


0 引言
胡杨(Populus euphratica)为高大落叶乔木,雌雄异株,具有耐高温、耐盐碱、耐湿、抗干旱、抗风沙等特性,在维持干旱荒漠区生态平衡中发挥着不可代替的作用[28-29]。胡杨具有异形叶性的生物学特性,这种特性与个体发育阶段相关,表现为幼树为条形叶,随着个体发育树冠上逐渐会出现除了条形叶以外的披针形、卵形及阔卵形叶,同时这些异形叶着生的枝条长度形态也存在变化,叶面积最小的条形叶着生在长枝上,叶面积最大的阔卵形叶着生在短枝上,呈现枝叶形态随发育阶段的变化,这种表型性状的明显变化不但存在于不同发育阶段,还存在于树冠的不同冠层[30-31]。基于胡杨异形叶性的生物学特性,我们推测胡杨枝叶性状间存在异速生长关系。但目前未见胡杨叶片发育过程叶性状异速生长关系的研究报道。胡杨枝叶性状是否存在异速生长关系,包括发育阶段、冠层高度对枝叶异速生长关系的影响尚不清楚。本文以同一立地条件下不同发育阶段的胡杨雌雄株为研究对象,拟解决以下问题:①随着胡杨个体的生长发育,其枝叶性状是保持一个固定的异速生长状态还是发生了异速生长状态的变化,若是发生了异速生长状态的变化,这种变化反映了不同发育阶段的胡杨对资源利用方式的哪些转变;②异速生长状态若随冠层高度变化,反映了胡杨不同冠层高度对资源利用方式的哪些转变;③胡杨雌雄株在不同生长发育阶段及树冠的垂直空间对资源利用方式的转变有什么样的差异。研究胡杨在不同发育阶段对资源利用方式的转变及雌雄差异,对揭示胡杨随发育阶段投资策略的变化有重要意义。
1 材料与方法
1.1 研究区概况
研究区(40°32′36.90″N、81°17′56.52″E,海拔980 m)位于新疆塔里木盆地西北缘,气候炎热干燥,终年干旱少雨,年均气温10.8 ℃,年降水量40.1~98.8 mm,年日照2 556~2 991 h,无霜期180~224 d,属于典型的暖温带大陆性干旱荒漠气候。研究区域距离塔里木河河道4~5 km,地下水位1.8 m,胡杨林面积180.6 hm2,林内包含355株胡杨个体,灰杨个体330棵,树龄3~19 a,胸径DBH(diameter at breast height)2.44~40 cm。
1.2 方法
在林地内选取生长良好、枝条和叶片无明显病虫害的胡杨植株,对胸径在2 cm以上的所有胡杨进行每木调查,其中以胸径4 cm为阶距进行整化,分为8、12、16、20 cm共4个径阶(发育阶段),每径阶3棵重复,雌雄株共24棵树(表1)。各径阶以离基部2 m为间隔,将树高(冠层高度)划分为2、4、6、8、10、12 m。
表1 胡杨雌雄株样本的基本信息
Table 1
| 性别 | 径阶 | 平均胸径/cm | 平均树高/m | 平均树龄/a |
|---|---|---|---|---|
| 雌株 | 8 cm | 8.33 | 7.53 | 8.10 |
| 12 cm | 14.30 | 9.47 | 9.30 | |
| 16 cm | 17.67 | 11.27 | 10.37 | |
| 20 cm | 23.23 | 12.87 | 11.17 | |
| 雄株 | 8 cm | 9.33 | 7.97 | 8.37 |
| 12 cm | 14.37 | 10.00 | 9.70 | |
| 16 cm | 17.33 | 10.93 | 10.13 | |
| 20 cm | 24.83 | 12.70 | 11.10 |
2021年,从东南西北4个方向在各径阶不同高度采取当年生枝条及其附着的所有叶片。利用围尺测量计算树木的胸径(cm),采用直尺测量枝长(cm),游标卡尺测量枝条粗(mm)和叶柄粗(mm);同时将叶片整齐摆在刻度纸板上拍照,之后利用Image J测定叶面积(cm2),最后将叶片和枝条进行烘干、打碎,测定枝条和叶片的干重。
1.3 数据分析
每种类型枝、叶性状的算术平均值为该种类型叶片的性状值,使用SPSS对不同发育阶段叶片性状差异进行显著性分析,然后将各性状值进行常用对数转换,使其满足正态分布后进行性状间异速生长关系的分析。采用异速生长方程Y=βMα拟合胡杨枝条和叶片与养分含量之间的关系,将等式两边同时取常用对数使其转化为lgY=lgβ+αlgM。其中Y、M分别代表因变量和自变量,β为线性关系的截距,α为线性关系的斜率,α=1则代表因变量和自变量呈等速关系,α>1或α<1时则表现为异速关系。利用SMART软件中的标准化主轴回归分析(SMA)方法计算各性状指标间的异速指数和常数,通过Pitman方法计算性状回归斜率的置信区间,然后对斜率进行异质性检验,且在斜率同质时计算共同斜率。采用Origin 9.1软件作图。
2 结果与分析
2.1 枝、叶性状及生物量随发育阶段及冠层高度的变化
随着径阶的增加,雌雄株的枝、叶性状及生物量均出现显著变化(表2)。随着径阶的增加,雌雄株枝长、叶片数显著减小,枝粗、叶柄长、叶柄粗、叶面积、叶干重显著增大。从8 cm径阶到20 cm径阶,枝长雄株相对雌株减小得更多,而叶片数雌株相对雄株减小得更多;同时,叶柄长、叶柄粗、叶面积、叶干重雌株分别增加了29.32%、32.94%、32.52%、78.57%,均大于雄株增加的23.47%、18.36%、28.17%、64.28%。结果表明,随着径阶的增加,雌株的叶性状和叶生物量变化较雄株更为明显。
表2 枝、叶性状及生物量在不同冠层高度间的差异
Table 2
| 14.51±4.66A | 11.68±3.66B | 10.50±4.19B | 8.24±2.66C | 8.24±2.66C | 12.36±4.06B | 8.58±4.17C | 6.98±1.23D | ||
| 2.23±0.33B | 2.22±0.30B | 1.97±0.31C | 2.56±0.30A | 2.55±0.45B | 2.44±0.36B | 2.52±0.43B | 2.73±0.30A | ||
| 3.24±0.72C | 3.74±0.71B | 2.88±0.54D | 4.19±0.51A | 3.45±0.84C | 4.13±0.58AB | 3.96±0.72B | 4.26±0.48A | ||
| 0.85±0.09C | 0.95±0.09B | 0.85±0.12C | 1.13±0.11A | 0.98±0.22B | 1.01±0.17B | 0.96±0.13B | 1.16±0.11A | ||
| 13.16±2.82C | 14.48±2.75B | 9.93±1.93D | 17.44±2.77A | 13.88±4.28B | 13.58±1.65B | 13.58±2.82B | 17.79±2.60A | ||
| 6.99±0.95A | 7.22±0.71A | 6.38±1.04B | 5.57±0.78C | 7.44±1.14A | 7.18±0.91A | 7.34±0.97A | 6.50±0.77B | ||
| 0.19±0.12A | 0.17±0.08A | 0.11±0.05B | 0.16±0.05A | 0.27±0.23A | 0.19±0.07B | 0.21±0.10B | 0.21±0.06B | ||
| 0.14±0.05C | 0.18±0.05B | 0.14±0.09C | 0.25±0.06A | 0.14±0.06C | 0.18±0.05B | 0.17±0.06B | 0.23±0.05A | ||
n=612;小写字母为同一径阶不同树高之间的差异显著性,大写字母为不同径阶之间的差异显著性,P<0.05。
随着冠层高度的增加,雌雄株叶柄长、叶柄粗显著增加,叶片数显著减小(表2)。雌株各径阶枝长随着冠层高度的增加显著减小,但雄株仅8 cm径阶显著减小;雌雄株各径阶枝粗除雌株20 cm径阶无显著变化外,其余均呈增加趋势;雌雄株8、16 cm径阶每枝叶面积随着冠层高度的增加显著增加,20 cm径阶的每枝叶面积无显著差异;而雌雄株叶干重在8 cm径阶随着冠层高度的增加无显著变化,同时雄株20 cm径阶的枝干重也无显著变化,其余各径阶均显著增加;20 cm径阶叶柄长、叶柄粗、枝干重及叶干重从冠层高度2 m到12 m处,雌株分别增加了45%、31%、38.46%及45%,而雄株分别增加了25.34%、40.19%、15.78%及71.42%。雌雄株各径阶枝、叶性状及生物量随着冠层高度的增加差异显著。
2.2 枝、叶形态间的生长关系
2.2.1 不同发育阶段枝、叶形态间的生长关系
图1
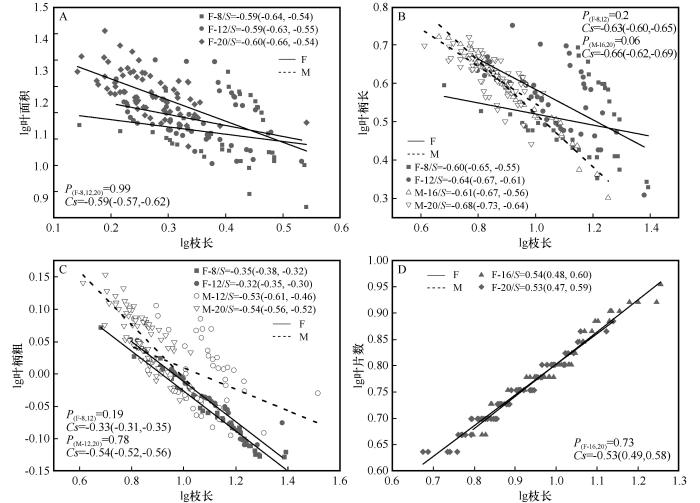
图1
不同发育阶段枝长与叶性状的生长关系
S为对应径阶斜率,括号内为95%置信区间;P为对应共同斜率的异质性;Cs代表共同斜率;F,雌株;M,雄株
Fig.1
Growth relationship between branch length with leaf traits at different developmental stages
图2
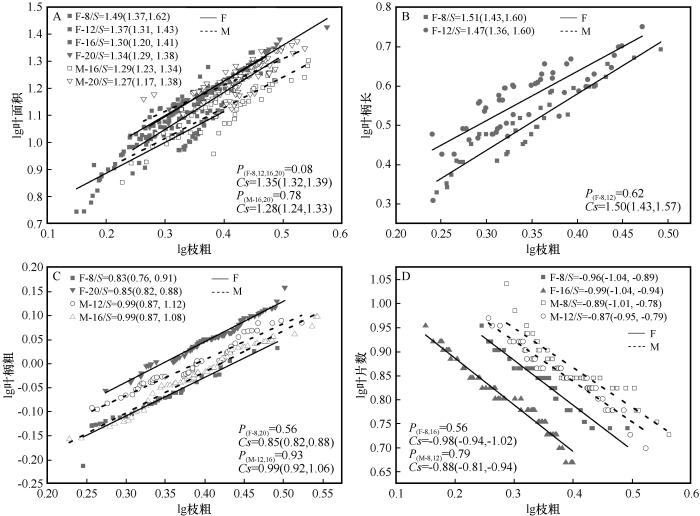
图2
不同发育阶段枝粗与叶性状的生长关系
S为对应径阶斜率,括号内为95%置信区间;P为对应共同斜率的异质性;Cs代表共同斜率;F,雌株;M,雄株
Fig.2
Growth relationship between branch thickness with leaf traits at different developmental stages
从共同斜率分析可以看出(图1、图2),雌株(8、12、20 cm径阶)枝长、枝粗分别与叶面积存在共同异速生长关系,斜率分别为-0.59(95% CI=-0.57~ -0.62)和1.35(95% CI=1.32~1.39);雄株(16、20 cm径阶)中,枝粗与叶面积存在共同斜率,为1.28(95% CI=1.24~1.33),说明雌株枝长生长减小的速度大于叶面积增加的速度,雄株大径阶(16、20 cm)的枝粗增加速度小于叶面积增加的速度。雌株(8、12 cm径阶)中枝长、枝粗与叶柄长分别存在着的共同斜率,为-0.63(95% CI=-0.60~-0.65)和1.50(95% CI=1.24~1.33);雄株16、20 cm径阶中枝长与叶柄长存在着共同斜率,为-0.66(95% CI=-0.62~ -0.69)。这表明雌株小径阶(8、12 cm)枝粗增加速度均大于叶柄长增加的速度,雌株小径阶(8、12 cm)和雄株大径阶(16、20 cm)枝长生长减小速度大于叶柄长增加的速度。雌雄株小径阶(8、12 cm)枝长、枝粗与叶柄粗在不同径阶上的共同斜率,雄株为-0.54(95% CI=-0.52~-0.56)和0.99(95% CI=0.92~1.06)相对分别大于雌株的 -0.33(95% CI=-0.31~-0.35)和0.85(95% CI=0.82~0.88),表明雌雄株小径阶枝长、枝粗与叶柄的变化速度雄株大于雌株。雌株中枝长、枝粗与叶片数在不同径阶上的共同斜率分别为0.53(95% CI=0.49~0.58)和-0.98(95% CI=-0.94~-1.02),而雄株中仅枝粗与叶片数间存在共同斜率,为-0.88(95% CI=-0.81~-0.94),说明雌雄株中枝长减小或枝粗增加的速度均大于叶片数减小的速度。整体上,不同径阶的雌雄株枝长、枝粗与叶面积、叶柄长、叶柄粗、叶片数存在共同异速生长关系。
2.2.2 同一发育阶段不同冠层高度枝、叶形态间的生长关系
同一径阶不同冠层高度,胡杨雌雄株枝、叶相关性状间存在显著相关性,枝长、枝粗与叶面积、叶柄长、叶柄粗、叶片数间存在异速生长关系。16 cm径阶雌株各高度和雄株8 m处冠层的枝长与叶片数的斜率与1.0无显著差异,存在等速生长关系。在8 cm径阶的不同高度上,雄株枝叶性状对应的异速生长斜率相对数目(n=15)大于雌株(n=9);而在12、16 cm径阶的不同高度上,雌株异速生长斜率的相对数目(n=18,n=25)均大于雄株(n=14,n=15);在20 cm径阶的不同高度上,雌株异速生长斜率的相对数目(n=24)趋于和雌株相等(n=24),说明胡杨雌雄株枝叶性状变化速度在不同冠层发育阶段存在差异,且在发育阶段的后期,雌雄株相对枝叶性状发育成熟,其生长斜率趋于稳定。
同一径阶不同冠层下枝长、枝粗与叶面积、叶柄长、叶柄粗、叶片数间存在着共同异速生长斜率。其中在8、12、16、20 cm径阶不同冠层上,雌雄株的枝长与叶性状间存在共同异速生长斜率,且斜率大部分小于1.0,而雌雄株的枝粗与叶性状间存在共同异速生长斜率大部分大于1.0,说明不同径阶各冠层枝长的减小速度大于叶性状的变化速度,枝粗的增加速度小于叶性状的变化速度。
2.3 枝、叶干重间的生长关系
2.3.1 不同发育阶段枝、叶干重间的生长关系
胡杨雌雄株在不同发育阶段下,枝干重与每枝叶干重间均存在极显著的相关关系(图3)。除了在雌株的16 cm径阶下,枝干重与每枝叶干重的斜率为1.04(95% CI=0.96~1.12),与1.0均无显著差异,存在共同斜率,显示出此时枝粗与叶面积存在等速生长关系外,其余雌雄各径阶枝干重与每枝叶干重间均存在异速生长关系,雌株枝干重与每枝叶干重间的异速指数数目相对大于雄株,8 cm径阶、20 cm径阶枝干重与每枝叶干重的斜率雌株大于雄株,即雌株枝干重与每枝叶干重变化速度大于雄株。
图3
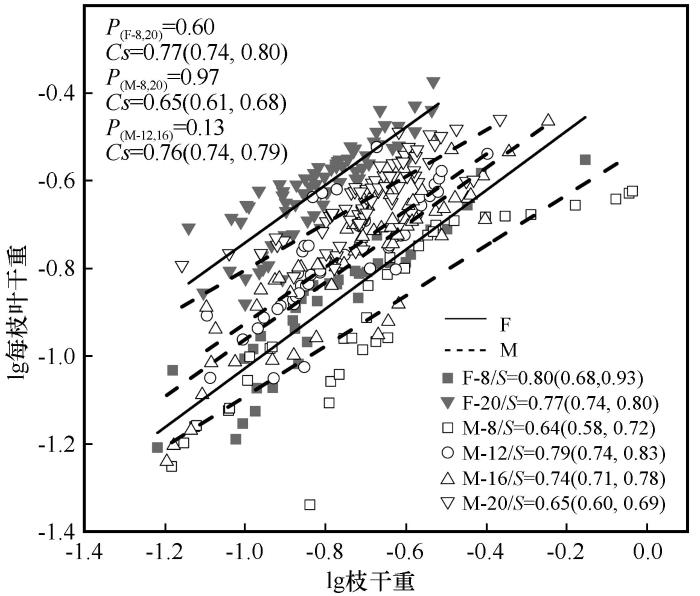
图3
不同发育阶段枝、叶干重间的生长关系
S为对应径阶斜率,括号内为95%置信区间;P为对应共同斜率的异质性;Cs代表共同斜率;F,雌株;M,雄株
Fig.3
Growth relationship of shoot and leaf biomass at different developmental stages
从图3可以看出,胡杨雌雄株枝干重与每枝叶干重在8 cm和20 cm径阶上存在共同斜率,分别为0.77(95% CI=0.74~0.80)和0.65(95% CI=0.61~0.68);同时雄株在12 cm和16 cm径阶上存在共同斜率,为0.76(95% CI=0.74~0.79)。总体上,胡杨雌雄株枝干重与每枝叶干重在不同径阶上的共同斜率均小于1.0,说明此时枝干重的增加速度大于每枝叶干重的增加速度。
2.3.2 同一发育阶段不同冠层高度枝、叶干重间的生长关系
胡杨雌雄株在同一径阶不同冠层高度下,枝干重与每枝叶干重间均存在显著的相关关系(图4)。8 cm径阶雌株枝、叶干重在2 m冠层高度处呈显著的等速生长关系,而雄株枝干重与每枝叶干重在6 m冠层高度处呈显著的等速生长关系;12 cm径阶仅雄株枝、叶干重在4、6、8 m冠层高度处存在显著的等速生长关系;16 cm径阶雌株枝、叶干重在4 m冠层高度处呈显著的等速生长关系;20 cm径阶雌株枝、叶干重在2、8 m冠层高度处呈显著的等速生长系,而雄株枝、叶干重在8、10 m高度处存在显著的等速生长关系,其余枝、叶干重不同冠层高度处均存在异速生长关系。
图4
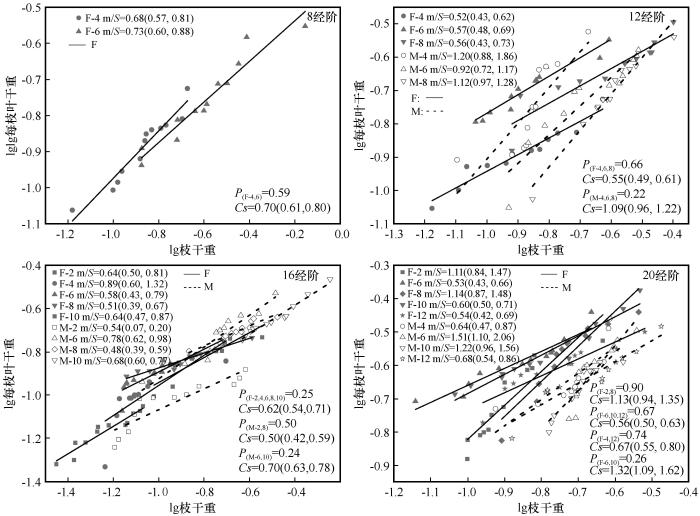
图4
不同冠层高度枝、叶干重间的异速生长关系
S为对应径阶斜率,括号内为95%置信区间;P为对应共同斜率的异质性;Cs代表共同斜率;F,雌株;M,雄株
Fig.4
Growth relationship of shoot and leaf biomass at different canopies
雌雄株枝、叶干重在同一径阶不同冠层高度上共同斜率均小于1.0,表明此时枝干重每单位的增加量大于每枝叶干重的增加量。其中在8 cm径阶雌株枝、叶干重在4、6 m冠层高度上存在共同异速生长斜率,为0.70(95% CI=0.61~0.80)。12 cm径阶雌株枝、叶干重在4、6、8 m冠层高度上均存在共同异速生长关系,而雄株枝、叶干重在4、6、8 m冠层高度上均存在共同等速生长关系。16 cm径阶雌株枝干重与每枝叶干重在不同冠层高度存在共同异速生长关系,而雄株枝干重与每枝叶干重在除4 m的其余冠层高度上存在共同异速生长关系。20 cm径阶雌株枝、叶干重在2、8 m冠层高度上存在共同等速生长关系,而在6、10、12冠层高度上均存在共同异速生长关系。雄株枝、叶干重在4、12 m及6、10 m冠层高度上均存在共同异速生长关系。
2.4 枝干重与叶性状间的关系
2.4.1 不同径阶枝干重与叶性状间的异速生长关系
不同径阶下雌雄株枝干重与叶性状间存在显著的相关关系,且雌雄株枝干重与叶性状间存在显著的异速生长关系(图5),斜率均小于1.0,表明在不同径阶下,雌雄株枝干重的增加速度大于叶面积、叶柄长、叶柄粗的增加速度和每枝叶片数的减小速度。
图5
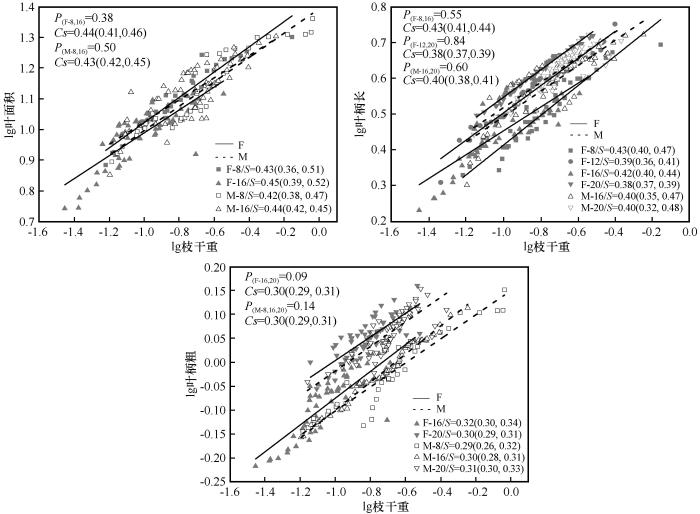
图5
不同径阶雌雄株枝干重与叶性状间的异速生长关系
S为对应径阶斜率,括号内为95%置信区间;P为对应共同斜率的异质性;Cs代表共同斜率;F,雌株;M,雄株
Fig.5
Growth relationship between shoot biomass and leaf traits at different developmental stages
雌雄株枝干重与叶面积在8、16 cm径阶上均存在共同异速生长斜率,分别为0.44(95% CI=0.41~0.46)和0.43(95% CI=0.42~0.45)。雌株枝干重与叶柄长在8、16 cm径阶和12、20 cm径阶存在共同异速生长斜率,而雄株仅在16、20 cm径阶上存在共同异速生长斜率。雌株枝干重与叶柄粗在16、20 cm径阶上存在共同斜率,为0.30(95% CI=0.27~0.33),而雄株枝干重与叶柄粗在8、16、20 cm径阶上存在共同异速生长斜率,为0.30(95% CI=0.29~0.31)。整体上,雌雄株枝干重与叶性状在不同径阶上的共同斜率均小于1.0,说明雌雄株枝干重的增加速度大于叶面积、叶柄长、叶柄粗的增加速度。
2.4.2 同一发育阶段不同冠层高长枝干重与叶性状间的生长关系
同一径阶不同冠层高度雌雄株枝干重与叶性状间存在显著的相关关系,其中在20 cm径阶的6 m和10 m冠层高度下,雄株枝干重与叶面积斜率分别为0.79(95% CI=0.56~1.10)和1.17(95% CI=0.98~1.41),与1.0无显著差异,表明在此高度冠层下,雄株枝干重的增加速度与叶面积的增加速度近似相等;6 m冠层高度,雄株枝干重与叶柄长的斜率为0.77(95% CI=0.56~1.06,与1.0无显著差异,表明此时雄株枝干重与叶柄长的增加速度近似相等。其余雌雄株各径阶不同冠层高度枝干重与叶性状间均存在异速生长关系。
8 cm径阶雌雄株枝干重与叶面积、叶柄长、叶柄粗在较低冠层高度(2、4、6 m)存在共同异速生长斜率;12 cm径阶雌株枝干重与叶面积在4、6、8 m冠层高度存在共同异速生长关系,而雄株在2、4 m和6、8 m冠层高度存在共同异速生长关系;雌株枝干重与叶柄长在2、4 m冠层高度存在共同异速生长关系,而雄株在6、8 m冠层高度处存在;在其他2个冠层高度下,雌雄株枝干重与叶面积、叶柄长、叶柄粗和每枝叶片数均存在共同异速生长斜率。
16 cm径阶雌株枝干重与叶柄长、叶柄粗和叶片数在3个冠层高度存在共同异速生长斜率,而雄株枝干重与叶面积、叶柄长和叶片数在3个冠层高度存在共同异速生长斜率。在其他2个冠层高度下,雌雄株枝干重与叶面积、叶柄长、叶柄粗和叶片数间存在共同异速生长斜率。20 cm径阶雌株枝干重与叶面积、叶柄长、叶柄粗、叶片数在3个冠层高度存在共同异速生长斜率,而雄株枝干重与叶面积、叶柄长、叶片数在3个冠层高度存在共同异速生长斜率,枝干重与叶柄粗在2个冠层高度存在共同异速生长斜率。总的来说,在同一径阶不同冠层高度下,雌雄株枝干重与叶面积、叶柄长、叶柄粗和叶片数间的共同异速生长斜率均小于1.0,说明雌雄株枝干重的增加速度大于叶面积、叶柄长、叶柄粗的增加速度和叶片数的减小速度。
3 讨论
3.1 胡杨雌雄株不同发育阶段及冠层高度枝叶性状及生物量变化特征
植物不同构件间的生长存在差异是普遍现象[9,32],也是植物体在特定生境下所采取的生态对策方式,反映了植物体各性状间的相关程度[33]。当然叶片面积增大的同时,叶柄长也会发生相应变化以提供与叶片相适应的支持能力和疏导能力,同时叶片面积的增加也需要更长的叶柄来减少个体内部对光的相互遮挡[34-36]。前人对胡杨的研究发现,随着径阶的增加,胡杨出现了由条形叶向阔卵形叶的转变,叶面积沿树冠基部向顶部方向逐渐增大[37-38]。本研究表明,随着径阶的增加,雌雄株枝长、叶片数显著减小,枝粗、叶柄长、叶柄粗、叶面积、叶干重显著增大,且叶性状和叶生物量变化雌株较雄株更为明显。雌雄株叶柄长、叶柄粗显著增加,叶片数显著减小。雌株各径阶枝长随着冠层高度的增加显著减小。较大的叶子为了更多地曝露于阳光下和更有效率地将水分运输到叶肉细胞,将会投资更多的生物量用于微管组织和厚壁组织的建设。胡杨的成熟伴随着光合能力的增强,叶面积的增加扩大了光合面积,这时投资更多的生物量输送给叶柄长、叶柄粗等支撑结构,同时枝长减小,减少了水分运输路程,叶干重的增大是多个叶性状协同变化使得光合能力增强的结果。
3.2 不同发育阶段及冠层高度枝、叶资源利用策略
由于水力和机械两方面的原因,枝条相关性状与其上面的附属物在植物生长的过程中保持一定的协调性。例如粗的枝条上有着更大的叶片存在[39-40]。本研究表明随着径阶或者同一径阶冠层高度的增加,胡杨雌雄株枝条长和枝条粗均与叶柄长和叶柄粗存在异速生长关系,可能原因是植物通过增加叶柄的长度,来减少叶片之间的相互遮荫,有效地提高叶片的光拦截效率,为促进光合作用奠定了基础,同时叶柄粗的增加也促进了枝条向叶片运输水分和养分的效率[41]。枝条粗与叶面积间存在异速生长关系,小枝直径与单叶面积之间为异速生长关系[42-43],小枝的横截面积与叶面积之间存在着异速生长关系[9,44]。本研究结果显示胡杨雌雄株枝条长度和粗均与叶面积存在异速生长关系。随着胡杨径阶的增加,植株需要光合作用制造更多的有机物来满足自身生存的需要,因此枝、叶相关性状发生快速的变化来有效地提高植物的光合作用,例如胡杨枝条变短、加粗加快了养分和水分运输到叶片,叶面积增加了捕获光能的有效面积,这些为植物在单位时间内积累更多的有机物质奠定了基础。
枝条生物量的分配不仅体现在枝叶大小的权衡上,也体现在枝叶水平生物量的分配上[9,40,45],研究枝条生物量的分配有利于揭示不同植物对环境的适应策略。本研究中在不同径阶,枝干重与每枝叶干重间雌株相对占优势的生长斜率数目(n=3)大于雄株(n=1),枝干重与叶性状间雌株相对占优势的生长斜率数目(n=9)大于雄株(n=6),而在同一径阶不同冠层高度,枝干重与每枝叶干重间雌株相对占优势的生长斜率数目(n=8)小于雄株(n=10),枝干重与叶性状间雌株相对占优势的生长斜率数目(n=35)小于雄株(n=37),这表明在不同径阶上雌株枝干重与每枝叶干重、叶性状间生物量的转变速度较快,而在同一径阶不同冠层高度上雄株枝干重与每枝叶干重、叶性状间生物量转变较快,其雌雄株异速生长斜率数目差异不显著,然而雌雄株枝干重与每枝叶干重、叶性状在不同径阶上或同一径阶不同冠层高度上均存在普遍的异速生长关系。具有较大叶和枝的物种叶生物量分配比例更高。产生异速生长的原因是植物体的输导组织网络的限制,植物在漫长的进化过程中形成不同等级的输导网络组织,营养选择的压力必然使其向最优化的方向进化,使得新陈代谢效率和与外界环境的交换面积最大化并使营养物质运输距离和时间最小化[24,46]。本研究中雌雄株枝干重与每枝叶干重、叶性状间存在着少数的等速生长关系,这表明随着发育阶段或冠层高度的变化,胡杨雌雄株生长存在着等速生长与异速生长间的转变,为胡杨利用环境资源更好地合成能量物质奠定基础。
3.3 胡杨雌雄株枝叶资源利用的差异
本研究发现在不同径阶上,胡杨雄株枝叶性状间占优势的生长斜率数目(n=18)相对大于雌株(n=14),这表明在不同发育阶段胡杨雄株枝、叶性状间的转变速度较快。而在同一径阶不同冠层高度,胡杨雌株枝叶性状间占优势生长斜率的数目(n=76)相对大于雄株(n=68),表明胡杨在同一发育阶段不同冠层高度上雌株枝、叶性状间的转变速度较快。胡杨雌雄株枝、叶相关性状在不同发育阶段或不同冠层高度下出现生长差异的原因,可能是胡杨为了对枝、叶不同器官间物质和能量进行协调分配,以最大化地利用环境资源进而达到雌雄株最适生长状态。
4 结论
随径阶的增加,雌雄株叶片数呈减小趋势,枝条粗、叶柄长、叶柄粗、叶面积、叶片数、枝干重和每枝叶干重呈增加趋势;不同径阶胡杨雌雄株枝、叶形态间存在异速生长关系,随着径阶的变化,雄株枝叶形态间转变速度较快,而雌株枝叶间的生物量转化效率更高;不同冠层高度胡杨雌雄株枝、叶形态间存在异速生长关系,随着冠层高度的变化,雌株枝叶形态间转变速度较快,而雄株生物量转化效率更高。雌雄株倾向于在高的冠层和成熟的发育阶段通过更短更粗的当年生枝条来为具有较大的叶柄长、叶柄粗、叶面积、较少的叶片数高效地提供水分和矿物质元素提供可能。通过胡杨枝叶性状及生长关系的研究,有助于进一步了解胡杨环境适应性的性别间的差异,为未来在种植胡杨中雌雄配置等问题提供理论依据。
参考文献
Identification of biomechanical actors involved in stem shape variability between pricot tree varieties
[J].
Optimal leaf size in relation to environment
[J].
Sizes and shapes of liane leaves
[J].
Corners rules revisited: ontogenetic and interspecific patterns in leaf-stem allometry
[J].
Let the concept of trait be functional!
[J].
The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient: an invariant allometric scaling relationship
[J].
低海拔常绿杜鹃小枝繁殖分配与异速生长关系研究
[J].
Branching pattern and leaf crown architecture of some tree and shrubs in northeast Mexico
[J].
Allometric models for leaf area and leaf mass predictions across different growing seasons of elm tree (Ulmus japonica)
[J].
武夷山不同海拔黄山松枝叶大小关系
[J].
长柄双花木叶性状异速生长关系随发育阶段和海拔梯度的变化
[J].
Adaptive and inevitable morphological plasticity of three herbaceous species in a multi-species community: field experiment with manipulated nutrients and light
[J].
Leaf traits show different relationships with shade tolerance in moist versus dry tropical forests
[J].
Allocation,plasticity and allometry in plants
[J].
Global allocation rules for patterns of biomass partitioning in seed plants
[J].
The interspecific size-density relationship among crowded plant stands and its implications for the -3/2 power rule of self-thinning
[J].
The scaling of leaf area and mass: the cost of light interception increases with leaf size
[J].
“Diminishing returns” in the scaling of functional leaf traits across and within species groups
[J].
Morphological,structural and physiological differences in heteromorphic leaves of Euphrates poplar during development stages and at crown scales
[J].
Leaf size and inflorescence size may be allometrically related traits
[J].
Allometric relationships between lamina area,lamina mass and petiole mass of 93 temperate woody species vary with leaf habit,leaf form and altitude
[J].
Effects of leaf blade narrowness and petiole length on the light capture efficiency of a shoot
[J].
The durian theory or the origin of the modern tree
[J].
The leaf size-twig size spectrum and its relationship to other important spectra of variation among species
[J].
Implications of opposite phyllotaxis for light interception efficiency of Mediterranean woody plants
[J].
Corner's rules in eastern deciduous trees: allometry and its implications for the adaptive architecture of trees
[J].
Evidence that temperate east North American evergreen woody plants follow Corner's rules
[J].
Plant ecological strategies: some leading dimensions of variation between species
[J].
A general model for the origin of allometric scaling laws in biology
[J].

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}