Global covariation of carbon turnover times with climate in terrestrial ecosystems
1
2014
... 土壤碳氮库动态及其机制是全球陆地生态系统研究的核心问题[1].全球土壤碳氮更趋向于分布在热带湿润区或者高寒永久冻土区[2],但占全球陆地面积约50%的干旱半干旱地区的土壤碳氮储量及其对全球气候影响的潜力仍不容忽视[3].近年来,干旱半干旱区土壤碳氮含量变化问题引起了部分国内外学者的关注.杨玉海等[4]研究了塔里木河流域土地利用变化对有机碳储量的影响;Dong等[5]以禁牧条件下的灌木荒漠土壤为研究对象,揭示了禁牧对土壤碳氮储量的影响;Wang等[6]研究了中国西北河西走廊荒漠草原的有机碳空间分布格局及其影响因素.干旱半干旱区土壤碳氮含量特征具有空间异质性,且造成这种差异的主导因素也不同. ...
Global soil carbon: understanding and managing the largest terrestrial carbon pool
1
2014
... 土壤碳氮库动态及其机制是全球陆地生态系统研究的核心问题[1].全球土壤碳氮更趋向于分布在热带湿润区或者高寒永久冻土区[2],但占全球陆地面积约50%的干旱半干旱地区的土壤碳氮储量及其对全球气候影响的潜力仍不容忽视[3].近年来,干旱半干旱区土壤碳氮含量变化问题引起了部分国内外学者的关注.杨玉海等[4]研究了塔里木河流域土地利用变化对有机碳储量的影响;Dong等[5]以禁牧条件下的灌木荒漠土壤为研究对象,揭示了禁牧对土壤碳氮储量的影响;Wang等[6]研究了中国西北河西走廊荒漠草原的有机碳空间分布格局及其影响因素.干旱半干旱区土壤碳氮含量特征具有空间异质性,且造成这种差异的主导因素也不同. ...
Hidden carbon sink beneath desert
1
2015
... 土壤碳氮库动态及其机制是全球陆地生态系统研究的核心问题[1].全球土壤碳氮更趋向于分布在热带湿润区或者高寒永久冻土区[2],但占全球陆地面积约50%的干旱半干旱地区的土壤碳氮储量及其对全球气候影响的潜力仍不容忽视[3].近年来,干旱半干旱区土壤碳氮含量变化问题引起了部分国内外学者的关注.杨玉海等[4]研究了塔里木河流域土地利用变化对有机碳储量的影响;Dong等[5]以禁牧条件下的灌木荒漠土壤为研究对象,揭示了禁牧对土壤碳氮储量的影响;Wang等[6]研究了中国西北河西走廊荒漠草原的有机碳空间分布格局及其影响因素.干旱半干旱区土壤碳氮含量特征具有空间异质性,且造成这种差异的主导因素也不同. ...
塔里木河干流土地利用/覆被变化对土壤有机碳储量的影响
1
2016
... 土壤碳氮库动态及其机制是全球陆地生态系统研究的核心问题[1].全球土壤碳氮更趋向于分布在热带湿润区或者高寒永久冻土区[2],但占全球陆地面积约50%的干旱半干旱地区的土壤碳氮储量及其对全球气候影响的潜力仍不容忽视[3].近年来,干旱半干旱区土壤碳氮含量变化问题引起了部分国内外学者的关注.杨玉海等[4]研究了塔里木河流域土地利用变化对有机碳储量的影响;Dong等[5]以禁牧条件下的灌木荒漠土壤为研究对象,揭示了禁牧对土壤碳氮储量的影响;Wang等[6]研究了中国西北河西走廊荒漠草原的有机碳空间分布格局及其影响因素.干旱半干旱区土壤碳氮含量特征具有空间异质性,且造成这种差异的主导因素也不同. ...
Community structure and carbon and nitrogen storage of sagebrush desert under grazing exclusion in Northwest China
1
2020
... 土壤碳氮库动态及其机制是全球陆地生态系统研究的核心问题[1].全球土壤碳氮更趋向于分布在热带湿润区或者高寒永久冻土区[2],但占全球陆地面积约50%的干旱半干旱地区的土壤碳氮储量及其对全球气候影响的潜力仍不容忽视[3].近年来,干旱半干旱区土壤碳氮含量变化问题引起了部分国内外学者的关注.杨玉海等[4]研究了塔里木河流域土地利用变化对有机碳储量的影响;Dong等[5]以禁牧条件下的灌木荒漠土壤为研究对象,揭示了禁牧对土壤碳氮储量的影响;Wang等[6]研究了中国西北河西走廊荒漠草原的有机碳空间分布格局及其影响因素.干旱半干旱区土壤碳氮含量特征具有空间异质性,且造成这种差异的主导因素也不同. ...
Spatial distribution of soil organic carbon and its influencing factors in desert grasslands of the Hexi Corridor,Northwest China
1
2014
... 土壤碳氮库动态及其机制是全球陆地生态系统研究的核心问题[1].全球土壤碳氮更趋向于分布在热带湿润区或者高寒永久冻土区[2],但占全球陆地面积约50%的干旱半干旱地区的土壤碳氮储量及其对全球气候影响的潜力仍不容忽视[3].近年来,干旱半干旱区土壤碳氮含量变化问题引起了部分国内外学者的关注.杨玉海等[4]研究了塔里木河流域土地利用变化对有机碳储量的影响;Dong等[5]以禁牧条件下的灌木荒漠土壤为研究对象,揭示了禁牧对土壤碳氮储量的影响;Wang等[6]研究了中国西北河西走廊荒漠草原的有机碳空间分布格局及其影响因素.干旱半干旱区土壤碳氮含量特征具有空间异质性,且造成这种差异的主导因素也不同. ...
冀北山地3种森林植被恢复类型对土壤质量的影响
1
2018
... 植被与土壤是陆地生态系统中相互制约、相互影响的统一体.植被从土壤中不断地吸收养分,并以枯落物的形式返还土壤,在不断往复的过程中对土壤的发育、养分循环等产生重要影响[7].在不同植被类型下,植被的特征、组成等差异引起土壤的环境、结构等不同,进而影响土壤养分的输入与消耗[8-9].生长在极端干旱环境下的荒漠河岸林为内陆河下游绿洲的主要组成部分[10],林区内生长的大量乔、灌、草等植被对于极端干旱区防风固沙、遏制沙漠化十分重要[11].目前关于干旱区荒漠河岸林研究多围绕植被与地下水关系[12]及土壤盐分、养分空间异质性[13-14]等方面,对于荒漠河岸林土壤碳氮分布特征尚未清晰,在一定程度上制约了人们对干旱区土壤碳氮循环过程及其机制的全面认识.基于此,本研究选取了黑河下游河岸林区3种典型植物群落下的土壤为研究对象,揭示土壤总碳、总氮沿土壤深度的变化特征,并分析其与土壤理化因子的关系,为荒漠河岸带生物地球化学循环过程以及土壤碳氮空间格局研究提供参考. ...
植被类型对黄土丘陵区土壤碳氮磷化学计量特征的季节变异
1
2021
... 植被与土壤是陆地生态系统中相互制约、相互影响的统一体.植被从土壤中不断地吸收养分,并以枯落物的形式返还土壤,在不断往复的过程中对土壤的发育、养分循环等产生重要影响[7].在不同植被类型下,植被的特征、组成等差异引起土壤的环境、结构等不同,进而影响土壤养分的输入与消耗[8-9].生长在极端干旱环境下的荒漠河岸林为内陆河下游绿洲的主要组成部分[10],林区内生长的大量乔、灌、草等植被对于极端干旱区防风固沙、遏制沙漠化十分重要[11].目前关于干旱区荒漠河岸林研究多围绕植被与地下水关系[12]及土壤盐分、养分空间异质性[13-14]等方面,对于荒漠河岸林土壤碳氮分布特征尚未清晰,在一定程度上制约了人们对干旱区土壤碳氮循环过程及其机制的全面认识.基于此,本研究选取了黑河下游河岸林区3种典型植物群落下的土壤为研究对象,揭示土壤总碳、总氮沿土壤深度的变化特征,并分析其与土壤理化因子的关系,为荒漠河岸带生物地球化学循环过程以及土壤碳氮空间格局研究提供参考. ...
陕北黄土区不同植被恢复模式植被与土壤耦合关系研究
1
2021
... 植被与土壤是陆地生态系统中相互制约、相互影响的统一体.植被从土壤中不断地吸收养分,并以枯落物的形式返还土壤,在不断往复的过程中对土壤的发育、养分循环等产生重要影响[7].在不同植被类型下,植被的特征、组成等差异引起土壤的环境、结构等不同,进而影响土壤养分的输入与消耗[8-9].生长在极端干旱环境下的荒漠河岸林为内陆河下游绿洲的主要组成部分[10],林区内生长的大量乔、灌、草等植被对于极端干旱区防风固沙、遏制沙漠化十分重要[11].目前关于干旱区荒漠河岸林研究多围绕植被与地下水关系[12]及土壤盐分、养分空间异质性[13-14]等方面,对于荒漠河岸林土壤碳氮分布特征尚未清晰,在一定程度上制约了人们对干旱区土壤碳氮循环过程及其机制的全面认识.基于此,本研究选取了黑河下游河岸林区3种典型植物群落下的土壤为研究对象,揭示土壤总碳、总氮沿土壤深度的变化特征,并分析其与土壤理化因子的关系,为荒漠河岸带生物地球化学循环过程以及土壤碳氮空间格局研究提供参考. ...
塔里木荒漠河岸林植物群落演替下的土壤理化性质研究
1
2010
... 植被与土壤是陆地生态系统中相互制约、相互影响的统一体.植被从土壤中不断地吸收养分,并以枯落物的形式返还土壤,在不断往复的过程中对土壤的发育、养分循环等产生重要影响[7].在不同植被类型下,植被的特征、组成等差异引起土壤的环境、结构等不同,进而影响土壤养分的输入与消耗[8-9].生长在极端干旱环境下的荒漠河岸林为内陆河下游绿洲的主要组成部分[10],林区内生长的大量乔、灌、草等植被对于极端干旱区防风固沙、遏制沙漠化十分重要[11].目前关于干旱区荒漠河岸林研究多围绕植被与地下水关系[12]及土壤盐分、养分空间异质性[13-14]等方面,对于荒漠河岸林土壤碳氮分布特征尚未清晰,在一定程度上制约了人们对干旱区土壤碳氮循环过程及其机制的全面认识.基于此,本研究选取了黑河下游河岸林区3种典型植物群落下的土壤为研究对象,揭示土壤总碳、总氮沿土壤深度的变化特征,并分析其与土壤理化因子的关系,为荒漠河岸带生物地球化学循环过程以及土壤碳氮空间格局研究提供参考. ...
Groundwater depth and soil properties are associated with variation in vegetation of a desert riparian ecosystem in an arid area of China
2
2018
... 植被与土壤是陆地生态系统中相互制约、相互影响的统一体.植被从土壤中不断地吸收养分,并以枯落物的形式返还土壤,在不断往复的过程中对土壤的发育、养分循环等产生重要影响[7].在不同植被类型下,植被的特征、组成等差异引起土壤的环境、结构等不同,进而影响土壤养分的输入与消耗[8-9].生长在极端干旱环境下的荒漠河岸林为内陆河下游绿洲的主要组成部分[10],林区内生长的大量乔、灌、草等植被对于极端干旱区防风固沙、遏制沙漠化十分重要[11].目前关于干旱区荒漠河岸林研究多围绕植被与地下水关系[12]及土壤盐分、养分空间异质性[13-14]等方面,对于荒漠河岸林土壤碳氮分布特征尚未清晰,在一定程度上制约了人们对干旱区土壤碳氮循环过程及其机制的全面认识.基于此,本研究选取了黑河下游河岸林区3种典型植物群落下的土壤为研究对象,揭示土壤总碳、总氮沿土壤深度的变化特征,并分析其与土壤理化因子的关系,为荒漠河岸带生物地球化学循环过程以及土壤碳氮空间格局研究提供参考. ...
... 本研究发现黑河下游荒漠河岸林土壤碳氮平均含量分别为17.94、0.59 mg·g-1,这与Zhang等[11]的研究结果接近,低于全国平均水平(19.33、1.61 mg·g-1)[18-19].与干旱区的部分沙地相比[20-21],黑河下游荒漠河岸带的土壤具有较高的碳氮含量,这主要与荒漠河岸带的植被有关.荒漠河岸林生长于河道两侧,由河水补给的地下水为河岸带内植物生长发育提供了重要的水分保障[22],在黑河下游形成了以乔、灌、草为主的荒漠植被带[23],成为土壤碳氮输入的主要来源,相较于其他荒漠生态系统,荒漠河岸林土壤可能形成干旱区的大型“沃岛”. ...
黑河下游绿洲NDVI对地下水位变化的响应研究
1
2013
... 植被与土壤是陆地生态系统中相互制约、相互影响的统一体.植被从土壤中不断地吸收养分,并以枯落物的形式返还土壤,在不断往复的过程中对土壤的发育、养分循环等产生重要影响[7].在不同植被类型下,植被的特征、组成等差异引起土壤的环境、结构等不同,进而影响土壤养分的输入与消耗[8-9].生长在极端干旱环境下的荒漠河岸林为内陆河下游绿洲的主要组成部分[10],林区内生长的大量乔、灌、草等植被对于极端干旱区防风固沙、遏制沙漠化十分重要[11].目前关于干旱区荒漠河岸林研究多围绕植被与地下水关系[12]及土壤盐分、养分空间异质性[13-14]等方面,对于荒漠河岸林土壤碳氮分布特征尚未清晰,在一定程度上制约了人们对干旱区土壤碳氮循环过程及其机制的全面认识.基于此,本研究选取了黑河下游河岸林区3种典型植物群落下的土壤为研究对象,揭示土壤总碳、总氮沿土壤深度的变化特征,并分析其与土壤理化因子的关系,为荒漠河岸带生物地球化学循环过程以及土壤碳氮空间格局研究提供参考. ...
黑河下游河流沿岸土壤养分和盐分的研究
1
2012
... 植被与土壤是陆地生态系统中相互制约、相互影响的统一体.植被从土壤中不断地吸收养分,并以枯落物的形式返还土壤,在不断往复的过程中对土壤的发育、养分循环等产生重要影响[7].在不同植被类型下,植被的特征、组成等差异引起土壤的环境、结构等不同,进而影响土壤养分的输入与消耗[8-9].生长在极端干旱环境下的荒漠河岸林为内陆河下游绿洲的主要组成部分[10],林区内生长的大量乔、灌、草等植被对于极端干旱区防风固沙、遏制沙漠化十分重要[11].目前关于干旱区荒漠河岸林研究多围绕植被与地下水关系[12]及土壤盐分、养分空间异质性[13-14]等方面,对于荒漠河岸林土壤碳氮分布特征尚未清晰,在一定程度上制约了人们对干旱区土壤碳氮循环过程及其机制的全面认识.基于此,本研究选取了黑河下游河岸林区3种典型植物群落下的土壤为研究对象,揭示土壤总碳、总氮沿土壤深度的变化特征,并分析其与土壤理化因子的关系,为荒漠河岸带生物地球化学循环过程以及土壤碳氮空间格局研究提供参考. ...
黑河下游土壤盐分分布特征
1
2017
... 植被与土壤是陆地生态系统中相互制约、相互影响的统一体.植被从土壤中不断地吸收养分,并以枯落物的形式返还土壤,在不断往复的过程中对土壤的发育、养分循环等产生重要影响[7].在不同植被类型下,植被的特征、组成等差异引起土壤的环境、结构等不同,进而影响土壤养分的输入与消耗[8-9].生长在极端干旱环境下的荒漠河岸林为内陆河下游绿洲的主要组成部分[10],林区内生长的大量乔、灌、草等植被对于极端干旱区防风固沙、遏制沙漠化十分重要[11].目前关于干旱区荒漠河岸林研究多围绕植被与地下水关系[12]及土壤盐分、养分空间异质性[13-14]等方面,对于荒漠河岸林土壤碳氮分布特征尚未清晰,在一定程度上制约了人们对干旱区土壤碳氮循环过程及其机制的全面认识.基于此,本研究选取了黑河下游河岸林区3种典型植物群落下的土壤为研究对象,揭示土壤总碳、总氮沿土壤深度的变化特征,并分析其与土壤理化因子的关系,为荒漠河岸带生物地球化学循环过程以及土壤碳氮空间格局研究提供参考. ...
额济纳绿洲沙漠化对柽柳群落影响的研究
1
2004
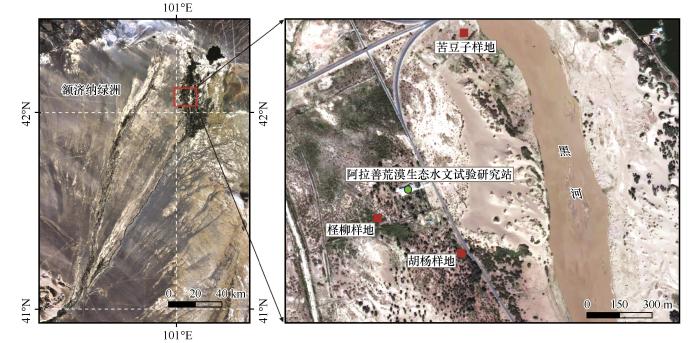
... 黑河下游荒漠河岸林内的乔、灌、草分别以胡杨、多枝柽柳以及苦豆子为优势种[15].多枝柽柳占研究区林地面积的72.3%[16],胡杨更是河岸林内唯一的乔木树种,是维持研究区生态系统稳定的关键物种.2021年9月沿垂直黑河下游东河河道方向选择以胡杨、多枝柽柳(简称柽柳)以及苦豆子为优势植被的3个样地(图1),每个样地内设置3个重复采样点.采用剖面挖掘采样法对土壤进行分层采样,3个样地的采样深度以达到地下水面为止,第1、2层土壤分别按0~10、10~40 cm间隔采样,其余层采样间隔为40 cm.胡杨、柽柳样地的采样深度为280 cm,距离河道50 m的苦豆子样地附近地下水位较浅,采样深度仅达到160 cm.采样时,在每一个采样点将每一层的土壤取出后装入已标记的自封袋中,分为两份,一份带回实验室自然风干,过筛(2 mm)备用,另一份4 ℃下保存备用.另外,每一土层取2个环刀样,用于测定土壤含水量及容重.同时记录采样点植被状况、经纬度等基本信息.本次采样共采集63份土壤样品,采样点基本情况信息见表1. ...
黑河下游绿洲植被优势种生物量空间分布及蒸腾耗水估算
1
2014
... 黑河下游荒漠河岸林内的乔、灌、草分别以胡杨、多枝柽柳以及苦豆子为优势种[15].多枝柽柳占研究区林地面积的72.3%[16],胡杨更是河岸林内唯一的乔木树种,是维持研究区生态系统稳定的关键物种.2021年9月沿垂直黑河下游东河河道方向选择以胡杨、多枝柽柳(简称柽柳)以及苦豆子为优势植被的3个样地(图1),每个样地内设置3个重复采样点.采用剖面挖掘采样法对土壤进行分层采样,3个样地的采样深度以达到地下水面为止,第1、2层土壤分别按0~10、10~40 cm间隔采样,其余层采样间隔为40 cm.胡杨、柽柳样地的采样深度为280 cm,距离河道50 m的苦豆子样地附近地下水位较浅,采样深度仅达到160 cm.采样时,在每一个采样点将每一层的土壤取出后装入已标记的自封袋中,分为两份,一份带回实验室自然风干,过筛(2 mm)备用,另一份4 ℃下保存备用.另外,每一土层取2个环刀样,用于测定土壤含水量及容重.同时记录采样点植被状况、经纬度等基本信息.本次采样共采集63份土壤样品,采样点基本情况信息见表1. ...
黄土高原中部三种典型绿化植物光合特性的季节变化
1
2020
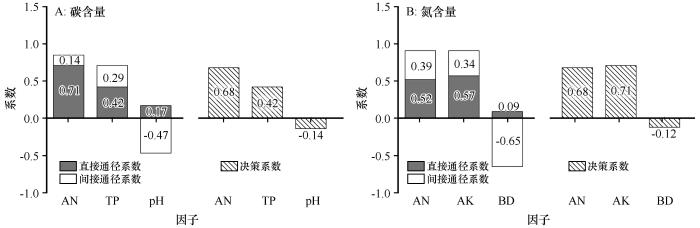
... 通径分析是一种能够定量解释自变量与因变量之间相关关系的方法.该方法计算得到简单相关系数、直接通径系数、间接通径系数以及决策系数用以衡量自变量xi (i=1,2,…,m)与因变量y之间的关系.直接通径系数与间接通经系数之和即为自变量xi 与因变量y的简单相关系数.决策系数R2通过公式(3)计算得出,它可以反映自变量对因变量的综合作用,当R2>0时,说明xi 对y起增强作用,当R2<0时,说明xi 对y起限制作用[17]. ...
Pattern and variation of C∶N∶P ratios in China's soils: a synthesis of observational data
1
2010
... 本研究发现黑河下游荒漠河岸林土壤碳氮平均含量分别为17.94、0.59 mg·g-1,这与Zhang等[11]的研究结果接近,低于全国平均水平(19.33、1.61 mg·g-1)[18-19].与干旱区的部分沙地相比[20-21],黑河下游荒漠河岸带的土壤具有较高的碳氮含量,这主要与荒漠河岸带的植被有关.荒漠河岸林生长于河道两侧,由河水补给的地下水为河岸带内植物生长发育提供了重要的水分保障[22],在黑河下游形成了以乔、灌、草为主的荒漠植被带[23],成为土壤碳氮输入的主要来源,相较于其他荒漠生态系统,荒漠河岸林土壤可能形成干旱区的大型“沃岛”. ...
江西省森林土壤碳氮磷含量及其化学计量比特征分析
1
2021
... 本研究发现黑河下游荒漠河岸林土壤碳氮平均含量分别为17.94、0.59 mg·g-1,这与Zhang等[11]的研究结果接近,低于全国平均水平(19.33、1.61 mg·g-1)[18-19].与干旱区的部分沙地相比[20-21],黑河下游荒漠河岸带的土壤具有较高的碳氮含量,这主要与荒漠河岸带的植被有关.荒漠河岸林生长于河道两侧,由河水补给的地下水为河岸带内植物生长发育提供了重要的水分保障[22],在黑河下游形成了以乔、灌、草为主的荒漠植被带[23],成为土壤碳氮输入的主要来源,相较于其他荒漠生态系统,荒漠河岸林土壤可能形成干旱区的大型“沃岛”. ...
科尔沁沙地4种典型灌木灌丛下土壤碳、氮、磷化学计量特征
1
2019
... 本研究发现黑河下游荒漠河岸林土壤碳氮平均含量分别为17.94、0.59 mg·g-1,这与Zhang等[11]的研究结果接近,低于全国平均水平(19.33、1.61 mg·g-1)[18-19].与干旱区的部分沙地相比[20-21],黑河下游荒漠河岸带的土壤具有较高的碳氮含量,这主要与荒漠河岸带的植被有关.荒漠河岸林生长于河道两侧,由河水补给的地下水为河岸带内植物生长发育提供了重要的水分保障[22],在黑河下游形成了以乔、灌、草为主的荒漠植被带[23],成为土壤碳氮输入的主要来源,相较于其他荒漠生态系统,荒漠河岸林土壤可能形成干旱区的大型“沃岛”. ...
农业利用对毛乌素沙地土壤碳氮含量及微生物量的影响
1
2020
... 本研究发现黑河下游荒漠河岸林土壤碳氮平均含量分别为17.94、0.59 mg·g-1,这与Zhang等[11]的研究结果接近,低于全国平均水平(19.33、1.61 mg·g-1)[18-19].与干旱区的部分沙地相比[20-21],黑河下游荒漠河岸带的土壤具有较高的碳氮含量,这主要与荒漠河岸带的植被有关.荒漠河岸林生长于河道两侧,由河水补给的地下水为河岸带内植物生长发育提供了重要的水分保障[22],在黑河下游形成了以乔、灌、草为主的荒漠植被带[23],成为土壤碳氮输入的主要来源,相较于其他荒漠生态系统,荒漠河岸林土壤可能形成干旱区的大型“沃岛”. ...
黑河下游典型生态系统水分补给源及优势植物水分来源研究
1
2012
... 本研究发现黑河下游荒漠河岸林土壤碳氮平均含量分别为17.94、0.59 mg·g-1,这与Zhang等[11]的研究结果接近,低于全国平均水平(19.33、1.61 mg·g-1)[18-19].与干旱区的部分沙地相比[20-21],黑河下游荒漠河岸带的土壤具有较高的碳氮含量,这主要与荒漠河岸带的植被有关.荒漠河岸林生长于河道两侧,由河水补给的地下水为河岸带内植物生长发育提供了重要的水分保障[22],在黑河下游形成了以乔、灌、草为主的荒漠植被带[23],成为土壤碳氮输入的主要来源,相较于其他荒漠生态系统,荒漠河岸林土壤可能形成干旱区的大型“沃岛”. ...
黑河下游极端干旱区胡杨净光合速率日变化及主要影响因子
1
2021
... 本研究发现黑河下游荒漠河岸林土壤碳氮平均含量分别为17.94、0.59 mg·g-1,这与Zhang等[11]的研究结果接近,低于全国平均水平(19.33、1.61 mg·g-1)[18-19].与干旱区的部分沙地相比[20-21],黑河下游荒漠河岸带的土壤具有较高的碳氮含量,这主要与荒漠河岸带的植被有关.荒漠河岸林生长于河道两侧,由河水补给的地下水为河岸带内植物生长发育提供了重要的水分保障[22],在黑河下游形成了以乔、灌、草为主的荒漠植被带[23],成为土壤碳氮输入的主要来源,相较于其他荒漠生态系统,荒漠河岸林土壤可能形成干旱区的大型“沃岛”. ...
旱化对浙江山地沼泽湿地土壤与植物碳氮磷含量的影响
1
2022
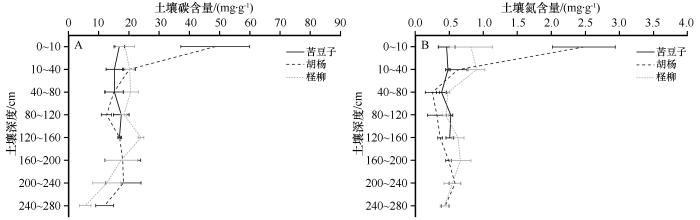
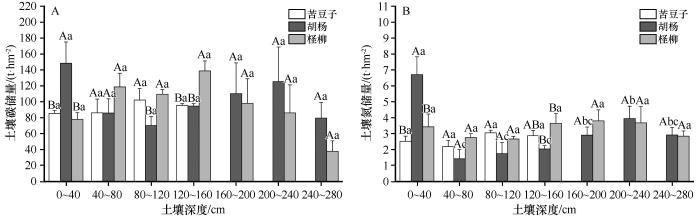
... 本次研究沿垂直河道方向采集了3种植被类型下的土壤.从水平方向上比较,深度0~10 cm的土层中,胡杨样地的碳氮含量显著高于其余两个样地,而深度10 cm以下的土层中,各样地之间的土壤碳氮含量差异并不明显,这主要与研究区植被类型差异有关.作为植物与土壤碳氮交换的场所,表层土壤的碳氮含量与植物碳氮输入量及转化效率有直接关系[24].根据之前的研究数据[25-27]可知,3个样地的优势植被类型的叶片碳含量较为相近,仅多枝柽柳叶片氮含量明显高于其他两种植被类型(表5).一方面,胡杨属于干旱区典型落叶乔木,其生物量往往大于灌、草植被[28],单位面积内为表层土壤提供更多的枝叶凋落物,增加了土壤碳氮输入量;另一方面,地表的凋落物具有调节水分、温度等土壤微气候的作用,进而能够影响土壤微生物活性[29],因此,胡杨植被的凋落物堆积为表层土壤碳氮转换创造了更好的条件.这种“植物效应”一般在土壤表层或者浅层中表现比较明显,随着土壤深度增加而逐渐减弱,直至某一深度基本消失[30],这也可以用来解释深度10 cm以下的土层中各样地之间的土壤碳氮含量差异并不明显的现象. ...
Rooting pattern and water relations of 3 pasture grasses growing in drying soil
2
1983
... 本次研究沿垂直河道方向采集了3种植被类型下的土壤.从水平方向上比较,深度0~10 cm的土层中,胡杨样地的碳氮含量显著高于其余两个样地,而深度10 cm以下的土层中,各样地之间的土壤碳氮含量差异并不明显,这主要与研究区植被类型差异有关.作为植物与土壤碳氮交换的场所,表层土壤的碳氮含量与植物碳氮输入量及转化效率有直接关系[24].根据之前的研究数据[25-27]可知,3个样地的优势植被类型的叶片碳含量较为相近,仅多枝柽柳叶片氮含量明显高于其他两种植被类型(表5).一方面,胡杨属于干旱区典型落叶乔木,其生物量往往大于灌、草植被[28],单位面积内为表层土壤提供更多的枝叶凋落物,增加了土壤碳氮输入量;另一方面,地表的凋落物具有调节水分、温度等土壤微气候的作用,进而能够影响土壤微生物活性[29],因此,胡杨植被的凋落物堆积为表层土壤碳氮转换创造了更好的条件.这种“植物效应”一般在土壤表层或者浅层中表现比较明显,随着土壤深度增加而逐渐减弱,直至某一深度基本消失[30],这也可以用来解释深度10 cm以下的土层中各样地之间的土壤碳氮含量差异并不明显的现象. ...
... Leaf carbon and nitrogen content of major vegetation
Table 5| 植物 | 碳含量/(mg·g-1) | 氮含量/(mg·g-1) | 数据来源 |
|---|
| 苦豆子 | 391.4 | 13.7 | [25] |
| 胡杨 | 435.1 | 14.9 | [26] |
| 多枝柽柳 | 380.27 | 30.42 | [27] |
从垂直方向上来看,随土壤深度增加,荒漠河岸林区土壤碳氮含量整体呈现波动下降趋势,0~40 cm深度的碳氮含量与160~280 cm深度相比降幅分别为39.98%、45.42%,说明荒漠河岸带土壤碳氮整体具有“表聚性”,这一变化规律与之前的研究结论相似[31-32].但是苦豆子与柽柳样地0~10 cm深度土壤碳氮含量与其下部10~40 cm深度土壤碳氮含量相比,“表聚性”不明显,究其原因可能包括以下几个方面:①表层以下土壤碳氮积累主要来源于根系及其分泌物[33].依赖土壤水或地下水生存的干旱区植物,受到水分胁迫影响,根冠比增大[34].因此,多枝柽柳及苦豆子的地下根系可能为其表层以下土壤碳氮提供了更多输入量.②研究区位于极端干旱区,大气降水极少,土壤水为依靠毛细管力上升的地下水[35],土壤含水量基本随深度增加而增加,表层以下的土壤水热条件在一定程度上优于表层,有利于土壤微生物对地下根系分解转化. ...
中国黑河下游荒漠河岸林植物群落水分利用策略研究
1
2014
... Leaf carbon and nitrogen content of major vegetation
Table 5| 植物 | 碳含量/(mg·g-1) | 氮含量/(mg·g-1) | 数据来源 |
|---|
| 苦豆子 | 391.4 | 13.7 | [25] |
| 胡杨 | 435.1 | 14.9 | [26] |
| 多枝柽柳 | 380.27 | 30.42 | [27] |
从垂直方向上来看,随土壤深度增加,荒漠河岸林区土壤碳氮含量整体呈现波动下降趋势,0~40 cm深度的碳氮含量与160~280 cm深度相比降幅分别为39.98%、45.42%,说明荒漠河岸带土壤碳氮整体具有“表聚性”,这一变化规律与之前的研究结论相似[31-32].但是苦豆子与柽柳样地0~10 cm深度土壤碳氮含量与其下部10~40 cm深度土壤碳氮含量相比,“表聚性”不明显,究其原因可能包括以下几个方面:①表层以下土壤碳氮积累主要来源于根系及其分泌物[33].依赖土壤水或地下水生存的干旱区植物,受到水分胁迫影响,根冠比增大[34].因此,多枝柽柳及苦豆子的地下根系可能为其表层以下土壤碳氮提供了更多输入量.②研究区位于极端干旱区,大气降水极少,土壤水为依靠毛细管力上升的地下水[35],土壤含水量基本随深度增加而增加,表层以下的土壤水热条件在一定程度上优于表层,有利于土壤微生物对地下根系分解转化. ...
伊犁河谷苦豆子C、N、P含量变化及化学计量特征
2
2018
... 本次研究沿垂直河道方向采集了3种植被类型下的土壤.从水平方向上比较,深度0~10 cm的土层中,胡杨样地的碳氮含量显著高于其余两个样地,而深度10 cm以下的土层中,各样地之间的土壤碳氮含量差异并不明显,这主要与研究区植被类型差异有关.作为植物与土壤碳氮交换的场所,表层土壤的碳氮含量与植物碳氮输入量及转化效率有直接关系[24].根据之前的研究数据[25-27]可知,3个样地的优势植被类型的叶片碳含量较为相近,仅多枝柽柳叶片氮含量明显高于其他两种植被类型(表5).一方面,胡杨属于干旱区典型落叶乔木,其生物量往往大于灌、草植被[28],单位面积内为表层土壤提供更多的枝叶凋落物,增加了土壤碳氮输入量;另一方面,地表的凋落物具有调节水分、温度等土壤微气候的作用,进而能够影响土壤微生物活性[29],因此,胡杨植被的凋落物堆积为表层土壤碳氮转换创造了更好的条件.这种“植物效应”一般在土壤表层或者浅层中表现比较明显,随着土壤深度增加而逐渐减弱,直至某一深度基本消失[30],这也可以用来解释深度10 cm以下的土层中各样地之间的土壤碳氮含量差异并不明显的现象. ...
... Leaf carbon and nitrogen content of major vegetation
Table 5| 植物 | 碳含量/(mg·g-1) | 氮含量/(mg·g-1) | 数据来源 |
|---|
| 苦豆子 | 391.4 | 13.7 | [25] |
| 胡杨 | 435.1 | 14.9 | [26] |
| 多枝柽柳 | 380.27 | 30.42 | [27] |
从垂直方向上来看,随土壤深度增加,荒漠河岸林区土壤碳氮含量整体呈现波动下降趋势,0~40 cm深度的碳氮含量与160~280 cm深度相比降幅分别为39.98%、45.42%,说明荒漠河岸带土壤碳氮整体具有“表聚性”,这一变化规律与之前的研究结论相似[31-32].但是苦豆子与柽柳样地0~10 cm深度土壤碳氮含量与其下部10~40 cm深度土壤碳氮含量相比,“表聚性”不明显,究其原因可能包括以下几个方面:①表层以下土壤碳氮积累主要来源于根系及其分泌物[33].依赖土壤水或地下水生存的干旱区植物,受到水分胁迫影响,根冠比增大[34].因此,多枝柽柳及苦豆子的地下根系可能为其表层以下土壤碳氮提供了更多输入量.②研究区位于极端干旱区,大气降水极少,土壤水为依靠毛细管力上升的地下水[35],土壤含水量基本随深度增加而增加,表层以下的土壤水热条件在一定程度上优于表层,有利于土壤微生物对地下根系分解转化. ...
Application of heuristic approaches for prediction of hydrological drought using multi-scalar streamflow drought index
1
2019
... 本次研究沿垂直河道方向采集了3种植被类型下的土壤.从水平方向上比较,深度0~10 cm的土层中,胡杨样地的碳氮含量显著高于其余两个样地,而深度10 cm以下的土层中,各样地之间的土壤碳氮含量差异并不明显,这主要与研究区植被类型差异有关.作为植物与土壤碳氮交换的场所,表层土壤的碳氮含量与植物碳氮输入量及转化效率有直接关系[24].根据之前的研究数据[25-27]可知,3个样地的优势植被类型的叶片碳含量较为相近,仅多枝柽柳叶片氮含量明显高于其他两种植被类型(表5).一方面,胡杨属于干旱区典型落叶乔木,其生物量往往大于灌、草植被[28],单位面积内为表层土壤提供更多的枝叶凋落物,增加了土壤碳氮输入量;另一方面,地表的凋落物具有调节水分、温度等土壤微气候的作用,进而能够影响土壤微生物活性[29],因此,胡杨植被的凋落物堆积为表层土壤碳氮转换创造了更好的条件.这种“植物效应”一般在土壤表层或者浅层中表现比较明显,随着土壤深度增加而逐渐减弱,直至某一深度基本消失[30],这也可以用来解释深度10 cm以下的土层中各样地之间的土壤碳氮含量差异并不明显的现象. ...
黄土高原森林带土壤养分和微生物量及其生态化学计量变化特征
1
2020
... 本次研究沿垂直河道方向采集了3种植被类型下的土壤.从水平方向上比较,深度0~10 cm的土层中,胡杨样地的碳氮含量显著高于其余两个样地,而深度10 cm以下的土层中,各样地之间的土壤碳氮含量差异并不明显,这主要与研究区植被类型差异有关.作为植物与土壤碳氮交换的场所,表层土壤的碳氮含量与植物碳氮输入量及转化效率有直接关系[24].根据之前的研究数据[25-27]可知,3个样地的优势植被类型的叶片碳含量较为相近,仅多枝柽柳叶片氮含量明显高于其他两种植被类型(表5).一方面,胡杨属于干旱区典型落叶乔木,其生物量往往大于灌、草植被[28],单位面积内为表层土壤提供更多的枝叶凋落物,增加了土壤碳氮输入量;另一方面,地表的凋落物具有调节水分、温度等土壤微气候的作用,进而能够影响土壤微生物活性[29],因此,胡杨植被的凋落物堆积为表层土壤碳氮转换创造了更好的条件.这种“植物效应”一般在土壤表层或者浅层中表现比较明显,随着土壤深度增加而逐渐减弱,直至某一深度基本消失[30],这也可以用来解释深度10 cm以下的土层中各样地之间的土壤碳氮含量差异并不明显的现象. ...
Floodplain biogeochemical mosaics: a multidimensional view of alluvial soils
1
2014
... 本次研究沿垂直河道方向采集了3种植被类型下的土壤.从水平方向上比较,深度0~10 cm的土层中,胡杨样地的碳氮含量显著高于其余两个样地,而深度10 cm以下的土层中,各样地之间的土壤碳氮含量差异并不明显,这主要与研究区植被类型差异有关.作为植物与土壤碳氮交换的场所,表层土壤的碳氮含量与植物碳氮输入量及转化效率有直接关系[24].根据之前的研究数据[25-27]可知,3个样地的优势植被类型的叶片碳含量较为相近,仅多枝柽柳叶片氮含量明显高于其他两种植被类型(表5).一方面,胡杨属于干旱区典型落叶乔木,其生物量往往大于灌、草植被[28],单位面积内为表层土壤提供更多的枝叶凋落物,增加了土壤碳氮输入量;另一方面,地表的凋落物具有调节水分、温度等土壤微气候的作用,进而能够影响土壤微生物活性[29],因此,胡杨植被的凋落物堆积为表层土壤碳氮转换创造了更好的条件.这种“植物效应”一般在土壤表层或者浅层中表现比较明显,随着土壤深度增加而逐渐减弱,直至某一深度基本消失[30],这也可以用来解释深度10 cm以下的土层中各样地之间的土壤碳氮含量差异并不明显的现象. ...
Multi-scale variability of soil carbon and nitrogen in the middle reaches of the Heihe River basin,northwestern China
1
2016
... 从垂直方向上来看,随土壤深度增加,荒漠河岸林区土壤碳氮含量整体呈现波动下降趋势,0~40 cm深度的碳氮含量与160~280 cm深度相比降幅分别为39.98%、45.42%,说明荒漠河岸带土壤碳氮整体具有“表聚性”,这一变化规律与之前的研究结论相似[31-32].但是苦豆子与柽柳样地0~10 cm深度土壤碳氮含量与其下部10~40 cm深度土壤碳氮含量相比,“表聚性”不明显,究其原因可能包括以下几个方面:①表层以下土壤碳氮积累主要来源于根系及其分泌物[33].依赖土壤水或地下水生存的干旱区植物,受到水分胁迫影响,根冠比增大[34].因此,多枝柽柳及苦豆子的地下根系可能为其表层以下土壤碳氮提供了更多输入量.②研究区位于极端干旱区,大气降水极少,土壤水为依靠毛细管力上升的地下水[35],土壤含水量基本随深度增加而增加,表层以下的土壤水热条件在一定程度上优于表层,有利于土壤微生物对地下根系分解转化. ...
艾比湖地区盐生植物群落土壤氮素的垂直分布特征
1
2014
... 从垂直方向上来看,随土壤深度增加,荒漠河岸林区土壤碳氮含量整体呈现波动下降趋势,0~40 cm深度的碳氮含量与160~280 cm深度相比降幅分别为39.98%、45.42%,说明荒漠河岸带土壤碳氮整体具有“表聚性”,这一变化规律与之前的研究结论相似[31-32].但是苦豆子与柽柳样地0~10 cm深度土壤碳氮含量与其下部10~40 cm深度土壤碳氮含量相比,“表聚性”不明显,究其原因可能包括以下几个方面:①表层以下土壤碳氮积累主要来源于根系及其分泌物[33].依赖土壤水或地下水生存的干旱区植物,受到水分胁迫影响,根冠比增大[34].因此,多枝柽柳及苦豆子的地下根系可能为其表层以下土壤碳氮提供了更多输入量.②研究区位于极端干旱区,大气降水极少,土壤水为依靠毛细管力上升的地下水[35],土壤含水量基本随深度增加而增加,表层以下的土壤水热条件在一定程度上优于表层,有利于土壤微生物对地下根系分解转化. ...
Understanding deep roots and their functions in ecosystems:an advocacy for more unconventional research
1
2016
... 从垂直方向上来看,随土壤深度增加,荒漠河岸林区土壤碳氮含量整体呈现波动下降趋势,0~40 cm深度的碳氮含量与160~280 cm深度相比降幅分别为39.98%、45.42%,说明荒漠河岸带土壤碳氮整体具有“表聚性”,这一变化规律与之前的研究结论相似[31-32].但是苦豆子与柽柳样地0~10 cm深度土壤碳氮含量与其下部10~40 cm深度土壤碳氮含量相比,“表聚性”不明显,究其原因可能包括以下几个方面:①表层以下土壤碳氮积累主要来源于根系及其分泌物[33].依赖土壤水或地下水生存的干旱区植物,受到水分胁迫影响,根冠比增大[34].因此,多枝柽柳及苦豆子的地下根系可能为其表层以下土壤碳氮提供了更多输入量.②研究区位于极端干旱区,大气降水极少,土壤水为依靠毛细管力上升的地下水[35],土壤含水量基本随深度增加而增加,表层以下的土壤水热条件在一定程度上优于表层,有利于土壤微生物对地下根系分解转化. ...
荒漠河岸林胡杨养分状况研究
1
2011
... 从垂直方向上来看,随土壤深度增加,荒漠河岸林区土壤碳氮含量整体呈现波动下降趋势,0~40 cm深度的碳氮含量与160~280 cm深度相比降幅分别为39.98%、45.42%,说明荒漠河岸带土壤碳氮整体具有“表聚性”,这一变化规律与之前的研究结论相似[31-32].但是苦豆子与柽柳样地0~10 cm深度土壤碳氮含量与其下部10~40 cm深度土壤碳氮含量相比,“表聚性”不明显,究其原因可能包括以下几个方面:①表层以下土壤碳氮积累主要来源于根系及其分泌物[33].依赖土壤水或地下水生存的干旱区植物,受到水分胁迫影响,根冠比增大[34].因此,多枝柽柳及苦豆子的地下根系可能为其表层以下土壤碳氮提供了更多输入量.②研究区位于极端干旱区,大气降水极少,土壤水为依靠毛细管力上升的地下水[35],土壤含水量基本随深度增加而增加,表层以下的土壤水热条件在一定程度上优于表层,有利于土壤微生物对地下根系分解转化. ...
荒漠河岸多枝柽柳灌丛碳氮磷化学计量特征及其影响因素
1
2021
... 从垂直方向上来看,随土壤深度增加,荒漠河岸林区土壤碳氮含量整体呈现波动下降趋势,0~40 cm深度的碳氮含量与160~280 cm深度相比降幅分别为39.98%、45.42%,说明荒漠河岸带土壤碳氮整体具有“表聚性”,这一变化规律与之前的研究结论相似[31-32].但是苦豆子与柽柳样地0~10 cm深度土壤碳氮含量与其下部10~40 cm深度土壤碳氮含量相比,“表聚性”不明显,究其原因可能包括以下几个方面:①表层以下土壤碳氮积累主要来源于根系及其分泌物[33].依赖土壤水或地下水生存的干旱区植物,受到水分胁迫影响,根冠比增大[34].因此,多枝柽柳及苦豆子的地下根系可能为其表层以下土壤碳氮提供了更多输入量.②研究区位于极端干旱区,大气降水极少,土壤水为依靠毛细管力上升的地下水[35],土壤含水量基本随深度增加而增加,表层以下的土壤水热条件在一定程度上优于表层,有利于土壤微生物对地下根系分解转化. ...
Vertical distribution of fine roots of Tamarix ramosissima in an arid region of southern Nevada
1
2013
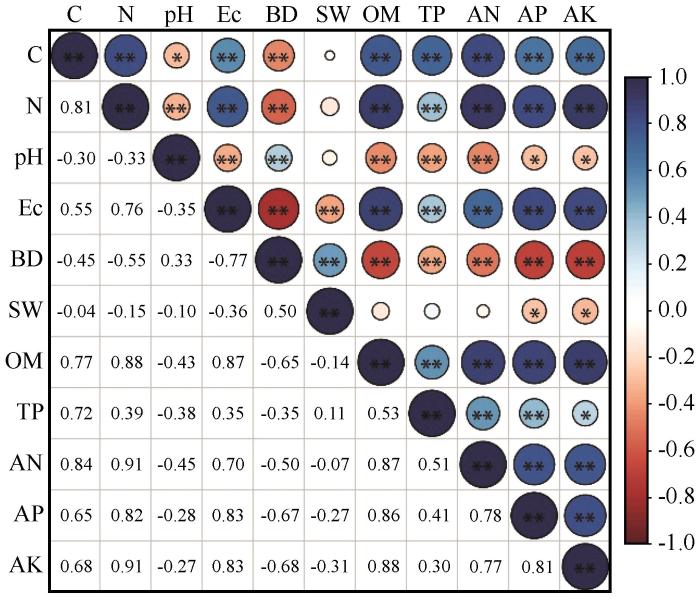
... 土壤碳氮含量是碳氮元素在土壤中积累和消耗平衡的结果,地上凋落物以及地下根系等为重要输入来源,直接影响土壤碳氮积累量[36-37],土壤水热条件、理化性质等也是影响碳氮积累转化效率的关键因素[38].本文对荒漠河岸林土壤碳氮含量与理化性质进行相关分析显示,电导率、有机质等与土壤碳氮含量均具有显著相关性(图5). ...
四川龙溪-虹口常绿阔叶林凋落物与土壤碳、氮、磷化学计量特征分析
1
2019
... 土壤碳氮含量是碳氮元素在土壤中积累和消耗平衡的结果,地上凋落物以及地下根系等为重要输入来源,直接影响土壤碳氮积累量[36-37],土壤水热条件、理化性质等也是影响碳氮积累转化效率的关键因素[38].本文对荒漠河岸林土壤碳氮含量与理化性质进行相关分析显示,电导率、有机质等与土壤碳氮含量均具有显著相关性(图5). ...
南滚河国家级自然保护区典型植被类型土壤有机碳及全氮储量的空间分布特征
1
2019
... 土壤碳氮含量是碳氮元素在土壤中积累和消耗平衡的结果,地上凋落物以及地下根系等为重要输入来源,直接影响土壤碳氮积累量[36-37],土壤水热条件、理化性质等也是影响碳氮积累转化效率的关键因素[38].本文对荒漠河岸林土壤碳氮含量与理化性质进行相关分析显示,电导率、有机质等与土壤碳氮含量均具有显著相关性(图5). ...
土壤有机碳研究进展
1
2008
... 有机质、总磷、速效磷、碱解氮、速效钾与总碳、总氮均属于土壤的重要物质,它们在迁移与转化过程中往往伴有一定的协同作用,在相关关系上常表现为显著正相关.土壤有机质主要是由动、植物残体分解转化而来,有机质经矿化后可转化为氮、磷等元素,另外,有机质自身较强的吸水能力,能够保持土壤水分,改善土壤环境[39].作为植物内部磷脂、核酸、核蛋白等的关键组分,磷元素具有促进植物生长、增加植物生物量的重要作用[40].钾元素是植物体内重要的酶活化剂,能够促进植物叶片的光合与呼吸作用,提高植物抗旱能力[41].随着土壤中的这些物质增多,植物的生长状态提升,固定碳氮的能力增强,进而影响土壤碳氮输入量以及转化效率. ...
土壤磷与植物关系研究进展
1
2013
... 有机质、总磷、速效磷、碱解氮、速效钾与总碳、总氮均属于土壤的重要物质,它们在迁移与转化过程中往往伴有一定的协同作用,在相关关系上常表现为显著正相关.土壤有机质主要是由动、植物残体分解转化而来,有机质经矿化后可转化为氮、磷等元素,另外,有机质自身较强的吸水能力,能够保持土壤水分,改善土壤环境[39].作为植物内部磷脂、核酸、核蛋白等的关键组分,磷元素具有促进植物生长、增加植物生物量的重要作用[40].钾元素是植物体内重要的酶活化剂,能够促进植物叶片的光合与呼吸作用,提高植物抗旱能力[41].随着土壤中的这些物质增多,植物的生长状态提升,固定碳氮的能力增强,进而影响土壤碳氮输入量以及转化效率. ...
新疆土壤有机碳与土壤理化性质的相关性
1
2017
... 有机质、总磷、速效磷、碱解氮、速效钾与总碳、总氮均属于土壤的重要物质,它们在迁移与转化过程中往往伴有一定的协同作用,在相关关系上常表现为显著正相关.土壤有机质主要是由动、植物残体分解转化而来,有机质经矿化后可转化为氮、磷等元素,另外,有机质自身较强的吸水能力,能够保持土壤水分,改善土壤环境[39].作为植物内部磷脂、核酸、核蛋白等的关键组分,磷元素具有促进植物生长、增加植物生物量的重要作用[40].钾元素是植物体内重要的酶活化剂,能够促进植物叶片的光合与呼吸作用,提高植物抗旱能力[41].随着土壤中的这些物质增多,植物的生长状态提升,固定碳氮的能力增强,进而影响土壤碳氮输入量以及转化效率. ...
祁连山南坡不同土地利用方式下土壤碳氮含量及通径分析
1
2021
... pH与土壤碳氮含量为显著的负相关关系,这与之前的研究一致[42].作为土壤环境关键参数,pH与土壤微生物活性紧密相关.Tang等[43]认为pH小于5.5或大于8.5时对土壤微生物活性具有抑制作用.本研究区内土壤pH多在8.5~9.5,高碱性环境不利于土壤微生物活动,因此,pH与土壤碳氮呈显著负相关关系.电导率是指示土壤盐分的基础指标,一般研究认为,土壤盐分含量增高会抑制植物生长,进而减少土壤碳氮的输入量,因此,两者之间为负相关关系[44-45],但是本研究结果显示电导率与土壤碳氮含量之间为正相关关系,这与蒋腊梅等[46]对干旱区草地土壤的研究结果一致.出现这种现象的原因可能是,与土壤盐分的抑制作用相比,极端干旱区内蒸发强导致的盐分在土壤表层大量积累的空间分布特征对电导率与土壤碳氮相关性的影响更大.容重变化对土壤碳氮含量的影响主要体现在调节土壤中的水、肥、气、热等要素,这一方面可以直接影响土壤微生物的活性,另一方面也可以影响地上植被的生长状况,进而影响土壤碳氮输入与转化[47-48].本研究显示土壤容重与碳氮含量呈现显著负相关关系,这与张旭冉等[49]在克氏针茅草原区的研究结果一致. ...
Effect of freeze-thaw cycles on carbon stocks of saline-alkali paddy soil
1
2016
... pH与土壤碳氮含量为显著的负相关关系,这与之前的研究一致[42].作为土壤环境关键参数,pH与土壤微生物活性紧密相关.Tang等[43]认为pH小于5.5或大于8.5时对土壤微生物活性具有抑制作用.本研究区内土壤pH多在8.5~9.5,高碱性环境不利于土壤微生物活动,因此,pH与土壤碳氮呈显著负相关关系.电导率是指示土壤盐分的基础指标,一般研究认为,土壤盐分含量增高会抑制植物生长,进而减少土壤碳氮的输入量,因此,两者之间为负相关关系[44-45],但是本研究结果显示电导率与土壤碳氮含量之间为正相关关系,这与蒋腊梅等[46]对干旱区草地土壤的研究结果一致.出现这种现象的原因可能是,与土壤盐分的抑制作用相比,极端干旱区内蒸发强导致的盐分在土壤表层大量积累的空间分布特征对电导率与土壤碳氮相关性的影响更大.容重变化对土壤碳氮含量的影响主要体现在调节土壤中的水、肥、气、热等要素,这一方面可以直接影响土壤微生物的活性,另一方面也可以影响地上植被的生长状况,进而影响土壤碳氮输入与转化[47-48].本研究显示土壤容重与碳氮含量呈现显著负相关关系,这与张旭冉等[49]在克氏针茅草原区的研究结果一致. ...
荒漠绿洲农田盐渍化过程中土壤环境的演变过程
1
2014
... pH与土壤碳氮含量为显著的负相关关系,这与之前的研究一致[42].作为土壤环境关键参数,pH与土壤微生物活性紧密相关.Tang等[43]认为pH小于5.5或大于8.5时对土壤微生物活性具有抑制作用.本研究区内土壤pH多在8.5~9.5,高碱性环境不利于土壤微生物活动,因此,pH与土壤碳氮呈显著负相关关系.电导率是指示土壤盐分的基础指标,一般研究认为,土壤盐分含量增高会抑制植物生长,进而减少土壤碳氮的输入量,因此,两者之间为负相关关系[44-45],但是本研究结果显示电导率与土壤碳氮含量之间为正相关关系,这与蒋腊梅等[46]对干旱区草地土壤的研究结果一致.出现这种现象的原因可能是,与土壤盐分的抑制作用相比,极端干旱区内蒸发强导致的盐分在土壤表层大量积累的空间分布特征对电导率与土壤碳氮相关性的影响更大.容重变化对土壤碳氮含量的影响主要体现在调节土壤中的水、肥、气、热等要素,这一方面可以直接影响土壤微生物的活性,另一方面也可以影响地上植被的生长状况,进而影响土壤碳氮输入与转化[47-48].本研究显示土壤容重与碳氮含量呈现显著负相关关系,这与张旭冉等[49]在克氏针茅草原区的研究结果一致. ...
盐碱地柽柳“盐岛”和“肥岛”效应及其碳氮磷生态化学计量学特征
1
2015
... pH与土壤碳氮含量为显著的负相关关系,这与之前的研究一致[42].作为土壤环境关键参数,pH与土壤微生物活性紧密相关.Tang等[43]认为pH小于5.5或大于8.5时对土壤微生物活性具有抑制作用.本研究区内土壤pH多在8.5~9.5,高碱性环境不利于土壤微生物活动,因此,pH与土壤碳氮呈显著负相关关系.电导率是指示土壤盐分的基础指标,一般研究认为,土壤盐分含量增高会抑制植物生长,进而减少土壤碳氮的输入量,因此,两者之间为负相关关系[44-45],但是本研究结果显示电导率与土壤碳氮含量之间为正相关关系,这与蒋腊梅等[46]对干旱区草地土壤的研究结果一致.出现这种现象的原因可能是,与土壤盐分的抑制作用相比,极端干旱区内蒸发强导致的盐分在土壤表层大量积累的空间分布特征对电导率与土壤碳氮相关性的影响更大.容重变化对土壤碳氮含量的影响主要体现在调节土壤中的水、肥、气、热等要素,这一方面可以直接影响土壤微生物的活性,另一方面也可以影响地上植被的生长状况,进而影响土壤碳氮输入与转化[47-48].本研究显示土壤容重与碳氮含量呈现显著负相关关系,这与张旭冉等[49]在克氏针茅草原区的研究结果一致. ...
不同管理模式对干旱区草地土壤有机碳氮库的影响及其影响因素探究
1
2018
... pH与土壤碳氮含量为显著的负相关关系,这与之前的研究一致[42].作为土壤环境关键参数,pH与土壤微生物活性紧密相关.Tang等[43]认为pH小于5.5或大于8.5时对土壤微生物活性具有抑制作用.本研究区内土壤pH多在8.5~9.5,高碱性环境不利于土壤微生物活动,因此,pH与土壤碳氮呈显著负相关关系.电导率是指示土壤盐分的基础指标,一般研究认为,土壤盐分含量增高会抑制植物生长,进而减少土壤碳氮的输入量,因此,两者之间为负相关关系[44-45],但是本研究结果显示电导率与土壤碳氮含量之间为正相关关系,这与蒋腊梅等[46]对干旱区草地土壤的研究结果一致.出现这种现象的原因可能是,与土壤盐分的抑制作用相比,极端干旱区内蒸发强导致的盐分在土壤表层大量积累的空间分布特征对电导率与土壤碳氮相关性的影响更大.容重变化对土壤碳氮含量的影响主要体现在调节土壤中的水、肥、气、热等要素,这一方面可以直接影响土壤微生物的活性,另一方面也可以影响地上植被的生长状况,进而影响土壤碳氮输入与转化[47-48].本研究显示土壤容重与碳氮含量呈现显著负相关关系,这与张旭冉等[49]在克氏针茅草原区的研究结果一致. ...
城市森林土壤有机碳密度及影响因子变化研究
1
2022
... pH与土壤碳氮含量为显著的负相关关系,这与之前的研究一致[42].作为土壤环境关键参数,pH与土壤微生物活性紧密相关.Tang等[43]认为pH小于5.5或大于8.5时对土壤微生物活性具有抑制作用.本研究区内土壤pH多在8.5~9.5,高碱性环境不利于土壤微生物活动,因此,pH与土壤碳氮呈显著负相关关系.电导率是指示土壤盐分的基础指标,一般研究认为,土壤盐分含量增高会抑制植物生长,进而减少土壤碳氮的输入量,因此,两者之间为负相关关系[44-45],但是本研究结果显示电导率与土壤碳氮含量之间为正相关关系,这与蒋腊梅等[46]对干旱区草地土壤的研究结果一致.出现这种现象的原因可能是,与土壤盐分的抑制作用相比,极端干旱区内蒸发强导致的盐分在土壤表层大量积累的空间分布特征对电导率与土壤碳氮相关性的影响更大.容重变化对土壤碳氮含量的影响主要体现在调节土壤中的水、肥、气、热等要素,这一方面可以直接影响土壤微生物的活性,另一方面也可以影响地上植被的生长状况,进而影响土壤碳氮输入与转化[47-48].本研究显示土壤容重与碳氮含量呈现显著负相关关系,这与张旭冉等[49]在克氏针茅草原区的研究结果一致. ...
荒漠草原沙漠化对土壤物理和化学特性的影响
1
2019
... pH与土壤碳氮含量为显著的负相关关系,这与之前的研究一致[42].作为土壤环境关键参数,pH与土壤微生物活性紧密相关.Tang等[43]认为pH小于5.5或大于8.5时对土壤微生物活性具有抑制作用.本研究区内土壤pH多在8.5~9.5,高碱性环境不利于土壤微生物活动,因此,pH与土壤碳氮呈显著负相关关系.电导率是指示土壤盐分的基础指标,一般研究认为,土壤盐分含量增高会抑制植物生长,进而减少土壤碳氮的输入量,因此,两者之间为负相关关系[44-45],但是本研究结果显示电导率与土壤碳氮含量之间为正相关关系,这与蒋腊梅等[46]对干旱区草地土壤的研究结果一致.出现这种现象的原因可能是,与土壤盐分的抑制作用相比,极端干旱区内蒸发强导致的盐分在土壤表层大量积累的空间分布特征对电导率与土壤碳氮相关性的影响更大.容重变化对土壤碳氮含量的影响主要体现在调节土壤中的水、肥、气、热等要素,这一方面可以直接影响土壤微生物的活性,另一方面也可以影响地上植被的生长状况,进而影响土壤碳氮输入与转化[47-48].本研究显示土壤容重与碳氮含量呈现显著负相关关系,这与张旭冉等[49]在克氏针茅草原区的研究结果一致. ...
克氏针茅草原区有机碳与土壤理化性质的关系
1
2020
... pH与土壤碳氮含量为显著的负相关关系,这与之前的研究一致[42].作为土壤环境关键参数,pH与土壤微生物活性紧密相关.Tang等[43]认为pH小于5.5或大于8.5时对土壤微生物活性具有抑制作用.本研究区内土壤pH多在8.5~9.5,高碱性环境不利于土壤微生物活动,因此,pH与土壤碳氮呈显著负相关关系.电导率是指示土壤盐分的基础指标,一般研究认为,土壤盐分含量增高会抑制植物生长,进而减少土壤碳氮的输入量,因此,两者之间为负相关关系[44-45],但是本研究结果显示电导率与土壤碳氮含量之间为正相关关系,这与蒋腊梅等[46]对干旱区草地土壤的研究结果一致.出现这种现象的原因可能是,与土壤盐分的抑制作用相比,极端干旱区内蒸发强导致的盐分在土壤表层大量积累的空间分布特征对电导率与土壤碳氮相关性的影响更大.容重变化对土壤碳氮含量的影响主要体现在调节土壤中的水、肥、气、热等要素,这一方面可以直接影响土壤微生物的活性,另一方面也可以影响地上植被的生长状况,进而影响土壤碳氮输入与转化[47-48].本研究显示土壤容重与碳氮含量呈现显著负相关关系,这与张旭冉等[49]在克氏针茅草原区的研究结果一致. ...





 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}