1
2017
... 氮(N)和磷(P)是影响植物生长发育的主要营养元素[1].N、P元素是构成有机体内蛋白质、磷酸等物质的基础,对调节植物生长和代谢具有重要作用[2].通常认为氮磷比(N∶P)是判断植物生长发育是否受N、P元素的限制,因此对植物N、P元素化学计量特征的研究有助于加深对植物生长和环境适应性的理解,进而促进对陆地生态系统养分调控和元素生物地球化学循环机制的深入理解[3].N∶P可以衡量植物生长对于N、P的限制程度,N、P含量较低的植物会倾向于在氮限制或磷限制的环境中成为优势种;传统的氮磷限制假说认为,当植物叶片N∶P小于14时,植物生长受氮限制;当N∶P大于16时,植物生长受磷元素所限制;当N∶P为14~16时,植物生长可能同时受或不受氮磷元素限制[4-6],但也有诸多研究认为不同的功能群植物受氮磷限制的临界氮磷比值存在差异[7],如常绿针叶林、豆科植物及草本植物的临界值分别为10、8~9、12.尽管氮磷限制假说饱受争议,但其依然广泛运用于荒漠[8-9]、草原[10]、森林[11]等多种生态系统中用于推断植物、土壤的养分限制.在养分受限的环境中,植物养分的分配对植物资源的吸收和利用起着关键作用,并与植物生长速率、繁殖、防御和适应环境异质性的策略有关.此外,植物的地上和地下部分直接相互依赖,与捕获和利用光、养分和水资源的吸收和利用密切相关[12],因此将植物器官间营养元素可塑性作为植物生长和环境适应性的重要研究内容.通常使用变异系数(CV)来描述植物器官间元素变异性或可塑性[13],由于植物不同器官所面临的环境不同,不同器官的适应能力可能不同.可塑性高的生物具有更大的生态幅度和适应性,不同器官间元素分配不同,可塑性不同,功能不同,因此植物不同器官间往往存在养分的幂指数关系. ...
氮添加对油蒿群落植物叶片生态化学计量特征的影响
1
2016
... 氮(N)和磷(P)是影响植物生长发育的主要营养元素[1].N、P元素是构成有机体内蛋白质、磷酸等物质的基础,对调节植物生长和代谢具有重要作用[2].通常认为氮磷比(N∶P)是判断植物生长发育是否受N、P元素的限制,因此对植物N、P元素化学计量特征的研究有助于加深对植物生长和环境适应性的理解,进而促进对陆地生态系统养分调控和元素生物地球化学循环机制的深入理解[3].N∶P可以衡量植物生长对于N、P的限制程度,N、P含量较低的植物会倾向于在氮限制或磷限制的环境中成为优势种;传统的氮磷限制假说认为,当植物叶片N∶P小于14时,植物生长受氮限制;当N∶P大于16时,植物生长受磷元素所限制;当N∶P为14~16时,植物生长可能同时受或不受氮磷元素限制[4-6],但也有诸多研究认为不同的功能群植物受氮磷限制的临界氮磷比值存在差异[7],如常绿针叶林、豆科植物及草本植物的临界值分别为10、8~9、12.尽管氮磷限制假说饱受争议,但其依然广泛运用于荒漠[8-9]、草原[10]、森林[11]等多种生态系统中用于推断植物、土壤的养分限制.在养分受限的环境中,植物养分的分配对植物资源的吸收和利用起着关键作用,并与植物生长速率、繁殖、防御和适应环境异质性的策略有关.此外,植物的地上和地下部分直接相互依赖,与捕获和利用光、养分和水资源的吸收和利用密切相关[12],因此将植物器官间营养元素可塑性作为植物生长和环境适应性的重要研究内容.通常使用变异系数(CV)来描述植物器官间元素变异性或可塑性[13],由于植物不同器官所面临的环境不同,不同器官的适应能力可能不同.可塑性高的生物具有更大的生态幅度和适应性,不同器官间元素分配不同,可塑性不同,功能不同,因此植物不同器官间往往存在养分的幂指数关系. ...
The vegetation N∶P ratio:a new tool to detect the nature of nutrient limitation
2
1996
... 氮(N)和磷(P)是影响植物生长发育的主要营养元素[1].N、P元素是构成有机体内蛋白质、磷酸等物质的基础,对调节植物生长和代谢具有重要作用[2].通常认为氮磷比(N∶P)是判断植物生长发育是否受N、P元素的限制,因此对植物N、P元素化学计量特征的研究有助于加深对植物生长和环境适应性的理解,进而促进对陆地生态系统养分调控和元素生物地球化学循环机制的深入理解[3].N∶P可以衡量植物生长对于N、P的限制程度,N、P含量较低的植物会倾向于在氮限制或磷限制的环境中成为优势种;传统的氮磷限制假说认为,当植物叶片N∶P小于14时,植物生长受氮限制;当N∶P大于16时,植物生长受磷元素所限制;当N∶P为14~16时,植物生长可能同时受或不受氮磷元素限制[4-6],但也有诸多研究认为不同的功能群植物受氮磷限制的临界氮磷比值存在差异[7],如常绿针叶林、豆科植物及草本植物的临界值分别为10、8~9、12.尽管氮磷限制假说饱受争议,但其依然广泛运用于荒漠[8-9]、草原[10]、森林[11]等多种生态系统中用于推断植物、土壤的养分限制.在养分受限的环境中,植物养分的分配对植物资源的吸收和利用起着关键作用,并与植物生长速率、繁殖、防御和适应环境异质性的策略有关.此外,植物的地上和地下部分直接相互依赖,与捕获和利用光、养分和水资源的吸收和利用密切相关[12],因此将植物器官间营养元素可塑性作为植物生长和环境适应性的重要研究内容.通常使用变异系数(CV)来描述植物器官间元素变异性或可塑性[13],由于植物不同器官所面临的环境不同,不同器官的适应能力可能不同.可塑性高的生物具有更大的生态幅度和适应性,不同器官间元素分配不同,可塑性不同,功能不同,因此植物不同器官间往往存在养分的幂指数关系. ...
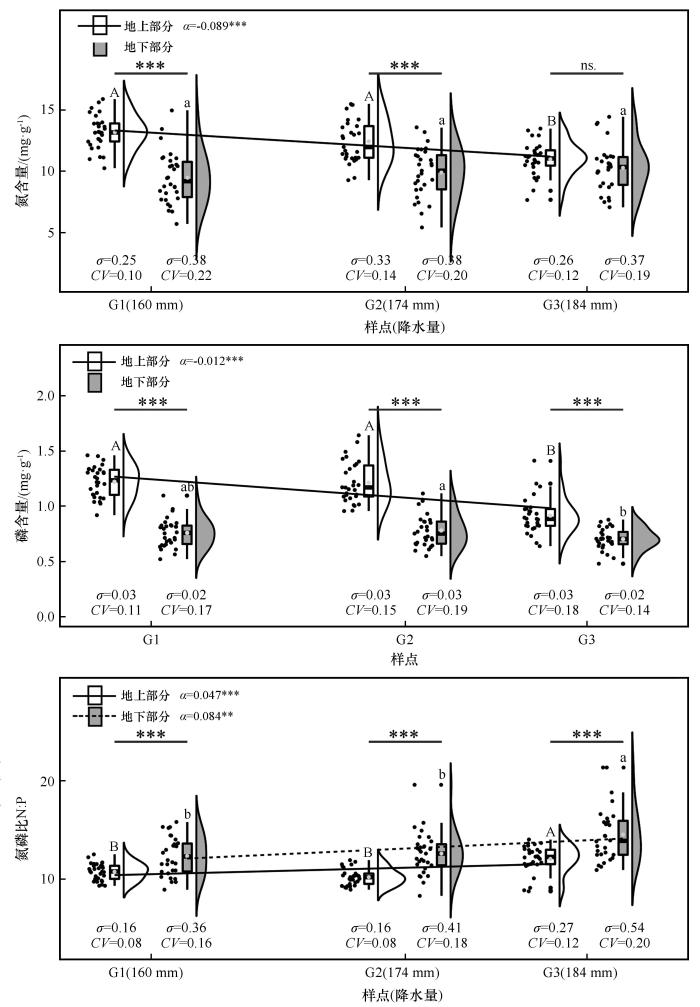
... 植物叶片的N、P元素含量及其N∶P可以用来判断植物生长过程中是否受到N、P元素的限制.本研究测得齿肋赤藓植物地上部分N、P元素含量及N∶P的均值分别为12.2±0.19、1.13±0.22 mg·g-1及10.96±0.14 mg·g-1,梅月磊等[27]在对沙漠齿肋赤藓的模拟氮添加试验中采用如下两种判断方式考量齿肋赤藓是否受到养分限制.①根据Koerselman等[3]的标准,当N∶P小于14时,植物受到氮限制.②根据Ellison[28]的标准,当N含量小于20 mg·g-1、P含量大于 1 mg·g-1时,植物受氮限制.参照上述标准推测齿肋赤藓在该沙漠的生长易受到氮限制,与梅月磊等[27]所得结果一致.齿肋赤藓植物地上部分的N、P元素含量低于古尔班通古特沙漠草本植物叶片N、P含量(36.78、2.13 mg·g-1)[29];低于中国北方典型荒漠及荒漠化地区维管植物叶片N、P含量(24.45±8.1、1.74±0.88 mg·g-1)[30]和全国植被N、P含量的平均水平(18.6、1.21 mg·g-1)[31].养分库差异和自生养分运输能力限制是造成维管植物(叶片)和苔藓植物地上部分养分差异的原因[31].维管植物含有较发达的根系,这些植物具有较强的N、P元素吸收、存储和运输能力,而藓类植物没有真正的根系,假根对水分和养分的吸收可能来自于虹吸作用,并不能大范围、高效地吸收和储存土壤中的养分[32].本研究测得齿肋赤藓植物地上部分N、P元素含量高于Shi等[33]在青藏高原高海拔地区所测得结果(N: 9.33 mg·g-1);与吴盼盼等[34]在北半球中高纬度泥炭沼泽所得泥炭藓结果接近(11.48、1.16 mg·g-1),但与非泥炭藓结果呈现差异(9.73、1.5 mg·g-1),它们的养分差异体现了藓类植物对于不同养分来源、环境、海拔、季节等因素的适应性变化.各样点的齿肋赤藓N、P含量均表现出地上部分显著高于地下部分(图1),研究结果与Li等[20]、梅月磊等[27]测得结果一致.造成这一结果的原因是地上部分主要由具有光合能力的绿色植株组成,而地下部分主要是由假根、被沙埋后所产生的植物组织残骸组成,生理活性相较于地上部分低.齿肋赤藓植物地上部分的N和N∶P比的变异系数(CV)明显低于地下部分(P<0.05),元素可塑性假说[35]认为,植物地下部分的化学计量特征比植物地上部分茎叶的化学计量特征有更高的变异性,主要由于植物地下部分所处环境较地上部分更为复杂导致.在恶劣的环境条件下,沙漠中的植物往往会为了生存而扩大根系,而不是支持地上部分的生长[36],非维管束植物齿肋赤藓则需要通过假根中菌丝体的生长提高其对土壤中水分和养分的吸收[37],菌丝体的发育状况和土壤可利用养分的差异导致齿肋赤藓地下部分的养分特征具有较高的变异性. ...
采伐剩余物管理对杉木人工林土壤和叶片δ 15N的长期效应
1
2014
... 氮(N)和磷(P)是影响植物生长发育的主要营养元素[1].N、P元素是构成有机体内蛋白质、磷酸等物质的基础,对调节植物生长和代谢具有重要作用[2].通常认为氮磷比(N∶P)是判断植物生长发育是否受N、P元素的限制,因此对植物N、P元素化学计量特征的研究有助于加深对植物生长和环境适应性的理解,进而促进对陆地生态系统养分调控和元素生物地球化学循环机制的深入理解[3].N∶P可以衡量植物生长对于N、P的限制程度,N、P含量较低的植物会倾向于在氮限制或磷限制的环境中成为优势种;传统的氮磷限制假说认为,当植物叶片N∶P小于14时,植物生长受氮限制;当N∶P大于16时,植物生长受磷元素所限制;当N∶P为14~16时,植物生长可能同时受或不受氮磷元素限制[4-6],但也有诸多研究认为不同的功能群植物受氮磷限制的临界氮磷比值存在差异[7],如常绿针叶林、豆科植物及草本植物的临界值分别为10、8~9、12.尽管氮磷限制假说饱受争议,但其依然广泛运用于荒漠[8-9]、草原[10]、森林[11]等多种生态系统中用于推断植物、土壤的养分限制.在养分受限的环境中,植物养分的分配对植物资源的吸收和利用起着关键作用,并与植物生长速率、繁殖、防御和适应环境异质性的策略有关.此外,植物的地上和地下部分直接相互依赖,与捕获和利用光、养分和水资源的吸收和利用密切相关[12],因此将植物器官间营养元素可塑性作为植物生长和环境适应性的重要研究内容.通常使用变异系数(CV)来描述植物器官间元素变异性或可塑性[13],由于植物不同器官所面临的环境不同,不同器官的适应能力可能不同.可塑性高的生物具有更大的生态幅度和适应性,不同器官间元素分配不同,可塑性不同,功能不同,因此植物不同器官间往往存在养分的幂指数关系. ...
中国典型生态脆弱区生态化学计量学研究进展
0
2021
北亚热带常绿阔叶林乔灌草物种的碳氮磷生态化学计量比
1
2019
... 氮(N)和磷(P)是影响植物生长发育的主要营养元素[1].N、P元素是构成有机体内蛋白质、磷酸等物质的基础,对调节植物生长和代谢具有重要作用[2].通常认为氮磷比(N∶P)是判断植物生长发育是否受N、P元素的限制,因此对植物N、P元素化学计量特征的研究有助于加深对植物生长和环境适应性的理解,进而促进对陆地生态系统养分调控和元素生物地球化学循环机制的深入理解[3].N∶P可以衡量植物生长对于N、P的限制程度,N、P含量较低的植物会倾向于在氮限制或磷限制的环境中成为优势种;传统的氮磷限制假说认为,当植物叶片N∶P小于14时,植物生长受氮限制;当N∶P大于16时,植物生长受磷元素所限制;当N∶P为14~16时,植物生长可能同时受或不受氮磷元素限制[4-6],但也有诸多研究认为不同的功能群植物受氮磷限制的临界氮磷比值存在差异[7],如常绿针叶林、豆科植物及草本植物的临界值分别为10、8~9、12.尽管氮磷限制假说饱受争议,但其依然广泛运用于荒漠[8-9]、草原[10]、森林[11]等多种生态系统中用于推断植物、土壤的养分限制.在养分受限的环境中,植物养分的分配对植物资源的吸收和利用起着关键作用,并与植物生长速率、繁殖、防御和适应环境异质性的策略有关.此外,植物的地上和地下部分直接相互依赖,与捕获和利用光、养分和水资源的吸收和利用密切相关[12],因此将植物器官间营养元素可塑性作为植物生长和环境适应性的重要研究内容.通常使用变异系数(CV)来描述植物器官间元素变异性或可塑性[13],由于植物不同器官所面临的环境不同,不同器官的适应能力可能不同.可塑性高的生物具有更大的生态幅度和适应性,不同器官间元素分配不同,可塑性不同,功能不同,因此植物不同器官间往往存在养分的幂指数关系. ...
植物生态化学计量特征及其主要假说
1
2021
... 氮(N)和磷(P)是影响植物生长发育的主要营养元素[1].N、P元素是构成有机体内蛋白质、磷酸等物质的基础,对调节植物生长和代谢具有重要作用[2].通常认为氮磷比(N∶P)是判断植物生长发育是否受N、P元素的限制,因此对植物N、P元素化学计量特征的研究有助于加深对植物生长和环境适应性的理解,进而促进对陆地生态系统养分调控和元素生物地球化学循环机制的深入理解[3].N∶P可以衡量植物生长对于N、P的限制程度,N、P含量较低的植物会倾向于在氮限制或磷限制的环境中成为优势种;传统的氮磷限制假说认为,当植物叶片N∶P小于14时,植物生长受氮限制;当N∶P大于16时,植物生长受磷元素所限制;当N∶P为14~16时,植物生长可能同时受或不受氮磷元素限制[4-6],但也有诸多研究认为不同的功能群植物受氮磷限制的临界氮磷比值存在差异[7],如常绿针叶林、豆科植物及草本植物的临界值分别为10、8~9、12.尽管氮磷限制假说饱受争议,但其依然广泛运用于荒漠[8-9]、草原[10]、森林[11]等多种生态系统中用于推断植物、土壤的养分限制.在养分受限的环境中,植物养分的分配对植物资源的吸收和利用起着关键作用,并与植物生长速率、繁殖、防御和适应环境异质性的策略有关.此外,植物的地上和地下部分直接相互依赖,与捕获和利用光、养分和水资源的吸收和利用密切相关[12],因此将植物器官间营养元素可塑性作为植物生长和环境适应性的重要研究内容.通常使用变异系数(CV)来描述植物器官间元素变异性或可塑性[13],由于植物不同器官所面临的环境不同,不同器官的适应能力可能不同.可塑性高的生物具有更大的生态幅度和适应性,不同器官间元素分配不同,可塑性不同,功能不同,因此植物不同器官间往往存在养分的幂指数关系. ...
Community characteristics and leaf stoichiometric traits of desert ecosystems regulated by precipitation and soil in an arid area of China
1
2018
... 氮(N)和磷(P)是影响植物生长发育的主要营养元素[1].N、P元素是构成有机体内蛋白质、磷酸等物质的基础,对调节植物生长和代谢具有重要作用[2].通常认为氮磷比(N∶P)是判断植物生长发育是否受N、P元素的限制,因此对植物N、P元素化学计量特征的研究有助于加深对植物生长和环境适应性的理解,进而促进对陆地生态系统养分调控和元素生物地球化学循环机制的深入理解[3].N∶P可以衡量植物生长对于N、P的限制程度,N、P含量较低的植物会倾向于在氮限制或磷限制的环境中成为优势种;传统的氮磷限制假说认为,当植物叶片N∶P小于14时,植物生长受氮限制;当N∶P大于16时,植物生长受磷元素所限制;当N∶P为14~16时,植物生长可能同时受或不受氮磷元素限制[4-6],但也有诸多研究认为不同的功能群植物受氮磷限制的临界氮磷比值存在差异[7],如常绿针叶林、豆科植物及草本植物的临界值分别为10、8~9、12.尽管氮磷限制假说饱受争议,但其依然广泛运用于荒漠[8-9]、草原[10]、森林[11]等多种生态系统中用于推断植物、土壤的养分限制.在养分受限的环境中,植物养分的分配对植物资源的吸收和利用起着关键作用,并与植物生长速率、繁殖、防御和适应环境异质性的策略有关.此外,植物的地上和地下部分直接相互依赖,与捕获和利用光、养分和水资源的吸收和利用密切相关[12],因此将植物器官间营养元素可塑性作为植物生长和环境适应性的重要研究内容.通常使用变异系数(CV)来描述植物器官间元素变异性或可塑性[13],由于植物不同器官所面临的环境不同,不同器官的适应能力可能不同.可塑性高的生物具有更大的生态幅度和适应性,不同器官间元素分配不同,可塑性不同,功能不同,因此植物不同器官间往往存在养分的幂指数关系. ...
新疆67种荒漠植物叶碳氮磷计量特征及其与气候的关系
1
2019
... 氮(N)和磷(P)是影响植物生长发育的主要营养元素[1].N、P元素是构成有机体内蛋白质、磷酸等物质的基础,对调节植物生长和代谢具有重要作用[2].通常认为氮磷比(N∶P)是判断植物生长发育是否受N、P元素的限制,因此对植物N、P元素化学计量特征的研究有助于加深对植物生长和环境适应性的理解,进而促进对陆地生态系统养分调控和元素生物地球化学循环机制的深入理解[3].N∶P可以衡量植物生长对于N、P的限制程度,N、P含量较低的植物会倾向于在氮限制或磷限制的环境中成为优势种;传统的氮磷限制假说认为,当植物叶片N∶P小于14时,植物生长受氮限制;当N∶P大于16时,植物生长受磷元素所限制;当N∶P为14~16时,植物生长可能同时受或不受氮磷元素限制[4-6],但也有诸多研究认为不同的功能群植物受氮磷限制的临界氮磷比值存在差异[7],如常绿针叶林、豆科植物及草本植物的临界值分别为10、8~9、12.尽管氮磷限制假说饱受争议,但其依然广泛运用于荒漠[8-9]、草原[10]、森林[11]等多种生态系统中用于推断植物、土壤的养分限制.在养分受限的环境中,植物养分的分配对植物资源的吸收和利用起着关键作用,并与植物生长速率、繁殖、防御和适应环境异质性的策略有关.此外,植物的地上和地下部分直接相互依赖,与捕获和利用光、养分和水资源的吸收和利用密切相关[12],因此将植物器官间营养元素可塑性作为植物生长和环境适应性的重要研究内容.通常使用变异系数(CV)来描述植物器官间元素变异性或可塑性[13],由于植物不同器官所面临的环境不同,不同器官的适应能力可能不同.可塑性高的生物具有更大的生态幅度和适应性,不同器官间元素分配不同,可塑性不同,功能不同,因此植物不同器官间往往存在养分的幂指数关系. ...
荒漠草原不同雨量带土壤-植物-微生物C、N、P及其化学计量特征
1
2020
... 氮(N)和磷(P)是影响植物生长发育的主要营养元素[1].N、P元素是构成有机体内蛋白质、磷酸等物质的基础,对调节植物生长和代谢具有重要作用[2].通常认为氮磷比(N∶P)是判断植物生长发育是否受N、P元素的限制,因此对植物N、P元素化学计量特征的研究有助于加深对植物生长和环境适应性的理解,进而促进对陆地生态系统养分调控和元素生物地球化学循环机制的深入理解[3].N∶P可以衡量植物生长对于N、P的限制程度,N、P含量较低的植物会倾向于在氮限制或磷限制的环境中成为优势种;传统的氮磷限制假说认为,当植物叶片N∶P小于14时,植物生长受氮限制;当N∶P大于16时,植物生长受磷元素所限制;当N∶P为14~16时,植物生长可能同时受或不受氮磷元素限制[4-6],但也有诸多研究认为不同的功能群植物受氮磷限制的临界氮磷比值存在差异[7],如常绿针叶林、豆科植物及草本植物的临界值分别为10、8~9、12.尽管氮磷限制假说饱受争议,但其依然广泛运用于荒漠[8-9]、草原[10]、森林[11]等多种生态系统中用于推断植物、土壤的养分限制.在养分受限的环境中,植物养分的分配对植物资源的吸收和利用起着关键作用,并与植物生长速率、繁殖、防御和适应环境异质性的策略有关.此外,植物的地上和地下部分直接相互依赖,与捕获和利用光、养分和水资源的吸收和利用密切相关[12],因此将植物器官间营养元素可塑性作为植物生长和环境适应性的重要研究内容.通常使用变异系数(CV)来描述植物器官间元素变异性或可塑性[13],由于植物不同器官所面临的环境不同,不同器官的适应能力可能不同.可塑性高的生物具有更大的生态幅度和适应性,不同器官间元素分配不同,可塑性不同,功能不同,因此植物不同器官间往往存在养分的幂指数关系. ...
C∶N∶P stoichiometry in China's forests:from organs to ecosystems
1
2018
... 氮(N)和磷(P)是影响植物生长发育的主要营养元素[1].N、P元素是构成有机体内蛋白质、磷酸等物质的基础,对调节植物生长和代谢具有重要作用[2].通常认为氮磷比(N∶P)是判断植物生长发育是否受N、P元素的限制,因此对植物N、P元素化学计量特征的研究有助于加深对植物生长和环境适应性的理解,进而促进对陆地生态系统养分调控和元素生物地球化学循环机制的深入理解[3].N∶P可以衡量植物生长对于N、P的限制程度,N、P含量较低的植物会倾向于在氮限制或磷限制的环境中成为优势种;传统的氮磷限制假说认为,当植物叶片N∶P小于14时,植物生长受氮限制;当N∶P大于16时,植物生长受磷元素所限制;当N∶P为14~16时,植物生长可能同时受或不受氮磷元素限制[4-6],但也有诸多研究认为不同的功能群植物受氮磷限制的临界氮磷比值存在差异[7],如常绿针叶林、豆科植物及草本植物的临界值分别为10、8~9、12.尽管氮磷限制假说饱受争议,但其依然广泛运用于荒漠[8-9]、草原[10]、森林[11]等多种生态系统中用于推断植物、土壤的养分限制.在养分受限的环境中,植物养分的分配对植物资源的吸收和利用起着关键作用,并与植物生长速率、繁殖、防御和适应环境异质性的策略有关.此外,植物的地上和地下部分直接相互依赖,与捕获和利用光、养分和水资源的吸收和利用密切相关[12],因此将植物器官间营养元素可塑性作为植物生长和环境适应性的重要研究内容.通常使用变异系数(CV)来描述植物器官间元素变异性或可塑性[13],由于植物不同器官所面临的环境不同,不同器官的适应能力可能不同.可塑性高的生物具有更大的生态幅度和适应性,不同器官间元素分配不同,可塑性不同,功能不同,因此植物不同器官间往往存在养分的幂指数关系. ...
Allometric scaling of biomass with nitrogen and phosphorus above-and below-ground in herbaceous plants varies along water-salinity gradients
1
2021
... 氮(N)和磷(P)是影响植物生长发育的主要营养元素[1].N、P元素是构成有机体内蛋白质、磷酸等物质的基础,对调节植物生长和代谢具有重要作用[2].通常认为氮磷比(N∶P)是判断植物生长发育是否受N、P元素的限制,因此对植物N、P元素化学计量特征的研究有助于加深对植物生长和环境适应性的理解,进而促进对陆地生态系统养分调控和元素生物地球化学循环机制的深入理解[3].N∶P可以衡量植物生长对于N、P的限制程度,N、P含量较低的植物会倾向于在氮限制或磷限制的环境中成为优势种;传统的氮磷限制假说认为,当植物叶片N∶P小于14时,植物生长受氮限制;当N∶P大于16时,植物生长受磷元素所限制;当N∶P为14~16时,植物生长可能同时受或不受氮磷元素限制[4-6],但也有诸多研究认为不同的功能群植物受氮磷限制的临界氮磷比值存在差异[7],如常绿针叶林、豆科植物及草本植物的临界值分别为10、8~9、12.尽管氮磷限制假说饱受争议,但其依然广泛运用于荒漠[8-9]、草原[10]、森林[11]等多种生态系统中用于推断植物、土壤的养分限制.在养分受限的环境中,植物养分的分配对植物资源的吸收和利用起着关键作用,并与植物生长速率、繁殖、防御和适应环境异质性的策略有关.此外,植物的地上和地下部分直接相互依赖,与捕获和利用光、养分和水资源的吸收和利用密切相关[12],因此将植物器官间营养元素可塑性作为植物生长和环境适应性的重要研究内容.通常使用变异系数(CV)来描述植物器官间元素变异性或可塑性[13],由于植物不同器官所面临的环境不同,不同器官的适应能力可能不同.可塑性高的生物具有更大的生态幅度和适应性,不同器官间元素分配不同,可塑性不同,功能不同,因此植物不同器官间往往存在养分的幂指数关系. ...
Quantitative estimation of phenotypic plasticity:bridging the gap between the evolutionary concept and its ecological applications
1
2006
... 氮(N)和磷(P)是影响植物生长发育的主要营养元素[1].N、P元素是构成有机体内蛋白质、磷酸等物质的基础,对调节植物生长和代谢具有重要作用[2].通常认为氮磷比(N∶P)是判断植物生长发育是否受N、P元素的限制,因此对植物N、P元素化学计量特征的研究有助于加深对植物生长和环境适应性的理解,进而促进对陆地生态系统养分调控和元素生物地球化学循环机制的深入理解[3].N∶P可以衡量植物生长对于N、P的限制程度,N、P含量较低的植物会倾向于在氮限制或磷限制的环境中成为优势种;传统的氮磷限制假说认为,当植物叶片N∶P小于14时,植物生长受氮限制;当N∶P大于16时,植物生长受磷元素所限制;当N∶P为14~16时,植物生长可能同时受或不受氮磷元素限制[4-6],但也有诸多研究认为不同的功能群植物受氮磷限制的临界氮磷比值存在差异[7],如常绿针叶林、豆科植物及草本植物的临界值分别为10、8~9、12.尽管氮磷限制假说饱受争议,但其依然广泛运用于荒漠[8-9]、草原[10]、森林[11]等多种生态系统中用于推断植物、土壤的养分限制.在养分受限的环境中,植物养分的分配对植物资源的吸收和利用起着关键作用,并与植物生长速率、繁殖、防御和适应环境异质性的策略有关.此外,植物的地上和地下部分直接相互依赖,与捕获和利用光、养分和水资源的吸收和利用密切相关[12],因此将植物器官间营养元素可塑性作为植物生长和环境适应性的重要研究内容.通常使用变异系数(CV)来描述植物器官间元素变异性或可塑性[13],由于植物不同器官所面临的环境不同,不同器官的适应能力可能不同.可塑性高的生物具有更大的生态幅度和适应性,不同器官间元素分配不同,可塑性不同,功能不同,因此植物不同器官间往往存在养分的幂指数关系. ...
Invariant allometric scaling of nitrogen and phosphorus in leaves,stems,and fine roots of woody plants along an altitudinal gradient
2
2016
... 养分的幂指数关系和营养元素化学计量动态平衡关系为研究植物器官间营养元素分配提供了解决方向,用于解释植物体资源分配的一般原则或分配规则.不同植物器官中N、P元素含量的变化常采用幂指数关系表示,其中常见的N-P幂指数显著小于1[14-16];Reich等[15]发现,在全球范围内,叶片N-P幂指数约为2/3;Niklas等[16]发现发育中的叶片中N-P幂指数约为3/4,且在植物体经历从初级生长到次级生长的过渡时,N-P幂指数会发生转变;Tian等[17]认为,植物叶片的N-P幂指数与全球范围内的温度、降水和植物功能类型等多种因素有关.其中,气候可以通过调节土壤中的养分条件来影响植物中的N、P元素含量.降水量的变化可以通过控制凋落物输入、分解和淋溶等过程来影响土壤N和P的有效性[18],进而影响植物营养元素的吸收和利用,最终引起植物生态化学计量特征的改变.近年来,国内外展开了大量有关降水变化对植物养分吸收和利用的研究.王珊等[19]发现在荒漠生态系统中,植物受水分胁迫的条件下均会表现出碳素积累、氮磷元素限制的格局,对N、P元素的利用不活跃.在荒漠生态系统中,藓类植物较维管束植物对荒漠降水更为敏感,因此在荒漠生态系统中藓类植物的养分吸收与利用是否更易受降水变化的影响是一个值得探索的问题. ...
... 利用植物N和P浓度的对数转换值使用模型Ⅱ回归估计N-P幂指数关系中的和参数.N-P幂指数关系的参数估计是通过使用R的“lmodel2”包中的“lmodel2”函数实现的.我们采用似然比检验(likelihood ratio test, LRT)成对比较各幂指数函数之间是否存在差异.似然比检验是通过使用R的“smatr”包中的“sma”函数实现[14]. ...
Evidence of a general 2/3-power law of scaling leaf nitrogen to phosphorus among major plant groups and biomes
2
2010
... 养分的幂指数关系和营养元素化学计量动态平衡关系为研究植物器官间营养元素分配提供了解决方向,用于解释植物体资源分配的一般原则或分配规则.不同植物器官中N、P元素含量的变化常采用幂指数关系表示,其中常见的N-P幂指数显著小于1[14-16];Reich等[15]发现,在全球范围内,叶片N-P幂指数约为2/3;Niklas等[16]发现发育中的叶片中N-P幂指数约为3/4,且在植物体经历从初级生长到次级生长的过渡时,N-P幂指数会发生转变;Tian等[17]认为,植物叶片的N-P幂指数与全球范围内的温度、降水和植物功能类型等多种因素有关.其中,气候可以通过调节土壤中的养分条件来影响植物中的N、P元素含量.降水量的变化可以通过控制凋落物输入、分解和淋溶等过程来影响土壤N和P的有效性[18],进而影响植物营养元素的吸收和利用,最终引起植物生态化学计量特征的改变.近年来,国内外展开了大量有关降水变化对植物养分吸收和利用的研究.王珊等[19]发现在荒漠生态系统中,植物受水分胁迫的条件下均会表现出碳素积累、氮磷元素限制的格局,对N、P元素的利用不活跃.在荒漠生态系统中,藓类植物较维管束植物对荒漠降水更为敏感,因此在荒漠生态系统中藓类植物的养分吸收与利用是否更易受降水变化的影响是一个值得探索的问题. ...
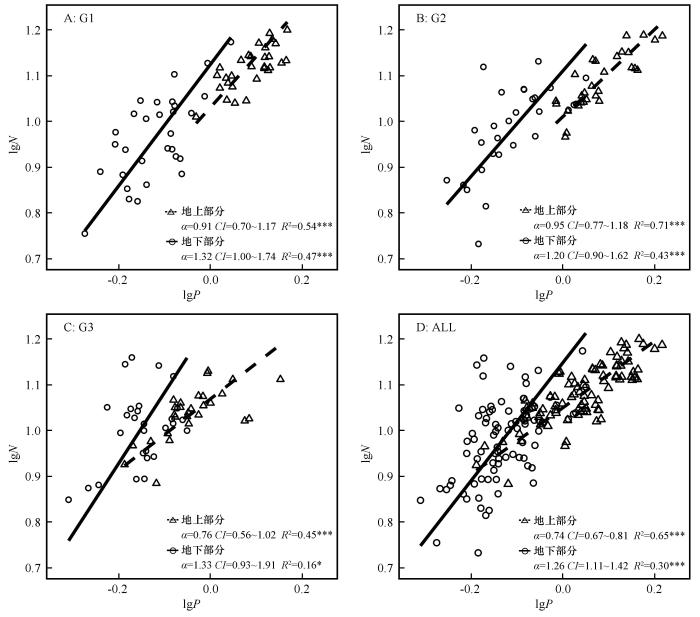
... 通过对各样点地上、地下部分的相关性分析发现:无论是地上部分还是地下部分,齿肋赤藓N、P元素含量均为显著正相关,这种相关性反映了植物体内营养元素间具有较强的线性关系[40],本文结果进一步验证了此观点.限制元素N、P呈异速关系是植物适应环境的重要策略[41-42],齿肋赤藓同部位N、P含量均表现出显著的异速关系,说明齿肋赤藓各部位N、P比例相对稳定.在众多大尺度的调查、养分添加的研究中,所得N和P的幂指数比均小于1,且斜率间无显著差异,齿肋赤藓植物养分吸收、利用和分配关系随环境改变呈现不显著的差异性变化,具有一定的保守性[35].本研究中齿肋赤藓植物地上部分N-P幂指数计量比均小于1,其结果均接近前人所提出的植物叶片2/3[15]或3/4[16]N-P幂指数计量关系;齿肋赤藓植物地下部分N-P幂指数计量关系均大于1,说明齿肋赤藓植物地下部分P含量占比相较于N含量高,这可能与藓类植物能大量吸收土壤P元素的特性有关[43]. ...
Nitrogen/phosphorus leaf stoichiometry and the scaling of plant growth
3
2005
... 养分的幂指数关系和营养元素化学计量动态平衡关系为研究植物器官间营养元素分配提供了解决方向,用于解释植物体资源分配的一般原则或分配规则.不同植物器官中N、P元素含量的变化常采用幂指数关系表示,其中常见的N-P幂指数显著小于1[14-16];Reich等[15]发现,在全球范围内,叶片N-P幂指数约为2/3;Niklas等[16]发现发育中的叶片中N-P幂指数约为3/4,且在植物体经历从初级生长到次级生长的过渡时,N-P幂指数会发生转变;Tian等[17]认为,植物叶片的N-P幂指数与全球范围内的温度、降水和植物功能类型等多种因素有关.其中,气候可以通过调节土壤中的养分条件来影响植物中的N、P元素含量.降水量的变化可以通过控制凋落物输入、分解和淋溶等过程来影响土壤N和P的有效性[18],进而影响植物营养元素的吸收和利用,最终引起植物生态化学计量特征的改变.近年来,国内外展开了大量有关降水变化对植物养分吸收和利用的研究.王珊等[19]发现在荒漠生态系统中,植物受水分胁迫的条件下均会表现出碳素积累、氮磷元素限制的格局,对N、P元素的利用不活跃.在荒漠生态系统中,藓类植物较维管束植物对荒漠降水更为敏感,因此在荒漠生态系统中藓类植物的养分吸收与利用是否更易受降水变化的影响是一个值得探索的问题. ...
... [16]发现发育中的叶片中N-P幂指数约为3/4,且在植物体经历从初级生长到次级生长的过渡时,N-P幂指数会发生转变;Tian等[17]认为,植物叶片的N-P幂指数与全球范围内的温度、降水和植物功能类型等多种因素有关.其中,气候可以通过调节土壤中的养分条件来影响植物中的N、P元素含量.降水量的变化可以通过控制凋落物输入、分解和淋溶等过程来影响土壤N和P的有效性[18],进而影响植物营养元素的吸收和利用,最终引起植物生态化学计量特征的改变.近年来,国内外展开了大量有关降水变化对植物养分吸收和利用的研究.王珊等[19]发现在荒漠生态系统中,植物受水分胁迫的条件下均会表现出碳素积累、氮磷元素限制的格局,对N、P元素的利用不活跃.在荒漠生态系统中,藓类植物较维管束植物对荒漠降水更为敏感,因此在荒漠生态系统中藓类植物的养分吸收与利用是否更易受降水变化的影响是一个值得探索的问题. ...
... 通过对各样点地上、地下部分的相关性分析发现:无论是地上部分还是地下部分,齿肋赤藓N、P元素含量均为显著正相关,这种相关性反映了植物体内营养元素间具有较强的线性关系[40],本文结果进一步验证了此观点.限制元素N、P呈异速关系是植物适应环境的重要策略[41-42],齿肋赤藓同部位N、P含量均表现出显著的异速关系,说明齿肋赤藓各部位N、P比例相对稳定.在众多大尺度的调查、养分添加的研究中,所得N和P的幂指数比均小于1,且斜率间无显著差异,齿肋赤藓植物养分吸收、利用和分配关系随环境改变呈现不显著的差异性变化,具有一定的保守性[35].本研究中齿肋赤藓植物地上部分N-P幂指数计量比均小于1,其结果均接近前人所提出的植物叶片2/3[15]或3/4[16]N-P幂指数计量关系;齿肋赤藓植物地下部分N-P幂指数计量关系均大于1,说明齿肋赤藓植物地下部分P含量占比相较于N含量高,这可能与藓类植物能大量吸收土壤P元素的特性有关[43]. ...
Global leaf nitrogen and phosphorus stoichiometry and their scaling exponent
1
2018
... 养分的幂指数关系和营养元素化学计量动态平衡关系为研究植物器官间营养元素分配提供了解决方向,用于解释植物体资源分配的一般原则或分配规则.不同植物器官中N、P元素含量的变化常采用幂指数关系表示,其中常见的N-P幂指数显著小于1[14-16];Reich等[15]发现,在全球范围内,叶片N-P幂指数约为2/3;Niklas等[16]发现发育中的叶片中N-P幂指数约为3/4,且在植物体经历从初级生长到次级生长的过渡时,N-P幂指数会发生转变;Tian等[17]认为,植物叶片的N-P幂指数与全球范围内的温度、降水和植物功能类型等多种因素有关.其中,气候可以通过调节土壤中的养分条件来影响植物中的N、P元素含量.降水量的变化可以通过控制凋落物输入、分解和淋溶等过程来影响土壤N和P的有效性[18],进而影响植物营养元素的吸收和利用,最终引起植物生态化学计量特征的改变.近年来,国内外展开了大量有关降水变化对植物养分吸收和利用的研究.王珊等[19]发现在荒漠生态系统中,植物受水分胁迫的条件下均会表现出碳素积累、氮磷元素限制的格局,对N、P元素的利用不活跃.在荒漠生态系统中,藓类植物较维管束植物对荒漠降水更为敏感,因此在荒漠生态系统中藓类植物的养分吸收与利用是否更易受降水变化的影响是一个值得探索的问题. ...
Decoupling of nitrogen and phosphorus in terrestrial plants associated with global changes
1
2015
... 养分的幂指数关系和营养元素化学计量动态平衡关系为研究植物器官间营养元素分配提供了解决方向,用于解释植物体资源分配的一般原则或分配规则.不同植物器官中N、P元素含量的变化常采用幂指数关系表示,其中常见的N-P幂指数显著小于1[14-16];Reich等[15]发现,在全球范围内,叶片N-P幂指数约为2/3;Niklas等[16]发现发育中的叶片中N-P幂指数约为3/4,且在植物体经历从初级生长到次级生长的过渡时,N-P幂指数会发生转变;Tian等[17]认为,植物叶片的N-P幂指数与全球范围内的温度、降水和植物功能类型等多种因素有关.其中,气候可以通过调节土壤中的养分条件来影响植物中的N、P元素含量.降水量的变化可以通过控制凋落物输入、分解和淋溶等过程来影响土壤N和P的有效性[18],进而影响植物营养元素的吸收和利用,最终引起植物生态化学计量特征的改变.近年来,国内外展开了大量有关降水变化对植物养分吸收和利用的研究.王珊等[19]发现在荒漠生态系统中,植物受水分胁迫的条件下均会表现出碳素积累、氮磷元素限制的格局,对N、P元素的利用不活跃.在荒漠生态系统中,藓类植物较维管束植物对荒漠降水更为敏感,因此在荒漠生态系统中藓类植物的养分吸收与利用是否更易受降水变化的影响是一个值得探索的问题. ...
降水变化对红砂-珍珠碳、氮、磷化学计量特征的影响
1
2020
... 养分的幂指数关系和营养元素化学计量动态平衡关系为研究植物器官间营养元素分配提供了解决方向,用于解释植物体资源分配的一般原则或分配规则.不同植物器官中N、P元素含量的变化常采用幂指数关系表示,其中常见的N-P幂指数显著小于1[14-16];Reich等[15]发现,在全球范围内,叶片N-P幂指数约为2/3;Niklas等[16]发现发育中的叶片中N-P幂指数约为3/4,且在植物体经历从初级生长到次级生长的过渡时,N-P幂指数会发生转变;Tian等[17]认为,植物叶片的N-P幂指数与全球范围内的温度、降水和植物功能类型等多种因素有关.其中,气候可以通过调节土壤中的养分条件来影响植物中的N、P元素含量.降水量的变化可以通过控制凋落物输入、分解和淋溶等过程来影响土壤N和P的有效性[18],进而影响植物营养元素的吸收和利用,最终引起植物生态化学计量特征的改变.近年来,国内外展开了大量有关降水变化对植物养分吸收和利用的研究.王珊等[19]发现在荒漠生态系统中,植物受水分胁迫的条件下均会表现出碳素积累、氮磷元素限制的格局,对N、P元素的利用不活跃.在荒漠生态系统中,藓类植物较维管束植物对荒漠降水更为敏感,因此在荒漠生态系统中藓类植物的养分吸收与利用是否更易受降水变化的影响是一个值得探索的问题. ...
Moss patch size and microhabitats influence stoichiometry of moss crusts in a temperate desert,Central Asia
2
2019
... 荒漠藓类植物对荒漠地表的稳定、水分分配、养分循环、能量平衡、抗风蚀和水蚀能力以及土壤微环境的改善具有重要作用,是干旱区土壤生物结皮的重要组成部分.荒漠藓类植物是典型的变水植物,干旱缺水时通常处于休眠状态,当环境适宜时,可以快速恢复光合生理活性,说明荒漠藓类植物对环境水分具有较强的敏感性.前期的研究发现荒漠藓类植物易受到微环境的影响,表现为灌丛下藓类植物具有较高的N、P含量等特征,主要由于灌丛的存在能够为藓类植物遮光、提供水分和养分等有利条件[20].在古尔班通古特沙漠中由东南到西北年降水量呈降低趋势,因此在该沙漠中藓类植物化学计量特征是否随年降水量的变化而变化?藓类植物化学计量特征是否能够响应该沙漠年降水量的变化?通过测定和比较不同降水背景下齿肋赤藓地上部分与地下部分N、P元素含量、N∶P间以及N-P幂指数间差异,阐明不同降水背景下荒漠藓类植物化学计量特征间差异以及其对环境适应性策略,有助于进一步加深荒漠藓类植物对荒漠水分的敏感性以及对荒漠不同降水环境的适应性特征的理解,为未来全球气候变化背景下预测荒漠藓类植物生长发育特征提供科学理论支持,同时为荒漠藓类植物在荒漠生态系统中拓植奠定科学数据基础. ...
... 植物叶片的N、P元素含量及其N∶P可以用来判断植物生长过程中是否受到N、P元素的限制.本研究测得齿肋赤藓植物地上部分N、P元素含量及N∶P的均值分别为12.2±0.19、1.13±0.22 mg·g-1及10.96±0.14 mg·g-1,梅月磊等[27]在对沙漠齿肋赤藓的模拟氮添加试验中采用如下两种判断方式考量齿肋赤藓是否受到养分限制.①根据Koerselman等[3]的标准,当N∶P小于14时,植物受到氮限制.②根据Ellison[28]的标准,当N含量小于20 mg·g-1、P含量大于 1 mg·g-1时,植物受氮限制.参照上述标准推测齿肋赤藓在该沙漠的生长易受到氮限制,与梅月磊等[27]所得结果一致.齿肋赤藓植物地上部分的N、P元素含量低于古尔班通古特沙漠草本植物叶片N、P含量(36.78、2.13 mg·g-1)[29];低于中国北方典型荒漠及荒漠化地区维管植物叶片N、P含量(24.45±8.1、1.74±0.88 mg·g-1)[30]和全国植被N、P含量的平均水平(18.6、1.21 mg·g-1)[31].养分库差异和自生养分运输能力限制是造成维管植物(叶片)和苔藓植物地上部分养分差异的原因[31].维管植物含有较发达的根系,这些植物具有较强的N、P元素吸收、存储和运输能力,而藓类植物没有真正的根系,假根对水分和养分的吸收可能来自于虹吸作用,并不能大范围、高效地吸收和储存土壤中的养分[32].本研究测得齿肋赤藓植物地上部分N、P元素含量高于Shi等[33]在青藏高原高海拔地区所测得结果(N: 9.33 mg·g-1);与吴盼盼等[34]在北半球中高纬度泥炭沼泽所得泥炭藓结果接近(11.48、1.16 mg·g-1),但与非泥炭藓结果呈现差异(9.73、1.5 mg·g-1),它们的养分差异体现了藓类植物对于不同养分来源、环境、海拔、季节等因素的适应性变化.各样点的齿肋赤藓N、P含量均表现出地上部分显著高于地下部分(图1),研究结果与Li等[20]、梅月磊等[27]测得结果一致.造成这一结果的原因是地上部分主要由具有光合能力的绿色植株组成,而地下部分主要是由假根、被沙埋后所产生的植物组织残骸组成,生理活性相较于地上部分低.齿肋赤藓植物地上部分的N和N∶P比的变异系数(CV)明显低于地下部分(P<0.05),元素可塑性假说[35]认为,植物地下部分的化学计量特征比植物地上部分茎叶的化学计量特征有更高的变异性,主要由于植物地下部分所处环境较地上部分更为复杂导致.在恶劣的环境条件下,沙漠中的植物往往会为了生存而扩大根系,而不是支持地上部分的生长[36],非维管束植物齿肋赤藓则需要通过假根中菌丝体的生长提高其对土壤中水分和养分的吸收[37],菌丝体的发育状况和土壤可利用养分的差异导致齿肋赤藓地下部分的养分特征具有较高的变异性. ...
新疆古尔班通古特沙漠生物结皮绿藻研究
1
2006
... 古尔班通古特沙漠位于新疆北部准噶尔盆地腹心(44°11′—46°20′N,84°31′—90°00′E)[21],固定半固定沙丘占总面积的87%[22],面积约为4.88×104 km2,是中国最大的固定半固定沙漠.年降水量为70~190 mm,属于温带大陆性气候,年潜在蒸发量达2 606.6 mm,夏季最高温度超过40 ℃,冬季最低温度低于-20 ℃,年日照时间为2 780~2 980 h[23].由于降水稀少、气候干燥、潜在水分蒸发量大、日照时间长等恶劣的环境条件,古尔班通古特沙漠植被结构及组成单一、分布稀疏[24].在古尔班通古特沙漠固定半固定沙丘上大面积覆盖有生物土壤结皮,苔藓结皮是生物土壤结皮发育的高级阶段,主要是由藓类植物组成,其中齿肋赤藓是该沙漠中藓类植物的优势物种[25]. ...
古尔班通古特沙漠沙丘主要灌木的种群数量动态
1
2021
... 古尔班通古特沙漠位于新疆北部准噶尔盆地腹心(44°11′—46°20′N,84°31′—90°00′E)[21],固定半固定沙丘占总面积的87%[22],面积约为4.88×104 km2,是中国最大的固定半固定沙漠.年降水量为70~190 mm,属于温带大陆性气候,年潜在蒸发量达2 606.6 mm,夏季最高温度超过40 ℃,冬季最低温度低于-20 ℃,年日照时间为2 780~2 980 h[23].由于降水稀少、气候干燥、潜在水分蒸发量大、日照时间长等恶劣的环境条件,古尔班通古特沙漠植被结构及组成单一、分布稀疏[24].在古尔班通古特沙漠固定半固定沙丘上大面积覆盖有生物土壤结皮,苔藓结皮是生物土壤结皮发育的高级阶段,主要是由藓类植物组成,其中齿肋赤藓是该沙漠中藓类植物的优势物种[25]. ...
Microbiotic crusts and their interrelations with environmental factors in the Gurbantonggut Desert,western China
1
2006
... 古尔班通古特沙漠位于新疆北部准噶尔盆地腹心(44°11′—46°20′N,84°31′—90°00′E)[21],固定半固定沙丘占总面积的87%[22],面积约为4.88×104 km2,是中国最大的固定半固定沙漠.年降水量为70~190 mm,属于温带大陆性气候,年潜在蒸发量达2 606.6 mm,夏季最高温度超过40 ℃,冬季最低温度低于-20 ℃,年日照时间为2 780~2 980 h[23].由于降水稀少、气候干燥、潜在水分蒸发量大、日照时间长等恶劣的环境条件,古尔班通古特沙漠植被结构及组成单一、分布稀疏[24].在古尔班通古特沙漠固定半固定沙丘上大面积覆盖有生物土壤结皮,苔藓结皮是生物土壤结皮发育的高级阶段,主要是由藓类植物组成,其中齿肋赤藓是该沙漠中藓类植物的优势物种[25]. ...
古尔班通古特沙漠南部植物多样性及群落分类
1
2012
... 古尔班通古特沙漠位于新疆北部准噶尔盆地腹心(44°11′—46°20′N,84°31′—90°00′E)[21],固定半固定沙丘占总面积的87%[22],面积约为4.88×104 km2,是中国最大的固定半固定沙漠.年降水量为70~190 mm,属于温带大陆性气候,年潜在蒸发量达2 606.6 mm,夏季最高温度超过40 ℃,冬季最低温度低于-20 ℃,年日照时间为2 780~2 980 h[23].由于降水稀少、气候干燥、潜在水分蒸发量大、日照时间长等恶劣的环境条件,古尔班通古特沙漠植被结构及组成单一、分布稀疏[24].在古尔班通古特沙漠固定半固定沙丘上大面积覆盖有生物土壤结皮,苔藓结皮是生物土壤结皮发育的高级阶段,主要是由藓类植物组成,其中齿肋赤藓是该沙漠中藓类植物的优势物种[25]. ...
干旱与半干旱地区苔藓植物生态学研究综述
1
2002
... 古尔班通古特沙漠位于新疆北部准噶尔盆地腹心(44°11′—46°20′N,84°31′—90°00′E)[21],固定半固定沙丘占总面积的87%[22],面积约为4.88×104 km2,是中国最大的固定半固定沙漠.年降水量为70~190 mm,属于温带大陆性气候,年潜在蒸发量达2 606.6 mm,夏季最高温度超过40 ℃,冬季最低温度低于-20 ℃,年日照时间为2 780~2 980 h[23].由于降水稀少、气候干燥、潜在水分蒸发量大、日照时间长等恶劣的环境条件,古尔班通古特沙漠植被结构及组成单一、分布稀疏[24].在古尔班通古特沙漠固定半固定沙丘上大面积覆盖有生物土壤结皮,苔藓结皮是生物土壤结皮发育的高级阶段,主要是由藓类植物组成,其中齿肋赤藓是该沙漠中藓类植物的优势物种[25]. ...
不同微生境下齿肋赤藓(Syntrichia caninervis)生理生化特性对不同季节的响应
1
2018
... 数据处理、统计分析、绘制图表均利用R4.1.2(R Development Core Team 2021)完成.将植物地上、地下部分N、P和N∶P采用单因素方差分析(ANOVA)和最小显著差异法(LSD)进行方差分析和显著性多重比较,采用配对T检验方法检验植物不同部位的养分化学计量是否存在差异,采用Pearson相关性分析法[26]对齿肋赤藓植物地上、地下部分氮磷化学计量和化学计量比进行相关性分析,利用模型Ⅱ回归 (Reduced major axis regression, RMA) 对N、P元素进行N-P幂指数关系分析. ...
氮沉降背景下藓类植物齿肋赤藓的氮磷特征及异速生长关系
3
2021
... 植物叶片的N、P元素含量及其N∶P可以用来判断植物生长过程中是否受到N、P元素的限制.本研究测得齿肋赤藓植物地上部分N、P元素含量及N∶P的均值分别为12.2±0.19、1.13±0.22 mg·g-1及10.96±0.14 mg·g-1,梅月磊等[27]在对沙漠齿肋赤藓的模拟氮添加试验中采用如下两种判断方式考量齿肋赤藓是否受到养分限制.①根据Koerselman等[3]的标准,当N∶P小于14时,植物受到氮限制.②根据Ellison[28]的标准,当N含量小于20 mg·g-1、P含量大于 1 mg·g-1时,植物受氮限制.参照上述标准推测齿肋赤藓在该沙漠的生长易受到氮限制,与梅月磊等[27]所得结果一致.齿肋赤藓植物地上部分的N、P元素含量低于古尔班通古特沙漠草本植物叶片N、P含量(36.78、2.13 mg·g-1)[29];低于中国北方典型荒漠及荒漠化地区维管植物叶片N、P含量(24.45±8.1、1.74±0.88 mg·g-1)[30]和全国植被N、P含量的平均水平(18.6、1.21 mg·g-1)[31].养分库差异和自生养分运输能力限制是造成维管植物(叶片)和苔藓植物地上部分养分差异的原因[31].维管植物含有较发达的根系,这些植物具有较强的N、P元素吸收、存储和运输能力,而藓类植物没有真正的根系,假根对水分和养分的吸收可能来自于虹吸作用,并不能大范围、高效地吸收和储存土壤中的养分[32].本研究测得齿肋赤藓植物地上部分N、P元素含量高于Shi等[33]在青藏高原高海拔地区所测得结果(N: 9.33 mg·g-1);与吴盼盼等[34]在北半球中高纬度泥炭沼泽所得泥炭藓结果接近(11.48、1.16 mg·g-1),但与非泥炭藓结果呈现差异(9.73、1.5 mg·g-1),它们的养分差异体现了藓类植物对于不同养分来源、环境、海拔、季节等因素的适应性变化.各样点的齿肋赤藓N、P含量均表现出地上部分显著高于地下部分(图1),研究结果与Li等[20]、梅月磊等[27]测得结果一致.造成这一结果的原因是地上部分主要由具有光合能力的绿色植株组成,而地下部分主要是由假根、被沙埋后所产生的植物组织残骸组成,生理活性相较于地上部分低.齿肋赤藓植物地上部分的N和N∶P比的变异系数(CV)明显低于地下部分(P<0.05),元素可塑性假说[35]认为,植物地下部分的化学计量特征比植物地上部分茎叶的化学计量特征有更高的变异性,主要由于植物地下部分所处环境较地上部分更为复杂导致.在恶劣的环境条件下,沙漠中的植物往往会为了生存而扩大根系,而不是支持地上部分的生长[36],非维管束植物齿肋赤藓则需要通过假根中菌丝体的生长提高其对土壤中水分和养分的吸收[37],菌丝体的发育状况和土壤可利用养分的差异导致齿肋赤藓地下部分的养分特征具有较高的变异性. ...
... [27]所得结果一致.齿肋赤藓植物地上部分的N、P元素含量低于古尔班通古特沙漠草本植物叶片N、P含量(36.78、2.13 mg·g-1)[29];低于中国北方典型荒漠及荒漠化地区维管植物叶片N、P含量(24.45±8.1、1.74±0.88 mg·g-1)[30]和全国植被N、P含量的平均水平(18.6、1.21 mg·g-1)[31].养分库差异和自生养分运输能力限制是造成维管植物(叶片)和苔藓植物地上部分养分差异的原因[31].维管植物含有较发达的根系,这些植物具有较强的N、P元素吸收、存储和运输能力,而藓类植物没有真正的根系,假根对水分和养分的吸收可能来自于虹吸作用,并不能大范围、高效地吸收和储存土壤中的养分[32].本研究测得齿肋赤藓植物地上部分N、P元素含量高于Shi等[33]在青藏高原高海拔地区所测得结果(N: 9.33 mg·g-1);与吴盼盼等[34]在北半球中高纬度泥炭沼泽所得泥炭藓结果接近(11.48、1.16 mg·g-1),但与非泥炭藓结果呈现差异(9.73、1.5 mg·g-1),它们的养分差异体现了藓类植物对于不同养分来源、环境、海拔、季节等因素的适应性变化.各样点的齿肋赤藓N、P含量均表现出地上部分显著高于地下部分(图1),研究结果与Li等[20]、梅月磊等[27]测得结果一致.造成这一结果的原因是地上部分主要由具有光合能力的绿色植株组成,而地下部分主要是由假根、被沙埋后所产生的植物组织残骸组成,生理活性相较于地上部分低.齿肋赤藓植物地上部分的N和N∶P比的变异系数(CV)明显低于地下部分(P<0.05),元素可塑性假说[35]认为,植物地下部分的化学计量特征比植物地上部分茎叶的化学计量特征有更高的变异性,主要由于植物地下部分所处环境较地上部分更为复杂导致.在恶劣的环境条件下,沙漠中的植物往往会为了生存而扩大根系,而不是支持地上部分的生长[36],非维管束植物齿肋赤藓则需要通过假根中菌丝体的生长提高其对土壤中水分和养分的吸收[37],菌丝体的发育状况和土壤可利用养分的差异导致齿肋赤藓地下部分的养分特征具有较高的变异性. ...
... [27]测得结果一致.造成这一结果的原因是地上部分主要由具有光合能力的绿色植株组成,而地下部分主要是由假根、被沙埋后所产生的植物组织残骸组成,生理活性相较于地上部分低.齿肋赤藓植物地上部分的N和N∶P比的变异系数(CV)明显低于地下部分(P<0.05),元素可塑性假说[35]认为,植物地下部分的化学计量特征比植物地上部分茎叶的化学计量特征有更高的变异性,主要由于植物地下部分所处环境较地上部分更为复杂导致.在恶劣的环境条件下,沙漠中的植物往往会为了生存而扩大根系,而不是支持地上部分的生长[36],非维管束植物齿肋赤藓则需要通过假根中菌丝体的生长提高其对土壤中水分和养分的吸收[37],菌丝体的发育状况和土壤可利用养分的差异导致齿肋赤藓地下部分的养分特征具有较高的变异性. ...
Nutrient limitation and stoichiometry of carnivorous plants
1
2006
... 植物叶片的N、P元素含量及其N∶P可以用来判断植物生长过程中是否受到N、P元素的限制.本研究测得齿肋赤藓植物地上部分N、P元素含量及N∶P的均值分别为12.2±0.19、1.13±0.22 mg·g-1及10.96±0.14 mg·g-1,梅月磊等[27]在对沙漠齿肋赤藓的模拟氮添加试验中采用如下两种判断方式考量齿肋赤藓是否受到养分限制.①根据Koerselman等[3]的标准,当N∶P小于14时,植物受到氮限制.②根据Ellison[28]的标准,当N含量小于20 mg·g-1、P含量大于 1 mg·g-1时,植物受氮限制.参照上述标准推测齿肋赤藓在该沙漠的生长易受到氮限制,与梅月磊等[27]所得结果一致.齿肋赤藓植物地上部分的N、P元素含量低于古尔班通古特沙漠草本植物叶片N、P含量(36.78、2.13 mg·g-1)[29];低于中国北方典型荒漠及荒漠化地区维管植物叶片N、P含量(24.45±8.1、1.74±0.88 mg·g-1)[30]和全国植被N、P含量的平均水平(18.6、1.21 mg·g-1)[31].养分库差异和自生养分运输能力限制是造成维管植物(叶片)和苔藓植物地上部分养分差异的原因[31].维管植物含有较发达的根系,这些植物具有较强的N、P元素吸收、存储和运输能力,而藓类植物没有真正的根系,假根对水分和养分的吸收可能来自于虹吸作用,并不能大范围、高效地吸收和储存土壤中的养分[32].本研究测得齿肋赤藓植物地上部分N、P元素含量高于Shi等[33]在青藏高原高海拔地区所测得结果(N: 9.33 mg·g-1);与吴盼盼等[34]在北半球中高纬度泥炭沼泽所得泥炭藓结果接近(11.48、1.16 mg·g-1),但与非泥炭藓结果呈现差异(9.73、1.5 mg·g-1),它们的养分差异体现了藓类植物对于不同养分来源、环境、海拔、季节等因素的适应性变化.各样点的齿肋赤藓N、P含量均表现出地上部分显著高于地下部分(图1),研究结果与Li等[20]、梅月磊等[27]测得结果一致.造成这一结果的原因是地上部分主要由具有光合能力的绿色植株组成,而地下部分主要是由假根、被沙埋后所产生的植物组织残骸组成,生理活性相较于地上部分低.齿肋赤藓植物地上部分的N和N∶P比的变异系数(CV)明显低于地下部分(P<0.05),元素可塑性假说[35]认为,植物地下部分的化学计量特征比植物地上部分茎叶的化学计量特征有更高的变异性,主要由于植物地下部分所处环境较地上部分更为复杂导致.在恶劣的环境条件下,沙漠中的植物往往会为了生存而扩大根系,而不是支持地上部分的生长[36],非维管束植物齿肋赤藓则需要通过假根中菌丝体的生长提高其对土壤中水分和养分的吸收[37],菌丝体的发育状况和土壤可利用养分的差异导致齿肋赤藓地下部分的养分特征具有较高的变异性. ...
古尔班通古特沙漠4种草本植物叶片与土壤的化学计量特征
1
2015
... 植物叶片的N、P元素含量及其N∶P可以用来判断植物生长过程中是否受到N、P元素的限制.本研究测得齿肋赤藓植物地上部分N、P元素含量及N∶P的均值分别为12.2±0.19、1.13±0.22 mg·g-1及10.96±0.14 mg·g-1,梅月磊等[27]在对沙漠齿肋赤藓的模拟氮添加试验中采用如下两种判断方式考量齿肋赤藓是否受到养分限制.①根据Koerselman等[3]的标准,当N∶P小于14时,植物受到氮限制.②根据Ellison[28]的标准,当N含量小于20 mg·g-1、P含量大于 1 mg·g-1时,植物受氮限制.参照上述标准推测齿肋赤藓在该沙漠的生长易受到氮限制,与梅月磊等[27]所得结果一致.齿肋赤藓植物地上部分的N、P元素含量低于古尔班通古特沙漠草本植物叶片N、P含量(36.78、2.13 mg·g-1)[29];低于中国北方典型荒漠及荒漠化地区维管植物叶片N、P含量(24.45±8.1、1.74±0.88 mg·g-1)[30]和全国植被N、P含量的平均水平(18.6、1.21 mg·g-1)[31].养分库差异和自生养分运输能力限制是造成维管植物(叶片)和苔藓植物地上部分养分差异的原因[31].维管植物含有较发达的根系,这些植物具有较强的N、P元素吸收、存储和运输能力,而藓类植物没有真正的根系,假根对水分和养分的吸收可能来自于虹吸作用,并不能大范围、高效地吸收和储存土壤中的养分[32].本研究测得齿肋赤藓植物地上部分N、P元素含量高于Shi等[33]在青藏高原高海拔地区所测得结果(N: 9.33 mg·g-1);与吴盼盼等[34]在北半球中高纬度泥炭沼泽所得泥炭藓结果接近(11.48、1.16 mg·g-1),但与非泥炭藓结果呈现差异(9.73、1.5 mg·g-1),它们的养分差异体现了藓类植物对于不同养分来源、环境、海拔、季节等因素的适应性变化.各样点的齿肋赤藓N、P含量均表现出地上部分显著高于地下部分(图1),研究结果与Li等[20]、梅月磊等[27]测得结果一致.造成这一结果的原因是地上部分主要由具有光合能力的绿色植株组成,而地下部分主要是由假根、被沙埋后所产生的植物组织残骸组成,生理活性相较于地上部分低.齿肋赤藓植物地上部分的N和N∶P比的变异系数(CV)明显低于地下部分(P<0.05),元素可塑性假说[35]认为,植物地下部分的化学计量特征比植物地上部分茎叶的化学计量特征有更高的变异性,主要由于植物地下部分所处环境较地上部分更为复杂导致.在恶劣的环境条件下,沙漠中的植物往往会为了生存而扩大根系,而不是支持地上部分的生长[36],非维管束植物齿肋赤藓则需要通过假根中菌丝体的生长提高其对土壤中水分和养分的吸收[37],菌丝体的发育状况和土壤可利用养分的差异导致齿肋赤藓地下部分的养分特征具有较高的变异性. ...
北方典型荒漠及荒漠化地区植物叶片氮磷化学计量特征研究
1
2010
... 植物叶片的N、P元素含量及其N∶P可以用来判断植物生长过程中是否受到N、P元素的限制.本研究测得齿肋赤藓植物地上部分N、P元素含量及N∶P的均值分别为12.2±0.19、1.13±0.22 mg·g-1及10.96±0.14 mg·g-1,梅月磊等[27]在对沙漠齿肋赤藓的模拟氮添加试验中采用如下两种判断方式考量齿肋赤藓是否受到养分限制.①根据Koerselman等[3]的标准,当N∶P小于14时,植物受到氮限制.②根据Ellison[28]的标准,当N含量小于20 mg·g-1、P含量大于 1 mg·g-1时,植物受氮限制.参照上述标准推测齿肋赤藓在该沙漠的生长易受到氮限制,与梅月磊等[27]所得结果一致.齿肋赤藓植物地上部分的N、P元素含量低于古尔班通古特沙漠草本植物叶片N、P含量(36.78、2.13 mg·g-1)[29];低于中国北方典型荒漠及荒漠化地区维管植物叶片N、P含量(24.45±8.1、1.74±0.88 mg·g-1)[30]和全国植被N、P含量的平均水平(18.6、1.21 mg·g-1)[31].养分库差异和自生养分运输能力限制是造成维管植物(叶片)和苔藓植物地上部分养分差异的原因[31].维管植物含有较发达的根系,这些植物具有较强的N、P元素吸收、存储和运输能力,而藓类植物没有真正的根系,假根对水分和养分的吸收可能来自于虹吸作用,并不能大范围、高效地吸收和储存土壤中的养分[32].本研究测得齿肋赤藓植物地上部分N、P元素含量高于Shi等[33]在青藏高原高海拔地区所测得结果(N: 9.33 mg·g-1);与吴盼盼等[34]在北半球中高纬度泥炭沼泽所得泥炭藓结果接近(11.48、1.16 mg·g-1),但与非泥炭藓结果呈现差异(9.73、1.5 mg·g-1),它们的养分差异体现了藓类植物对于不同养分来源、环境、海拔、季节等因素的适应性变化.各样点的齿肋赤藓N、P含量均表现出地上部分显著高于地下部分(图1),研究结果与Li等[20]、梅月磊等[27]测得结果一致.造成这一结果的原因是地上部分主要由具有光合能力的绿色植株组成,而地下部分主要是由假根、被沙埋后所产生的植物组织残骸组成,生理活性相较于地上部分低.齿肋赤藓植物地上部分的N和N∶P比的变异系数(CV)明显低于地下部分(P<0.05),元素可塑性假说[35]认为,植物地下部分的化学计量特征比植物地上部分茎叶的化学计量特征有更高的变异性,主要由于植物地下部分所处环境较地上部分更为复杂导致.在恶劣的环境条件下,沙漠中的植物往往会为了生存而扩大根系,而不是支持地上部分的生长[36],非维管束植物齿肋赤藓则需要通过假根中菌丝体的生长提高其对土壤中水分和养分的吸收[37],菌丝体的发育状况和土壤可利用养分的差异导致齿肋赤藓地下部分的养分特征具有较高的变异性. ...
Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China
2
2005
... 植物叶片的N、P元素含量及其N∶P可以用来判断植物生长过程中是否受到N、P元素的限制.本研究测得齿肋赤藓植物地上部分N、P元素含量及N∶P的均值分别为12.2±0.19、1.13±0.22 mg·g-1及10.96±0.14 mg·g-1,梅月磊等[27]在对沙漠齿肋赤藓的模拟氮添加试验中采用如下两种判断方式考量齿肋赤藓是否受到养分限制.①根据Koerselman等[3]的标准,当N∶P小于14时,植物受到氮限制.②根据Ellison[28]的标准,当N含量小于20 mg·g-1、P含量大于 1 mg·g-1时,植物受氮限制.参照上述标准推测齿肋赤藓在该沙漠的生长易受到氮限制,与梅月磊等[27]所得结果一致.齿肋赤藓植物地上部分的N、P元素含量低于古尔班通古特沙漠草本植物叶片N、P含量(36.78、2.13 mg·g-1)[29];低于中国北方典型荒漠及荒漠化地区维管植物叶片N、P含量(24.45±8.1、1.74±0.88 mg·g-1)[30]和全国植被N、P含量的平均水平(18.6、1.21 mg·g-1)[31].养分库差异和自生养分运输能力限制是造成维管植物(叶片)和苔藓植物地上部分养分差异的原因[31].维管植物含有较发达的根系,这些植物具有较强的N、P元素吸收、存储和运输能力,而藓类植物没有真正的根系,假根对水分和养分的吸收可能来自于虹吸作用,并不能大范围、高效地吸收和储存土壤中的养分[32].本研究测得齿肋赤藓植物地上部分N、P元素含量高于Shi等[33]在青藏高原高海拔地区所测得结果(N: 9.33 mg·g-1);与吴盼盼等[34]在北半球中高纬度泥炭沼泽所得泥炭藓结果接近(11.48、1.16 mg·g-1),但与非泥炭藓结果呈现差异(9.73、1.5 mg·g-1),它们的养分差异体现了藓类植物对于不同养分来源、环境、海拔、季节等因素的适应性变化.各样点的齿肋赤藓N、P含量均表现出地上部分显著高于地下部分(图1),研究结果与Li等[20]、梅月磊等[27]测得结果一致.造成这一结果的原因是地上部分主要由具有光合能力的绿色植株组成,而地下部分主要是由假根、被沙埋后所产生的植物组织残骸组成,生理活性相较于地上部分低.齿肋赤藓植物地上部分的N和N∶P比的变异系数(CV)明显低于地下部分(P<0.05),元素可塑性假说[35]认为,植物地下部分的化学计量特征比植物地上部分茎叶的化学计量特征有更高的变异性,主要由于植物地下部分所处环境较地上部分更为复杂导致.在恶劣的环境条件下,沙漠中的植物往往会为了生存而扩大根系,而不是支持地上部分的生长[36],非维管束植物齿肋赤藓则需要通过假根中菌丝体的生长提高其对土壤中水分和养分的吸收[37],菌丝体的发育状况和土壤可利用养分的差异导致齿肋赤藓地下部分的养分特征具有较高的变异性. ...
... [31].维管植物含有较发达的根系,这些植物具有较强的N、P元素吸收、存储和运输能力,而藓类植物没有真正的根系,假根对水分和养分的吸收可能来自于虹吸作用,并不能大范围、高效地吸收和储存土壤中的养分[32].本研究测得齿肋赤藓植物地上部分N、P元素含量高于Shi等[33]在青藏高原高海拔地区所测得结果(N: 9.33 mg·g-1);与吴盼盼等[34]在北半球中高纬度泥炭沼泽所得泥炭藓结果接近(11.48、1.16 mg·g-1),但与非泥炭藓结果呈现差异(9.73、1.5 mg·g-1),它们的养分差异体现了藓类植物对于不同养分来源、环境、海拔、季节等因素的适应性变化.各样点的齿肋赤藓N、P含量均表现出地上部分显著高于地下部分(图1),研究结果与Li等[20]、梅月磊等[27]测得结果一致.造成这一结果的原因是地上部分主要由具有光合能力的绿色植株组成,而地下部分主要是由假根、被沙埋后所产生的植物组织残骸组成,生理活性相较于地上部分低.齿肋赤藓植物地上部分的N和N∶P比的变异系数(CV)明显低于地下部分(P<0.05),元素可塑性假说[35]认为,植物地下部分的化学计量特征比植物地上部分茎叶的化学计量特征有更高的变异性,主要由于植物地下部分所处环境较地上部分更为复杂导致.在恶劣的环境条件下,沙漠中的植物往往会为了生存而扩大根系,而不是支持地上部分的生长[36],非维管束植物齿肋赤藓则需要通过假根中菌丝体的生长提高其对土壤中水分和养分的吸收[37],菌丝体的发育状况和土壤可利用养分的差异导致齿肋赤藓地下部分的养分特征具有较高的变异性. ...
Shrub modulates the stoichiometry of moss and soil in desert ecosystems,China
1
2019
... 植物叶片的N、P元素含量及其N∶P可以用来判断植物生长过程中是否受到N、P元素的限制.本研究测得齿肋赤藓植物地上部分N、P元素含量及N∶P的均值分别为12.2±0.19、1.13±0.22 mg·g-1及10.96±0.14 mg·g-1,梅月磊等[27]在对沙漠齿肋赤藓的模拟氮添加试验中采用如下两种判断方式考量齿肋赤藓是否受到养分限制.①根据Koerselman等[3]的标准,当N∶P小于14时,植物受到氮限制.②根据Ellison[28]的标准,当N含量小于20 mg·g-1、P含量大于 1 mg·g-1时,植物受氮限制.参照上述标准推测齿肋赤藓在该沙漠的生长易受到氮限制,与梅月磊等[27]所得结果一致.齿肋赤藓植物地上部分的N、P元素含量低于古尔班通古特沙漠草本植物叶片N、P含量(36.78、2.13 mg·g-1)[29];低于中国北方典型荒漠及荒漠化地区维管植物叶片N、P含量(24.45±8.1、1.74±0.88 mg·g-1)[30]和全国植被N、P含量的平均水平(18.6、1.21 mg·g-1)[31].养分库差异和自生养分运输能力限制是造成维管植物(叶片)和苔藓植物地上部分养分差异的原因[31].维管植物含有较发达的根系,这些植物具有较强的N、P元素吸收、存储和运输能力,而藓类植物没有真正的根系,假根对水分和养分的吸收可能来自于虹吸作用,并不能大范围、高效地吸收和储存土壤中的养分[32].本研究测得齿肋赤藓植物地上部分N、P元素含量高于Shi等[33]在青藏高原高海拔地区所测得结果(N: 9.33 mg·g-1);与吴盼盼等[34]在北半球中高纬度泥炭沼泽所得泥炭藓结果接近(11.48、1.16 mg·g-1),但与非泥炭藓结果呈现差异(9.73、1.5 mg·g-1),它们的养分差异体现了藓类植物对于不同养分来源、环境、海拔、季节等因素的适应性变化.各样点的齿肋赤藓N、P含量均表现出地上部分显著高于地下部分(图1),研究结果与Li等[20]、梅月磊等[27]测得结果一致.造成这一结果的原因是地上部分主要由具有光合能力的绿色植株组成,而地下部分主要是由假根、被沙埋后所产生的植物组织残骸组成,生理活性相较于地上部分低.齿肋赤藓植物地上部分的N和N∶P比的变异系数(CV)明显低于地下部分(P<0.05),元素可塑性假说[35]认为,植物地下部分的化学计量特征比植物地上部分茎叶的化学计量特征有更高的变异性,主要由于植物地下部分所处环境较地上部分更为复杂导致.在恶劣的环境条件下,沙漠中的植物往往会为了生存而扩大根系,而不是支持地上部分的生长[36],非维管束植物齿肋赤藓则需要通过假根中菌丝体的生长提高其对土壤中水分和养分的吸收[37],菌丝体的发育状况和土壤可利用养分的差异导致齿肋赤藓地下部分的养分特征具有较高的变异性. ...
Altitudinal variation in leaf nitrogen concentration on the Eastern Slope of Mount Gongga on the Tibetan Plateau,China
1
2012
... 植物叶片的N、P元素含量及其N∶P可以用来判断植物生长过程中是否受到N、P元素的限制.本研究测得齿肋赤藓植物地上部分N、P元素含量及N∶P的均值分别为12.2±0.19、1.13±0.22 mg·g-1及10.96±0.14 mg·g-1,梅月磊等[27]在对沙漠齿肋赤藓的模拟氮添加试验中采用如下两种判断方式考量齿肋赤藓是否受到养分限制.①根据Koerselman等[3]的标准,当N∶P小于14时,植物受到氮限制.②根据Ellison[28]的标准,当N含量小于20 mg·g-1、P含量大于 1 mg·g-1时,植物受氮限制.参照上述标准推测齿肋赤藓在该沙漠的生长易受到氮限制,与梅月磊等[27]所得结果一致.齿肋赤藓植物地上部分的N、P元素含量低于古尔班通古特沙漠草本植物叶片N、P含量(36.78、2.13 mg·g-1)[29];低于中国北方典型荒漠及荒漠化地区维管植物叶片N、P含量(24.45±8.1、1.74±0.88 mg·g-1)[30]和全国植被N、P含量的平均水平(18.6、1.21 mg·g-1)[31].养分库差异和自生养分运输能力限制是造成维管植物(叶片)和苔藓植物地上部分养分差异的原因[31].维管植物含有较发达的根系,这些植物具有较强的N、P元素吸收、存储和运输能力,而藓类植物没有真正的根系,假根对水分和养分的吸收可能来自于虹吸作用,并不能大范围、高效地吸收和储存土壤中的养分[32].本研究测得齿肋赤藓植物地上部分N、P元素含量高于Shi等[33]在青藏高原高海拔地区所测得结果(N: 9.33 mg·g-1);与吴盼盼等[34]在北半球中高纬度泥炭沼泽所得泥炭藓结果接近(11.48、1.16 mg·g-1),但与非泥炭藓结果呈现差异(9.73、1.5 mg·g-1),它们的养分差异体现了藓类植物对于不同养分来源、环境、海拔、季节等因素的适应性变化.各样点的齿肋赤藓N、P含量均表现出地上部分显著高于地下部分(图1),研究结果与Li等[20]、梅月磊等[27]测得结果一致.造成这一结果的原因是地上部分主要由具有光合能力的绿色植株组成,而地下部分主要是由假根、被沙埋后所产生的植物组织残骸组成,生理活性相较于地上部分低.齿肋赤藓植物地上部分的N和N∶P比的变异系数(CV)明显低于地下部分(P<0.05),元素可塑性假说[35]认为,植物地下部分的化学计量特征比植物地上部分茎叶的化学计量特征有更高的变异性,主要由于植物地下部分所处环境较地上部分更为复杂导致.在恶劣的环境条件下,沙漠中的植物往往会为了生存而扩大根系,而不是支持地上部分的生长[36],非维管束植物齿肋赤藓则需要通过假根中菌丝体的生长提高其对土壤中水分和养分的吸收[37],菌丝体的发育状况和土壤可利用养分的差异导致齿肋赤藓地下部分的养分特征具有较高的变异性. ...
大兴安岭贫营养泥炭地维管束和非维管束植物叶片碳氮磷化学计量学特征
1
2020
... 植物叶片的N、P元素含量及其N∶P可以用来判断植物生长过程中是否受到N、P元素的限制.本研究测得齿肋赤藓植物地上部分N、P元素含量及N∶P的均值分别为12.2±0.19、1.13±0.22 mg·g-1及10.96±0.14 mg·g-1,梅月磊等[27]在对沙漠齿肋赤藓的模拟氮添加试验中采用如下两种判断方式考量齿肋赤藓是否受到养分限制.①根据Koerselman等[3]的标准,当N∶P小于14时,植物受到氮限制.②根据Ellison[28]的标准,当N含量小于20 mg·g-1、P含量大于 1 mg·g-1时,植物受氮限制.参照上述标准推测齿肋赤藓在该沙漠的生长易受到氮限制,与梅月磊等[27]所得结果一致.齿肋赤藓植物地上部分的N、P元素含量低于古尔班通古特沙漠草本植物叶片N、P含量(36.78、2.13 mg·g-1)[29];低于中国北方典型荒漠及荒漠化地区维管植物叶片N、P含量(24.45±8.1、1.74±0.88 mg·g-1)[30]和全国植被N、P含量的平均水平(18.6、1.21 mg·g-1)[31].养分库差异和自生养分运输能力限制是造成维管植物(叶片)和苔藓植物地上部分养分差异的原因[31].维管植物含有较发达的根系,这些植物具有较强的N、P元素吸收、存储和运输能力,而藓类植物没有真正的根系,假根对水分和养分的吸收可能来自于虹吸作用,并不能大范围、高效地吸收和储存土壤中的养分[32].本研究测得齿肋赤藓植物地上部分N、P元素含量高于Shi等[33]在青藏高原高海拔地区所测得结果(N: 9.33 mg·g-1);与吴盼盼等[34]在北半球中高纬度泥炭沼泽所得泥炭藓结果接近(11.48、1.16 mg·g-1),但与非泥炭藓结果呈现差异(9.73、1.5 mg·g-1),它们的养分差异体现了藓类植物对于不同养分来源、环境、海拔、季节等因素的适应性变化.各样点的齿肋赤藓N、P含量均表现出地上部分显著高于地下部分(图1),研究结果与Li等[20]、梅月磊等[27]测得结果一致.造成这一结果的原因是地上部分主要由具有光合能力的绿色植株组成,而地下部分主要是由假根、被沙埋后所产生的植物组织残骸组成,生理活性相较于地上部分低.齿肋赤藓植物地上部分的N和N∶P比的变异系数(CV)明显低于地下部分(P<0.05),元素可塑性假说[35]认为,植物地下部分的化学计量特征比植物地上部分茎叶的化学计量特征有更高的变异性,主要由于植物地下部分所处环境较地上部分更为复杂导致.在恶劣的环境条件下,沙漠中的植物往往会为了生存而扩大根系,而不是支持地上部分的生长[36],非维管束植物齿肋赤藓则需要通过假根中菌丝体的生长提高其对土壤中水分和养分的吸收[37],菌丝体的发育状况和土壤可利用养分的差异导致齿肋赤藓地下部分的养分特征具有较高的变异性. ...
Allocation strategies for nitrogen and phosphorus in forest plants
2
2018
... 植物叶片的N、P元素含量及其N∶P可以用来判断植物生长过程中是否受到N、P元素的限制.本研究测得齿肋赤藓植物地上部分N、P元素含量及N∶P的均值分别为12.2±0.19、1.13±0.22 mg·g-1及10.96±0.14 mg·g-1,梅月磊等[27]在对沙漠齿肋赤藓的模拟氮添加试验中采用如下两种判断方式考量齿肋赤藓是否受到养分限制.①根据Koerselman等[3]的标准,当N∶P小于14时,植物受到氮限制.②根据Ellison[28]的标准,当N含量小于20 mg·g-1、P含量大于 1 mg·g-1时,植物受氮限制.参照上述标准推测齿肋赤藓在该沙漠的生长易受到氮限制,与梅月磊等[27]所得结果一致.齿肋赤藓植物地上部分的N、P元素含量低于古尔班通古特沙漠草本植物叶片N、P含量(36.78、2.13 mg·g-1)[29];低于中国北方典型荒漠及荒漠化地区维管植物叶片N、P含量(24.45±8.1、1.74±0.88 mg·g-1)[30]和全国植被N、P含量的平均水平(18.6、1.21 mg·g-1)[31].养分库差异和自生养分运输能力限制是造成维管植物(叶片)和苔藓植物地上部分养分差异的原因[31].维管植物含有较发达的根系,这些植物具有较强的N、P元素吸收、存储和运输能力,而藓类植物没有真正的根系,假根对水分和养分的吸收可能来自于虹吸作用,并不能大范围、高效地吸收和储存土壤中的养分[32].本研究测得齿肋赤藓植物地上部分N、P元素含量高于Shi等[33]在青藏高原高海拔地区所测得结果(N: 9.33 mg·g-1);与吴盼盼等[34]在北半球中高纬度泥炭沼泽所得泥炭藓结果接近(11.48、1.16 mg·g-1),但与非泥炭藓结果呈现差异(9.73、1.5 mg·g-1),它们的养分差异体现了藓类植物对于不同养分来源、环境、海拔、季节等因素的适应性变化.各样点的齿肋赤藓N、P含量均表现出地上部分显著高于地下部分(图1),研究结果与Li等[20]、梅月磊等[27]测得结果一致.造成这一结果的原因是地上部分主要由具有光合能力的绿色植株组成,而地下部分主要是由假根、被沙埋后所产生的植物组织残骸组成,生理活性相较于地上部分低.齿肋赤藓植物地上部分的N和N∶P比的变异系数(CV)明显低于地下部分(P<0.05),元素可塑性假说[35]认为,植物地下部分的化学计量特征比植物地上部分茎叶的化学计量特征有更高的变异性,主要由于植物地下部分所处环境较地上部分更为复杂导致.在恶劣的环境条件下,沙漠中的植物往往会为了生存而扩大根系,而不是支持地上部分的生长[36],非维管束植物齿肋赤藓则需要通过假根中菌丝体的生长提高其对土壤中水分和养分的吸收[37],菌丝体的发育状况和土壤可利用养分的差异导致齿肋赤藓地下部分的养分特征具有较高的变异性. ...
... 通过对各样点地上、地下部分的相关性分析发现:无论是地上部分还是地下部分,齿肋赤藓N、P元素含量均为显著正相关,这种相关性反映了植物体内营养元素间具有较强的线性关系[40],本文结果进一步验证了此观点.限制元素N、P呈异速关系是植物适应环境的重要策略[41-42],齿肋赤藓同部位N、P含量均表现出显著的异速关系,说明齿肋赤藓各部位N、P比例相对稳定.在众多大尺度的调查、养分添加的研究中,所得N和P的幂指数比均小于1,且斜率间无显著差异,齿肋赤藓植物养分吸收、利用和分配关系随环境改变呈现不显著的差异性变化,具有一定的保守性[35].本研究中齿肋赤藓植物地上部分N-P幂指数计量比均小于1,其结果均接近前人所提出的植物叶片2/3[15]或3/4[16]N-P幂指数计量关系;齿肋赤藓植物地下部分N-P幂指数计量关系均大于1,说明齿肋赤藓植物地下部分P含量占比相较于N含量高,这可能与藓类植物能大量吸收土壤P元素的特性有关[43]. ...
Root-shoot growth responses during interspecific competition quantified using allometric modelling
1
2010
... 植物叶片的N、P元素含量及其N∶P可以用来判断植物生长过程中是否受到N、P元素的限制.本研究测得齿肋赤藓植物地上部分N、P元素含量及N∶P的均值分别为12.2±0.19、1.13±0.22 mg·g-1及10.96±0.14 mg·g-1,梅月磊等[27]在对沙漠齿肋赤藓的模拟氮添加试验中采用如下两种判断方式考量齿肋赤藓是否受到养分限制.①根据Koerselman等[3]的标准,当N∶P小于14时,植物受到氮限制.②根据Ellison[28]的标准,当N含量小于20 mg·g-1、P含量大于 1 mg·g-1时,植物受氮限制.参照上述标准推测齿肋赤藓在该沙漠的生长易受到氮限制,与梅月磊等[27]所得结果一致.齿肋赤藓植物地上部分的N、P元素含量低于古尔班通古特沙漠草本植物叶片N、P含量(36.78、2.13 mg·g-1)[29];低于中国北方典型荒漠及荒漠化地区维管植物叶片N、P含量(24.45±8.1、1.74±0.88 mg·g-1)[30]和全国植被N、P含量的平均水平(18.6、1.21 mg·g-1)[31].养分库差异和自生养分运输能力限制是造成维管植物(叶片)和苔藓植物地上部分养分差异的原因[31].维管植物含有较发达的根系,这些植物具有较强的N、P元素吸收、存储和运输能力,而藓类植物没有真正的根系,假根对水分和养分的吸收可能来自于虹吸作用,并不能大范围、高效地吸收和储存土壤中的养分[32].本研究测得齿肋赤藓植物地上部分N、P元素含量高于Shi等[33]在青藏高原高海拔地区所测得结果(N: 9.33 mg·g-1);与吴盼盼等[34]在北半球中高纬度泥炭沼泽所得泥炭藓结果接近(11.48、1.16 mg·g-1),但与非泥炭藓结果呈现差异(9.73、1.5 mg·g-1),它们的养分差异体现了藓类植物对于不同养分来源、环境、海拔、季节等因素的适应性变化.各样点的齿肋赤藓N、P含量均表现出地上部分显著高于地下部分(图1),研究结果与Li等[20]、梅月磊等[27]测得结果一致.造成这一结果的原因是地上部分主要由具有光合能力的绿色植株组成,而地下部分主要是由假根、被沙埋后所产生的植物组织残骸组成,生理活性相较于地上部分低.齿肋赤藓植物地上部分的N和N∶P比的变异系数(CV)明显低于地下部分(P<0.05),元素可塑性假说[35]认为,植物地下部分的化学计量特征比植物地上部分茎叶的化学计量特征有更高的变异性,主要由于植物地下部分所处环境较地上部分更为复杂导致.在恶劣的环境条件下,沙漠中的植物往往会为了生存而扩大根系,而不是支持地上部分的生长[36],非维管束植物齿肋赤藓则需要通过假根中菌丝体的生长提高其对土壤中水分和养分的吸收[37],菌丝体的发育状况和土壤可利用养分的差异导致齿肋赤藓地下部分的养分特征具有较高的变异性. ...
生物土壤结皮发育及其影响因素研究进展
1
2017
... 植物叶片的N、P元素含量及其N∶P可以用来判断植物生长过程中是否受到N、P元素的限制.本研究测得齿肋赤藓植物地上部分N、P元素含量及N∶P的均值分别为12.2±0.19、1.13±0.22 mg·g-1及10.96±0.14 mg·g-1,梅月磊等[27]在对沙漠齿肋赤藓的模拟氮添加试验中采用如下两种判断方式考量齿肋赤藓是否受到养分限制.①根据Koerselman等[3]的标准,当N∶P小于14时,植物受到氮限制.②根据Ellison[28]的标准,当N含量小于20 mg·g-1、P含量大于 1 mg·g-1时,植物受氮限制.参照上述标准推测齿肋赤藓在该沙漠的生长易受到氮限制,与梅月磊等[27]所得结果一致.齿肋赤藓植物地上部分的N、P元素含量低于古尔班通古特沙漠草本植物叶片N、P含量(36.78、2.13 mg·g-1)[29];低于中国北方典型荒漠及荒漠化地区维管植物叶片N、P含量(24.45±8.1、1.74±0.88 mg·g-1)[30]和全国植被N、P含量的平均水平(18.6、1.21 mg·g-1)[31].养分库差异和自生养分运输能力限制是造成维管植物(叶片)和苔藓植物地上部分养分差异的原因[31].维管植物含有较发达的根系,这些植物具有较强的N、P元素吸收、存储和运输能力,而藓类植物没有真正的根系,假根对水分和养分的吸收可能来自于虹吸作用,并不能大范围、高效地吸收和储存土壤中的养分[32].本研究测得齿肋赤藓植物地上部分N、P元素含量高于Shi等[33]在青藏高原高海拔地区所测得结果(N: 9.33 mg·g-1);与吴盼盼等[34]在北半球中高纬度泥炭沼泽所得泥炭藓结果接近(11.48、1.16 mg·g-1),但与非泥炭藓结果呈现差异(9.73、1.5 mg·g-1),它们的养分差异体现了藓类植物对于不同养分来源、环境、海拔、季节等因素的适应性变化.各样点的齿肋赤藓N、P含量均表现出地上部分显著高于地下部分(图1),研究结果与Li等[20]、梅月磊等[27]测得结果一致.造成这一结果的原因是地上部分主要由具有光合能力的绿色植株组成,而地下部分主要是由假根、被沙埋后所产生的植物组织残骸组成,生理活性相较于地上部分低.齿肋赤藓植物地上部分的N和N∶P比的变异系数(CV)明显低于地下部分(P<0.05),元素可塑性假说[35]认为,植物地下部分的化学计量特征比植物地上部分茎叶的化学计量特征有更高的变异性,主要由于植物地下部分所处环境较地上部分更为复杂导致.在恶劣的环境条件下,沙漠中的植物往往会为了生存而扩大根系,而不是支持地上部分的生长[36],非维管束植物齿肋赤藓则需要通过假根中菌丝体的生长提高其对土壤中水分和养分的吸收[37],菌丝体的发育状况和土壤可利用养分的差异导致齿肋赤藓地下部分的养分特征具有较高的变异性. ...
模拟增雨对白刺和油蒿幼苗化学计量特征的影响
1
2020
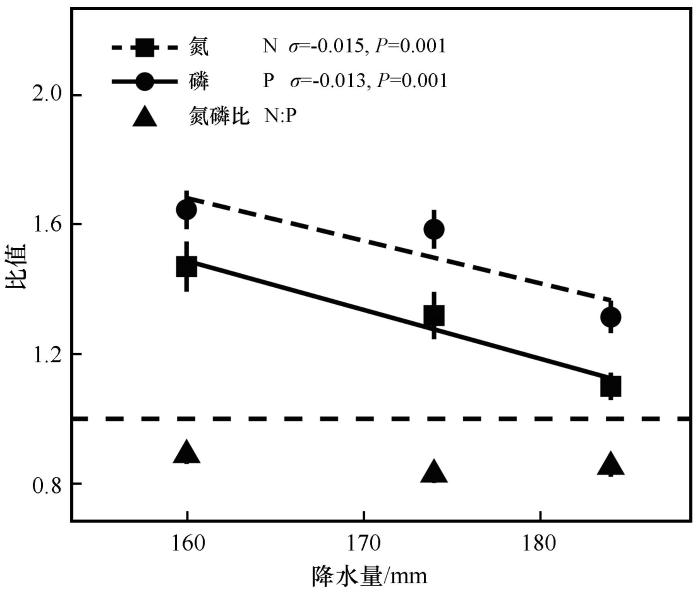
... 本研究中,年降水量最小的样点(G1)齿肋赤藓植物地上部分N、P元素含量最高,可能的原因是:①在干旱条件下,植物需要提高细胞内部的渗透压,增强对体内水分的保持、提高水分利用效率,适应干旱环境[38];②干旱条件可能会降低养分的质量流量或扩散率,从而限制养分在地上部与地下部之间的运输[39],导致地上、地下的N、P元素含量差异较大.随着年降水量的增加,水分胁迫作用减缓,养分的扩散率增加,植物地上部分发育限制减弱,N、P元素含量均呈现显著的线性下降状态.降水量变化对齿肋赤藓地上部分功能性物质N、P含量的显著性影响,可能由于齿肋赤藓常以斑块状聚集生长,在斑块水平上对环境变化则表现出种群的适应能力.而就齿肋赤藓地下部分的N、P养分含量而言,其变化无明显规律,可以认为该植物地下部分的养分含量在斑块水平上是稳定的.本试验发现,随着年降水量的增加,地上、地下部分的N∶P呈线性增大,一方面原因为降水量的增加减少了齿肋赤藓的水分胁迫,地上部分的N、P元素含量显著下降且P元素含量下降得更快,导致地上部分N∶P呈线性增大;另一方面原因为年降水量的增加也会增加土壤中硝酸还原酶等的活性,可利用养分增加,齿肋赤藓地下部分既可以吸收大量土壤中可利用氮,同时能够获得来源于地上部分的氮,导致地下部分N∶P呈线性增大. ...
Drought effect on plant nitrogen and phosphorus:a metaanalysis
1
2014
... 本研究中,年降水量最小的样点(G1)齿肋赤藓植物地上部分N、P元素含量最高,可能的原因是:①在干旱条件下,植物需要提高细胞内部的渗透压,增强对体内水分的保持、提高水分利用效率,适应干旱环境[38];②干旱条件可能会降低养分的质量流量或扩散率,从而限制养分在地上部与地下部之间的运输[39],导致地上、地下的N、P元素含量差异较大.随着年降水量的增加,水分胁迫作用减缓,养分的扩散率增加,植物地上部分发育限制减弱,N、P元素含量均呈现显著的线性下降状态.降水量变化对齿肋赤藓地上部分功能性物质N、P含量的显著性影响,可能由于齿肋赤藓常以斑块状聚集生长,在斑块水平上对环境变化则表现出种群的适应能力.而就齿肋赤藓地下部分的N、P养分含量而言,其变化无明显规律,可以认为该植物地下部分的养分含量在斑块水平上是稳定的.本试验发现,随着年降水量的增加,地上、地下部分的N∶P呈线性增大,一方面原因为降水量的增加减少了齿肋赤藓的水分胁迫,地上部分的N、P元素含量显著下降且P元素含量下降得更快,导致地上部分N∶P呈线性增大;另一方面原因为年降水量的增加也会增加土壤中硝酸还原酶等的活性,可利用养分增加,齿肋赤藓地下部分既可以吸收大量土壤中可利用氮,同时能够获得来源于地上部分的氮,导致地下部分N∶P呈线性增大. ...
内蒙古典型草原植物叶片碳氮磷化学计量特征的季节动态
1
2020
... 通过对各样点地上、地下部分的相关性分析发现:无论是地上部分还是地下部分,齿肋赤藓N、P元素含量均为显著正相关,这种相关性反映了植物体内营养元素间具有较强的线性关系[40],本文结果进一步验证了此观点.限制元素N、P呈异速关系是植物适应环境的重要策略[41-42],齿肋赤藓同部位N、P含量均表现出显著的异速关系,说明齿肋赤藓各部位N、P比例相对稳定.在众多大尺度的调查、养分添加的研究中,所得N和P的幂指数比均小于1,且斜率间无显著差异,齿肋赤藓植物养分吸收、利用和分配关系随环境改变呈现不显著的差异性变化,具有一定的保守性[35].本研究中齿肋赤藓植物地上部分N-P幂指数计量比均小于1,其结果均接近前人所提出的植物叶片2/3[15]或3/4[16]N-P幂指数计量关系;齿肋赤藓植物地下部分N-P幂指数计量关系均大于1,说明齿肋赤藓植物地下部分P含量占比相较于N含量高,这可能与藓类植物能大量吸收土壤P元素的特性有关[43]. ...
新疆45种荒漠植物粗根碳、氮、磷计量特征及其与环境的关系
1
2019
... 通过对各样点地上、地下部分的相关性分析发现:无论是地上部分还是地下部分,齿肋赤藓N、P元素含量均为显著正相关,这种相关性反映了植物体内营养元素间具有较强的线性关系[40],本文结果进一步验证了此观点.限制元素N、P呈异速关系是植物适应环境的重要策略[41-42],齿肋赤藓同部位N、P含量均表现出显著的异速关系,说明齿肋赤藓各部位N、P比例相对稳定.在众多大尺度的调查、养分添加的研究中,所得N和P的幂指数比均小于1,且斜率间无显著差异,齿肋赤藓植物养分吸收、利用和分配关系随环境改变呈现不显著的差异性变化,具有一定的保守性[35].本研究中齿肋赤藓植物地上部分N-P幂指数计量比均小于1,其结果均接近前人所提出的植物叶片2/3[15]或3/4[16]N-P幂指数计量关系;齿肋赤藓植物地下部分N-P幂指数计量关系均大于1,说明齿肋赤藓植物地下部分P含量占比相较于N含量高,这可能与藓类植物能大量吸收土壤P元素的特性有关[43]. ...
古尔班通古特沙漠中4种荒漠草本植物的生物量与化学计量特征
1
2019
... 通过对各样点地上、地下部分的相关性分析发现:无论是地上部分还是地下部分,齿肋赤藓N、P元素含量均为显著正相关,这种相关性反映了植物体内营养元素间具有较强的线性关系[40],本文结果进一步验证了此观点.限制元素N、P呈异速关系是植物适应环境的重要策略[41-42],齿肋赤藓同部位N、P含量均表现出显著的异速关系,说明齿肋赤藓各部位N、P比例相对稳定.在众多大尺度的调查、养分添加的研究中,所得N和P的幂指数比均小于1,且斜率间无显著差异,齿肋赤藓植物养分吸收、利用和分配关系随环境改变呈现不显著的差异性变化,具有一定的保守性[35].本研究中齿肋赤藓植物地上部分N-P幂指数计量比均小于1,其结果均接近前人所提出的植物叶片2/3[15]或3/4[16]N-P幂指数计量关系;齿肋赤藓植物地下部分N-P幂指数计量关系均大于1,说明齿肋赤藓植物地下部分P含量占比相较于N含量高,这可能与藓类植物能大量吸收土壤P元素的特性有关[43]. ...
长白山暗针叶林苔藓植物在养分循环中的作用
1
2005
... 通过对各样点地上、地下部分的相关性分析发现:无论是地上部分还是地下部分,齿肋赤藓N、P元素含量均为显著正相关,这种相关性反映了植物体内营养元素间具有较强的线性关系[40],本文结果进一步验证了此观点.限制元素N、P呈异速关系是植物适应环境的重要策略[41-42],齿肋赤藓同部位N、P含量均表现出显著的异速关系,说明齿肋赤藓各部位N、P比例相对稳定.在众多大尺度的调查、养分添加的研究中,所得N和P的幂指数比均小于1,且斜率间无显著差异,齿肋赤藓植物养分吸收、利用和分配关系随环境改变呈现不显著的差异性变化,具有一定的保守性[35].本研究中齿肋赤藓植物地上部分N-P幂指数计量比均小于1,其结果均接近前人所提出的植物叶片2/3[15]或3/4[16]N-P幂指数计量关系;齿肋赤藓植物地下部分N-P幂指数计量关系均大于1,说明齿肋赤藓植物地下部分P含量占比相较于N含量高,这可能与藓类植物能大量吸收土壤P元素的特性有关[43]. ...
黄土丘陵区刺槐叶片-土壤-微生物碳氮磷化学计量学及其稳态性特征
1
2019
... 植物地上与地下构件的许多性状间如比叶面积与比根长、叶片质量比与细根质量比等都存在一定的异速关系.这种异速关系为不同器官生长发育的协调性提供重要依据,但各构件间的营养成分可能会受到植物自身及环境的相互作用的高度影响.地上、地下部分的N-P幂指数关系呈现显著性差异的原因可能是由于齿肋赤藓地上部分是光合、呼吸以及生长的主要器官,而地下部分则是由假根和茎组成,作为非维管束植物,假根和茎经沙埋后主要起到固定和少量水分和养分运输的作用.不同降水梯度下齿肋赤藓植物同一部位的N-P幂指数计量关系无显著变化,因此认为齿肋赤藓植物对N-P异速关系指数对环境变化响应不敏感,说明齿肋赤藓氮磷养分化学计量比较高内稳性[44].本研究结果表明在同一降水量条件下齿肋赤藓植物不同部位的N-P异速关系指数呈显著不同,说明齿肋赤藓植物能够通过调节不同部位氮磷元素的吸收、分配来适应荒漠极端环境的改变. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}