Note sur les algues qui colourent en rouge certaines eaux des marais salants méditerranéens
1
1837
... 有关粉色盐湖的最早记录出现于《本草纲目》,其中记载了制盐所用卤水久置后会变红的现象.而针对此类湖泊的科学研究则开始于19世纪,其中杜氏藻最初被认为是造成盐湖变粉的原因,在地中海沿岸红色盐池中被分离出,由Teodoresco将其命名并归类为杜氏藻属(Dunaliella)[1-2].杜氏藻(D.salina)可在体内积累较多的β-胡萝卜素,最高可达细胞干重的16%,因此细胞呈现粉红色[3].研究者也分别从光照、盐度以及水中氮磷含量等角度研究了影响杜氏藻积累类胡萝卜素的原因[4-5].此后有学者认为水体中的嗜盐细菌与嗜盐古菌是造成水体变粉的主要原因.对以色列红色盐水的研究表明,红色盐水中以嗜盐古菌及杜氏藻所含的菌红素以及类胡萝卜素为主[6-8].同时证明虽然杜氏藻所含β-胡萝卜素占比较大,但因其在叶绿素内密集堆叠,光学面积小,因此嗜盐古菌的存在才是导致盐水变红的主要因素.而1999年通过荧光原位杂交技术(FISH)在西班牙盐池发现一种名为Salinibacter的红色细菌在盐池生态系统中占主导地位,证明嗜盐古菌不是唯一造成盐湖变粉的菌类,并在2002年实现菌种分离[9-11].而对Salinibacter培养物的实验室研究证明,杜氏藻光合作用的产物甘油易被Salinibacter吸收并刺激其生长[12].因此造成高盐水体变粉的微生物往往不止一种,可能是杜氏藻、嗜盐古菌与嗜盐细菌的共同作用. ...
Organisation et développement du Dunaliella,nouveau genre de Volvocacée-Polyblépharidée
1
1905
... 有关粉色盐湖的最早记录出现于《本草纲目》,其中记载了制盐所用卤水久置后会变红的现象.而针对此类湖泊的科学研究则开始于19世纪,其中杜氏藻最初被认为是造成盐湖变粉的原因,在地中海沿岸红色盐池中被分离出,由Teodoresco将其命名并归类为杜氏藻属(Dunaliella)[1-2].杜氏藻(D.salina)可在体内积累较多的β-胡萝卜素,最高可达细胞干重的16%,因此细胞呈现粉红色[3].研究者也分别从光照、盐度以及水中氮磷含量等角度研究了影响杜氏藻积累类胡萝卜素的原因[4-5].此后有学者认为水体中的嗜盐细菌与嗜盐古菌是造成水体变粉的主要原因.对以色列红色盐水的研究表明,红色盐水中以嗜盐古菌及杜氏藻所含的菌红素以及类胡萝卜素为主[6-8].同时证明虽然杜氏藻所含β-胡萝卜素占比较大,但因其在叶绿素内密集堆叠,光学面积小,因此嗜盐古菌的存在才是导致盐水变红的主要因素.而1999年通过荧光原位杂交技术(FISH)在西班牙盐池发现一种名为Salinibacter的红色细菌在盐池生态系统中占主导地位,证明嗜盐古菌不是唯一造成盐湖变粉的菌类,并在2002年实现菌种分离[9-11].而对Salinibacter培养物的实验室研究证明,杜氏藻光合作用的产物甘油易被Salinibacter吸收并刺激其生长[12].因此造成高盐水体变粉的微生物往往不止一种,可能是杜氏藻、嗜盐古菌与嗜盐细菌的共同作用. ...
1
2019
... 有关粉色盐湖的最早记录出现于《本草纲目》,其中记载了制盐所用卤水久置后会变红的现象.而针对此类湖泊的科学研究则开始于19世纪,其中杜氏藻最初被认为是造成盐湖变粉的原因,在地中海沿岸红色盐池中被分离出,由Teodoresco将其命名并归类为杜氏藻属(Dunaliella)[1-2].杜氏藻(D.salina)可在体内积累较多的β-胡萝卜素,最高可达细胞干重的16%,因此细胞呈现粉红色[3].研究者也分别从光照、盐度以及水中氮磷含量等角度研究了影响杜氏藻积累类胡萝卜素的原因[4-5].此后有学者认为水体中的嗜盐细菌与嗜盐古菌是造成水体变粉的主要原因.对以色列红色盐水的研究表明,红色盐水中以嗜盐古菌及杜氏藻所含的菌红素以及类胡萝卜素为主[6-8].同时证明虽然杜氏藻所含β-胡萝卜素占比较大,但因其在叶绿素内密集堆叠,光学面积小,因此嗜盐古菌的存在才是导致盐水变红的主要因素.而1999年通过荧光原位杂交技术(FISH)在西班牙盐池发现一种名为Salinibacter的红色细菌在盐池生态系统中占主导地位,证明嗜盐古菌不是唯一造成盐湖变粉的菌类,并在2002年实现菌种分离[9-11].而对Salinibacter培养物的实验室研究证明,杜氏藻光合作用的产物甘油易被Salinibacter吸收并刺激其生长[12].因此造成高盐水体变粉的微生物往往不止一种,可能是杜氏藻、嗜盐古菌与嗜盐细菌的共同作用. ...
Photosynthesis and pigments influenced by light intensity and salinity in the halophile Dunaliella salina (Chlorophyta)
1
1982
... 有关粉色盐湖的最早记录出现于《本草纲目》,其中记载了制盐所用卤水久置后会变红的现象.而针对此类湖泊的科学研究则开始于19世纪,其中杜氏藻最初被认为是造成盐湖变粉的原因,在地中海沿岸红色盐池中被分离出,由Teodoresco将其命名并归类为杜氏藻属(Dunaliella)[1-2].杜氏藻(D.salina)可在体内积累较多的β-胡萝卜素,最高可达细胞干重的16%,因此细胞呈现粉红色[3].研究者也分别从光照、盐度以及水中氮磷含量等角度研究了影响杜氏藻积累类胡萝卜素的原因[4-5].此后有学者认为水体中的嗜盐细菌与嗜盐古菌是造成水体变粉的主要原因.对以色列红色盐水的研究表明,红色盐水中以嗜盐古菌及杜氏藻所含的菌红素以及类胡萝卜素为主[6-8].同时证明虽然杜氏藻所含β-胡萝卜素占比较大,但因其在叶绿素内密集堆叠,光学面积小,因此嗜盐古菌的存在才是导致盐水变红的主要因素.而1999年通过荧光原位杂交技术(FISH)在西班牙盐池发现一种名为Salinibacter的红色细菌在盐池生态系统中占主导地位,证明嗜盐古菌不是唯一造成盐湖变粉的菌类,并在2002年实现菌种分离[9-11].而对Salinibacter培养物的实验室研究证明,杜氏藻光合作用的产物甘油易被Salinibacter吸收并刺激其生长[12].因此造成高盐水体变粉的微生物往往不止一种,可能是杜氏藻、嗜盐古菌与嗜盐细菌的共同作用. ...
氮、磷对盐藻生长及其β-胡萝卜素积累的影响
1
1997
... 有关粉色盐湖的最早记录出现于《本草纲目》,其中记载了制盐所用卤水久置后会变红的现象.而针对此类湖泊的科学研究则开始于19世纪,其中杜氏藻最初被认为是造成盐湖变粉的原因,在地中海沿岸红色盐池中被分离出,由Teodoresco将其命名并归类为杜氏藻属(Dunaliella)[1-2].杜氏藻(D.salina)可在体内积累较多的β-胡萝卜素,最高可达细胞干重的16%,因此细胞呈现粉红色[3].研究者也分别从光照、盐度以及水中氮磷含量等角度研究了影响杜氏藻积累类胡萝卜素的原因[4-5].此后有学者认为水体中的嗜盐细菌与嗜盐古菌是造成水体变粉的主要原因.对以色列红色盐水的研究表明,红色盐水中以嗜盐古菌及杜氏藻所含的菌红素以及类胡萝卜素为主[6-8].同时证明虽然杜氏藻所含β-胡萝卜素占比较大,但因其在叶绿素内密集堆叠,光学面积小,因此嗜盐古菌的存在才是导致盐水变红的主要因素.而1999年通过荧光原位杂交技术(FISH)在西班牙盐池发现一种名为Salinibacter的红色细菌在盐池生态系统中占主导地位,证明嗜盐古菌不是唯一造成盐湖变粉的菌类,并在2002年实现菌种分离[9-11].而对Salinibacter培养物的实验室研究证明,杜氏藻光合作用的产物甘油易被Salinibacter吸收并刺激其生长[12].因此造成高盐水体变粉的微生物往往不止一种,可能是杜氏藻、嗜盐古菌与嗜盐细菌的共同作用. ...
On the red coloration of saltern crystallizer ponds
1
1992
... 有关粉色盐湖的最早记录出现于《本草纲目》,其中记载了制盐所用卤水久置后会变红的现象.而针对此类湖泊的科学研究则开始于19世纪,其中杜氏藻最初被认为是造成盐湖变粉的原因,在地中海沿岸红色盐池中被分离出,由Teodoresco将其命名并归类为杜氏藻属(Dunaliella)[1-2].杜氏藻(D.salina)可在体内积累较多的β-胡萝卜素,最高可达细胞干重的16%,因此细胞呈现粉红色[3].研究者也分别从光照、盐度以及水中氮磷含量等角度研究了影响杜氏藻积累类胡萝卜素的原因[4-5].此后有学者认为水体中的嗜盐细菌与嗜盐古菌是造成水体变粉的主要原因.对以色列红色盐水的研究表明,红色盐水中以嗜盐古菌及杜氏藻所含的菌红素以及类胡萝卜素为主[6-8].同时证明虽然杜氏藻所含β-胡萝卜素占比较大,但因其在叶绿素内密集堆叠,光学面积小,因此嗜盐古菌的存在才是导致盐水变红的主要因素.而1999年通过荧光原位杂交技术(FISH)在西班牙盐池发现一种名为Salinibacter的红色细菌在盐池生态系统中占主导地位,证明嗜盐古菌不是唯一造成盐湖变粉的菌类,并在2002年实现菌种分离[9-11].而对Salinibacter培养物的实验室研究证明,杜氏藻光合作用的产物甘油易被Salinibacter吸收并刺激其生长[12].因此造成高盐水体变粉的微生物往往不止一种,可能是杜氏藻、嗜盐古菌与嗜盐细菌的共同作用. ...
The Wadi Natrun:chemical composition and microbial mass developments in alkaline brines of eutrophic desert lakes
0
1979
The microbiology of red brines
1
2020
... 有关粉色盐湖的最早记录出现于《本草纲目》,其中记载了制盐所用卤水久置后会变红的现象.而针对此类湖泊的科学研究则开始于19世纪,其中杜氏藻最初被认为是造成盐湖变粉的原因,在地中海沿岸红色盐池中被分离出,由Teodoresco将其命名并归类为杜氏藻属(Dunaliella)[1-2].杜氏藻(D.salina)可在体内积累较多的β-胡萝卜素,最高可达细胞干重的16%,因此细胞呈现粉红色[3].研究者也分别从光照、盐度以及水中氮磷含量等角度研究了影响杜氏藻积累类胡萝卜素的原因[4-5].此后有学者认为水体中的嗜盐细菌与嗜盐古菌是造成水体变粉的主要原因.对以色列红色盐水的研究表明,红色盐水中以嗜盐古菌及杜氏藻所含的菌红素以及类胡萝卜素为主[6-8].同时证明虽然杜氏藻所含β-胡萝卜素占比较大,但因其在叶绿素内密集堆叠,光学面积小,因此嗜盐古菌的存在才是导致盐水变红的主要因素.而1999年通过荧光原位杂交技术(FISH)在西班牙盐池发现一种名为Salinibacter的红色细菌在盐池生态系统中占主导地位,证明嗜盐古菌不是唯一造成盐湖变粉的菌类,并在2002年实现菌种分离[9-11].而对Salinibacter培养物的实验室研究证明,杜氏藻光合作用的产物甘油易被Salinibacter吸收并刺激其生长[12].因此造成高盐水体变粉的微生物往往不止一种,可能是杜氏藻、嗜盐古菌与嗜盐细菌的共同作用. ...
Fluorescence in situ hybridization analysis of the prokaryotic community inhabiting crystallizer ponds
1
1999
... 有关粉色盐湖的最早记录出现于《本草纲目》,其中记载了制盐所用卤水久置后会变红的现象.而针对此类湖泊的科学研究则开始于19世纪,其中杜氏藻最初被认为是造成盐湖变粉的原因,在地中海沿岸红色盐池中被分离出,由Teodoresco将其命名并归类为杜氏藻属(Dunaliella)[1-2].杜氏藻(D.salina)可在体内积累较多的β-胡萝卜素,最高可达细胞干重的16%,因此细胞呈现粉红色[3].研究者也分别从光照、盐度以及水中氮磷含量等角度研究了影响杜氏藻积累类胡萝卜素的原因[4-5].此后有学者认为水体中的嗜盐细菌与嗜盐古菌是造成水体变粉的主要原因.对以色列红色盐水的研究表明,红色盐水中以嗜盐古菌及杜氏藻所含的菌红素以及类胡萝卜素为主[6-8].同时证明虽然杜氏藻所含β-胡萝卜素占比较大,但因其在叶绿素内密集堆叠,光学面积小,因此嗜盐古菌的存在才是导致盐水变红的主要因素.而1999年通过荧光原位杂交技术(FISH)在西班牙盐池发现一种名为Salinibacter的红色细菌在盐池生态系统中占主导地位,证明嗜盐古菌不是唯一造成盐湖变粉的菌类,并在2002年实现菌种分离[9-11].而对Salinibacter培养物的实验室研究证明,杜氏藻光合作用的产物甘油易被Salinibacter吸收并刺激其生长[12].因此造成高盐水体变粉的微生物往往不止一种,可能是杜氏藻、嗜盐古菌与嗜盐细菌的共同作用. ...
Salinibacter ruber gen.nov.,sp.nov.,a novel,extremely halophilic member of the Bacteria from saltern crystallizer ponds
0
2002
The genera Rhodothermus,Thermonema,Hymenobacter and Salinibacter
1
2006
... 有关粉色盐湖的最早记录出现于《本草纲目》,其中记载了制盐所用卤水久置后会变红的现象.而针对此类湖泊的科学研究则开始于19世纪,其中杜氏藻最初被认为是造成盐湖变粉的原因,在地中海沿岸红色盐池中被分离出,由Teodoresco将其命名并归类为杜氏藻属(Dunaliella)[1-2].杜氏藻(D.salina)可在体内积累较多的β-胡萝卜素,最高可达细胞干重的16%,因此细胞呈现粉红色[3].研究者也分别从光照、盐度以及水中氮磷含量等角度研究了影响杜氏藻积累类胡萝卜素的原因[4-5].此后有学者认为水体中的嗜盐细菌与嗜盐古菌是造成水体变粉的主要原因.对以色列红色盐水的研究表明,红色盐水中以嗜盐古菌及杜氏藻所含的菌红素以及类胡萝卜素为主[6-8].同时证明虽然杜氏藻所含β-胡萝卜素占比较大,但因其在叶绿素内密集堆叠,光学面积小,因此嗜盐古菌的存在才是导致盐水变红的主要因素.而1999年通过荧光原位杂交技术(FISH)在西班牙盐池发现一种名为Salinibacter的红色细菌在盐池生态系统中占主导地位,证明嗜盐古菌不是唯一造成盐湖变粉的菌类,并在2002年实现菌种分离[9-11].而对Salinibacter培养物的实验室研究证明,杜氏藻光合作用的产物甘油易被Salinibacter吸收并刺激其生长[12].因此造成高盐水体变粉的微生物往往不止一种,可能是杜氏藻、嗜盐古菌与嗜盐细菌的共同作用. ...
Inter relationships between Dunaliella and halophilic prokaryotes in saltern crystallizer ponds
1
2008
... 有关粉色盐湖的最早记录出现于《本草纲目》,其中记载了制盐所用卤水久置后会变红的现象.而针对此类湖泊的科学研究则开始于19世纪,其中杜氏藻最初被认为是造成盐湖变粉的原因,在地中海沿岸红色盐池中被分离出,由Teodoresco将其命名并归类为杜氏藻属(Dunaliella)[1-2].杜氏藻(D.salina)可在体内积累较多的β-胡萝卜素,最高可达细胞干重的16%,因此细胞呈现粉红色[3].研究者也分别从光照、盐度以及水中氮磷含量等角度研究了影响杜氏藻积累类胡萝卜素的原因[4-5].此后有学者认为水体中的嗜盐细菌与嗜盐古菌是造成水体变粉的主要原因.对以色列红色盐水的研究表明,红色盐水中以嗜盐古菌及杜氏藻所含的菌红素以及类胡萝卜素为主[6-8].同时证明虽然杜氏藻所含β-胡萝卜素占比较大,但因其在叶绿素内密集堆叠,光学面积小,因此嗜盐古菌的存在才是导致盐水变红的主要因素.而1999年通过荧光原位杂交技术(FISH)在西班牙盐池发现一种名为Salinibacter的红色细菌在盐池生态系统中占主导地位,证明嗜盐古菌不是唯一造成盐湖变粉的菌类,并在2002年实现菌种分离[9-11].而对Salinibacter培养物的实验室研究证明,杜氏藻光合作用的产物甘油易被Salinibacter吸收并刺激其生长[12].因此造成高盐水体变粉的微生物往往不止一种,可能是杜氏藻、嗜盐古菌与嗜盐细菌的共同作用. ...
试论巴丹吉林沙漠地下水库的发现对西部调水计划的影响
1
2004
... 巴丹吉林沙漠作为中国第二大流动性沙漠,地势南高北低、东高西低.沙漠处于中纬度干旱区,具有典型的大陆性气候特征.多年平均降水量40~80 mm,湖区一带最大不超过90 mm,且降水集中在6—8月,多年平均潜在蒸发量超过1 000 mm,因此沙漠严重缺水[13-14].夏季和冬季的平均气温分别为25.3 ℃和-9.1 ℃,被称为“寒冷的沙漠”[15].年平均风速为2.8~4.6 m·s-1,由南向北逐渐递增,每年4、5月风速最大[16]. ...
Spatial variations in the chemical composition of eolian sediments in hyperarid regions:a case study from the Badain Jaran Desert,northwestern China
1
2018
... 巴丹吉林沙漠作为中国第二大流动性沙漠,地势南高北低、东高西低.沙漠处于中纬度干旱区,具有典型的大陆性气候特征.多年平均降水量40~80 mm,湖区一带最大不超过90 mm,且降水集中在6—8月,多年平均潜在蒸发量超过1 000 mm,因此沙漠严重缺水[13-14].夏季和冬季的平均气温分别为25.3 ℃和-9.1 ℃,被称为“寒冷的沙漠”[15].年平均风速为2.8~4.6 m·s-1,由南向北逐渐递增,每年4、5月风速最大[16]. ...
Surface energy and water vapor fluxes observed on a megadune in the Badain Jaran Desert,China
1
2015
... 巴丹吉林沙漠作为中国第二大流动性沙漠,地势南高北低、东高西低.沙漠处于中纬度干旱区,具有典型的大陆性气候特征.多年平均降水量40~80 mm,湖区一带最大不超过90 mm,且降水集中在6—8月,多年平均潜在蒸发量超过1 000 mm,因此沙漠严重缺水[13-14].夏季和冬季的平均气温分别为25.3 ℃和-9.1 ℃,被称为“寒冷的沙漠”[15].年平均风速为2.8~4.6 m·s-1,由南向北逐渐递增,每年4、5月风速最大[16]. ...
Observation of mega-dune evaporation after various rain events in the hinterland of Badain Jaran Desert,China
1
2014
... 巴丹吉林沙漠作为中国第二大流动性沙漠,地势南高北低、东高西低.沙漠处于中纬度干旱区,具有典型的大陆性气候特征.多年平均降水量40~80 mm,湖区一带最大不超过90 mm,且降水集中在6—8月,多年平均潜在蒸发量超过1 000 mm,因此沙漠严重缺水[13-14].夏季和冬季的平均气温分别为25.3 ℃和-9.1 ℃,被称为“寒冷的沙漠”[15].年平均风速为2.8~4.6 m·s-1,由南向北逐渐递增,每年4、5月风速最大[16]. ...
近40 a巴丹吉林沙漠腹地湖泊面积变化及其影响因素
1
2012
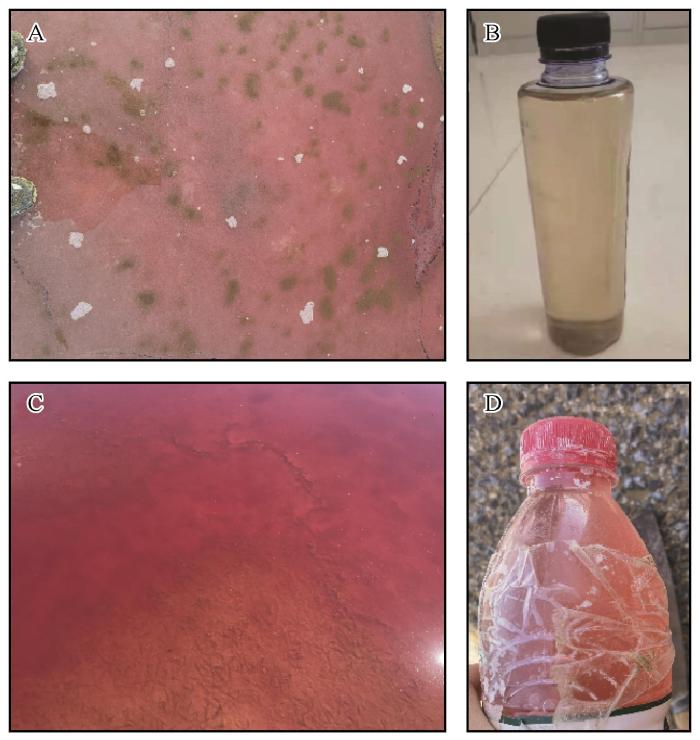
... 巴丹吉林沙漠以世界上最高大的沙山与沙山间的众多湖泊闻名,现共有湖泊146个,常年有水湖泊119个,主要分布在沙漠东南部的沙丘背风坡丘间地[17].这些湖泊多为盐湖,湖水含盐量10~450 g·L-1[18].达格图(39°48′40″N、102°19′58″E,图1)位于沙漠东南部,是巴丹吉林沙漠最为著名的粉色湖泊. ...
巴丹吉林沙漠湖泊水化学类型与钙华沉积关系
1
2022
... 巴丹吉林沙漠以世界上最高大的沙山与沙山间的众多湖泊闻名,现共有湖泊146个,常年有水湖泊119个,主要分布在沙漠东南部的沙丘背风坡丘间地[17].这些湖泊多为盐湖,湖水含盐量10~450 g·L-1[18].达格图(39°48′40″N、102°19′58″E,图1)位于沙漠东南部,是巴丹吉林沙漠最为著名的粉色湖泊. ...
响应土壤阴离子类型的盐碱土古细菌群落多样性研究
1
2018
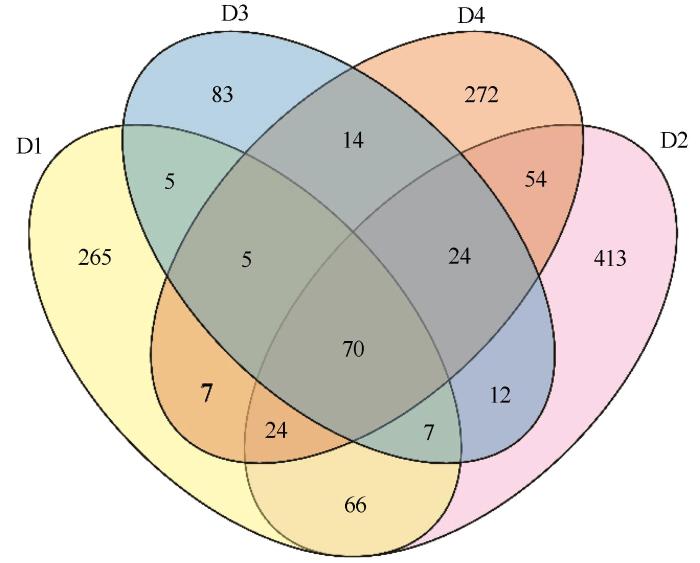
... 从表2可以看出,4个采样点多样性测序结果覆盖率均在99.96%以上,表明当前测序深度下,测序文库接近饱和,数据有效可靠.通过比较4个采样点细菌多样性指数可以看出,D2中ASV数量与Chao1指数均高于其他样品,但其Shannon指数较低,Simpson指数为4个样品中最低,说明D2中细菌群落多样性较低且分布不均[19].因此,D2中虽然具有最多的细菌种类,但其群落多样性较其他采样点较差,表明其细菌群落优势种具有明显的优势性. ...
Signature of the Himalayan salt
1
2020
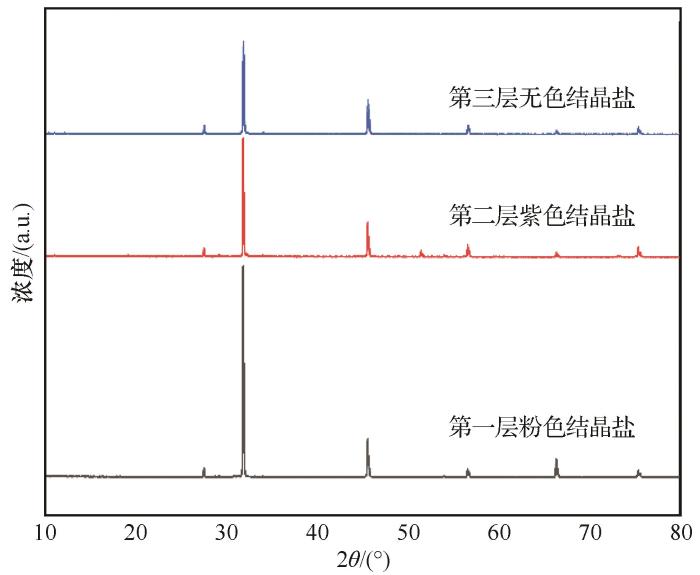
... 盐通常是白色的,但若夹杂了其他矿物,即使是在微量水平也可能会改变它的颜色[20].以上所有矿物中除赤铁矿外,其余矿物通常表现为白色或无色,而赤铁矿通常表现为红色,且其含量变化与结晶盐颜色变化规律类似.因此掺杂了赤铁矿可能是造成结晶盐变粉的主要原因. ...
New C40-carotenoid acyl glycoside as principal carotenoid in Salinibacter ruber,an extremely halophilic eubacterium
1
2002
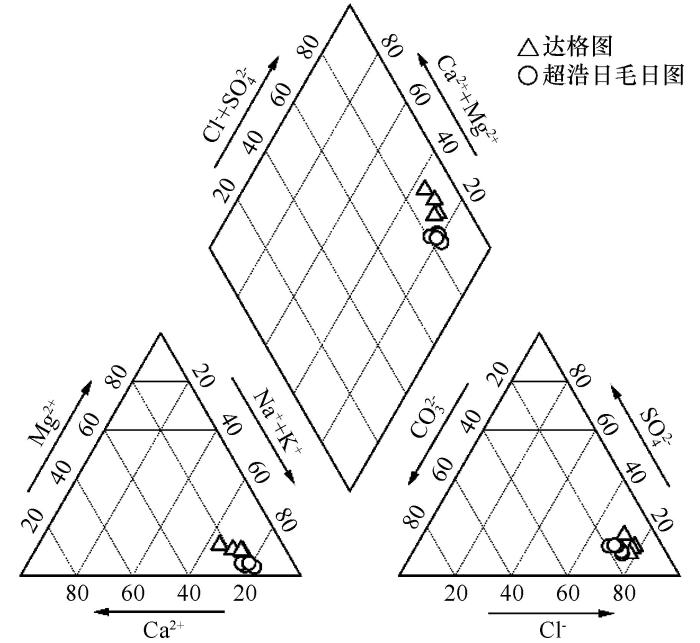
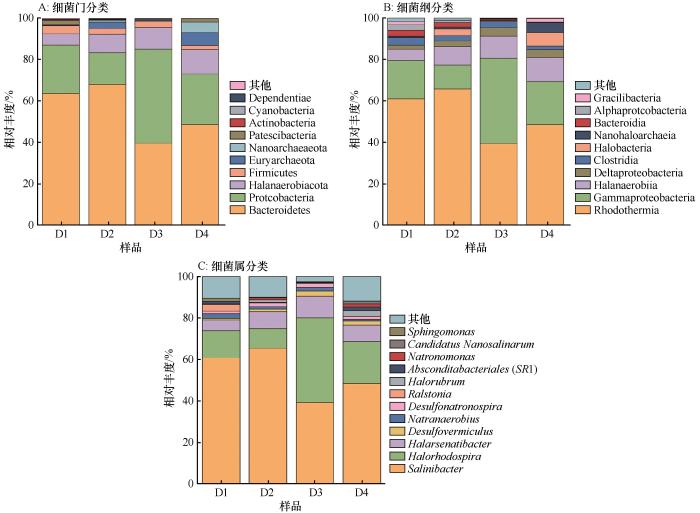
... 夏季湖水中微生物优势种群为Salinibacter、Halorhodospira与Halobacteria,其中Salinibacter与Halorhodospira占比最高,具有明显的优势性.Salinibacte对于高光照环境具有较强的耐受性,这种特性源于其含有的胡萝卜素salinixanthin与视网膜色素xanthorhodopsin,这两类色素的存在使细胞呈现红色[21-22].同属细菌的Halorhodospira属光合厌氧菌,光合色素包含类胡萝卜素,因此其细胞悬液可表现为粉红色.Halobacteria几乎所有成员都因可以合成并积累C50类胡萝卜素(菌红素),而使菌落呈现橙色或亮粉色[23],因此这三类微生物被认为是造成湖水变粉的主要原因[24-25].Salinibacter、Halorhodospira与Halobacteria生长发育都需要较高的盐浓度.Salinibacter无法在15%的盐浓度以下生长,在37 ℃、20%~25%直至盐度饱和情况下生长情况最好[26].Halorhodospira主要存在于高盐度至极端盐度环境中,可忍受轻度至较高碱性(pH最高可达12)[26].而Halobacteria中嗜碱类群生存需要高浓度Na+和Cl-、高pH值和低浓度的Mg2+,它们的生长至少需要1.5 mol·L-1 NaCl,大多数菌种在3.5~4 mol·L-1 NaCl条件下生长最好[27].达格图夏季湖水pH可达9.3以上,全盐量236~443 g·L-1,水中离子以Na+和Cl-为主,同时又具有较低的Mg2+浓度,属于高盐高碱的氯化物型盐湖;湖中全年存在NaCl结晶,说明水体常年盐度饱和.适宜的水环境条件与夏季较高水温,为以上嗜盐嗜碱微生物生存创造适宜的环境[28],因此这类有色微生物在夏季湖水中大量繁殖,造成水体变粉.超浩日毛日图湖水无色透明,表明有色微生物无法大量生存.湖中无结晶盐析出,湖水盐度及总氮、总磷含量较达格图低,Ca2+、Mg2+浓度较高,说明Salinibacter、Halorhodospira与Halobacteria的生长繁殖除需较高的NaCl浓度外,充足的N、P等营养物质会对其产生促进作用,而Ca2+、Mg2+等金属阳离子会抑制生长[29]. ...
The genome of Salinibacter ruber:convergence and gene exchange among hyperhalophilic bacteria and archaea
1
2005
... 夏季湖水中微生物优势种群为Salinibacter、Halorhodospira与Halobacteria,其中Salinibacter与Halorhodospira占比最高,具有明显的优势性.Salinibacte对于高光照环境具有较强的耐受性,这种特性源于其含有的胡萝卜素salinixanthin与视网膜色素xanthorhodopsin,这两类色素的存在使细胞呈现红色[21-22].同属细菌的Halorhodospira属光合厌氧菌,光合色素包含类胡萝卜素,因此其细胞悬液可表现为粉红色.Halobacteria几乎所有成员都因可以合成并积累C50类胡萝卜素(菌红素),而使菌落呈现橙色或亮粉色[23],因此这三类微生物被认为是造成湖水变粉的主要原因[24-25].Salinibacter、Halorhodospira与Halobacteria生长发育都需要较高的盐浓度.Salinibacter无法在15%的盐浓度以下生长,在37 ℃、20%~25%直至盐度饱和情况下生长情况最好[26].Halorhodospira主要存在于高盐度至极端盐度环境中,可忍受轻度至较高碱性(pH最高可达12)[26].而Halobacteria中嗜碱类群生存需要高浓度Na+和Cl-、高pH值和低浓度的Mg2+,它们的生长至少需要1.5 mol·L-1 NaCl,大多数菌种在3.5~4 mol·L-1 NaCl条件下生长最好[27].达格图夏季湖水pH可达9.3以上,全盐量236~443 g·L-1,水中离子以Na+和Cl-为主,同时又具有较低的Mg2+浓度,属于高盐高碱的氯化物型盐湖;湖中全年存在NaCl结晶,说明水体常年盐度饱和.适宜的水环境条件与夏季较高水温,为以上嗜盐嗜碱微生物生存创造适宜的环境[28],因此这类有色微生物在夏季湖水中大量繁殖,造成水体变粉.超浩日毛日图湖水无色透明,表明有色微生物无法大量生存.湖中无结晶盐析出,湖水盐度及总氮、总磷含量较达格图低,Ca2+、Mg2+浓度较高,说明Salinibacter、Halorhodospira与Halobacteria的生长繁殖除需较高的NaCl浓度外,充足的N、P等营养物质会对其产生促进作用,而Ca2+、Mg2+等金属阳离子会抑制生长[29]. ...
Etophysiology of the aerobic halophilic archaebacteria
1
1986
... 夏季湖水中微生物优势种群为Salinibacter、Halorhodospira与Halobacteria,其中Salinibacter与Halorhodospira占比最高,具有明显的优势性.Salinibacte对于高光照环境具有较强的耐受性,这种特性源于其含有的胡萝卜素salinixanthin与视网膜色素xanthorhodopsin,这两类色素的存在使细胞呈现红色[21-22].同属细菌的Halorhodospira属光合厌氧菌,光合色素包含类胡萝卜素,因此其细胞悬液可表现为粉红色.Halobacteria几乎所有成员都因可以合成并积累C50类胡萝卜素(菌红素),而使菌落呈现橙色或亮粉色[23],因此这三类微生物被认为是造成湖水变粉的主要原因[24-25].Salinibacter、Halorhodospira与Halobacteria生长发育都需要较高的盐浓度.Salinibacter无法在15%的盐浓度以下生长,在37 ℃、20%~25%直至盐度饱和情况下生长情况最好[26].Halorhodospira主要存在于高盐度至极端盐度环境中,可忍受轻度至较高碱性(pH最高可达12)[26].而Halobacteria中嗜碱类群生存需要高浓度Na+和Cl-、高pH值和低浓度的Mg2+,它们的生长至少需要1.5 mol·L-1 NaCl,大多数菌种在3.5~4 mol·L-1 NaCl条件下生长最好[27].达格图夏季湖水pH可达9.3以上,全盐量236~443 g·L-1,水中离子以Na+和Cl-为主,同时又具有较低的Mg2+浓度,属于高盐高碱的氯化物型盐湖;湖中全年存在NaCl结晶,说明水体常年盐度饱和.适宜的水环境条件与夏季较高水温,为以上嗜盐嗜碱微生物生存创造适宜的环境[28],因此这类有色微生物在夏季湖水中大量繁殖,造成水体变粉.超浩日毛日图湖水无色透明,表明有色微生物无法大量生存.湖中无结晶盐析出,湖水盐度及总氮、总磷含量较达格图低,Ca2+、Mg2+浓度较高,说明Salinibacter、Halorhodospira与Halobacteria的生长繁殖除需较高的NaCl浓度外,充足的N、P等营养物质会对其产生促进作用,而Ca2+、Mg2+等金属阳离子会抑制生长[29]. ...
On the red coloration of saltern crystallizer ponds:II.additional evidence for the contribution of halobacterial pigments
1
1994
... 夏季湖水中微生物优势种群为Salinibacter、Halorhodospira与Halobacteria,其中Salinibacter与Halorhodospira占比最高,具有明显的优势性.Salinibacte对于高光照环境具有较强的耐受性,这种特性源于其含有的胡萝卜素salinixanthin与视网膜色素xanthorhodopsin,这两类色素的存在使细胞呈现红色[21-22].同属细菌的Halorhodospira属光合厌氧菌,光合色素包含类胡萝卜素,因此其细胞悬液可表现为粉红色.Halobacteria几乎所有成员都因可以合成并积累C50类胡萝卜素(菌红素),而使菌落呈现橙色或亮粉色[23],因此这三类微生物被认为是造成湖水变粉的主要原因[24-25].Salinibacter、Halorhodospira与Halobacteria生长发育都需要较高的盐浓度.Salinibacter无法在15%的盐浓度以下生长,在37 ℃、20%~25%直至盐度饱和情况下生长情况最好[26].Halorhodospira主要存在于高盐度至极端盐度环境中,可忍受轻度至较高碱性(pH最高可达12)[26].而Halobacteria中嗜碱类群生存需要高浓度Na+和Cl-、高pH值和低浓度的Mg2+,它们的生长至少需要1.5 mol·L-1 NaCl,大多数菌种在3.5~4 mol·L-1 NaCl条件下生长最好[27].达格图夏季湖水pH可达9.3以上,全盐量236~443 g·L-1,水中离子以Na+和Cl-为主,同时又具有较低的Mg2+浓度,属于高盐高碱的氯化物型盐湖;湖中全年存在NaCl结晶,说明水体常年盐度饱和.适宜的水环境条件与夏季较高水温,为以上嗜盐嗜碱微生物生存创造适宜的环境[28],因此这类有色微生物在夏季湖水中大量繁殖,造成水体变粉.超浩日毛日图湖水无色透明,表明有色微生物无法大量生存.湖中无结晶盐析出,湖水盐度及总氮、总磷含量较达格图低,Ca2+、Mg2+浓度较高,说明Salinibacter、Halorhodospira与Halobacteria的生长繁殖除需较高的NaCl浓度外,充足的N、P等营养物质会对其产生促进作用,而Ca2+、Mg2+等金属阳离子会抑制生长[29]. ...
The contribution of halophilic Bacteria to the red coloration of saltern crystallizer ponds
1
2001
... 夏季湖水中微生物优势种群为Salinibacter、Halorhodospira与Halobacteria,其中Salinibacter与Halorhodospira占比最高,具有明显的优势性.Salinibacte对于高光照环境具有较强的耐受性,这种特性源于其含有的胡萝卜素salinixanthin与视网膜色素xanthorhodopsin,这两类色素的存在使细胞呈现红色[21-22].同属细菌的Halorhodospira属光合厌氧菌,光合色素包含类胡萝卜素,因此其细胞悬液可表现为粉红色.Halobacteria几乎所有成员都因可以合成并积累C50类胡萝卜素(菌红素),而使菌落呈现橙色或亮粉色[23],因此这三类微生物被认为是造成湖水变粉的主要原因[24-25].Salinibacter、Halorhodospira与Halobacteria生长发育都需要较高的盐浓度.Salinibacter无法在15%的盐浓度以下生长,在37 ℃、20%~25%直至盐度饱和情况下生长情况最好[26].Halorhodospira主要存在于高盐度至极端盐度环境中,可忍受轻度至较高碱性(pH最高可达12)[26].而Halobacteria中嗜碱类群生存需要高浓度Na+和Cl-、高pH值和低浓度的Mg2+,它们的生长至少需要1.5 mol·L-1 NaCl,大多数菌种在3.5~4 mol·L-1 NaCl条件下生长最好[27].达格图夏季湖水pH可达9.3以上,全盐量236~443 g·L-1,水中离子以Na+和Cl-为主,同时又具有较低的Mg2+浓度,属于高盐高碱的氯化物型盐湖;湖中全年存在NaCl结晶,说明水体常年盐度饱和.适宜的水环境条件与夏季较高水温,为以上嗜盐嗜碱微生物生存创造适宜的环境[28],因此这类有色微生物在夏季湖水中大量繁殖,造成水体变粉.超浩日毛日图湖水无色透明,表明有色微生物无法大量生存.湖中无结晶盐析出,湖水盐度及总氮、总磷含量较达格图低,Ca2+、Mg2+浓度较高,说明Salinibacter、Halorhodospira与Halobacteria的生长繁殖除需较高的NaCl浓度外,充足的N、P等营养物质会对其产生促进作用,而Ca2+、Mg2+等金属阳离子会抑制生长[29]. ...
Halorhodospira
2
2015
... 夏季湖水中微生物优势种群为Salinibacter、Halorhodospira与Halobacteria,其中Salinibacter与Halorhodospira占比最高,具有明显的优势性.Salinibacte对于高光照环境具有较强的耐受性,这种特性源于其含有的胡萝卜素salinixanthin与视网膜色素xanthorhodopsin,这两类色素的存在使细胞呈现红色[21-22].同属细菌的Halorhodospira属光合厌氧菌,光合色素包含类胡萝卜素,因此其细胞悬液可表现为粉红色.Halobacteria几乎所有成员都因可以合成并积累C50类胡萝卜素(菌红素),而使菌落呈现橙色或亮粉色[23],因此这三类微生物被认为是造成湖水变粉的主要原因[24-25].Salinibacter、Halorhodospira与Halobacteria生长发育都需要较高的盐浓度.Salinibacter无法在15%的盐浓度以下生长,在37 ℃、20%~25%直至盐度饱和情况下生长情况最好[26].Halorhodospira主要存在于高盐度至极端盐度环境中,可忍受轻度至较高碱性(pH最高可达12)[26].而Halobacteria中嗜碱类群生存需要高浓度Na+和Cl-、高pH值和低浓度的Mg2+,它们的生长至少需要1.5 mol·L-1 NaCl,大多数菌种在3.5~4 mol·L-1 NaCl条件下生长最好[27].达格图夏季湖水pH可达9.3以上,全盐量236~443 g·L-1,水中离子以Na+和Cl-为主,同时又具有较低的Mg2+浓度,属于高盐高碱的氯化物型盐湖;湖中全年存在NaCl结晶,说明水体常年盐度饱和.适宜的水环境条件与夏季较高水温,为以上嗜盐嗜碱微生物生存创造适宜的环境[28],因此这类有色微生物在夏季湖水中大量繁殖,造成水体变粉.超浩日毛日图湖水无色透明,表明有色微生物无法大量生存.湖中无结晶盐析出,湖水盐度及总氮、总磷含量较达格图低,Ca2+、Mg2+浓度较高,说明Salinibacter、Halorhodospira与Halobacteria的生长繁殖除需较高的NaCl浓度外,充足的N、P等营养物质会对其产生促进作用,而Ca2+、Mg2+等金属阳离子会抑制生长[29]. ...
... [26].而Halobacteria中嗜碱类群生存需要高浓度Na+和Cl-、高pH值和低浓度的Mg2+,它们的生长至少需要1.5 mol·L-1 NaCl,大多数菌种在3.5~4 mol·L-1 NaCl条件下生长最好[27].达格图夏季湖水pH可达9.3以上,全盐量236~443 g·L-1,水中离子以Na+和Cl-为主,同时又具有较低的Mg2+浓度,属于高盐高碱的氯化物型盐湖;湖中全年存在NaCl结晶,说明水体常年盐度饱和.适宜的水环境条件与夏季较高水温,为以上嗜盐嗜碱微生物生存创造适宜的环境[28],因此这类有色微生物在夏季湖水中大量繁殖,造成水体变粉.超浩日毛日图湖水无色透明,表明有色微生物无法大量生存.湖中无结晶盐析出,湖水盐度及总氮、总磷含量较达格图低,Ca2+、Mg2+浓度较高,说明Salinibacter、Halorhodospira与Halobacteria的生长繁殖除需较高的NaCl浓度外,充足的N、P等营养物质会对其产生促进作用,而Ca2+、Mg2+等金属阳离子会抑制生长[29]. ...
Extremely halophilic bacteria in crystallizer ponds from solar salterns
1
2000
... 夏季湖水中微生物优势种群为Salinibacter、Halorhodospira与Halobacteria,其中Salinibacter与Halorhodospira占比最高,具有明显的优势性.Salinibacte对于高光照环境具有较强的耐受性,这种特性源于其含有的胡萝卜素salinixanthin与视网膜色素xanthorhodopsin,这两类色素的存在使细胞呈现红色[21-22].同属细菌的Halorhodospira属光合厌氧菌,光合色素包含类胡萝卜素,因此其细胞悬液可表现为粉红色.Halobacteria几乎所有成员都因可以合成并积累C50类胡萝卜素(菌红素),而使菌落呈现橙色或亮粉色[23],因此这三类微生物被认为是造成湖水变粉的主要原因[24-25].Salinibacter、Halorhodospira与Halobacteria生长发育都需要较高的盐浓度.Salinibacter无法在15%的盐浓度以下生长,在37 ℃、20%~25%直至盐度饱和情况下生长情况最好[26].Halorhodospira主要存在于高盐度至极端盐度环境中,可忍受轻度至较高碱性(pH最高可达12)[26].而Halobacteria中嗜碱类群生存需要高浓度Na+和Cl-、高pH值和低浓度的Mg2+,它们的生长至少需要1.5 mol·L-1 NaCl,大多数菌种在3.5~4 mol·L-1 NaCl条件下生长最好[27].达格图夏季湖水pH可达9.3以上,全盐量236~443 g·L-1,水中离子以Na+和Cl-为主,同时又具有较低的Mg2+浓度,属于高盐高碱的氯化物型盐湖;湖中全年存在NaCl结晶,说明水体常年盐度饱和.适宜的水环境条件与夏季较高水温,为以上嗜盐嗜碱微生物生存创造适宜的环境[28],因此这类有色微生物在夏季湖水中大量繁殖,造成水体变粉.超浩日毛日图湖水无色透明,表明有色微生物无法大量生存.湖中无结晶盐析出,湖水盐度及总氮、总磷含量较达格图低,Ca2+、Mg2+浓度较高,说明Salinibacter、Halorhodospira与Halobacteria的生长繁殖除需较高的NaCl浓度外,充足的N、P等营养物质会对其产生促进作用,而Ca2+、Mg2+等金属阳离子会抑制生长[29]. ...
极端嗜盐菌的特性及其应用前景
1
2005
... 夏季湖水中微生物优势种群为Salinibacter、Halorhodospira与Halobacteria,其中Salinibacter与Halorhodospira占比最高,具有明显的优势性.Salinibacte对于高光照环境具有较强的耐受性,这种特性源于其含有的胡萝卜素salinixanthin与视网膜色素xanthorhodopsin,这两类色素的存在使细胞呈现红色[21-22].同属细菌的Halorhodospira属光合厌氧菌,光合色素包含类胡萝卜素,因此其细胞悬液可表现为粉红色.Halobacteria几乎所有成员都因可以合成并积累C50类胡萝卜素(菌红素),而使菌落呈现橙色或亮粉色[23],因此这三类微生物被认为是造成湖水变粉的主要原因[24-25].Salinibacter、Halorhodospira与Halobacteria生长发育都需要较高的盐浓度.Salinibacter无法在15%的盐浓度以下生长,在37 ℃、20%~25%直至盐度饱和情况下生长情况最好[26].Halorhodospira主要存在于高盐度至极端盐度环境中,可忍受轻度至较高碱性(pH最高可达12)[26].而Halobacteria中嗜碱类群生存需要高浓度Na+和Cl-、高pH值和低浓度的Mg2+,它们的生长至少需要1.5 mol·L-1 NaCl,大多数菌种在3.5~4 mol·L-1 NaCl条件下生长最好[27].达格图夏季湖水pH可达9.3以上,全盐量236~443 g·L-1,水中离子以Na+和Cl-为主,同时又具有较低的Mg2+浓度,属于高盐高碱的氯化物型盐湖;湖中全年存在NaCl结晶,说明水体常年盐度饱和.适宜的水环境条件与夏季较高水温,为以上嗜盐嗜碱微生物生存创造适宜的环境[28],因此这类有色微生物在夏季湖水中大量繁殖,造成水体变粉.超浩日毛日图湖水无色透明,表明有色微生物无法大量生存.湖中无结晶盐析出,湖水盐度及总氮、总磷含量较达格图低,Ca2+、Mg2+浓度较高,说明Salinibacter、Halorhodospira与Halobacteria的生长繁殖除需较高的NaCl浓度外,充足的N、P等营养物质会对其产生促进作用,而Ca2+、Mg2+等金属阳离子会抑制生长[29]. ...
短期增温和降水减少对沙质草地土壤微生物量碳氮和酶活性的影响
1
2022
... 夏季湖水中微生物优势种群为Salinibacter、Halorhodospira与Halobacteria,其中Salinibacter与Halorhodospira占比最高,具有明显的优势性.Salinibacte对于高光照环境具有较强的耐受性,这种特性源于其含有的胡萝卜素salinixanthin与视网膜色素xanthorhodopsin,这两类色素的存在使细胞呈现红色[21-22].同属细菌的Halorhodospira属光合厌氧菌,光合色素包含类胡萝卜素,因此其细胞悬液可表现为粉红色.Halobacteria几乎所有成员都因可以合成并积累C50类胡萝卜素(菌红素),而使菌落呈现橙色或亮粉色[23],因此这三类微生物被认为是造成湖水变粉的主要原因[24-25].Salinibacter、Halorhodospira与Halobacteria生长发育都需要较高的盐浓度.Salinibacter无法在15%的盐浓度以下生长,在37 ℃、20%~25%直至盐度饱和情况下生长情况最好[26].Halorhodospira主要存在于高盐度至极端盐度环境中,可忍受轻度至较高碱性(pH最高可达12)[26].而Halobacteria中嗜碱类群生存需要高浓度Na+和Cl-、高pH值和低浓度的Mg2+,它们的生长至少需要1.5 mol·L-1 NaCl,大多数菌种在3.5~4 mol·L-1 NaCl条件下生长最好[27].达格图夏季湖水pH可达9.3以上,全盐量236~443 g·L-1,水中离子以Na+和Cl-为主,同时又具有较低的Mg2+浓度,属于高盐高碱的氯化物型盐湖;湖中全年存在NaCl结晶,说明水体常年盐度饱和.适宜的水环境条件与夏季较高水温,为以上嗜盐嗜碱微生物生存创造适宜的环境[28],因此这类有色微生物在夏季湖水中大量繁殖,造成水体变粉.超浩日毛日图湖水无色透明,表明有色微生物无法大量生存.湖中无结晶盐析出,湖水盐度及总氮、总磷含量较达格图低,Ca2+、Mg2+浓度较高,说明Salinibacter、Halorhodospira与Halobacteria的生长繁殖除需较高的NaCl浓度外,充足的N、P等营养物质会对其产生促进作用,而Ca2+、Mg2+等金属阳离子会抑制生长[29]. ...
巴丹吉林沙漠湖泊季节变化的遥感监测
1
2011
... 秋季达格图湖水盐度仍处于饱和状态,但随着温度降低NaCl溶解度有所下降,湖水盐度较夏季减小.盐度与温度的改变使环境不再适宜Salinibacter与Halobacteria生长繁殖,因此水中有色微生物减少或消失,水体变为无色透明.同时湖泊水量随季节变化逐渐减小[30],大量氯化钠结晶析出并在湖底形成较厚盐层,其中因掺杂赤铁矿使盐晶变为粉色,这些结晶盐透过无色透明的水面使湖泊整体呈现淡粉色. ...
New observational and experimental evidence for the recharge mechanism of the lake group in the Alxa Desert,north-central China
1
2016
... 巴丹吉林沙漠湖泊主要依靠地下水源补给[31-32],外源矿物主要依靠风力作用携带进入湖泊中[33].沙漠表层风积砂中存在赤铁矿,随风进入湖泊后因密度较大沉淀于湖底并在碱性环境下保持化学性质稳定,后随氯化钠一同析出.夏季湖水中厌氧微生物Halorhodospira的大量存在证明湖中为缺氧环境.缺氧条件下可能发生异化铁还原反应[34],使得湖中赤铁矿的Fe(Ⅲ)被还原为Fe(Ⅱ),因此湖中赤铁矿浓度减小.而秋季微生物数量减少造成水中氧浓度升高,Fe(Ⅱ)再次被氧化为Fe(Ⅲ),同时伴随湖泊水量的减少,赤铁矿浓度因此增大,进入氯化钠结晶盐数量随之增多,使结晶盐颜色更深. ...
巴丹吉林沙漠及其高大沙山研究评述
1
2021
... 巴丹吉林沙漠湖泊主要依靠地下水源补给[31-32],外源矿物主要依靠风力作用携带进入湖泊中[33].沙漠表层风积砂中存在赤铁矿,随风进入湖泊后因密度较大沉淀于湖底并在碱性环境下保持化学性质稳定,后随氯化钠一同析出.夏季湖水中厌氧微生物Halorhodospira的大量存在证明湖中为缺氧环境.缺氧条件下可能发生异化铁还原反应[34],使得湖中赤铁矿的Fe(Ⅲ)被还原为Fe(Ⅱ),因此湖中赤铁矿浓度减小.而秋季微生物数量减少造成水中氧浓度升高,Fe(Ⅱ)再次被氧化为Fe(Ⅲ),同时伴随湖泊水量的减少,赤铁矿浓度因此增大,进入氯化钠结晶盐数量随之增多,使结晶盐颜色更深. ...
巴丹吉林沙漠湖泊表层沉积物盐类矿物分布及对气候环境的指示
1
... 巴丹吉林沙漠湖泊主要依靠地下水源补给[31-32],外源矿物主要依靠风力作用携带进入湖泊中[33].沙漠表层风积砂中存在赤铁矿,随风进入湖泊后因密度较大沉淀于湖底并在碱性环境下保持化学性质稳定,后随氯化钠一同析出.夏季湖水中厌氧微生物Halorhodospira的大量存在证明湖中为缺氧环境.缺氧条件下可能发生异化铁还原反应[34],使得湖中赤铁矿的Fe(Ⅲ)被还原为Fe(Ⅱ),因此湖中赤铁矿浓度减小.而秋季微生物数量减少造成水中氧浓度升高,Fe(Ⅱ)再次被氧化为Fe(Ⅲ),同时伴随湖泊水量的减少,赤铁矿浓度因此增大,进入氯化钠结晶盐数量随之增多,使结晶盐颜色更深. ...
厌氧条件下微生物还原含铁矿物实验研究
1
2013
... 巴丹吉林沙漠湖泊主要依靠地下水源补给[31-32],外源矿物主要依靠风力作用携带进入湖泊中[33].沙漠表层风积砂中存在赤铁矿,随风进入湖泊后因密度较大沉淀于湖底并在碱性环境下保持化学性质稳定,后随氯化钠一同析出.夏季湖水中厌氧微生物Halorhodospira的大量存在证明湖中为缺氧环境.缺氧条件下可能发生异化铁还原反应[34],使得湖中赤铁矿的Fe(Ⅲ)被还原为Fe(Ⅱ),因此湖中赤铁矿浓度减小.而秋季微生物数量减少造成水中氧浓度升高,Fe(Ⅱ)再次被氧化为Fe(Ⅲ),同时伴随湖泊水量的减少,赤铁矿浓度因此增大,进入氯化钠结晶盐数量随之增多,使结晶盐颜色更深. ...






 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}