Cloning and expression analysis of sugar transporter gene SpSWEET3 from the desert plant Stipagrostis pennata
Sun Bohan,1, Yang Dan1, Wang Fei1,2,3, Li Rong,1,2,3, Li Hongbin,1,2,3
1.College of Life Science /, Shihezi University,Shihezi 832003,Xinjiang,China
2.Xinjiang Production and Construction Corps Key Laboratory of Oasis Town and Mountain-basin System Ecology /, Shihezi University,Shihezi 832003,Xinjiang,China
3.Key Laboratory of Xinjiang Phytomedicine Resource and Utilization of Ministry of Education, Shihezi University,Shihezi 832003,Xinjiang,China
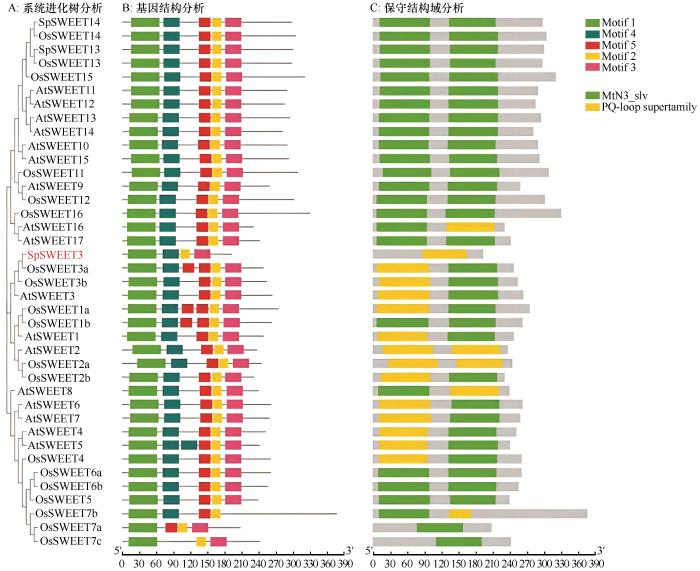
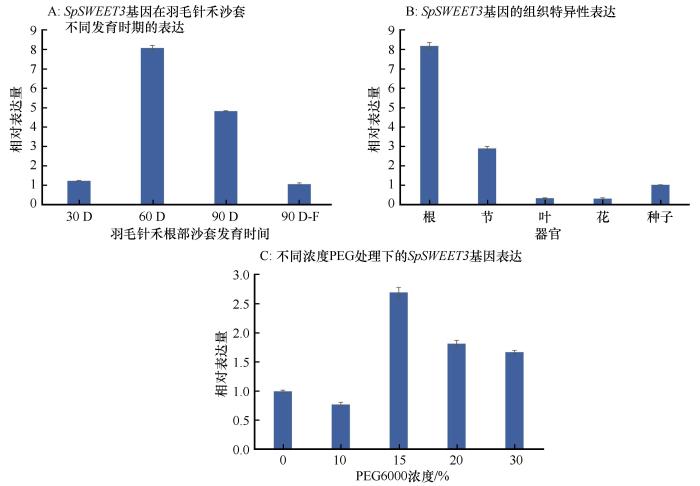
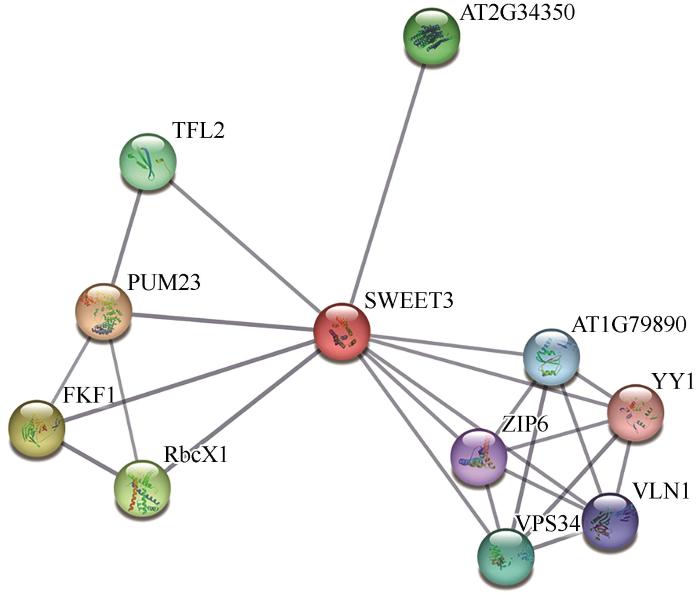
Sugar transporter (SWEET) plays an important role in plant growth and development and stress response. The purpose of this study is to clone and analyze the expression characteristics of Stipagrosis pennata sugar transporter SpSWEET3 gene, and lay a foundation for further exploring the mechanism and function of its influence on the development of S. pennata root sand trap. The full-length open reading frame sequence of S. pennata SpSWEET3 gene was cloned by molecular cloning technology; The physical and chemical properties of SpSWEET3 gene were analyzed by bioinformatics; The gene expression characteristics were analyzed by qRT-PCR; The subcellular localization was observed and analyzed by laser confocal microscope. SpSWEET3was cloned from the desert plant S. pennata, which includes a 576-bp full-length open reading frame and encodes a basic strongly hydrophobic protein containing 191 amino acids with a relative molecular mass of 21.619 kD. SpSWEET3 belongs to the PQ-loop superfamily and contains the typical MtN3_slv transmembrane domain that catalyzes intracellular sugar efflux. Subcellular localization analysis revealed localization of the gene to the plasma membrane, implying that it may function as a transmembrane protein. qRT-PCR results showed that the SpSWEET3 gene had a higher cumulative expression in S. pennata root tissues, indicating its important role for sand rhizosheath development. Expression of the gene SpSWEET3 was significantly induced by drought stress with PEG, indicating its importance for drought adaptation. Protein interaction analysis showed that SpSWEET3 may transport sugar by interacting with proteins involved in the maintenance of nucleic acid structural stability, transcription factors, and transmembrane transport. The results of this study provide a foundation for in-depth investigation of the functions of SWEET genes and the mechanisms by which they regulate plant tissue development and adaptation to stress.
Sun Bohan, Yang Dan, Wang Fei, Li Rong, Li Hongbin. Cloning and expression analysis of sugar transporter gene SpSWEET3 from the desert plant Stipagrostis pennata. Journal of Desert Research[J], 2023, 43(5): 129-138 doi:10.7522/j.issn.1000-694X.2023.00037
可溶性糖在植物生长发育和逆境胁迫响应调控中发挥着重要作用。在逆境胁迫下,植物为了维持细胞渗透势的平衡,通过对体内可溶性糖进行再分配,从而利于植物在逆境胁迫下维持正常生长[3]。植物沙套的形成需要沙粒和根表分泌物的共同作用[4]。沙套代谢组学研究显示,在干旱胁迫处理后,根部大多数糖类物质和部分氨基酸在沙套根中高度积累[5];尤其是糖类物质在干旱处理的根中表现出显著的累积,表明其与干旱条件下的沙套生成密切相关[6]。糖转运蛋白是调控可溶性糖再分配的关键因子,能够响应多种逆境胁迫,与植物逆境适应密切相关。SWEET(Sugars will Eventually be Exported Transporter)是一种新型糖转运蛋白,广泛存在于动植物中[7]。SWEET糖转运蛋白具有7个跨膜结构域,SWEET蛋白的结构同源性比较高,在结构上高度保守[8]。在植物体内,SWEET糖转运蛋白的存在很好地解释了糖分转运模式,研究表明 SWEET转运蛋白可以促进糖类顺浓度梯度扩散以及调控植物体内糖类化合物的运输[9-11]。也有研究表明,SWEET基因家族成员在植物应对干旱胁迫过程中发挥重要作用。在拟南芥中,在干旱胁迫条件下,AtSWEET11、AtSWEET12和AtSWEET15在叶片中的表达量上调,提高蔗糖转运效率,促进糖向根系转移,从而降低根系渗透压,提高根系吸水量[12]。
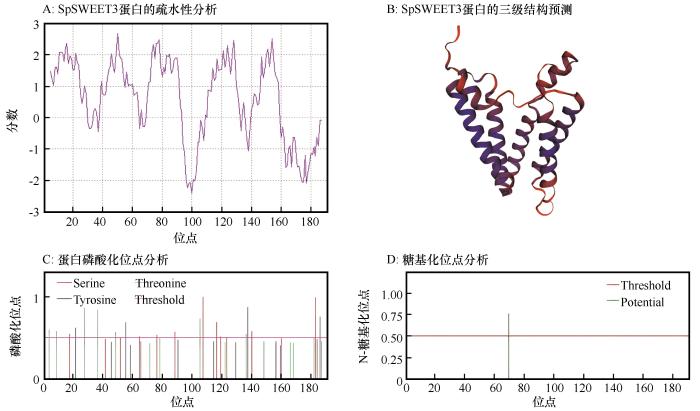
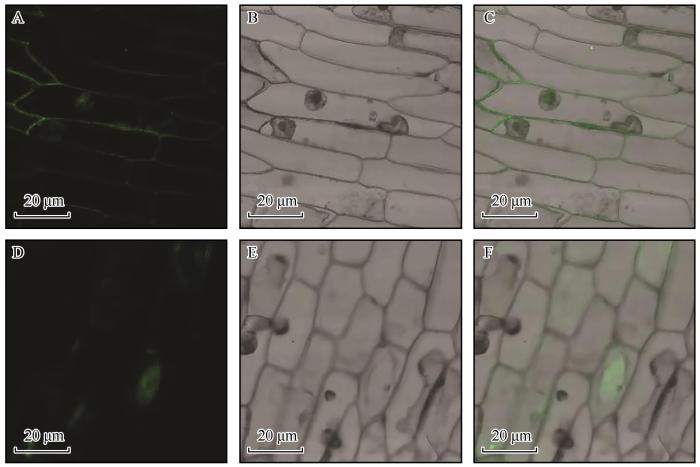
利用ExPSAy在线服务器中的PortParam在线软件对SpSWEET3进行疏水性分析(图4)。SpSWEET3蛋白的亲水性总平均值(Grand average of hydropathicity,GRAVY)为0.635 (GRAVY在-2与2之间,大于0为疏水性,小于0为亲水性),脂溶指数 (Aliphatic index,AI)为110.73。分析结果显示SpSWEET3蛋白具有强疏水性和高脂溶性;亚细胞定位预测结果显示其定位于质膜。SpSWEET3蛋白包含11个α-螺旋,其余皆为无规卷曲,主要由这两种二级结构组成。对SpSWEET3蛋白的磷酸化和糖基化位点分析结果显示:SpSWEET3含有41个磷酸化位点,包含15个丝氨酸位点、15个苏氨酸位点和11个酪氨酸位点。SpSWEET3含有1个潜在的糖基化位点。
Functional analysis of an Arabidopsis thaliana abiotic stress-inducible facilitated diffusion transporter for monosaccharides
1
2010
... 可溶性糖在植物生长发育和逆境胁迫响应调控中发挥着重要作用.在逆境胁迫下,植物为了维持细胞渗透势的平衡,通过对体内可溶性糖进行再分配,从而利于植物在逆境胁迫下维持正常生长[3].植物沙套的形成需要沙粒和根表分泌物的共同作用[4].沙套代谢组学研究显示,在干旱胁迫处理后,根部大多数糖类物质和部分氨基酸在沙套根中高度积累[5];尤其是糖类物质在干旱处理的根中表现出显著的累积,表明其与干旱条件下的沙套生成密切相关[6].糖转运蛋白是调控可溶性糖再分配的关键因子,能够响应多种逆境胁迫,与植物逆境适应密切相关.SWEET(Sugars will Eventually be Exported Transporter)是一种新型糖转运蛋白,广泛存在于动植物中[7].SWEET糖转运蛋白具有7个跨膜结构域,SWEET蛋白的结构同源性比较高,在结构上高度保守[8].在植物体内,SWEET糖转运蛋白的存在很好地解释了糖分转运模式,研究表明 SWEET转运蛋白可以促进糖类顺浓度梯度扩散以及调控植物体内糖类化合物的运输[9-11].也有研究表明,SWEET基因家族成员在植物应对干旱胁迫过程中发挥重要作用.在拟南芥中,在干旱胁迫条件下,AtSWEET11、AtSWEET12和AtSWEET15在叶片中的表达量上调,提高蔗糖转运效率,促进糖向根系转移,从而降低根系渗透压,提高根系吸水量[12]. ...
Soil sterilization effects on root growth and formation of rhizosheaths in wheat seedlings
1
2014
... 可溶性糖在植物生长发育和逆境胁迫响应调控中发挥着重要作用.在逆境胁迫下,植物为了维持细胞渗透势的平衡,通过对体内可溶性糖进行再分配,从而利于植物在逆境胁迫下维持正常生长[3].植物沙套的形成需要沙粒和根表分泌物的共同作用[4].沙套代谢组学研究显示,在干旱胁迫处理后,根部大多数糖类物质和部分氨基酸在沙套根中高度积累[5];尤其是糖类物质在干旱处理的根中表现出显著的累积,表明其与干旱条件下的沙套生成密切相关[6].糖转运蛋白是调控可溶性糖再分配的关键因子,能够响应多种逆境胁迫,与植物逆境适应密切相关.SWEET(Sugars will Eventually be Exported Transporter)是一种新型糖转运蛋白,广泛存在于动植物中[7].SWEET糖转运蛋白具有7个跨膜结构域,SWEET蛋白的结构同源性比较高,在结构上高度保守[8].在植物体内,SWEET糖转运蛋白的存在很好地解释了糖分转运模式,研究表明 SWEET转运蛋白可以促进糖类顺浓度梯度扩散以及调控植物体内糖类化合物的运输[9-11].也有研究表明,SWEET基因家族成员在植物应对干旱胁迫过程中发挥重要作用.在拟南芥中,在干旱胁迫条件下,AtSWEET11、AtSWEET12和AtSWEET15在叶片中的表达量上调,提高蔗糖转运效率,促进糖向根系转移,从而降低根系渗透压,提高根系吸水量[12]. ...
Uptake of Zn65from dry soil by plants
1
1976
... 可溶性糖在植物生长发育和逆境胁迫响应调控中发挥着重要作用.在逆境胁迫下,植物为了维持细胞渗透势的平衡,通过对体内可溶性糖进行再分配,从而利于植物在逆境胁迫下维持正常生长[3].植物沙套的形成需要沙粒和根表分泌物的共同作用[4].沙套代谢组学研究显示,在干旱胁迫处理后,根部大多数糖类物质和部分氨基酸在沙套根中高度积累[5];尤其是糖类物质在干旱处理的根中表现出显著的累积,表明其与干旱条件下的沙套生成密切相关[6].糖转运蛋白是调控可溶性糖再分配的关键因子,能够响应多种逆境胁迫,与植物逆境适应密切相关.SWEET(Sugars will Eventually be Exported Transporter)是一种新型糖转运蛋白,广泛存在于动植物中[7].SWEET糖转运蛋白具有7个跨膜结构域,SWEET蛋白的结构同源性比较高,在结构上高度保守[8].在植物体内,SWEET糖转运蛋白的存在很好地解释了糖分转运模式,研究表明 SWEET转运蛋白可以促进糖类顺浓度梯度扩散以及调控植物体内糖类化合物的运输[9-11].也有研究表明,SWEET基因家族成员在植物应对干旱胁迫过程中发挥重要作用.在拟南芥中,在干旱胁迫条件下,AtSWEET11、AtSWEET12和AtSWEET15在叶片中的表达量上调,提高蔗糖转运效率,促进糖向根系转移,从而降低根系渗透压,提高根系吸水量[12]. ...
Comparative metabolite profiling of two switchgrass ecotypes reveals differences in drought stress responses and rhizosheath weight
1
2019
... 可溶性糖在植物生长发育和逆境胁迫响应调控中发挥着重要作用.在逆境胁迫下,植物为了维持细胞渗透势的平衡,通过对体内可溶性糖进行再分配,从而利于植物在逆境胁迫下维持正常生长[3].植物沙套的形成需要沙粒和根表分泌物的共同作用[4].沙套代谢组学研究显示,在干旱胁迫处理后,根部大多数糖类物质和部分氨基酸在沙套根中高度积累[5];尤其是糖类物质在干旱处理的根中表现出显著的累积,表明其与干旱条件下的沙套生成密切相关[6].糖转运蛋白是调控可溶性糖再分配的关键因子,能够响应多种逆境胁迫,与植物逆境适应密切相关.SWEET(Sugars will Eventually be Exported Transporter)是一种新型糖转运蛋白,广泛存在于动植物中[7].SWEET糖转运蛋白具有7个跨膜结构域,SWEET蛋白的结构同源性比较高,在结构上高度保守[8].在植物体内,SWEET糖转运蛋白的存在很好地解释了糖分转运模式,研究表明 SWEET转运蛋白可以促进糖类顺浓度梯度扩散以及调控植物体内糖类化合物的运输[9-11].也有研究表明,SWEET基因家族成员在植物应对干旱胁迫过程中发挥重要作用.在拟南芥中,在干旱胁迫条件下,AtSWEET11、AtSWEET12和AtSWEET15在叶片中的表达量上调,提高蔗糖转运效率,促进糖向根系转移,从而降低根系渗透压,提高根系吸水量[12]. ...
植物SWEET基因家族的相关研究进展
1
2017
... 可溶性糖在植物生长发育和逆境胁迫响应调控中发挥着重要作用.在逆境胁迫下,植物为了维持细胞渗透势的平衡,通过对体内可溶性糖进行再分配,从而利于植物在逆境胁迫下维持正常生长[3].植物沙套的形成需要沙粒和根表分泌物的共同作用[4].沙套代谢组学研究显示,在干旱胁迫处理后,根部大多数糖类物质和部分氨基酸在沙套根中高度积累[5];尤其是糖类物质在干旱处理的根中表现出显著的累积,表明其与干旱条件下的沙套生成密切相关[6].糖转运蛋白是调控可溶性糖再分配的关键因子,能够响应多种逆境胁迫,与植物逆境适应密切相关.SWEET(Sugars will Eventually be Exported Transporter)是一种新型糖转运蛋白,广泛存在于动植物中[7].SWEET糖转运蛋白具有7个跨膜结构域,SWEET蛋白的结构同源性比较高,在结构上高度保守[8].在植物体内,SWEET糖转运蛋白的存在很好地解释了糖分转运模式,研究表明 SWEET转运蛋白可以促进糖类顺浓度梯度扩散以及调控植物体内糖类化合物的运输[9-11].也有研究表明,SWEET基因家族成员在植物应对干旱胁迫过程中发挥重要作用.在拟南芥中,在干旱胁迫条件下,AtSWEET11、AtSWEET12和AtSWEET15在叶片中的表达量上调,提高蔗糖转运效率,促进糖向根系转移,从而降低根系渗透压,提高根系吸水量[12]. ...
Sugar transporters for intercellular exchange and nutrition of pathogens
1
2010
... 可溶性糖在植物生长发育和逆境胁迫响应调控中发挥着重要作用.在逆境胁迫下,植物为了维持细胞渗透势的平衡,通过对体内可溶性糖进行再分配,从而利于植物在逆境胁迫下维持正常生长[3].植物沙套的形成需要沙粒和根表分泌物的共同作用[4].沙套代谢组学研究显示,在干旱胁迫处理后,根部大多数糖类物质和部分氨基酸在沙套根中高度积累[5];尤其是糖类物质在干旱处理的根中表现出显著的累积,表明其与干旱条件下的沙套生成密切相关[6].糖转运蛋白是调控可溶性糖再分配的关键因子,能够响应多种逆境胁迫,与植物逆境适应密切相关.SWEET(Sugars will Eventually be Exported Transporter)是一种新型糖转运蛋白,广泛存在于动植物中[7].SWEET糖转运蛋白具有7个跨膜结构域,SWEET蛋白的结构同源性比较高,在结构上高度保守[8].在植物体内,SWEET糖转运蛋白的存在很好地解释了糖分转运模式,研究表明 SWEET转运蛋白可以促进糖类顺浓度梯度扩散以及调控植物体内糖类化合物的运输[9-11].也有研究表明,SWEET基因家族成员在植物应对干旱胁迫过程中发挥重要作用.在拟南芥中,在干旱胁迫条件下,AtSWEET11、AtSWEET12和AtSWEET15在叶片中的表达量上调,提高蔗糖转运效率,促进糖向根系转移,从而降低根系渗透压,提高根系吸水量[12]. ...
SWEET as sugar: new sucrose effluxers in plants
1
2012
... 可溶性糖在植物生长发育和逆境胁迫响应调控中发挥着重要作用.在逆境胁迫下,植物为了维持细胞渗透势的平衡,通过对体内可溶性糖进行再分配,从而利于植物在逆境胁迫下维持正常生长[3].植物沙套的形成需要沙粒和根表分泌物的共同作用[4].沙套代谢组学研究显示,在干旱胁迫处理后,根部大多数糖类物质和部分氨基酸在沙套根中高度积累[5];尤其是糖类物质在干旱处理的根中表现出显著的累积,表明其与干旱条件下的沙套生成密切相关[6].糖转运蛋白是调控可溶性糖再分配的关键因子,能够响应多种逆境胁迫,与植物逆境适应密切相关.SWEET(Sugars will Eventually be Exported Transporter)是一种新型糖转运蛋白,广泛存在于动植物中[7].SWEET糖转运蛋白具有7个跨膜结构域,SWEET蛋白的结构同源性比较高,在结构上高度保守[8].在植物体内,SWEET糖转运蛋白的存在很好地解释了糖分转运模式,研究表明 SWEET转运蛋白可以促进糖类顺浓度梯度扩散以及调控植物体内糖类化合物的运输[9-11].也有研究表明,SWEET基因家族成员在植物应对干旱胁迫过程中发挥重要作用.在拟南芥中,在干旱胁迫条件下,AtSWEET11、AtSWEET12和AtSWEET15在叶片中的表达量上调,提高蔗糖转运效率,促进糖向根系转移,从而降低根系渗透压,提高根系吸水量[12]. ...
SWEET sugar transporters for phloem transport and pathogen nutrition
0
2014
Co-option of developmentally regulated plant SWEET transporters for pathogen nutrition and abiotic stress tolerance
1
2015
... 可溶性糖在植物生长发育和逆境胁迫响应调控中发挥着重要作用.在逆境胁迫下,植物为了维持细胞渗透势的平衡,通过对体内可溶性糖进行再分配,从而利于植物在逆境胁迫下维持正常生长[3].植物沙套的形成需要沙粒和根表分泌物的共同作用[4].沙套代谢组学研究显示,在干旱胁迫处理后,根部大多数糖类物质和部分氨基酸在沙套根中高度积累[5];尤其是糖类物质在干旱处理的根中表现出显著的累积,表明其与干旱条件下的沙套生成密切相关[6].糖转运蛋白是调控可溶性糖再分配的关键因子,能够响应多种逆境胁迫,与植物逆境适应密切相关.SWEET(Sugars will Eventually be Exported Transporter)是一种新型糖转运蛋白,广泛存在于动植物中[7].SWEET糖转运蛋白具有7个跨膜结构域,SWEET蛋白的结构同源性比较高,在结构上高度保守[8].在植物体内,SWEET糖转运蛋白的存在很好地解释了糖分转运模式,研究表明 SWEET转运蛋白可以促进糖类顺浓度梯度扩散以及调控植物体内糖类化合物的运输[9-11].也有研究表明,SWEET基因家族成员在植物应对干旱胁迫过程中发挥重要作用.在拟南芥中,在干旱胁迫条件下,AtSWEET11、AtSWEET12和AtSWEET15在叶片中的表达量上调,提高蔗糖转运效率,促进糖向根系转移,从而降低根系渗透压,提高根系吸水量[12]. ...
Water deficit enhances c export to the roots in Arabidopsis thaliana plants with contribution of sucrose transporters in both shoot and roots
1
2016
... 可溶性糖在植物生长发育和逆境胁迫响应调控中发挥着重要作用.在逆境胁迫下,植物为了维持细胞渗透势的平衡,通过对体内可溶性糖进行再分配,从而利于植物在逆境胁迫下维持正常生长[3].植物沙套的形成需要沙粒和根表分泌物的共同作用[4].沙套代谢组学研究显示,在干旱胁迫处理后,根部大多数糖类物质和部分氨基酸在沙套根中高度积累[5];尤其是糖类物质在干旱处理的根中表现出显著的累积,表明其与干旱条件下的沙套生成密切相关[6].糖转运蛋白是调控可溶性糖再分配的关键因子,能够响应多种逆境胁迫,与植物逆境适应密切相关.SWEET(Sugars will Eventually be Exported Transporter)是一种新型糖转运蛋白,广泛存在于动植物中[7].SWEET糖转运蛋白具有7个跨膜结构域,SWEET蛋白的结构同源性比较高,在结构上高度保守[8].在植物体内,SWEET糖转运蛋白的存在很好地解释了糖分转运模式,研究表明 SWEET转运蛋白可以促进糖类顺浓度梯度扩散以及调控植物体内糖类化合物的运输[9-11].也有研究表明,SWEET基因家族成员在植物应对干旱胁迫过程中发挥重要作用.在拟南芥中,在干旱胁迫条件下,AtSWEET11、AtSWEET12和AtSWEET15在叶片中的表达量上调,提高蔗糖转运效率,促进糖向根系转移,从而降低根系渗透压,提高根系吸水量[12]. ...
Selection of the reference genes for quantitative gene expression by RT-qPCR in the desert plant Stipagrostis pennata







 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}