巴丹吉林沙漠东缘主要灌木数量特征及空间分布
姜亚东 , 1 , 吕世杰 , 2 , 刘红梅 3 , 娜仁花 4 , 刘昕宇 5
1.内蒙古自治区地质调查研究院,内蒙古 呼和浩特 010018
2.内蒙古农业大学 理学院,内蒙古 呼和浩特 010018
3.内蒙古自治区林业科学研究院,内蒙古 呼和浩特 010010
4.内蒙古自治区自然资源督察保障中心,内蒙古 呼和浩特 010020
5.广西师范大学,广西 桂林 541006
Analysis of quantitative characteristics and spatial distribution for main shrubs on the eastern edge of the Badain Jaran Desert
Jiang Yadong , 1 , Lv Shijie , 2 , Liu Hongmei 3 , Narenhua 4 , Liu Xinyu 5
1.Inner Mongolia Autonomous Region Geological Survey Institute,Hohhot 010018,China
2.Science Faculty,Inner Mongolia Agricultural University,Hohhot 010018,China
3.Inner Mongolia Autonomous Region Forestry Sciences Institute,Hohhot 010010,China
4.Inner Mongolia Autonomous Region Natural Resources Supervision and Protection Center,Hohhot 010020,China
5.Guangxi Normal University,Guilin 541006,Guangxi,China
通讯作者: 吕世杰(E-mail: lshj123@imau.edu.cn )
收稿日期: 2023-02-27
修回日期: 2023-04-03
基金资助:
内蒙古自治区自然科学基金项目 . 2019MS03035 . 2021MS03042
Received: 2023-02-27
Revised: 2023-04-03
作者简介 About authors
姜亚东(1979—),男,内蒙古呼和浩特人,硕士研究生,主要从事生态保护与修复研究E-mail:838166735@qq.com
, E-mail:838166735@qq.com
摘要
荒漠植被是极端大陆性干旱气候下形成的以灌木、半灌木为主的植被类型,基于对严酷环境的耐受能力和生态适应过程,形成了特有的表型可塑性和生态响应策略。以巴丹吉林沙漠东缘硬质沙地植被为研究对象,采用样地和尺度构造双因素试验设计,对比分析了主要种群适应基础和空间分布状态。结果表明:梭梭(Haloxylon ammodendron )耐贫瘠、抗风沙使得冠幅直径和高度表现较高,白刺(Nitraria tangutorum )通过捕获流沙增加自己的株丛茎,红砂(Reaumuria songarica )种子萌发和幼苗定植能力强导致出现频次较高;三者在植物群落地位和作用依次降低。白刺和梭梭种群空间分布主要以母株为中心,沿着合成风向扩散;红砂种群空间分布尽管受结构性因素影响,但是集中连片分布形式较弱;3个植物种群整体空间分布主要受结构因素影响,呈现出斑块镶嵌分布,物种界限消失。梭梭、白刺和红砂植物种群对环境的适应基础差异能够通过数量特征相关指标体现,其在植物群落中的地位和作用依次降低;单一种群空间分布能够反映母株为中心的扩散形式,复合种群空间分布反映的空间协同适应情况,呈现出斑块镶嵌分布,物种界限消失。
关键词:
巴丹吉林沙漠 主要种群 数量特征 空间分布
Abstract
The Badain Jaran Desert is the third largest desert in China, which typical desert vegetation is a shrub and semi-shrub dominated vegetation type formed under extreme continental arid climate, and its tolerance to harsh environments and ecological adaptation process have formed unique phenotypic plasticity and ecological response strategies. In order to explore the adaptive basis and spatial distribution status of major populations, this study focused on the vegetation grown on the hard surface of sandy land on the eastern edge of the Badain Jaran Desert, using a two-factor experimental design of sample area and scale construction, combined with comparative and geostatistical analysis, the preliminary conclusions are as follows. Haloxylon ammodendron is resistant to barrenness and sandstorms, resulting in a high crown diameter and height performance. Nitraria tangutorum increases its own diameter of bunch by capturing flowing sand. Reaumuria songarica has strong seed germination and seedling planting ability, resulting in a higher frequency of occurrence. The status and role of the three species in the plant community decrease in order. The spatial distribution of N. tangutorum and H. ammodendron populations is mainly centered on the mother plant and spreads along the prevailing wind direction, while the spatial distribution of R.songarica populations is weakly concentrated even under the effect of structural factors. The overall spatial distribution of the three plant populations is mainly affected by structural factors, presenting a mosaic distribution pattern, and the boundaries between species disappear. In summary, the differences in the adaptation basis of H. ammodendron , N. tangutorum , and R. songarica populations to the environment can be reflected by quantitative characteristic indicators. Their status and role in the plant community decrease in order. The spatial distribution of a single population can reflect the diffusion pattern centered on the mother plant, and the spatial distribution of composite populations reflects the spatial coordination and adaptation situation, presenting a mosaic distribution pattern, and the boundaries between species disappear.
Keywords:
Badain Jaran Desert main populations quantitative characteristics spatial distribution
本文引用格式
姜亚东, 吕世杰, 刘红梅, 娜仁花, 刘昕宇. 巴丹吉林沙漠东缘主要灌木数量特征及空间分布 . 中国沙漠 [J], 2023, 43(3): 295-304 doi:10.7522/j.issn.1000-694X.2023.00027
Jiang Yadong, Lv Shijie, Liu Hongmei, Narenhua , Liu Xinyu. Analysis of quantitative characteristics and spatial distribution for main shrubs on the eastern edge of the Badain Jaran Desert . Journal of Desert Research
0 引言
巴丹吉林沙漠东缘荒漠区气候干旱,植被稀疏,以旱生、超旱生的灌木和小灌木为主[1 -2 ] 。这些灌木种群是巴丹吉林沙漠防风固沙的主要植物,对改善生态环境具有重要意义[1 ] 。红砂(Reaumuria songarica )植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] 。白刺(Nitraria tangutorum )能够构造自己的灌丛小生境,容易形成荒漠白刺土丘独有的自然景观,在干旱少雨时主要利用深层地下水,当降雨量超过年平均降水量时会及时利用地表水[6 -8 ] ;梭梭(Haloxylon ammodendron )对于土壤养分的需求低于红砂和白刺,所以可以在半流动沙丘上存活,成为沙漠流沙治理的先锋植物种群[2 ,9 -10 ] 。3种主要灌木具有不同适应策略,其数量特征变化以及空间分布状态必然存在自己的独特性,因此掌握主要灌木种群数量特征和空间分布差异不仅有利于揭示其适应状态,更有利于利用这一适应基础为荒漠化防治提供理论依据和数据支撑。
刘红梅等[2 ,10 ] 研究巴丹吉林半流动沙地白刺、红砂和梭梭植物种群空间分布关系时发现,3个种群存在小尺度竞争、大尺度协同的空间镶嵌分布形式,且整体呈现以梭梭种群为主导的植被扩散格局和发展趋势。梭梭的空间分布与半流动沙丘微地形存在明显相关关系,且存在一定程度的沙埋适应[9 -12 ] 。白刺植物种群空间分布在景观上表现出密度大而体型小的特点,灌丛沙堆处于发育的中早期阶段[13 ] ;红砂植物种群的密度、高度和冠幅空间异质性处于中等强度,且其密度空间分布与细沙空间分布紧密相关[14 ] 。不同植物种群空间分布状态及其数量特征不仅与其自身适应性有关,也与生境砂层粒径存在相关性;然而这些研究均是基于流动、半流动沙地展开的研究,其在硬质沙地上相关研究鲜有报道。
本研究在巴丹吉林沙漠东缘硬质沙地对主要灌木种群展开研究,采用样地调查方法测定其高度、冠幅直径、频次以及株丛径,结合对比分析和地统计分析方法,拟解决以下问题:①植被构成的主要灌木有哪些?②其基本数量特征和空间分布伴随尺度效应如何变化?③种群间的数量特征和空间分布差异具有什么指示意义?相关问题的解决不仅能够阐明巴丹吉林沙漠东缘灌木种群适应基础和适应能力差异,也能够为荒漠区植被恢复与重建提供理论基础和数据支撑。
1 方法
1.1 试验设计及主要灌木种群
试验地行政区划属于阿拉善右旗塔木素苏木格勒图嘎查的朝恒扎干(塔木素野生肉苁蓉及梭梭产籽基地),地理位置为40°33.059′—40°35.531′N、103°25.162′—103°27.546′E,海拔1 255.8~1 264.3 m。研究区属典型温带干旱荒漠性气候,年降水量40~80 mm,集中于5—9月,年潜在蒸发量达到年降水量的40倍以上,干燥度7~12;年平均气温8~8.9 ℃,绝对最高气温37~43 ℃,平均无霜期150 d;风沙频繁,年均风速4 m·s-1 ;研究区流沙带以新月形沙丘和简单横向沙丘为主,沙丘主要走向为SW-NE。自然植被稀少,种属贫乏、覆盖率低,地表裸露[9 -10 ,14 ] 。
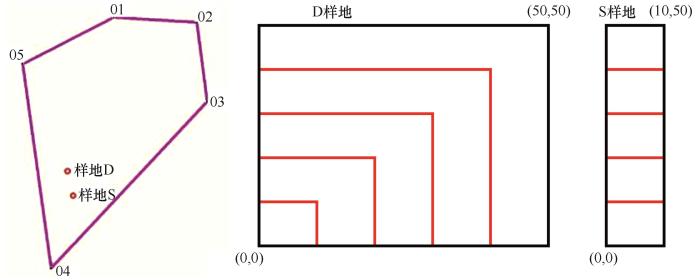
在研究区内硬质沙地选择2个小样地(图1 ),其中一个50 m×50 m(标记为D样地),另一个10 m×50 m(标记为S样地),D样地居于S样地和流动沙地之间,D样地和S样地均为硬质沙地。两样地物种接近,且为该区域硬质沙地代表性植被,分布面积较广。两样地西南角定义为(0,0)坐标点,分别向北向东拉样线,构建平面直角坐标系,即样地图形上下左右分别对应北南西东地理方位。在样地中分种测定每一株灌木的高度(cm)、冠幅直径(cm)、株丛径(cm),同时记录该灌木相对位置(x ,y )。根据样地划分尺度,D样地划分为10 m×10 m、20 m×20 m、30 m×30 m、40 m×40 m、50 m×50 m尺度,S样地划分为10 m×10 m、10 m×20 m、10 m×30 m、10 m×40 m、10 m×50 m尺度,然后两个样地与5个尺度(统一命名为Sca1~Sca5)构造双因素无重复试验设计。
图1
图1
小样地及尺度划分
Fig.1
The small sample plots and scale division
对硬质沙地2个小样地的灌木种群进行统计,其数量特征见表1 。由表1 可以看到,主要灌木种群为白刺、红砂和梭梭,霸王(Zygophyllum xanthoxylon )偶尔出现。
1.2 数据分析
R F ¯ = F i / ∑ i = 1 n F (1)
R H ¯ = H ¯ i / ∑ i = 1 n H ¯ (2)
R C ¯ = C ¯ i / ∑ i = 1 n C ¯ (3)
R D ¯ = D ¯ i / ∑ i = 1 n D ¯ (4)
I V P = R F ¯ + R H ¯ + R C ¯ + R D ¯ 4 × 100 % (5)
式中:F ¯ H ¯ C ¯ D ¯ R F ¯ R H ¯ R C ¯ R D ¯ n 为出现的灌木种群数;IVP 代表种群的重要值。
H ' = H s / ∑ s = 1 t H (6)
C ' = C s / ∑ s = 1 t C (7)
D ' = D s / ∑ s = 1 t D (8)
I V I = H ' + C ' + D ' 3 × 100 % (9)
式中:H C D H ' C ' D ' t 为出现的灌木个体总数;IVI 代表个体的重要值。
采用双因素无重复试验设计的方差分析模型检验灌木种群基本数量特征及其重要值的样地间和尺度间差异显著性,同时记录模型拟合率判断线性模型的可加性。分析过程在SAS 9.2中调用ANOVA过程,多重比较结果选用Duncan法。分析结果在Microsoft Excel 2019中绘制成表格,同时对同一灌木种群不同尺度下的变化规律在Sigmaplot 14.0 (Systat Software Inc.)中绘制图形。
分别对两个小样地白刺、红砂、梭梭种群及其整体进行半方差函数分析(数据采用个体重要值指数),分析过程在GS+9.0软件中进行,并在Microsoft Excel 2019中整理函数参数呈表格形式[15 ] ,然后再Surfur 12.0软件中绘制各种群及其整体空间分布状态[10 ] 。
2 结果与分析
2.1 不同灌木基本数量特征
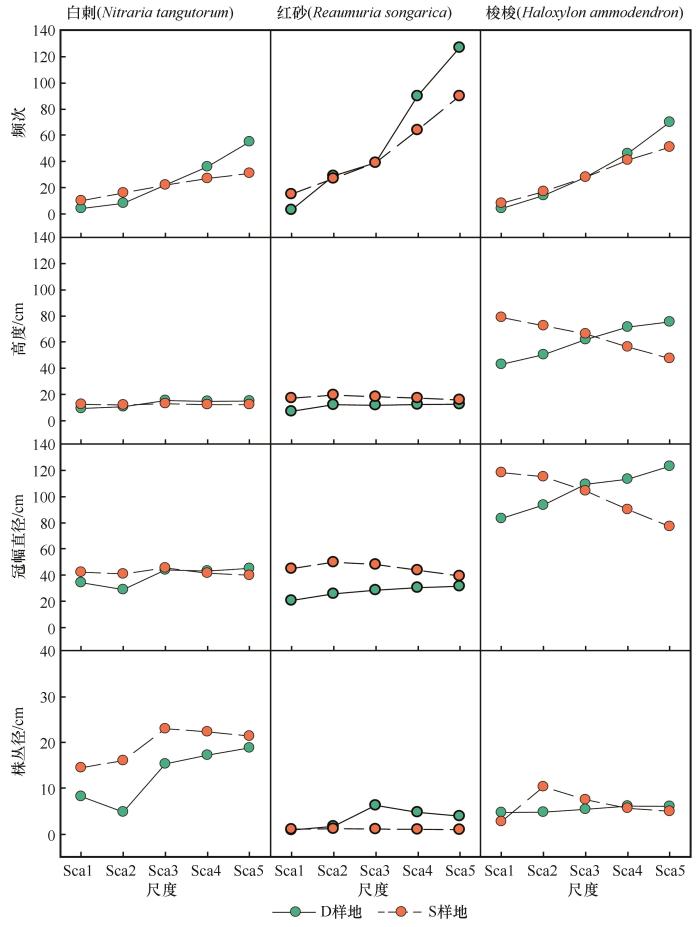
白刺种群在不同尺度上频度存在较弱的变化趋势(0.05≤P <0.15,表2 ),即伴随尺度增大白刺种群频次增加的幅度较小(图2 ),红砂和梭梭植物种群伴随尺度增大频次显著增加(P <0.05,表2 ,图2 )。3个植物种群尺度效应远远大于样地效应(平方和差异),说明正方形样地(D)和长方形样地(S)之间频次差异较小,反映出靠近流动沙地方向的样地物种出现频次较低,而远离流动沙地灌木种群出现频次较高,因为二者之间存在面积5倍关系。
图2
图2
灌木种群基本数量特征伴随尺度的变化
Fig.2
The change rule of the basic quantitative characteristics of different shrub populations with specific scales
在高度上,只有红砂植物种群高度在D样地和S样地之间存在显著差异(P <0.05),S样地红砂高度始终高于D样地。在冠幅直径上,同样只有红砂冠幅直径在D样地和S样地之间存在显著差异,S样地红砂冠幅直径始终高于D样地。在株丛径上,白刺种群无论是在样地之间,还是在尺度之间,均存在显著性差异,表现出S样地株丛径始终高于D样地,且30 m以下的尺度波动较大,30 m以上的尺度趋于接近;红砂植物种群仍然存在较弱的变化趋势,即D样地株丛径高于S样地(0.05≤P <0.15)。
综合来看,样地和尺度差异均不显著的方差分析模型主要是线性可加性不存在(拟合率较小,小于70%),当存在样地或者尺度之间的差异时,方差分析模型均具有较好的拟合效果,拟合率高于70%。同时可以看出,样地之间的差异主要体现在红砂植物种群,而尺度差异则体现在白刺和梭梭种群。红砂在样地内的频次较其他种群高,梭梭植物种群的高度和冠幅直径较其他种群高,而白刺植物种群株丛径则更占优势。
2.2 不同灌木在植物群落中的地位和作用
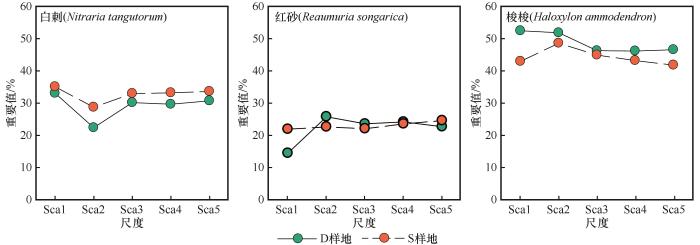
在巴丹吉林沙漠东缘硬质沙地上,白刺植物种群重要值无论是在尺度变化上,还是在样地之间,均存在显著性差异(表3 ),即S样地白刺重要值高于D样地,且二者具有相同的变化规律(图3 )。红砂植物种群无论是在尺度之间,还是在样地之间,均不存在显著性差异;二者之间的重要值在30 m尺度之下,呈现交错状态,当大于30 m时,样地之间的重要值几乎一致且比较平稳。梭梭植物种群重要值在样地之间存在显著性差异,但尺度之间差异不显著(P >0.05%),即D样地梭梭重要值始终高于S样地。
图3
图3
灌木种群重要值的变化
Fig.3
The change rule of important values of different shrub populations
综合来看,梭梭种群在植物群落的重要值最大,对植物群落的结构和功能具有主导地位,其次是白刺植物种群,最后是红砂植物种群。梭梭植物种群重要值较大,主要受冠幅直径和高度影响,样地间差异是数量特征的整体结果;白刺重要值次之,主要受株丛径的影响较大,且株丛径在样地之间的差异导致重要值在样地之间也产生显著性差异。红砂植物种群重要值相对较小,但其样地出现频次较高,说明尽管红砂植物种群在严酷环境下能够进行有效定植,但是其株丛低矮瘦小,难以对植物群落结构和功能产生较大的影响。
2.3 不同灌木在植物群落中的空间分布特征
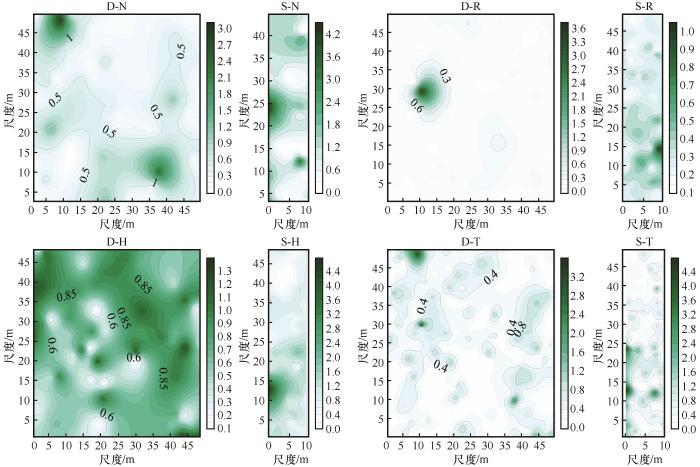
白刺植物种群在两个样地的空间分布主要受结构性因素影响,D样地和S样地的结构比分别为0.999和0.948,最适拟合模型依次为高斯模型(Gaussian)和球形模型(Spherical);自相关尺度分别为2.41和1.95,二者相差不大(表4 )。红砂植物种群在D样地和S样地其空间分布主要受结构性因素影响,结构比分别为0.911和0.872,最适拟合模型分别为指数模型(Exponential)和高斯模型;自相关尺度S样地远大于D样地,分别为51.00和2.97。梭梭植物种群空间分布在D样地处于完全随机状态(受随机因素控制),在S样地受结构因素和随机因素共同影响,结构比分别为0.000和0.700;最适拟合模型为线性模型(Linear)和指数模型;空间自相关尺度S样地大于D样地,分别为51.00和31.60。3个植物种群整体上在D样地受结构性因素影响主导,在S样地受随机因素和结构因素共同决定,结构比分别为0.947和0.521,最适拟合模型分别为球形模型和高斯模型;自相关尺度S样地远大于D样地,分别为16.43和1.40。具体分析过程数据转换形式见表4 。
D样地白刺植物种群空间分布在东南和西北区域具有重要值较大的分布区,并以这两分布区形成连续的空间分布条带;在S样地,白刺在西部中间区域具有较大的重要值分布区,且在南北区域也存在连片的重要值较大的空间分布区,使得其空间分布状态形成南部、中部和北部3个重要值较大分布区(图4 )。红砂在D样地西北部区域存在一个重要值较大的分布区,除此之外,红砂在样地内的分布比较分散;在S样地,红砂重要值较大的分布区域集中在南部区域。梭梭在D样地的空间分布比较集中连片,整体呈现由西北向东南的主条带分布形式;在S样地,梭梭重要值较大的空间分布区域在西南部,北部区域重要值较小。当将白刺、红砂和梭梭3个植物种群进行整体空间分布分析时发现,D样地白刺和梭梭空间分布的连续性消失,取而代之的是空间小斑块镶嵌分布,且这些小斑块没有种群限制,说明斑块内种群协同空间分布形式居于主导地位;在S样地,白刺、红砂和梭梭3个植物种群集中分布区域消失,取而代之的是以原集中分布区中心形成的高重要值变化区,且相互之间的联结性较弱。整体来看,当以单一植物种群分析空间分布状态时,其往往形成集中连片区域,这反映了灌木种群以母株为中心的空间扩散形式;当以3个植物种群整体分析,其空间分布呈现出斑块镶嵌分布,物种界限消失,反映出物种协同作用适应严酷环境的一种空间分布状态。
图4
图4
不同灌木种群及其整体空间分布状态(重要值)
D-N、D-R、D-H、D-T、S-N、S-R、S-H、S-T中,D代表双向样地,S代表单向样地,N代表白刺,R代表红砂,H代表梭梭,T代表3个种群整体
Fig.4
The spatial distribution of different shrub populations and the whole
3 讨论
3.1 灌木数量特征变化与其适应性
“压力优势假说”[16 -18 ] 认为,环境(非生物因素)过滤因子在更大环境梯度上对于种群构建植物群落具有重要意义,而竞争(生物因素)过滤因子在非生物压力较低的情况下更为重要。“唯利假说”[19 -20 ] 也预测了这种类似的现象,即当生存条件不太有利时,环境过滤能力更强。白刺、红砂和梭梭尽管生活型相同,均属于灌木,具备应对高风沙、高温、极端干旱严酷环境的能力,但是其适应基础存在较大的差异。对于白刺种群而言,其植株低矮,但是能够捕获流沙对自己进行掩埋形成沙丘,然后通过有性和无性繁殖扩大株丛,导致其株丛径大小本质上是沙丘的大小,尽管试验样地没有出现白刺为主的高大沙丘连续分布的地貌景观,但是其适应基础没有改变,因此其数量特征株丛径大小在3个种群占据优势[6 -8 ] 。红砂植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] ;而研究区红砂种群株丛几乎没有形成沙丘,但是其出现频次在3个种群最高,印证了红砂对严酷环境具有高度的适应性,能够生长在砾石基质沙地上,由于其相互之间没有沙丘连接,均是具有主干形式的小灌木株丛,说明其有性繁殖的种子萌发能力和幼苗成功定植能力均比较强。梭梭植物种群对于土壤养分的需求低于红砂和白刺,所以能够在半流动沙地生存[2 ,9 -10 ] ,然而本研究区属于硬质沙地,梭梭植物种群不仅能够存活,而且具有相对较高的株高和冠幅直径(图2 ),说明在有限的土壤养分资源情况下,梭梭能够快速生长,体现在株高和冠幅直径上,同时又因本区域属于沙漠边缘区域,风大沙多,所以梭梭种群高大株型对风沙的耐受能力也要优于白刺和红砂。这也间接揭示了巴丹吉林沙漠东缘植被以梭梭主导的扩散格局和发展趋势[10 ] ,本研究重要值也显示梭梭植物种群在群落中的地位和作用最大,而白刺植物种群在植物群落中的地位和作用次之,红砂在植物群落中地位和作用在3个种群中最小;进而也证明“压力优势假说”和“唯利假说”导致不同灌木同样在干旱环境条件下,适应策略不同,进而表现在不同数量特征指标上。
一般认为,植物的高度和冠幅直径反映了植物种群对光的捕获能力[21 ] ,而该区域植被稀疏,群落结构简单,且光照周期长,所以主要植物种群株高和冠幅直径并不能反映其对光的敏感性和适应性,因此植物群落物种构成以及地理区域不同,可能会导致植物种群同一指标反映的适应基础不同。密度或者频次,往往反映了植物种群繁衍的能力,进而对于多年生物种判断有性繁殖和无性繁殖的适应能力[22 ] 。高强度干扰可导致有性繁殖能力下降,但也会导致有性繁殖质量增加[23 ] 。本研究红砂种群具有较强的有性繁殖能力,但是究竟是有性繁殖质量增加所致,还是有性繁殖能力不变或者适应严酷环境的结果有待于进一步研究,进而白刺以母株为中心捕获流沙,是否获得无性繁殖体扩展导致其具有较大的株丛径也需要进一步明确。种群生物量反映了植物种群在一定空间内,通过繁殖体获取资源的能力[24 ] ,尽管本研究没有相关研究,但是从重要值角度进行了初步分析,因重要值反映了其在植物群落的地位和作用,也反映了其在植物群落的功能强弱,所以初步判定梭梭、白刺和红砂在群落中的整体作用依次降低。
3.2 灌木空间分布状态与其适应性
结构比反映空间变量空间分布的主要影响因素。当结构比>75%,空间变量空间自相关性较强,主要受结构因素影响;结构比位于25%~75%,空间变量空间自相关居于中等强度,主要受结构因素和随机因素共同控制;当结构比<25%,空间变量空间自相关性最弱,主要受随机因素影响[25 ] 。白刺和红砂植物种群均主要受结构因素影响,空间自相关尺度在不同样地的差别较大,特别是红砂植物种群差别更大,这可能与样地面积有关。贾利娟等[26 ] 研究荒漠草原时指出,样地样方法的样方数最好大于64个,样地10 m间隔取样最小面积应大于6 400 m2 。本研究是样地连续取样,且划分5个尺度,发现30 m尺度可能是该研究区最佳样地大小,既节约人力、财力和物力,也能有效地获得种群空间分布相对稳定的信息(图2 ),同时D样地设计对于种群监测更为有利[27 ] 。白刺和梭梭的空间分布主要呈集中连片分布,这与刘红梅等[2 ,10 ] 前期的研究结果一致,但是整体上的空间分布在流动沙地呈大的斑块形式,而本研究尽管分析方法不同,二者仍然存在这一变化形式,但是红砂种群却存在较大的差异,主要是取样方法和研究区土壤基质差异所致。当对3个种群整体进行空间分布分析时,S样地随机性和结构性因子共同控制灌木空间分布,而D样地则显示结构性因素占据主导地位(表4 ),尽管二者决定因素不同,但是空间分布状态几乎一致,即呈现出斑块镶嵌分布,物种界限消失,这说明物种间采用协同方式来适应严酷环境[8 ,28 ] 。
在研究区域,随机因素主导的空间分布状态主要指植物种群以母株为中心形成的集中连片分布形式,具有这种分布形式的种群为白刺和梭梭种群,其中梭梭的分布方向由西北向东南形成的条带分布符合该区域常年的合成风向,而梭梭种子传播在盛行风向具有较强的方向依赖性,所以这种基于合成风向的分布条带和其他风向形成的随机过程,导致梭梭种群形成当前的空间分布状态[2 ,29 -30 ] 。结构性因素决定的空间分布状态主要是指由研究区气候、水文、土壤基质等决定的空间分布形式[31 ] ,D样地3个植物种群空间分布受结构性因素影响(表4 ),但是其空间自相关尺度仅为1.40,说明其空间分布的复杂性较低,空间异质性较弱。结合植被稀疏低矮、物种匮乏[2 ,9 ] ,可知其植物群落结构简单,进而在季相上形成灌木半灌木功能群构成的单一功能群灌丛群落,受“夏雨型”植物层片影响,虫实(Corispermum hyssopifolium )和雾冰藜(Grubovia dasyphylla )等大量出现,丰富了群落物种丰富度,但植物群落的简单结构变化不大,因此该群落的抗干扰能力和干扰后的恢复能力很弱,需要加强保护[2 ,32 -33 ] 。
4 结论
巴丹吉林沙漠东缘植物群落物种构成简单,梭梭、红砂和白刺是主要种群,且梭梭对植物群落的调控作用较大。
梭梭适应基础是耐贫瘠、抗风沙,体现在冠幅直径和高度指标上;白刺的适应基础是捕获流沙的能力,体现在株丛径上;红砂适应基础是种子萌发和幼苗定植能力,体现在出现频次上。3个植物种群在植物群落的地位和作用依次降低。
梭梭和白刺空间分布均呈现以母株为中心的扩散形式,且梭梭扩散趋势与研究区合成风向相关;3个植物种群整体空间分布受结构性因素主导,呈现出斑块镶嵌分布状态,物种界限消失。
参考文献
View Option
[1]
王蒙 ,逯军峰 ,付鹏 ,等 巴丹吉林沙漠周边地区土壤养分和粒度特征
[J].中国沙漠 ,2022 ,42 (5 ):232 -244 .
[本文引用: 2]
[2]
刘红梅 ,吕世杰 ,刘清泉 ,等 塔木素天然梭梭林空间分布及其与林地地貌起伏变化的关系
[J].生态学杂志 ,2015 ,34 (9 ):2415 -2423 .
[本文引用: 8]
[3]
刘家琼 ,邱明新 ,蒲锦春 ,等 我国荒漠典型超旱生植物:红砂
[J].植物学报 ,1982 ,24 (5 ):485 -488 .
[本文引用: 2]
[4]
曾彦军 ,王彦荣 ,张宝林 ,等 红砂种群繁殖特性的研究
[J].草业学报 ,2002 ,11 (2 ):66 -71 .
[5]
朱恭 ,李正平 ,王万鹏 ,等 红砂属植物研究进展
[J].甘肃林业科技 ,2004 ,29 (3 ):1 -6 .
[本文引用: 2]
[6]
杜建会 ,严平 ,展秀丽 ,等 民勤绿洲白刺灌丛沙堆不同演化阶段表面抗蚀性及其影响因素
[J].应用生态学报 ,2008 ,19 (4 ):71 -76 .
[本文引用: 2]
[7]
朱雅娟 ,卢琦 ,吴波 ,等 增雨对巴丹吉林沙漠东南缘白刺水分利用的影响
[J].应用生态学报 ,2013 ,23 (1 ):45 -52 .
[8]
丁延龙 白刺灌丛沙堆演化对地表蚀积的影响及其作用机制
[D].呼和浩特 :内蒙古农业大学 ,2019 .
[本文引用: 3]
[9]
王猛 ,汪季 ,蒙仲举 ,等 巴丹吉林沙漠东缘天然梭梭种群空间分布异质性
[J].生态学报 ,2016 ,36 (13 ):4055 -4063 .
[本文引用: 5]
[10]
刘红梅 ,吕世杰 ,刘青泉 ,等 巴丹吉林沙漠东缘主要植物种群空间分布关系
[J].生态学杂志 ,2021 ,40 (4 ):959 -967 .
[本文引用: 7]
[11]
Kang J J Wang S M Zhao M et al ffects of separate and mutual application of Na and Si on drought resistance of Haloxylon ammodendron
[J].Sciences in Cold and Arid Regions ,2012 ,4 (6 ):490 -497 .
[12]
He P Li Y F Xu N et al Predicting the suitable habitats of parasitic desert species based on a niche model with Haloxylon ammodendron and Cistanche deserticola as examples
[J].Ecology and Evolution ,2021 ,11 (24 ):17817 -17834 .
[本文引用: 1]
[13]
马瑞 ,赵锦梅 ,马彦军 ,等 巴丹吉林沙漠南缘近流沙区白刺灌丛沙堆形态特征与空间分布
[J].水土保持学报 ,2021 ,35 (4 ):217 -221 .
[本文引用: 1]
[14]
刘红梅 ,吕世杰 ,任倩楠 ,等 红砂植物种群空间分布与沙子粒级及地貌起伏之间的关系
[J].生态环境学报 ,2020 ,29 (10 ):1971 -1978 .
[本文引用: 2]
[15]
Lv S J Yan B L Wang Z W et al Grazing intensity enhances spatial aggregation of dominant species in a desert steppe
[J].Ecology and Evolution ,2019 ,9 (10 ):6138 -6147 .
[本文引用: 1]
[16]
Swenson N G Enquist B J Ecological and evolutionary determinants of a key plant functional trait: Wood density and its community-wide variation across latitude and elevation
[J].American Journal of Botany ,2007 ,94 (3 ):451 -459 .
[本文引用: 1]
[17]
Coyle J R Halliday F W Lopez B E et al Using trait and phylogenetic diversity to evaluate the generality of the stress-dominance hypothesis in Eastern North American tree communities
[J].Ecography ,2014 ,37 (9 ):814 -826 .
[18]
Balazs K R Munson S M Havrilla C A et al Directional selection shifts trait distributions of planted species in dryland restoration
[J].Journal of Ecology ,2022 , 110 (3 ):540 -552 .
[本文引用: 1]
[19]
Swenson N G Enquist B J Pither J et al The biogeography and filtering of woody plant functional diversityin North and South America
[J].Global Ecology and Biogeography ,2012 ,21 (8 ):798 -808 .
[本文引用: 1]
[20]
Wieczynski D J Boyle B Buzzard V et al Correction: climate shapes and shifts functional biodiversity in forests worldwide
[J].Proceedings of the National Academy of Sciences of the United States of America ,2019 ,116 :587 -592 .
[本文引用: 1]
[21]
Moles A T Warton D I Warman L et al Global patterns in plant height
[J].Journal of Ecology ,2009 ,97 (5 ):923 -932 .
[本文引用: 1]
[22]
孟雅冰 ,李新蓉 多年生植物欧夏至草集合繁殖体特性
[J].西北植物学报 ,2015 ,35 (2 ):397 -402 .
[本文引用: 1]
[23]
Liu H M Wu Y L Li Y X et al Free-grazing versus enclosure lead to an increase in the germination of the Leymus chinensis seed bank in the Hulunbuir Grassland
[J].Agronomy ,2023 ,13 (1 ):22 .
[本文引用: 1]
[24]
王子龙 ,胡斌 ,包维楷 ,等 西南干旱河谷植物群落组分生物量的纬度格局及其影响因素
[J].植物生态学报 ,2022 ,46 (5 ):539 -551 .
[本文引用: 1]
[25]
Cambardella C A Moorman T B Novak J M Fieldscale variability of soil properties in central Iowa soils
[J].Soil Science Society of America Journal ,1994 ,58 (5 ):1501 -1511 .
[本文引用: 1]
[26]
贾利娟 ,桑雪颖 ,李慧娟 ,等 短花针茅荒漠草原物种数空间变化特点及其与取样方法的关系
[J].草原与草业 ,2014 ,26 (4 ):30 -34 .
[本文引用: 1]
[28]
吕世杰 ,刘红梅 ,吴艳玲 ,等 放牧对短花针茅荒漠草原建群种与优势种空间分布关系的影响
[J].应用生态学报 ,2014 ,25 (12 ):3469 -3474 .
[本文引用: 1]
[29]
Keitt T H Spectral representation of neutral landscapes
[J].Landscape Ecology ,2000 ,15 :479 -494 .
[本文引用: 1]
[30]
刘冰 ,赵文智 漠绿洲过渡带泡泡刺灌丛沙堆形态特征及其空间异质性
[J].应用生态学报 ,2007 ,18 (12 ):2814 -2820 .
[本文引用: 1]
[31]
吴艳玲 ,陈立波 ,卫智军 ,等 不同放牧压短花针茅荒漠草原群落植物种的空间异质特征
[J].干旱区资源与环境 ,2012 ,26 (7 ):110 -115 .
[本文引用: 1]
[32]
刘铮瑶 巴丹吉林沙漠边缘地区植物群落特征及资源利用分析
[D].西安 :陕西师范大学 ,2015 .
[本文引用: 1]
[33]
崔徐甲 ,董治宝 ,逯军峰 ,等 巴丹吉林沙漠高大沙山区植被特征与地貌形态的关系
[J].水土保持通报 ,2014 ,34 (5 ):278 -283 .
[本文引用: 1]
巴丹吉林沙漠周边地区土壤养分和粒度特征
2
2022
... 巴丹吉林沙漠东缘荒漠区气候干旱,植被稀疏,以旱生、超旱生的灌木和小灌木为主[1 -2 ] .这些灌木种群是巴丹吉林沙漠防风固沙的主要植物,对改善生态环境具有重要意义[1 ] .红砂(Reaumuria songarica )植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] .白刺(Nitraria tangutorum )能够构造自己的灌丛小生境,容易形成荒漠白刺土丘独有的自然景观,在干旱少雨时主要利用深层地下水,当降雨量超过年平均降水量时会及时利用地表水[6 -8 ] ;梭梭(Haloxylon ammodendron )对于土壤养分的需求低于红砂和白刺,所以可以在半流动沙丘上存活,成为沙漠流沙治理的先锋植物种群[2 ,9 -10 ] .3种主要灌木具有不同适应策略,其数量特征变化以及空间分布状态必然存在自己的独特性,因此掌握主要灌木种群数量特征和空间分布差异不仅有利于揭示其适应状态,更有利于利用这一适应基础为荒漠化防治提供理论依据和数据支撑. ...
... [1 ].红砂(Reaumuria songarica )植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] .白刺(Nitraria tangutorum )能够构造自己的灌丛小生境,容易形成荒漠白刺土丘独有的自然景观,在干旱少雨时主要利用深层地下水,当降雨量超过年平均降水量时会及时利用地表水[6 -8 ] ;梭梭(Haloxylon ammodendron )对于土壤养分的需求低于红砂和白刺,所以可以在半流动沙丘上存活,成为沙漠流沙治理的先锋植物种群[2 ,9 -10 ] .3种主要灌木具有不同适应策略,其数量特征变化以及空间分布状态必然存在自己的独特性,因此掌握主要灌木种群数量特征和空间分布差异不仅有利于揭示其适应状态,更有利于利用这一适应基础为荒漠化防治提供理论依据和数据支撑. ...
塔木素天然梭梭林空间分布及其与林地地貌起伏变化的关系
8
2015
... 巴丹吉林沙漠东缘荒漠区气候干旱,植被稀疏,以旱生、超旱生的灌木和小灌木为主[1 -2 ] .这些灌木种群是巴丹吉林沙漠防风固沙的主要植物,对改善生态环境具有重要意义[1 ] .红砂(Reaumuria songarica )植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] .白刺(Nitraria tangutorum )能够构造自己的灌丛小生境,容易形成荒漠白刺土丘独有的自然景观,在干旱少雨时主要利用深层地下水,当降雨量超过年平均降水量时会及时利用地表水[6 -8 ] ;梭梭(Haloxylon ammodendron )对于土壤养分的需求低于红砂和白刺,所以可以在半流动沙丘上存活,成为沙漠流沙治理的先锋植物种群[2 ,9 -10 ] .3种主要灌木具有不同适应策略,其数量特征变化以及空间分布状态必然存在自己的独特性,因此掌握主要灌木种群数量特征和空间分布差异不仅有利于揭示其适应状态,更有利于利用这一适应基础为荒漠化防治提供理论依据和数据支撑. ...
... [2 ,9 -10 ].3种主要灌木具有不同适应策略,其数量特征变化以及空间分布状态必然存在自己的独特性,因此掌握主要灌木种群数量特征和空间分布差异不仅有利于揭示其适应状态,更有利于利用这一适应基础为荒漠化防治提供理论依据和数据支撑. ...
... 刘红梅等[2 ,10 ] 研究巴丹吉林半流动沙地白刺、红砂和梭梭植物种群空间分布关系时发现,3个种群存在小尺度竞争、大尺度协同的空间镶嵌分布形式,且整体呈现以梭梭种群为主导的植被扩散格局和发展趋势.梭梭的空间分布与半流动沙丘微地形存在明显相关关系,且存在一定程度的沙埋适应[9 -12 ] .白刺植物种群空间分布在景观上表现出密度大而体型小的特点,灌丛沙堆处于发育的中早期阶段[13 ] ;红砂植物种群的密度、高度和冠幅空间异质性处于中等强度,且其密度空间分布与细沙空间分布紧密相关[14 ] .不同植物种群空间分布状态及其数量特征不仅与其自身适应性有关,也与生境砂层粒径存在相关性;然而这些研究均是基于流动、半流动沙地展开的研究,其在硬质沙地上相关研究鲜有报道. ...
... “压力优势假说”[16 -18 ] 认为,环境(非生物因素)过滤因子在更大环境梯度上对于种群构建植物群落具有重要意义,而竞争(生物因素)过滤因子在非生物压力较低的情况下更为重要.“唯利假说”[19 -20 ] 也预测了这种类似的现象,即当生存条件不太有利时,环境过滤能力更强.白刺、红砂和梭梭尽管生活型相同,均属于灌木,具备应对高风沙、高温、极端干旱严酷环境的能力,但是其适应基础存在较大的差异.对于白刺种群而言,其植株低矮,但是能够捕获流沙对自己进行掩埋形成沙丘,然后通过有性和无性繁殖扩大株丛,导致其株丛径大小本质上是沙丘的大小,尽管试验样地没有出现白刺为主的高大沙丘连续分布的地貌景观,但是其适应基础没有改变,因此其数量特征株丛径大小在3个种群占据优势[6 -8 ] .红砂植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] ;而研究区红砂种群株丛几乎没有形成沙丘,但是其出现频次在3个种群最高,印证了红砂对严酷环境具有高度的适应性,能够生长在砾石基质沙地上,由于其相互之间没有沙丘连接,均是具有主干形式的小灌木株丛,说明其有性繁殖的种子萌发能力和幼苗成功定植能力均比较强.梭梭植物种群对于土壤养分的需求低于红砂和白刺,所以能够在半流动沙地生存[2 ,9 -10 ] ,然而本研究区属于硬质沙地,梭梭植物种群不仅能够存活,而且具有相对较高的株高和冠幅直径(图2 ),说明在有限的土壤养分资源情况下,梭梭能够快速生长,体现在株高和冠幅直径上,同时又因本区域属于沙漠边缘区域,风大沙多,所以梭梭种群高大株型对风沙的耐受能力也要优于白刺和红砂.这也间接揭示了巴丹吉林沙漠东缘植被以梭梭主导的扩散格局和发展趋势[10 ] ,本研究重要值也显示梭梭植物种群在群落中的地位和作用最大,而白刺植物种群在植物群落中的地位和作用次之,红砂在植物群落中地位和作用在3个种群中最小;进而也证明“压力优势假说”和“唯利假说”导致不同灌木同样在干旱环境条件下,适应策略不同,进而表现在不同数量特征指标上. ...
... 结构比反映空间变量空间分布的主要影响因素.当结构比>75%,空间变量空间自相关性较强,主要受结构因素影响;结构比位于25%~75%,空间变量空间自相关居于中等强度,主要受结构因素和随机因素共同控制;当结构比<25%,空间变量空间自相关性最弱,主要受随机因素影响[25 ] .白刺和红砂植物种群均主要受结构因素影响,空间自相关尺度在不同样地的差别较大,特别是红砂植物种群差别更大,这可能与样地面积有关.贾利娟等[26 ] 研究荒漠草原时指出,样地样方法的样方数最好大于64个,样地10 m间隔取样最小面积应大于6 400 m2 .本研究是样地连续取样,且划分5个尺度,发现30 m尺度可能是该研究区最佳样地大小,既节约人力、财力和物力,也能有效地获得种群空间分布相对稳定的信息(图2 ),同时D样地设计对于种群监测更为有利[27 ] .白刺和梭梭的空间分布主要呈集中连片分布,这与刘红梅等[2 ,10 ] 前期的研究结果一致,但是整体上的空间分布在流动沙地呈大的斑块形式,而本研究尽管分析方法不同,二者仍然存在这一变化形式,但是红砂种群却存在较大的差异,主要是取样方法和研究区土壤基质差异所致.当对3个种群整体进行空间分布分析时,S样地随机性和结构性因子共同控制灌木空间分布,而D样地则显示结构性因素占据主导地位(表4 ),尽管二者决定因素不同,但是空间分布状态几乎一致,即呈现出斑块镶嵌分布,物种界限消失,这说明物种间采用协同方式来适应严酷环境[8 ,28 ] . ...
... 在研究区域,随机因素主导的空间分布状态主要指植物种群以母株为中心形成的集中连片分布形式,具有这种分布形式的种群为白刺和梭梭种群,其中梭梭的分布方向由西北向东南形成的条带分布符合该区域常年的合成风向,而梭梭种子传播在盛行风向具有较强的方向依赖性,所以这种基于合成风向的分布条带和其他风向形成的随机过程,导致梭梭种群形成当前的空间分布状态[2 ,29 -30 ] .结构性因素决定的空间分布状态主要是指由研究区气候、水文、土壤基质等决定的空间分布形式[31 ] ,D样地3个植物种群空间分布受结构性因素影响(表4 ),但是其空间自相关尺度仅为1.40,说明其空间分布的复杂性较低,空间异质性较弱.结合植被稀疏低矮、物种匮乏[2 ,9 ] ,可知其植物群落结构简单,进而在季相上形成灌木半灌木功能群构成的单一功能群灌丛群落,受“夏雨型”植物层片影响,虫实(Corispermum hyssopifolium )和雾冰藜(Grubovia dasyphylla )等大量出现,丰富了群落物种丰富度,但植物群落的简单结构变化不大,因此该群落的抗干扰能力和干扰后的恢复能力很弱,需要加强保护[2 ,32 -33 ] . ...
... [2 ,9 ],可知其植物群落结构简单,进而在季相上形成灌木半灌木功能群构成的单一功能群灌丛群落,受“夏雨型”植物层片影响,虫实(Corispermum hyssopifolium )和雾冰藜(Grubovia dasyphylla )等大量出现,丰富了群落物种丰富度,但植物群落的简单结构变化不大,因此该群落的抗干扰能力和干扰后的恢复能力很弱,需要加强保护[2 ,32 -33 ] . ...
... [2 ,32 -33 ]. ...
我国荒漠典型超旱生植物:红砂
2
1982
... 巴丹吉林沙漠东缘荒漠区气候干旱,植被稀疏,以旱生、超旱生的灌木和小灌木为主[1 -2 ] .这些灌木种群是巴丹吉林沙漠防风固沙的主要植物,对改善生态环境具有重要意义[1 ] .红砂(Reaumuria songarica )植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] .白刺(Nitraria tangutorum )能够构造自己的灌丛小生境,容易形成荒漠白刺土丘独有的自然景观,在干旱少雨时主要利用深层地下水,当降雨量超过年平均降水量时会及时利用地表水[6 -8 ] ;梭梭(Haloxylon ammodendron )对于土壤养分的需求低于红砂和白刺,所以可以在半流动沙丘上存活,成为沙漠流沙治理的先锋植物种群[2 ,9 -10 ] .3种主要灌木具有不同适应策略,其数量特征变化以及空间分布状态必然存在自己的独特性,因此掌握主要灌木种群数量特征和空间分布差异不仅有利于揭示其适应状态,更有利于利用这一适应基础为荒漠化防治提供理论依据和数据支撑. ...
... “压力优势假说”[16 -18 ] 认为,环境(非生物因素)过滤因子在更大环境梯度上对于种群构建植物群落具有重要意义,而竞争(生物因素)过滤因子在非生物压力较低的情况下更为重要.“唯利假说”[19 -20 ] 也预测了这种类似的现象,即当生存条件不太有利时,环境过滤能力更强.白刺、红砂和梭梭尽管生活型相同,均属于灌木,具备应对高风沙、高温、极端干旱严酷环境的能力,但是其适应基础存在较大的差异.对于白刺种群而言,其植株低矮,但是能够捕获流沙对自己进行掩埋形成沙丘,然后通过有性和无性繁殖扩大株丛,导致其株丛径大小本质上是沙丘的大小,尽管试验样地没有出现白刺为主的高大沙丘连续分布的地貌景观,但是其适应基础没有改变,因此其数量特征株丛径大小在3个种群占据优势[6 -8 ] .红砂植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] ;而研究区红砂种群株丛几乎没有形成沙丘,但是其出现频次在3个种群最高,印证了红砂对严酷环境具有高度的适应性,能够生长在砾石基质沙地上,由于其相互之间没有沙丘连接,均是具有主干形式的小灌木株丛,说明其有性繁殖的种子萌发能力和幼苗成功定植能力均比较强.梭梭植物种群对于土壤养分的需求低于红砂和白刺,所以能够在半流动沙地生存[2 ,9 -10 ] ,然而本研究区属于硬质沙地,梭梭植物种群不仅能够存活,而且具有相对较高的株高和冠幅直径(图2 ),说明在有限的土壤养分资源情况下,梭梭能够快速生长,体现在株高和冠幅直径上,同时又因本区域属于沙漠边缘区域,风大沙多,所以梭梭种群高大株型对风沙的耐受能力也要优于白刺和红砂.这也间接揭示了巴丹吉林沙漠东缘植被以梭梭主导的扩散格局和发展趋势[10 ] ,本研究重要值也显示梭梭植物种群在群落中的地位和作用最大,而白刺植物种群在植物群落中的地位和作用次之,红砂在植物群落中地位和作用在3个种群中最小;进而也证明“压力优势假说”和“唯利假说”导致不同灌木同样在干旱环境条件下,适应策略不同,进而表现在不同数量特征指标上. ...
红砂属植物研究进展
2
2004
... 巴丹吉林沙漠东缘荒漠区气候干旱,植被稀疏,以旱生、超旱生的灌木和小灌木为主[1 -2 ] .这些灌木种群是巴丹吉林沙漠防风固沙的主要植物,对改善生态环境具有重要意义[1 ] .红砂(Reaumuria songarica )植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] .白刺(Nitraria tangutorum )能够构造自己的灌丛小生境,容易形成荒漠白刺土丘独有的自然景观,在干旱少雨时主要利用深层地下水,当降雨量超过年平均降水量时会及时利用地表水[6 -8 ] ;梭梭(Haloxylon ammodendron )对于土壤养分的需求低于红砂和白刺,所以可以在半流动沙丘上存活,成为沙漠流沙治理的先锋植物种群[2 ,9 -10 ] .3种主要灌木具有不同适应策略,其数量特征变化以及空间分布状态必然存在自己的独特性,因此掌握主要灌木种群数量特征和空间分布差异不仅有利于揭示其适应状态,更有利于利用这一适应基础为荒漠化防治提供理论依据和数据支撑. ...
... “压力优势假说”[16 -18 ] 认为,环境(非生物因素)过滤因子在更大环境梯度上对于种群构建植物群落具有重要意义,而竞争(生物因素)过滤因子在非生物压力较低的情况下更为重要.“唯利假说”[19 -20 ] 也预测了这种类似的现象,即当生存条件不太有利时,环境过滤能力更强.白刺、红砂和梭梭尽管生活型相同,均属于灌木,具备应对高风沙、高温、极端干旱严酷环境的能力,但是其适应基础存在较大的差异.对于白刺种群而言,其植株低矮,但是能够捕获流沙对自己进行掩埋形成沙丘,然后通过有性和无性繁殖扩大株丛,导致其株丛径大小本质上是沙丘的大小,尽管试验样地没有出现白刺为主的高大沙丘连续分布的地貌景观,但是其适应基础没有改变,因此其数量特征株丛径大小在3个种群占据优势[6 -8 ] .红砂植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] ;而研究区红砂种群株丛几乎没有形成沙丘,但是其出现频次在3个种群最高,印证了红砂对严酷环境具有高度的适应性,能够生长在砾石基质沙地上,由于其相互之间没有沙丘连接,均是具有主干形式的小灌木株丛,说明其有性繁殖的种子萌发能力和幼苗成功定植能力均比较强.梭梭植物种群对于土壤养分的需求低于红砂和白刺,所以能够在半流动沙地生存[2 ,9 -10 ] ,然而本研究区属于硬质沙地,梭梭植物种群不仅能够存活,而且具有相对较高的株高和冠幅直径(图2 ),说明在有限的土壤养分资源情况下,梭梭能够快速生长,体现在株高和冠幅直径上,同时又因本区域属于沙漠边缘区域,风大沙多,所以梭梭种群高大株型对风沙的耐受能力也要优于白刺和红砂.这也间接揭示了巴丹吉林沙漠东缘植被以梭梭主导的扩散格局和发展趋势[10 ] ,本研究重要值也显示梭梭植物种群在群落中的地位和作用最大,而白刺植物种群在植物群落中的地位和作用次之,红砂在植物群落中地位和作用在3个种群中最小;进而也证明“压力优势假说”和“唯利假说”导致不同灌木同样在干旱环境条件下,适应策略不同,进而表现在不同数量特征指标上. ...
民勤绿洲白刺灌丛沙堆不同演化阶段表面抗蚀性及其影响因素
2
2008
... 巴丹吉林沙漠东缘荒漠区气候干旱,植被稀疏,以旱生、超旱生的灌木和小灌木为主[1 -2 ] .这些灌木种群是巴丹吉林沙漠防风固沙的主要植物,对改善生态环境具有重要意义[1 ] .红砂(Reaumuria songarica )植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] .白刺(Nitraria tangutorum )能够构造自己的灌丛小生境,容易形成荒漠白刺土丘独有的自然景观,在干旱少雨时主要利用深层地下水,当降雨量超过年平均降水量时会及时利用地表水[6 -8 ] ;梭梭(Haloxylon ammodendron )对于土壤养分的需求低于红砂和白刺,所以可以在半流动沙丘上存活,成为沙漠流沙治理的先锋植物种群[2 ,9 -10 ] .3种主要灌木具有不同适应策略,其数量特征变化以及空间分布状态必然存在自己的独特性,因此掌握主要灌木种群数量特征和空间分布差异不仅有利于揭示其适应状态,更有利于利用这一适应基础为荒漠化防治提供理论依据和数据支撑. ...
... “压力优势假说”[16 -18 ] 认为,环境(非生物因素)过滤因子在更大环境梯度上对于种群构建植物群落具有重要意义,而竞争(生物因素)过滤因子在非生物压力较低的情况下更为重要.“唯利假说”[19 -20 ] 也预测了这种类似的现象,即当生存条件不太有利时,环境过滤能力更强.白刺、红砂和梭梭尽管生活型相同,均属于灌木,具备应对高风沙、高温、极端干旱严酷环境的能力,但是其适应基础存在较大的差异.对于白刺种群而言,其植株低矮,但是能够捕获流沙对自己进行掩埋形成沙丘,然后通过有性和无性繁殖扩大株丛,导致其株丛径大小本质上是沙丘的大小,尽管试验样地没有出现白刺为主的高大沙丘连续分布的地貌景观,但是其适应基础没有改变,因此其数量特征株丛径大小在3个种群占据优势[6 -8 ] .红砂植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] ;而研究区红砂种群株丛几乎没有形成沙丘,但是其出现频次在3个种群最高,印证了红砂对严酷环境具有高度的适应性,能够生长在砾石基质沙地上,由于其相互之间没有沙丘连接,均是具有主干形式的小灌木株丛,说明其有性繁殖的种子萌发能力和幼苗成功定植能力均比较强.梭梭植物种群对于土壤养分的需求低于红砂和白刺,所以能够在半流动沙地生存[2 ,9 -10 ] ,然而本研究区属于硬质沙地,梭梭植物种群不仅能够存活,而且具有相对较高的株高和冠幅直径(图2 ),说明在有限的土壤养分资源情况下,梭梭能够快速生长,体现在株高和冠幅直径上,同时又因本区域属于沙漠边缘区域,风大沙多,所以梭梭种群高大株型对风沙的耐受能力也要优于白刺和红砂.这也间接揭示了巴丹吉林沙漠东缘植被以梭梭主导的扩散格局和发展趋势[10 ] ,本研究重要值也显示梭梭植物种群在群落中的地位和作用最大,而白刺植物种群在植物群落中的地位和作用次之,红砂在植物群落中地位和作用在3个种群中最小;进而也证明“压力优势假说”和“唯利假说”导致不同灌木同样在干旱环境条件下,适应策略不同,进而表现在不同数量特征指标上. ...
增雨对巴丹吉林沙漠东南缘白刺水分利用的影响
0
2013
白刺灌丛沙堆演化对地表蚀积的影响及其作用机制
3
2019
... 巴丹吉林沙漠东缘荒漠区气候干旱,植被稀疏,以旱生、超旱生的灌木和小灌木为主[1 -2 ] .这些灌木种群是巴丹吉林沙漠防风固沙的主要植物,对改善生态环境具有重要意义[1 ] .红砂(Reaumuria songarica )植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] .白刺(Nitraria tangutorum )能够构造自己的灌丛小生境,容易形成荒漠白刺土丘独有的自然景观,在干旱少雨时主要利用深层地下水,当降雨量超过年平均降水量时会及时利用地表水[6 -8 ] ;梭梭(Haloxylon ammodendron )对于土壤养分的需求低于红砂和白刺,所以可以在半流动沙丘上存活,成为沙漠流沙治理的先锋植物种群[2 ,9 -10 ] .3种主要灌木具有不同适应策略,其数量特征变化以及空间分布状态必然存在自己的独特性,因此掌握主要灌木种群数量特征和空间分布差异不仅有利于揭示其适应状态,更有利于利用这一适应基础为荒漠化防治提供理论依据和数据支撑. ...
... “压力优势假说”[16 -18 ] 认为,环境(非生物因素)过滤因子在更大环境梯度上对于种群构建植物群落具有重要意义,而竞争(生物因素)过滤因子在非生物压力较低的情况下更为重要.“唯利假说”[19 -20 ] 也预测了这种类似的现象,即当生存条件不太有利时,环境过滤能力更强.白刺、红砂和梭梭尽管生活型相同,均属于灌木,具备应对高风沙、高温、极端干旱严酷环境的能力,但是其适应基础存在较大的差异.对于白刺种群而言,其植株低矮,但是能够捕获流沙对自己进行掩埋形成沙丘,然后通过有性和无性繁殖扩大株丛,导致其株丛径大小本质上是沙丘的大小,尽管试验样地没有出现白刺为主的高大沙丘连续分布的地貌景观,但是其适应基础没有改变,因此其数量特征株丛径大小在3个种群占据优势[6 -8 ] .红砂植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] ;而研究区红砂种群株丛几乎没有形成沙丘,但是其出现频次在3个种群最高,印证了红砂对严酷环境具有高度的适应性,能够生长在砾石基质沙地上,由于其相互之间没有沙丘连接,均是具有主干形式的小灌木株丛,说明其有性繁殖的种子萌发能力和幼苗成功定植能力均比较强.梭梭植物种群对于土壤养分的需求低于红砂和白刺,所以能够在半流动沙地生存[2 ,9 -10 ] ,然而本研究区属于硬质沙地,梭梭植物种群不仅能够存活,而且具有相对较高的株高和冠幅直径(图2 ),说明在有限的土壤养分资源情况下,梭梭能够快速生长,体现在株高和冠幅直径上,同时又因本区域属于沙漠边缘区域,风大沙多,所以梭梭种群高大株型对风沙的耐受能力也要优于白刺和红砂.这也间接揭示了巴丹吉林沙漠东缘植被以梭梭主导的扩散格局和发展趋势[10 ] ,本研究重要值也显示梭梭植物种群在群落中的地位和作用最大,而白刺植物种群在植物群落中的地位和作用次之,红砂在植物群落中地位和作用在3个种群中最小;进而也证明“压力优势假说”和“唯利假说”导致不同灌木同样在干旱环境条件下,适应策略不同,进而表现在不同数量特征指标上. ...
... 结构比反映空间变量空间分布的主要影响因素.当结构比>75%,空间变量空间自相关性较强,主要受结构因素影响;结构比位于25%~75%,空间变量空间自相关居于中等强度,主要受结构因素和随机因素共同控制;当结构比<25%,空间变量空间自相关性最弱,主要受随机因素影响[25 ] .白刺和红砂植物种群均主要受结构因素影响,空间自相关尺度在不同样地的差别较大,特别是红砂植物种群差别更大,这可能与样地面积有关.贾利娟等[26 ] 研究荒漠草原时指出,样地样方法的样方数最好大于64个,样地10 m间隔取样最小面积应大于6 400 m2 .本研究是样地连续取样,且划分5个尺度,发现30 m尺度可能是该研究区最佳样地大小,既节约人力、财力和物力,也能有效地获得种群空间分布相对稳定的信息(图2 ),同时D样地设计对于种群监测更为有利[27 ] .白刺和梭梭的空间分布主要呈集中连片分布,这与刘红梅等[2 ,10 ] 前期的研究结果一致,但是整体上的空间分布在流动沙地呈大的斑块形式,而本研究尽管分析方法不同,二者仍然存在这一变化形式,但是红砂种群却存在较大的差异,主要是取样方法和研究区土壤基质差异所致.当对3个种群整体进行空间分布分析时,S样地随机性和结构性因子共同控制灌木空间分布,而D样地则显示结构性因素占据主导地位(表4 ),尽管二者决定因素不同,但是空间分布状态几乎一致,即呈现出斑块镶嵌分布,物种界限消失,这说明物种间采用协同方式来适应严酷环境[8 ,28 ] . ...
巴丹吉林沙漠东缘天然梭梭种群空间分布异质性
5
2016
... 巴丹吉林沙漠东缘荒漠区气候干旱,植被稀疏,以旱生、超旱生的灌木和小灌木为主[1 -2 ] .这些灌木种群是巴丹吉林沙漠防风固沙的主要植物,对改善生态环境具有重要意义[1 ] .红砂(Reaumuria songarica )植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] .白刺(Nitraria tangutorum )能够构造自己的灌丛小生境,容易形成荒漠白刺土丘独有的自然景观,在干旱少雨时主要利用深层地下水,当降雨量超过年平均降水量时会及时利用地表水[6 -8 ] ;梭梭(Haloxylon ammodendron )对于土壤养分的需求低于红砂和白刺,所以可以在半流动沙丘上存活,成为沙漠流沙治理的先锋植物种群[2 ,9 -10 ] .3种主要灌木具有不同适应策略,其数量特征变化以及空间分布状态必然存在自己的独特性,因此掌握主要灌木种群数量特征和空间分布差异不仅有利于揭示其适应状态,更有利于利用这一适应基础为荒漠化防治提供理论依据和数据支撑. ...
... 刘红梅等[2 ,10 ] 研究巴丹吉林半流动沙地白刺、红砂和梭梭植物种群空间分布关系时发现,3个种群存在小尺度竞争、大尺度协同的空间镶嵌分布形式,且整体呈现以梭梭种群为主导的植被扩散格局和发展趋势.梭梭的空间分布与半流动沙丘微地形存在明显相关关系,且存在一定程度的沙埋适应[9 -12 ] .白刺植物种群空间分布在景观上表现出密度大而体型小的特点,灌丛沙堆处于发育的中早期阶段[13 ] ;红砂植物种群的密度、高度和冠幅空间异质性处于中等强度,且其密度空间分布与细沙空间分布紧密相关[14 ] .不同植物种群空间分布状态及其数量特征不仅与其自身适应性有关,也与生境砂层粒径存在相关性;然而这些研究均是基于流动、半流动沙地展开的研究,其在硬质沙地上相关研究鲜有报道. ...
... 试验地行政区划属于阿拉善右旗塔木素苏木格勒图嘎查的朝恒扎干(塔木素野生肉苁蓉及梭梭产籽基地),地理位置为40°33.059′—40°35.531′N、103°25.162′—103°27.546′E,海拔1 255.8~1 264.3 m.研究区属典型温带干旱荒漠性气候,年降水量40~80 mm,集中于5—9月,年潜在蒸发量达到年降水量的40倍以上,干燥度7~12;年平均气温8~8.9 ℃,绝对最高气温37~43 ℃,平均无霜期150 d;风沙频繁,年均风速4 m·s-1 ;研究区流沙带以新月形沙丘和简单横向沙丘为主,沙丘主要走向为SW-NE.自然植被稀少,种属贫乏、覆盖率低,地表裸露[9 -10 ,14 ] . ...
... “压力优势假说”[16 -18 ] 认为,环境(非生物因素)过滤因子在更大环境梯度上对于种群构建植物群落具有重要意义,而竞争(生物因素)过滤因子在非生物压力较低的情况下更为重要.“唯利假说”[19 -20 ] 也预测了这种类似的现象,即当生存条件不太有利时,环境过滤能力更强.白刺、红砂和梭梭尽管生活型相同,均属于灌木,具备应对高风沙、高温、极端干旱严酷环境的能力,但是其适应基础存在较大的差异.对于白刺种群而言,其植株低矮,但是能够捕获流沙对自己进行掩埋形成沙丘,然后通过有性和无性繁殖扩大株丛,导致其株丛径大小本质上是沙丘的大小,尽管试验样地没有出现白刺为主的高大沙丘连续分布的地貌景观,但是其适应基础没有改变,因此其数量特征株丛径大小在3个种群占据优势[6 -8 ] .红砂植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] ;而研究区红砂种群株丛几乎没有形成沙丘,但是其出现频次在3个种群最高,印证了红砂对严酷环境具有高度的适应性,能够生长在砾石基质沙地上,由于其相互之间没有沙丘连接,均是具有主干形式的小灌木株丛,说明其有性繁殖的种子萌发能力和幼苗成功定植能力均比较强.梭梭植物种群对于土壤养分的需求低于红砂和白刺,所以能够在半流动沙地生存[2 ,9 -10 ] ,然而本研究区属于硬质沙地,梭梭植物种群不仅能够存活,而且具有相对较高的株高和冠幅直径(图2 ),说明在有限的土壤养分资源情况下,梭梭能够快速生长,体现在株高和冠幅直径上,同时又因本区域属于沙漠边缘区域,风大沙多,所以梭梭种群高大株型对风沙的耐受能力也要优于白刺和红砂.这也间接揭示了巴丹吉林沙漠东缘植被以梭梭主导的扩散格局和发展趋势[10 ] ,本研究重要值也显示梭梭植物种群在群落中的地位和作用最大,而白刺植物种群在植物群落中的地位和作用次之,红砂在植物群落中地位和作用在3个种群中最小;进而也证明“压力优势假说”和“唯利假说”导致不同灌木同样在干旱环境条件下,适应策略不同,进而表现在不同数量特征指标上. ...
... 在研究区域,随机因素主导的空间分布状态主要指植物种群以母株为中心形成的集中连片分布形式,具有这种分布形式的种群为白刺和梭梭种群,其中梭梭的分布方向由西北向东南形成的条带分布符合该区域常年的合成风向,而梭梭种子传播在盛行风向具有较强的方向依赖性,所以这种基于合成风向的分布条带和其他风向形成的随机过程,导致梭梭种群形成当前的空间分布状态[2 ,29 -30 ] .结构性因素决定的空间分布状态主要是指由研究区气候、水文、土壤基质等决定的空间分布形式[31 ] ,D样地3个植物种群空间分布受结构性因素影响(表4 ),但是其空间自相关尺度仅为1.40,说明其空间分布的复杂性较低,空间异质性较弱.结合植被稀疏低矮、物种匮乏[2 ,9 ] ,可知其植物群落结构简单,进而在季相上形成灌木半灌木功能群构成的单一功能群灌丛群落,受“夏雨型”植物层片影响,虫实(Corispermum hyssopifolium )和雾冰藜(Grubovia dasyphylla )等大量出现,丰富了群落物种丰富度,但植物群落的简单结构变化不大,因此该群落的抗干扰能力和干扰后的恢复能力很弱,需要加强保护[2 ,32 -33 ] . ...
巴丹吉林沙漠东缘主要植物种群空间分布关系
7
2021
... 巴丹吉林沙漠东缘荒漠区气候干旱,植被稀疏,以旱生、超旱生的灌木和小灌木为主[1 -2 ] .这些灌木种群是巴丹吉林沙漠防风固沙的主要植物,对改善生态环境具有重要意义[1 ] .红砂(Reaumuria songarica )植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] .白刺(Nitraria tangutorum )能够构造自己的灌丛小生境,容易形成荒漠白刺土丘独有的自然景观,在干旱少雨时主要利用深层地下水,当降雨量超过年平均降水量时会及时利用地表水[6 -8 ] ;梭梭(Haloxylon ammodendron )对于土壤养分的需求低于红砂和白刺,所以可以在半流动沙丘上存活,成为沙漠流沙治理的先锋植物种群[2 ,9 -10 ] .3种主要灌木具有不同适应策略,其数量特征变化以及空间分布状态必然存在自己的独特性,因此掌握主要灌木种群数量特征和空间分布差异不仅有利于揭示其适应状态,更有利于利用这一适应基础为荒漠化防治提供理论依据和数据支撑. ...
... 刘红梅等[2 ,10 ] 研究巴丹吉林半流动沙地白刺、红砂和梭梭植物种群空间分布关系时发现,3个种群存在小尺度竞争、大尺度协同的空间镶嵌分布形式,且整体呈现以梭梭种群为主导的植被扩散格局和发展趋势.梭梭的空间分布与半流动沙丘微地形存在明显相关关系,且存在一定程度的沙埋适应[9 -12 ] .白刺植物种群空间分布在景观上表现出密度大而体型小的特点,灌丛沙堆处于发育的中早期阶段[13 ] ;红砂植物种群的密度、高度和冠幅空间异质性处于中等强度,且其密度空间分布与细沙空间分布紧密相关[14 ] .不同植物种群空间分布状态及其数量特征不仅与其自身适应性有关,也与生境砂层粒径存在相关性;然而这些研究均是基于流动、半流动沙地展开的研究,其在硬质沙地上相关研究鲜有报道. ...
... 试验地行政区划属于阿拉善右旗塔木素苏木格勒图嘎查的朝恒扎干(塔木素野生肉苁蓉及梭梭产籽基地),地理位置为40°33.059′—40°35.531′N、103°25.162′—103°27.546′E,海拔1 255.8~1 264.3 m.研究区属典型温带干旱荒漠性气候,年降水量40~80 mm,集中于5—9月,年潜在蒸发量达到年降水量的40倍以上,干燥度7~12;年平均气温8~8.9 ℃,绝对最高气温37~43 ℃,平均无霜期150 d;风沙频繁,年均风速4 m·s-1 ;研究区流沙带以新月形沙丘和简单横向沙丘为主,沙丘主要走向为SW-NE.自然植被稀少,种属贫乏、覆盖率低,地表裸露[9 -10 ,14 ] . ...
... 分别对两个小样地白刺、红砂、梭梭种群及其整体进行半方差函数分析(数据采用个体重要值指数),分析过程在GS+9.0软件中进行,并在Microsoft Excel 2019中整理函数参数呈表格形式[15 ] ,然后再Surfur 12.0软件中绘制各种群及其整体空间分布状态[10 ] . ...
... “压力优势假说”[16 -18 ] 认为,环境(非生物因素)过滤因子在更大环境梯度上对于种群构建植物群落具有重要意义,而竞争(生物因素)过滤因子在非生物压力较低的情况下更为重要.“唯利假说”[19 -20 ] 也预测了这种类似的现象,即当生存条件不太有利时,环境过滤能力更强.白刺、红砂和梭梭尽管生活型相同,均属于灌木,具备应对高风沙、高温、极端干旱严酷环境的能力,但是其适应基础存在较大的差异.对于白刺种群而言,其植株低矮,但是能够捕获流沙对自己进行掩埋形成沙丘,然后通过有性和无性繁殖扩大株丛,导致其株丛径大小本质上是沙丘的大小,尽管试验样地没有出现白刺为主的高大沙丘连续分布的地貌景观,但是其适应基础没有改变,因此其数量特征株丛径大小在3个种群占据优势[6 -8 ] .红砂植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] ;而研究区红砂种群株丛几乎没有形成沙丘,但是其出现频次在3个种群最高,印证了红砂对严酷环境具有高度的适应性,能够生长在砾石基质沙地上,由于其相互之间没有沙丘连接,均是具有主干形式的小灌木株丛,说明其有性繁殖的种子萌发能力和幼苗成功定植能力均比较强.梭梭植物种群对于土壤养分的需求低于红砂和白刺,所以能够在半流动沙地生存[2 ,9 -10 ] ,然而本研究区属于硬质沙地,梭梭植物种群不仅能够存活,而且具有相对较高的株高和冠幅直径(图2 ),说明在有限的土壤养分资源情况下,梭梭能够快速生长,体现在株高和冠幅直径上,同时又因本区域属于沙漠边缘区域,风大沙多,所以梭梭种群高大株型对风沙的耐受能力也要优于白刺和红砂.这也间接揭示了巴丹吉林沙漠东缘植被以梭梭主导的扩散格局和发展趋势[10 ] ,本研究重要值也显示梭梭植物种群在群落中的地位和作用最大,而白刺植物种群在植物群落中的地位和作用次之,红砂在植物群落中地位和作用在3个种群中最小;进而也证明“压力优势假说”和“唯利假说”导致不同灌木同样在干旱环境条件下,适应策略不同,进而表现在不同数量特征指标上. ...
... [10 ],本研究重要值也显示梭梭植物种群在群落中的地位和作用最大,而白刺植物种群在植物群落中的地位和作用次之,红砂在植物群落中地位和作用在3个种群中最小;进而也证明“压力优势假说”和“唯利假说”导致不同灌木同样在干旱环境条件下,适应策略不同,进而表现在不同数量特征指标上. ...
... 结构比反映空间变量空间分布的主要影响因素.当结构比>75%,空间变量空间自相关性较强,主要受结构因素影响;结构比位于25%~75%,空间变量空间自相关居于中等强度,主要受结构因素和随机因素共同控制;当结构比<25%,空间变量空间自相关性最弱,主要受随机因素影响[25 ] .白刺和红砂植物种群均主要受结构因素影响,空间自相关尺度在不同样地的差别较大,特别是红砂植物种群差别更大,这可能与样地面积有关.贾利娟等[26 ] 研究荒漠草原时指出,样地样方法的样方数最好大于64个,样地10 m间隔取样最小面积应大于6 400 m2 .本研究是样地连续取样,且划分5个尺度,发现30 m尺度可能是该研究区最佳样地大小,既节约人力、财力和物力,也能有效地获得种群空间分布相对稳定的信息(图2 ),同时D样地设计对于种群监测更为有利[27 ] .白刺和梭梭的空间分布主要呈集中连片分布,这与刘红梅等[2 ,10 ] 前期的研究结果一致,但是整体上的空间分布在流动沙地呈大的斑块形式,而本研究尽管分析方法不同,二者仍然存在这一变化形式,但是红砂种群却存在较大的差异,主要是取样方法和研究区土壤基质差异所致.当对3个种群整体进行空间分布分析时,S样地随机性和结构性因子共同控制灌木空间分布,而D样地则显示结构性因素占据主导地位(表4 ),尽管二者决定因素不同,但是空间分布状态几乎一致,即呈现出斑块镶嵌分布,物种界限消失,这说明物种间采用协同方式来适应严酷环境[8 ,28 ] . ...
ffects of separate and mutual application of Na and Si on drought resistance of Haloxylon ammodendron
0
2012
Predicting the suitable habitats of parasitic desert species based on a niche model with Haloxylon ammodendron and Cistanche deserticola as examples
1
2021
... 刘红梅等[2 ,10 ] 研究巴丹吉林半流动沙地白刺、红砂和梭梭植物种群空间分布关系时发现,3个种群存在小尺度竞争、大尺度协同的空间镶嵌分布形式,且整体呈现以梭梭种群为主导的植被扩散格局和发展趋势.梭梭的空间分布与半流动沙丘微地形存在明显相关关系,且存在一定程度的沙埋适应[9 -12 ] .白刺植物种群空间分布在景观上表现出密度大而体型小的特点,灌丛沙堆处于发育的中早期阶段[13 ] ;红砂植物种群的密度、高度和冠幅空间异质性处于中等强度,且其密度空间分布与细沙空间分布紧密相关[14 ] .不同植物种群空间分布状态及其数量特征不仅与其自身适应性有关,也与生境砂层粒径存在相关性;然而这些研究均是基于流动、半流动沙地展开的研究,其在硬质沙地上相关研究鲜有报道. ...
巴丹吉林沙漠南缘近流沙区白刺灌丛沙堆形态特征与空间分布
1
2021
... 刘红梅等[2 ,10 ] 研究巴丹吉林半流动沙地白刺、红砂和梭梭植物种群空间分布关系时发现,3个种群存在小尺度竞争、大尺度协同的空间镶嵌分布形式,且整体呈现以梭梭种群为主导的植被扩散格局和发展趋势.梭梭的空间分布与半流动沙丘微地形存在明显相关关系,且存在一定程度的沙埋适应[9 -12 ] .白刺植物种群空间分布在景观上表现出密度大而体型小的特点,灌丛沙堆处于发育的中早期阶段[13 ] ;红砂植物种群的密度、高度和冠幅空间异质性处于中等强度,且其密度空间分布与细沙空间分布紧密相关[14 ] .不同植物种群空间分布状态及其数量特征不仅与其自身适应性有关,也与生境砂层粒径存在相关性;然而这些研究均是基于流动、半流动沙地展开的研究,其在硬质沙地上相关研究鲜有报道. ...
红砂植物种群空间分布与沙子粒级及地貌起伏之间的关系
2
2020
... 刘红梅等[2 ,10 ] 研究巴丹吉林半流动沙地白刺、红砂和梭梭植物种群空间分布关系时发现,3个种群存在小尺度竞争、大尺度协同的空间镶嵌分布形式,且整体呈现以梭梭种群为主导的植被扩散格局和发展趋势.梭梭的空间分布与半流动沙丘微地形存在明显相关关系,且存在一定程度的沙埋适应[9 -12 ] .白刺植物种群空间分布在景观上表现出密度大而体型小的特点,灌丛沙堆处于发育的中早期阶段[13 ] ;红砂植物种群的密度、高度和冠幅空间异质性处于中等强度,且其密度空间分布与细沙空间分布紧密相关[14 ] .不同植物种群空间分布状态及其数量特征不仅与其自身适应性有关,也与生境砂层粒径存在相关性;然而这些研究均是基于流动、半流动沙地展开的研究,其在硬质沙地上相关研究鲜有报道. ...
... 试验地行政区划属于阿拉善右旗塔木素苏木格勒图嘎查的朝恒扎干(塔木素野生肉苁蓉及梭梭产籽基地),地理位置为40°33.059′—40°35.531′N、103°25.162′—103°27.546′E,海拔1 255.8~1 264.3 m.研究区属典型温带干旱荒漠性气候,年降水量40~80 mm,集中于5—9月,年潜在蒸发量达到年降水量的40倍以上,干燥度7~12;年平均气温8~8.9 ℃,绝对最高气温37~43 ℃,平均无霜期150 d;风沙频繁,年均风速4 m·s-1 ;研究区流沙带以新月形沙丘和简单横向沙丘为主,沙丘主要走向为SW-NE.自然植被稀少,种属贫乏、覆盖率低,地表裸露[9 -10 ,14 ] . ...
Grazing intensity enhances spatial aggregation of dominant species in a desert steppe
1
2019
... 分别对两个小样地白刺、红砂、梭梭种群及其整体进行半方差函数分析(数据采用个体重要值指数),分析过程在GS+9.0软件中进行,并在Microsoft Excel 2019中整理函数参数呈表格形式[15 ] ,然后再Surfur 12.0软件中绘制各种群及其整体空间分布状态[10 ] . ...
Ecological and evolutionary determinants of a key plant functional trait: Wood density and its community-wide variation across latitude and elevation
1
2007
... “压力优势假说”[16 -18 ] 认为,环境(非生物因素)过滤因子在更大环境梯度上对于种群构建植物群落具有重要意义,而竞争(生物因素)过滤因子在非生物压力较低的情况下更为重要.“唯利假说”[19 -20 ] 也预测了这种类似的现象,即当生存条件不太有利时,环境过滤能力更强.白刺、红砂和梭梭尽管生活型相同,均属于灌木,具备应对高风沙、高温、极端干旱严酷环境的能力,但是其适应基础存在较大的差异.对于白刺种群而言,其植株低矮,但是能够捕获流沙对自己进行掩埋形成沙丘,然后通过有性和无性繁殖扩大株丛,导致其株丛径大小本质上是沙丘的大小,尽管试验样地没有出现白刺为主的高大沙丘连续分布的地貌景观,但是其适应基础没有改变,因此其数量特征株丛径大小在3个种群占据优势[6 -8 ] .红砂植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] ;而研究区红砂种群株丛几乎没有形成沙丘,但是其出现频次在3个种群最高,印证了红砂对严酷环境具有高度的适应性,能够生长在砾石基质沙地上,由于其相互之间没有沙丘连接,均是具有主干形式的小灌木株丛,说明其有性繁殖的种子萌发能力和幼苗成功定植能力均比较强.梭梭植物种群对于土壤养分的需求低于红砂和白刺,所以能够在半流动沙地生存[2 ,9 -10 ] ,然而本研究区属于硬质沙地,梭梭植物种群不仅能够存活,而且具有相对较高的株高和冠幅直径(图2 ),说明在有限的土壤养分资源情况下,梭梭能够快速生长,体现在株高和冠幅直径上,同时又因本区域属于沙漠边缘区域,风大沙多,所以梭梭种群高大株型对风沙的耐受能力也要优于白刺和红砂.这也间接揭示了巴丹吉林沙漠东缘植被以梭梭主导的扩散格局和发展趋势[10 ] ,本研究重要值也显示梭梭植物种群在群落中的地位和作用最大,而白刺植物种群在植物群落中的地位和作用次之,红砂在植物群落中地位和作用在3个种群中最小;进而也证明“压力优势假说”和“唯利假说”导致不同灌木同样在干旱环境条件下,适应策略不同,进而表现在不同数量特征指标上. ...
Using trait and phylogenetic diversity to evaluate the generality of the stress-dominance hypothesis in Eastern North American tree communities
0
2014
Directional selection shifts trait distributions of planted species in dryland restoration
1
2022
... “压力优势假说”[16 -18 ] 认为,环境(非生物因素)过滤因子在更大环境梯度上对于种群构建植物群落具有重要意义,而竞争(生物因素)过滤因子在非生物压力较低的情况下更为重要.“唯利假说”[19 -20 ] 也预测了这种类似的现象,即当生存条件不太有利时,环境过滤能力更强.白刺、红砂和梭梭尽管生活型相同,均属于灌木,具备应对高风沙、高温、极端干旱严酷环境的能力,但是其适应基础存在较大的差异.对于白刺种群而言,其植株低矮,但是能够捕获流沙对自己进行掩埋形成沙丘,然后通过有性和无性繁殖扩大株丛,导致其株丛径大小本质上是沙丘的大小,尽管试验样地没有出现白刺为主的高大沙丘连续分布的地貌景观,但是其适应基础没有改变,因此其数量特征株丛径大小在3个种群占据优势[6 -8 ] .红砂植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] ;而研究区红砂种群株丛几乎没有形成沙丘,但是其出现频次在3个种群最高,印证了红砂对严酷环境具有高度的适应性,能够生长在砾石基质沙地上,由于其相互之间没有沙丘连接,均是具有主干形式的小灌木株丛,说明其有性繁殖的种子萌发能力和幼苗成功定植能力均比较强.梭梭植物种群对于土壤养分的需求低于红砂和白刺,所以能够在半流动沙地生存[2 ,9 -10 ] ,然而本研究区属于硬质沙地,梭梭植物种群不仅能够存活,而且具有相对较高的株高和冠幅直径(图2 ),说明在有限的土壤养分资源情况下,梭梭能够快速生长,体现在株高和冠幅直径上,同时又因本区域属于沙漠边缘区域,风大沙多,所以梭梭种群高大株型对风沙的耐受能力也要优于白刺和红砂.这也间接揭示了巴丹吉林沙漠东缘植被以梭梭主导的扩散格局和发展趋势[10 ] ,本研究重要值也显示梭梭植物种群在群落中的地位和作用最大,而白刺植物种群在植物群落中的地位和作用次之,红砂在植物群落中地位和作用在3个种群中最小;进而也证明“压力优势假说”和“唯利假说”导致不同灌木同样在干旱环境条件下,适应策略不同,进而表现在不同数量特征指标上. ...
The biogeography and filtering of woody plant functional diversityin North and South America
1
2012
... “压力优势假说”[16 -18 ] 认为,环境(非生物因素)过滤因子在更大环境梯度上对于种群构建植物群落具有重要意义,而竞争(生物因素)过滤因子在非生物压力较低的情况下更为重要.“唯利假说”[19 -20 ] 也预测了这种类似的现象,即当生存条件不太有利时,环境过滤能力更强.白刺、红砂和梭梭尽管生活型相同,均属于灌木,具备应对高风沙、高温、极端干旱严酷环境的能力,但是其适应基础存在较大的差异.对于白刺种群而言,其植株低矮,但是能够捕获流沙对自己进行掩埋形成沙丘,然后通过有性和无性繁殖扩大株丛,导致其株丛径大小本质上是沙丘的大小,尽管试验样地没有出现白刺为主的高大沙丘连续分布的地貌景观,但是其适应基础没有改变,因此其数量特征株丛径大小在3个种群占据优势[6 -8 ] .红砂植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] ;而研究区红砂种群株丛几乎没有形成沙丘,但是其出现频次在3个种群最高,印证了红砂对严酷环境具有高度的适应性,能够生长在砾石基质沙地上,由于其相互之间没有沙丘连接,均是具有主干形式的小灌木株丛,说明其有性繁殖的种子萌发能力和幼苗成功定植能力均比较强.梭梭植物种群对于土壤养分的需求低于红砂和白刺,所以能够在半流动沙地生存[2 ,9 -10 ] ,然而本研究区属于硬质沙地,梭梭植物种群不仅能够存活,而且具有相对较高的株高和冠幅直径(图2 ),说明在有限的土壤养分资源情况下,梭梭能够快速生长,体现在株高和冠幅直径上,同时又因本区域属于沙漠边缘区域,风大沙多,所以梭梭种群高大株型对风沙的耐受能力也要优于白刺和红砂.这也间接揭示了巴丹吉林沙漠东缘植被以梭梭主导的扩散格局和发展趋势[10 ] ,本研究重要值也显示梭梭植物种群在群落中的地位和作用最大,而白刺植物种群在植物群落中的地位和作用次之,红砂在植物群落中地位和作用在3个种群中最小;进而也证明“压力优势假说”和“唯利假说”导致不同灌木同样在干旱环境条件下,适应策略不同,进而表现在不同数量特征指标上. ...
Correction: climate shapes and shifts functional biodiversity in forests worldwide
1
2019
... “压力优势假说”[16 -18 ] 认为,环境(非生物因素)过滤因子在更大环境梯度上对于种群构建植物群落具有重要意义,而竞争(生物因素)过滤因子在非生物压力较低的情况下更为重要.“唯利假说”[19 -20 ] 也预测了这种类似的现象,即当生存条件不太有利时,环境过滤能力更强.白刺、红砂和梭梭尽管生活型相同,均属于灌木,具备应对高风沙、高温、极端干旱严酷环境的能力,但是其适应基础存在较大的差异.对于白刺种群而言,其植株低矮,但是能够捕获流沙对自己进行掩埋形成沙丘,然后通过有性和无性繁殖扩大株丛,导致其株丛径大小本质上是沙丘的大小,尽管试验样地没有出现白刺为主的高大沙丘连续分布的地貌景观,但是其适应基础没有改变,因此其数量特征株丛径大小在3个种群占据优势[6 -8 ] .红砂植物种群是超旱生小灌木,对严酷环境具有高度的适应性,可以生长在砾石基质沙地上,由于独特的无性繁殖方式形成或不形成土丘[3 -5 ] ;而研究区红砂种群株丛几乎没有形成沙丘,但是其出现频次在3个种群最高,印证了红砂对严酷环境具有高度的适应性,能够生长在砾石基质沙地上,由于其相互之间没有沙丘连接,均是具有主干形式的小灌木株丛,说明其有性繁殖的种子萌发能力和幼苗成功定植能力均比较强.梭梭植物种群对于土壤养分的需求低于红砂和白刺,所以能够在半流动沙地生存[2 ,9 -10 ] ,然而本研究区属于硬质沙地,梭梭植物种群不仅能够存活,而且具有相对较高的株高和冠幅直径(图2 ),说明在有限的土壤养分资源情况下,梭梭能够快速生长,体现在株高和冠幅直径上,同时又因本区域属于沙漠边缘区域,风大沙多,所以梭梭种群高大株型对风沙的耐受能力也要优于白刺和红砂.这也间接揭示了巴丹吉林沙漠东缘植被以梭梭主导的扩散格局和发展趋势[10 ] ,本研究重要值也显示梭梭植物种群在群落中的地位和作用最大,而白刺植物种群在植物群落中的地位和作用次之,红砂在植物群落中地位和作用在3个种群中最小;进而也证明“压力优势假说”和“唯利假说”导致不同灌木同样在干旱环境条件下,适应策略不同,进而表现在不同数量特征指标上. ...
Global patterns in plant height
1
2009
... 一般认为,植物的高度和冠幅直径反映了植物种群对光的捕获能力[21 ] ,而该区域植被稀疏,群落结构简单,且光照周期长,所以主要植物种群株高和冠幅直径并不能反映其对光的敏感性和适应性,因此植物群落物种构成以及地理区域不同,可能会导致植物种群同一指标反映的适应基础不同.密度或者频次,往往反映了植物种群繁衍的能力,进而对于多年生物种判断有性繁殖和无性繁殖的适应能力[22 ] .高强度干扰可导致有性繁殖能力下降,但也会导致有性繁殖质量增加[23 ] .本研究红砂种群具有较强的有性繁殖能力,但是究竟是有性繁殖质量增加所致,还是有性繁殖能力不变或者适应严酷环境的结果有待于进一步研究,进而白刺以母株为中心捕获流沙,是否获得无性繁殖体扩展导致其具有较大的株丛径也需要进一步明确.种群生物量反映了植物种群在一定空间内,通过繁殖体获取资源的能力[24 ] ,尽管本研究没有相关研究,但是从重要值角度进行了初步分析,因重要值反映了其在植物群落的地位和作用,也反映了其在植物群落的功能强弱,所以初步判定梭梭、白刺和红砂在群落中的整体作用依次降低. ...
多年生植物欧夏至草集合繁殖体特性
1
2015
... 一般认为,植物的高度和冠幅直径反映了植物种群对光的捕获能力[21 ] ,而该区域植被稀疏,群落结构简单,且光照周期长,所以主要植物种群株高和冠幅直径并不能反映其对光的敏感性和适应性,因此植物群落物种构成以及地理区域不同,可能会导致植物种群同一指标反映的适应基础不同.密度或者频次,往往反映了植物种群繁衍的能力,进而对于多年生物种判断有性繁殖和无性繁殖的适应能力[22 ] .高强度干扰可导致有性繁殖能力下降,但也会导致有性繁殖质量增加[23 ] .本研究红砂种群具有较强的有性繁殖能力,但是究竟是有性繁殖质量增加所致,还是有性繁殖能力不变或者适应严酷环境的结果有待于进一步研究,进而白刺以母株为中心捕获流沙,是否获得无性繁殖体扩展导致其具有较大的株丛径也需要进一步明确.种群生物量反映了植物种群在一定空间内,通过繁殖体获取资源的能力[24 ] ,尽管本研究没有相关研究,但是从重要值角度进行了初步分析,因重要值反映了其在植物群落的地位和作用,也反映了其在植物群落的功能强弱,所以初步判定梭梭、白刺和红砂在群落中的整体作用依次降低. ...
Free-grazing versus enclosure lead to an increase in the germination of the Leymus chinensis seed bank in the Hulunbuir Grassland
1
2023
... 一般认为,植物的高度和冠幅直径反映了植物种群对光的捕获能力[21 ] ,而该区域植被稀疏,群落结构简单,且光照周期长,所以主要植物种群株高和冠幅直径并不能反映其对光的敏感性和适应性,因此植物群落物种构成以及地理区域不同,可能会导致植物种群同一指标反映的适应基础不同.密度或者频次,往往反映了植物种群繁衍的能力,进而对于多年生物种判断有性繁殖和无性繁殖的适应能力[22 ] .高强度干扰可导致有性繁殖能力下降,但也会导致有性繁殖质量增加[23 ] .本研究红砂种群具有较强的有性繁殖能力,但是究竟是有性繁殖质量增加所致,还是有性繁殖能力不变或者适应严酷环境的结果有待于进一步研究,进而白刺以母株为中心捕获流沙,是否获得无性繁殖体扩展导致其具有较大的株丛径也需要进一步明确.种群生物量反映了植物种群在一定空间内,通过繁殖体获取资源的能力[24 ] ,尽管本研究没有相关研究,但是从重要值角度进行了初步分析,因重要值反映了其在植物群落的地位和作用,也反映了其在植物群落的功能强弱,所以初步判定梭梭、白刺和红砂在群落中的整体作用依次降低. ...
西南干旱河谷植物群落组分生物量的纬度格局及其影响因素
1
2022
... 一般认为,植物的高度和冠幅直径反映了植物种群对光的捕获能力[21 ] ,而该区域植被稀疏,群落结构简单,且光照周期长,所以主要植物种群株高和冠幅直径并不能反映其对光的敏感性和适应性,因此植物群落物种构成以及地理区域不同,可能会导致植物种群同一指标反映的适应基础不同.密度或者频次,往往反映了植物种群繁衍的能力,进而对于多年生物种判断有性繁殖和无性繁殖的适应能力[22 ] .高强度干扰可导致有性繁殖能力下降,但也会导致有性繁殖质量增加[23 ] .本研究红砂种群具有较强的有性繁殖能力,但是究竟是有性繁殖质量增加所致,还是有性繁殖能力不变或者适应严酷环境的结果有待于进一步研究,进而白刺以母株为中心捕获流沙,是否获得无性繁殖体扩展导致其具有较大的株丛径也需要进一步明确.种群生物量反映了植物种群在一定空间内,通过繁殖体获取资源的能力[24 ] ,尽管本研究没有相关研究,但是从重要值角度进行了初步分析,因重要值反映了其在植物群落的地位和作用,也反映了其在植物群落的功能强弱,所以初步判定梭梭、白刺和红砂在群落中的整体作用依次降低. ...
Fieldscale variability of soil properties in central Iowa soils
1
1994
... 结构比反映空间变量空间分布的主要影响因素.当结构比>75%,空间变量空间自相关性较强,主要受结构因素影响;结构比位于25%~75%,空间变量空间自相关居于中等强度,主要受结构因素和随机因素共同控制;当结构比<25%,空间变量空间自相关性最弱,主要受随机因素影响[25 ] .白刺和红砂植物种群均主要受结构因素影响,空间自相关尺度在不同样地的差别较大,特别是红砂植物种群差别更大,这可能与样地面积有关.贾利娟等[26 ] 研究荒漠草原时指出,样地样方法的样方数最好大于64个,样地10 m间隔取样最小面积应大于6 400 m2 .本研究是样地连续取样,且划分5个尺度,发现30 m尺度可能是该研究区最佳样地大小,既节约人力、财力和物力,也能有效地获得种群空间分布相对稳定的信息(图2 ),同时D样地设计对于种群监测更为有利[27 ] .白刺和梭梭的空间分布主要呈集中连片分布,这与刘红梅等[2 ,10 ] 前期的研究结果一致,但是整体上的空间分布在流动沙地呈大的斑块形式,而本研究尽管分析方法不同,二者仍然存在这一变化形式,但是红砂种群却存在较大的差异,主要是取样方法和研究区土壤基质差异所致.当对3个种群整体进行空间分布分析时,S样地随机性和结构性因子共同控制灌木空间分布,而D样地则显示结构性因素占据主导地位(表4 ),尽管二者决定因素不同,但是空间分布状态几乎一致,即呈现出斑块镶嵌分布,物种界限消失,这说明物种间采用协同方式来适应严酷环境[8 ,28 ] . ...
短花针茅荒漠草原物种数空间变化特点及其与取样方法的关系
1
2014
... 结构比反映空间变量空间分布的主要影响因素.当结构比>75%,空间变量空间自相关性较强,主要受结构因素影响;结构比位于25%~75%,空间变量空间自相关居于中等强度,主要受结构因素和随机因素共同控制;当结构比<25%,空间变量空间自相关性最弱,主要受随机因素影响[25 ] .白刺和红砂植物种群均主要受结构因素影响,空间自相关尺度在不同样地的差别较大,特别是红砂植物种群差别更大,这可能与样地面积有关.贾利娟等[26 ] 研究荒漠草原时指出,样地样方法的样方数最好大于64个,样地10 m间隔取样最小面积应大于6 400 m2 .本研究是样地连续取样,且划分5个尺度,发现30 m尺度可能是该研究区最佳样地大小,既节约人力、财力和物力,也能有效地获得种群空间分布相对稳定的信息(图2 ),同时D样地设计对于种群监测更为有利[27 ] .白刺和梭梭的空间分布主要呈集中连片分布,这与刘红梅等[2 ,10 ] 前期的研究结果一致,但是整体上的空间分布在流动沙地呈大的斑块形式,而本研究尽管分析方法不同,二者仍然存在这一变化形式,但是红砂种群却存在较大的差异,主要是取样方法和研究区土壤基质差异所致.当对3个种群整体进行空间分布分析时,S样地随机性和结构性因子共同控制灌木空间分布,而D样地则显示结构性因素占据主导地位(表4 ),尽管二者决定因素不同,但是空间分布状态几乎一致,即呈现出斑块镶嵌分布,物种界限消失,这说明物种间采用协同方式来适应严酷环境[8 ,28 ] . ...
1
2022
... 结构比反映空间变量空间分布的主要影响因素.当结构比>75%,空间变量空间自相关性较强,主要受结构因素影响;结构比位于25%~75%,空间变量空间自相关居于中等强度,主要受结构因素和随机因素共同控制;当结构比<25%,空间变量空间自相关性最弱,主要受随机因素影响[25 ] .白刺和红砂植物种群均主要受结构因素影响,空间自相关尺度在不同样地的差别较大,特别是红砂植物种群差别更大,这可能与样地面积有关.贾利娟等[26 ] 研究荒漠草原时指出,样地样方法的样方数最好大于64个,样地10 m间隔取样最小面积应大于6 400 m2 .本研究是样地连续取样,且划分5个尺度,发现30 m尺度可能是该研究区最佳样地大小,既节约人力、财力和物力,也能有效地获得种群空间分布相对稳定的信息(图2 ),同时D样地设计对于种群监测更为有利[27 ] .白刺和梭梭的空间分布主要呈集中连片分布,这与刘红梅等[2 ,10 ] 前期的研究结果一致,但是整体上的空间分布在流动沙地呈大的斑块形式,而本研究尽管分析方法不同,二者仍然存在这一变化形式,但是红砂种群却存在较大的差异,主要是取样方法和研究区土壤基质差异所致.当对3个种群整体进行空间分布分析时,S样地随机性和结构性因子共同控制灌木空间分布,而D样地则显示结构性因素占据主导地位(表4 ),尽管二者决定因素不同,但是空间分布状态几乎一致,即呈现出斑块镶嵌分布,物种界限消失,这说明物种间采用协同方式来适应严酷环境[8 ,28 ] . ...
放牧对短花针茅荒漠草原建群种与优势种空间分布关系的影响
1
2014
... 结构比反映空间变量空间分布的主要影响因素.当结构比>75%,空间变量空间自相关性较强,主要受结构因素影响;结构比位于25%~75%,空间变量空间自相关居于中等强度,主要受结构因素和随机因素共同控制;当结构比<25%,空间变量空间自相关性最弱,主要受随机因素影响[25 ] .白刺和红砂植物种群均主要受结构因素影响,空间自相关尺度在不同样地的差别较大,特别是红砂植物种群差别更大,这可能与样地面积有关.贾利娟等[26 ] 研究荒漠草原时指出,样地样方法的样方数最好大于64个,样地10 m间隔取样最小面积应大于6 400 m2 .本研究是样地连续取样,且划分5个尺度,发现30 m尺度可能是该研究区最佳样地大小,既节约人力、财力和物力,也能有效地获得种群空间分布相对稳定的信息(图2 ),同时D样地设计对于种群监测更为有利[27 ] .白刺和梭梭的空间分布主要呈集中连片分布,这与刘红梅等[2 ,10 ] 前期的研究结果一致,但是整体上的空间分布在流动沙地呈大的斑块形式,而本研究尽管分析方法不同,二者仍然存在这一变化形式,但是红砂种群却存在较大的差异,主要是取样方法和研究区土壤基质差异所致.当对3个种群整体进行空间分布分析时,S样地随机性和结构性因子共同控制灌木空间分布,而D样地则显示结构性因素占据主导地位(表4 ),尽管二者决定因素不同,但是空间分布状态几乎一致,即呈现出斑块镶嵌分布,物种界限消失,这说明物种间采用协同方式来适应严酷环境[8 ,28 ] . ...
Spectral representation of neutral landscapes
1
2000
... 在研究区域,随机因素主导的空间分布状态主要指植物种群以母株为中心形成的集中连片分布形式,具有这种分布形式的种群为白刺和梭梭种群,其中梭梭的分布方向由西北向东南形成的条带分布符合该区域常年的合成风向,而梭梭种子传播在盛行风向具有较强的方向依赖性,所以这种基于合成风向的分布条带和其他风向形成的随机过程,导致梭梭种群形成当前的空间分布状态[2 ,29 -30 ] .结构性因素决定的空间分布状态主要是指由研究区气候、水文、土壤基质等决定的空间分布形式[31 ] ,D样地3个植物种群空间分布受结构性因素影响(表4 ),但是其空间自相关尺度仅为1.40,说明其空间分布的复杂性较低,空间异质性较弱.结合植被稀疏低矮、物种匮乏[2 ,9 ] ,可知其植物群落结构简单,进而在季相上形成灌木半灌木功能群构成的单一功能群灌丛群落,受“夏雨型”植物层片影响,虫实(Corispermum hyssopifolium )和雾冰藜(Grubovia dasyphylla )等大量出现,丰富了群落物种丰富度,但植物群落的简单结构变化不大,因此该群落的抗干扰能力和干扰后的恢复能力很弱,需要加强保护[2 ,32 -33 ] . ...
漠绿洲过渡带泡泡刺灌丛沙堆形态特征及其空间异质性
1
2007
... 在研究区域,随机因素主导的空间分布状态主要指植物种群以母株为中心形成的集中连片分布形式,具有这种分布形式的种群为白刺和梭梭种群,其中梭梭的分布方向由西北向东南形成的条带分布符合该区域常年的合成风向,而梭梭种子传播在盛行风向具有较强的方向依赖性,所以这种基于合成风向的分布条带和其他风向形成的随机过程,导致梭梭种群形成当前的空间分布状态[2 ,29 -30 ] .结构性因素决定的空间分布状态主要是指由研究区气候、水文、土壤基质等决定的空间分布形式[31 ] ,D样地3个植物种群空间分布受结构性因素影响(表4 ),但是其空间自相关尺度仅为1.40,说明其空间分布的复杂性较低,空间异质性较弱.结合植被稀疏低矮、物种匮乏[2 ,9 ] ,可知其植物群落结构简单,进而在季相上形成灌木半灌木功能群构成的单一功能群灌丛群落,受“夏雨型”植物层片影响,虫实(Corispermum hyssopifolium )和雾冰藜(Grubovia dasyphylla )等大量出现,丰富了群落物种丰富度,但植物群落的简单结构变化不大,因此该群落的抗干扰能力和干扰后的恢复能力很弱,需要加强保护[2 ,32 -33 ] . ...
不同放牧压短花针茅荒漠草原群落植物种的空间异质特征
1
2012
... 在研究区域,随机因素主导的空间分布状态主要指植物种群以母株为中心形成的集中连片分布形式,具有这种分布形式的种群为白刺和梭梭种群,其中梭梭的分布方向由西北向东南形成的条带分布符合该区域常年的合成风向,而梭梭种子传播在盛行风向具有较强的方向依赖性,所以这种基于合成风向的分布条带和其他风向形成的随机过程,导致梭梭种群形成当前的空间分布状态[2 ,29 -30 ] .结构性因素决定的空间分布状态主要是指由研究区气候、水文、土壤基质等决定的空间分布形式[31 ] ,D样地3个植物种群空间分布受结构性因素影响(表4 ),但是其空间自相关尺度仅为1.40,说明其空间分布的复杂性较低,空间异质性较弱.结合植被稀疏低矮、物种匮乏[2 ,9 ] ,可知其植物群落结构简单,进而在季相上形成灌木半灌木功能群构成的单一功能群灌丛群落,受“夏雨型”植物层片影响,虫实(Corispermum hyssopifolium )和雾冰藜(Grubovia dasyphylla )等大量出现,丰富了群落物种丰富度,但植物群落的简单结构变化不大,因此该群落的抗干扰能力和干扰后的恢复能力很弱,需要加强保护[2 ,32 -33 ] . ...
巴丹吉林沙漠边缘地区植物群落特征及资源利用分析
1
2015
... 在研究区域,随机因素主导的空间分布状态主要指植物种群以母株为中心形成的集中连片分布形式,具有这种分布形式的种群为白刺和梭梭种群,其中梭梭的分布方向由西北向东南形成的条带分布符合该区域常年的合成风向,而梭梭种子传播在盛行风向具有较强的方向依赖性,所以这种基于合成风向的分布条带和其他风向形成的随机过程,导致梭梭种群形成当前的空间分布状态[2 ,29 -30 ] .结构性因素决定的空间分布状态主要是指由研究区气候、水文、土壤基质等决定的空间分布形式[31 ] ,D样地3个植物种群空间分布受结构性因素影响(表4 ),但是其空间自相关尺度仅为1.40,说明其空间分布的复杂性较低,空间异质性较弱.结合植被稀疏低矮、物种匮乏[2 ,9 ] ,可知其植物群落结构简单,进而在季相上形成灌木半灌木功能群构成的单一功能群灌丛群落,受“夏雨型”植物层片影响,虫实(Corispermum hyssopifolium )和雾冰藜(Grubovia dasyphylla )等大量出现,丰富了群落物种丰富度,但植物群落的简单结构变化不大,因此该群落的抗干扰能力和干扰后的恢复能力很弱,需要加强保护[2 ,32 -33 ] . ...
巴丹吉林沙漠高大沙山区植被特征与地貌形态的关系
1
2014
... 在研究区域,随机因素主导的空间分布状态主要指植物种群以母株为中心形成的集中连片分布形式,具有这种分布形式的种群为白刺和梭梭种群,其中梭梭的分布方向由西北向东南形成的条带分布符合该区域常年的合成风向,而梭梭种子传播在盛行风向具有较强的方向依赖性,所以这种基于合成风向的分布条带和其他风向形成的随机过程,导致梭梭种群形成当前的空间分布状态[2 ,29 -30 ] .结构性因素决定的空间分布状态主要是指由研究区气候、水文、土壤基质等决定的空间分布形式[31 ] ,D样地3个植物种群空间分布受结构性因素影响(表4 ),但是其空间自相关尺度仅为1.40,说明其空间分布的复杂性较低,空间异质性较弱.结合植被稀疏低矮、物种匮乏[2 ,9 ] ,可知其植物群落结构简单,进而在季相上形成灌木半灌木功能群构成的单一功能群灌丛群落,受“夏雨型”植物层片影响,虫实(Corispermum hyssopifolium )和雾冰藜(Grubovia dasyphylla )等大量出现,丰富了群落物种丰富度,但植物群落的简单结构变化不大,因此该群落的抗干扰能力和干扰后的恢复能力很弱,需要加强保护[2 ,32 -33 ] . ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}