疏勒河源冻土区高寒草甸土壤碳氮含量特征
贾映兰 , 1 , 2 , 吴明辉 1 , 2 , 魏培洁 1 , 2 , 赵建中 4 , 高雅月 5 , 陈生云 , 1 , 3 , 5
1.中国科学院西北生态环境资源研究院 冰冻圈科学国家重点实验室/疏勒河源冰冻圈与生态环境综合监测研究站,甘肃 兰州 730000
2.中国科学院大学,北京 100049
3.青海师范大学 高原科学与可持续发展研究院/地理科学学院,青海 西宁 810008
4.青海农牧科技职业学院,青海 西宁 812100
5.祁连山国家公园国家长期科研基地,青海 西宁 810000
Characteristics of soil carbon and nitrogen contents in permafrost region of the Shule River Headwaters
Jia Yinglan , 1 , 2 , Wu Minghui 1 , 2 , Wei Peijie 1 , 2 , Zhao Jianzhong 4 , Gao Yayue 5 , Chen Shengyun , 1 , 3 , 5
1.State Key Laboratory of Cryospheric Science / Cryosphere and Eco-Environment Research Station of Shule River Headwaters,Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
2.University of Chinese Academy of Sciences,Beijing 100049,China
3.Academy of Plateau Science and Sustainability /School of Geographical Sciences,Qinghai Normal University,Xining 810008,China
4.Qinghai Agri -animal Husbandry Vocational College,Xining 812100,China
5.Long-term National Scientific Research Base of the Qilian Mountain National Park,Xining 810000,China
通讯作者: 陈生云(E-mail: sychen@lzb.ac.cn )
收稿日期: 2021-11-17
修回日期: 2022-01-19
基金资助:
国家自然科学基金项目 . 41871064 青海省重点研发与转化计划项目 . 2020-SF-146 青海省“高端创新人才千人计划”高层次人才项目
Received: 2021-11-17
Revised: 2022-01-19
作者简介 About authors
贾映兰(1996—),女,甘肃陇西人,硕士研究生,研究方向为冻土碳氮循环E-mail:yinglanj@163.com
, E-mail:yinglanj@163.com
摘要
土壤碳氮是高寒植被响应多年冻土区生态环境变化的重要营养和能源物质,但对其调查仍以生长季的单次采样为主,缺乏对其他季节的研究,这对于准确把握多年冻土区土壤碳氮含量及储量评估存在明显局限性。为此,本研究以青藏高原东北缘祁连山西段疏勒河源多年冻土区高寒草甸为对象,对0—50 cm土层土壤有机碳(Soil Organic Carbon, SOC)、全氮(Total Nitrogen, TN)含量及其比值(C/N)的剖面分布和季节变化及其影响因素进行分析。结果表明:(1)SOC、TN剖面分布规律一致,0—10 cm土层均显著高于10—50 cm各层(P <0.05),0—50 cm深度仅秋季逐渐下降,而春夏冬季0—30 cm递减。(2)SOC、TN含量存在季节变化,SOC表现为夏季>冬季>春季>秋季,TN表现为春秋冬季含量一致,夏季略低。(3)C/N季节变化显著,夏季显著最高,秋季显著最低(P <0.05)。(4)土壤含水量和生物量是影响SOC、TN及C/N剖面分布和季节变化的关键因素。(5)夏季土壤碳氮密度均高于全年平均。可见,仅单一节点(生长季为主)调查以表征全年土壤碳氮储量存在高估趋势。
关键词:
多年冻土区 高寒草甸 土壤碳氮 剖面变化 季节特征
Abstract
Soil carbon and nitrogen are important nutrient and energy substances for alpine vegetation to respond to the changes of ecological environment in permafrost regions. However, their surveys are still based on single-pass sampling during the growing season, and there is a lack of researches on seasonal changes, which is very important but exists obvious limitation for accurately grasping contents and estimating storages of soil carbon and nitrogen in permafrost area. Thus, in the study, the alpine meadow, located at the permafrost regions of Shule River headwaters in the western part of the Qilian Mountains and the northeast margin of the Qinghai-Tibet Plateau, was selected as the study area. The vertical (0-50 cm profile) and seasonal variations of soil organic carbon (SOC), total nitrogen (TN) contents and carbon to nitrogen ratio (C/N), as well as their influencing factors were analyzed. The results showed that: (1) The profile distribution of SOC and TN are consistent as, SOC and TN in the 0-10 cm soil layer were significantly higher than those in the 10-50 cm (P <0.05). However, the SOC and TN decreased gradually from 0 to 50 cm only in autumn, while 0-30 cm in spring, summer and winter; (2) Seasonal changes were found in SOC and TN contents, and SOC showed summer > winter > spring > autumn, while TN was consistent in spring, autumn and winter, but slightly lower in summer; (3) The seasonal variation of C/N was remarkable (P <0.05), which highest in summer and lowest in autumn; 4) Soil moisture content and vegetation biomass were key factors for vertical and seasonal changes in the SOC content, TN content and C/N; (5) The soil carbon and nitrogen density in summer were higher than the annual average. It can be seen that only the survey of a single node (mainly in the growing season) indicates that there is an overvalued trend of the annual soil carbon and nitrogen storage.
Keywords:
permafrost regions alpine meadow soil carbon and nitrogen vertical change seasonal pattern
本文引用格式
贾映兰, 吴明辉, 魏培洁, 赵建中, 高雅月, 陈生云. 疏勒河源冻土区高寒草甸土壤碳氮含量特征 . 中国沙漠 [J], 2022, 42(4): 219-228 doi:10.7522/j.issn.1000-694X.2022.00013
Jia Yinglan, Wu Minghui, Wei Peijie, Zhao Jianzhong, Gao Yayue, Chen Shengyun. Characteristics of soil carbon and nitrogen contents in permafrost region of the Shule River Headwaters . Journal of Desert Research
0 引言
高寒草甸是青藏高原分布最广的植被类型,约占高原总面积的49.81%[1 ] ,也是多年冻土区的优势植被类型(约占冻土区总面积的80%)[2 ] 。近年来,由于气候变暖,高原多年冻土区生态环境变化显著[3 ] 。一方面,多年冻土广泛、持续和快速退化[4 ] ,冻土的隔水作用下降,入渗能力增强[5 ] ,地表植被可利用水分及养分流失,导致依赖于冻结层上水的短根系植物枯死[3 ] ,造成其植被盖度下降和组成变化[6 ] ;另一方面,多年冻土融化改变地形地貌,如热熔塌陷和热融沟[7 ] ,不仅会使土壤中冻结的有机质暴露于空气中被持续氧化,甚至导致草地出现大面积“斑块化”现象[8 ] 。然而,冻土环境的改变与植被变化并不是相互独立的,而是耦合形成植被-土壤反馈系统协同变化[9 ] 。在这个系统中,土壤碳氮作为主要的营养和能源物质[10 ] ,在调节植被生长、改善土壤质量、参与土壤呼吸和养分循环等[11 ] 生物和非生物重要过程中扮演重要角色,其含量和分布受冻土环境变化影响十分显著[7 ,12 ] ,且直接关系到高寒植被的演替趋势[13 ] ,并最终影响整个高原土壤碳氮储藏和多年冻土生态系统碳氮库。
然而,目前诸多研究仅集中于比较高原多年冻土区不同采样点之间土壤碳氮的含量大小及剖面分布特征。李新星等[14 ] 对马衔山的研究表明,表层土壤有机碳(Soil Organic Carbon, SOC)和全氮(Total Nitrogen, TN)含量高于下层;Zhao等[15 ] 发现土壤碳氮储量随土壤深度增加呈现降低趋势;马素辉等[16 ] 以黑河上游俄博岭各草地类型为研究对象,比较发现高海拔的高寒沼泽草甸SOC密度(Soil Organic Carbon Density, SOCD)最高,而高寒草甸海拔低且SOCD显著降低;Wang等[17 ] 模拟得出,青藏高原东部SOCD空间上由东向西递减;Xie等[18 ] 发现TN含量季节上基本保持稳定,但高寒草甸显著高于其他植被类型。尽管各研究关于多年冻土区土壤碳氮分布具有较高的一致性,但其对于了解高寒区土壤碳氮分布甚至于评估碳氮库储量存在明显的局限性。首先,多年冻土区由于年内温度变化剧烈,浅层土壤存在明显的季节性冻融现象[19 ] ,表现为春秋季冻融频繁,夏冬季则分别完全融化和冻结[20 ] 。已有研究表明,季节性冻融会对土壤碳氮矿化速率[21 ] 及土壤水溶性有机碳[22 ] 和微生物碳氮[23 ] 的含量产生显著影响;其次,大量的基于单一时间节点的采样调查忽视了季节变化对土壤碳氮的影响,导致相关研究对土壤碳氮库储量的估计仍存在明显的不确定性。寇丹[24 ] 、Yuan等[25 ] 均是以一年中单采样节点(生长季)对青藏高原、黑河上游多年冻土区土壤碳氮储量进行了评估。然而,非生长季对土壤碳、氮循环影响在年际尺度上是不可忽视的[12 ] 。因此,在多年冻土区开展以空间分布和季节变化相结合的生态调查对于准确把握土壤碳氮含量分布及评估碳氮储量具有重要意义。
位于青藏高原东北缘祁连山西段的疏勒河源区多年冻土广泛发育,其多年冻土面积约占源区面积的73%[9 ] ,源区内优势植被类型为高寒草甸,覆盖面积约为42%[22 ] 。近20年观测资料显示,源区冻土退化明显且土壤季节性冻融活动显著[9 , 22 ] 。目前该区域的最近土壤碳氮调查资料仅为2012年Liu等[26 ] 对生长季高寒草甸内单一样点的剖面分布研究,缺乏年内季节变化研究。因此,本研究拟开展源区浅层SOC、TN含量在季节尺度上的变化研究,揭示其垂直分布规律及主要的影响因素,并估算疏勒河源多年冻土区高寒草甸土壤碳氮储量,以期为全球气候变化背景下青藏高原多年冻土区高寒草甸土壤碳、氮研究提供数据支撑和科学指导。
1 材料与方法
1.1 样地概况
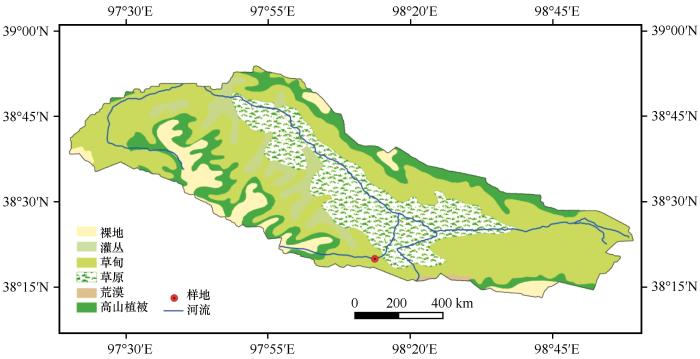
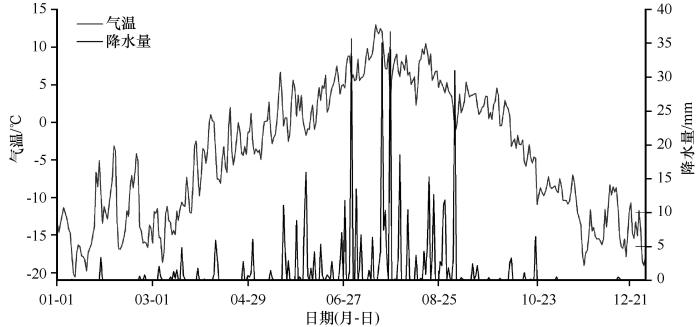
疏勒河源区位于青藏高原东北缘祁连山西段,面积约为5 123 km2 ,约占祁连山国家公园青海片区的19.68%。气候类型为大陆性干旱荒漠气候,干冷多风[27 ] 。该区域主要植被类型为灌丛、草甸、草原、荒漠及高山稀疏植被,其中高寒草甸面积为2 393.57 km2 ,约占源区总面积的46.72%(图1 )。研究样地位于疏勒河源多年冻土区高寒草甸生态系统综合观测场(38°19′58″N、98°13′44″E,海拔4 036 m,图1 )。基于观测场2017年的气象观测数据资料显示,样地内年均气温为-4.06 ℃,年降水量为466.6 mm且集中在6—9月(图2 ),空气相对湿度为55.49%,年均太阳总辐射为194.3 W·m-2 。0—50 cm土壤砂粒、粉粒和黏粒含量分别为53.80%、35.69%和9.97%。多年冻土类型为阿尔金山-祁连山高寒带山地多年冻土[27 ] ,活动层厚度约为1.5 m,土壤类型为冷钙土。植被覆盖度80%以上,优势植物种有藏嵩草(Kobresia tibetica )和高山嵩草(Kobresia pygmaea )[9 ] 。
图1
图1
研究区及采样点位置图
Fig.1
Location of study area and sampling point
图2
图2
样地2017年气温和降水量日变化
Fig.2
Daily variations of air temperature and precipitation in the sampling site in 2017
1.2 研究方法
1.2.1 样品采集
2017年4月29日与5月7日、7月29日、9月29日与10月3日和12月30日分别采样,用以表征春、夏、秋季和冬季样品。在样地内随机选取6个50 cm×50 cm样方,样方内的植物用剪刀齐地收集并装入信封(鲜草和凋落物),用于测定地上生物量。由于地下生物量集中在0— 30 cm层[27 ] ,用土钻(内半径2.75 cm)自上而下对0— 50 cm土壤分5层(0— 10、10— 20、20— 30、30— 40 cm和40— 50 cm)取样。每层样品取3次混合后,分为3份:1份待自然风干后用于测定SOC、TN含量;1份放入4 ℃便携式保温箱带回实验室,用于测定pH值和氧化还原电位;剩余1份自然风干后清洗,用于测定地下生物量。环刀(100 g·cm-3 )采集土壤容重并用于计算SOCD和TN密度(Total Nitrogen Density, TND)。
1.2.2 环境因子监测和样品分析
研究样地内布设有全自动监测系统。其中,气温由HMP155A型温湿度传感器(Vaisala,芬兰)测定,降水数据采用人工雨量筒进行测定。0—10、10—20、20—30、30—40、40—50 cm各层土壤温度和含水量均利用Hydra-Probe II型土壤传感器(Stevens公司,美国)测定,上述传感器连接至CR1000型数据采集器(Campbell公司,美国),每10 min记录1次数据[28 ] 。
SOC、TN含量分别用重铬酸钾氧化-外加热法和微量凯氏定氮法测量,C/N则由SOC除以TN所得;pH值和氧化还原电位用酸度计(PHBJ-260)进行测定(水土比为5∶1)[9 ] 。地下生物量是将土壤样品先去除杂物再经过2 mm筛洗净,将其与地上生物量放入烘箱80 ℃烘干至恒重称重所得[23 ] 。样地0—50 cm土壤理化性质及生物量概况如表1 所示。
1.2.3 数据分析
S O C D = ∑ i = 1 N h i × B D i × S O C i × ( 1 - C i ) / 100 (1)
式中:SOCD 代表土壤有机碳密度(kg·m-2 );hi 代表土壤厚度(cm);BDi 代表土壤容重(g·cm-3 );SOCi 代表土壤有机碳含量(g·kg-1 );Ci 代表>2 mm土壤石砾百分含量(%);i 为参与计算的土壤层次;n 为参与计算的土壤层总数。TND 的计算公式同SOCD [29 ] ,土壤碳、氮储量由研究区高寒草甸面积分别与其年均密度的乘积计算所得,0—50 cm土层有机碳和全氮含量是0—10、10—20、20—30、30—40、40—50 cm各层次土壤有机碳和全氮含量的加权平均值[30 ] ,年均SOCD 、TND 由四季求平均值所得。
采用Excel 2019和SPSS 22.0对所测数据进行整理及统计分析,利用单因素方差分析(One-way ANOVA)及最小显著性差异法(Least-Significant Difference,LSD)检验SOC、TN及C/N在不同季节及土层之间的差异;用双因素方差分析(Two-way ANOVA)检验剖面和季节变化及其交互作用对SOC、TN及C/N的影响。用Pearson相关和逐步回归分析环境因子(气象、土壤理化性质及生物量)与剖面和季节上SOC、TN及C/N之间的关系,采用Origin 2018制图。
2 结果与分析
2.1 SOC 、TN 剖面和季节变化特征
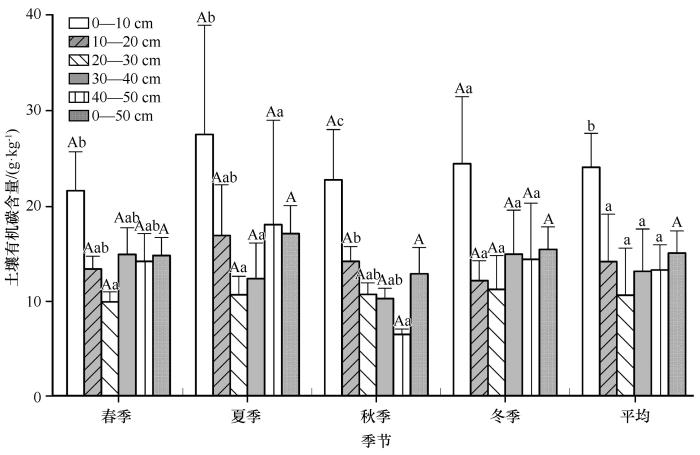
0—50 cm层SOC含量表现出“N”型变化特征,即夏季>冬季>春季>秋季(图3 )。在各土层深度中,不同季节SOC变化特征表现不同,其中,0—10、10—20 cm SOC也在夏季达到最大值,但最小值分别出现在春季和冬季;20—30、30—40 cm SOC含量最大值出现在冬季,最低值分别在春、秋季;而40—50 cm层SOC含量变化同0—50 cm分布一致。此外,相同土层不同季节间SOC含量均没有显著差异(P >0.05)。
图3
图3
土壤有机碳(SOC)含量的变化特征
不同大写字母表示同一土层不同季节间的差异显著(P <0.05),不同小写字母表示同一季节不同土层间的差异显著(P <0.05)
Fig.3
The variation characteristic of SOC content
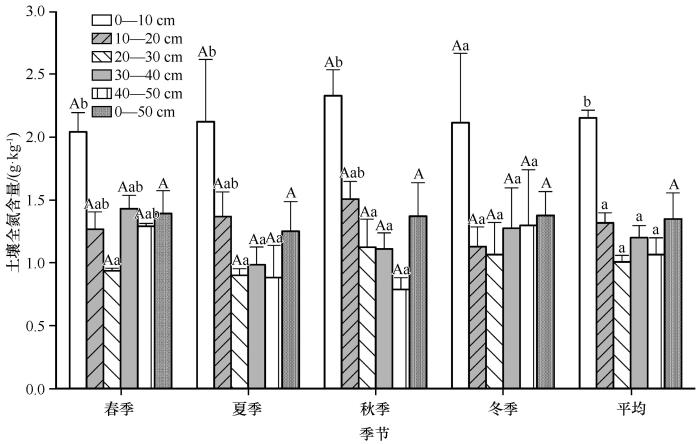
0—50 cm层TN含量季节变化规律呈“V”型分布(图4 ),即春、秋季和冬季基本一致,夏季略低。春季0—50 cm TN含量平均值为1.39±0.18 g·kg-1 ,分别是夏、秋、冬季的1.11、1.02、1.01倍。各层TN的季节分布情况为:0—10、10—20、20—30 cm层TN含量最大值出现在秋季,最小值分别出现在春、冬季和夏季;30—40 cm TN季节变化同0—50 cm一致;40—50 cm TN冬季值最大,秋季值最小。此外,相同土层不同季节间SOC含量均没有显著差异(P >0.05)。
图4
图4
土壤全氮(TN)含量的变化特征
不同大写字母表示同一土层不同季节间的差异显著(P <0.05),不同小写字母表示同一季节不同土层间的差异显著(P <0.05)
Fig.4
The variation characteristic of TN content
SOC和TN的垂直分布特征因季节不同产生差异,即随土壤深度增加,SOC、TN含量秋季递减,春、夏季和冬季仅在0—30 cm内递减。具体而言,除冬季SOC和TN含量各层差异不显著外(P >0.05),春季0—10 cm层SOC、TN含量均显著高于20—30 cm(P <0.05),秋冬季节0—10 cm层SOC、TN含量均显著高于20—30、30—40、40—50 cm(P <0.05)。0—50 cm层年平均SOC、TN含量分别为14.41±2.33、1.35±0.21 g·kg-1 ,且在0—30 cm呈递减趋势,而30—40 cm的SOC、TN均有略微升高。此外,0—50 cm土壤,春、夏、秋、冬季和年平均SOCD及TND分别为8.63±0.15、8.98±0.25、7.38±0.23、8.98±0.18、8.49±0.38 kg·m-2 及0.8121±0.10、0.7855±0.14、0.7202±0.11、0.8011±0.19、0.7802±0.02 kg·m-2 。0—50 cm年均土壤碳、氮储量分别为20.33±0.91、1.89±0.03 Tg。
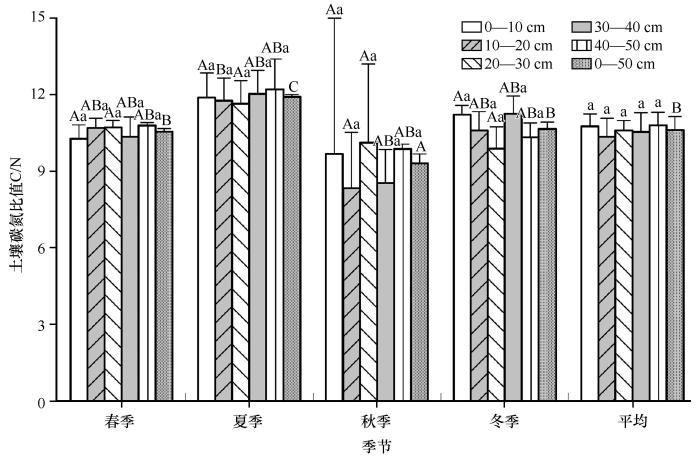
2.2 C/N 剖面及季节变化特征
如图5 所示,0—50 cm土壤C/N季节分布特征与SOC含量相似(夏季>冬季>春季>秋季),且夏季C/N显著高于春、秋季和冬季,秋季显著低于春、冬季(P <0.05)。此外,0—10、10—20、30—40 cm和40—50 cm层C/N的季节变化趋势与0—50 cm层的相同,且除0—10 cm外,其他3层C/N夏季显著高于秋季(P <0.05);20—30 cm C/N最低值出现在了冬季且各季节间差异不显著(P >0.05)。
图5
图5
C/N变化特征
不同大写字母表示同一土层不同季节间的差异显著(P <0.05),不同小写字母表示同一季节不同土层间的差异显著(P <0.05)
Fig.5
The variation "characteristic of C/N
各季节C/N随土层深度的变化形式各不相同。整体而言,春、夏季和秋季是波动上升,分别在40—50、40—50、20—30 cm层中最高,这说明表层土壤氮矿化速率较中下层更快;冬季则波动下降,且不同层间的C/N没有显著差异(P >0.05)。年平均C/N总体有上升的趋势,变幅10.35—10.80。
2.3 SOC 、TN 及C/N 的影响因素
方差分析结果(表2 )显示,剖面变化对SOC、TN含量有极显著的影响(P <0.01),对C/N无显著影响(P >0.05);季节变化仅对C/N的影响显著(P <0.05);剖面和季节变化的交互作用对SOC、TN和C/N均没有显著的影响(P >0.05)。
SOC、TN含量及C/N与各环境因子之间相关性如表3 所示。剖面变化上SOC、TN均与土壤含水量(SM)、生物量(AGB和BGB)显著正相关(P <0.05),与土壤pH极显著负相关(P <0.01),而C/N与各因子均无明显相关性(P >0.05);季节变化上SOC、TN均与AGB和BGB呈显著正相关关系(P <0.05),TN还与pH呈显著负相关关系(P <0.05),而C/N仅与BGB呈正相关关系(P <0.05)。
逐步回归分析表明,SOC、TN及C/N季节变化的主控因子均为BGB,BGB分别能够解释它们季节变异性的50.4%、55.7%和24.5%(表4 )。SOC剖面变化的关键因子为SM和BGB,其能够解释剖面变异性的76.7%(P <0.001),而TN剖面变化的驱动因子除SM和BGB,还有AGB,它们共同解释81.5%的变异性(P <0.001)。
3 讨论
3.1 SOC 、TN 的剖面变化
SOC含量取决于进入土壤的植物残体量及其土壤微生物分解作用下损失量的平衡,TN含量主要来源于植物残体归还和生物固氮[31 ] 。本研究中,SOC、TN剖面分布呈一定规律,0—10 cm层高于其他层,这与以往的研究结果一致[14 ] 。首先,地下生物量(根系及其分泌物、细根周转产生的碎屑等)是SOC和TN主要来源[29 ] 。研究表明,高海拔植物会将更多的生物量分配在地下部器官[32 ] ,且植被的凋落物在地表形成凋落物层[22 ] ,这决定了SOC、TN剖面分布特征;其次,土壤含水量是影响根系分布和土壤微生物分解的重要环境因子[29 ] 。春夏季土壤表层含水量高于下层[22 ] ,促进了植被生产力,进而输入土壤的碳氮增加,且高的水分可能会造成厌氧微生物群落丰度的增加,而厌氧生物对SOC的分解效率低于好氧生物,从而导致SOC的积累[33 ] ,秋冬季节下层含水量是高于表层的[22 ] ,表层土壤为物质交换提供畅通环境,为土壤碳氮的积累、分解和转化创造了良好的基础[34 ] 。此外,SOC、TN还与地上生物量显著正相关,与pH显著负相关,这说明地表枯落物对土壤碳、氮的输入是极其重要的,而pH通过影响植被的生长和土壤微生物的活性、数量来间接影响碳、氮的累积[9 ] 。
此外,SOC和TN的垂直分布还因季节不同产生差异,即随土壤深度增加,秋季0—50 cm内呈递减趋势,春、夏季和冬季仅在0—30 cm内递减,30—50 cm含量有所回升,这种递减趋势可能与该区域植被根系的分布在0—30 cm有关[27 ] ,而季节差异则可能是淋溶过程中养分从表层向下层迁移的动态结果[35 ] 。高寒草甸下发育着的土壤,质地为沙壤,养分易淋溶[36 ] ,表层的土壤碳氮会不同比例地向下层流失[37 ] 。但是,这种养分向深层的运移通道会因季节性冻融过程和融化深度不同而产生差异。冻融过程和融化深度主要通过改变土壤水热动态、孔隙结构及根系与土壤间的缝隙促进或抑制大孔隙流形成[38 ] 。具体来看,除秋季外,春夏冬季均在30—50 cm层SOC、TN含量有升高,这是由于30 cm层以下根系分布少且土壤紧实[39 ] ,一定程度上抑制了养分向下迁移的速率,并且冻土活动层在秋季达到最大融化深度[40 ] ,且中层土壤是高能水区[22 ] ,可能促进了表层SOC、TN持续向下层(30—50 cm)淋溶,因而秋季表现出空间的连续性降低。
3.2 SOC 、TN 的季节变化
生物量及环境因素的季节变化使SOC含量具有季节差异。陈梦棋等[41 ] 对比名山县生长季各植被类型SOC发现,其存在差异但不显著,这与本研究的结果相符。本研究表明,影响SOC季节差异的主控因子是地下生物量,这表明碳输入量的不同会影响SOC含量大小,但变幅相对较小。春季植被返青期主要消耗上一年份累积的土壤养分来度过[42 ] ,加之土壤融化水分和温度增加促进微生物活性并消耗碳源,因而春季含量低于冬季;夏季优越的水热条件植被生长旺盛,强烈的光合作用使根系和根茎固定了更多的碳,大量的有机物质(凋落物、根系分泌物及其脱落物等)进入土壤并转化为SOC,与此同时土壤微生物的新陈代谢虽会消耗一部分SOC,但SOC积累量远大于消耗量,表现为夏季最高;秋季地表枯黄产生凋落物、根系衰减及动物残体(气温骤降大量细菌因低温环境休眠或死亡)等还未来得及被分解而以腐殖质的形式储存在土体中,致秋季最低;冬季植被凋亡,地下生物量急剧降低,但来源于秋季的大量腐殖质为适应低温环境而幸存下来的真菌(微生物群体中约87%的细菌休眠或死亡)微生物提供了较为丰富的代谢底物[43 ] ,再加上积雪覆盖在土壤表面起隔热作用[23 ] ,以及低气温抑制土壤呼吸,减缓有机质分解速率[44 ] ,SOC积累胜过消耗,冬季高于秋季。此外,生长季温度高,土壤酶催化过程以及微生物代谢过程均加速[45 ] ,从而加快SOC矿化过程,这可能是造成各季节间SOC含量差异不显著的原因之一。本研究中,年均SOCD为8.49±0.25 kg·m-2 ,低于黑河上游多年冻土区高寒草甸以生长季采样所得结果(11.22 kg·m-2 )[16 ] ,可能原因是影响其变化的因素如植被盖度、凋落物、土壤呼吸速率因所处环境的不同均存在差异[46 -47 ] 。本研究通过全年采样得出,夏季SOCD比年平均密度约高5.77%,可见,利用生长盛季估算全年SOCD及储量会存在高估的趋势。
土壤TN的季节特征表现为夏季最少,春秋和冬季基本一致。这是由于各季节植被对氮素的消耗不同所致,夏季具有良好的代谢环境(碳源充足、温度适宜),微生物和根系活性高,更多的有机态氮转化为无机态氮[48 ] ,植物生长消耗大量氮素,再加上冻土融化土壤水分增加抑制了氧气进入,加速厌氧环境形成,反硝化过程增强,释放N2 O气体[12 ] ,使得夏季含量低于其他季节,而秋冬季消耗氮速率下降,氮矿化能力也随之降低,土壤中氮能够被累积下来。Liu等[29 ] 的研究表明,北麓河土壤平均TND为0.47±0.07 kg·m-2 ,本研究对应值为0.16±0.02 kg·m-2 ,海拔差异可能是造成两区域差异较大的主要原因。北麓河海拔相比疏勒河更高,据报道,海拔与TN之间具有极显著正相关性[14 ] ,海拔增加、温度下降,土壤中微生物和酶活性下降,分解速率减慢利于TN的积累[13 ] 。此外,本研究中SOC、TN含量的季节变化差异不显著,但存在季节变化趋势,这可能与观测时间仅为一年且植被类型单一有关,因而多年冻土区长期的连续观测及不同植被类型下各季节间的差异比较,将是进一步需要探究的问题。
3.3 C/N 的变化
C/N是反映土壤有机质分解速率及氮素矿化能力的标志[49 ] ,其比值低说明微生物分解有机质迅速,且氮矿化作用强。反之,有机质分解过程受氮素限制,微生物与植物存在对无机态氮的竞争,不利于植物的生长。本研究得出,剖面变化对C/N影响不显著,这与以往的研究结果一致[49 ] 。本研究中,0—50 cm年平均C/N为10.35—10.80,这与青藏高原北麓河的研究结果接近[29 ] ,略低于祁连山东段[50 ] ,表明多年冻土区土壤微生物对有机质的分解过程随地理位置的变化较小,可能与多年冻土区整体温度较低有关。
本研究中,季节变化对C/N有显著的影响,且地下生物量是主要影响因素,可能还与土壤含氧量有关。夏季C/N最高说明夏季微生物分解过程受氮的限制[51 ] ,而碳源充足,更多的SOC转化为土壤微生物碳[9 ] ,较少的TN转化为土壤微生物氮,刘放等[23 ] 的研究也证实,夏季疏勒河源高寒草甸微生物生物量碳最多,而微生物生物量氮含量最少;另一方面,有机质矿化过程中部分碳在土壤呼吸的作用下以CO2 的形式排放至大气,夏季土壤完全解冻,土壤中含氧量充足成为有机质的矿化能力升高的必要条件,有研究表明,疏勒河土壤CO2 通量夏季最高[28 ] 。秋季C/N最低,这说明该季节TN含量是相对充足的,有多余的有机氮转化为无机氮储存在土壤中,但土壤微生物同化1份氮的同时需要5份碳构建自身细胞,20份碳作为生命活动的能源,此时可利用的土壤碳却是较少的(图3 )。此外,地下生物量中大多为死根,能提供碳源的根系减少,土壤碳供应不足且土壤呼吸较弱,抑制了有机质矿化。
4 结论
本研究通过对青藏高原祁连山疏勒河源多年冻土区高寒草甸不同季节土壤0—50 cm SOC、TN含量的测定,发现各季节以及不同土层之间SOC、TN存在差异。在该研究区内,SOC、TN存在表层聚集性;SOC、TN随剖面深度增加的变化趋势因季节而异:春、夏季和冬季呈波动变化,秋季逐渐递减;春、夏季和冬季的C/N显著高于秋季。SOC、TN对土壤剖面的变化响应较季节更加敏感,而C/N受季节影响显著;SOC、TN剖面变化受土壤含水量和生物量的影响,地下生物量是影响SOC、TN及C/N季节差异的关键因子。生物量季节的不同导致土壤碳氮储量存在差异,且以生长盛季估算区域碳氮储量存在高估趋势。本研究发现各季节SOC、TN差异虽不显著,但存在动态变化趋势,因而长时间序列的观测及不同植被类型对比下SOC、TN季节动态变化将是未来继续探究的问题。
参考文献
View Option
[1]
李文宇 ,张扬建 ,沈若楠 ,等 氮磷共限制青藏高原高寒草甸生态系统碳吸收
[J].应用生态学报 ,2022 ,33 (1 ):51 -58 .
[本文引用: 1]
[2]
Wang Z W Wang Q Zhou L et al Mapping the vegetation distribution of the permafrost zone on the Qinghai-Tibet Plateau
[J].Journal of Mountain Science ,2016 ,13 (6 ):1035 -1046 .
[本文引用: 1]
[3]
程国栋 ,赵林 ,李韧 ,等 青藏高原多年冻土特征、变化及影响
[J].科学通报 ,2019 ,64 (27 ):2783 -2795 .
[本文引用: 2]
[4]
Jin H J Wu Q B Romanovsky V E Degrading permafrost and its impacts
[J].Advances in Climate Change Research ,2021 ,5 (1 ):1 -5 .
[本文引用: 1]
[5]
秦越 青藏高原东北部典型流域冻土退化及其生态水文效应研究
[D].北京 :清华大学 ,2018 .
[本文引用: 1]
[6]
Chen S Y Liu W J Qin X et al Response characteristics of vegetation and soil environment to permafrost degradation in the upstream regions of the Shule River Basin
[J].Environmental Research Letters ,2012 ,7 (4 ):45406 .
[本文引用: 1]
[7]
马蔷 ,金会军 气候变暖对多年冻土区土壤有机碳库的影响
[J].冰川冻土 ,2020 ,42 (1 ):91 -103 .
[本文引用: 2]
[8]
韩立辉 ,尚占环 ,任国华 ,等 青藏高原“黑土滩”退化草地植物和土壤对秃斑面积变化的响应
[J].草业学报 ,2011 ,20 (1 ):1 -6 .
[本文引用: 1]
[9]
吴明辉 ,瞿德业 ,李婷 ,等 祁连山疏勒河源区冻土退化对土壤微生物生物量碳氮的影响
[J].地理科学 ,2021 ,41 (1 ):177 -186 .
[本文引用: 7]
[10]
耿增超 ,姜林 ,李珊珊 ,等 祁连山中段土壤有机碳和氮素的剖面分布
[J].应用生态学报 ,2011 ,22 (3 ):665 -672 .
[本文引用: 1]
[11]
张鹏 ,张涛 ,陈年来 祁连山北麓山体垂直带土壤碳氮分布特征及影响因素
[J].应用生态学报 ,2009 ,20 (3 ):518 -524 .
[本文引用: 1]
[12]
宋阳 ,于晓菲 ,邹元春 ,等 冻融作用对土壤碳、氮、磷循环的影响
[J].土壤与作物 ,2016 ,5 (2 ):78 -90 .
[本文引用: 3]
[13]
王翀 高寒草地土壤有机碳影响因子与模拟模型的研究
[D].兰州 :兰州大学 ,2018 .
[本文引用: 2]
[14]
李新星 ,刘桂民 ,吴小丽 ,等 马衔山不同海拔土壤碳、氮、磷含量及生态化学计量特征
[J].生态学杂志 ,2020 ,39 (3 ):758 -765 .
[本文引用: 3]
[15]
Zhao L Wu X D Wang Z W et al Soil organic carbon and total nitrogen pools in permafrost zones of the Qinghai-Tibetan Plateau
[J].Scientific Reports ,2018 ,8 :3656 .
[本文引用: 1]
[16]
马素辉 ,牟翠翠 ,郭红 ,等 祁连山黑河上游多年冻土区不同植被类型土壤有机碳密度分布特征
[J].冰川冻土 ,2018 ,40 (3 ):426 -433 .
[本文引用: 2]
[17]
Wang D Li X X Zou D F et al Modeling soil organic carbon spatial distribution for a complex terrain based on geographically weighted regression in the eastern Qinghai-Tibetan Plateau
[J].Catena ,2020 ,187 (12 ):104399 .
[本文引用: 1]
[18]
Xie M Z Zhao L Wu X D et al Seasonal variations of nitrogen in permafrost-affected soils of the Qinghai-Tibetan Plateau
[J].Catena ,2020 ,195 :104793 .
[本文引用: 1]
[19]
刘鑫 青藏高原多年冻土区冻融循环作用下土壤水热特征研究
[D].兰州 :兰州大学 ,2020 .
[本文引用: 1]
[20]
陈瑞 ,杨梅学 ,万国宁 ,等 基于水热变化的青藏高原土壤冻融过程研究进展
[J].地理科学进展 ,2020 ,39 (11 ):1944 -1958 .
[本文引用: 1]
[21]
鲁博权 冻融作用对大兴安岭森林土壤碳氮矿化的影响
[D].哈尔滨 :哈尔滨师范大学 ,2019 .
[本文引用: 1]
[22]
魏培洁 ,刘放 ,吴明辉 ,等 疏勒河源多年冻土区土壤水溶性有机碳变化特征
[J].草业科学 ,2021 ,38 (4 ):605 -617 .
[本文引用: 7]
[23]
刘放 ,吴明辉 ,魏培洁 ,等 疏勒河源高寒草甸土壤微生物生物量碳氮变化特征
[J].生态学报 ,2020 ,40 (18 ):6416 -6426 .
[本文引用: 4]
[24]
寇丹 青藏高原冻土区土壤氮循环:储量、过程及动态变化
[D].北京 :中国科学院大学 ,2019 .
[本文引用: 1]
[25]
Yuan Z Q Jin H J Wang Q F et al Profile distributions of soil organic carbon fractions in a permafrost region of the Qinghai-Tibet Plateau
[J].Permafrost and Periglacial Processes ,2020 ,31 (4 ):538 -547 .
[本文引用: 1]
[26]
Liu W J Chen S Y Qin X et al Storage,patterns,and control of soil organic carbon and nitrogen in the northeastern margin of the Qinghai-Tibetan Plateau
[J].Environmental Research Letters ,2012 ,7 (3 ):35401 .
[本文引用: 1]
[27]
陈生云 ,刘文杰 ,叶柏生 ,等 疏勒河上游地区植被物种多样性和生物量及其与环境因子的关系
[J].草业学报 ,2011 ,20 (3 ):70 -83 .
[本文引用: 4]
[28]
赵倩 ,刘文杰 ,陈生云 ,等 祁连山疏勒河上游多年冻土区高寒草甸土壤CO2 通量特征
[J].冰川冻土 ,2014 ,36 (6 ):1572 -1581 .
[本文引用: 2]
[29]
Liu W J Chen S Y Zhao Q et al Variation and control of soil organic carbon and other nutrients in permafrost regions on central Qinghai-Tibetan Plateau
[J].Environmental Research Letters ,2014 ,9 (11 ):114013 .
[本文引用: 6]
[30]
牛赟 ,刘贤德 ,赵维俊 ,等 祁连山青海云杉(Picea crassifolia )林浅层土壤碳、氮含量特征及其相互关系
[J].中国沙漠 ,2014 ,34 (2 ):371 -377 .
[本文引用: 1]
[31]
孙小东 ,宁志英 ,杨红玲 ,等 中国北方典型风沙区土壤碳氮磷化学计量特征
[J].中国沙漠 ,2018 ,38 (6 ):1209 -1218 .
[本文引用: 1]
[32]
马维玲 ,石培礼 ,李文华 ,等 青藏高原高寒草甸植株性状和生物量分配的海拔梯度变异
[J].中国科学:生命科学 ,2010 ,40 (6 ):533 -543 .
[本文引用: 1]
[33]
史飞 ,徐梦 ,张旭博 ,等 藏东南色季拉山西坡不同植被土壤有机碳垂直分布特征及其影响因素
[J].植物营养与肥料学报 ,2020 ,26 (5 ):942 -953 .
[本文引用: 1]
[34]
李龙 ,秦富仓 ,姜丽娜 ,等 赤峰市敖汉旗土壤有机碳含量的垂直分布及其影响因素
[J].生态学报 ,2019 ,39 (1 ):345 -354 .
[本文引用: 1]
[35]
王丽华 ,薛晶月 ,谢雨 ,等 不同气候类型下四川草地土壤有机碳空间分布及影响因素
[J].植物生态学报 ,2018 ,42 (3 ):297 -306 .
[本文引用: 1]
[36]
李元寿 ,张人禾 ,王根绪 ,等 青藏高原典型高寒草甸区土壤有机碳氮的变异特性
[J].环境科学 ,2009 ,30 (6 ):1826 -1831 .
[本文引用: 1]
[37]
汪智军 ,梁轩 ,袁道先 岩溶流域不同土壤剖面溶解性碳氮分布和淋失特征
[J].水土保持学报 ,2010 ,24 (6 ):83 -87 ,93 .
[本文引用: 1]
[38]
邓芳博 ,鲍雪莲 ,梁超 ,等 冻融交替对农田氮磷淋溶影响的研究进展
[J].中国生态农业学报(中英文) ,2021 ,29 (1 ):128 -140 .
[本文引用: 1]
[39]
赵锦梅 ,张德罡 ,刘长仲 ,等 祁连山东段高寒地区土地利用方式对土壤性状的影响
[J].生态学报 ,2012 ,32 (2 ):548 -556 .
[本文引用: 1]
[40]
赵林 ,程国栋 ,李述训 ,等 青藏高原五道梁附近多年冻土活动层冻结和融化过程
[J].科学通报 ,2000 ,45 (11 ):1205 -1211 .
[本文引用: 1]
[41]
陈梦棋 ,彭滔 名山县不同土壤类型下有机碳组成和季节性变化研究
[J].安徽农业科学 ,2014 ,42 (33 ):11707 -11710 .
[本文引用: 1]
[42]
Groffman P M Driscoll C T Fahey T J et al Effects of mild winter freezing on soil nitrogen and carbon dynamics in a northern hardwood forest
[J].Biogeochemistry ,2001 ,56 (2 ):191 -213 .
[本文引用: 1]
[43]
Robinson C H Cold adaptation in Arctic and Antarctic fungi
[J].New Phytologist ,2001 ,151 (2 ):341 -353 .
[本文引用: 1]
[44]
黄锦学 ,熊德成 ,刘小飞 ,等 增温对土壤有机碳矿化的影响研究综述
[J].生态学报 ,2017 ,37 (1 ):12 -24 .
[本文引用: 1]
[45]
李云飞 ,谢婷 ,石万里 ,等 腾格里沙漠东南缘人工固沙植被区表层土壤有机碳矿化对凋落物添加的响应
[J].中国沙漠 ,2019 ,39 (5 ):200 -209 .
[本文引用: 1]
[46]
Yang Y H Fang J Y Tang Y H et al Storage,patterns and controls of soil organic carbon in the Tibetan grasslands
[J].Global Change Biology ,2008 ,14 (77 ):1592 -1599 .
[本文引用: 1]
[47]
吴雅琼 ,刘国华 ,傅伯杰 ,等 青藏高原土壤有机碳密度垂直分布研究
[J].环境科学学报 ,2008 ,28 (2 ):362 -367 .
[本文引用: 1]
[48]
陈静 ,李玉霖 ,冯静 ,等 温度和水分对科尔沁沙质草地土壤氮矿化的影响
[J].中国沙漠 ,2016 ,36 (1 ):103 -110 .
[本文引用: 1]
[49]
Yang Y H Fang J Y Guo D L et al Vertical patterns of soil carbon,nitrogen and carbon:nitrogen stoichiometry in Tibetan grasslands
[J].Biogeosciences Discussions ,2010 ,7 (1 ):1 -24 .
[本文引用: 2]
[50]
薛晓娟 ,李英年 ,杜明远 ,等 祁连山东段南麓不同海拔土壤有机质及全氮的分布状况
[J].冰川冻土 ,2009 ,31 (4 ):642 -649 .
[本文引用: 1]
[51]
武小钢 ,郭晋平 ,田旭平 ,等 芦芽山土壤有机碳和全氮沿海拔梯度变化规律
[J].生态环境学报 ,2014 ,23 (1 ):50 -57 .
[本文引用: 1]
氮磷共限制青藏高原高寒草甸生态系统碳吸收
1
2022
... 高寒草甸是青藏高原分布最广的植被类型,约占高原总面积的49.81%[1 ] ,也是多年冻土区的优势植被类型(约占冻土区总面积的80%)[2 ] .近年来,由于气候变暖,高原多年冻土区生态环境变化显著[3 ] .一方面,多年冻土广泛、持续和快速退化[4 ] ,冻土的隔水作用下降,入渗能力增强[5 ] ,地表植被可利用水分及养分流失,导致依赖于冻结层上水的短根系植物枯死[3 ] ,造成其植被盖度下降和组成变化[6 ] ;另一方面,多年冻土融化改变地形地貌,如热熔塌陷和热融沟[7 ] ,不仅会使土壤中冻结的有机质暴露于空气中被持续氧化,甚至导致草地出现大面积“斑块化”现象[8 ] .然而,冻土环境的改变与植被变化并不是相互独立的,而是耦合形成植被-土壤反馈系统协同变化[9 ] .在这个系统中,土壤碳氮作为主要的营养和能源物质[10 ] ,在调节植被生长、改善土壤质量、参与土壤呼吸和养分循环等[11 ] 生物和非生物重要过程中扮演重要角色,其含量和分布受冻土环境变化影响十分显著[7 ,12 ] ,且直接关系到高寒植被的演替趋势[13 ] ,并最终影响整个高原土壤碳氮储藏和多年冻土生态系统碳氮库. ...
Mapping the vegetation distribution of the permafrost zone on the Qinghai-Tibet Plateau
1
2016
... 高寒草甸是青藏高原分布最广的植被类型,约占高原总面积的49.81%[1 ] ,也是多年冻土区的优势植被类型(约占冻土区总面积的80%)[2 ] .近年来,由于气候变暖,高原多年冻土区生态环境变化显著[3 ] .一方面,多年冻土广泛、持续和快速退化[4 ] ,冻土的隔水作用下降,入渗能力增强[5 ] ,地表植被可利用水分及养分流失,导致依赖于冻结层上水的短根系植物枯死[3 ] ,造成其植被盖度下降和组成变化[6 ] ;另一方面,多年冻土融化改变地形地貌,如热熔塌陷和热融沟[7 ] ,不仅会使土壤中冻结的有机质暴露于空气中被持续氧化,甚至导致草地出现大面积“斑块化”现象[8 ] .然而,冻土环境的改变与植被变化并不是相互独立的,而是耦合形成植被-土壤反馈系统协同变化[9 ] .在这个系统中,土壤碳氮作为主要的营养和能源物质[10 ] ,在调节植被生长、改善土壤质量、参与土壤呼吸和养分循环等[11 ] 生物和非生物重要过程中扮演重要角色,其含量和分布受冻土环境变化影响十分显著[7 ,12 ] ,且直接关系到高寒植被的演替趋势[13 ] ,并最终影响整个高原土壤碳氮储藏和多年冻土生态系统碳氮库. ...
青藏高原多年冻土特征、变化及影响
2
2019
... 高寒草甸是青藏高原分布最广的植被类型,约占高原总面积的49.81%[1 ] ,也是多年冻土区的优势植被类型(约占冻土区总面积的80%)[2 ] .近年来,由于气候变暖,高原多年冻土区生态环境变化显著[3 ] .一方面,多年冻土广泛、持续和快速退化[4 ] ,冻土的隔水作用下降,入渗能力增强[5 ] ,地表植被可利用水分及养分流失,导致依赖于冻结层上水的短根系植物枯死[3 ] ,造成其植被盖度下降和组成变化[6 ] ;另一方面,多年冻土融化改变地形地貌,如热熔塌陷和热融沟[7 ] ,不仅会使土壤中冻结的有机质暴露于空气中被持续氧化,甚至导致草地出现大面积“斑块化”现象[8 ] .然而,冻土环境的改变与植被变化并不是相互独立的,而是耦合形成植被-土壤反馈系统协同变化[9 ] .在这个系统中,土壤碳氮作为主要的营养和能源物质[10 ] ,在调节植被生长、改善土壤质量、参与土壤呼吸和养分循环等[11 ] 生物和非生物重要过程中扮演重要角色,其含量和分布受冻土环境变化影响十分显著[7 ,12 ] ,且直接关系到高寒植被的演替趋势[13 ] ,并最终影响整个高原土壤碳氮储藏和多年冻土生态系统碳氮库. ...
... [3 ],造成其植被盖度下降和组成变化[6 ] ;另一方面,多年冻土融化改变地形地貌,如热熔塌陷和热融沟[7 ] ,不仅会使土壤中冻结的有机质暴露于空气中被持续氧化,甚至导致草地出现大面积“斑块化”现象[8 ] .然而,冻土环境的改变与植被变化并不是相互独立的,而是耦合形成植被-土壤反馈系统协同变化[9 ] .在这个系统中,土壤碳氮作为主要的营养和能源物质[10 ] ,在调节植被生长、改善土壤质量、参与土壤呼吸和养分循环等[11 ] 生物和非生物重要过程中扮演重要角色,其含量和分布受冻土环境变化影响十分显著[7 ,12 ] ,且直接关系到高寒植被的演替趋势[13 ] ,并最终影响整个高原土壤碳氮储藏和多年冻土生态系统碳氮库. ...
Degrading permafrost and its impacts
1
2021
... 高寒草甸是青藏高原分布最广的植被类型,约占高原总面积的49.81%[1 ] ,也是多年冻土区的优势植被类型(约占冻土区总面积的80%)[2 ] .近年来,由于气候变暖,高原多年冻土区生态环境变化显著[3 ] .一方面,多年冻土广泛、持续和快速退化[4 ] ,冻土的隔水作用下降,入渗能力增强[5 ] ,地表植被可利用水分及养分流失,导致依赖于冻结层上水的短根系植物枯死[3 ] ,造成其植被盖度下降和组成变化[6 ] ;另一方面,多年冻土融化改变地形地貌,如热熔塌陷和热融沟[7 ] ,不仅会使土壤中冻结的有机质暴露于空气中被持续氧化,甚至导致草地出现大面积“斑块化”现象[8 ] .然而,冻土环境的改变与植被变化并不是相互独立的,而是耦合形成植被-土壤反馈系统协同变化[9 ] .在这个系统中,土壤碳氮作为主要的营养和能源物质[10 ] ,在调节植被生长、改善土壤质量、参与土壤呼吸和养分循环等[11 ] 生物和非生物重要过程中扮演重要角色,其含量和分布受冻土环境变化影响十分显著[7 ,12 ] ,且直接关系到高寒植被的演替趋势[13 ] ,并最终影响整个高原土壤碳氮储藏和多年冻土生态系统碳氮库. ...
青藏高原东北部典型流域冻土退化及其生态水文效应研究
1
2018
... 高寒草甸是青藏高原分布最广的植被类型,约占高原总面积的49.81%[1 ] ,也是多年冻土区的优势植被类型(约占冻土区总面积的80%)[2 ] .近年来,由于气候变暖,高原多年冻土区生态环境变化显著[3 ] .一方面,多年冻土广泛、持续和快速退化[4 ] ,冻土的隔水作用下降,入渗能力增强[5 ] ,地表植被可利用水分及养分流失,导致依赖于冻结层上水的短根系植物枯死[3 ] ,造成其植被盖度下降和组成变化[6 ] ;另一方面,多年冻土融化改变地形地貌,如热熔塌陷和热融沟[7 ] ,不仅会使土壤中冻结的有机质暴露于空气中被持续氧化,甚至导致草地出现大面积“斑块化”现象[8 ] .然而,冻土环境的改变与植被变化并不是相互独立的,而是耦合形成植被-土壤反馈系统协同变化[9 ] .在这个系统中,土壤碳氮作为主要的营养和能源物质[10 ] ,在调节植被生长、改善土壤质量、参与土壤呼吸和养分循环等[11 ] 生物和非生物重要过程中扮演重要角色,其含量和分布受冻土环境变化影响十分显著[7 ,12 ] ,且直接关系到高寒植被的演替趋势[13 ] ,并最终影响整个高原土壤碳氮储藏和多年冻土生态系统碳氮库. ...
Response characteristics of vegetation and soil environment to permafrost degradation in the upstream regions of the Shule River Basin
1
2012
... 高寒草甸是青藏高原分布最广的植被类型,约占高原总面积的49.81%[1 ] ,也是多年冻土区的优势植被类型(约占冻土区总面积的80%)[2 ] .近年来,由于气候变暖,高原多年冻土区生态环境变化显著[3 ] .一方面,多年冻土广泛、持续和快速退化[4 ] ,冻土的隔水作用下降,入渗能力增强[5 ] ,地表植被可利用水分及养分流失,导致依赖于冻结层上水的短根系植物枯死[3 ] ,造成其植被盖度下降和组成变化[6 ] ;另一方面,多年冻土融化改变地形地貌,如热熔塌陷和热融沟[7 ] ,不仅会使土壤中冻结的有机质暴露于空气中被持续氧化,甚至导致草地出现大面积“斑块化”现象[8 ] .然而,冻土环境的改变与植被变化并不是相互独立的,而是耦合形成植被-土壤反馈系统协同变化[9 ] .在这个系统中,土壤碳氮作为主要的营养和能源物质[10 ] ,在调节植被生长、改善土壤质量、参与土壤呼吸和养分循环等[11 ] 生物和非生物重要过程中扮演重要角色,其含量和分布受冻土环境变化影响十分显著[7 ,12 ] ,且直接关系到高寒植被的演替趋势[13 ] ,并最终影响整个高原土壤碳氮储藏和多年冻土生态系统碳氮库. ...
气候变暖对多年冻土区土壤有机碳库的影响
2
2020
... 高寒草甸是青藏高原分布最广的植被类型,约占高原总面积的49.81%[1 ] ,也是多年冻土区的优势植被类型(约占冻土区总面积的80%)[2 ] .近年来,由于气候变暖,高原多年冻土区生态环境变化显著[3 ] .一方面,多年冻土广泛、持续和快速退化[4 ] ,冻土的隔水作用下降,入渗能力增强[5 ] ,地表植被可利用水分及养分流失,导致依赖于冻结层上水的短根系植物枯死[3 ] ,造成其植被盖度下降和组成变化[6 ] ;另一方面,多年冻土融化改变地形地貌,如热熔塌陷和热融沟[7 ] ,不仅会使土壤中冻结的有机质暴露于空气中被持续氧化,甚至导致草地出现大面积“斑块化”现象[8 ] .然而,冻土环境的改变与植被变化并不是相互独立的,而是耦合形成植被-土壤反馈系统协同变化[9 ] .在这个系统中,土壤碳氮作为主要的营养和能源物质[10 ] ,在调节植被生长、改善土壤质量、参与土壤呼吸和养分循环等[11 ] 生物和非生物重要过程中扮演重要角色,其含量和分布受冻土环境变化影响十分显著[7 ,12 ] ,且直接关系到高寒植被的演替趋势[13 ] ,并最终影响整个高原土壤碳氮储藏和多年冻土生态系统碳氮库. ...
... [7 ,12 ],且直接关系到高寒植被的演替趋势[13 ] ,并最终影响整个高原土壤碳氮储藏和多年冻土生态系统碳氮库. ...
青藏高原“黑土滩”退化草地植物和土壤对秃斑面积变化的响应
1
2011
... 高寒草甸是青藏高原分布最广的植被类型,约占高原总面积的49.81%[1 ] ,也是多年冻土区的优势植被类型(约占冻土区总面积的80%)[2 ] .近年来,由于气候变暖,高原多年冻土区生态环境变化显著[3 ] .一方面,多年冻土广泛、持续和快速退化[4 ] ,冻土的隔水作用下降,入渗能力增强[5 ] ,地表植被可利用水分及养分流失,导致依赖于冻结层上水的短根系植物枯死[3 ] ,造成其植被盖度下降和组成变化[6 ] ;另一方面,多年冻土融化改变地形地貌,如热熔塌陷和热融沟[7 ] ,不仅会使土壤中冻结的有机质暴露于空气中被持续氧化,甚至导致草地出现大面积“斑块化”现象[8 ] .然而,冻土环境的改变与植被变化并不是相互独立的,而是耦合形成植被-土壤反馈系统协同变化[9 ] .在这个系统中,土壤碳氮作为主要的营养和能源物质[10 ] ,在调节植被生长、改善土壤质量、参与土壤呼吸和养分循环等[11 ] 生物和非生物重要过程中扮演重要角色,其含量和分布受冻土环境变化影响十分显著[7 ,12 ] ,且直接关系到高寒植被的演替趋势[13 ] ,并最终影响整个高原土壤碳氮储藏和多年冻土生态系统碳氮库. ...
祁连山疏勒河源区冻土退化对土壤微生物生物量碳氮的影响
7
2021
... 高寒草甸是青藏高原分布最广的植被类型,约占高原总面积的49.81%[1 ] ,也是多年冻土区的优势植被类型(约占冻土区总面积的80%)[2 ] .近年来,由于气候变暖,高原多年冻土区生态环境变化显著[3 ] .一方面,多年冻土广泛、持续和快速退化[4 ] ,冻土的隔水作用下降,入渗能力增强[5 ] ,地表植被可利用水分及养分流失,导致依赖于冻结层上水的短根系植物枯死[3 ] ,造成其植被盖度下降和组成变化[6 ] ;另一方面,多年冻土融化改变地形地貌,如热熔塌陷和热融沟[7 ] ,不仅会使土壤中冻结的有机质暴露于空气中被持续氧化,甚至导致草地出现大面积“斑块化”现象[8 ] .然而,冻土环境的改变与植被变化并不是相互独立的,而是耦合形成植被-土壤反馈系统协同变化[9 ] .在这个系统中,土壤碳氮作为主要的营养和能源物质[10 ] ,在调节植被生长、改善土壤质量、参与土壤呼吸和养分循环等[11 ] 生物和非生物重要过程中扮演重要角色,其含量和分布受冻土环境变化影响十分显著[7 ,12 ] ,且直接关系到高寒植被的演替趋势[13 ] ,并最终影响整个高原土壤碳氮储藏和多年冻土生态系统碳氮库. ...
... 位于青藏高原东北缘祁连山西段的疏勒河源区多年冻土广泛发育,其多年冻土面积约占源区面积的73%[9 ] ,源区内优势植被类型为高寒草甸,覆盖面积约为42%[22 ] .近20年观测资料显示,源区冻土退化明显且土壤季节性冻融活动显著[9 , 22 ] .目前该区域的最近土壤碳氮调查资料仅为2012年Liu等[26 ] 对生长季高寒草甸内单一样点的剖面分布研究,缺乏年内季节变化研究.因此,本研究拟开展源区浅层SOC、TN含量在季节尺度上的变化研究,揭示其垂直分布规律及主要的影响因素,并估算疏勒河源多年冻土区高寒草甸土壤碳氮储量,以期为全球气候变化背景下青藏高原多年冻土区高寒草甸土壤碳、氮研究提供数据支撑和科学指导. ...
... [9 , 22 ].目前该区域的最近土壤碳氮调查资料仅为2012年Liu等[26 ] 对生长季高寒草甸内单一样点的剖面分布研究,缺乏年内季节变化研究.因此,本研究拟开展源区浅层SOC、TN含量在季节尺度上的变化研究,揭示其垂直分布规律及主要的影响因素,并估算疏勒河源多年冻土区高寒草甸土壤碳氮储量,以期为全球气候变化背景下青藏高原多年冻土区高寒草甸土壤碳、氮研究提供数据支撑和科学指导. ...
... 疏勒河源区位于青藏高原东北缘祁连山西段,面积约为5 123 km2 ,约占祁连山国家公园青海片区的19.68%.气候类型为大陆性干旱荒漠气候,干冷多风[27 ] .该区域主要植被类型为灌丛、草甸、草原、荒漠及高山稀疏植被,其中高寒草甸面积为2 393.57 km2 ,约占源区总面积的46.72%(图1 ).研究样地位于疏勒河源多年冻土区高寒草甸生态系统综合观测场(38°19′58″N、98°13′44″E,海拔4 036 m,图1 ).基于观测场2017年的气象观测数据资料显示,样地内年均气温为-4.06 ℃,年降水量为466.6 mm且集中在6—9月(图2 ),空气相对湿度为55.49%,年均太阳总辐射为194.3 W·m-2 .0—50 cm土壤砂粒、粉粒和黏粒含量分别为53.80%、35.69%和9.97%.多年冻土类型为阿尔金山-祁连山高寒带山地多年冻土[27 ] ,活动层厚度约为1.5 m,土壤类型为冷钙土.植被覆盖度80%以上,优势植物种有藏嵩草(Kobresia tibetica )和高山嵩草(Kobresia pygmaea )[9 ] . ...
... SOC、TN含量分别用重铬酸钾氧化-外加热法和微量凯氏定氮法测量,C/N则由SOC除以TN所得;pH值和氧化还原电位用酸度计(PHBJ-260)进行测定(水土比为5∶1)[9 ] .地下生物量是将土壤样品先去除杂物再经过2 mm筛洗净,将其与地上生物量放入烘箱80 ℃烘干至恒重称重所得[23 ] .样地0—50 cm土壤理化性质及生物量概况如表1 所示. ...
... SOC含量取决于进入土壤的植物残体量及其土壤微生物分解作用下损失量的平衡,TN含量主要来源于植物残体归还和生物固氮[31 ] .本研究中,SOC、TN剖面分布呈一定规律,0—10 cm层高于其他层,这与以往的研究结果一致[14 ] .首先,地下生物量(根系及其分泌物、细根周转产生的碎屑等)是SOC和TN主要来源[29 ] .研究表明,高海拔植物会将更多的生物量分配在地下部器官[32 ] ,且植被的凋落物在地表形成凋落物层[22 ] ,这决定了SOC、TN剖面分布特征;其次,土壤含水量是影响根系分布和土壤微生物分解的重要环境因子[29 ] .春夏季土壤表层含水量高于下层[22 ] ,促进了植被生产力,进而输入土壤的碳氮增加,且高的水分可能会造成厌氧微生物群落丰度的增加,而厌氧生物对SOC的分解效率低于好氧生物,从而导致SOC的积累[33 ] ,秋冬季节下层含水量是高于表层的[22 ] ,表层土壤为物质交换提供畅通环境,为土壤碳氮的积累、分解和转化创造了良好的基础[34 ] .此外,SOC、TN还与地上生物量显著正相关,与pH显著负相关,这说明地表枯落物对土壤碳、氮的输入是极其重要的,而pH通过影响植被的生长和土壤微生物的活性、数量来间接影响碳、氮的累积[9 ] . ...
... 本研究中,季节变化对C/N有显著的影响,且地下生物量是主要影响因素,可能还与土壤含氧量有关.夏季C/N最高说明夏季微生物分解过程受氮的限制[51 ] ,而碳源充足,更多的SOC转化为土壤微生物碳[9 ] ,较少的TN转化为土壤微生物氮,刘放等[23 ] 的研究也证实,夏季疏勒河源高寒草甸微生物生物量碳最多,而微生物生物量氮含量最少;另一方面,有机质矿化过程中部分碳在土壤呼吸的作用下以CO2 的形式排放至大气,夏季土壤完全解冻,土壤中含氧量充足成为有机质的矿化能力升高的必要条件,有研究表明,疏勒河土壤CO2 通量夏季最高[28 ] .秋季C/N最低,这说明该季节TN含量是相对充足的,有多余的有机氮转化为无机氮储存在土壤中,但土壤微生物同化1份氮的同时需要5份碳构建自身细胞,20份碳作为生命活动的能源,此时可利用的土壤碳却是较少的(图3 ).此外,地下生物量中大多为死根,能提供碳源的根系减少,土壤碳供应不足且土壤呼吸较弱,抑制了有机质矿化. ...
祁连山中段土壤有机碳和氮素的剖面分布
1
2011
... 高寒草甸是青藏高原分布最广的植被类型,约占高原总面积的49.81%[1 ] ,也是多年冻土区的优势植被类型(约占冻土区总面积的80%)[2 ] .近年来,由于气候变暖,高原多年冻土区生态环境变化显著[3 ] .一方面,多年冻土广泛、持续和快速退化[4 ] ,冻土的隔水作用下降,入渗能力增强[5 ] ,地表植被可利用水分及养分流失,导致依赖于冻结层上水的短根系植物枯死[3 ] ,造成其植被盖度下降和组成变化[6 ] ;另一方面,多年冻土融化改变地形地貌,如热熔塌陷和热融沟[7 ] ,不仅会使土壤中冻结的有机质暴露于空气中被持续氧化,甚至导致草地出现大面积“斑块化”现象[8 ] .然而,冻土环境的改变与植被变化并不是相互独立的,而是耦合形成植被-土壤反馈系统协同变化[9 ] .在这个系统中,土壤碳氮作为主要的营养和能源物质[10 ] ,在调节植被生长、改善土壤质量、参与土壤呼吸和养分循环等[11 ] 生物和非生物重要过程中扮演重要角色,其含量和分布受冻土环境变化影响十分显著[7 ,12 ] ,且直接关系到高寒植被的演替趋势[13 ] ,并最终影响整个高原土壤碳氮储藏和多年冻土生态系统碳氮库. ...
祁连山北麓山体垂直带土壤碳氮分布特征及影响因素
1
2009
... 高寒草甸是青藏高原分布最广的植被类型,约占高原总面积的49.81%[1 ] ,也是多年冻土区的优势植被类型(约占冻土区总面积的80%)[2 ] .近年来,由于气候变暖,高原多年冻土区生态环境变化显著[3 ] .一方面,多年冻土广泛、持续和快速退化[4 ] ,冻土的隔水作用下降,入渗能力增强[5 ] ,地表植被可利用水分及养分流失,导致依赖于冻结层上水的短根系植物枯死[3 ] ,造成其植被盖度下降和组成变化[6 ] ;另一方面,多年冻土融化改变地形地貌,如热熔塌陷和热融沟[7 ] ,不仅会使土壤中冻结的有机质暴露于空气中被持续氧化,甚至导致草地出现大面积“斑块化”现象[8 ] .然而,冻土环境的改变与植被变化并不是相互独立的,而是耦合形成植被-土壤反馈系统协同变化[9 ] .在这个系统中,土壤碳氮作为主要的营养和能源物质[10 ] ,在调节植被生长、改善土壤质量、参与土壤呼吸和养分循环等[11 ] 生物和非生物重要过程中扮演重要角色,其含量和分布受冻土环境变化影响十分显著[7 ,12 ] ,且直接关系到高寒植被的演替趋势[13 ] ,并最终影响整个高原土壤碳氮储藏和多年冻土生态系统碳氮库. ...
冻融作用对土壤碳、氮、磷循环的影响
3
2016
... 高寒草甸是青藏高原分布最广的植被类型,约占高原总面积的49.81%[1 ] ,也是多年冻土区的优势植被类型(约占冻土区总面积的80%)[2 ] .近年来,由于气候变暖,高原多年冻土区生态环境变化显著[3 ] .一方面,多年冻土广泛、持续和快速退化[4 ] ,冻土的隔水作用下降,入渗能力增强[5 ] ,地表植被可利用水分及养分流失,导致依赖于冻结层上水的短根系植物枯死[3 ] ,造成其植被盖度下降和组成变化[6 ] ;另一方面,多年冻土融化改变地形地貌,如热熔塌陷和热融沟[7 ] ,不仅会使土壤中冻结的有机质暴露于空气中被持续氧化,甚至导致草地出现大面积“斑块化”现象[8 ] .然而,冻土环境的改变与植被变化并不是相互独立的,而是耦合形成植被-土壤反馈系统协同变化[9 ] .在这个系统中,土壤碳氮作为主要的营养和能源物质[10 ] ,在调节植被生长、改善土壤质量、参与土壤呼吸和养分循环等[11 ] 生物和非生物重要过程中扮演重要角色,其含量和分布受冻土环境变化影响十分显著[7 ,12 ] ,且直接关系到高寒植被的演替趋势[13 ] ,并最终影响整个高原土壤碳氮储藏和多年冻土生态系统碳氮库. ...
... 然而,目前诸多研究仅集中于比较高原多年冻土区不同采样点之间土壤碳氮的含量大小及剖面分布特征.李新星等[14 ] 对马衔山的研究表明,表层土壤有机碳(Soil Organic Carbon, SOC)和全氮(Total Nitrogen, TN)含量高于下层;Zhao等[15 ] 发现土壤碳氮储量随土壤深度增加呈现降低趋势;马素辉等[16 ] 以黑河上游俄博岭各草地类型为研究对象,比较发现高海拔的高寒沼泽草甸SOC密度(Soil Organic Carbon Density, SOCD)最高,而高寒草甸海拔低且SOCD显著降低;Wang等[17 ] 模拟得出,青藏高原东部SOCD空间上由东向西递减;Xie等[18 ] 发现TN含量季节上基本保持稳定,但高寒草甸显著高于其他植被类型.尽管各研究关于多年冻土区土壤碳氮分布具有较高的一致性,但其对于了解高寒区土壤碳氮分布甚至于评估碳氮库储量存在明显的局限性.首先,多年冻土区由于年内温度变化剧烈,浅层土壤存在明显的季节性冻融现象[19 ] ,表现为春秋季冻融频繁,夏冬季则分别完全融化和冻结[20 ] .已有研究表明,季节性冻融会对土壤碳氮矿化速率[21 ] 及土壤水溶性有机碳[22 ] 和微生物碳氮[23 ] 的含量产生显著影响;其次,大量的基于单一时间节点的采样调查忽视了季节变化对土壤碳氮的影响,导致相关研究对土壤碳氮库储量的估计仍存在明显的不确定性.寇丹[24 ] 、Yuan等[25 ] 均是以一年中单采样节点(生长季)对青藏高原、黑河上游多年冻土区土壤碳氮储量进行了评估.然而,非生长季对土壤碳、氮循环影响在年际尺度上是不可忽视的[12 ] .因此,在多年冻土区开展以空间分布和季节变化相结合的生态调查对于准确把握土壤碳氮含量分布及评估碳氮储量具有重要意义. ...
... 土壤TN的季节特征表现为夏季最少,春秋和冬季基本一致.这是由于各季节植被对氮素的消耗不同所致,夏季具有良好的代谢环境(碳源充足、温度适宜),微生物和根系活性高,更多的有机态氮转化为无机态氮[48 ] ,植物生长消耗大量氮素,再加上冻土融化土壤水分增加抑制了氧气进入,加速厌氧环境形成,反硝化过程增强,释放N2 O气体[12 ] ,使得夏季含量低于其他季节,而秋冬季消耗氮速率下降,氮矿化能力也随之降低,土壤中氮能够被累积下来.Liu等[29 ] 的研究表明,北麓河土壤平均TND为0.47±0.07 kg·m-2 ,本研究对应值为0.16±0.02 kg·m-2 ,海拔差异可能是造成两区域差异较大的主要原因.北麓河海拔相比疏勒河更高,据报道,海拔与TN之间具有极显著正相关性[14 ] ,海拔增加、温度下降,土壤中微生物和酶活性下降,分解速率减慢利于TN的积累[13 ] .此外,本研究中SOC、TN含量的季节变化差异不显著,但存在季节变化趋势,这可能与观测时间仅为一年且植被类型单一有关,因而多年冻土区长期的连续观测及不同植被类型下各季节间的差异比较,将是进一步需要探究的问题. ...
高寒草地土壤有机碳影响因子与模拟模型的研究
2
2018
... 高寒草甸是青藏高原分布最广的植被类型,约占高原总面积的49.81%[1 ] ,也是多年冻土区的优势植被类型(约占冻土区总面积的80%)[2 ] .近年来,由于气候变暖,高原多年冻土区生态环境变化显著[3 ] .一方面,多年冻土广泛、持续和快速退化[4 ] ,冻土的隔水作用下降,入渗能力增强[5 ] ,地表植被可利用水分及养分流失,导致依赖于冻结层上水的短根系植物枯死[3 ] ,造成其植被盖度下降和组成变化[6 ] ;另一方面,多年冻土融化改变地形地貌,如热熔塌陷和热融沟[7 ] ,不仅会使土壤中冻结的有机质暴露于空气中被持续氧化,甚至导致草地出现大面积“斑块化”现象[8 ] .然而,冻土环境的改变与植被变化并不是相互独立的,而是耦合形成植被-土壤反馈系统协同变化[9 ] .在这个系统中,土壤碳氮作为主要的营养和能源物质[10 ] ,在调节植被生长、改善土壤质量、参与土壤呼吸和养分循环等[11 ] 生物和非生物重要过程中扮演重要角色,其含量和分布受冻土环境变化影响十分显著[7 ,12 ] ,且直接关系到高寒植被的演替趋势[13 ] ,并最终影响整个高原土壤碳氮储藏和多年冻土生态系统碳氮库. ...
... 土壤TN的季节特征表现为夏季最少,春秋和冬季基本一致.这是由于各季节植被对氮素的消耗不同所致,夏季具有良好的代谢环境(碳源充足、温度适宜),微生物和根系活性高,更多的有机态氮转化为无机态氮[48 ] ,植物生长消耗大量氮素,再加上冻土融化土壤水分增加抑制了氧气进入,加速厌氧环境形成,反硝化过程增强,释放N2 O气体[12 ] ,使得夏季含量低于其他季节,而秋冬季消耗氮速率下降,氮矿化能力也随之降低,土壤中氮能够被累积下来.Liu等[29 ] 的研究表明,北麓河土壤平均TND为0.47±0.07 kg·m-2 ,本研究对应值为0.16±0.02 kg·m-2 ,海拔差异可能是造成两区域差异较大的主要原因.北麓河海拔相比疏勒河更高,据报道,海拔与TN之间具有极显著正相关性[14 ] ,海拔增加、温度下降,土壤中微生物和酶活性下降,分解速率减慢利于TN的积累[13 ] .此外,本研究中SOC、TN含量的季节变化差异不显著,但存在季节变化趋势,这可能与观测时间仅为一年且植被类型单一有关,因而多年冻土区长期的连续观测及不同植被类型下各季节间的差异比较,将是进一步需要探究的问题. ...
马衔山不同海拔土壤碳、氮、磷含量及生态化学计量特征
3
2020
... 然而,目前诸多研究仅集中于比较高原多年冻土区不同采样点之间土壤碳氮的含量大小及剖面分布特征.李新星等[14 ] 对马衔山的研究表明,表层土壤有机碳(Soil Organic Carbon, SOC)和全氮(Total Nitrogen, TN)含量高于下层;Zhao等[15 ] 发现土壤碳氮储量随土壤深度增加呈现降低趋势;马素辉等[16 ] 以黑河上游俄博岭各草地类型为研究对象,比较发现高海拔的高寒沼泽草甸SOC密度(Soil Organic Carbon Density, SOCD)最高,而高寒草甸海拔低且SOCD显著降低;Wang等[17 ] 模拟得出,青藏高原东部SOCD空间上由东向西递减;Xie等[18 ] 发现TN含量季节上基本保持稳定,但高寒草甸显著高于其他植被类型.尽管各研究关于多年冻土区土壤碳氮分布具有较高的一致性,但其对于了解高寒区土壤碳氮分布甚至于评估碳氮库储量存在明显的局限性.首先,多年冻土区由于年内温度变化剧烈,浅层土壤存在明显的季节性冻融现象[19 ] ,表现为春秋季冻融频繁,夏冬季则分别完全融化和冻结[20 ] .已有研究表明,季节性冻融会对土壤碳氮矿化速率[21 ] 及土壤水溶性有机碳[22 ] 和微生物碳氮[23 ] 的含量产生显著影响;其次,大量的基于单一时间节点的采样调查忽视了季节变化对土壤碳氮的影响,导致相关研究对土壤碳氮库储量的估计仍存在明显的不确定性.寇丹[24 ] 、Yuan等[25 ] 均是以一年中单采样节点(生长季)对青藏高原、黑河上游多年冻土区土壤碳氮储量进行了评估.然而,非生长季对土壤碳、氮循环影响在年际尺度上是不可忽视的[12 ] .因此,在多年冻土区开展以空间分布和季节变化相结合的生态调查对于准确把握土壤碳氮含量分布及评估碳氮储量具有重要意义. ...
... SOC含量取决于进入土壤的植物残体量及其土壤微生物分解作用下损失量的平衡,TN含量主要来源于植物残体归还和生物固氮[31 ] .本研究中,SOC、TN剖面分布呈一定规律,0—10 cm层高于其他层,这与以往的研究结果一致[14 ] .首先,地下生物量(根系及其分泌物、细根周转产生的碎屑等)是SOC和TN主要来源[29 ] .研究表明,高海拔植物会将更多的生物量分配在地下部器官[32 ] ,且植被的凋落物在地表形成凋落物层[22 ] ,这决定了SOC、TN剖面分布特征;其次,土壤含水量是影响根系分布和土壤微生物分解的重要环境因子[29 ] .春夏季土壤表层含水量高于下层[22 ] ,促进了植被生产力,进而输入土壤的碳氮增加,且高的水分可能会造成厌氧微生物群落丰度的增加,而厌氧生物对SOC的分解效率低于好氧生物,从而导致SOC的积累[33 ] ,秋冬季节下层含水量是高于表层的[22 ] ,表层土壤为物质交换提供畅通环境,为土壤碳氮的积累、分解和转化创造了良好的基础[34 ] .此外,SOC、TN还与地上生物量显著正相关,与pH显著负相关,这说明地表枯落物对土壤碳、氮的输入是极其重要的,而pH通过影响植被的生长和土壤微生物的活性、数量来间接影响碳、氮的累积[9 ] . ...
... 土壤TN的季节特征表现为夏季最少,春秋和冬季基本一致.这是由于各季节植被对氮素的消耗不同所致,夏季具有良好的代谢环境(碳源充足、温度适宜),微生物和根系活性高,更多的有机态氮转化为无机态氮[48 ] ,植物生长消耗大量氮素,再加上冻土融化土壤水分增加抑制了氧气进入,加速厌氧环境形成,反硝化过程增强,释放N2 O气体[12 ] ,使得夏季含量低于其他季节,而秋冬季消耗氮速率下降,氮矿化能力也随之降低,土壤中氮能够被累积下来.Liu等[29 ] 的研究表明,北麓河土壤平均TND为0.47±0.07 kg·m-2 ,本研究对应值为0.16±0.02 kg·m-2 ,海拔差异可能是造成两区域差异较大的主要原因.北麓河海拔相比疏勒河更高,据报道,海拔与TN之间具有极显著正相关性[14 ] ,海拔增加、温度下降,土壤中微生物和酶活性下降,分解速率减慢利于TN的积累[13 ] .此外,本研究中SOC、TN含量的季节变化差异不显著,但存在季节变化趋势,这可能与观测时间仅为一年且植被类型单一有关,因而多年冻土区长期的连续观测及不同植被类型下各季节间的差异比较,将是进一步需要探究的问题. ...
Soil organic carbon and total nitrogen pools in permafrost zones of the Qinghai-Tibetan Plateau
1
2018
... 然而,目前诸多研究仅集中于比较高原多年冻土区不同采样点之间土壤碳氮的含量大小及剖面分布特征.李新星等[14 ] 对马衔山的研究表明,表层土壤有机碳(Soil Organic Carbon, SOC)和全氮(Total Nitrogen, TN)含量高于下层;Zhao等[15 ] 发现土壤碳氮储量随土壤深度增加呈现降低趋势;马素辉等[16 ] 以黑河上游俄博岭各草地类型为研究对象,比较发现高海拔的高寒沼泽草甸SOC密度(Soil Organic Carbon Density, SOCD)最高,而高寒草甸海拔低且SOCD显著降低;Wang等[17 ] 模拟得出,青藏高原东部SOCD空间上由东向西递减;Xie等[18 ] 发现TN含量季节上基本保持稳定,但高寒草甸显著高于其他植被类型.尽管各研究关于多年冻土区土壤碳氮分布具有较高的一致性,但其对于了解高寒区土壤碳氮分布甚至于评估碳氮库储量存在明显的局限性.首先,多年冻土区由于年内温度变化剧烈,浅层土壤存在明显的季节性冻融现象[19 ] ,表现为春秋季冻融频繁,夏冬季则分别完全融化和冻结[20 ] .已有研究表明,季节性冻融会对土壤碳氮矿化速率[21 ] 及土壤水溶性有机碳[22 ] 和微生物碳氮[23 ] 的含量产生显著影响;其次,大量的基于单一时间节点的采样调查忽视了季节变化对土壤碳氮的影响,导致相关研究对土壤碳氮库储量的估计仍存在明显的不确定性.寇丹[24 ] 、Yuan等[25 ] 均是以一年中单采样节点(生长季)对青藏高原、黑河上游多年冻土区土壤碳氮储量进行了评估.然而,非生长季对土壤碳、氮循环影响在年际尺度上是不可忽视的[12 ] .因此,在多年冻土区开展以空间分布和季节变化相结合的生态调查对于准确把握土壤碳氮含量分布及评估碳氮储量具有重要意义. ...
祁连山黑河上游多年冻土区不同植被类型土壤有机碳密度分布特征
2
2018
... 然而,目前诸多研究仅集中于比较高原多年冻土区不同采样点之间土壤碳氮的含量大小及剖面分布特征.李新星等[14 ] 对马衔山的研究表明,表层土壤有机碳(Soil Organic Carbon, SOC)和全氮(Total Nitrogen, TN)含量高于下层;Zhao等[15 ] 发现土壤碳氮储量随土壤深度增加呈现降低趋势;马素辉等[16 ] 以黑河上游俄博岭各草地类型为研究对象,比较发现高海拔的高寒沼泽草甸SOC密度(Soil Organic Carbon Density, SOCD)最高,而高寒草甸海拔低且SOCD显著降低;Wang等[17 ] 模拟得出,青藏高原东部SOCD空间上由东向西递减;Xie等[18 ] 发现TN含量季节上基本保持稳定,但高寒草甸显著高于其他植被类型.尽管各研究关于多年冻土区土壤碳氮分布具有较高的一致性,但其对于了解高寒区土壤碳氮分布甚至于评估碳氮库储量存在明显的局限性.首先,多年冻土区由于年内温度变化剧烈,浅层土壤存在明显的季节性冻融现象[19 ] ,表现为春秋季冻融频繁,夏冬季则分别完全融化和冻结[20 ] .已有研究表明,季节性冻融会对土壤碳氮矿化速率[21 ] 及土壤水溶性有机碳[22 ] 和微生物碳氮[23 ] 的含量产生显著影响;其次,大量的基于单一时间节点的采样调查忽视了季节变化对土壤碳氮的影响,导致相关研究对土壤碳氮库储量的估计仍存在明显的不确定性.寇丹[24 ] 、Yuan等[25 ] 均是以一年中单采样节点(生长季)对青藏高原、黑河上游多年冻土区土壤碳氮储量进行了评估.然而,非生长季对土壤碳、氮循环影响在年际尺度上是不可忽视的[12 ] .因此,在多年冻土区开展以空间分布和季节变化相结合的生态调查对于准确把握土壤碳氮含量分布及评估碳氮储量具有重要意义. ...
... 生物量及环境因素的季节变化使SOC含量具有季节差异.陈梦棋等[41 ] 对比名山县生长季各植被类型SOC发现,其存在差异但不显著,这与本研究的结果相符.本研究表明,影响SOC季节差异的主控因子是地下生物量,这表明碳输入量的不同会影响SOC含量大小,但变幅相对较小.春季植被返青期主要消耗上一年份累积的土壤养分来度过[42 ] ,加之土壤融化水分和温度增加促进微生物活性并消耗碳源,因而春季含量低于冬季;夏季优越的水热条件植被生长旺盛,强烈的光合作用使根系和根茎固定了更多的碳,大量的有机物质(凋落物、根系分泌物及其脱落物等)进入土壤并转化为SOC,与此同时土壤微生物的新陈代谢虽会消耗一部分SOC,但SOC积累量远大于消耗量,表现为夏季最高;秋季地表枯黄产生凋落物、根系衰减及动物残体(气温骤降大量细菌因低温环境休眠或死亡)等还未来得及被分解而以腐殖质的形式储存在土体中,致秋季最低;冬季植被凋亡,地下生物量急剧降低,但来源于秋季的大量腐殖质为适应低温环境而幸存下来的真菌(微生物群体中约87%的细菌休眠或死亡)微生物提供了较为丰富的代谢底物[43 ] ,再加上积雪覆盖在土壤表面起隔热作用[23 ] ,以及低气温抑制土壤呼吸,减缓有机质分解速率[44 ] ,SOC积累胜过消耗,冬季高于秋季.此外,生长季温度高,土壤酶催化过程以及微生物代谢过程均加速[45 ] ,从而加快SOC矿化过程,这可能是造成各季节间SOC含量差异不显著的原因之一.本研究中,年均SOCD为8.49±0.25 kg·m-2 ,低于黑河上游多年冻土区高寒草甸以生长季采样所得结果(11.22 kg·m-2 )[16 ] ,可能原因是影响其变化的因素如植被盖度、凋落物、土壤呼吸速率因所处环境的不同均存在差异[46 -47 ] .本研究通过全年采样得出,夏季SOCD比年平均密度约高5.77%,可见,利用生长盛季估算全年SOCD及储量会存在高估的趋势. ...
Modeling soil organic carbon spatial distribution for a complex terrain based on geographically weighted regression in the eastern Qinghai-Tibetan Plateau
1
2020
... 然而,目前诸多研究仅集中于比较高原多年冻土区不同采样点之间土壤碳氮的含量大小及剖面分布特征.李新星等[14 ] 对马衔山的研究表明,表层土壤有机碳(Soil Organic Carbon, SOC)和全氮(Total Nitrogen, TN)含量高于下层;Zhao等[15 ] 发现土壤碳氮储量随土壤深度增加呈现降低趋势;马素辉等[16 ] 以黑河上游俄博岭各草地类型为研究对象,比较发现高海拔的高寒沼泽草甸SOC密度(Soil Organic Carbon Density, SOCD)最高,而高寒草甸海拔低且SOCD显著降低;Wang等[17 ] 模拟得出,青藏高原东部SOCD空间上由东向西递减;Xie等[18 ] 发现TN含量季节上基本保持稳定,但高寒草甸显著高于其他植被类型.尽管各研究关于多年冻土区土壤碳氮分布具有较高的一致性,但其对于了解高寒区土壤碳氮分布甚至于评估碳氮库储量存在明显的局限性.首先,多年冻土区由于年内温度变化剧烈,浅层土壤存在明显的季节性冻融现象[19 ] ,表现为春秋季冻融频繁,夏冬季则分别完全融化和冻结[20 ] .已有研究表明,季节性冻融会对土壤碳氮矿化速率[21 ] 及土壤水溶性有机碳[22 ] 和微生物碳氮[23 ] 的含量产生显著影响;其次,大量的基于单一时间节点的采样调查忽视了季节变化对土壤碳氮的影响,导致相关研究对土壤碳氮库储量的估计仍存在明显的不确定性.寇丹[24 ] 、Yuan等[25 ] 均是以一年中单采样节点(生长季)对青藏高原、黑河上游多年冻土区土壤碳氮储量进行了评估.然而,非生长季对土壤碳、氮循环影响在年际尺度上是不可忽视的[12 ] .因此,在多年冻土区开展以空间分布和季节变化相结合的生态调查对于准确把握土壤碳氮含量分布及评估碳氮储量具有重要意义. ...
Seasonal variations of nitrogen in permafrost-affected soils of the Qinghai-Tibetan Plateau
1
2020
... 然而,目前诸多研究仅集中于比较高原多年冻土区不同采样点之间土壤碳氮的含量大小及剖面分布特征.李新星等[14 ] 对马衔山的研究表明,表层土壤有机碳(Soil Organic Carbon, SOC)和全氮(Total Nitrogen, TN)含量高于下层;Zhao等[15 ] 发现土壤碳氮储量随土壤深度增加呈现降低趋势;马素辉等[16 ] 以黑河上游俄博岭各草地类型为研究对象,比较发现高海拔的高寒沼泽草甸SOC密度(Soil Organic Carbon Density, SOCD)最高,而高寒草甸海拔低且SOCD显著降低;Wang等[17 ] 模拟得出,青藏高原东部SOCD空间上由东向西递减;Xie等[18 ] 发现TN含量季节上基本保持稳定,但高寒草甸显著高于其他植被类型.尽管各研究关于多年冻土区土壤碳氮分布具有较高的一致性,但其对于了解高寒区土壤碳氮分布甚至于评估碳氮库储量存在明显的局限性.首先,多年冻土区由于年内温度变化剧烈,浅层土壤存在明显的季节性冻融现象[19 ] ,表现为春秋季冻融频繁,夏冬季则分别完全融化和冻结[20 ] .已有研究表明,季节性冻融会对土壤碳氮矿化速率[21 ] 及土壤水溶性有机碳[22 ] 和微生物碳氮[23 ] 的含量产生显著影响;其次,大量的基于单一时间节点的采样调查忽视了季节变化对土壤碳氮的影响,导致相关研究对土壤碳氮库储量的估计仍存在明显的不确定性.寇丹[24 ] 、Yuan等[25 ] 均是以一年中单采样节点(生长季)对青藏高原、黑河上游多年冻土区土壤碳氮储量进行了评估.然而,非生长季对土壤碳、氮循环影响在年际尺度上是不可忽视的[12 ] .因此,在多年冻土区开展以空间分布和季节变化相结合的生态调查对于准确把握土壤碳氮含量分布及评估碳氮储量具有重要意义. ...
青藏高原多年冻土区冻融循环作用下土壤水热特征研究
1
2020
... 然而,目前诸多研究仅集中于比较高原多年冻土区不同采样点之间土壤碳氮的含量大小及剖面分布特征.李新星等[14 ] 对马衔山的研究表明,表层土壤有机碳(Soil Organic Carbon, SOC)和全氮(Total Nitrogen, TN)含量高于下层;Zhao等[15 ] 发现土壤碳氮储量随土壤深度增加呈现降低趋势;马素辉等[16 ] 以黑河上游俄博岭各草地类型为研究对象,比较发现高海拔的高寒沼泽草甸SOC密度(Soil Organic Carbon Density, SOCD)最高,而高寒草甸海拔低且SOCD显著降低;Wang等[17 ] 模拟得出,青藏高原东部SOCD空间上由东向西递减;Xie等[18 ] 发现TN含量季节上基本保持稳定,但高寒草甸显著高于其他植被类型.尽管各研究关于多年冻土区土壤碳氮分布具有较高的一致性,但其对于了解高寒区土壤碳氮分布甚至于评估碳氮库储量存在明显的局限性.首先,多年冻土区由于年内温度变化剧烈,浅层土壤存在明显的季节性冻融现象[19 ] ,表现为春秋季冻融频繁,夏冬季则分别完全融化和冻结[20 ] .已有研究表明,季节性冻融会对土壤碳氮矿化速率[21 ] 及土壤水溶性有机碳[22 ] 和微生物碳氮[23 ] 的含量产生显著影响;其次,大量的基于单一时间节点的采样调查忽视了季节变化对土壤碳氮的影响,导致相关研究对土壤碳氮库储量的估计仍存在明显的不确定性.寇丹[24 ] 、Yuan等[25 ] 均是以一年中单采样节点(生长季)对青藏高原、黑河上游多年冻土区土壤碳氮储量进行了评估.然而,非生长季对土壤碳、氮循环影响在年际尺度上是不可忽视的[12 ] .因此,在多年冻土区开展以空间分布和季节变化相结合的生态调查对于准确把握土壤碳氮含量分布及评估碳氮储量具有重要意义. ...
基于水热变化的青藏高原土壤冻融过程研究进展
1
2020
... 然而,目前诸多研究仅集中于比较高原多年冻土区不同采样点之间土壤碳氮的含量大小及剖面分布特征.李新星等[14 ] 对马衔山的研究表明,表层土壤有机碳(Soil Organic Carbon, SOC)和全氮(Total Nitrogen, TN)含量高于下层;Zhao等[15 ] 发现土壤碳氮储量随土壤深度增加呈现降低趋势;马素辉等[16 ] 以黑河上游俄博岭各草地类型为研究对象,比较发现高海拔的高寒沼泽草甸SOC密度(Soil Organic Carbon Density, SOCD)最高,而高寒草甸海拔低且SOCD显著降低;Wang等[17 ] 模拟得出,青藏高原东部SOCD空间上由东向西递减;Xie等[18 ] 发现TN含量季节上基本保持稳定,但高寒草甸显著高于其他植被类型.尽管各研究关于多年冻土区土壤碳氮分布具有较高的一致性,但其对于了解高寒区土壤碳氮分布甚至于评估碳氮库储量存在明显的局限性.首先,多年冻土区由于年内温度变化剧烈,浅层土壤存在明显的季节性冻融现象[19 ] ,表现为春秋季冻融频繁,夏冬季则分别完全融化和冻结[20 ] .已有研究表明,季节性冻融会对土壤碳氮矿化速率[21 ] 及土壤水溶性有机碳[22 ] 和微生物碳氮[23 ] 的含量产生显著影响;其次,大量的基于单一时间节点的采样调查忽视了季节变化对土壤碳氮的影响,导致相关研究对土壤碳氮库储量的估计仍存在明显的不确定性.寇丹[24 ] 、Yuan等[25 ] 均是以一年中单采样节点(生长季)对青藏高原、黑河上游多年冻土区土壤碳氮储量进行了评估.然而,非生长季对土壤碳、氮循环影响在年际尺度上是不可忽视的[12 ] .因此,在多年冻土区开展以空间分布和季节变化相结合的生态调查对于准确把握土壤碳氮含量分布及评估碳氮储量具有重要意义. ...
冻融作用对大兴安岭森林土壤碳氮矿化的影响
1
2019
... 然而,目前诸多研究仅集中于比较高原多年冻土区不同采样点之间土壤碳氮的含量大小及剖面分布特征.李新星等[14 ] 对马衔山的研究表明,表层土壤有机碳(Soil Organic Carbon, SOC)和全氮(Total Nitrogen, TN)含量高于下层;Zhao等[15 ] 发现土壤碳氮储量随土壤深度增加呈现降低趋势;马素辉等[16 ] 以黑河上游俄博岭各草地类型为研究对象,比较发现高海拔的高寒沼泽草甸SOC密度(Soil Organic Carbon Density, SOCD)最高,而高寒草甸海拔低且SOCD显著降低;Wang等[17 ] 模拟得出,青藏高原东部SOCD空间上由东向西递减;Xie等[18 ] 发现TN含量季节上基本保持稳定,但高寒草甸显著高于其他植被类型.尽管各研究关于多年冻土区土壤碳氮分布具有较高的一致性,但其对于了解高寒区土壤碳氮分布甚至于评估碳氮库储量存在明显的局限性.首先,多年冻土区由于年内温度变化剧烈,浅层土壤存在明显的季节性冻融现象[19 ] ,表现为春秋季冻融频繁,夏冬季则分别完全融化和冻结[20 ] .已有研究表明,季节性冻融会对土壤碳氮矿化速率[21 ] 及土壤水溶性有机碳[22 ] 和微生物碳氮[23 ] 的含量产生显著影响;其次,大量的基于单一时间节点的采样调查忽视了季节变化对土壤碳氮的影响,导致相关研究对土壤碳氮库储量的估计仍存在明显的不确定性.寇丹[24 ] 、Yuan等[25 ] 均是以一年中单采样节点(生长季)对青藏高原、黑河上游多年冻土区土壤碳氮储量进行了评估.然而,非生长季对土壤碳、氮循环影响在年际尺度上是不可忽视的[12 ] .因此,在多年冻土区开展以空间分布和季节变化相结合的生态调查对于准确把握土壤碳氮含量分布及评估碳氮储量具有重要意义. ...
疏勒河源多年冻土区土壤水溶性有机碳变化特征
7
2021
... 然而,目前诸多研究仅集中于比较高原多年冻土区不同采样点之间土壤碳氮的含量大小及剖面分布特征.李新星等[14 ] 对马衔山的研究表明,表层土壤有机碳(Soil Organic Carbon, SOC)和全氮(Total Nitrogen, TN)含量高于下层;Zhao等[15 ] 发现土壤碳氮储量随土壤深度增加呈现降低趋势;马素辉等[16 ] 以黑河上游俄博岭各草地类型为研究对象,比较发现高海拔的高寒沼泽草甸SOC密度(Soil Organic Carbon Density, SOCD)最高,而高寒草甸海拔低且SOCD显著降低;Wang等[17 ] 模拟得出,青藏高原东部SOCD空间上由东向西递减;Xie等[18 ] 发现TN含量季节上基本保持稳定,但高寒草甸显著高于其他植被类型.尽管各研究关于多年冻土区土壤碳氮分布具有较高的一致性,但其对于了解高寒区土壤碳氮分布甚至于评估碳氮库储量存在明显的局限性.首先,多年冻土区由于年内温度变化剧烈,浅层土壤存在明显的季节性冻融现象[19 ] ,表现为春秋季冻融频繁,夏冬季则分别完全融化和冻结[20 ] .已有研究表明,季节性冻融会对土壤碳氮矿化速率[21 ] 及土壤水溶性有机碳[22 ] 和微生物碳氮[23 ] 的含量产生显著影响;其次,大量的基于单一时间节点的采样调查忽视了季节变化对土壤碳氮的影响,导致相关研究对土壤碳氮库储量的估计仍存在明显的不确定性.寇丹[24 ] 、Yuan等[25 ] 均是以一年中单采样节点(生长季)对青藏高原、黑河上游多年冻土区土壤碳氮储量进行了评估.然而,非生长季对土壤碳、氮循环影响在年际尺度上是不可忽视的[12 ] .因此,在多年冻土区开展以空间分布和季节变化相结合的生态调查对于准确把握土壤碳氮含量分布及评估碳氮储量具有重要意义. ...
... 位于青藏高原东北缘祁连山西段的疏勒河源区多年冻土广泛发育,其多年冻土面积约占源区面积的73%[9 ] ,源区内优势植被类型为高寒草甸,覆盖面积约为42%[22 ] .近20年观测资料显示,源区冻土退化明显且土壤季节性冻融活动显著[9 , 22 ] .目前该区域的最近土壤碳氮调查资料仅为2012年Liu等[26 ] 对生长季高寒草甸内单一样点的剖面分布研究,缺乏年内季节变化研究.因此,本研究拟开展源区浅层SOC、TN含量在季节尺度上的变化研究,揭示其垂直分布规律及主要的影响因素,并估算疏勒河源多年冻土区高寒草甸土壤碳氮储量,以期为全球气候变化背景下青藏高原多年冻土区高寒草甸土壤碳、氮研究提供数据支撑和科学指导. ...
... , 22 ].目前该区域的最近土壤碳氮调查资料仅为2012年Liu等[26 ] 对生长季高寒草甸内单一样点的剖面分布研究,缺乏年内季节变化研究.因此,本研究拟开展源区浅层SOC、TN含量在季节尺度上的变化研究,揭示其垂直分布规律及主要的影响因素,并估算疏勒河源多年冻土区高寒草甸土壤碳氮储量,以期为全球气候变化背景下青藏高原多年冻土区高寒草甸土壤碳、氮研究提供数据支撑和科学指导. ...
... SOC含量取决于进入土壤的植物残体量及其土壤微生物分解作用下损失量的平衡,TN含量主要来源于植物残体归还和生物固氮[31 ] .本研究中,SOC、TN剖面分布呈一定规律,0—10 cm层高于其他层,这与以往的研究结果一致[14 ] .首先,地下生物量(根系及其分泌物、细根周转产生的碎屑等)是SOC和TN主要来源[29 ] .研究表明,高海拔植物会将更多的生物量分配在地下部器官[32 ] ,且植被的凋落物在地表形成凋落物层[22 ] ,这决定了SOC、TN剖面分布特征;其次,土壤含水量是影响根系分布和土壤微生物分解的重要环境因子[29 ] .春夏季土壤表层含水量高于下层[22 ] ,促进了植被生产力,进而输入土壤的碳氮增加,且高的水分可能会造成厌氧微生物群落丰度的增加,而厌氧生物对SOC的分解效率低于好氧生物,从而导致SOC的积累[33 ] ,秋冬季节下层含水量是高于表层的[22 ] ,表层土壤为物质交换提供畅通环境,为土壤碳氮的积累、分解和转化创造了良好的基础[34 ] .此外,SOC、TN还与地上生物量显著正相关,与pH显著负相关,这说明地表枯落物对土壤碳、氮的输入是极其重要的,而pH通过影响植被的生长和土壤微生物的活性、数量来间接影响碳、氮的累积[9 ] . ...
... [22 ],促进了植被生产力,进而输入土壤的碳氮增加,且高的水分可能会造成厌氧微生物群落丰度的增加,而厌氧生物对SOC的分解效率低于好氧生物,从而导致SOC的积累[33 ] ,秋冬季节下层含水量是高于表层的[22 ] ,表层土壤为物质交换提供畅通环境,为土壤碳氮的积累、分解和转化创造了良好的基础[34 ] .此外,SOC、TN还与地上生物量显著正相关,与pH显著负相关,这说明地表枯落物对土壤碳、氮的输入是极其重要的,而pH通过影响植被的生长和土壤微生物的活性、数量来间接影响碳、氮的累积[9 ] . ...
... [22 ],表层土壤为物质交换提供畅通环境,为土壤碳氮的积累、分解和转化创造了良好的基础[34 ] .此外,SOC、TN还与地上生物量显著正相关,与pH显著负相关,这说明地表枯落物对土壤碳、氮的输入是极其重要的,而pH通过影响植被的生长和土壤微生物的活性、数量来间接影响碳、氮的累积[9 ] . ...
... 此外,SOC和TN的垂直分布还因季节不同产生差异,即随土壤深度增加,秋季0—50 cm内呈递减趋势,春、夏季和冬季仅在0—30 cm内递减,30—50 cm含量有所回升,这种递减趋势可能与该区域植被根系的分布在0—30 cm有关[27 ] ,而季节差异则可能是淋溶过程中养分从表层向下层迁移的动态结果[35 ] .高寒草甸下发育着的土壤,质地为沙壤,养分易淋溶[36 ] ,表层的土壤碳氮会不同比例地向下层流失[37 ] .但是,这种养分向深层的运移通道会因季节性冻融过程和融化深度不同而产生差异.冻融过程和融化深度主要通过改变土壤水热动态、孔隙结构及根系与土壤间的缝隙促进或抑制大孔隙流形成[38 ] .具体来看,除秋季外,春夏冬季均在30—50 cm层SOC、TN含量有升高,这是由于30 cm层以下根系分布少且土壤紧实[39 ] ,一定程度上抑制了养分向下迁移的速率,并且冻土活动层在秋季达到最大融化深度[40 ] ,且中层土壤是高能水区[22 ] ,可能促进了表层SOC、TN持续向下层(30—50 cm)淋溶,因而秋季表现出空间的连续性降低. ...
疏勒河源高寒草甸土壤微生物生物量碳氮变化特征
4
2020
... 然而,目前诸多研究仅集中于比较高原多年冻土区不同采样点之间土壤碳氮的含量大小及剖面分布特征.李新星等[14 ] 对马衔山的研究表明,表层土壤有机碳(Soil Organic Carbon, SOC)和全氮(Total Nitrogen, TN)含量高于下层;Zhao等[15 ] 发现土壤碳氮储量随土壤深度增加呈现降低趋势;马素辉等[16 ] 以黑河上游俄博岭各草地类型为研究对象,比较发现高海拔的高寒沼泽草甸SOC密度(Soil Organic Carbon Density, SOCD)最高,而高寒草甸海拔低且SOCD显著降低;Wang等[17 ] 模拟得出,青藏高原东部SOCD空间上由东向西递减;Xie等[18 ] 发现TN含量季节上基本保持稳定,但高寒草甸显著高于其他植被类型.尽管各研究关于多年冻土区土壤碳氮分布具有较高的一致性,但其对于了解高寒区土壤碳氮分布甚至于评估碳氮库储量存在明显的局限性.首先,多年冻土区由于年内温度变化剧烈,浅层土壤存在明显的季节性冻融现象[19 ] ,表现为春秋季冻融频繁,夏冬季则分别完全融化和冻结[20 ] .已有研究表明,季节性冻融会对土壤碳氮矿化速率[21 ] 及土壤水溶性有机碳[22 ] 和微生物碳氮[23 ] 的含量产生显著影响;其次,大量的基于单一时间节点的采样调查忽视了季节变化对土壤碳氮的影响,导致相关研究对土壤碳氮库储量的估计仍存在明显的不确定性.寇丹[24 ] 、Yuan等[25 ] 均是以一年中单采样节点(生长季)对青藏高原、黑河上游多年冻土区土壤碳氮储量进行了评估.然而,非生长季对土壤碳、氮循环影响在年际尺度上是不可忽视的[12 ] .因此,在多年冻土区开展以空间分布和季节变化相结合的生态调查对于准确把握土壤碳氮含量分布及评估碳氮储量具有重要意义. ...
... SOC、TN含量分别用重铬酸钾氧化-外加热法和微量凯氏定氮法测量,C/N则由SOC除以TN所得;pH值和氧化还原电位用酸度计(PHBJ-260)进行测定(水土比为5∶1)[9 ] .地下生物量是将土壤样品先去除杂物再经过2 mm筛洗净,将其与地上生物量放入烘箱80 ℃烘干至恒重称重所得[23 ] .样地0—50 cm土壤理化性质及生物量概况如表1 所示. ...
... 生物量及环境因素的季节变化使SOC含量具有季节差异.陈梦棋等[41 ] 对比名山县生长季各植被类型SOC发现,其存在差异但不显著,这与本研究的结果相符.本研究表明,影响SOC季节差异的主控因子是地下生物量,这表明碳输入量的不同会影响SOC含量大小,但变幅相对较小.春季植被返青期主要消耗上一年份累积的土壤养分来度过[42 ] ,加之土壤融化水分和温度增加促进微生物活性并消耗碳源,因而春季含量低于冬季;夏季优越的水热条件植被生长旺盛,强烈的光合作用使根系和根茎固定了更多的碳,大量的有机物质(凋落物、根系分泌物及其脱落物等)进入土壤并转化为SOC,与此同时土壤微生物的新陈代谢虽会消耗一部分SOC,但SOC积累量远大于消耗量,表现为夏季最高;秋季地表枯黄产生凋落物、根系衰减及动物残体(气温骤降大量细菌因低温环境休眠或死亡)等还未来得及被分解而以腐殖质的形式储存在土体中,致秋季最低;冬季植被凋亡,地下生物量急剧降低,但来源于秋季的大量腐殖质为适应低温环境而幸存下来的真菌(微生物群体中约87%的细菌休眠或死亡)微生物提供了较为丰富的代谢底物[43 ] ,再加上积雪覆盖在土壤表面起隔热作用[23 ] ,以及低气温抑制土壤呼吸,减缓有机质分解速率[44 ] ,SOC积累胜过消耗,冬季高于秋季.此外,生长季温度高,土壤酶催化过程以及微生物代谢过程均加速[45 ] ,从而加快SOC矿化过程,这可能是造成各季节间SOC含量差异不显著的原因之一.本研究中,年均SOCD为8.49±0.25 kg·m-2 ,低于黑河上游多年冻土区高寒草甸以生长季采样所得结果(11.22 kg·m-2 )[16 ] ,可能原因是影响其变化的因素如植被盖度、凋落物、土壤呼吸速率因所处环境的不同均存在差异[46 -47 ] .本研究通过全年采样得出,夏季SOCD比年平均密度约高5.77%,可见,利用生长盛季估算全年SOCD及储量会存在高估的趋势. ...
... 本研究中,季节变化对C/N有显著的影响,且地下生物量是主要影响因素,可能还与土壤含氧量有关.夏季C/N最高说明夏季微生物分解过程受氮的限制[51 ] ,而碳源充足,更多的SOC转化为土壤微生物碳[9 ] ,较少的TN转化为土壤微生物氮,刘放等[23 ] 的研究也证实,夏季疏勒河源高寒草甸微生物生物量碳最多,而微生物生物量氮含量最少;另一方面,有机质矿化过程中部分碳在土壤呼吸的作用下以CO2 的形式排放至大气,夏季土壤完全解冻,土壤中含氧量充足成为有机质的矿化能力升高的必要条件,有研究表明,疏勒河土壤CO2 通量夏季最高[28 ] .秋季C/N最低,这说明该季节TN含量是相对充足的,有多余的有机氮转化为无机氮储存在土壤中,但土壤微生物同化1份氮的同时需要5份碳构建自身细胞,20份碳作为生命活动的能源,此时可利用的土壤碳却是较少的(图3 ).此外,地下生物量中大多为死根,能提供碳源的根系减少,土壤碳供应不足且土壤呼吸较弱,抑制了有机质矿化. ...
青藏高原冻土区土壤氮循环:储量、过程及动态变化
1
2019
... 然而,目前诸多研究仅集中于比较高原多年冻土区不同采样点之间土壤碳氮的含量大小及剖面分布特征.李新星等[14 ] 对马衔山的研究表明,表层土壤有机碳(Soil Organic Carbon, SOC)和全氮(Total Nitrogen, TN)含量高于下层;Zhao等[15 ] 发现土壤碳氮储量随土壤深度增加呈现降低趋势;马素辉等[16 ] 以黑河上游俄博岭各草地类型为研究对象,比较发现高海拔的高寒沼泽草甸SOC密度(Soil Organic Carbon Density, SOCD)最高,而高寒草甸海拔低且SOCD显著降低;Wang等[17 ] 模拟得出,青藏高原东部SOCD空间上由东向西递减;Xie等[18 ] 发现TN含量季节上基本保持稳定,但高寒草甸显著高于其他植被类型.尽管各研究关于多年冻土区土壤碳氮分布具有较高的一致性,但其对于了解高寒区土壤碳氮分布甚至于评估碳氮库储量存在明显的局限性.首先,多年冻土区由于年内温度变化剧烈,浅层土壤存在明显的季节性冻融现象[19 ] ,表现为春秋季冻融频繁,夏冬季则分别完全融化和冻结[20 ] .已有研究表明,季节性冻融会对土壤碳氮矿化速率[21 ] 及土壤水溶性有机碳[22 ] 和微生物碳氮[23 ] 的含量产生显著影响;其次,大量的基于单一时间节点的采样调查忽视了季节变化对土壤碳氮的影响,导致相关研究对土壤碳氮库储量的估计仍存在明显的不确定性.寇丹[24 ] 、Yuan等[25 ] 均是以一年中单采样节点(生长季)对青藏高原、黑河上游多年冻土区土壤碳氮储量进行了评估.然而,非生长季对土壤碳、氮循环影响在年际尺度上是不可忽视的[12 ] .因此,在多年冻土区开展以空间分布和季节变化相结合的生态调查对于准确把握土壤碳氮含量分布及评估碳氮储量具有重要意义. ...
Profile distributions of soil organic carbon fractions in a permafrost region of the Qinghai-Tibet Plateau
1
2020
... 然而,目前诸多研究仅集中于比较高原多年冻土区不同采样点之间土壤碳氮的含量大小及剖面分布特征.李新星等[14 ] 对马衔山的研究表明,表层土壤有机碳(Soil Organic Carbon, SOC)和全氮(Total Nitrogen, TN)含量高于下层;Zhao等[15 ] 发现土壤碳氮储量随土壤深度增加呈现降低趋势;马素辉等[16 ] 以黑河上游俄博岭各草地类型为研究对象,比较发现高海拔的高寒沼泽草甸SOC密度(Soil Organic Carbon Density, SOCD)最高,而高寒草甸海拔低且SOCD显著降低;Wang等[17 ] 模拟得出,青藏高原东部SOCD空间上由东向西递减;Xie等[18 ] 发现TN含量季节上基本保持稳定,但高寒草甸显著高于其他植被类型.尽管各研究关于多年冻土区土壤碳氮分布具有较高的一致性,但其对于了解高寒区土壤碳氮分布甚至于评估碳氮库储量存在明显的局限性.首先,多年冻土区由于年内温度变化剧烈,浅层土壤存在明显的季节性冻融现象[19 ] ,表现为春秋季冻融频繁,夏冬季则分别完全融化和冻结[20 ] .已有研究表明,季节性冻融会对土壤碳氮矿化速率[21 ] 及土壤水溶性有机碳[22 ] 和微生物碳氮[23 ] 的含量产生显著影响;其次,大量的基于单一时间节点的采样调查忽视了季节变化对土壤碳氮的影响,导致相关研究对土壤碳氮库储量的估计仍存在明显的不确定性.寇丹[24 ] 、Yuan等[25 ] 均是以一年中单采样节点(生长季)对青藏高原、黑河上游多年冻土区土壤碳氮储量进行了评估.然而,非生长季对土壤碳、氮循环影响在年际尺度上是不可忽视的[12 ] .因此,在多年冻土区开展以空间分布和季节变化相结合的生态调查对于准确把握土壤碳氮含量分布及评估碳氮储量具有重要意义. ...
Storage,patterns,and control of soil organic carbon and nitrogen in the northeastern margin of the Qinghai-Tibetan Plateau
1
2012
... 位于青藏高原东北缘祁连山西段的疏勒河源区多年冻土广泛发育,其多年冻土面积约占源区面积的73%[9 ] ,源区内优势植被类型为高寒草甸,覆盖面积约为42%[22 ] .近20年观测资料显示,源区冻土退化明显且土壤季节性冻融活动显著[9 , 22 ] .目前该区域的最近土壤碳氮调查资料仅为2012年Liu等[26 ] 对生长季高寒草甸内单一样点的剖面分布研究,缺乏年内季节变化研究.因此,本研究拟开展源区浅层SOC、TN含量在季节尺度上的变化研究,揭示其垂直分布规律及主要的影响因素,并估算疏勒河源多年冻土区高寒草甸土壤碳氮储量,以期为全球气候变化背景下青藏高原多年冻土区高寒草甸土壤碳、氮研究提供数据支撑和科学指导. ...
疏勒河上游地区植被物种多样性和生物量及其与环境因子的关系
4
2011
... 疏勒河源区位于青藏高原东北缘祁连山西段,面积约为5 123 km2 ,约占祁连山国家公园青海片区的19.68%.气候类型为大陆性干旱荒漠气候,干冷多风[27 ] .该区域主要植被类型为灌丛、草甸、草原、荒漠及高山稀疏植被,其中高寒草甸面积为2 393.57 km2 ,约占源区总面积的46.72%(图1 ).研究样地位于疏勒河源多年冻土区高寒草甸生态系统综合观测场(38°19′58″N、98°13′44″E,海拔4 036 m,图1 ).基于观测场2017年的气象观测数据资料显示,样地内年均气温为-4.06 ℃,年降水量为466.6 mm且集中在6—9月(图2 ),空气相对湿度为55.49%,年均太阳总辐射为194.3 W·m-2 .0—50 cm土壤砂粒、粉粒和黏粒含量分别为53.80%、35.69%和9.97%.多年冻土类型为阿尔金山-祁连山高寒带山地多年冻土[27 ] ,活动层厚度约为1.5 m,土壤类型为冷钙土.植被覆盖度80%以上,优势植物种有藏嵩草(Kobresia tibetica )和高山嵩草(Kobresia pygmaea )[9 ] . ...
... [27 ],活动层厚度约为1.5 m,土壤类型为冷钙土.植被覆盖度80%以上,优势植物种有藏嵩草(Kobresia tibetica )和高山嵩草(Kobresia pygmaea )[9 ] . ...
... 2017年4月29日与5月7日、7月29日、9月29日与10月3日和12月30日分别采样,用以表征春、夏、秋季和冬季样品.在样地内随机选取6个50 cm×50 cm样方,样方内的植物用剪刀齐地收集并装入信封(鲜草和凋落物),用于测定地上生物量.由于地下生物量集中在0— 30 cm层[27 ] ,用土钻(内半径2.75 cm)自上而下对0— 50 cm土壤分5层(0— 10、10— 20、20— 30、30— 40 cm和40— 50 cm)取样.每层样品取3次混合后,分为3份:1份待自然风干后用于测定SOC、TN含量;1份放入4 ℃便携式保温箱带回实验室,用于测定pH值和氧化还原电位;剩余1份自然风干后清洗,用于测定地下生物量.环刀(100 g·cm-3 )采集土壤容重并用于计算SOCD和TN密度(Total Nitrogen Density, TND). ...
... 此外,SOC和TN的垂直分布还因季节不同产生差异,即随土壤深度增加,秋季0—50 cm内呈递减趋势,春、夏季和冬季仅在0—30 cm内递减,30—50 cm含量有所回升,这种递减趋势可能与该区域植被根系的分布在0—30 cm有关[27 ] ,而季节差异则可能是淋溶过程中养分从表层向下层迁移的动态结果[35 ] .高寒草甸下发育着的土壤,质地为沙壤,养分易淋溶[36 ] ,表层的土壤碳氮会不同比例地向下层流失[37 ] .但是,这种养分向深层的运移通道会因季节性冻融过程和融化深度不同而产生差异.冻融过程和融化深度主要通过改变土壤水热动态、孔隙结构及根系与土壤间的缝隙促进或抑制大孔隙流形成[38 ] .具体来看,除秋季外,春夏冬季均在30—50 cm层SOC、TN含量有升高,这是由于30 cm层以下根系分布少且土壤紧实[39 ] ,一定程度上抑制了养分向下迁移的速率,并且冻土活动层在秋季达到最大融化深度[40 ] ,且中层土壤是高能水区[22 ] ,可能促进了表层SOC、TN持续向下层(30—50 cm)淋溶,因而秋季表现出空间的连续性降低. ...
祁连山疏勒河上游多年冻土区高寒草甸土壤CO2 通量特征
2
2014
... 研究样地内布设有全自动监测系统.其中,气温由HMP155A型温湿度传感器(Vaisala,芬兰)测定,降水数据采用人工雨量筒进行测定.0—10、10—20、20—30、30—40、40—50 cm各层土壤温度和含水量均利用Hydra-Probe II型土壤传感器(Stevens公司,美国)测定,上述传感器连接至CR1000型数据采集器(Campbell公司,美国),每10 min记录1次数据[28 ] . ...
... 本研究中,季节变化对C/N有显著的影响,且地下生物量是主要影响因素,可能还与土壤含氧量有关.夏季C/N最高说明夏季微生物分解过程受氮的限制[51 ] ,而碳源充足,更多的SOC转化为土壤微生物碳[9 ] ,较少的TN转化为土壤微生物氮,刘放等[23 ] 的研究也证实,夏季疏勒河源高寒草甸微生物生物量碳最多,而微生物生物量氮含量最少;另一方面,有机质矿化过程中部分碳在土壤呼吸的作用下以CO2 的形式排放至大气,夏季土壤完全解冻,土壤中含氧量充足成为有机质的矿化能力升高的必要条件,有研究表明,疏勒河土壤CO2 通量夏季最高[28 ] .秋季C/N最低,这说明该季节TN含量是相对充足的,有多余的有机氮转化为无机氮储存在土壤中,但土壤微生物同化1份氮的同时需要5份碳构建自身细胞,20份碳作为生命活动的能源,此时可利用的土壤碳却是较少的(图3 ).此外,地下生物量中大多为死根,能提供碳源的根系减少,土壤碳供应不足且土壤呼吸较弱,抑制了有机质矿化. ...
Variation and control of soil organic carbon and other nutrients in permafrost regions on central Qinghai-Tibetan Plateau
6
2014
... SOCD 用下列公式计算[29 ] : ...
... 式中:SOCD 代表土壤有机碳密度(kg·m-2 );hi 代表土壤厚度(cm);BDi 代表土壤容重(g·cm-3 );SOCi 代表土壤有机碳含量(g·kg-1 );Ci 代表>2 mm土壤石砾百分含量(%);i 为参与计算的土壤层次;n 为参与计算的土壤层总数.TND 的计算公式同SOCD [29 ] ,土壤碳、氮储量由研究区高寒草甸面积分别与其年均密度的乘积计算所得,0—50 cm土层有机碳和全氮含量是0—10、10—20、20—30、30—40、40—50 cm各层次土壤有机碳和全氮含量的加权平均值[30 ] ,年均SOCD 、TND 由四季求平均值所得. ...
... SOC含量取决于进入土壤的植物残体量及其土壤微生物分解作用下损失量的平衡,TN含量主要来源于植物残体归还和生物固氮[31 ] .本研究中,SOC、TN剖面分布呈一定规律,0—10 cm层高于其他层,这与以往的研究结果一致[14 ] .首先,地下生物量(根系及其分泌物、细根周转产生的碎屑等)是SOC和TN主要来源[29 ] .研究表明,高海拔植物会将更多的生物量分配在地下部器官[32 ] ,且植被的凋落物在地表形成凋落物层[22 ] ,这决定了SOC、TN剖面分布特征;其次,土壤含水量是影响根系分布和土壤微生物分解的重要环境因子[29 ] .春夏季土壤表层含水量高于下层[22 ] ,促进了植被生产力,进而输入土壤的碳氮增加,且高的水分可能会造成厌氧微生物群落丰度的增加,而厌氧生物对SOC的分解效率低于好氧生物,从而导致SOC的积累[33 ] ,秋冬季节下层含水量是高于表层的[22 ] ,表层土壤为物质交换提供畅通环境,为土壤碳氮的积累、分解和转化创造了良好的基础[34 ] .此外,SOC、TN还与地上生物量显著正相关,与pH显著负相关,这说明地表枯落物对土壤碳、氮的输入是极其重要的,而pH通过影响植被的生长和土壤微生物的活性、数量来间接影响碳、氮的累积[9 ] . ...
... [29 ].春夏季土壤表层含水量高于下层[22 ] ,促进了植被生产力,进而输入土壤的碳氮增加,且高的水分可能会造成厌氧微生物群落丰度的增加,而厌氧生物对SOC的分解效率低于好氧生物,从而导致SOC的积累[33 ] ,秋冬季节下层含水量是高于表层的[22 ] ,表层土壤为物质交换提供畅通环境,为土壤碳氮的积累、分解和转化创造了良好的基础[34 ] .此外,SOC、TN还与地上生物量显著正相关,与pH显著负相关,这说明地表枯落物对土壤碳、氮的输入是极其重要的,而pH通过影响植被的生长和土壤微生物的活性、数量来间接影响碳、氮的累积[9 ] . ...
... 土壤TN的季节特征表现为夏季最少,春秋和冬季基本一致.这是由于各季节植被对氮素的消耗不同所致,夏季具有良好的代谢环境(碳源充足、温度适宜),微生物和根系活性高,更多的有机态氮转化为无机态氮[48 ] ,植物生长消耗大量氮素,再加上冻土融化土壤水分增加抑制了氧气进入,加速厌氧环境形成,反硝化过程增强,释放N2 O气体[12 ] ,使得夏季含量低于其他季节,而秋冬季消耗氮速率下降,氮矿化能力也随之降低,土壤中氮能够被累积下来.Liu等[29 ] 的研究表明,北麓河土壤平均TND为0.47±0.07 kg·m-2 ,本研究对应值为0.16±0.02 kg·m-2 ,海拔差异可能是造成两区域差异较大的主要原因.北麓河海拔相比疏勒河更高,据报道,海拔与TN之间具有极显著正相关性[14 ] ,海拔增加、温度下降,土壤中微生物和酶活性下降,分解速率减慢利于TN的积累[13 ] .此外,本研究中SOC、TN含量的季节变化差异不显著,但存在季节变化趋势,这可能与观测时间仅为一年且植被类型单一有关,因而多年冻土区长期的连续观测及不同植被类型下各季节间的差异比较,将是进一步需要探究的问题. ...
... C/N是反映土壤有机质分解速率及氮素矿化能力的标志[49 ] ,其比值低说明微生物分解有机质迅速,且氮矿化作用强.反之,有机质分解过程受氮素限制,微生物与植物存在对无机态氮的竞争,不利于植物的生长.本研究得出,剖面变化对C/N影响不显著,这与以往的研究结果一致[49 ] .本研究中,0—50 cm年平均C/N为10.35—10.80,这与青藏高原北麓河的研究结果接近[29 ] ,略低于祁连山东段[50 ] ,表明多年冻土区土壤微生物对有机质的分解过程随地理位置的变化较小,可能与多年冻土区整体温度较低有关. ...
祁连山青海云杉(Picea crassifolia )林浅层土壤碳、氮含量特征及其相互关系
1
2014
... 式中:SOCD 代表土壤有机碳密度(kg·m-2 );hi 代表土壤厚度(cm);BDi 代表土壤容重(g·cm-3 );SOCi 代表土壤有机碳含量(g·kg-1 );Ci 代表>2 mm土壤石砾百分含量(%);i 为参与计算的土壤层次;n 为参与计算的土壤层总数.TND 的计算公式同SOCD [29 ] ,土壤碳、氮储量由研究区高寒草甸面积分别与其年均密度的乘积计算所得,0—50 cm土层有机碳和全氮含量是0—10、10—20、20—30、30—40、40—50 cm各层次土壤有机碳和全氮含量的加权平均值[30 ] ,年均SOCD 、TND 由四季求平均值所得. ...
中国北方典型风沙区土壤碳氮磷化学计量特征
1
2018
... SOC含量取决于进入土壤的植物残体量及其土壤微生物分解作用下损失量的平衡,TN含量主要来源于植物残体归还和生物固氮[31 ] .本研究中,SOC、TN剖面分布呈一定规律,0—10 cm层高于其他层,这与以往的研究结果一致[14 ] .首先,地下生物量(根系及其分泌物、细根周转产生的碎屑等)是SOC和TN主要来源[29 ] .研究表明,高海拔植物会将更多的生物量分配在地下部器官[32 ] ,且植被的凋落物在地表形成凋落物层[22 ] ,这决定了SOC、TN剖面分布特征;其次,土壤含水量是影响根系分布和土壤微生物分解的重要环境因子[29 ] .春夏季土壤表层含水量高于下层[22 ] ,促进了植被生产力,进而输入土壤的碳氮增加,且高的水分可能会造成厌氧微生物群落丰度的增加,而厌氧生物对SOC的分解效率低于好氧生物,从而导致SOC的积累[33 ] ,秋冬季节下层含水量是高于表层的[22 ] ,表层土壤为物质交换提供畅通环境,为土壤碳氮的积累、分解和转化创造了良好的基础[34 ] .此外,SOC、TN还与地上生物量显著正相关,与pH显著负相关,这说明地表枯落物对土壤碳、氮的输入是极其重要的,而pH通过影响植被的生长和土壤微生物的活性、数量来间接影响碳、氮的累积[9 ] . ...
青藏高原高寒草甸植株性状和生物量分配的海拔梯度变异
1
2010
... SOC含量取决于进入土壤的植物残体量及其土壤微生物分解作用下损失量的平衡,TN含量主要来源于植物残体归还和生物固氮[31 ] .本研究中,SOC、TN剖面分布呈一定规律,0—10 cm层高于其他层,这与以往的研究结果一致[14 ] .首先,地下生物量(根系及其分泌物、细根周转产生的碎屑等)是SOC和TN主要来源[29 ] .研究表明,高海拔植物会将更多的生物量分配在地下部器官[32 ] ,且植被的凋落物在地表形成凋落物层[22 ] ,这决定了SOC、TN剖面分布特征;其次,土壤含水量是影响根系分布和土壤微生物分解的重要环境因子[29 ] .春夏季土壤表层含水量高于下层[22 ] ,促进了植被生产力,进而输入土壤的碳氮增加,且高的水分可能会造成厌氧微生物群落丰度的增加,而厌氧生物对SOC的分解效率低于好氧生物,从而导致SOC的积累[33 ] ,秋冬季节下层含水量是高于表层的[22 ] ,表层土壤为物质交换提供畅通环境,为土壤碳氮的积累、分解和转化创造了良好的基础[34 ] .此外,SOC、TN还与地上生物量显著正相关,与pH显著负相关,这说明地表枯落物对土壤碳、氮的输入是极其重要的,而pH通过影响植被的生长和土壤微生物的活性、数量来间接影响碳、氮的累积[9 ] . ...
藏东南色季拉山西坡不同植被土壤有机碳垂直分布特征及其影响因素
1
2020
... SOC含量取决于进入土壤的植物残体量及其土壤微生物分解作用下损失量的平衡,TN含量主要来源于植物残体归还和生物固氮[31 ] .本研究中,SOC、TN剖面分布呈一定规律,0—10 cm层高于其他层,这与以往的研究结果一致[14 ] .首先,地下生物量(根系及其分泌物、细根周转产生的碎屑等)是SOC和TN主要来源[29 ] .研究表明,高海拔植物会将更多的生物量分配在地下部器官[32 ] ,且植被的凋落物在地表形成凋落物层[22 ] ,这决定了SOC、TN剖面分布特征;其次,土壤含水量是影响根系分布和土壤微生物分解的重要环境因子[29 ] .春夏季土壤表层含水量高于下层[22 ] ,促进了植被生产力,进而输入土壤的碳氮增加,且高的水分可能会造成厌氧微生物群落丰度的增加,而厌氧生物对SOC的分解效率低于好氧生物,从而导致SOC的积累[33 ] ,秋冬季节下层含水量是高于表层的[22 ] ,表层土壤为物质交换提供畅通环境,为土壤碳氮的积累、分解和转化创造了良好的基础[34 ] .此外,SOC、TN还与地上生物量显著正相关,与pH显著负相关,这说明地表枯落物对土壤碳、氮的输入是极其重要的,而pH通过影响植被的生长和土壤微生物的活性、数量来间接影响碳、氮的累积[9 ] . ...
赤峰市敖汉旗土壤有机碳含量的垂直分布及其影响因素
1
2019
... SOC含量取决于进入土壤的植物残体量及其土壤微生物分解作用下损失量的平衡,TN含量主要来源于植物残体归还和生物固氮[31 ] .本研究中,SOC、TN剖面分布呈一定规律,0—10 cm层高于其他层,这与以往的研究结果一致[14 ] .首先,地下生物量(根系及其分泌物、细根周转产生的碎屑等)是SOC和TN主要来源[29 ] .研究表明,高海拔植物会将更多的生物量分配在地下部器官[32 ] ,且植被的凋落物在地表形成凋落物层[22 ] ,这决定了SOC、TN剖面分布特征;其次,土壤含水量是影响根系分布和土壤微生物分解的重要环境因子[29 ] .春夏季土壤表层含水量高于下层[22 ] ,促进了植被生产力,进而输入土壤的碳氮增加,且高的水分可能会造成厌氧微生物群落丰度的增加,而厌氧生物对SOC的分解效率低于好氧生物,从而导致SOC的积累[33 ] ,秋冬季节下层含水量是高于表层的[22 ] ,表层土壤为物质交换提供畅通环境,为土壤碳氮的积累、分解和转化创造了良好的基础[34 ] .此外,SOC、TN还与地上生物量显著正相关,与pH显著负相关,这说明地表枯落物对土壤碳、氮的输入是极其重要的,而pH通过影响植被的生长和土壤微生物的活性、数量来间接影响碳、氮的累积[9 ] . ...
不同气候类型下四川草地土壤有机碳空间分布及影响因素
1
2018
... 此外,SOC和TN的垂直分布还因季节不同产生差异,即随土壤深度增加,秋季0—50 cm内呈递减趋势,春、夏季和冬季仅在0—30 cm内递减,30—50 cm含量有所回升,这种递减趋势可能与该区域植被根系的分布在0—30 cm有关[27 ] ,而季节差异则可能是淋溶过程中养分从表层向下层迁移的动态结果[35 ] .高寒草甸下发育着的土壤,质地为沙壤,养分易淋溶[36 ] ,表层的土壤碳氮会不同比例地向下层流失[37 ] .但是,这种养分向深层的运移通道会因季节性冻融过程和融化深度不同而产生差异.冻融过程和融化深度主要通过改变土壤水热动态、孔隙结构及根系与土壤间的缝隙促进或抑制大孔隙流形成[38 ] .具体来看,除秋季外,春夏冬季均在30—50 cm层SOC、TN含量有升高,这是由于30 cm层以下根系分布少且土壤紧实[39 ] ,一定程度上抑制了养分向下迁移的速率,并且冻土活动层在秋季达到最大融化深度[40 ] ,且中层土壤是高能水区[22 ] ,可能促进了表层SOC、TN持续向下层(30—50 cm)淋溶,因而秋季表现出空间的连续性降低. ...
青藏高原典型高寒草甸区土壤有机碳氮的变异特性
1
2009
... 此外,SOC和TN的垂直分布还因季节不同产生差异,即随土壤深度增加,秋季0—50 cm内呈递减趋势,春、夏季和冬季仅在0—30 cm内递减,30—50 cm含量有所回升,这种递减趋势可能与该区域植被根系的分布在0—30 cm有关[27 ] ,而季节差异则可能是淋溶过程中养分从表层向下层迁移的动态结果[35 ] .高寒草甸下发育着的土壤,质地为沙壤,养分易淋溶[36 ] ,表层的土壤碳氮会不同比例地向下层流失[37 ] .但是,这种养分向深层的运移通道会因季节性冻融过程和融化深度不同而产生差异.冻融过程和融化深度主要通过改变土壤水热动态、孔隙结构及根系与土壤间的缝隙促进或抑制大孔隙流形成[38 ] .具体来看,除秋季外,春夏冬季均在30—50 cm层SOC、TN含量有升高,这是由于30 cm层以下根系分布少且土壤紧实[39 ] ,一定程度上抑制了养分向下迁移的速率,并且冻土活动层在秋季达到最大融化深度[40 ] ,且中层土壤是高能水区[22 ] ,可能促进了表层SOC、TN持续向下层(30—50 cm)淋溶,因而秋季表现出空间的连续性降低. ...
岩溶流域不同土壤剖面溶解性碳氮分布和淋失特征
1
2010
... 此外,SOC和TN的垂直分布还因季节不同产生差异,即随土壤深度增加,秋季0—50 cm内呈递减趋势,春、夏季和冬季仅在0—30 cm内递减,30—50 cm含量有所回升,这种递减趋势可能与该区域植被根系的分布在0—30 cm有关[27 ] ,而季节差异则可能是淋溶过程中养分从表层向下层迁移的动态结果[35 ] .高寒草甸下发育着的土壤,质地为沙壤,养分易淋溶[36 ] ,表层的土壤碳氮会不同比例地向下层流失[37 ] .但是,这种养分向深层的运移通道会因季节性冻融过程和融化深度不同而产生差异.冻融过程和融化深度主要通过改变土壤水热动态、孔隙结构及根系与土壤间的缝隙促进或抑制大孔隙流形成[38 ] .具体来看,除秋季外,春夏冬季均在30—50 cm层SOC、TN含量有升高,这是由于30 cm层以下根系分布少且土壤紧实[39 ] ,一定程度上抑制了养分向下迁移的速率,并且冻土活动层在秋季达到最大融化深度[40 ] ,且中层土壤是高能水区[22 ] ,可能促进了表层SOC、TN持续向下层(30—50 cm)淋溶,因而秋季表现出空间的连续性降低. ...
冻融交替对农田氮磷淋溶影响的研究进展
1
2021
... 此外,SOC和TN的垂直分布还因季节不同产生差异,即随土壤深度增加,秋季0—50 cm内呈递减趋势,春、夏季和冬季仅在0—30 cm内递减,30—50 cm含量有所回升,这种递减趋势可能与该区域植被根系的分布在0—30 cm有关[27 ] ,而季节差异则可能是淋溶过程中养分从表层向下层迁移的动态结果[35 ] .高寒草甸下发育着的土壤,质地为沙壤,养分易淋溶[36 ] ,表层的土壤碳氮会不同比例地向下层流失[37 ] .但是,这种养分向深层的运移通道会因季节性冻融过程和融化深度不同而产生差异.冻融过程和融化深度主要通过改变土壤水热动态、孔隙结构及根系与土壤间的缝隙促进或抑制大孔隙流形成[38 ] .具体来看,除秋季外,春夏冬季均在30—50 cm层SOC、TN含量有升高,这是由于30 cm层以下根系分布少且土壤紧实[39 ] ,一定程度上抑制了养分向下迁移的速率,并且冻土活动层在秋季达到最大融化深度[40 ] ,且中层土壤是高能水区[22 ] ,可能促进了表层SOC、TN持续向下层(30—50 cm)淋溶,因而秋季表现出空间的连续性降低. ...
祁连山东段高寒地区土地利用方式对土壤性状的影响
1
2012
... 此外,SOC和TN的垂直分布还因季节不同产生差异,即随土壤深度增加,秋季0—50 cm内呈递减趋势,春、夏季和冬季仅在0—30 cm内递减,30—50 cm含量有所回升,这种递减趋势可能与该区域植被根系的分布在0—30 cm有关[27 ] ,而季节差异则可能是淋溶过程中养分从表层向下层迁移的动态结果[35 ] .高寒草甸下发育着的土壤,质地为沙壤,养分易淋溶[36 ] ,表层的土壤碳氮会不同比例地向下层流失[37 ] .但是,这种养分向深层的运移通道会因季节性冻融过程和融化深度不同而产生差异.冻融过程和融化深度主要通过改变土壤水热动态、孔隙结构及根系与土壤间的缝隙促进或抑制大孔隙流形成[38 ] .具体来看,除秋季外,春夏冬季均在30—50 cm层SOC、TN含量有升高,这是由于30 cm层以下根系分布少且土壤紧实[39 ] ,一定程度上抑制了养分向下迁移的速率,并且冻土活动层在秋季达到最大融化深度[40 ] ,且中层土壤是高能水区[22 ] ,可能促进了表层SOC、TN持续向下层(30—50 cm)淋溶,因而秋季表现出空间的连续性降低. ...
青藏高原五道梁附近多年冻土活动层冻结和融化过程
1
2000
... 此外,SOC和TN的垂直分布还因季节不同产生差异,即随土壤深度增加,秋季0—50 cm内呈递减趋势,春、夏季和冬季仅在0—30 cm内递减,30—50 cm含量有所回升,这种递减趋势可能与该区域植被根系的分布在0—30 cm有关[27 ] ,而季节差异则可能是淋溶过程中养分从表层向下层迁移的动态结果[35 ] .高寒草甸下发育着的土壤,质地为沙壤,养分易淋溶[36 ] ,表层的土壤碳氮会不同比例地向下层流失[37 ] .但是,这种养分向深层的运移通道会因季节性冻融过程和融化深度不同而产生差异.冻融过程和融化深度主要通过改变土壤水热动态、孔隙结构及根系与土壤间的缝隙促进或抑制大孔隙流形成[38 ] .具体来看,除秋季外,春夏冬季均在30—50 cm层SOC、TN含量有升高,这是由于30 cm层以下根系分布少且土壤紧实[39 ] ,一定程度上抑制了养分向下迁移的速率,并且冻土活动层在秋季达到最大融化深度[40 ] ,且中层土壤是高能水区[22 ] ,可能促进了表层SOC、TN持续向下层(30—50 cm)淋溶,因而秋季表现出空间的连续性降低. ...
名山县不同土壤类型下有机碳组成和季节性变化研究
1
2014
... 生物量及环境因素的季节变化使SOC含量具有季节差异.陈梦棋等[41 ] 对比名山县生长季各植被类型SOC发现,其存在差异但不显著,这与本研究的结果相符.本研究表明,影响SOC季节差异的主控因子是地下生物量,这表明碳输入量的不同会影响SOC含量大小,但变幅相对较小.春季植被返青期主要消耗上一年份累积的土壤养分来度过[42 ] ,加之土壤融化水分和温度增加促进微生物活性并消耗碳源,因而春季含量低于冬季;夏季优越的水热条件植被生长旺盛,强烈的光合作用使根系和根茎固定了更多的碳,大量的有机物质(凋落物、根系分泌物及其脱落物等)进入土壤并转化为SOC,与此同时土壤微生物的新陈代谢虽会消耗一部分SOC,但SOC积累量远大于消耗量,表现为夏季最高;秋季地表枯黄产生凋落物、根系衰减及动物残体(气温骤降大量细菌因低温环境休眠或死亡)等还未来得及被分解而以腐殖质的形式储存在土体中,致秋季最低;冬季植被凋亡,地下生物量急剧降低,但来源于秋季的大量腐殖质为适应低温环境而幸存下来的真菌(微生物群体中约87%的细菌休眠或死亡)微生物提供了较为丰富的代谢底物[43 ] ,再加上积雪覆盖在土壤表面起隔热作用[23 ] ,以及低气温抑制土壤呼吸,减缓有机质分解速率[44 ] ,SOC积累胜过消耗,冬季高于秋季.此外,生长季温度高,土壤酶催化过程以及微生物代谢过程均加速[45 ] ,从而加快SOC矿化过程,这可能是造成各季节间SOC含量差异不显著的原因之一.本研究中,年均SOCD为8.49±0.25 kg·m-2 ,低于黑河上游多年冻土区高寒草甸以生长季采样所得结果(11.22 kg·m-2 )[16 ] ,可能原因是影响其变化的因素如植被盖度、凋落物、土壤呼吸速率因所处环境的不同均存在差异[46 -47 ] .本研究通过全年采样得出,夏季SOCD比年平均密度约高5.77%,可见,利用生长盛季估算全年SOCD及储量会存在高估的趋势. ...
Effects of mild winter freezing on soil nitrogen and carbon dynamics in a northern hardwood forest
1
2001
... 生物量及环境因素的季节变化使SOC含量具有季节差异.陈梦棋等[41 ] 对比名山县生长季各植被类型SOC发现,其存在差异但不显著,这与本研究的结果相符.本研究表明,影响SOC季节差异的主控因子是地下生物量,这表明碳输入量的不同会影响SOC含量大小,但变幅相对较小.春季植被返青期主要消耗上一年份累积的土壤养分来度过[42 ] ,加之土壤融化水分和温度增加促进微生物活性并消耗碳源,因而春季含量低于冬季;夏季优越的水热条件植被生长旺盛,强烈的光合作用使根系和根茎固定了更多的碳,大量的有机物质(凋落物、根系分泌物及其脱落物等)进入土壤并转化为SOC,与此同时土壤微生物的新陈代谢虽会消耗一部分SOC,但SOC积累量远大于消耗量,表现为夏季最高;秋季地表枯黄产生凋落物、根系衰减及动物残体(气温骤降大量细菌因低温环境休眠或死亡)等还未来得及被分解而以腐殖质的形式储存在土体中,致秋季最低;冬季植被凋亡,地下生物量急剧降低,但来源于秋季的大量腐殖质为适应低温环境而幸存下来的真菌(微生物群体中约87%的细菌休眠或死亡)微生物提供了较为丰富的代谢底物[43 ] ,再加上积雪覆盖在土壤表面起隔热作用[23 ] ,以及低气温抑制土壤呼吸,减缓有机质分解速率[44 ] ,SOC积累胜过消耗,冬季高于秋季.此外,生长季温度高,土壤酶催化过程以及微生物代谢过程均加速[45 ] ,从而加快SOC矿化过程,这可能是造成各季节间SOC含量差异不显著的原因之一.本研究中,年均SOCD为8.49±0.25 kg·m-2 ,低于黑河上游多年冻土区高寒草甸以生长季采样所得结果(11.22 kg·m-2 )[16 ] ,可能原因是影响其变化的因素如植被盖度、凋落物、土壤呼吸速率因所处环境的不同均存在差异[46 -47 ] .本研究通过全年采样得出,夏季SOCD比年平均密度约高5.77%,可见,利用生长盛季估算全年SOCD及储量会存在高估的趋势. ...
Cold adaptation in Arctic and Antarctic fungi
1
2001
... 生物量及环境因素的季节变化使SOC含量具有季节差异.陈梦棋等[41 ] 对比名山县生长季各植被类型SOC发现,其存在差异但不显著,这与本研究的结果相符.本研究表明,影响SOC季节差异的主控因子是地下生物量,这表明碳输入量的不同会影响SOC含量大小,但变幅相对较小.春季植被返青期主要消耗上一年份累积的土壤养分来度过[42 ] ,加之土壤融化水分和温度增加促进微生物活性并消耗碳源,因而春季含量低于冬季;夏季优越的水热条件植被生长旺盛,强烈的光合作用使根系和根茎固定了更多的碳,大量的有机物质(凋落物、根系分泌物及其脱落物等)进入土壤并转化为SOC,与此同时土壤微生物的新陈代谢虽会消耗一部分SOC,但SOC积累量远大于消耗量,表现为夏季最高;秋季地表枯黄产生凋落物、根系衰减及动物残体(气温骤降大量细菌因低温环境休眠或死亡)等还未来得及被分解而以腐殖质的形式储存在土体中,致秋季最低;冬季植被凋亡,地下生物量急剧降低,但来源于秋季的大量腐殖质为适应低温环境而幸存下来的真菌(微生物群体中约87%的细菌休眠或死亡)微生物提供了较为丰富的代谢底物[43 ] ,再加上积雪覆盖在土壤表面起隔热作用[23 ] ,以及低气温抑制土壤呼吸,减缓有机质分解速率[44 ] ,SOC积累胜过消耗,冬季高于秋季.此外,生长季温度高,土壤酶催化过程以及微生物代谢过程均加速[45 ] ,从而加快SOC矿化过程,这可能是造成各季节间SOC含量差异不显著的原因之一.本研究中,年均SOCD为8.49±0.25 kg·m-2 ,低于黑河上游多年冻土区高寒草甸以生长季采样所得结果(11.22 kg·m-2 )[16 ] ,可能原因是影响其变化的因素如植被盖度、凋落物、土壤呼吸速率因所处环境的不同均存在差异[46 -47 ] .本研究通过全年采样得出,夏季SOCD比年平均密度约高5.77%,可见,利用生长盛季估算全年SOCD及储量会存在高估的趋势. ...
增温对土壤有机碳矿化的影响研究综述
1
2017
... 生物量及环境因素的季节变化使SOC含量具有季节差异.陈梦棋等[41 ] 对比名山县生长季各植被类型SOC发现,其存在差异但不显著,这与本研究的结果相符.本研究表明,影响SOC季节差异的主控因子是地下生物量,这表明碳输入量的不同会影响SOC含量大小,但变幅相对较小.春季植被返青期主要消耗上一年份累积的土壤养分来度过[42 ] ,加之土壤融化水分和温度增加促进微生物活性并消耗碳源,因而春季含量低于冬季;夏季优越的水热条件植被生长旺盛,强烈的光合作用使根系和根茎固定了更多的碳,大量的有机物质(凋落物、根系分泌物及其脱落物等)进入土壤并转化为SOC,与此同时土壤微生物的新陈代谢虽会消耗一部分SOC,但SOC积累量远大于消耗量,表现为夏季最高;秋季地表枯黄产生凋落物、根系衰减及动物残体(气温骤降大量细菌因低温环境休眠或死亡)等还未来得及被分解而以腐殖质的形式储存在土体中,致秋季最低;冬季植被凋亡,地下生物量急剧降低,但来源于秋季的大量腐殖质为适应低温环境而幸存下来的真菌(微生物群体中约87%的细菌休眠或死亡)微生物提供了较为丰富的代谢底物[43 ] ,再加上积雪覆盖在土壤表面起隔热作用[23 ] ,以及低气温抑制土壤呼吸,减缓有机质分解速率[44 ] ,SOC积累胜过消耗,冬季高于秋季.此外,生长季温度高,土壤酶催化过程以及微生物代谢过程均加速[45 ] ,从而加快SOC矿化过程,这可能是造成各季节间SOC含量差异不显著的原因之一.本研究中,年均SOCD为8.49±0.25 kg·m-2 ,低于黑河上游多年冻土区高寒草甸以生长季采样所得结果(11.22 kg·m-2 )[16 ] ,可能原因是影响其变化的因素如植被盖度、凋落物、土壤呼吸速率因所处环境的不同均存在差异[46 -47 ] .本研究通过全年采样得出,夏季SOCD比年平均密度约高5.77%,可见,利用生长盛季估算全年SOCD及储量会存在高估的趋势. ...
腾格里沙漠东南缘人工固沙植被区表层土壤有机碳矿化对凋落物添加的响应
1
2019
... 生物量及环境因素的季节变化使SOC含量具有季节差异.陈梦棋等[41 ] 对比名山县生长季各植被类型SOC发现,其存在差异但不显著,这与本研究的结果相符.本研究表明,影响SOC季节差异的主控因子是地下生物量,这表明碳输入量的不同会影响SOC含量大小,但变幅相对较小.春季植被返青期主要消耗上一年份累积的土壤养分来度过[42 ] ,加之土壤融化水分和温度增加促进微生物活性并消耗碳源,因而春季含量低于冬季;夏季优越的水热条件植被生长旺盛,强烈的光合作用使根系和根茎固定了更多的碳,大量的有机物质(凋落物、根系分泌物及其脱落物等)进入土壤并转化为SOC,与此同时土壤微生物的新陈代谢虽会消耗一部分SOC,但SOC积累量远大于消耗量,表现为夏季最高;秋季地表枯黄产生凋落物、根系衰减及动物残体(气温骤降大量细菌因低温环境休眠或死亡)等还未来得及被分解而以腐殖质的形式储存在土体中,致秋季最低;冬季植被凋亡,地下生物量急剧降低,但来源于秋季的大量腐殖质为适应低温环境而幸存下来的真菌(微生物群体中约87%的细菌休眠或死亡)微生物提供了较为丰富的代谢底物[43 ] ,再加上积雪覆盖在土壤表面起隔热作用[23 ] ,以及低气温抑制土壤呼吸,减缓有机质分解速率[44 ] ,SOC积累胜过消耗,冬季高于秋季.此外,生长季温度高,土壤酶催化过程以及微生物代谢过程均加速[45 ] ,从而加快SOC矿化过程,这可能是造成各季节间SOC含量差异不显著的原因之一.本研究中,年均SOCD为8.49±0.25 kg·m-2 ,低于黑河上游多年冻土区高寒草甸以生长季采样所得结果(11.22 kg·m-2 )[16 ] ,可能原因是影响其变化的因素如植被盖度、凋落物、土壤呼吸速率因所处环境的不同均存在差异[46 -47 ] .本研究通过全年采样得出,夏季SOCD比年平均密度约高5.77%,可见,利用生长盛季估算全年SOCD及储量会存在高估的趋势. ...
Storage,patterns and controls of soil organic carbon in the Tibetan grasslands
1
2008
... 生物量及环境因素的季节变化使SOC含量具有季节差异.陈梦棋等[41 ] 对比名山县生长季各植被类型SOC发现,其存在差异但不显著,这与本研究的结果相符.本研究表明,影响SOC季节差异的主控因子是地下生物量,这表明碳输入量的不同会影响SOC含量大小,但变幅相对较小.春季植被返青期主要消耗上一年份累积的土壤养分来度过[42 ] ,加之土壤融化水分和温度增加促进微生物活性并消耗碳源,因而春季含量低于冬季;夏季优越的水热条件植被生长旺盛,强烈的光合作用使根系和根茎固定了更多的碳,大量的有机物质(凋落物、根系分泌物及其脱落物等)进入土壤并转化为SOC,与此同时土壤微生物的新陈代谢虽会消耗一部分SOC,但SOC积累量远大于消耗量,表现为夏季最高;秋季地表枯黄产生凋落物、根系衰减及动物残体(气温骤降大量细菌因低温环境休眠或死亡)等还未来得及被分解而以腐殖质的形式储存在土体中,致秋季最低;冬季植被凋亡,地下生物量急剧降低,但来源于秋季的大量腐殖质为适应低温环境而幸存下来的真菌(微生物群体中约87%的细菌休眠或死亡)微生物提供了较为丰富的代谢底物[43 ] ,再加上积雪覆盖在土壤表面起隔热作用[23 ] ,以及低气温抑制土壤呼吸,减缓有机质分解速率[44 ] ,SOC积累胜过消耗,冬季高于秋季.此外,生长季温度高,土壤酶催化过程以及微生物代谢过程均加速[45 ] ,从而加快SOC矿化过程,这可能是造成各季节间SOC含量差异不显著的原因之一.本研究中,年均SOCD为8.49±0.25 kg·m-2 ,低于黑河上游多年冻土区高寒草甸以生长季采样所得结果(11.22 kg·m-2 )[16 ] ,可能原因是影响其变化的因素如植被盖度、凋落物、土壤呼吸速率因所处环境的不同均存在差异[46 -47 ] .本研究通过全年采样得出,夏季SOCD比年平均密度约高5.77%,可见,利用生长盛季估算全年SOCD及储量会存在高估的趋势. ...
青藏高原土壤有机碳密度垂直分布研究
1
2008
... 生物量及环境因素的季节变化使SOC含量具有季节差异.陈梦棋等[41 ] 对比名山县生长季各植被类型SOC发现,其存在差异但不显著,这与本研究的结果相符.本研究表明,影响SOC季节差异的主控因子是地下生物量,这表明碳输入量的不同会影响SOC含量大小,但变幅相对较小.春季植被返青期主要消耗上一年份累积的土壤养分来度过[42 ] ,加之土壤融化水分和温度增加促进微生物活性并消耗碳源,因而春季含量低于冬季;夏季优越的水热条件植被生长旺盛,强烈的光合作用使根系和根茎固定了更多的碳,大量的有机物质(凋落物、根系分泌物及其脱落物等)进入土壤并转化为SOC,与此同时土壤微生物的新陈代谢虽会消耗一部分SOC,但SOC积累量远大于消耗量,表现为夏季最高;秋季地表枯黄产生凋落物、根系衰减及动物残体(气温骤降大量细菌因低温环境休眠或死亡)等还未来得及被分解而以腐殖质的形式储存在土体中,致秋季最低;冬季植被凋亡,地下生物量急剧降低,但来源于秋季的大量腐殖质为适应低温环境而幸存下来的真菌(微生物群体中约87%的细菌休眠或死亡)微生物提供了较为丰富的代谢底物[43 ] ,再加上积雪覆盖在土壤表面起隔热作用[23 ] ,以及低气温抑制土壤呼吸,减缓有机质分解速率[44 ] ,SOC积累胜过消耗,冬季高于秋季.此外,生长季温度高,土壤酶催化过程以及微生物代谢过程均加速[45 ] ,从而加快SOC矿化过程,这可能是造成各季节间SOC含量差异不显著的原因之一.本研究中,年均SOCD为8.49±0.25 kg·m-2 ,低于黑河上游多年冻土区高寒草甸以生长季采样所得结果(11.22 kg·m-2 )[16 ] ,可能原因是影响其变化的因素如植被盖度、凋落物、土壤呼吸速率因所处环境的不同均存在差异[46 -47 ] .本研究通过全年采样得出,夏季SOCD比年平均密度约高5.77%,可见,利用生长盛季估算全年SOCD及储量会存在高估的趋势. ...
温度和水分对科尔沁沙质草地土壤氮矿化的影响
1
2016
... 土壤TN的季节特征表现为夏季最少,春秋和冬季基本一致.这是由于各季节植被对氮素的消耗不同所致,夏季具有良好的代谢环境(碳源充足、温度适宜),微生物和根系活性高,更多的有机态氮转化为无机态氮[48 ] ,植物生长消耗大量氮素,再加上冻土融化土壤水分增加抑制了氧气进入,加速厌氧环境形成,反硝化过程增强,释放N2 O气体[12 ] ,使得夏季含量低于其他季节,而秋冬季消耗氮速率下降,氮矿化能力也随之降低,土壤中氮能够被累积下来.Liu等[29 ] 的研究表明,北麓河土壤平均TND为0.47±0.07 kg·m-2 ,本研究对应值为0.16±0.02 kg·m-2 ,海拔差异可能是造成两区域差异较大的主要原因.北麓河海拔相比疏勒河更高,据报道,海拔与TN之间具有极显著正相关性[14 ] ,海拔增加、温度下降,土壤中微生物和酶活性下降,分解速率减慢利于TN的积累[13 ] .此外,本研究中SOC、TN含量的季节变化差异不显著,但存在季节变化趋势,这可能与观测时间仅为一年且植被类型单一有关,因而多年冻土区长期的连续观测及不同植被类型下各季节间的差异比较,将是进一步需要探究的问题. ...
Vertical patterns of soil carbon,nitrogen and carbon:nitrogen stoichiometry in Tibetan grasslands
2
2010
... C/N是反映土壤有机质分解速率及氮素矿化能力的标志[49 ] ,其比值低说明微生物分解有机质迅速,且氮矿化作用强.反之,有机质分解过程受氮素限制,微生物与植物存在对无机态氮的竞争,不利于植物的生长.本研究得出,剖面变化对C/N影响不显著,这与以往的研究结果一致[49 ] .本研究中,0—50 cm年平均C/N为10.35—10.80,这与青藏高原北麓河的研究结果接近[29 ] ,略低于祁连山东段[50 ] ,表明多年冻土区土壤微生物对有机质的分解过程随地理位置的变化较小,可能与多年冻土区整体温度较低有关. ...
... [49 ].本研究中,0—50 cm年平均C/N为10.35—10.80,这与青藏高原北麓河的研究结果接近[29 ] ,略低于祁连山东段[50 ] ,表明多年冻土区土壤微生物对有机质的分解过程随地理位置的变化较小,可能与多年冻土区整体温度较低有关. ...
祁连山东段南麓不同海拔土壤有机质及全氮的分布状况
1
2009
... C/N是反映土壤有机质分解速率及氮素矿化能力的标志[49 ] ,其比值低说明微生物分解有机质迅速,且氮矿化作用强.反之,有机质分解过程受氮素限制,微生物与植物存在对无机态氮的竞争,不利于植物的生长.本研究得出,剖面变化对C/N影响不显著,这与以往的研究结果一致[49 ] .本研究中,0—50 cm年平均C/N为10.35—10.80,这与青藏高原北麓河的研究结果接近[29 ] ,略低于祁连山东段[50 ] ,表明多年冻土区土壤微生物对有机质的分解过程随地理位置的变化较小,可能与多年冻土区整体温度较低有关. ...
芦芽山土壤有机碳和全氮沿海拔梯度变化规律
1
2014
... 本研究中,季节变化对C/N有显著的影响,且地下生物量是主要影响因素,可能还与土壤含氧量有关.夏季C/N最高说明夏季微生物分解过程受氮的限制[51 ] ,而碳源充足,更多的SOC转化为土壤微生物碳[9 ] ,较少的TN转化为土壤微生物氮,刘放等[23 ] 的研究也证实,夏季疏勒河源高寒草甸微生物生物量碳最多,而微生物生物量氮含量最少;另一方面,有机质矿化过程中部分碳在土壤呼吸的作用下以CO2 的形式排放至大气,夏季土壤完全解冻,土壤中含氧量充足成为有机质的矿化能力升高的必要条件,有研究表明,疏勒河土壤CO2 通量夏季最高[28 ] .秋季C/N最低,这说明该季节TN含量是相对充足的,有多余的有机氮转化为无机氮储存在土壤中,但土壤微生物同化1份氮的同时需要5份碳构建自身细胞,20份碳作为生命活动的能源,此时可利用的土壤碳却是较少的(图3 ).此外,地下生物量中大多为死根,能提供碳源的根系减少,土壤碳供应不足且土壤呼吸较弱,抑制了有机质矿化. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}