阶段变化也表现明显差异,因此本文未采用绝对根径划分灌丛的粗根与细根,而是依据灌丛的径级将其区分为粗根与细根,其中3级根代表粗根,1级根、2级根代表细根.本试验结果显示,灌丛生长发育过程中分支率不断增加,灌丛细根不断生长,其生物量占比不断提高.雏形阶段灌丛为更好地固定灌丛沙堆以及保存营养适应贫瘠土壤,不定根系多为粗根,而发育后期灌丛逐渐稳定,因此将更多生物量分配到细根以求提高资源吸收利用范围及效率.这表明灌丛随生长发育在生态策略上会做出调整. ...
水曲柳和落叶松不同根序之间细根直径的变异研究
1
2005
... 灌丛根系是固定植物以及吸收利用荒漠环境中养分、水分的重要器官,同时也是地上部分正常生长发育的基本保障[36 ] .研究灌丛根系生物量可以了解灌丛的生产潜力,揭示灌丛对该区域自然资源的利用能力与效率.不同径级根系的功能不同[37 ] ,粗根用于支撑固定植物和贮存营养物质[36 ] ,细根负责养分与水分的吸收和运输[38 ] .因此,除根冠比和相对生长关系外,各级根系生物量占比也可反映植物对环境的适应策略.由于灌丛的根径大小随生长阶段变化也表现明显差异,因此本文未采用绝对根径划分灌丛的粗根与细根,而是依据灌丛的径级将其区分为粗根与细根,其中3级根代表粗根,1级根、2级根代表细根.本试验结果显示,灌丛生长发育过程中分支率不断增加,灌丛细根不断生长,其生物量占比不断提高.雏形阶段灌丛为更好地固定灌丛沙堆以及保存营养适应贫瘠土壤,不定根系多为粗根,而发育后期灌丛逐渐稳定,因此将更多生物量分配到细根以求提高资源吸收利用范围及效率.这表明灌丛随生长发育在生态策略上会做出调整. ...





 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}