0 引言
种群空间分布格局是植物种群自身生物学特性与环境因素相互作用的结果,一直是生态学研究的热点[1 -4 ] 。点格局方法能够以由小到大的一系列连续尺度分析点状物体平面分布格局[5 -6 ] ,对于揭示植物种群对外部环境的生态学适应机制,展现物种在群落中的生态学地位,指示该物种之间的相互关系有着较大的意义[7 -9 ] 。植物功能性状是植物适应环境的一种表现形式,会随外界环境变化而呈现出相应的形态结构与生理特性,是研究植物响应环境变化的有力工具[10 ] 。植物种群在面对不同风沙条件时,需要不断地调整株高、叶片、种群密度等特征。因此,开展沙鞭地上分株分布格局对不同风沙环境响应的研究能够反映出植物种群的资源投入、选择和适应策略,对于揭示植物种群应对干旱风沙等极端环境的响应机制具有重要意义。
沙鞭(Psammochloa villosa )作为干旱地区流动沙丘先锋物种,对于干旱风沙环境具有极强生态适应能力,在流动沙丘上分布的沙鞭斑块与流沙斑块相互镶嵌,已成为流动沙丘的独特景观[1 ,3 ,5 ,11 ] ,主要分布在内蒙古库布齐沙漠、浑善达克沙地西部、巴丹吉林沙漠和甘肃河西走廊沙地等地区。目前关于沙鞭的研究主要包括有性繁殖[12 -13 ] 与无性繁殖[5 ,7 ,14 ] 、不同居群形态变异[15 ] 、遗传多样性[16 -17 ] 、改造沙化景观[18 ] ,极大地丰富和发展了沙鞭的种群特征研究,但是关于不同沙丘生境下沙鞭种群空间分布与繁殖策略特征的研究尚未涉及。基于此,本研究选择流动、半流动、半固定和固定沙丘,开展沙鞭种群地上分株分布格局研究,旨在研究沙丘流动性对于沙鞭种群空间分布格局的影响,为了解极端干旱地区植物种群对环境胁迫的响应机理提供科学依据。
1 研究方法
1.1 研究区概况
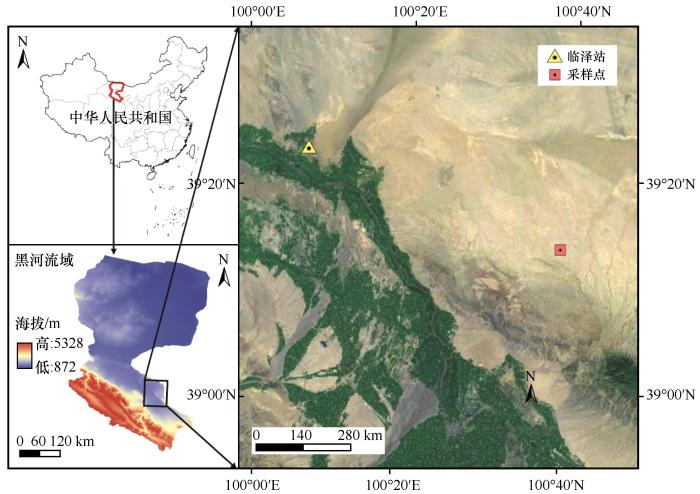
研究区位于甘肃省河西走廊中部临泽县北部(39°13′19.92″N,100°39′00″E),依托中国科学院临泽内陆河流域研究站开展相关研究(图1 )。研究区位于欧亚大陆腹地,远离海洋,属典型大陆性干旱荒漠气候,辐射强,热量丰富,降水少,年均气温7.6 ℃,极端最高气温可达39.1 ℃,极端最低气温为-27.3 ℃,无霜期150—165 d,季节性冻土厚度109—123 cm。多年平均降水量117.4 mm (1965—2018年),年均潜在蒸发量2 390 mm,干旱指数达15.9,年日照时数3 045 h,太阳辐射总量高达6.1×105 J·cm-2 ·a-1 [19 ] 。区域内降水存在明显季节变化,夏季(7—9月)降水占全年降水总量的65%左右,冬季降雪量占全年降水总量的3%;以小降水事件为主,≤5 mm小降水事件占全年总降水事件的56.1%,并占年降水量的46.7%(1967—2017年)。天然植被总体特征组成简单、类型单一、群落不郁闭、分布稀疏。地带性植被为以超旱生半灌木和多年生草本为优势种或建群种形成的沙丘植被类型,旱生和盐生灌木为主要植物群落,梭梭(Haloxylon ammodendron )、沙拐枣(Calligonum mongolicum )、柽柳(Tamarix chinensis )、花棒(Hedysarum scoparium )、柠条(Caragana korshinskii )和泡泡刺(Nitraria sphaerocarpa )、红柳(Tamarix ramosissima )是广泛分布于临泽荒漠绿洲固定、半固定沙丘上的灌木和半灌木。一年生草本植物如虫实(Corispermum candelabrum )、雾冰藜(Bassia dasyphylla )、白茎盐生草(Halogeton arachnoideus )、碱蓬(Suaeda glauca )、沙蓬(Agriophyllum squarrosum )和画眉草(Eragrostis pilosa )等,多年生草本植物如沙鞭、沙蒿(Artemisia desertorum )等[19 ] 。沙鞭为广布于流动沙丘的重要先锋物种,其种群消长对于沙丘生态系统群落结构和功能的稳定性具有不可替代的生态作用。
图1
图1
研究区位置图
Fig.1
The location of the study area
1.2 样地设置与调查分析
根据赵哈林等[20 ] 和左小安等[21 ] 对沙丘类型的划分标准,在地貌相对一致、地形平缓的巴丹吉林沙漠边缘的甘肃省张掖市临泽县平川镇,结合实地踏查和群落学调查,根据沙丘植被盖度、植被和流沙特征、沙丘表面结皮情况,以及群落优势种和伴生种等指标选取了4处围栏沙地作为实验样地(表1 ),分别为流动沙丘(A)、半流动沙丘(B)、半固定沙丘(C)、固定沙丘(D)。
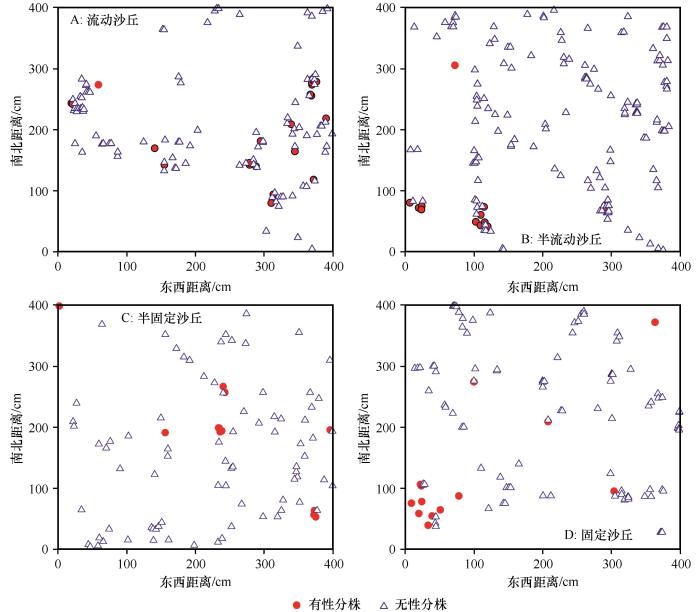
试验在2020年6月植物生长盛期进行,在每个取样点内随机设置3个4 m×4 m的样方,共设置12个样方,用1 m×1 m的样方框(样方框内小格子的规格为10 cm×10 cm)按从左到右、从上到下的顺序依次放置(样方框按正南正北方向放置)16次,进行草地群落调查、沙鞭种群的空间格局调查。实测并记录每个样方(4 m×4 m)内的植物种类、密度、株高和地上生物量;以样方框左下角顶点为基点,记录样方(4 m×4 m)中每个沙鞭地上分株的地面投影和相对位置,用坐标值直接表示距离。在室内对每一个样方(4 m×4 m)按照取样时样方框放置的顺序进行拼接、合并,确定各群落优势种(图2 )。
图2
图2
不同沙丘类型沙鞭(Psammochloa villosa )种群分株分布点位图
Fig.2
Bitmap of Psammochloa villosa population ramet distribution in different dune types
1.3 研究方法
1.3.1 点格局分析
处于研究区域中二维空间的点构成点事件,以k(r) 函数揭示点事件空间布局的依赖程度[22 ] ,定义为:
k ( r ) = λ - 1 E ( # ( r i j ≤ r ) ) (1)
式中:λ 代表着研究区域中每个单位面积点的事件密度;E 代表着在一定的距离尺度中点事件的数量期望;# 代表数量;i 和j 代表研究区域中两个拥有相同特征的点事件;rij 代表某个点与其他点之间的距离;r 为距离尺度。如果假定在一定的距离尺度中,点事件是随机分布的,则有:
E ( # ( r i j ≤ d ) ) = λ π r 2 (2)
那么此时,k (r )=πr2 。在点事件处于聚集分布时,如果距离尺度比较小,则k (r )的观测值就会大于πr 。估计式为:
K ̂ ( r ) = A n 2 ∑ i n ∑ j n I i j ( r ) W i j ( r ) (3)
式中:A 代表着研究区域的面积;λ ̂ = n A E ̂ = n - 1 ∑ i n ∑ j n I i j ( r ) W i j ( r ) n 代表研究区域中的点事件数量;i 和j 代表样方中任意两个拥有相同特征的点事件;Iij (r )=1(当dij ≤r 时成立,rij 为i 和j 之间的距离);Wij (r )代表边缘校正的权重,等于以点事件i 作圆心、以rij 作半径的圆位于研究区域中的弧长与整个圆周长的比值。
一般情况下在一定尺度中点事件的分布类型是以L (r )函数的估计值来确定的。在距离尺度d 中,如果L ̂ ( r ) > 0 L ̂ ( r ) = 0 L ̂ ( r ) < 0
L ̂ ( r ) = k ̂ ( r ) / π - r (4)
采取Monte Carlo方法求解得到随机分布的上下两条包迹线,如果研究对象的实际值位于上下包迹线之间的区间内,那么在这个距离尺度中的点事件就是随机分布;如果研究对象的实际值位于上下包迹线的上方,那么在这个距离尺度中的点事件则为聚集分布;如果研究对象的实际值位于上下包迹线的下方,那么在这个距离尺度中的点事件则为均匀分布。实际的操作方法为选取m n L (r )值,之后把对应在每个d 的m 个L (r )进行从小至大的排序,各自选取最大值与最小值用作上下包迹线的取值范围。
1.3.2 零模型的选择
成功地运用Ripley's K函数分析所要解决的生态学问题的关键是选择合适的零模型,并且能够合理地解释实测数据与零模型的偏离。在本项研究中,为了揭示沙鞭种群空间格局的分布特征,选择了完全空间随机模型、泊松聚块模型和嵌套双聚块模型。
1.3.3 完全空间随机模型
完全空间随机模型(CSR)是简单、常用的均质泊松过程 (homogeneous Poisson process,HP)。在这个模型中任何一点(或个体)在研究区域内任何一个位置上出现(或发生)的机会是相同的;同时,点间(或个体间)是相互独立的,也就是说,任何两点间均不发生相互作用。常作为零假设(null test)来检验物种的空间分布格局,CSR模型k 函数同上文定义。
1.3.4 泊松聚块模型
泊松聚块模型(NS)亦被称作均质托马斯过程(homogeneous Thomas process,TH),用以描述聚块机制。其假设母体事件以完全空间随机过程发生,而每一个母体在其周围按一定的概率分布产生随机数量的子代个体,且这些子代个体在空间的分布上遵循某双变量概率密度函数。如果子代个体的数量遵循泊松分布且其在空间的位置相对母体而言符合双变量高斯分布,那么,子代个体的发生就符合泊松聚块过程。泊松聚块模型的k 函数表达式如下:
k r , σ , ρ = π r 2 + 1 - e x p ( - r 2 / 4 σ 2 ) ρ (5)
式中:r 为尺度;ρ 为该过程中母体的密度;σ 2
1.3.5 嵌套双聚块模型
嵌套双聚块模型(Nested double-cluster process,DC)是泊松聚块模型的多代扩展,泊松聚块过程的子代产生自己的后代个体。详细的描述参考文献[22 ],嵌套双聚块模型的k 函数表达式如下:
k r , σ 1 , ρ 1 , σ 2 , ρ 2 = π r 2 + 1 - e x p ( - r 2 / 4 σ 2 2 ) ρ 2 + π r 2 + 1 - e x p ( - r 2 / 4 σ 2 s u m ) ρ 2
且r 2 s u m = σ 1 + σ 2 (6)
参数r 、 ρ σ 2
1.3.6 数据处理
采用CAD软件来绘制植物株丛在地面的投影分布图,使用GetData Graph Digitizer 2.22软件将植物株丛在地面投影的分布图里各株丛的实际坐标值提取出来,最后把得到的坐标值录入Excel软件里,在这之后即可把数据应用于种间空间格局及其关联分析中。使用Thorsten Wiegand教授提供的Programita 2018软件进行计算。文中采用的步长为1 cm,Monte Carlo随机模拟的数目为199次,得到上下两条包迹线围成的95%置信区间。
2 结果与分析
2.1 基于完全随机模型的点格局分析
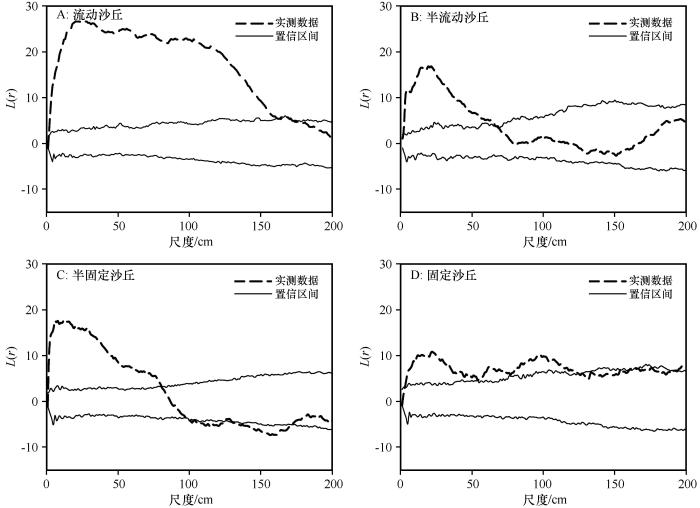
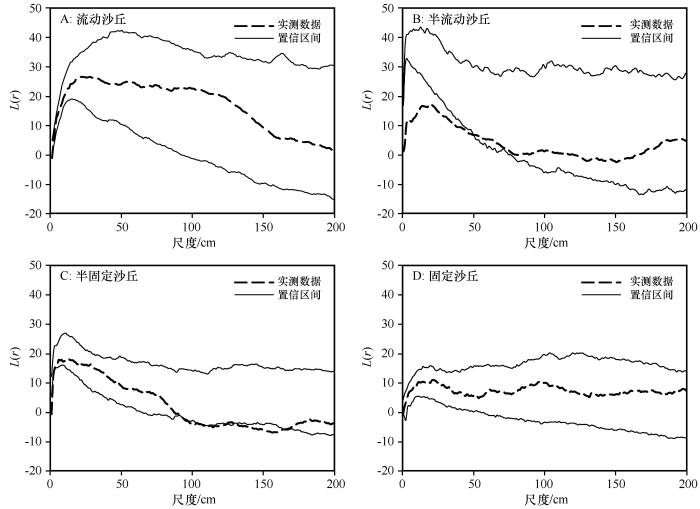
流动沙丘中,0—162 cm和169—170 cm尺度沙鞭种群地上分株表现为聚集分布,162—169 cm和170—200 cm尺度表现为随机分布(图3 A);半流动沙丘中,0—65 cm尺度沙鞭种群地上分株表现为聚集分布,65—200 cm尺度表现为随机分布(图3 B);半固定沙丘中,0—80 cm尺度沙鞭种群地上分株表现为聚集分布,98—124 cm和130—171 cm尺度表现为均匀分布,80—98、124—130 cm和171—200 cm尺度表现为随机分布(图3 C);固定沙丘中,4—127、161—163 cm和195—200 cm尺度沙鞭种群地上分株表现为聚集分布,0—4、127—161 cm和163—195 cm尺度表现为随机分布(图3 D)。
图3
图3
不同沙丘类型沙鞭(Psammochloa villosa )种群基于完全空间随机模型的点格局分析
Fig.3
Point pattern analysis of Psammochloa villosa in different dune types based on complete atial randomness
2.2 基于泊松聚块模型的点格局分析
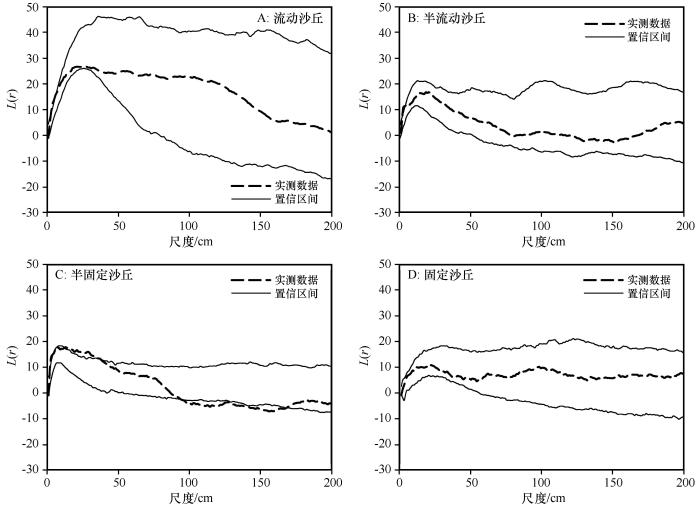
流动沙丘、半流动沙丘和固定沙丘环境下,沙鞭种群地上分株分布格局符合泊松聚块模型(图4 A、B、D),仅在半固定沙丘,2—37 cm尺度符合正向偏离泊松聚块模型,95—168 cm尺度符合负向偏离泊松聚块模型(图4 C)。
图4
图4
不同沙丘类型沙鞭(Psammochloa villosa )种群基于泊松聚块模型的点格局分析
Fig.4
Point pattern analysis of Psammochloa villosa in different dune types based on Poisson cluster process
2.3 基于嵌套双聚块模型的点格局分析
流动沙丘和固定沙丘环境下,沙鞭种群地上分株分布格局符合嵌套双聚块模型(图5 A、D),半流动沙丘,0—50 cm尺度符合负向偏离嵌套双聚块模型,其他尺度符合离嵌套双聚块模型(图5 B);半固定沙丘,0—3、98—105、110—123 cm和130—168 cm尺度符合负向偏离嵌套双聚块模型(图5 C)。
图5
图5
不同沙丘类型沙鞭(Psammochloa villosa )种群基于嵌套双聚块模型的点格局分析
Fig.5
Point pattern analysis of Psammochloa villosa in different dune types based on double cluster process
3 讨论与结论
空间分布格局研究的不断进步,为揭示植物种群的内部结构、扩散规律、更新机制以及共存问题等提供了重要手段[23 -27 ] 。其中,基于各种机理的模型中,完全空间随机模型最为常见,这种模型可以揭示植物种群在不同尺度上呈现的分布格局类型,如聚集分布、均匀分布和随机分布,对于认识植被空间分布的机理具有一定的价值和作用。
基于完全空间随机模型点格局分析的结果显示,沙鞭种群在不同尺度上均表现出了聚集分布(图3 ),这可能与沙鞭种群是兼性克隆植物相关,沙鞭母体可以通过根茎繁殖,在其周围产生分株,分株又产生新的分株,形成分株系统,分株系统中的植株可以继续根茎繁殖,形成下一级分株系统,所有的分株系统构成了一个大聚块,各分株系统之间可以通过克隆整合进行资源共享和风险分摊[11 ,27 -28 ] ,故而沙鞭种群在不同风沙环境下均有较大尺度的聚集分布。从不同风沙环境中聚集尺度大小来看,沙鞭种群存在从流动沙丘至固定沙丘聚集尺度先减小后增大的变化规律,这可能与沙鞭种群获得群落资源的能力,以及群落物种增加导致的种间竞争加剧相关。在流动沙丘中,沙鞭可以通过分蘖和根茎繁殖方式在适应异质性强且水分匮乏环境中具有独特优势[5 ,7 ] ,一旦在沙地定居,就能够迅速向周围裸沙扩张[11 ,18 ] ,并可通过根茎和分株形成植被斑块。同时,流动沙丘中沙鞭个体需要抱团来获取资源以确保种群存活,导致沙鞭在0—162 cm和169—170 cm尺度呈现为聚集分布格局。
在半流动沙丘中,沙鞭种群在0—65 cm尺度范围内表现为聚集分布,65—200 cm尺度随机分布,沙鞭聚集分布的尺度显著低于流动沙丘。这可能与半流动沙丘中的水资源不足以支持沙鞭种群在大尺度上聚集分布并形成更大的植被斑块,在一定程度上限制了沙鞭种群的根茎和种子繁殖过程。原有植被斑块内个体种内合作继续占据群落资源,而较大尺度上的个体则可能通过根茎拓展和种子扩散的方式将子代输送到距离母株相对较远的地方,表现为种内个体之间的随机分布,这也与植物的向外扩张基因(遗传)导致克隆生长能够将子代扩散到更广泛的区域而避免与来自相同个体的其他配子和后代竞争的研究结果一致[25 -26 ,29 -30 ] 。
半固定沙丘中,沙鞭种群在0—80 cm尺度表现为聚集分布,而在98—124 cm和130—171 cm尺度表现为均匀分布,这种均匀分布格局可能与半流动沙丘中沙鞭种内种间关系变化相一致。有限水分资源加剧了较大尺度上沙鞭植被斑块内个体的竞争关系,引起了种内自疏效应。半流动沙丘中随机分布格局转变为均匀分布格局,以牺牲沙鞭斑块外围个体的策略来确保原有植被斑块的稳定,这种策略与西北针茅等植物种群受到资源限制采取的适应策略一致[31 ] 。在半固定沙丘中聚集分布的格局从半流动沙丘0—65 cm尺度增长到在0—80 cm尺度,这可能与沙鞭、沙米(Agriophyllum squarrosum )、虫实(Corispermum hyssopifolium )等一二年生草本物种展开种间竞争相关。在群落调查中也发现了比较特殊的群落属性:在流动和半流动沙丘中,沙鞭多为单种群落;在半固定沙丘中,除沙米、虫实等一二年生草本物种外,出现了沙蒿等多年生草本,而在固定沙丘中则出现了泡泡刺(Nitraria sphaerocarpa )、霸王(Zygophyllum xanthoxylon )等灌木或半灌木物种。沙鞭与多年生草本和灌木物种之间的种间竞争关系,可能会减弱沙鞭种内竞争导致的自疏现象,种内分株之间表现出合作关系争夺群落资源。因此,固定沙丘环境中,沙鞭在4—127、161—163 cm和195—200 cm尺度表现出聚集分布格局。
从聚集分布格局强度来看,表现出流动沙丘聚集强度最高,且从流动沙丘向固定沙丘减弱的趋势。这可能与两个原因相关:①流动沙丘中沙鞭种群需要高强度聚集分布才能确保物种获得群落中的水分资源,进而保障个体存活和种群延续,而在其他风沙环境下,沙鞭种群个体面临的群落环境有所好转,种内自疏和个体外移行为造成个体间较低的聚集分布强度;②其他物种的种间竞争关系和有限的水土资源限制了沙鞭个体间聚集分布格局,沙鞭个体间聚集强度被迫降低。
基于完全空间随机模型的点格局分析可以揭示种群内个体的空间分布方式或配置特点,但无法直接揭示种群聚集分布或均匀分布格局的内在机制或者形成机理,泊松聚块模型和嵌套双聚块模型描述的是种群聚集分布的内在形成机制,二者紧密联系。使用泊松聚块模型检验的种群格局在一定尺度位于置信区间之上,表明在种群聚集分布的聚块内可能存在较高密度的小聚块,且可通过嵌套双聚块模型加以进一步验证[32 -33 ] 。
流动沙丘、半流动沙丘和固定沙丘环境下,沙鞭种群地上分株分布格局符合泊松聚块模型(图4 A、B、D),仅在半固定沙丘,2—37 cm尺度符合正向偏离泊松聚块模型,95—168 cm尺度符合负向偏离泊松聚块模型(图4 C)。这表明流动沙丘、半流动沙丘和固定沙丘环境下,沙鞭种群空间格局在大聚块中不存在聚集密度较高的小聚块,仅在半固定沙丘中,沙鞭种群聚集分布的聚块内可能存在较高密度的小聚块。这可能与不同风沙环境下沙鞭种群小尺度上空间格局形成与变化受到植物生长环境和植物自身的生物学特征影响有关。在流动沙丘、半流动沙丘中未受到其他物种的种间竞争胁迫,种群分株系统是一个整体[11 ,34 -36 ] ,所有的分株系统构成了一个大聚块,沙鞭种群以一个大的斑块整体(大聚块)共同应对极端干旱风沙环境以争取群落水分等资源[37 ] ,从而表现为符合泊松聚块模型;而在半固定沙丘中,沙鞭大聚块被沙米、虫实和沙蒿等草本物种切割,原有植被斑块内部出现了沙鞭未占据的“资源空斑”,沙鞭种群个体为了应对种间竞争和“空斑资源”争夺聚集在一起,在较小尺度上形成了小聚块实现资源共享和风险分摊,从而表现出牺牲斑块外围个体,斑块内部个体聚集确保原有植被斑块稳定的策略,种群格局表现为嵌套双聚块过程(图5 C)。但在固定沙丘中,霸王和泡泡刺等物种与沙鞭在不同尺度上出现了种间竞争关系,群落内的“空斑资源”和剩余资源消失,小尺度上形成的小聚块再度被切割,从而种群格局表现为符合泊松聚块模型。这与笔者在陕西榆林调研沙鞭种群分布格局发现的现象相一致:当沙柳(Salix cheilophila )、西北针茅(Stipa krylovii )、紫菀(Aster tataricus )等物种成为群落优势种后,沙鞭植被斑块会彻底破碎,沙鞭个体呈现零星分布,与流动和半流动沙丘中沙鞭分株连线成片、斑块状分布的空间分布格局完全不同。
沙鞭种群在半流动沙丘中0—50 cm尺度符合负向偏离嵌套双聚块模型,半固定沙丘中95—168 cm尺度符合负向偏离泊松聚块模型,0—3、98—105、110—123 cm和130—168 cm尺度符合负向偏离嵌套双聚块模型,这与泊松聚块模型和嵌套双聚块模型所代表的含义不一致,可能是干旱风沙环境形成的特殊植物生长环境和植物自身生物学特性所致,需要下一步进行深入研究。
综上所述,流动沙丘中分蘖和根茎繁殖方式形成植被斑块促进种群占据和获取群落资源,但在半固定和固定沙丘中加剧了种内个体间的竞争压力;沙米、沙蒿和泡泡刺等物种切割沙鞭斑块,从而影响沙鞭斑块的稳定性,确保斑块内部个体高度聚集,牺牲斑块外围个体是沙鞭种群空间布局和资源占据的生态适应策略之一。
参考文献
View Option
[1]
Ghani A E Bornkamm M Sawaf R E et al Plant species distribution and spatial habitat heterogeneity in the landscape of urbanizing desert ecosystems in Egypt
[J].Journal of Arid Environments ,2011 ,14 (4 ):585 -616 .
[本文引用: 2]
[2]
Li Y P He X L Zhao L L Tempo-spatial dynamics of arbuscular mycorrhizal fungi under clonal plant Psammochloa villosa Trin.Bor in Mu Us Sandland
[J].European Journal of Soil Biology ,2010 ,46 (5 ):295 -301 .
[3]
Huang Z Y Dong M Gutterman Y Factors influencing seed dormancy and germination in sand,and seedling survival under desiccation,of Psammochloa villosa (Poaceae),inhabiting the moving sand dunes of Ordos,China
[J].Plant and Soil ,2004 ,259 (1/2 ):231 -241 .
[本文引用: 1]
[4]
任珩 ,赵成章 高寒退化草狼毒与赖草种群空间格局及竞争关系研究
[J].生态学报 ,2013 ,33 (2 ):435 -442 .
[本文引用: 1]
[5]
Dong M Alaten B Clonal plasticity in response to rhizome severing and heterogeneous resource supply in the rhizomatous grass Psammochloa villosa in an Inner Mongolian dune,China
[J].Plant Ecology ,1999 ,141 (1/2 ):53 -58 .
[本文引用: 4]
[6]
杨洪晓 ,张金屯 ,吴波 ,等 毛乌素沙地油蒿种群点格局分析
[J].植物生态学报 ,2006 ,30 (4 ):563 -570 .
[本文引用: 1]
[7]
Yu F H Dong M Krusi B Clonal integration helps Psammochloa villosa survive sand burial in an inland dune
[J].New Phytologist ,2004 ,162 (3 ):697 -704 .
[本文引用: 3]
[8]
Wang Y H Dong M Yu F H et al Mechanical shaking and soil water affect the growth of Psammochloa villosa in the Mu Us Sandland
[J].Journal of Arid Environments ,2011 ,75 (10 ):974 -977 .
[9]
操国兴 ,钟章成 ,刘芸 ,等 缙云山川鄂连蕊茶种群空间分布格局研究
[J].生物学杂志 ,2003 (1 ):10 -12 .
[本文引用: 1]
[10]
许爱云 ,曹兵 ,谢云 干旱风沙区煤炭基地12种草本植物对干旱胁迫的生理生态响应及抗旱性评价
[J].草业学报 ,2020 ,29 (10 ):22 -34 .
[本文引用: 1]
[11]
叶学华 ,董鸣 毛乌素沙地克隆植物对风蚀坑的修复
[J].生态学报 ,2011 ,31 (19 ):5505 -5511 .
[本文引用: 4]
[12]
朱雅娟 ,董鸣 ,黄振英 沙埋和种子大小对固沙禾草沙鞭的种子萌发与幼苗出土的影响
[J].植物生态学报 ,2005 ,29 (5 ):730 -739 .
[本文引用: 1]
[13]
朱雅娟 ,董鸣 ,黄振英 种子萌发和幼苗生长对沙丘环境的适应机制
[J].应用生态学报 ,2006 ,17 (1 ):137 -142 .
[本文引用: 1]
[14]
葛颂 ,王可青 ,董鸣 毛乌素沙地根茎灌木羊柴的遗传多样性和克隆结构
[J].植物学报 ,1999 (3 ):75 -80 .
[本文引用: 1]
[15]
吕婷 ,刘涛 ,梁瑞芳 ,等 沙生植物沙鞭不同居群形态变异研究
[J].植物研究 ,2021 ,41 (1 ):60 -66 .
[本文引用: 1]
[16]
吕婷 沙生牧草沙鞭的遗传多样性研究
[D].西宁 :青海师范大学 ,2019 .
[本文引用: 1]
[17]
吕婷 ,刘玉萍 ,周勇辉 ,等 荒漠植物沙鞭(Psammochloa villosa )种质资源收集及谱系遗传分化初探
[J].草地学报 ,2018 ,26 (3 ):733 -740 .
[本文引用: 1]
[18]
陈玉福 ,于飞海 ,张称意 ,等 根茎禾草沙鞭的克隆生长在毛乌素沙地斑块动态中的作用
[J].生态学报 ,2001 ,21 (11 ):1745 -1750 .
[本文引用: 2]
[19]
赵文智 ,刘鹄 干旱、半干旱环境降水脉动对生态系统的影响
[J].应用生态学报 ,2011 ,22 (1 ):243 -249 .
[本文引用: 2]
[20]
赵哈林 ,周瑞莲 ,赵学勇 ,等 科尔沁沙地沙漠化正、逆过程的地面判别方法
[J].中国沙漠 ,2008 ,28 (1 ):8 -15 .
[本文引用: 1]
[21]
左小安 ,赵哈林 ,赵学勇 ,等 科尔沁沙地不同恢复年限退化植被的物种多样性
[J].草业学报 ,2009 ,18 (4 ):9 -16 .
[本文引用: 1]
[22]
Diggle P J Statistical Analysis of Spatial Point Patterns [M].New York,USA :Academic Press ,1983 .
[本文引用: 2]
[23]
王鑫厅 ,侯亚丽 ,刘芳 ,等 羊草+大针茅草原退化群落优势种群空间点格局分析
[J].植物生态学报 ,2011 ,35 (12 ):1281 -1289 .
[本文引用: 1]
[24]
高福元 ,赵成章 高寒退化草地狼毒种群株丛间格局控制机理
[J].生态学报 ,2013 ,33 (10 ):3114 -3121 .
[25]
Verdú M Traveset A Early emergence enhances plant fitness:a phylogenetically controlled meta-analysis
[J].Ecology ,2005 ,86 (6 ):1385 -1394 .
[本文引用: 1]
[26]
Howe H F Smallwood J Ecology of seed dispersal
[J].Annual Review of Ecology and Systematics ,1982 ,13 :201 -228 .
[本文引用: 1]
[27]
Cheplick G P Sibling competition in plants
[J].Journal of Ecology ,1992 ,80 (3 ):567 -575 .
[本文引用: 2]
[28]
Nilsson P Fagerström T Tuomi J et al Does seed dormancy benefit the mother plant by reducing sib competition?
[J].Evolutionary Ecology ,1994 ,8 (4 ):422 -430 .
[本文引用: 1]
[29]
Dyer A R Maternal and sibling factors induce dormancy in dimorphic seed pairs of Aegilops triuncialis
[J].Plant Ecology ,2004 ,172 (2 ):211 -218 .
[本文引用: 1]
[30]
Venable D L Brown J S The population-dynamic functions of seed dispersal
[J].Vegetatio ,1993 ,107/108 (1 ):31 -55 .
[本文引用: 1]
[31]
任珩 ,赵成章 ,高福元 ,等 退化草地阿尔泰针茅生殖株丛与非生殖株丛的空间格局
[J].生态学报 ,2012 ,32 (21 ):6909 -6916 .
[本文引用: 1]
[32]
王鑫厅 ,侯亚丽 ,梁存柱 ,等 基于不同零模型的点格局分析
[J].生物多样性 ,2012 ,20 (2 ):151 -158 .
[本文引用: 1]
[33]
Hellstrom K Kytoviita M M Tuomi J et al Plasticity of clonal integration in the perennial herb Linaria vulgaris after damage
[J].Functional Ecology ,2006 ,20 (3 ):413 -420 .
[本文引用: 1]
[34]
Pluess A R Stöcklin J The importance of population origin and environment on clonal and sexual reproduction in the alpine plant Geum reptans
[J].Functional Ecology ,2005 ,19 (2 ):228 -237 .
[本文引用: 1]
[35]
祁永 放牧对草原群落特征及种群繁殖特性的影响
[D].北京 :中国农业大学 ,2005 .
[36]
Ikegami M Whigham D F Werger M J A Responses of rhizome length and ramet production to resource availability in the clonal sedge Scirpus olneyi A.Gray
[J].Plant Ecology ,2007 ,189 (2 ):247 -259 .
[本文引用: 1]
[37]
张晓娜 ,哈达朝鲁 ,潘庆民 刈割干扰下内蒙古草原两种丛生禾草繁殖策略的适应性调节
[J].植物生态学报 ,2010 ,34 (3 ):253 -262 .
[本文引用: 1]
Plant species distribution and spatial habitat heterogeneity in the landscape of urbanizing desert ecosystems in Egypt
2
2011
... 种群空间分布格局是植物种群自身生物学特性与环境因素相互作用的结果,一直是生态学研究的热点[1 -4 ] .点格局方法能够以由小到大的一系列连续尺度分析点状物体平面分布格局[5 -6 ] ,对于揭示植物种群对外部环境的生态学适应机制,展现物种在群落中的生态学地位,指示该物种之间的相互关系有着较大的意义[7 -9 ] .植物功能性状是植物适应环境的一种表现形式,会随外界环境变化而呈现出相应的形态结构与生理特性,是研究植物响应环境变化的有力工具[10 ] .植物种群在面对不同风沙条件时,需要不断地调整株高、叶片、种群密度等特征.因此,开展沙鞭地上分株分布格局对不同风沙环境响应的研究能够反映出植物种群的资源投入、选择和适应策略,对于揭示植物种群应对干旱风沙等极端环境的响应机制具有重要意义. ...
... 沙鞭(Psammochloa villosa )作为干旱地区流动沙丘先锋物种,对于干旱风沙环境具有极强生态适应能力,在流动沙丘上分布的沙鞭斑块与流沙斑块相互镶嵌,已成为流动沙丘的独特景观[1 ,3 ,5 ,11 ] ,主要分布在内蒙古库布齐沙漠、浑善达克沙地西部、巴丹吉林沙漠和甘肃河西走廊沙地等地区.目前关于沙鞭的研究主要包括有性繁殖[12 -13 ] 与无性繁殖[5 ,7 ,14 ] 、不同居群形态变异[15 ] 、遗传多样性[16 -17 ] 、改造沙化景观[18 ] ,极大地丰富和发展了沙鞭的种群特征研究,但是关于不同沙丘生境下沙鞭种群空间分布与繁殖策略特征的研究尚未涉及.基于此,本研究选择流动、半流动、半固定和固定沙丘,开展沙鞭种群地上分株分布格局研究,旨在研究沙丘流动性对于沙鞭种群空间分布格局的影响,为了解极端干旱地区植物种群对环境胁迫的响应机理提供科学依据. ...
Tempo-spatial dynamics of arbuscular mycorrhizal fungi under clonal plant Psammochloa villosa Trin.Bor in Mu Us Sandland
0
2010
Factors influencing seed dormancy and germination in sand,and seedling survival under desiccation,of Psammochloa villosa (Poaceae),inhabiting the moving sand dunes of Ordos,China
1
2004
... 沙鞭(Psammochloa villosa )作为干旱地区流动沙丘先锋物种,对于干旱风沙环境具有极强生态适应能力,在流动沙丘上分布的沙鞭斑块与流沙斑块相互镶嵌,已成为流动沙丘的独特景观[1 ,3 ,5 ,11 ] ,主要分布在内蒙古库布齐沙漠、浑善达克沙地西部、巴丹吉林沙漠和甘肃河西走廊沙地等地区.目前关于沙鞭的研究主要包括有性繁殖[12 -13 ] 与无性繁殖[5 ,7 ,14 ] 、不同居群形态变异[15 ] 、遗传多样性[16 -17 ] 、改造沙化景观[18 ] ,极大地丰富和发展了沙鞭的种群特征研究,但是关于不同沙丘生境下沙鞭种群空间分布与繁殖策略特征的研究尚未涉及.基于此,本研究选择流动、半流动、半固定和固定沙丘,开展沙鞭种群地上分株分布格局研究,旨在研究沙丘流动性对于沙鞭种群空间分布格局的影响,为了解极端干旱地区植物种群对环境胁迫的响应机理提供科学依据. ...
高寒退化草狼毒与赖草种群空间格局及竞争关系研究
1
2013
... 种群空间分布格局是植物种群自身生物学特性与环境因素相互作用的结果,一直是生态学研究的热点[1 -4 ] .点格局方法能够以由小到大的一系列连续尺度分析点状物体平面分布格局[5 -6 ] ,对于揭示植物种群对外部环境的生态学适应机制,展现物种在群落中的生态学地位,指示该物种之间的相互关系有着较大的意义[7 -9 ] .植物功能性状是植物适应环境的一种表现形式,会随外界环境变化而呈现出相应的形态结构与生理特性,是研究植物响应环境变化的有力工具[10 ] .植物种群在面对不同风沙条件时,需要不断地调整株高、叶片、种群密度等特征.因此,开展沙鞭地上分株分布格局对不同风沙环境响应的研究能够反映出植物种群的资源投入、选择和适应策略,对于揭示植物种群应对干旱风沙等极端环境的响应机制具有重要意义. ...
Clonal plasticity in response to rhizome severing and heterogeneous resource supply in the rhizomatous grass Psammochloa villosa in an Inner Mongolian dune,China
4
1999
... 种群空间分布格局是植物种群自身生物学特性与环境因素相互作用的结果,一直是生态学研究的热点[1 -4 ] .点格局方法能够以由小到大的一系列连续尺度分析点状物体平面分布格局[5 -6 ] ,对于揭示植物种群对外部环境的生态学适应机制,展现物种在群落中的生态学地位,指示该物种之间的相互关系有着较大的意义[7 -9 ] .植物功能性状是植物适应环境的一种表现形式,会随外界环境变化而呈现出相应的形态结构与生理特性,是研究植物响应环境变化的有力工具[10 ] .植物种群在面对不同风沙条件时,需要不断地调整株高、叶片、种群密度等特征.因此,开展沙鞭地上分株分布格局对不同风沙环境响应的研究能够反映出植物种群的资源投入、选择和适应策略,对于揭示植物种群应对干旱风沙等极端环境的响应机制具有重要意义. ...
... 沙鞭(Psammochloa villosa )作为干旱地区流动沙丘先锋物种,对于干旱风沙环境具有极强生态适应能力,在流动沙丘上分布的沙鞭斑块与流沙斑块相互镶嵌,已成为流动沙丘的独特景观[1 ,3 ,5 ,11 ] ,主要分布在内蒙古库布齐沙漠、浑善达克沙地西部、巴丹吉林沙漠和甘肃河西走廊沙地等地区.目前关于沙鞭的研究主要包括有性繁殖[12 -13 ] 与无性繁殖[5 ,7 ,14 ] 、不同居群形态变异[15 ] 、遗传多样性[16 -17 ] 、改造沙化景观[18 ] ,极大地丰富和发展了沙鞭的种群特征研究,但是关于不同沙丘生境下沙鞭种群空间分布与繁殖策略特征的研究尚未涉及.基于此,本研究选择流动、半流动、半固定和固定沙丘,开展沙鞭种群地上分株分布格局研究,旨在研究沙丘流动性对于沙鞭种群空间分布格局的影响,为了解极端干旱地区植物种群对环境胁迫的响应机理提供科学依据. ...
... [5 ,7 ,14 ]、不同居群形态变异[15 ] 、遗传多样性[16 -17 ] 、改造沙化景观[18 ] ,极大地丰富和发展了沙鞭的种群特征研究,但是关于不同沙丘生境下沙鞭种群空间分布与繁殖策略特征的研究尚未涉及.基于此,本研究选择流动、半流动、半固定和固定沙丘,开展沙鞭种群地上分株分布格局研究,旨在研究沙丘流动性对于沙鞭种群空间分布格局的影响,为了解极端干旱地区植物种群对环境胁迫的响应机理提供科学依据. ...
... 基于完全空间随机模型点格局分析的结果显示,沙鞭种群在不同尺度上均表现出了聚集分布(图3 ),这可能与沙鞭种群是兼性克隆植物相关,沙鞭母体可以通过根茎繁殖,在其周围产生分株,分株又产生新的分株,形成分株系统,分株系统中的植株可以继续根茎繁殖,形成下一级分株系统,所有的分株系统构成了一个大聚块,各分株系统之间可以通过克隆整合进行资源共享和风险分摊[11 ,27 -28 ] ,故而沙鞭种群在不同风沙环境下均有较大尺度的聚集分布.从不同风沙环境中聚集尺度大小来看,沙鞭种群存在从流动沙丘至固定沙丘聚集尺度先减小后增大的变化规律,这可能与沙鞭种群获得群落资源的能力,以及群落物种增加导致的种间竞争加剧相关.在流动沙丘中,沙鞭可以通过分蘖和根茎繁殖方式在适应异质性强且水分匮乏环境中具有独特优势[5 ,7 ] ,一旦在沙地定居,就能够迅速向周围裸沙扩张[11 ,18 ] ,并可通过根茎和分株形成植被斑块.同时,流动沙丘中沙鞭个体需要抱团来获取资源以确保种群存活,导致沙鞭在0—162 cm和169—170 cm尺度呈现为聚集分布格局. ...
毛乌素沙地油蒿种群点格局分析
1
2006
... 种群空间分布格局是植物种群自身生物学特性与环境因素相互作用的结果,一直是生态学研究的热点[1 -4 ] .点格局方法能够以由小到大的一系列连续尺度分析点状物体平面分布格局[5 -6 ] ,对于揭示植物种群对外部环境的生态学适应机制,展现物种在群落中的生态学地位,指示该物种之间的相互关系有着较大的意义[7 -9 ] .植物功能性状是植物适应环境的一种表现形式,会随外界环境变化而呈现出相应的形态结构与生理特性,是研究植物响应环境变化的有力工具[10 ] .植物种群在面对不同风沙条件时,需要不断地调整株高、叶片、种群密度等特征.因此,开展沙鞭地上分株分布格局对不同风沙环境响应的研究能够反映出植物种群的资源投入、选择和适应策略,对于揭示植物种群应对干旱风沙等极端环境的响应机制具有重要意义. ...
Clonal integration helps Psammochloa villosa survive sand burial in an inland dune
3
2004
... 种群空间分布格局是植物种群自身生物学特性与环境因素相互作用的结果,一直是生态学研究的热点[1 -4 ] .点格局方法能够以由小到大的一系列连续尺度分析点状物体平面分布格局[5 -6 ] ,对于揭示植物种群对外部环境的生态学适应机制,展现物种在群落中的生态学地位,指示该物种之间的相互关系有着较大的意义[7 -9 ] .植物功能性状是植物适应环境的一种表现形式,会随外界环境变化而呈现出相应的形态结构与生理特性,是研究植物响应环境变化的有力工具[10 ] .植物种群在面对不同风沙条件时,需要不断地调整株高、叶片、种群密度等特征.因此,开展沙鞭地上分株分布格局对不同风沙环境响应的研究能够反映出植物种群的资源投入、选择和适应策略,对于揭示植物种群应对干旱风沙等极端环境的响应机制具有重要意义. ...
... 沙鞭(Psammochloa villosa )作为干旱地区流动沙丘先锋物种,对于干旱风沙环境具有极强生态适应能力,在流动沙丘上分布的沙鞭斑块与流沙斑块相互镶嵌,已成为流动沙丘的独特景观[1 ,3 ,5 ,11 ] ,主要分布在内蒙古库布齐沙漠、浑善达克沙地西部、巴丹吉林沙漠和甘肃河西走廊沙地等地区.目前关于沙鞭的研究主要包括有性繁殖[12 -13 ] 与无性繁殖[5 ,7 ,14 ] 、不同居群形态变异[15 ] 、遗传多样性[16 -17 ] 、改造沙化景观[18 ] ,极大地丰富和发展了沙鞭的种群特征研究,但是关于不同沙丘生境下沙鞭种群空间分布与繁殖策略特征的研究尚未涉及.基于此,本研究选择流动、半流动、半固定和固定沙丘,开展沙鞭种群地上分株分布格局研究,旨在研究沙丘流动性对于沙鞭种群空间分布格局的影响,为了解极端干旱地区植物种群对环境胁迫的响应机理提供科学依据. ...
... 基于完全空间随机模型点格局分析的结果显示,沙鞭种群在不同尺度上均表现出了聚集分布(图3 ),这可能与沙鞭种群是兼性克隆植物相关,沙鞭母体可以通过根茎繁殖,在其周围产生分株,分株又产生新的分株,形成分株系统,分株系统中的植株可以继续根茎繁殖,形成下一级分株系统,所有的分株系统构成了一个大聚块,各分株系统之间可以通过克隆整合进行资源共享和风险分摊[11 ,27 -28 ] ,故而沙鞭种群在不同风沙环境下均有较大尺度的聚集分布.从不同风沙环境中聚集尺度大小来看,沙鞭种群存在从流动沙丘至固定沙丘聚集尺度先减小后增大的变化规律,这可能与沙鞭种群获得群落资源的能力,以及群落物种增加导致的种间竞争加剧相关.在流动沙丘中,沙鞭可以通过分蘖和根茎繁殖方式在适应异质性强且水分匮乏环境中具有独特优势[5 ,7 ] ,一旦在沙地定居,就能够迅速向周围裸沙扩张[11 ,18 ] ,并可通过根茎和分株形成植被斑块.同时,流动沙丘中沙鞭个体需要抱团来获取资源以确保种群存活,导致沙鞭在0—162 cm和169—170 cm尺度呈现为聚集分布格局. ...
Mechanical shaking and soil water affect the growth of Psammochloa villosa in the Mu Us Sandland
0
2011
缙云山川鄂连蕊茶种群空间分布格局研究
1
2003
... 种群空间分布格局是植物种群自身生物学特性与环境因素相互作用的结果,一直是生态学研究的热点[1 -4 ] .点格局方法能够以由小到大的一系列连续尺度分析点状物体平面分布格局[5 -6 ] ,对于揭示植物种群对外部环境的生态学适应机制,展现物种在群落中的生态学地位,指示该物种之间的相互关系有着较大的意义[7 -9 ] .植物功能性状是植物适应环境的一种表现形式,会随外界环境变化而呈现出相应的形态结构与生理特性,是研究植物响应环境变化的有力工具[10 ] .植物种群在面对不同风沙条件时,需要不断地调整株高、叶片、种群密度等特征.因此,开展沙鞭地上分株分布格局对不同风沙环境响应的研究能够反映出植物种群的资源投入、选择和适应策略,对于揭示植物种群应对干旱风沙等极端环境的响应机制具有重要意义. ...
干旱风沙区煤炭基地12种草本植物对干旱胁迫的生理生态响应及抗旱性评价
1
2020
... 种群空间分布格局是植物种群自身生物学特性与环境因素相互作用的结果,一直是生态学研究的热点[1 -4 ] .点格局方法能够以由小到大的一系列连续尺度分析点状物体平面分布格局[5 -6 ] ,对于揭示植物种群对外部环境的生态学适应机制,展现物种在群落中的生态学地位,指示该物种之间的相互关系有着较大的意义[7 -9 ] .植物功能性状是植物适应环境的一种表现形式,会随外界环境变化而呈现出相应的形态结构与生理特性,是研究植物响应环境变化的有力工具[10 ] .植物种群在面对不同风沙条件时,需要不断地调整株高、叶片、种群密度等特征.因此,开展沙鞭地上分株分布格局对不同风沙环境响应的研究能够反映出植物种群的资源投入、选择和适应策略,对于揭示植物种群应对干旱风沙等极端环境的响应机制具有重要意义. ...
毛乌素沙地克隆植物对风蚀坑的修复
4
2011
... 沙鞭(Psammochloa villosa )作为干旱地区流动沙丘先锋物种,对于干旱风沙环境具有极强生态适应能力,在流动沙丘上分布的沙鞭斑块与流沙斑块相互镶嵌,已成为流动沙丘的独特景观[1 ,3 ,5 ,11 ] ,主要分布在内蒙古库布齐沙漠、浑善达克沙地西部、巴丹吉林沙漠和甘肃河西走廊沙地等地区.目前关于沙鞭的研究主要包括有性繁殖[12 -13 ] 与无性繁殖[5 ,7 ,14 ] 、不同居群形态变异[15 ] 、遗传多样性[16 -17 ] 、改造沙化景观[18 ] ,极大地丰富和发展了沙鞭的种群特征研究,但是关于不同沙丘生境下沙鞭种群空间分布与繁殖策略特征的研究尚未涉及.基于此,本研究选择流动、半流动、半固定和固定沙丘,开展沙鞭种群地上分株分布格局研究,旨在研究沙丘流动性对于沙鞭种群空间分布格局的影响,为了解极端干旱地区植物种群对环境胁迫的响应机理提供科学依据. ...
... 基于完全空间随机模型点格局分析的结果显示,沙鞭种群在不同尺度上均表现出了聚集分布(图3 ),这可能与沙鞭种群是兼性克隆植物相关,沙鞭母体可以通过根茎繁殖,在其周围产生分株,分株又产生新的分株,形成分株系统,分株系统中的植株可以继续根茎繁殖,形成下一级分株系统,所有的分株系统构成了一个大聚块,各分株系统之间可以通过克隆整合进行资源共享和风险分摊[11 ,27 -28 ] ,故而沙鞭种群在不同风沙环境下均有较大尺度的聚集分布.从不同风沙环境中聚集尺度大小来看,沙鞭种群存在从流动沙丘至固定沙丘聚集尺度先减小后增大的变化规律,这可能与沙鞭种群获得群落资源的能力,以及群落物种增加导致的种间竞争加剧相关.在流动沙丘中,沙鞭可以通过分蘖和根茎繁殖方式在适应异质性强且水分匮乏环境中具有独特优势[5 ,7 ] ,一旦在沙地定居,就能够迅速向周围裸沙扩张[11 ,18 ] ,并可通过根茎和分株形成植被斑块.同时,流动沙丘中沙鞭个体需要抱团来获取资源以确保种群存活,导致沙鞭在0—162 cm和169—170 cm尺度呈现为聚集分布格局. ...
... [11 ,18 ],并可通过根茎和分株形成植被斑块.同时,流动沙丘中沙鞭个体需要抱团来获取资源以确保种群存活,导致沙鞭在0—162 cm和169—170 cm尺度呈现为聚集分布格局. ...
... 流动沙丘、半流动沙丘和固定沙丘环境下,沙鞭种群地上分株分布格局符合泊松聚块模型(图4 A、B、D),仅在半固定沙丘,2—37 cm尺度符合正向偏离泊松聚块模型,95—168 cm尺度符合负向偏离泊松聚块模型(图4 C).这表明流动沙丘、半流动沙丘和固定沙丘环境下,沙鞭种群空间格局在大聚块中不存在聚集密度较高的小聚块,仅在半固定沙丘中,沙鞭种群聚集分布的聚块内可能存在较高密度的小聚块.这可能与不同风沙环境下沙鞭种群小尺度上空间格局形成与变化受到植物生长环境和植物自身的生物学特征影响有关.在流动沙丘、半流动沙丘中未受到其他物种的种间竞争胁迫,种群分株系统是一个整体[11 ,34 -36 ] ,所有的分株系统构成了一个大聚块,沙鞭种群以一个大的斑块整体(大聚块)共同应对极端干旱风沙环境以争取群落水分等资源[37 ] ,从而表现为符合泊松聚块模型;而在半固定沙丘中,沙鞭大聚块被沙米、虫实和沙蒿等草本物种切割,原有植被斑块内部出现了沙鞭未占据的“资源空斑”,沙鞭种群个体为了应对种间竞争和“空斑资源”争夺聚集在一起,在较小尺度上形成了小聚块实现资源共享和风险分摊,从而表现出牺牲斑块外围个体,斑块内部个体聚集确保原有植被斑块稳定的策略,种群格局表现为嵌套双聚块过程(图5 C).但在固定沙丘中,霸王和泡泡刺等物种与沙鞭在不同尺度上出现了种间竞争关系,群落内的“空斑资源”和剩余资源消失,小尺度上形成的小聚块再度被切割,从而种群格局表现为符合泊松聚块模型.这与笔者在陕西榆林调研沙鞭种群分布格局发现的现象相一致:当沙柳(Salix cheilophila )、西北针茅(Stipa krylovii )、紫菀(Aster tataricus )等物种成为群落优势种后,沙鞭植被斑块会彻底破碎,沙鞭个体呈现零星分布,与流动和半流动沙丘中沙鞭分株连线成片、斑块状分布的空间分布格局完全不同. ...
沙埋和种子大小对固沙禾草沙鞭的种子萌发与幼苗出土的影响
1
2005
... 沙鞭(Psammochloa villosa )作为干旱地区流动沙丘先锋物种,对于干旱风沙环境具有极强生态适应能力,在流动沙丘上分布的沙鞭斑块与流沙斑块相互镶嵌,已成为流动沙丘的独特景观[1 ,3 ,5 ,11 ] ,主要分布在内蒙古库布齐沙漠、浑善达克沙地西部、巴丹吉林沙漠和甘肃河西走廊沙地等地区.目前关于沙鞭的研究主要包括有性繁殖[12 -13 ] 与无性繁殖[5 ,7 ,14 ] 、不同居群形态变异[15 ] 、遗传多样性[16 -17 ] 、改造沙化景观[18 ] ,极大地丰富和发展了沙鞭的种群特征研究,但是关于不同沙丘生境下沙鞭种群空间分布与繁殖策略特征的研究尚未涉及.基于此,本研究选择流动、半流动、半固定和固定沙丘,开展沙鞭种群地上分株分布格局研究,旨在研究沙丘流动性对于沙鞭种群空间分布格局的影响,为了解极端干旱地区植物种群对环境胁迫的响应机理提供科学依据. ...
种子萌发和幼苗生长对沙丘环境的适应机制
1
2006
... 沙鞭(Psammochloa villosa )作为干旱地区流动沙丘先锋物种,对于干旱风沙环境具有极强生态适应能力,在流动沙丘上分布的沙鞭斑块与流沙斑块相互镶嵌,已成为流动沙丘的独特景观[1 ,3 ,5 ,11 ] ,主要分布在内蒙古库布齐沙漠、浑善达克沙地西部、巴丹吉林沙漠和甘肃河西走廊沙地等地区.目前关于沙鞭的研究主要包括有性繁殖[12 -13 ] 与无性繁殖[5 ,7 ,14 ] 、不同居群形态变异[15 ] 、遗传多样性[16 -17 ] 、改造沙化景观[18 ] ,极大地丰富和发展了沙鞭的种群特征研究,但是关于不同沙丘生境下沙鞭种群空间分布与繁殖策略特征的研究尚未涉及.基于此,本研究选择流动、半流动、半固定和固定沙丘,开展沙鞭种群地上分株分布格局研究,旨在研究沙丘流动性对于沙鞭种群空间分布格局的影响,为了解极端干旱地区植物种群对环境胁迫的响应机理提供科学依据. ...
毛乌素沙地根茎灌木羊柴的遗传多样性和克隆结构
1
1999
... 沙鞭(Psammochloa villosa )作为干旱地区流动沙丘先锋物种,对于干旱风沙环境具有极强生态适应能力,在流动沙丘上分布的沙鞭斑块与流沙斑块相互镶嵌,已成为流动沙丘的独特景观[1 ,3 ,5 ,11 ] ,主要分布在内蒙古库布齐沙漠、浑善达克沙地西部、巴丹吉林沙漠和甘肃河西走廊沙地等地区.目前关于沙鞭的研究主要包括有性繁殖[12 -13 ] 与无性繁殖[5 ,7 ,14 ] 、不同居群形态变异[15 ] 、遗传多样性[16 -17 ] 、改造沙化景观[18 ] ,极大地丰富和发展了沙鞭的种群特征研究,但是关于不同沙丘生境下沙鞭种群空间分布与繁殖策略特征的研究尚未涉及.基于此,本研究选择流动、半流动、半固定和固定沙丘,开展沙鞭种群地上分株分布格局研究,旨在研究沙丘流动性对于沙鞭种群空间分布格局的影响,为了解极端干旱地区植物种群对环境胁迫的响应机理提供科学依据. ...
沙生植物沙鞭不同居群形态变异研究
1
2021
... 沙鞭(Psammochloa villosa )作为干旱地区流动沙丘先锋物种,对于干旱风沙环境具有极强生态适应能力,在流动沙丘上分布的沙鞭斑块与流沙斑块相互镶嵌,已成为流动沙丘的独特景观[1 ,3 ,5 ,11 ] ,主要分布在内蒙古库布齐沙漠、浑善达克沙地西部、巴丹吉林沙漠和甘肃河西走廊沙地等地区.目前关于沙鞭的研究主要包括有性繁殖[12 -13 ] 与无性繁殖[5 ,7 ,14 ] 、不同居群形态变异[15 ] 、遗传多样性[16 -17 ] 、改造沙化景观[18 ] ,极大地丰富和发展了沙鞭的种群特征研究,但是关于不同沙丘生境下沙鞭种群空间分布与繁殖策略特征的研究尚未涉及.基于此,本研究选择流动、半流动、半固定和固定沙丘,开展沙鞭种群地上分株分布格局研究,旨在研究沙丘流动性对于沙鞭种群空间分布格局的影响,为了解极端干旱地区植物种群对环境胁迫的响应机理提供科学依据. ...
沙生牧草沙鞭的遗传多样性研究
1
2019
... 沙鞭(Psammochloa villosa )作为干旱地区流动沙丘先锋物种,对于干旱风沙环境具有极强生态适应能力,在流动沙丘上分布的沙鞭斑块与流沙斑块相互镶嵌,已成为流动沙丘的独特景观[1 ,3 ,5 ,11 ] ,主要分布在内蒙古库布齐沙漠、浑善达克沙地西部、巴丹吉林沙漠和甘肃河西走廊沙地等地区.目前关于沙鞭的研究主要包括有性繁殖[12 -13 ] 与无性繁殖[5 ,7 ,14 ] 、不同居群形态变异[15 ] 、遗传多样性[16 -17 ] 、改造沙化景观[18 ] ,极大地丰富和发展了沙鞭的种群特征研究,但是关于不同沙丘生境下沙鞭种群空间分布与繁殖策略特征的研究尚未涉及.基于此,本研究选择流动、半流动、半固定和固定沙丘,开展沙鞭种群地上分株分布格局研究,旨在研究沙丘流动性对于沙鞭种群空间分布格局的影响,为了解极端干旱地区植物种群对环境胁迫的响应机理提供科学依据. ...
荒漠植物沙鞭(Psammochloa villosa )种质资源收集及谱系遗传分化初探
1
2018
... 沙鞭(Psammochloa villosa )作为干旱地区流动沙丘先锋物种,对于干旱风沙环境具有极强生态适应能力,在流动沙丘上分布的沙鞭斑块与流沙斑块相互镶嵌,已成为流动沙丘的独特景观[1 ,3 ,5 ,11 ] ,主要分布在内蒙古库布齐沙漠、浑善达克沙地西部、巴丹吉林沙漠和甘肃河西走廊沙地等地区.目前关于沙鞭的研究主要包括有性繁殖[12 -13 ] 与无性繁殖[5 ,7 ,14 ] 、不同居群形态变异[15 ] 、遗传多样性[16 -17 ] 、改造沙化景观[18 ] ,极大地丰富和发展了沙鞭的种群特征研究,但是关于不同沙丘生境下沙鞭种群空间分布与繁殖策略特征的研究尚未涉及.基于此,本研究选择流动、半流动、半固定和固定沙丘,开展沙鞭种群地上分株分布格局研究,旨在研究沙丘流动性对于沙鞭种群空间分布格局的影响,为了解极端干旱地区植物种群对环境胁迫的响应机理提供科学依据. ...
根茎禾草沙鞭的克隆生长在毛乌素沙地斑块动态中的作用
2
2001
... 沙鞭(Psammochloa villosa )作为干旱地区流动沙丘先锋物种,对于干旱风沙环境具有极强生态适应能力,在流动沙丘上分布的沙鞭斑块与流沙斑块相互镶嵌,已成为流动沙丘的独特景观[1 ,3 ,5 ,11 ] ,主要分布在内蒙古库布齐沙漠、浑善达克沙地西部、巴丹吉林沙漠和甘肃河西走廊沙地等地区.目前关于沙鞭的研究主要包括有性繁殖[12 -13 ] 与无性繁殖[5 ,7 ,14 ] 、不同居群形态变异[15 ] 、遗传多样性[16 -17 ] 、改造沙化景观[18 ] ,极大地丰富和发展了沙鞭的种群特征研究,但是关于不同沙丘生境下沙鞭种群空间分布与繁殖策略特征的研究尚未涉及.基于此,本研究选择流动、半流动、半固定和固定沙丘,开展沙鞭种群地上分株分布格局研究,旨在研究沙丘流动性对于沙鞭种群空间分布格局的影响,为了解极端干旱地区植物种群对环境胁迫的响应机理提供科学依据. ...
... 基于完全空间随机模型点格局分析的结果显示,沙鞭种群在不同尺度上均表现出了聚集分布(图3 ),这可能与沙鞭种群是兼性克隆植物相关,沙鞭母体可以通过根茎繁殖,在其周围产生分株,分株又产生新的分株,形成分株系统,分株系统中的植株可以继续根茎繁殖,形成下一级分株系统,所有的分株系统构成了一个大聚块,各分株系统之间可以通过克隆整合进行资源共享和风险分摊[11 ,27 -28 ] ,故而沙鞭种群在不同风沙环境下均有较大尺度的聚集分布.从不同风沙环境中聚集尺度大小来看,沙鞭种群存在从流动沙丘至固定沙丘聚集尺度先减小后增大的变化规律,这可能与沙鞭种群获得群落资源的能力,以及群落物种增加导致的种间竞争加剧相关.在流动沙丘中,沙鞭可以通过分蘖和根茎繁殖方式在适应异质性强且水分匮乏环境中具有独特优势[5 ,7 ] ,一旦在沙地定居,就能够迅速向周围裸沙扩张[11 ,18 ] ,并可通过根茎和分株形成植被斑块.同时,流动沙丘中沙鞭个体需要抱团来获取资源以确保种群存活,导致沙鞭在0—162 cm和169—170 cm尺度呈现为聚集分布格局. ...
干旱、半干旱环境降水脉动对生态系统的影响
2
2011
... 研究区位于甘肃省河西走廊中部临泽县北部(39°13′19.92″N,100°39′00″E),依托中国科学院临泽内陆河流域研究站开展相关研究(图1 ).研究区位于欧亚大陆腹地,远离海洋,属典型大陆性干旱荒漠气候,辐射强,热量丰富,降水少,年均气温7.6 ℃,极端最高气温可达39.1 ℃,极端最低气温为-27.3 ℃,无霜期150—165 d,季节性冻土厚度109—123 cm.多年平均降水量117.4 mm (1965—2018年),年均潜在蒸发量2 390 mm,干旱指数达15.9,年日照时数3 045 h,太阳辐射总量高达6.1×105 J·cm-2 ·a-1 [19 ] .区域内降水存在明显季节变化,夏季(7—9月)降水占全年降水总量的65%左右,冬季降雪量占全年降水总量的3%;以小降水事件为主,≤5 mm小降水事件占全年总降水事件的56.1%,并占年降水量的46.7%(1967—2017年).天然植被总体特征组成简单、类型单一、群落不郁闭、分布稀疏.地带性植被为以超旱生半灌木和多年生草本为优势种或建群种形成的沙丘植被类型,旱生和盐生灌木为主要植物群落,梭梭(Haloxylon ammodendron )、沙拐枣(Calligonum mongolicum )、柽柳(Tamarix chinensis )、花棒(Hedysarum scoparium )、柠条(Caragana korshinskii )和泡泡刺(Nitraria sphaerocarpa )、红柳(Tamarix ramosissima )是广泛分布于临泽荒漠绿洲固定、半固定沙丘上的灌木和半灌木.一年生草本植物如虫实(Corispermum candelabrum )、雾冰藜(Bassia dasyphylla )、白茎盐生草(Halogeton arachnoideus )、碱蓬(Suaeda glauca )、沙蓬(Agriophyllum squarrosum )和画眉草(Eragrostis pilosa )等,多年生草本植物如沙鞭、沙蒿(Artemisia desertorum )等[19 ] .沙鞭为广布于流动沙丘的重要先锋物种,其种群消长对于沙丘生态系统群落结构和功能的稳定性具有不可替代的生态作用. ...
... [19 ].沙鞭为广布于流动沙丘的重要先锋物种,其种群消长对于沙丘生态系统群落结构和功能的稳定性具有不可替代的生态作用. ...
科尔沁沙地沙漠化正、逆过程的地面判别方法
1
2008
... 根据赵哈林等[20 ] 和左小安等[21 ] 对沙丘类型的划分标准,在地貌相对一致、地形平缓的巴丹吉林沙漠边缘的甘肃省张掖市临泽县平川镇,结合实地踏查和群落学调查,根据沙丘植被盖度、植被和流沙特征、沙丘表面结皮情况,以及群落优势种和伴生种等指标选取了4处围栏沙地作为实验样地(表1 ),分别为流动沙丘(A)、半流动沙丘(B)、半固定沙丘(C)、固定沙丘(D). ...
科尔沁沙地不同恢复年限退化植被的物种多样性
1
2009
... 根据赵哈林等[20 ] 和左小安等[21 ] 对沙丘类型的划分标准,在地貌相对一致、地形平缓的巴丹吉林沙漠边缘的甘肃省张掖市临泽县平川镇,结合实地踏查和群落学调查,根据沙丘植被盖度、植被和流沙特征、沙丘表面结皮情况,以及群落优势种和伴生种等指标选取了4处围栏沙地作为实验样地(表1 ),分别为流动沙丘(A)、半流动沙丘(B)、半固定沙丘(C)、固定沙丘(D). ...
2
1983
... 处于研究区域中二维空间的点构成点事件,以k(r) 函数揭示点事件空间布局的依赖程度[22 ] ,定义为: ...
... 嵌套双聚块模型(Nested double-cluster process,DC)是泊松聚块模型的多代扩展,泊松聚块过程的子代产生自己的后代个体.详细的描述参考文献[22 ],嵌套双聚块模型的k 函数表达式如下: ...
羊草+大针茅草原退化群落优势种群空间点格局分析
1
2011
... 空间分布格局研究的不断进步,为揭示植物种群的内部结构、扩散规律、更新机制以及共存问题等提供了重要手段[23 -27 ] .其中,基于各种机理的模型中,完全空间随机模型最为常见,这种模型可以揭示植物种群在不同尺度上呈现的分布格局类型,如聚集分布、均匀分布和随机分布,对于认识植被空间分布的机理具有一定的价值和作用. ...
高寒退化草地狼毒种群株丛间格局控制机理
0
2013
Early emergence enhances plant fitness:a phylogenetically controlled meta-analysis
1
2005
... 在半流动沙丘中,沙鞭种群在0—65 cm尺度范围内表现为聚集分布,65—200 cm尺度随机分布,沙鞭聚集分布的尺度显著低于流动沙丘.这可能与半流动沙丘中的水资源不足以支持沙鞭种群在大尺度上聚集分布并形成更大的植被斑块,在一定程度上限制了沙鞭种群的根茎和种子繁殖过程.原有植被斑块内个体种内合作继续占据群落资源,而较大尺度上的个体则可能通过根茎拓展和种子扩散的方式将子代输送到距离母株相对较远的地方,表现为种内个体之间的随机分布,这也与植物的向外扩张基因(遗传)导致克隆生长能够将子代扩散到更广泛的区域而避免与来自相同个体的其他配子和后代竞争的研究结果一致[25 -26 ,29 -30 ] . ...
Ecology of seed dispersal
1
1982
... 在半流动沙丘中,沙鞭种群在0—65 cm尺度范围内表现为聚集分布,65—200 cm尺度随机分布,沙鞭聚集分布的尺度显著低于流动沙丘.这可能与半流动沙丘中的水资源不足以支持沙鞭种群在大尺度上聚集分布并形成更大的植被斑块,在一定程度上限制了沙鞭种群的根茎和种子繁殖过程.原有植被斑块内个体种内合作继续占据群落资源,而较大尺度上的个体则可能通过根茎拓展和种子扩散的方式将子代输送到距离母株相对较远的地方,表现为种内个体之间的随机分布,这也与植物的向外扩张基因(遗传)导致克隆生长能够将子代扩散到更广泛的区域而避免与来自相同个体的其他配子和后代竞争的研究结果一致[25 -26 ,29 -30 ] . ...
Sibling competition in plants
2
1992
... 空间分布格局研究的不断进步,为揭示植物种群的内部结构、扩散规律、更新机制以及共存问题等提供了重要手段[23 -27 ] .其中,基于各种机理的模型中,完全空间随机模型最为常见,这种模型可以揭示植物种群在不同尺度上呈现的分布格局类型,如聚集分布、均匀分布和随机分布,对于认识植被空间分布的机理具有一定的价值和作用. ...
... 基于完全空间随机模型点格局分析的结果显示,沙鞭种群在不同尺度上均表现出了聚集分布(图3 ),这可能与沙鞭种群是兼性克隆植物相关,沙鞭母体可以通过根茎繁殖,在其周围产生分株,分株又产生新的分株,形成分株系统,分株系统中的植株可以继续根茎繁殖,形成下一级分株系统,所有的分株系统构成了一个大聚块,各分株系统之间可以通过克隆整合进行资源共享和风险分摊[11 ,27 -28 ] ,故而沙鞭种群在不同风沙环境下均有较大尺度的聚集分布.从不同风沙环境中聚集尺度大小来看,沙鞭种群存在从流动沙丘至固定沙丘聚集尺度先减小后增大的变化规律,这可能与沙鞭种群获得群落资源的能力,以及群落物种增加导致的种间竞争加剧相关.在流动沙丘中,沙鞭可以通过分蘖和根茎繁殖方式在适应异质性强且水分匮乏环境中具有独特优势[5 ,7 ] ,一旦在沙地定居,就能够迅速向周围裸沙扩张[11 ,18 ] ,并可通过根茎和分株形成植被斑块.同时,流动沙丘中沙鞭个体需要抱团来获取资源以确保种群存活,导致沙鞭在0—162 cm和169—170 cm尺度呈现为聚集分布格局. ...
Does seed dormancy benefit the mother plant by reducing sib competition?
1
1994
... 基于完全空间随机模型点格局分析的结果显示,沙鞭种群在不同尺度上均表现出了聚集分布(图3 ),这可能与沙鞭种群是兼性克隆植物相关,沙鞭母体可以通过根茎繁殖,在其周围产生分株,分株又产生新的分株,形成分株系统,分株系统中的植株可以继续根茎繁殖,形成下一级分株系统,所有的分株系统构成了一个大聚块,各分株系统之间可以通过克隆整合进行资源共享和风险分摊[11 ,27 -28 ] ,故而沙鞭种群在不同风沙环境下均有较大尺度的聚集分布.从不同风沙环境中聚集尺度大小来看,沙鞭种群存在从流动沙丘至固定沙丘聚集尺度先减小后增大的变化规律,这可能与沙鞭种群获得群落资源的能力,以及群落物种增加导致的种间竞争加剧相关.在流动沙丘中,沙鞭可以通过分蘖和根茎繁殖方式在适应异质性强且水分匮乏环境中具有独特优势[5 ,7 ] ,一旦在沙地定居,就能够迅速向周围裸沙扩张[11 ,18 ] ,并可通过根茎和分株形成植被斑块.同时,流动沙丘中沙鞭个体需要抱团来获取资源以确保种群存活,导致沙鞭在0—162 cm和169—170 cm尺度呈现为聚集分布格局. ...
Maternal and sibling factors induce dormancy in dimorphic seed pairs of Aegilops triuncialis
1
2004
... 在半流动沙丘中,沙鞭种群在0—65 cm尺度范围内表现为聚集分布,65—200 cm尺度随机分布,沙鞭聚集分布的尺度显著低于流动沙丘.这可能与半流动沙丘中的水资源不足以支持沙鞭种群在大尺度上聚集分布并形成更大的植被斑块,在一定程度上限制了沙鞭种群的根茎和种子繁殖过程.原有植被斑块内个体种内合作继续占据群落资源,而较大尺度上的个体则可能通过根茎拓展和种子扩散的方式将子代输送到距离母株相对较远的地方,表现为种内个体之间的随机分布,这也与植物的向外扩张基因(遗传)导致克隆生长能够将子代扩散到更广泛的区域而避免与来自相同个体的其他配子和后代竞争的研究结果一致[25 -26 ,29 -30 ] . ...
The population-dynamic functions of seed dispersal
1
1993
... 在半流动沙丘中,沙鞭种群在0—65 cm尺度范围内表现为聚集分布,65—200 cm尺度随机分布,沙鞭聚集分布的尺度显著低于流动沙丘.这可能与半流动沙丘中的水资源不足以支持沙鞭种群在大尺度上聚集分布并形成更大的植被斑块,在一定程度上限制了沙鞭种群的根茎和种子繁殖过程.原有植被斑块内个体种内合作继续占据群落资源,而较大尺度上的个体则可能通过根茎拓展和种子扩散的方式将子代输送到距离母株相对较远的地方,表现为种内个体之间的随机分布,这也与植物的向外扩张基因(遗传)导致克隆生长能够将子代扩散到更广泛的区域而避免与来自相同个体的其他配子和后代竞争的研究结果一致[25 -26 ,29 -30 ] . ...
退化草地阿尔泰针茅生殖株丛与非生殖株丛的空间格局
1
2012
... 半固定沙丘中,沙鞭种群在0—80 cm尺度表现为聚集分布,而在98—124 cm和130—171 cm尺度表现为均匀分布,这种均匀分布格局可能与半流动沙丘中沙鞭种内种间关系变化相一致.有限水分资源加剧了较大尺度上沙鞭植被斑块内个体的竞争关系,引起了种内自疏效应.半流动沙丘中随机分布格局转变为均匀分布格局,以牺牲沙鞭斑块外围个体的策略来确保原有植被斑块的稳定,这种策略与西北针茅等植物种群受到资源限制采取的适应策略一致[31 ] .在半固定沙丘中聚集分布的格局从半流动沙丘0—65 cm尺度增长到在0—80 cm尺度,这可能与沙鞭、沙米(Agriophyllum squarrosum )、虫实(Corispermum hyssopifolium )等一二年生草本物种展开种间竞争相关.在群落调查中也发现了比较特殊的群落属性:在流动和半流动沙丘中,沙鞭多为单种群落;在半固定沙丘中,除沙米、虫实等一二年生草本物种外,出现了沙蒿等多年生草本,而在固定沙丘中则出现了泡泡刺(Nitraria sphaerocarpa )、霸王(Zygophyllum xanthoxylon )等灌木或半灌木物种.沙鞭与多年生草本和灌木物种之间的种间竞争关系,可能会减弱沙鞭种内竞争导致的自疏现象,种内分株之间表现出合作关系争夺群落资源.因此,固定沙丘环境中,沙鞭在4—127、161—163 cm和195—200 cm尺度表现出聚集分布格局. ...
基于不同零模型的点格局分析
1
2012
... 基于完全空间随机模型的点格局分析可以揭示种群内个体的空间分布方式或配置特点,但无法直接揭示种群聚集分布或均匀分布格局的内在机制或者形成机理,泊松聚块模型和嵌套双聚块模型描述的是种群聚集分布的内在形成机制,二者紧密联系.使用泊松聚块模型检验的种群格局在一定尺度位于置信区间之上,表明在种群聚集分布的聚块内可能存在较高密度的小聚块,且可通过嵌套双聚块模型加以进一步验证[32 -33 ] . ...
Plasticity of clonal integration in the perennial herb Linaria vulgaris after damage
1
2006
... 基于完全空间随机模型的点格局分析可以揭示种群内个体的空间分布方式或配置特点,但无法直接揭示种群聚集分布或均匀分布格局的内在机制或者形成机理,泊松聚块模型和嵌套双聚块模型描述的是种群聚集分布的内在形成机制,二者紧密联系.使用泊松聚块模型检验的种群格局在一定尺度位于置信区间之上,表明在种群聚集分布的聚块内可能存在较高密度的小聚块,且可通过嵌套双聚块模型加以进一步验证[32 -33 ] . ...
The importance of population origin and environment on clonal and sexual reproduction in the alpine plant Geum reptans
1
2005
... 流动沙丘、半流动沙丘和固定沙丘环境下,沙鞭种群地上分株分布格局符合泊松聚块模型(图4 A、B、D),仅在半固定沙丘,2—37 cm尺度符合正向偏离泊松聚块模型,95—168 cm尺度符合负向偏离泊松聚块模型(图4 C).这表明流动沙丘、半流动沙丘和固定沙丘环境下,沙鞭种群空间格局在大聚块中不存在聚集密度较高的小聚块,仅在半固定沙丘中,沙鞭种群聚集分布的聚块内可能存在较高密度的小聚块.这可能与不同风沙环境下沙鞭种群小尺度上空间格局形成与变化受到植物生长环境和植物自身的生物学特征影响有关.在流动沙丘、半流动沙丘中未受到其他物种的种间竞争胁迫,种群分株系统是一个整体[11 ,34 -36 ] ,所有的分株系统构成了一个大聚块,沙鞭种群以一个大的斑块整体(大聚块)共同应对极端干旱风沙环境以争取群落水分等资源[37 ] ,从而表现为符合泊松聚块模型;而在半固定沙丘中,沙鞭大聚块被沙米、虫实和沙蒿等草本物种切割,原有植被斑块内部出现了沙鞭未占据的“资源空斑”,沙鞭种群个体为了应对种间竞争和“空斑资源”争夺聚集在一起,在较小尺度上形成了小聚块实现资源共享和风险分摊,从而表现出牺牲斑块外围个体,斑块内部个体聚集确保原有植被斑块稳定的策略,种群格局表现为嵌套双聚块过程(图5 C).但在固定沙丘中,霸王和泡泡刺等物种与沙鞭在不同尺度上出现了种间竞争关系,群落内的“空斑资源”和剩余资源消失,小尺度上形成的小聚块再度被切割,从而种群格局表现为符合泊松聚块模型.这与笔者在陕西榆林调研沙鞭种群分布格局发现的现象相一致:当沙柳(Salix cheilophila )、西北针茅(Stipa krylovii )、紫菀(Aster tataricus )等物种成为群落优势种后,沙鞭植被斑块会彻底破碎,沙鞭个体呈现零星分布,与流动和半流动沙丘中沙鞭分株连线成片、斑块状分布的空间分布格局完全不同. ...
放牧对草原群落特征及种群繁殖特性的影响
0
2005
Responses of rhizome length and ramet production to resource availability in the clonal sedge Scirpus olneyi A.Gray
1
2007
... 流动沙丘、半流动沙丘和固定沙丘环境下,沙鞭种群地上分株分布格局符合泊松聚块模型(图4 A、B、D),仅在半固定沙丘,2—37 cm尺度符合正向偏离泊松聚块模型,95—168 cm尺度符合负向偏离泊松聚块模型(图4 C).这表明流动沙丘、半流动沙丘和固定沙丘环境下,沙鞭种群空间格局在大聚块中不存在聚集密度较高的小聚块,仅在半固定沙丘中,沙鞭种群聚集分布的聚块内可能存在较高密度的小聚块.这可能与不同风沙环境下沙鞭种群小尺度上空间格局形成与变化受到植物生长环境和植物自身的生物学特征影响有关.在流动沙丘、半流动沙丘中未受到其他物种的种间竞争胁迫,种群分株系统是一个整体[11 ,34 -36 ] ,所有的分株系统构成了一个大聚块,沙鞭种群以一个大的斑块整体(大聚块)共同应对极端干旱风沙环境以争取群落水分等资源[37 ] ,从而表现为符合泊松聚块模型;而在半固定沙丘中,沙鞭大聚块被沙米、虫实和沙蒿等草本物种切割,原有植被斑块内部出现了沙鞭未占据的“资源空斑”,沙鞭种群个体为了应对种间竞争和“空斑资源”争夺聚集在一起,在较小尺度上形成了小聚块实现资源共享和风险分摊,从而表现出牺牲斑块外围个体,斑块内部个体聚集确保原有植被斑块稳定的策略,种群格局表现为嵌套双聚块过程(图5 C).但在固定沙丘中,霸王和泡泡刺等物种与沙鞭在不同尺度上出现了种间竞争关系,群落内的“空斑资源”和剩余资源消失,小尺度上形成的小聚块再度被切割,从而种群格局表现为符合泊松聚块模型.这与笔者在陕西榆林调研沙鞭种群分布格局发现的现象相一致:当沙柳(Salix cheilophila )、西北针茅(Stipa krylovii )、紫菀(Aster tataricus )等物种成为群落优势种后,沙鞭植被斑块会彻底破碎,沙鞭个体呈现零星分布,与流动和半流动沙丘中沙鞭分株连线成片、斑块状分布的空间分布格局完全不同. ...
刈割干扰下内蒙古草原两种丛生禾草繁殖策略的适应性调节
1
2010
... 流动沙丘、半流动沙丘和固定沙丘环境下,沙鞭种群地上分株分布格局符合泊松聚块模型(图4 A、B、D),仅在半固定沙丘,2—37 cm尺度符合正向偏离泊松聚块模型,95—168 cm尺度符合负向偏离泊松聚块模型(图4 C).这表明流动沙丘、半流动沙丘和固定沙丘环境下,沙鞭种群空间格局在大聚块中不存在聚集密度较高的小聚块,仅在半固定沙丘中,沙鞭种群聚集分布的聚块内可能存在较高密度的小聚块.这可能与不同风沙环境下沙鞭种群小尺度上空间格局形成与变化受到植物生长环境和植物自身的生物学特征影响有关.在流动沙丘、半流动沙丘中未受到其他物种的种间竞争胁迫,种群分株系统是一个整体[11 ,34 -36 ] ,所有的分株系统构成了一个大聚块,沙鞭种群以一个大的斑块整体(大聚块)共同应对极端干旱风沙环境以争取群落水分等资源[37 ] ,从而表现为符合泊松聚块模型;而在半固定沙丘中,沙鞭大聚块被沙米、虫实和沙蒿等草本物种切割,原有植被斑块内部出现了沙鞭未占据的“资源空斑”,沙鞭种群个体为了应对种间竞争和“空斑资源”争夺聚集在一起,在较小尺度上形成了小聚块实现资源共享和风险分摊,从而表现出牺牲斑块外围个体,斑块内部个体聚集确保原有植被斑块稳定的策略,种群格局表现为嵌套双聚块过程(图5 C).但在固定沙丘中,霸王和泡泡刺等物种与沙鞭在不同尺度上出现了种间竞争关系,群落内的“空斑资源”和剩余资源消失,小尺度上形成的小聚块再度被切割,从而种群格局表现为符合泊松聚块模型.这与笔者在陕西榆林调研沙鞭种群分布格局发现的现象相一致:当沙柳(Salix cheilophila )、西北针茅(Stipa krylovii )、紫菀(Aster tataricus )等物种成为群落优势种后,沙鞭植被斑块会彻底破碎,沙鞭个体呈现零星分布,与流动和半流动沙丘中沙鞭分株连线成片、斑块状分布的空间分布格局完全不同. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}