Abiotic stress ssignaling and responses in plants
1
2016
... 在干旱半干旱地区,土壤盐分是限制灌溉农业发展和威胁粮食安全的主要环境因素[1],受盐分影响的耕地面积约占全球耕地总面积的20%[2].同时,受全球气候变化影响,未来受干旱影响的范围将会增大,干旱持续时间将会延长,降水和河流水量将经历更加短暂和剧烈的变动,致使土壤盐分随着水分变化而剧烈波动[3-4],从而加剧土壤次生盐渍化[5-6].土壤水盐失衡进而影响土壤养分的物理扩散和化学形态转化[6],阻碍土壤生产潜力的发挥和抑制植物的生长发育[7-8]. ...
Progress and perspective on drought and salt stress tolerance in cotton
4
2019
... 在干旱半干旱地区,土壤盐分是限制灌溉农业发展和威胁粮食安全的主要环境因素[1],受盐分影响的耕地面积约占全球耕地总面积的20%[2].同时,受全球气候变化影响,未来受干旱影响的范围将会增大,干旱持续时间将会延长,降水和河流水量将经历更加短暂和剧烈的变动,致使土壤盐分随着水分变化而剧烈波动[3-4],从而加剧土壤次生盐渍化[5-6].土壤水盐失衡进而影响土壤养分的物理扩散和化学形态转化[6],阻碍土壤生产潜力的发挥和抑制植物的生长发育[7-8]. ...
... 盐胁迫下土壤中可被根系吸收和利用的水分较少,土壤溶液渗透压超过植物细胞液正常渗透压就会造成植物吸水困难和体内水分亏缺,使渗透胁迫成为植物最先感受到的胁迫[2,28].植物对土壤水分变化的响应极为敏感[29],轻微胁迫也会造成植物生长率大大降低[30],且在营养生长阶段的水分亏缺对作物产量、种子质量和营养价值的影响远大于生殖生长阶段[5]. ...
... 在盐渍土壤中,盐离子胁迫对植物的影响晚于渗透胁迫[2].Na+和Cl-是盐胁迫下对植物产生毒害的主要盐离子[34].保持细胞质低水平的Na+和Cl-含量是降低盐离子毒害的重要方式[40]. ...
... 盐胁迫导致的渗透胁迫和离子胁迫会影响土壤中养分元素的有效性,对植物造成营养亏缺[2].氮(Nitrogen,N)不仅是植物生命活动中最重要的元素,也是植物生长发育最重要的构成组分,植物对N的吸收、运输、同化和分配都要消耗能量[45].植物在生长发育过程中吸收利用的氮源主要为硝态氮(NO)和铵态氮(NH),且吸收NO比NH更消耗能量[46-47].土壤中盐分含量较高时,Cl-与NO间的拮抗作用会限制植物对NO的吸收,Na+与NH间的拮抗作用会限制植物对NH的吸收,而N代谢过程的改变导致植物生存需要消耗更多的ATP[48-49]. ...
Radically rethinking agriculture for the 21st century
1
2010
... 在干旱半干旱地区,土壤盐分是限制灌溉农业发展和威胁粮食安全的主要环境因素[1],受盐分影响的耕地面积约占全球耕地总面积的20%[2].同时,受全球气候变化影响,未来受干旱影响的范围将会增大,干旱持续时间将会延长,降水和河流水量将经历更加短暂和剧烈的变动,致使土壤盐分随着水分变化而剧烈波动[3-4],从而加剧土壤次生盐渍化[5-6].土壤水盐失衡进而影响土壤养分的物理扩散和化学形态转化[6],阻碍土壤生产潜力的发挥和抑制植物的生长发育[7-8]. ...
Alleviation of soil salinization and the management of saline soils,climate change,and soil interactions
1
2020
... 在干旱半干旱地区,土壤盐分是限制灌溉农业发展和威胁粮食安全的主要环境因素[1],受盐分影响的耕地面积约占全球耕地总面积的20%[2].同时,受全球气候变化影响,未来受干旱影响的范围将会增大,干旱持续时间将会延长,降水和河流水量将经历更加短暂和剧烈的变动,致使土壤盐分随着水分变化而剧烈波动[3-4],从而加剧土壤次生盐渍化[5-6].土壤水盐失衡进而影响土壤养分的物理扩散和化学形态转化[6],阻碍土壤生产潜力的发挥和抑制植物的生长发育[7-8]. ...
Plant survival under drought stress:implications,adaptive responses,and integrated rhizosphere management strategy for stress mitigation
2
2021
... 在干旱半干旱地区,土壤盐分是限制灌溉农业发展和威胁粮食安全的主要环境因素[1],受盐分影响的耕地面积约占全球耕地总面积的20%[2].同时,受全球气候变化影响,未来受干旱影响的范围将会增大,干旱持续时间将会延长,降水和河流水量将经历更加短暂和剧烈的变动,致使土壤盐分随着水分变化而剧烈波动[3-4],从而加剧土壤次生盐渍化[5-6].土壤水盐失衡进而影响土壤养分的物理扩散和化学形态转化[6],阻碍土壤生产潜力的发挥和抑制植物的生长发育[7-8]. ...
... 盐胁迫下土壤中可被根系吸收和利用的水分较少,土壤溶液渗透压超过植物细胞液正常渗透压就会造成植物吸水困难和体内水分亏缺,使渗透胁迫成为植物最先感受到的胁迫[2,28].植物对土壤水分变化的响应极为敏感[29],轻微胁迫也会造成植物生长率大大降低[30],且在营养生长阶段的水分亏缺对作物产量、种子质量和营养价值的影响远大于生殖生长阶段[5]. ...
The physiology of plant responses to drought
2
2020
... 在干旱半干旱地区,土壤盐分是限制灌溉农业发展和威胁粮食安全的主要环境因素[1],受盐分影响的耕地面积约占全球耕地总面积的20%[2].同时,受全球气候变化影响,未来受干旱影响的范围将会增大,干旱持续时间将会延长,降水和河流水量将经历更加短暂和剧烈的变动,致使土壤盐分随着水分变化而剧烈波动[3-4],从而加剧土壤次生盐渍化[5-6].土壤水盐失衡进而影响土壤养分的物理扩散和化学形态转化[6],阻碍土壤生产潜力的发挥和抑制植物的生长发育[7-8]. ...
... [6],阻碍土壤生产潜力的发挥和抑制植物的生长发育[7-8]. ...
Microbial inoculum development for ameliorating crop drought stress:a case study of Variovorax paradoxus 5C-2
1
2020
... 在干旱半干旱地区,土壤盐分是限制灌溉农业发展和威胁粮食安全的主要环境因素[1],受盐分影响的耕地面积约占全球耕地总面积的20%[2].同时,受全球气候变化影响,未来受干旱影响的范围将会增大,干旱持续时间将会延长,降水和河流水量将经历更加短暂和剧烈的变动,致使土壤盐分随着水分变化而剧烈波动[3-4],从而加剧土壤次生盐渍化[5-6].土壤水盐失衡进而影响土壤养分的物理扩散和化学形态转化[6],阻碍土壤生产潜力的发挥和抑制植物的生长发育[7-8]. ...
Climate change microbiology-problems and perspectives
1
2019
... 在干旱半干旱地区,土壤盐分是限制灌溉农业发展和威胁粮食安全的主要环境因素[1],受盐分影响的耕地面积约占全球耕地总面积的20%[2].同时,受全球气候变化影响,未来受干旱影响的范围将会增大,干旱持续时间将会延长,降水和河流水量将经历更加短暂和剧烈的变动,致使土壤盐分随着水分变化而剧烈波动[3-4],从而加剧土壤次生盐渍化[5-6].土壤水盐失衡进而影响土壤养分的物理扩散和化学形态转化[6],阻碍土壤生产潜力的发挥和抑制植物的生长发育[7-8]. ...
植物雄性不育与能量代谢的关系
1
2014
... 能量代谢活动的正常运行是支撑植物生长发育的前提[9].然而,土壤中盐分增加会干扰植物种子萌发、营养生长和生殖生长阶段的能量代谢[10-11],导致植物能量产生减少、分解缓慢和消耗增多[11-13].植物通过调整能量代谢对胁迫做出响应,导致用于生长的能量减少而用于防御的能量增加,致使矿质元素的吸收以及矿质元素与同化物的运输受到抑制,最终影响作物产量和品质[12,14]. ...
盐胁迫对植物的影响及AMF提高植物耐盐性的机制
3
2018
... 能量代谢活动的正常运行是支撑植物生长发育的前提[9].然而,土壤中盐分增加会干扰植物种子萌发、营养生长和生殖生长阶段的能量代谢[10-11],导致植物能量产生减少、分解缓慢和消耗增多[11-13].植物通过调整能量代谢对胁迫做出响应,导致用于生长的能量减少而用于防御的能量增加,致使矿质元素的吸收以及矿质元素与同化物的运输受到抑制,最终影响作物产量和品质[12,14]. ...
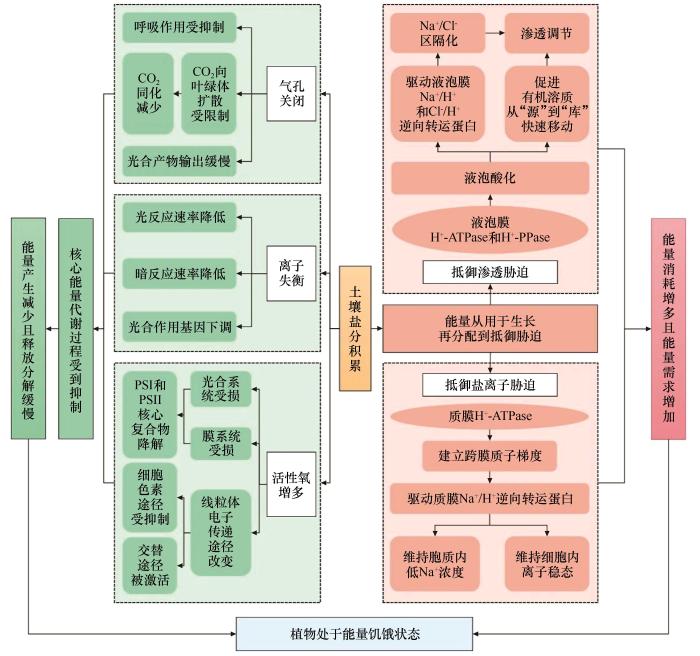
... 盐胁迫会对处于营养生长阶段的植物造成渗透胁迫、离子胁迫和营养亏缺,其共同作用产生的次级氧化胁迫会进而影响光合作用的正常进行[10],导致能量产生减少和释放分解缓慢[26].同时,当植物将用于生长的能量转用于抵御盐胁迫时,能量消耗增多且需求增加[11-13],植物处于能量饥饿状态,甚至会导致植物因能量耗尽而死亡[27](图2). ...
... 盐胁迫下,植物通过从外界环境中吸收无机离子和在体内合成有机溶质的方式进行渗透调节,促进根对水分的吸收[10].K+和Na+等无机离子可作为渗透调节剂对植物进行渗透调节[31].K+在降低细胞渗透势和增加植物耐盐性等方面发挥着重要作用[32].盐逆境胁迫下,以ATP为主要形式的能量供应不足时,K+能够促进以焦磷酸(Pyrophosphate,PPi)作为能量来源的液泡膜H+-PPase的高表达,通过液泡膜H+-ATPase和H+-PPase共同作用使液泡酸化和通过Na+/H+和Cl-/H+逆向转运蛋白对液泡中Na+和Cl-进行区隔化,以保证细胞内能量收支平衡,同时促进有机渗透调节物质(如可溶性糖等)从“源”到“库”的快速移动[13].可见,将盐离子从细胞质中主动隔离到液泡中对植物组织的耐盐性起着至关重要的作用[33].Na+也可作为渗透调节剂来保持植物体内较高的渗透势[34].然而,当液泡膜H+-ATPase和H+-PPase通过驱动Na+/H+逆向转运蛋白将Na+储存在液泡中进行渗透调节时,需要消耗以ATP为主要形式的能量[12],使得植物用于抵御渗透胁迫的能量成本升高[13].同时,当Na+被区隔化到液泡中时需要将其保留,否则Na+会通过慢速和快速液泡通道跨过液泡膜再次进入细胞质,这将会导致Na+进入无效循环,从而使植物消耗更多能量[33]. ...
Can alternative metabolic pathways and shunts overcome salinity induced inhibition of central carbon metabolism in crops?
5
2020
... 能量代谢活动的正常运行是支撑植物生长发育的前提[9].然而,土壤中盐分增加会干扰植物种子萌发、营养生长和生殖生长阶段的能量代谢[10-11],导致植物能量产生减少、分解缓慢和消耗增多[11-13].植物通过调整能量代谢对胁迫做出响应,导致用于生长的能量减少而用于防御的能量增加,致使矿质元素的吸收以及矿质元素与同化物的运输受到抑制,最终影响作物产量和品质[12,14]. ...
... [11-13].植物通过调整能量代谢对胁迫做出响应,导致用于生长的能量减少而用于防御的能量增加,致使矿质元素的吸收以及矿质元素与同化物的运输受到抑制,最终影响作物产量和品质[12,14]. ...
... 为揭示干旱区植物的耐盐机理,学者们已从形态学、解剖学、遗传学、生理生化和分子生物学等方面进行了大量研究,而从能量代谢方面进行的研究相对较少.对盐胁迫下植物的能量代谢过程进行研究,理解盐胁迫下植物能量需求,对于保证植物充足的能源供应和提高植物耐盐性能至关重要[11-12,15].基于此,本文从植物种子萌发、营养生长和生殖生长等3个阶段入手,系统梳理了渗透胁迫、离子胁迫、营养亏缺、氧化胁迫和光合损伤等对植物能量代谢过程的影响,以期为提高干旱区植物的耐盐性能和合理利用盐碱地提供理论依据. ...
... 盐胁迫会对处于营养生长阶段的植物造成渗透胁迫、离子胁迫和营养亏缺,其共同作用产生的次级氧化胁迫会进而影响光合作用的正常进行[10],导致能量产生减少和释放分解缓慢[26].同时,当植物将用于生长的能量转用于抵御盐胁迫时,能量消耗增多且需求增加[11-13],植物处于能量饥饿状态,甚至会导致植物因能量耗尽而死亡[27](图2). ...
... 光合作用是植物生长的基础和新陈代谢的重要途径[54],也是植物吸收、固定和积累能量的重要过程[27,70].在正常条件下,植物将光合作用获得的能量大部分用于植物生长和维持各种生命活动的进行[11].在逆境胁迫下,当植物光合作用积累的能量高于生长和抵御胁迫所消耗的能量时,植物才能成功生长与繁殖[71]. ...
Comprehensive biophysical model of ion and water transport in plant roots.III.quantifying the energy costs of ion transport in salt-stressed roots of Arabidopsis
9
2020
... 能量代谢活动的正常运行是支撑植物生长发育的前提[9].然而,土壤中盐分增加会干扰植物种子萌发、营养生长和生殖生长阶段的能量代谢[10-11],导致植物能量产生减少、分解缓慢和消耗增多[11-13].植物通过调整能量代谢对胁迫做出响应,导致用于生长的能量减少而用于防御的能量增加,致使矿质元素的吸收以及矿质元素与同化物的运输受到抑制,最终影响作物产量和品质[12,14]. ...
... 为揭示干旱区植物的耐盐机理,学者们已从形态学、解剖学、遗传学、生理生化和分子生物学等方面进行了大量研究,而从能量代谢方面进行的研究相对较少.对盐胁迫下植物的能量代谢过程进行研究,理解盐胁迫下植物能量需求,对于保证植物充足的能源供应和提高植物耐盐性能至关重要[11-12,15].基于此,本文从植物种子萌发、营养生长和生殖生长等3个阶段入手,系统梳理了渗透胁迫、离子胁迫、营养亏缺、氧化胁迫和光合损伤等对植物能量代谢过程的影响,以期为提高干旱区植物的耐盐性能和合理利用盐碱地提供理论依据. ...
... 盐胁迫下,植物通过从外界环境中吸收无机离子和在体内合成有机溶质的方式进行渗透调节,促进根对水分的吸收[10].K+和Na+等无机离子可作为渗透调节剂对植物进行渗透调节[31].K+在降低细胞渗透势和增加植物耐盐性等方面发挥着重要作用[32].盐逆境胁迫下,以ATP为主要形式的能量供应不足时,K+能够促进以焦磷酸(Pyrophosphate,PPi)作为能量来源的液泡膜H+-PPase的高表达,通过液泡膜H+-ATPase和H+-PPase共同作用使液泡酸化和通过Na+/H+和Cl-/H+逆向转运蛋白对液泡中Na+和Cl-进行区隔化,以保证细胞内能量收支平衡,同时促进有机渗透调节物质(如可溶性糖等)从“源”到“库”的快速移动[13].可见,将盐离子从细胞质中主动隔离到液泡中对植物组织的耐盐性起着至关重要的作用[33].Na+也可作为渗透调节剂来保持植物体内较高的渗透势[34].然而,当液泡膜H+-ATPase和H+-PPase通过驱动Na+/H+逆向转运蛋白将Na+储存在液泡中进行渗透调节时,需要消耗以ATP为主要形式的能量[12],使得植物用于抵御渗透胁迫的能量成本升高[13].同时,当Na+被区隔化到液泡中时需要将其保留,否则Na+会通过慢速和快速液泡通道跨过液泡膜再次进入细胞质,这将会导致Na+进入无效循环,从而使植物消耗更多能量[33]. ...
... 为应对Na+胁迫和降低Na+毒害,植物根部需要利用ATP为主要形式的能量,通过质膜H+-ATPase产生pH梯度,驱动质膜Na+/H+逆向转运蛋白将Na+泵出细胞外,防止根细胞质中的Na+积累到毒害水平,维持细胞内离子稳态[12,15,34,41].提高质膜H+-ATPase酶活性进行根系皮层细胞质膜的Na+外排可能是整个植物Na+转运中最耗能和最重要的步骤,其能量消耗大于液泡中进行Na+区隔化所消耗的能量[12,42].随盐胁迫程度的升高,Na+外排时消耗的能量和所造成的生物量损失增多.因此,加强根系液泡中Na+的储存、控制Na+从土壤到根的吸收和降低Na+从根到茎中的运输是降低植物能量消耗的有益途径[12]. ...
... [12,42].随盐胁迫程度的升高,Na+外排时消耗的能量和所造成的生物量损失增多.因此,加强根系液泡中Na+的储存、控制Na+从土壤到根的吸收和降低Na+从根到茎中的运输是降低植物能量消耗的有益途径[12]. ...
... [12]. ...
... Cl-是大多数盐渍土壤中最重要的阴离子,它的过度积累不仅会直接对植物细胞造成毒害,而且会改变植物细胞和组织中Na+和K+的可用性和分布[43].因此,保持Cl-稳态对于盐胁迫下植物的生长极为重要.由于植物能量消耗受非原质体和细胞质之间浓度梯度的影响,为保持植物体内电中性,Na+进入引起的势能差可能促进Cl-进入细胞,或Na+和Cl-可能以相同数量同时进入细胞[38],从而对植物造成毒害.然而,Na+和Cl-的吸收、运输和区隔化所消耗的能量是不同的.低盐胁迫下,植物运输Cl-最为消耗能量,而在高盐胁迫下,根质膜上Na+的主动外排和Cl-的主动吸收对能量的需求最大[12].因此,在低盐胁迫下降低Cl-运输和在高盐胁迫下减少根质膜上Na+外排和Cl-吸收,是减少植物能量消耗并提高植物耐盐性的重要途径[12].有趣的是,盐离子在液泡的区隔化使受液泡膜质子泵产生的跨膜电化学梯度驱动液泡膜上的Na+/H+逆向转运蛋白得以实现,但由于液泡膜内外两侧存在的电势差,Na+进行区隔化所要消耗的能量比Cl-更多[44]. ...
... [12].有趣的是,盐离子在液泡的区隔化使受液泡膜质子泵产生的跨膜电化学梯度驱动液泡膜上的Na+/H+逆向转运蛋白得以实现,但由于液泡膜内外两侧存在的电势差,Na+进行区隔化所要消耗的能量比Cl-更多[44]. ...
... 然而,也有研究发现,在植物特定生长阶段施加适度胁迫有利于促进作物生长、提高作物产量和改善籽粒品质[80-81].碳水化合物是植物进行光合作用的主要产物,可分为参与植物结构形态构建的结构性碳水化合物和影响植物生长代谢过程中能量供应的非结构性碳水化合物(Non-structural carbohydrate,NSC)[82].适度干旱胁迫诱导NSC合成与转运相关基因的表达,促进NSC的积累,提高NSC转运量、转运率和表现贡献率,从而为植物生长提供能量,进而提高植物抗性、产量和品质[83-85].因此,充分认识干旱区植物能量需求和植物抵御胁迫的能量消耗,提高胁迫条件下植物代谢和生理过程中能量利用效率、增加外界能量物质的输入和促进生殖生长阶段物质能量从“源”到“库”的运输,是提高产量和品质的重要途径[12,77,86]. ...
Energy costs of salt tolerance in crop plants
9
2020
... 能量代谢活动的正常运行是支撑植物生长发育的前提[9].然而,土壤中盐分增加会干扰植物种子萌发、营养生长和生殖生长阶段的能量代谢[10-11],导致植物能量产生减少、分解缓慢和消耗增多[11-13].植物通过调整能量代谢对胁迫做出响应,导致用于生长的能量减少而用于防御的能量增加,致使矿质元素的吸收以及矿质元素与同化物的运输受到抑制,最终影响作物产量和品质[12,14]. ...
... 盐胁迫会对处于营养生长阶段的植物造成渗透胁迫、离子胁迫和营养亏缺,其共同作用产生的次级氧化胁迫会进而影响光合作用的正常进行[10],导致能量产生减少和释放分解缓慢[26].同时,当植物将用于生长的能量转用于抵御盐胁迫时,能量消耗增多且需求增加[11-13],植物处于能量饥饿状态,甚至会导致植物因能量耗尽而死亡[27](图2). ...
... 盐胁迫下,植物通过从外界环境中吸收无机离子和在体内合成有机溶质的方式进行渗透调节,促进根对水分的吸收[10].K+和Na+等无机离子可作为渗透调节剂对植物进行渗透调节[31].K+在降低细胞渗透势和增加植物耐盐性等方面发挥着重要作用[32].盐逆境胁迫下,以ATP为主要形式的能量供应不足时,K+能够促进以焦磷酸(Pyrophosphate,PPi)作为能量来源的液泡膜H+-PPase的高表达,通过液泡膜H+-ATPase和H+-PPase共同作用使液泡酸化和通过Na+/H+和Cl-/H+逆向转运蛋白对液泡中Na+和Cl-进行区隔化,以保证细胞内能量收支平衡,同时促进有机渗透调节物质(如可溶性糖等)从“源”到“库”的快速移动[13].可见,将盐离子从细胞质中主动隔离到液泡中对植物组织的耐盐性起着至关重要的作用[33].Na+也可作为渗透调节剂来保持植物体内较高的渗透势[34].然而,当液泡膜H+-ATPase和H+-PPase通过驱动Na+/H+逆向转运蛋白将Na+储存在液泡中进行渗透调节时,需要消耗以ATP为主要形式的能量[12],使得植物用于抵御渗透胁迫的能量成本升高[13].同时,当Na+被区隔化到液泡中时需要将其保留,否则Na+会通过慢速和快速液泡通道跨过液泡膜再次进入细胞质,这将会导致Na+进入无效循环,从而使植物消耗更多能量[33]. ...
... [13].同时,当Na+被区隔化到液泡中时需要将其保留,否则Na+会通过慢速和快速液泡通道跨过液泡膜再次进入细胞质,这将会导致Na+进入无效循环,从而使植物消耗更多能量[33]. ...
... 正常生境下,植物依靠细胞器的ROS清除系统维持自身ROS的动态平衡.线粒体是参与植物能量代谢的细胞器和植物根部产生ROS的主要部位[60],也是细胞呼吸链(Respiration chain,RC)和氧化磷酸化的中心[61-62].线粒体电子输运链(Mitochondrial electron transport chain,mETC)是由镶嵌在线粒体内囊体膜上的蛋白复合体组成,是ROS产生的主要位点,其正常运行对维持细胞膜结构的完整性和稳定性至关重要.盐胁迫下,植物细胞中的ROS过多会抑制mETC[63].mETC包含两种途径.第一种途径是以细胞色素氧化酶(Cytochrome oxidase, COX)作为末端氧化酶的细胞色素途径(Cytochrome pathway,CP).CP是产生跨膜质子梯度的主要电子传递途径[64],制造了植物体中95%的能量;CP降低会导致mETC的过分还原,造成ROS产生和ATP降低[65].第二种途径是以交替氧化酶(Alternative oxidase,AOX)作为末端氧化酶的交替途径(Alternative pathway,AP)[13].AP是植物特有的耗能电子传递途径,能加强底物水平磷酸化,部分弥补CP下降造成的ATP不足,可缓解ROS对植物的伤害[65]. ...
... 在盐逆境中产生的渗透胁迫和离子胁迫的相互作用下,植物细胞器中ROS产生与清除间的动态平衡被破坏,过量ROS对植物造成氧化胁迫[66],破坏了细胞膜结构的完整性和稳定性,导致RC的主要途径——CP受抑制和COX活性降低,ATP合成迅速减少[13,67].同时,ROS作为触发植物胁迫防御途径和适应性反应的关键信号[62],激活AP和AOX相关基因[60].AP的激活加强了底物水平磷酸化,调节了细胞能量代谢平衡,部分弥补了CP受抑制后造成的ATP供应不足状况,有利于降低线粒体中ROS含量,使TCA循环能够继续为新陈代谢提高碳价和能量[65,68].因此,AP的激活有利于维持细胞线粒体与叶绿体间、植物生长与抵抗盐胁迫间的能量代谢平衡,建立起光合作用和呼吸作用间的联系[13,67,69].然而,当胁迫程度不断增加后,ROS含量持续增多也会对AOX带来不可逆损伤,破坏ROS对AP的诱导与AP降低ROS生成之间的平衡状态,不利于胁迫条件下的能量供应[60]. ...
... [13,67,69].然而,当胁迫程度不断增加后,ROS含量持续增多也会对AOX带来不可逆损伤,破坏ROS对AP的诱导与AP降低ROS生成之间的平衡状态,不利于胁迫条件下的能量供应[60]. ...
... 盐胁迫下,①水分亏缺致使植物气孔关闭,限制了CO2向叶绿体的扩散,导致CO2同化减少,植物正常物质积累过程受阻,光合产物输出缓慢;②叶绿体中离子失衡导致光反应和暗反应速率降低,光合作用基因和核心能量代谢过程下调,降低了光合作用中能量的产生;③ROS产生过多,对光合系统和膜系统造成氧化损伤,致使光系统I(PSI)和光系统II(PSII)核心复合物降解,最终导致光合速率降低和能量代谢过程受到抑制[13,54,72-73].对沙枣光合作用相关基因进行转录组分析发现,盐胁迫下8个定位于光捕获蛋白的基因表达和8个定位于光合途径的基因表达均被下调,可能会抑制光能的吸收、转移、分布与电子传递,最终导致沙枣体内能量减少[74].可见,盐胁迫下的植物需要更多的周转蛋白对光合系统进行修复和保持叶绿体内离子平衡,使得植物体内能量供应增加[13].然而,在干旱区,当植物处于干旱胁迫时添加一定的盐分反而能够有效减弱干旱对植物光合系统造成的损伤和降低生物量建成成本[39].因此,旱盐胁迫下,盐分对光合系统的影响以及盐分在植物能量代谢过程所起的作用值得今后深入研究. ...
... [13].然而,在干旱区,当植物处于干旱胁迫时添加一定的盐分反而能够有效减弱干旱对植物光合系统造成的损伤和降低生物量建成成本[39].因此,旱盐胁迫下,盐分对光合系统的影响以及盐分在植物能量代谢过程所起的作用值得今后深入研究. ...
Dynamics in plant roots and shoots minimize stress,save energy and maintain water and nutrient uptake
1
2020
... 能量代谢活动的正常运行是支撑植物生长发育的前提[9].然而,土壤中盐分增加会干扰植物种子萌发、营养生长和生殖生长阶段的能量代谢[10-11],导致植物能量产生减少、分解缓慢和消耗增多[11-13].植物通过调整能量代谢对胁迫做出响应,导致用于生长的能量减少而用于防御的能量增加,致使矿质元素的吸收以及矿质元素与同化物的运输受到抑制,最终影响作物产量和品质[12,14]. ...
Energy costs of salinity tolerance in crop plants
2
2018
... 为揭示干旱区植物的耐盐机理,学者们已从形态学、解剖学、遗传学、生理生化和分子生物学等方面进行了大量研究,而从能量代谢方面进行的研究相对较少.对盐胁迫下植物的能量代谢过程进行研究,理解盐胁迫下植物能量需求,对于保证植物充足的能源供应和提高植物耐盐性能至关重要[11-12,15].基于此,本文从植物种子萌发、营养生长和生殖生长等3个阶段入手,系统梳理了渗透胁迫、离子胁迫、营养亏缺、氧化胁迫和光合损伤等对植物能量代谢过程的影响,以期为提高干旱区植物的耐盐性能和合理利用盐碱地提供理论依据. ...
... 为应对Na+胁迫和降低Na+毒害,植物根部需要利用ATP为主要形式的能量,通过质膜H+-ATPase产生pH梯度,驱动质膜Na+/H+逆向转运蛋白将Na+泵出细胞外,防止根细胞质中的Na+积累到毒害水平,维持细胞内离子稳态[12,15,34,41].提高质膜H+-ATPase酶活性进行根系皮层细胞质膜的Na+外排可能是整个植物Na+转运中最耗能和最重要的步骤,其能量消耗大于液泡中进行Na+区隔化所消耗的能量[12,42].随盐胁迫程度的升高,Na+外排时消耗的能量和所造成的生物量损失增多.因此,加强根系液泡中Na+的储存、控制Na+从土壤到根的吸收和降低Na+从根到茎中的运输是降低植物能量消耗的有益途径[12]. ...
水盐浸种对NaCl胁迫下向日葵幼苗渗透调节物质的影响
1
2013
... 种子萌发是植物生长发育的最初阶段.种子迅速萌发和萌发后整齐出苗是保证农业生产的关键[16].种子萌发易受到土壤中水分和盐分的影响,能量在调节种子萌发过程中具有关键作用,并且与种子萌动过程中的吸水速率、物质转化、酶活动和新器官形成等密切相关[17-18].腺苷三磷酸(Adenosine triphosphate,ATP)是植物最重要的能量携带者,也是植物维持生命活动的最重要物质[19],其代谢水平决定着种子萌发的启动[18].种子吸胀是萌发开始的首要步骤.正常生长环境中,种子吸水后酶系统不断修复和活化,种子呼吸速率明显加强,吸涨种子通过细胞质酵解途径、三羧酸循环(Tricarboxylic acid cycle,TCA循环)和线粒体氧化电子传递等3个途径产生ATP[20],从而调节胚胎生长[21].种子可通过调控能量代谢过程和调节能量储存物质等形式确保萌发[22-23]. ...
油松种子萌发初始阶段物质代谢的研究
2
2010
... 种子萌发是植物生长发育的最初阶段.种子迅速萌发和萌发后整齐出苗是保证农业生产的关键[16].种子萌发易受到土壤中水分和盐分的影响,能量在调节种子萌发过程中具有关键作用,并且与种子萌动过程中的吸水速率、物质转化、酶活动和新器官形成等密切相关[17-18].腺苷三磷酸(Adenosine triphosphate,ATP)是植物最重要的能量携带者,也是植物维持生命活动的最重要物质[19],其代谢水平决定着种子萌发的启动[18].种子吸胀是萌发开始的首要步骤.正常生长环境中,种子吸水后酶系统不断修复和活化,种子呼吸速率明显加强,吸涨种子通过细胞质酵解途径、三羧酸循环(Tricarboxylic acid cycle,TCA循环)和线粒体氧化电子传递等3个途径产生ATP[20],从而调节胚胎生长[21].种子可通过调控能量代谢过程和调节能量储存物质等形式确保萌发[22-23]. ...
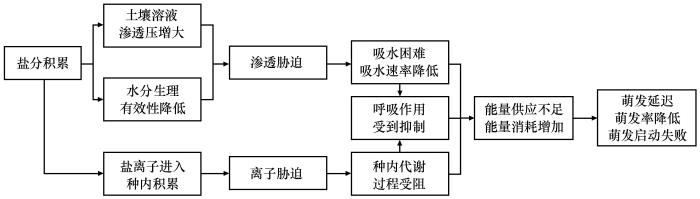
... 土壤盐分积累会对种子萌发过程中的能量代谢产生重要影响[22].首先,渗透胁迫会降低种子的吸水量和吸水速度,造成种子水分吸胀启动延迟,阻碍胚乳水解、种子呼吸代谢和种内物质的储备动员[18,21],使种子萌发时ATP合成和利用受到抑制,导致能量供应不足[18,21-22,24].其次,盐离子对种子细胞膜结构造成破坏,使大量细胞液外渗而盐离子进入种子,进而影响种子内部物质代谢和能量转化[17,25].再次,受渗透胁迫和离子胁迫的影响,种内活性氧(Reactive oxygen species,ROS)平衡体系被破坏,多种细胞成分受到氧化损伤,最终降低种子活力[22].因此,盐胁迫下,种子需要消耗大量能量进行有机渗透调节、盐离子外排和降低ROS对细胞成分的伤害,最终导致种子萌发延迟、萌发率降低,甚至萌发启动失败[20](图1). ...
ACC促进盐胁迫下野生大豆种子萌发与其能量代谢的关系
4
2001
... 种子萌发是植物生长发育的最初阶段.种子迅速萌发和萌发后整齐出苗是保证农业生产的关键[16].种子萌发易受到土壤中水分和盐分的影响,能量在调节种子萌发过程中具有关键作用,并且与种子萌动过程中的吸水速率、物质转化、酶活动和新器官形成等密切相关[17-18].腺苷三磷酸(Adenosine triphosphate,ATP)是植物最重要的能量携带者,也是植物维持生命活动的最重要物质[19],其代谢水平决定着种子萌发的启动[18].种子吸胀是萌发开始的首要步骤.正常生长环境中,种子吸水后酶系统不断修复和活化,种子呼吸速率明显加强,吸涨种子通过细胞质酵解途径、三羧酸循环(Tricarboxylic acid cycle,TCA循环)和线粒体氧化电子传递等3个途径产生ATP[20],从而调节胚胎生长[21].种子可通过调控能量代谢过程和调节能量储存物质等形式确保萌发[22-23]. ...
... [18].种子吸胀是萌发开始的首要步骤.正常生长环境中,种子吸水后酶系统不断修复和活化,种子呼吸速率明显加强,吸涨种子通过细胞质酵解途径、三羧酸循环(Tricarboxylic acid cycle,TCA循环)和线粒体氧化电子传递等3个途径产生ATP[20],从而调节胚胎生长[21].种子可通过调控能量代谢过程和调节能量储存物质等形式确保萌发[22-23]. ...
... 土壤盐分积累会对种子萌发过程中的能量代谢产生重要影响[22].首先,渗透胁迫会降低种子的吸水量和吸水速度,造成种子水分吸胀启动延迟,阻碍胚乳水解、种子呼吸代谢和种内物质的储备动员[18,21],使种子萌发时ATP合成和利用受到抑制,导致能量供应不足[18,21-22,24].其次,盐离子对种子细胞膜结构造成破坏,使大量细胞液外渗而盐离子进入种子,进而影响种子内部物质代谢和能量转化[17,25].再次,受渗透胁迫和离子胁迫的影响,种内活性氧(Reactive oxygen species,ROS)平衡体系被破坏,多种细胞成分受到氧化损伤,最终降低种子活力[22].因此,盐胁迫下,种子需要消耗大量能量进行有机渗透调节、盐离子外排和降低ROS对细胞成分的伤害,最终导致种子萌发延迟、萌发率降低,甚至萌发启动失败[20](图1). ...
... [18,21-22,24].其次,盐离子对种子细胞膜结构造成破坏,使大量细胞液外渗而盐离子进入种子,进而影响种子内部物质代谢和能量转化[17,25].再次,受渗透胁迫和离子胁迫的影响,种内活性氧(Reactive oxygen species,ROS)平衡体系被破坏,多种细胞成分受到氧化损伤,最终降低种子活力[22].因此,盐胁迫下,种子需要消耗大量能量进行有机渗透调节、盐离子外排和降低ROS对细胞成分的伤害,最终导致种子萌发延迟、萌发率降低,甚至萌发启动失败[20](图1). ...
甘薯可溶性蛋白、叶绿素及ATP含量变化与品种抗旱性关系的研究
1
2003
... 种子萌发是植物生长发育的最初阶段.种子迅速萌发和萌发后整齐出苗是保证农业生产的关键[16].种子萌发易受到土壤中水分和盐分的影响,能量在调节种子萌发过程中具有关键作用,并且与种子萌动过程中的吸水速率、物质转化、酶活动和新器官形成等密切相关[17-18].腺苷三磷酸(Adenosine triphosphate,ATP)是植物最重要的能量携带者,也是植物维持生命活动的最重要物质[19],其代谢水平决定着种子萌发的启动[18].种子吸胀是萌发开始的首要步骤.正常生长环境中,种子吸水后酶系统不断修复和活化,种子呼吸速率明显加强,吸涨种子通过细胞质酵解途径、三羧酸循环(Tricarboxylic acid cycle,TCA循环)和线粒体氧化电子传递等3个途径产生ATP[20],从而调节胚胎生长[21].种子可通过调控能量代谢过程和调节能量储存物质等形式确保萌发[22-23]. ...
乙烯消减盐抑制苜蓿种子萌发的机理研究
2
2001
... 种子萌发是植物生长发育的最初阶段.种子迅速萌发和萌发后整齐出苗是保证农业生产的关键[16].种子萌发易受到土壤中水分和盐分的影响,能量在调节种子萌发过程中具有关键作用,并且与种子萌动过程中的吸水速率、物质转化、酶活动和新器官形成等密切相关[17-18].腺苷三磷酸(Adenosine triphosphate,ATP)是植物最重要的能量携带者,也是植物维持生命活动的最重要物质[19],其代谢水平决定着种子萌发的启动[18].种子吸胀是萌发开始的首要步骤.正常生长环境中,种子吸水后酶系统不断修复和活化,种子呼吸速率明显加强,吸涨种子通过细胞质酵解途径、三羧酸循环(Tricarboxylic acid cycle,TCA循环)和线粒体氧化电子传递等3个途径产生ATP[20],从而调节胚胎生长[21].种子可通过调控能量代谢过程和调节能量储存物质等形式确保萌发[22-23]. ...
... 土壤盐分积累会对种子萌发过程中的能量代谢产生重要影响[22].首先,渗透胁迫会降低种子的吸水量和吸水速度,造成种子水分吸胀启动延迟,阻碍胚乳水解、种子呼吸代谢和种内物质的储备动员[18,21],使种子萌发时ATP合成和利用受到抑制,导致能量供应不足[18,21-22,24].其次,盐离子对种子细胞膜结构造成破坏,使大量细胞液外渗而盐离子进入种子,进而影响种子内部物质代谢和能量转化[17,25].再次,受渗透胁迫和离子胁迫的影响,种内活性氧(Reactive oxygen species,ROS)平衡体系被破坏,多种细胞成分受到氧化损伤,最终降低种子活力[22].因此,盐胁迫下,种子需要消耗大量能量进行有机渗透调节、盐离子外排和降低ROS对细胞成分的伤害,最终导致种子萌发延迟、萌发率降低,甚至萌发启动失败[20](图1). ...
呼吸途径和能量对种子萌发的调节
3
1990
... 种子萌发是植物生长发育的最初阶段.种子迅速萌发和萌发后整齐出苗是保证农业生产的关键[16].种子萌发易受到土壤中水分和盐分的影响,能量在调节种子萌发过程中具有关键作用,并且与种子萌动过程中的吸水速率、物质转化、酶活动和新器官形成等密切相关[17-18].腺苷三磷酸(Adenosine triphosphate,ATP)是植物最重要的能量携带者,也是植物维持生命活动的最重要物质[19],其代谢水平决定着种子萌发的启动[18].种子吸胀是萌发开始的首要步骤.正常生长环境中,种子吸水后酶系统不断修复和活化,种子呼吸速率明显加强,吸涨种子通过细胞质酵解途径、三羧酸循环(Tricarboxylic acid cycle,TCA循环)和线粒体氧化电子传递等3个途径产生ATP[20],从而调节胚胎生长[21].种子可通过调控能量代谢过程和调节能量储存物质等形式确保萌发[22-23]. ...
... 土壤盐分积累会对种子萌发过程中的能量代谢产生重要影响[22].首先,渗透胁迫会降低种子的吸水量和吸水速度,造成种子水分吸胀启动延迟,阻碍胚乳水解、种子呼吸代谢和种内物质的储备动员[18,21],使种子萌发时ATP合成和利用受到抑制,导致能量供应不足[18,21-22,24].其次,盐离子对种子细胞膜结构造成破坏,使大量细胞液外渗而盐离子进入种子,进而影响种子内部物质代谢和能量转化[17,25].再次,受渗透胁迫和离子胁迫的影响,种内活性氧(Reactive oxygen species,ROS)平衡体系被破坏,多种细胞成分受到氧化损伤,最终降低种子活力[22].因此,盐胁迫下,种子需要消耗大量能量进行有机渗透调节、盐离子外排和降低ROS对细胞成分的伤害,最终导致种子萌发延迟、萌发率降低,甚至萌发启动失败[20](图1). ...
... ,21-22,24].其次,盐离子对种子细胞膜结构造成破坏,使大量细胞液外渗而盐离子进入种子,进而影响种子内部物质代谢和能量转化[17,25].再次,受渗透胁迫和离子胁迫的影响,种内活性氧(Reactive oxygen species,ROS)平衡体系被破坏,多种细胞成分受到氧化损伤,最终降低种子活力[22].因此,盐胁迫下,种子需要消耗大量能量进行有机渗透调节、盐离子外排和降低ROS对细胞成分的伤害,最终导致种子萌发延迟、萌发率降低,甚至萌发启动失败[20](图1). ...
非生物逆境胁迫下的种子萌发调控机制研究进展
4
2021
... 种子萌发是植物生长发育的最初阶段.种子迅速萌发和萌发后整齐出苗是保证农业生产的关键[16].种子萌发易受到土壤中水分和盐分的影响,能量在调节种子萌发过程中具有关键作用,并且与种子萌动过程中的吸水速率、物质转化、酶活动和新器官形成等密切相关[17-18].腺苷三磷酸(Adenosine triphosphate,ATP)是植物最重要的能量携带者,也是植物维持生命活动的最重要物质[19],其代谢水平决定着种子萌发的启动[18].种子吸胀是萌发开始的首要步骤.正常生长环境中,种子吸水后酶系统不断修复和活化,种子呼吸速率明显加强,吸涨种子通过细胞质酵解途径、三羧酸循环(Tricarboxylic acid cycle,TCA循环)和线粒体氧化电子传递等3个途径产生ATP[20],从而调节胚胎生长[21].种子可通过调控能量代谢过程和调节能量储存物质等形式确保萌发[22-23]. ...
... 土壤盐分积累会对种子萌发过程中的能量代谢产生重要影响[22].首先,渗透胁迫会降低种子的吸水量和吸水速度,造成种子水分吸胀启动延迟,阻碍胚乳水解、种子呼吸代谢和种内物质的储备动员[18,21],使种子萌发时ATP合成和利用受到抑制,导致能量供应不足[18,21-22,24].其次,盐离子对种子细胞膜结构造成破坏,使大量细胞液外渗而盐离子进入种子,进而影响种子内部物质代谢和能量转化[17,25].再次,受渗透胁迫和离子胁迫的影响,种内活性氧(Reactive oxygen species,ROS)平衡体系被破坏,多种细胞成分受到氧化损伤,最终降低种子活力[22].因此,盐胁迫下,种子需要消耗大量能量进行有机渗透调节、盐离子外排和降低ROS对细胞成分的伤害,最终导致种子萌发延迟、萌发率降低,甚至萌发启动失败[20](图1). ...
... -22,24].其次,盐离子对种子细胞膜结构造成破坏,使大量细胞液外渗而盐离子进入种子,进而影响种子内部物质代谢和能量转化[17,25].再次,受渗透胁迫和离子胁迫的影响,种内活性氧(Reactive oxygen species,ROS)平衡体系被破坏,多种细胞成分受到氧化损伤,最终降低种子活力[22].因此,盐胁迫下,种子需要消耗大量能量进行有机渗透调节、盐离子外排和降低ROS对细胞成分的伤害,最终导致种子萌发延迟、萌发率降低,甚至萌发启动失败[20](图1). ...
... [22].因此,盐胁迫下,种子需要消耗大量能量进行有机渗透调节、盐离子外排和降低ROS对细胞成分的伤害,最终导致种子萌发延迟、萌发率降低,甚至萌发启动失败[20](图1). ...
种子活力研究进展
1
2006
... 种子萌发是植物生长发育的最初阶段.种子迅速萌发和萌发后整齐出苗是保证农业生产的关键[16].种子萌发易受到土壤中水分和盐分的影响,能量在调节种子萌发过程中具有关键作用,并且与种子萌动过程中的吸水速率、物质转化、酶活动和新器官形成等密切相关[17-18].腺苷三磷酸(Adenosine triphosphate,ATP)是植物最重要的能量携带者,也是植物维持生命活动的最重要物质[19],其代谢水平决定着种子萌发的启动[18].种子吸胀是萌发开始的首要步骤.正常生长环境中,种子吸水后酶系统不断修复和活化,种子呼吸速率明显加强,吸涨种子通过细胞质酵解途径、三羧酸循环(Tricarboxylic acid cycle,TCA循环)和线粒体氧化电子传递等3个途径产生ATP[20],从而调节胚胎生长[21].种子可通过调控能量代谢过程和调节能量储存物质等形式确保萌发[22-23]. ...
人工模拟盐、碱环境对向日葵种子萌发及幼苗生长的影响
1
2008
... 土壤盐分积累会对种子萌发过程中的能量代谢产生重要影响[22].首先,渗透胁迫会降低种子的吸水量和吸水速度,造成种子水分吸胀启动延迟,阻碍胚乳水解、种子呼吸代谢和种内物质的储备动员[18,21],使种子萌发时ATP合成和利用受到抑制,导致能量供应不足[18,21-22,24].其次,盐离子对种子细胞膜结构造成破坏,使大量细胞液外渗而盐离子进入种子,进而影响种子内部物质代谢和能量转化[17,25].再次,受渗透胁迫和离子胁迫的影响,种内活性氧(Reactive oxygen species,ROS)平衡体系被破坏,多种细胞成分受到氧化损伤,最终降低种子活力[22].因此,盐胁迫下,种子需要消耗大量能量进行有机渗透调节、盐离子外排和降低ROS对细胞成分的伤害,最终导致种子萌发延迟、萌发率降低,甚至萌发启动失败[20](图1). ...
盐胁迫对油葵种子活力和幼苗生理生化特性的影响
1
2004
... 土壤盐分积累会对种子萌发过程中的能量代谢产生重要影响[22].首先,渗透胁迫会降低种子的吸水量和吸水速度,造成种子水分吸胀启动延迟,阻碍胚乳水解、种子呼吸代谢和种内物质的储备动员[18,21],使种子萌发时ATP合成和利用受到抑制,导致能量供应不足[18,21-22,24].其次,盐离子对种子细胞膜结构造成破坏,使大量细胞液外渗而盐离子进入种子,进而影响种子内部物质代谢和能量转化[17,25].再次,受渗透胁迫和离子胁迫的影响,种内活性氧(Reactive oxygen species,ROS)平衡体系被破坏,多种细胞成分受到氧化损伤,最终降低种子活力[22].因此,盐胁迫下,种子需要消耗大量能量进行有机渗透调节、盐离子外排和降低ROS对细胞成分的伤害,最终导致种子萌发延迟、萌发率降低,甚至萌发启动失败[20](图1). ...
拟南芥光信号蛋白FHY3/FAR1调控淀粉合成及能量饥饿响应功能分析
1
2017
... 盐胁迫会对处于营养生长阶段的植物造成渗透胁迫、离子胁迫和营养亏缺,其共同作用产生的次级氧化胁迫会进而影响光合作用的正常进行[10],导致能量产生减少和释放分解缓慢[26].同时,当植物将用于生长的能量转用于抵御盐胁迫时,能量消耗增多且需求增加[11-13],植物处于能量饥饿状态,甚至会导致植物因能量耗尽而死亡[27](图2). ...
基于转录组学的植物响应盐胁迫调控机制研究进展
2
2022
... 盐胁迫会对处于营养生长阶段的植物造成渗透胁迫、离子胁迫和营养亏缺,其共同作用产生的次级氧化胁迫会进而影响光合作用的正常进行[10],导致能量产生减少和释放分解缓慢[26].同时,当植物将用于生长的能量转用于抵御盐胁迫时,能量消耗增多且需求增加[11-13],植物处于能量饥饿状态,甚至会导致植物因能量耗尽而死亡[27](图2). ...
... 光合作用是植物生长的基础和新陈代谢的重要途径[54],也是植物吸收、固定和积累能量的重要过程[27,70].在正常条件下,植物将光合作用获得的能量大部分用于植物生长和维持各种生命活动的进行[11].在逆境胁迫下,当植物光合作用积累的能量高于生长和抵御胁迫所消耗的能量时,植物才能成功生长与繁殖[71]. ...
Mechanisms of salinity tolerance
2
2008
... 盐胁迫下土壤中可被根系吸收和利用的水分较少,土壤溶液渗透压超过植物细胞液正常渗透压就会造成植物吸水困难和体内水分亏缺,使渗透胁迫成为植物最先感受到的胁迫[2,28].植物对土壤水分变化的响应极为敏感[29],轻微胁迫也会造成植物生长率大大降低[30],且在营养生长阶段的水分亏缺对作物产量、种子质量和营养价值的影响远大于生殖生长阶段[5]. ...
... 盐胁迫下,虽然有机渗透物质的合成与运输有利于促进植物对水分的吸收[35],但是其合成与运输都需要消耗能量[36],最终导致植物生长变缓甚至停滞[28,37].植物使用有机溶质作为渗透调节物质所消耗的碳架和能量成本远高于无机离子,且导致植物用于生长的能量减少[36-38].同时,不同渗透调节物质的合成所要消耗的能量也存在差异.从ATP利用的角度来看,甜菜碱和葡萄糖的生物合成消耗能量最少,而蔗糖、脯氨酸和海藻糖等的合成消耗ATP则更多[34,38].因此,在单一盐胁迫条件下,优先使用无机离子作为渗透调节剂,减少合成有机溶质的能量消耗,有助于降低盐胁迫下植物能量损失和提高能量利用效率.然而,在干旱区,干旱和盐渍化常常相伴出现,对植物造成旱盐复合胁迫,植物的能量代谢过程不同于单一盐胁迫下的过程.已有研究表明,在干旱胁迫下,土壤中一定浓度的盐分反而有利于大豆利用更多的无机离子进行渗透调节,从而减少合成有机渗透溶质的能量消耗,降低植物生长所需的能量成本[39]. ...
植物干旱胁迫下水分代谢、碳饥饿与死亡机理
1
2013
... 盐胁迫下土壤中可被根系吸收和利用的水分较少,土壤溶液渗透压超过植物细胞液正常渗透压就会造成植物吸水困难和体内水分亏缺,使渗透胁迫成为植物最先感受到的胁迫[2,28].植物对土壤水分变化的响应极为敏感[29],轻微胁迫也会造成植物生长率大大降低[30],且在营养生长阶段的水分亏缺对作物产量、种子质量和营养价值的影响远大于生殖生长阶段[5]. ...
Molecular aspects of osmotic stress in plants
1
1997
... 盐胁迫下土壤中可被根系吸收和利用的水分较少,土壤溶液渗透压超过植物细胞液正常渗透压就会造成植物吸水困难和体内水分亏缺,使渗透胁迫成为植物最先感受到的胁迫[2,28].植物对土壤水分变化的响应极为敏感[29],轻微胁迫也会造成植物生长率大大降低[30],且在营养生长阶段的水分亏缺对作物产量、种子质量和营养价值的影响远大于生殖生长阶段[5]. ...
盐生植物耐盐分子机制的研究进展
1
2013
... 盐胁迫下,植物通过从外界环境中吸收无机离子和在体内合成有机溶质的方式进行渗透调节,促进根对水分的吸收[10].K+和Na+等无机离子可作为渗透调节剂对植物进行渗透调节[31].K+在降低细胞渗透势和增加植物耐盐性等方面发挥着重要作用[32].盐逆境胁迫下,以ATP为主要形式的能量供应不足时,K+能够促进以焦磷酸(Pyrophosphate,PPi)作为能量来源的液泡膜H+-PPase的高表达,通过液泡膜H+-ATPase和H+-PPase共同作用使液泡酸化和通过Na+/H+和Cl-/H+逆向转运蛋白对液泡中Na+和Cl-进行区隔化,以保证细胞内能量收支平衡,同时促进有机渗透调节物质(如可溶性糖等)从“源”到“库”的快速移动[13].可见,将盐离子从细胞质中主动隔离到液泡中对植物组织的耐盐性起着至关重要的作用[33].Na+也可作为渗透调节剂来保持植物体内较高的渗透势[34].然而,当液泡膜H+-ATPase和H+-PPase通过驱动Na+/H+逆向转运蛋白将Na+储存在液泡中进行渗透调节时,需要消耗以ATP为主要形式的能量[12],使得植物用于抵御渗透胁迫的能量成本升高[13].同时,当Na+被区隔化到液泡中时需要将其保留,否则Na+会通过慢速和快速液泡通道跨过液泡膜再次进入细胞质,这将会导致Na+进入无效循环,从而使植物消耗更多能量[33]. ...
植物高亲和钾离子转运蛋白HAK功能研究进展
2
2020
... 盐胁迫下,植物通过从外界环境中吸收无机离子和在体内合成有机溶质的方式进行渗透调节,促进根对水分的吸收[10].K+和Na+等无机离子可作为渗透调节剂对植物进行渗透调节[31].K+在降低细胞渗透势和增加植物耐盐性等方面发挥着重要作用[32].盐逆境胁迫下,以ATP为主要形式的能量供应不足时,K+能够促进以焦磷酸(Pyrophosphate,PPi)作为能量来源的液泡膜H+-PPase的高表达,通过液泡膜H+-ATPase和H+-PPase共同作用使液泡酸化和通过Na+/H+和Cl-/H+逆向转运蛋白对液泡中Na+和Cl-进行区隔化,以保证细胞内能量收支平衡,同时促进有机渗透调节物质(如可溶性糖等)从“源”到“库”的快速移动[13].可见,将盐离子从细胞质中主动隔离到液泡中对植物组织的耐盐性起着至关重要的作用[33].Na+也可作为渗透调节剂来保持植物体内较高的渗透势[34].然而,当液泡膜H+-ATPase和H+-PPase通过驱动Na+/H+逆向转运蛋白将Na+储存在液泡中进行渗透调节时,需要消耗以ATP为主要形式的能量[12],使得植物用于抵御渗透胁迫的能量成本升高[13].同时,当Na+被区隔化到液泡中时需要将其保留,否则Na+会通过慢速和快速液泡通道跨过液泡膜再次进入细胞质,这将会导致Na+进入无效循环,从而使植物消耗更多能量[33]. ...
... K+是植物生长发育所必需的营养元素、“抗逆元素”和“品质元素”[32].在盐胁迫下,Na+会通过抑制高亲和性K+转运系统和内向整流型K+通道来抑制植物对K+的吸收[57],从而导致植物体内K+亏缺[58].减少K+流失和保持细胞内K+浓度是植物耐盐的重要机制[57,59].一般,植物利用单向转运通道和K+/H+同向转运体吸收1 mol的K+需要分别消耗1 mol和2 mol的ATP[57].然而,在盐胁迫下,由于Na+使质膜电位去极化,显著降低了K+的电化学梯度,这不仅影响了植物对K+的吸收,而且K+吸收更多依赖于K+/H+同向转运体,导致植物吸收K+所要消耗的能量增多[57].因此,Na+的增加会导致植物吸收K+所要消耗的能量增多且能量投入增大.K+的外排已成为植物能量由维持生长转为抵御盐胁迫的“开关”[57]. ...
The energy cost of the tonoplast futile sodium
2
2020
... 盐胁迫下,植物通过从外界环境中吸收无机离子和在体内合成有机溶质的方式进行渗透调节,促进根对水分的吸收[10].K+和Na+等无机离子可作为渗透调节剂对植物进行渗透调节[31].K+在降低细胞渗透势和增加植物耐盐性等方面发挥着重要作用[32].盐逆境胁迫下,以ATP为主要形式的能量供应不足时,K+能够促进以焦磷酸(Pyrophosphate,PPi)作为能量来源的液泡膜H+-PPase的高表达,通过液泡膜H+-ATPase和H+-PPase共同作用使液泡酸化和通过Na+/H+和Cl-/H+逆向转运蛋白对液泡中Na+和Cl-进行区隔化,以保证细胞内能量收支平衡,同时促进有机渗透调节物质(如可溶性糖等)从“源”到“库”的快速移动[13].可见,将盐离子从细胞质中主动隔离到液泡中对植物组织的耐盐性起着至关重要的作用[33].Na+也可作为渗透调节剂来保持植物体内较高的渗透势[34].然而,当液泡膜H+-ATPase和H+-PPase通过驱动Na+/H+逆向转运蛋白将Na+储存在液泡中进行渗透调节时,需要消耗以ATP为主要形式的能量[12],使得植物用于抵御渗透胁迫的能量成本升高[13].同时,当Na+被区隔化到液泡中时需要将其保留,否则Na+会通过慢速和快速液泡通道跨过液泡膜再次进入细胞质,这将会导致Na+进入无效循环,从而使植物消耗更多能量[33]. ...
... [33]. ...
Tissue tolerance coupled with ionic discrimination can potentially minimize the energy cost of salinity tolerance in rice
4
2020
... 盐胁迫下,植物通过从外界环境中吸收无机离子和在体内合成有机溶质的方式进行渗透调节,促进根对水分的吸收[10].K+和Na+等无机离子可作为渗透调节剂对植物进行渗透调节[31].K+在降低细胞渗透势和增加植物耐盐性等方面发挥着重要作用[32].盐逆境胁迫下,以ATP为主要形式的能量供应不足时,K+能够促进以焦磷酸(Pyrophosphate,PPi)作为能量来源的液泡膜H+-PPase的高表达,通过液泡膜H+-ATPase和H+-PPase共同作用使液泡酸化和通过Na+/H+和Cl-/H+逆向转运蛋白对液泡中Na+和Cl-进行区隔化,以保证细胞内能量收支平衡,同时促进有机渗透调节物质(如可溶性糖等)从“源”到“库”的快速移动[13].可见,将盐离子从细胞质中主动隔离到液泡中对植物组织的耐盐性起着至关重要的作用[33].Na+也可作为渗透调节剂来保持植物体内较高的渗透势[34].然而,当液泡膜H+-ATPase和H+-PPase通过驱动Na+/H+逆向转运蛋白将Na+储存在液泡中进行渗透调节时,需要消耗以ATP为主要形式的能量[12],使得植物用于抵御渗透胁迫的能量成本升高[13].同时,当Na+被区隔化到液泡中时需要将其保留,否则Na+会通过慢速和快速液泡通道跨过液泡膜再次进入细胞质,这将会导致Na+进入无效循环,从而使植物消耗更多能量[33]. ...
... 盐胁迫下,虽然有机渗透物质的合成与运输有利于促进植物对水分的吸收[35],但是其合成与运输都需要消耗能量[36],最终导致植物生长变缓甚至停滞[28,37].植物使用有机溶质作为渗透调节物质所消耗的碳架和能量成本远高于无机离子,且导致植物用于生长的能量减少[36-38].同时,不同渗透调节物质的合成所要消耗的能量也存在差异.从ATP利用的角度来看,甜菜碱和葡萄糖的生物合成消耗能量最少,而蔗糖、脯氨酸和海藻糖等的合成消耗ATP则更多[34,38].因此,在单一盐胁迫条件下,优先使用无机离子作为渗透调节剂,减少合成有机溶质的能量消耗,有助于降低盐胁迫下植物能量损失和提高能量利用效率.然而,在干旱区,干旱和盐渍化常常相伴出现,对植物造成旱盐复合胁迫,植物的能量代谢过程不同于单一盐胁迫下的过程.已有研究表明,在干旱胁迫下,土壤中一定浓度的盐分反而有利于大豆利用更多的无机离子进行渗透调节,从而减少合成有机渗透溶质的能量消耗,降低植物生长所需的能量成本[39]. ...
... 在盐渍土壤中,盐离子胁迫对植物的影响晚于渗透胁迫[2].Na+和Cl-是盐胁迫下对植物产生毒害的主要盐离子[34].保持细胞质低水平的Na+和Cl-含量是降低盐离子毒害的重要方式[40]. ...
... 为应对Na+胁迫和降低Na+毒害,植物根部需要利用ATP为主要形式的能量,通过质膜H+-ATPase产生pH梯度,驱动质膜Na+/H+逆向转运蛋白将Na+泵出细胞外,防止根细胞质中的Na+积累到毒害水平,维持细胞内离子稳态[12,15,34,41].提高质膜H+-ATPase酶活性进行根系皮层细胞质膜的Na+外排可能是整个植物Na+转运中最耗能和最重要的步骤,其能量消耗大于液泡中进行Na+区隔化所消耗的能量[12,42].随盐胁迫程度的升高,Na+外排时消耗的能量和所造成的生物量损失增多.因此,加强根系液泡中Na+的储存、控制Na+从土壤到根的吸收和降低Na+从根到茎中的运输是降低植物能量消耗的有益途径[12]. ...
Do halophytes and glycophytes differ in their interactions with arbuscular mycorrhizal fungi under salt stress?a meta-analysis
1
2020
... 盐胁迫下,虽然有机渗透物质的合成与运输有利于促进植物对水分的吸收[35],但是其合成与运输都需要消耗能量[36],最终导致植物生长变缓甚至停滞[28,37].植物使用有机溶质作为渗透调节物质所消耗的碳架和能量成本远高于无机离子,且导致植物用于生长的能量减少[36-38].同时,不同渗透调节物质的合成所要消耗的能量也存在差异.从ATP利用的角度来看,甜菜碱和葡萄糖的生物合成消耗能量最少,而蔗糖、脯氨酸和海藻糖等的合成消耗ATP则更多[34,38].因此,在单一盐胁迫条件下,优先使用无机离子作为渗透调节剂,减少合成有机溶质的能量消耗,有助于降低盐胁迫下植物能量损失和提高能量利用效率.然而,在干旱区,干旱和盐渍化常常相伴出现,对植物造成旱盐复合胁迫,植物的能量代谢过程不同于单一盐胁迫下的过程.已有研究表明,在干旱胁迫下,土壤中一定浓度的盐分反而有利于大豆利用更多的无机离子进行渗透调节,从而减少合成有机渗透溶质的能量消耗,降低植物生长所需的能量成本[39]. ...
Role of vacuolar membrane transport systems in plant salinity tolerance
2
2023
... 盐胁迫下,虽然有机渗透物质的合成与运输有利于促进植物对水分的吸收[35],但是其合成与运输都需要消耗能量[36],最终导致植物生长变缓甚至停滞[28,37].植物使用有机溶质作为渗透调节物质所消耗的碳架和能量成本远高于无机离子,且导致植物用于生长的能量减少[36-38].同时,不同渗透调节物质的合成所要消耗的能量也存在差异.从ATP利用的角度来看,甜菜碱和葡萄糖的生物合成消耗能量最少,而蔗糖、脯氨酸和海藻糖等的合成消耗ATP则更多[34,38].因此,在单一盐胁迫条件下,优先使用无机离子作为渗透调节剂,减少合成有机溶质的能量消耗,有助于降低盐胁迫下植物能量损失和提高能量利用效率.然而,在干旱区,干旱和盐渍化常常相伴出现,对植物造成旱盐复合胁迫,植物的能量代谢过程不同于单一盐胁迫下的过程.已有研究表明,在干旱胁迫下,土壤中一定浓度的盐分反而有利于大豆利用更多的无机离子进行渗透调节,从而减少合成有机渗透溶质的能量消耗,降低植物生长所需的能量成本[39]. ...
... [36-38].同时,不同渗透调节物质的合成所要消耗的能量也存在差异.从ATP利用的角度来看,甜菜碱和葡萄糖的生物合成消耗能量最少,而蔗糖、脯氨酸和海藻糖等的合成消耗ATP则更多[34,38].因此,在单一盐胁迫条件下,优先使用无机离子作为渗透调节剂,减少合成有机溶质的能量消耗,有助于降低盐胁迫下植物能量损失和提高能量利用效率.然而,在干旱区,干旱和盐渍化常常相伴出现,对植物造成旱盐复合胁迫,植物的能量代谢过程不同于单一盐胁迫下的过程.已有研究表明,在干旱胁迫下,土壤中一定浓度的盐分反而有利于大豆利用更多的无机离子进行渗透调节,从而减少合成有机渗透溶质的能量消耗,降低植物生长所需的能量成本[39]. ...
1
2011
... 盐胁迫下,虽然有机渗透物质的合成与运输有利于促进植物对水分的吸收[35],但是其合成与运输都需要消耗能量[36],最终导致植物生长变缓甚至停滞[28,37].植物使用有机溶质作为渗透调节物质所消耗的碳架和能量成本远高于无机离子,且导致植物用于生长的能量减少[36-38].同时,不同渗透调节物质的合成所要消耗的能量也存在差异.从ATP利用的角度来看,甜菜碱和葡萄糖的生物合成消耗能量最少,而蔗糖、脯氨酸和海藻糖等的合成消耗ATP则更多[34,38].因此,在单一盐胁迫条件下,优先使用无机离子作为渗透调节剂,减少合成有机溶质的能量消耗,有助于降低盐胁迫下植物能量损失和提高能量利用效率.然而,在干旱区,干旱和盐渍化常常相伴出现,对植物造成旱盐复合胁迫,植物的能量代谢过程不同于单一盐胁迫下的过程.已有研究表明,在干旱胁迫下,土壤中一定浓度的盐分反而有利于大豆利用更多的无机离子进行渗透调节,从而减少合成有机渗透溶质的能量消耗,降低植物生长所需的能量成本[39]. ...
Osmotic adjustment and energy limitations to plant growth in saline soil
3
2019
... 盐胁迫下,虽然有机渗透物质的合成与运输有利于促进植物对水分的吸收[35],但是其合成与运输都需要消耗能量[36],最终导致植物生长变缓甚至停滞[28,37].植物使用有机溶质作为渗透调节物质所消耗的碳架和能量成本远高于无机离子,且导致植物用于生长的能量减少[36-38].同时,不同渗透调节物质的合成所要消耗的能量也存在差异.从ATP利用的角度来看,甜菜碱和葡萄糖的生物合成消耗能量最少,而蔗糖、脯氨酸和海藻糖等的合成消耗ATP则更多[34,38].因此,在单一盐胁迫条件下,优先使用无机离子作为渗透调节剂,减少合成有机溶质的能量消耗,有助于降低盐胁迫下植物能量损失和提高能量利用效率.然而,在干旱区,干旱和盐渍化常常相伴出现,对植物造成旱盐复合胁迫,植物的能量代谢过程不同于单一盐胁迫下的过程.已有研究表明,在干旱胁迫下,土壤中一定浓度的盐分反而有利于大豆利用更多的无机离子进行渗透调节,从而减少合成有机渗透溶质的能量消耗,降低植物生长所需的能量成本[39]. ...
... ,38].因此,在单一盐胁迫条件下,优先使用无机离子作为渗透调节剂,减少合成有机溶质的能量消耗,有助于降低盐胁迫下植物能量损失和提高能量利用效率.然而,在干旱区,干旱和盐渍化常常相伴出现,对植物造成旱盐复合胁迫,植物的能量代谢过程不同于单一盐胁迫下的过程.已有研究表明,在干旱胁迫下,土壤中一定浓度的盐分反而有利于大豆利用更多的无机离子进行渗透调节,从而减少合成有机渗透溶质的能量消耗,降低植物生长所需的能量成本[39]. ...
... Cl-是大多数盐渍土壤中最重要的阴离子,它的过度积累不仅会直接对植物细胞造成毒害,而且会改变植物细胞和组织中Na+和K+的可用性和分布[43].因此,保持Cl-稳态对于盐胁迫下植物的生长极为重要.由于植物能量消耗受非原质体和细胞质之间浓度梯度的影响,为保持植物体内电中性,Na+进入引起的势能差可能促进Cl-进入细胞,或Na+和Cl-可能以相同数量同时进入细胞[38],从而对植物造成毒害.然而,Na+和Cl-的吸收、运输和区隔化所消耗的能量是不同的.低盐胁迫下,植物运输Cl-最为消耗能量,而在高盐胁迫下,根质膜上Na+的主动外排和Cl-的主动吸收对能量的需求最大[12].因此,在低盐胁迫下降低Cl-运输和在高盐胁迫下减少根质膜上Na+外排和Cl-吸收,是减少植物能量消耗并提高植物耐盐性的重要途径[12].有趣的是,盐离子在液泡的区隔化使受液泡膜质子泵产生的跨膜电化学梯度驱动液泡膜上的Na+/H+逆向转运蛋白得以实现,但由于液泡膜内外两侧存在的电势差,Na+进行区隔化所要消耗的能量比Cl-更多[44]. ...
旱盐胁迫下大豆的适应性反应及Na+的调节作用
2
2018
... 盐胁迫下,虽然有机渗透物质的合成与运输有利于促进植物对水分的吸收[35],但是其合成与运输都需要消耗能量[36],最终导致植物生长变缓甚至停滞[28,37].植物使用有机溶质作为渗透调节物质所消耗的碳架和能量成本远高于无机离子,且导致植物用于生长的能量减少[36-38].同时,不同渗透调节物质的合成所要消耗的能量也存在差异.从ATP利用的角度来看,甜菜碱和葡萄糖的生物合成消耗能量最少,而蔗糖、脯氨酸和海藻糖等的合成消耗ATP则更多[34,38].因此,在单一盐胁迫条件下,优先使用无机离子作为渗透调节剂,减少合成有机溶质的能量消耗,有助于降低盐胁迫下植物能量损失和提高能量利用效率.然而,在干旱区,干旱和盐渍化常常相伴出现,对植物造成旱盐复合胁迫,植物的能量代谢过程不同于单一盐胁迫下的过程.已有研究表明,在干旱胁迫下,土壤中一定浓度的盐分反而有利于大豆利用更多的无机离子进行渗透调节,从而减少合成有机渗透溶质的能量消耗,降低植物生长所需的能量成本[39]. ...
... 盐胁迫下,①水分亏缺致使植物气孔关闭,限制了CO2向叶绿体的扩散,导致CO2同化减少,植物正常物质积累过程受阻,光合产物输出缓慢;②叶绿体中离子失衡导致光反应和暗反应速率降低,光合作用基因和核心能量代谢过程下调,降低了光合作用中能量的产生;③ROS产生过多,对光合系统和膜系统造成氧化损伤,致使光系统I(PSI)和光系统II(PSII)核心复合物降解,最终导致光合速率降低和能量代谢过程受到抑制[13,54,72-73].对沙枣光合作用相关基因进行转录组分析发现,盐胁迫下8个定位于光捕获蛋白的基因表达和8个定位于光合途径的基因表达均被下调,可能会抑制光能的吸收、转移、分布与电子传递,最终导致沙枣体内能量减少[74].可见,盐胁迫下的植物需要更多的周转蛋白对光合系统进行修复和保持叶绿体内离子平衡,使得植物体内能量供应增加[13].然而,在干旱区,当植物处于干旱胁迫时添加一定的盐分反而能够有效减弱干旱对植物光合系统造成的损伤和降低生物量建成成本[39].因此,旱盐胁迫下,盐分对光合系统的影响以及盐分在植物能量代谢过程所起的作用值得今后深入研究. ...
NaCl胁迫对高粱根、叶鞘和叶片液泡膜ATP酶和焦磷酸酶活性的影响
1
2000
... 在盐渍土壤中,盐离子胁迫对植物的影响晚于渗透胁迫[2].Na+和Cl-是盐胁迫下对植物产生毒害的主要盐离子[34].保持细胞质低水平的Na+和Cl-含量是降低盐离子毒害的重要方式[40]. ...
The plasmamembrane transport systems and adaptation to salinity
1
2014
... 为应对Na+胁迫和降低Na+毒害,植物根部需要利用ATP为主要形式的能量,通过质膜H+-ATPase产生pH梯度,驱动质膜Na+/H+逆向转运蛋白将Na+泵出细胞外,防止根细胞质中的Na+积累到毒害水平,维持细胞内离子稳态[12,15,34,41].提高质膜H+-ATPase酶活性进行根系皮层细胞质膜的Na+外排可能是整个植物Na+转运中最耗能和最重要的步骤,其能量消耗大于液泡中进行Na+区隔化所消耗的能量[12,42].随盐胁迫程度的升高,Na+外排时消耗的能量和所造成的生物量损失增多.因此,加强根系液泡中Na+的储存、控制Na+从土壤到根的吸收和降低Na+从根到茎中的运输是降低植物能量消耗的有益途径[12]. ...
沙漠植物羽毛针禾(Stipagrostis pennata)糖转运蛋白基因SpSWEET3的克隆与表达
1
2023
... 为应对Na+胁迫和降低Na+毒害,植物根部需要利用ATP为主要形式的能量,通过质膜H+-ATPase产生pH梯度,驱动质膜Na+/H+逆向转运蛋白将Na+泵出细胞外,防止根细胞质中的Na+积累到毒害水平,维持细胞内离子稳态[12,15,34,41].提高质膜H+-ATPase酶活性进行根系皮层细胞质膜的Na+外排可能是整个植物Na+转运中最耗能和最重要的步骤,其能量消耗大于液泡中进行Na+区隔化所消耗的能量[12,42].随盐胁迫程度的升高,Na+外排时消耗的能量和所造成的生物量损失增多.因此,加强根系液泡中Na+的储存、控制Na+从土壤到根的吸收和降低Na+从根到茎中的运输是降低植物能量消耗的有益途径[12]. ...
Identification and functional characterization of cation-chloride cotransporters in plants
1
2007
... Cl-是大多数盐渍土壤中最重要的阴离子,它的过度积累不仅会直接对植物细胞造成毒害,而且会改变植物细胞和组织中Na+和K+的可用性和分布[43].因此,保持Cl-稳态对于盐胁迫下植物的生长极为重要.由于植物能量消耗受非原质体和细胞质之间浓度梯度的影响,为保持植物体内电中性,Na+进入引起的势能差可能促进Cl-进入细胞,或Na+和Cl-可能以相同数量同时进入细胞[38],从而对植物造成毒害.然而,Na+和Cl-的吸收、运输和区隔化所消耗的能量是不同的.低盐胁迫下,植物运输Cl-最为消耗能量,而在高盐胁迫下,根质膜上Na+的主动外排和Cl-的主动吸收对能量的需求最大[12].因此,在低盐胁迫下降低Cl-运输和在高盐胁迫下减少根质膜上Na+外排和Cl-吸收,是减少植物能量消耗并提高植物耐盐性的重要途径[12].有趣的是,盐离子在液泡的区隔化使受液泡膜质子泵产生的跨膜电化学梯度驱动液泡膜上的Na+/H+逆向转运蛋白得以实现,但由于液泡膜内外两侧存在的电势差,Na+进行区隔化所要消耗的能量比Cl-更多[44]. ...
盐生草盐分区隔化耐盐机制研究
1
2017
... Cl-是大多数盐渍土壤中最重要的阴离子,它的过度积累不仅会直接对植物细胞造成毒害,而且会改变植物细胞和组织中Na+和K+的可用性和分布[43].因此,保持Cl-稳态对于盐胁迫下植物的生长极为重要.由于植物能量消耗受非原质体和细胞质之间浓度梯度的影响,为保持植物体内电中性,Na+进入引起的势能差可能促进Cl-进入细胞,或Na+和Cl-可能以相同数量同时进入细胞[38],从而对植物造成毒害.然而,Na+和Cl-的吸收、运输和区隔化所消耗的能量是不同的.低盐胁迫下,植物运输Cl-最为消耗能量,而在高盐胁迫下,根质膜上Na+的主动外排和Cl-的主动吸收对能量的需求最大[12].因此,在低盐胁迫下降低Cl-运输和在高盐胁迫下减少根质膜上Na+外排和Cl-吸收,是减少植物能量消耗并提高植物耐盐性的重要途径[12].有趣的是,盐离子在液泡的区隔化使受液泡膜质子泵产生的跨膜电化学梯度驱动液泡膜上的Na+/H+逆向转运蛋白得以实现,但由于液泡膜内外两侧存在的电势差,Na+进行区隔化所要消耗的能量比Cl-更多[44]. ...
氮磷对植物光合作用及碳分配的影响
1
2005
... 盐胁迫导致的渗透胁迫和离子胁迫会影响土壤中养分元素的有效性,对植物造成营养亏缺[2].氮(Nitrogen,N)不仅是植物生命活动中最重要的元素,也是植物生长发育最重要的构成组分,植物对N的吸收、运输、同化和分配都要消耗能量[45].植物在生长发育过程中吸收利用的氮源主要为硝态氮(NO)和铵态氮(NH),且吸收NO比NH更消耗能量[46-47].土壤中盐分含量较高时,Cl-与NO间的拮抗作用会限制植物对NO的吸收,Na+与NH间的拮抗作用会限制植物对NH的吸收,而N代谢过程的改变导致植物生存需要消耗更多的ATP[48-49]. ...
植物硝态氮吸收和转运的调控研究进展
1
2021
... 盐胁迫导致的渗透胁迫和离子胁迫会影响土壤中养分元素的有效性,对植物造成营养亏缺[2].氮(Nitrogen,N)不仅是植物生命活动中最重要的元素,也是植物生长发育最重要的构成组分,植物对N的吸收、运输、同化和分配都要消耗能量[45].植物在生长发育过程中吸收利用的氮源主要为硝态氮(NO)和铵态氮(NH),且吸收NO比NH更消耗能量[46-47].土壤中盐分含量较高时,Cl-与NO间的拮抗作用会限制植物对NO的吸收,Na+与NH间的拮抗作用会限制植物对NH的吸收,而N代谢过程的改变导致植物生存需要消耗更多的ATP[48-49]. ...
植物吸收转运无机氮的生理及分子机制
1
2007
... 盐胁迫导致的渗透胁迫和离子胁迫会影响土壤中养分元素的有效性,对植物造成营养亏缺[2].氮(Nitrogen,N)不仅是植物生命活动中最重要的元素,也是植物生长发育最重要的构成组分,植物对N的吸收、运输、同化和分配都要消耗能量[45].植物在生长发育过程中吸收利用的氮源主要为硝态氮(NO)和铵态氮(NH),且吸收NO比NH更消耗能量[46-47].土壤中盐分含量较高时,Cl-与NO间的拮抗作用会限制植物对NO的吸收,Na+与NH间的拮抗作用会限制植物对NH的吸收,而N代谢过程的改变导致植物生存需要消耗更多的ATP[48-49]. ...
氮代谢参与植物逆境抵抗的作用机理研究进展
1
2020
... 盐胁迫导致的渗透胁迫和离子胁迫会影响土壤中养分元素的有效性,对植物造成营养亏缺[2].氮(Nitrogen,N)不仅是植物生命活动中最重要的元素,也是植物生长发育最重要的构成组分,植物对N的吸收、运输、同化和分配都要消耗能量[45].植物在生长发育过程中吸收利用的氮源主要为硝态氮(NO)和铵态氮(NH),且吸收NO比NH更消耗能量[46-47].土壤中盐分含量较高时,Cl-与NO间的拮抗作用会限制植物对NO的吸收,Na+与NH间的拮抗作用会限制植物对NH的吸收,而N代谢过程的改变导致植物生存需要消耗更多的ATP[48-49]. ...
Nitrogen nutrition and adaptation of glycophytes to saline environment:a review
1
2018
... 盐胁迫导致的渗透胁迫和离子胁迫会影响土壤中养分元素的有效性,对植物造成营养亏缺[2].氮(Nitrogen,N)不仅是植物生命活动中最重要的元素,也是植物生长发育最重要的构成组分,植物对N的吸收、运输、同化和分配都要消耗能量[45].植物在生长发育过程中吸收利用的氮源主要为硝态氮(NO)和铵态氮(NH),且吸收NO比NH更消耗能量[46-47].土壤中盐分含量较高时,Cl-与NO间的拮抗作用会限制植物对NO的吸收,Na+与NH间的拮抗作用会限制植物对NH的吸收,而N代谢过程的改变导致植物生存需要消耗更多的ATP[48-49]. ...
植物吸收和转运磷素的分子机理研究进展
1
2011
... 磷(Phosphorus,P)是核酸、磷脂和ATP等生命大分子的重要组成部分,也是植物体内能量代谢的关键底物,对植物的生长代谢至关重要[50].根系对P的吸收和P在植物细胞和组织间的运输是与H+共运输(H+/Pi)为主要方式,需要能量介导且受质子载体效率限制[51].土壤中Na+与H2PO竞争使PO活性降低,土壤吸附作用也会影响植物对P的有效吸收利用[52-53],导致CO2同化受抑制、光合作用相关基因下调和核心能量代谢过程受阻,抑制了ATP的合成[54],导致植株生长变缓和发育延迟,进而影响植物耐盐性能[55].当植物处于缺P状态时,细胞无法通过正常途径提供充足的能量,细胞质中的PPi可以作为潜在的第二能量供体,来部分替代ATP的功用参与能量代谢过程,为细胞提供能量[56].因此,盐胁迫对植物造成的P饥饿会改变植物体内能量代谢过程[54],PPi作为植物细胞代谢能源为盐胁迫下植物提供的选择性代谢系统是植物适应土壤盐环境的策略[56]. ...
植物磷转运子 PHT1家族研究进展
1
2017
... 磷(Phosphorus,P)是核酸、磷脂和ATP等生命大分子的重要组成部分,也是植物体内能量代谢的关键底物,对植物的生长代谢至关重要[50].根系对P的吸收和P在植物细胞和组织间的运输是与H+共运输(H+/Pi)为主要方式,需要能量介导且受质子载体效率限制[51].土壤中Na+与H2PO竞争使PO活性降低,土壤吸附作用也会影响植物对P的有效吸收利用[52-53],导致CO2同化受抑制、光合作用相关基因下调和核心能量代谢过程受阻,抑制了ATP的合成[54],导致植株生长变缓和发育延迟,进而影响植物耐盐性能[55].当植物处于缺P状态时,细胞无法通过正常途径提供充足的能量,细胞质中的PPi可以作为潜在的第二能量供体,来部分替代ATP的功用参与能量代谢过程,为细胞提供能量[56].因此,盐胁迫对植物造成的P饥饿会改变植物体内能量代谢过程[54],PPi作为植物细胞代谢能源为盐胁迫下植物提供的选择性代谢系统是植物适应土壤盐环境的策略[56]. ...
1
2013
... 磷(Phosphorus,P)是核酸、磷脂和ATP等生命大分子的重要组成部分,也是植物体内能量代谢的关键底物,对植物的生长代谢至关重要[50].根系对P的吸收和P在植物细胞和组织间的运输是与H+共运输(H+/Pi)为主要方式,需要能量介导且受质子载体效率限制[51].土壤中Na+与H2PO竞争使PO活性降低,土壤吸附作用也会影响植物对P的有效吸收利用[52-53],导致CO2同化受抑制、光合作用相关基因下调和核心能量代谢过程受阻,抑制了ATP的合成[54],导致植株生长变缓和发育延迟,进而影响植物耐盐性能[55].当植物处于缺P状态时,细胞无法通过正常途径提供充足的能量,细胞质中的PPi可以作为潜在的第二能量供体,来部分替代ATP的功用参与能量代谢过程,为细胞提供能量[56].因此,盐胁迫对植物造成的P饥饿会改变植物体内能量代谢过程[54],PPi作为植物细胞代谢能源为盐胁迫下植物提供的选择性代谢系统是植物适应土壤盐环境的策略[56]. ...
Na+ tolerance and Na+ transport in higher plants
1
2003
... 磷(Phosphorus,P)是核酸、磷脂和ATP等生命大分子的重要组成部分,也是植物体内能量代谢的关键底物,对植物的生长代谢至关重要[50].根系对P的吸收和P在植物细胞和组织间的运输是与H+共运输(H+/Pi)为主要方式,需要能量介导且受质子载体效率限制[51].土壤中Na+与H2PO竞争使PO活性降低,土壤吸附作用也会影响植物对P的有效吸收利用[52-53],导致CO2同化受抑制、光合作用相关基因下调和核心能量代谢过程受阻,抑制了ATP的合成[54],导致植株生长变缓和发育延迟,进而影响植物耐盐性能[55].当植物处于缺P状态时,细胞无法通过正常途径提供充足的能量,细胞质中的PPi可以作为潜在的第二能量供体,来部分替代ATP的功用参与能量代谢过程,为细胞提供能量[56].因此,盐胁迫对植物造成的P饥饿会改变植物体内能量代谢过程[54],PPi作为植物细胞代谢能源为盐胁迫下植物提供的选择性代谢系统是植物适应土壤盐环境的策略[56]. ...
植物磷转运蛋白基因的研究进展
4
2017
... 磷(Phosphorus,P)是核酸、磷脂和ATP等生命大分子的重要组成部分,也是植物体内能量代谢的关键底物,对植物的生长代谢至关重要[50].根系对P的吸收和P在植物细胞和组织间的运输是与H+共运输(H+/Pi)为主要方式,需要能量介导且受质子载体效率限制[51].土壤中Na+与H2PO竞争使PO活性降低,土壤吸附作用也会影响植物对P的有效吸收利用[52-53],导致CO2同化受抑制、光合作用相关基因下调和核心能量代谢过程受阻,抑制了ATP的合成[54],导致植株生长变缓和发育延迟,进而影响植物耐盐性能[55].当植物处于缺P状态时,细胞无法通过正常途径提供充足的能量,细胞质中的PPi可以作为潜在的第二能量供体,来部分替代ATP的功用参与能量代谢过程,为细胞提供能量[56].因此,盐胁迫对植物造成的P饥饿会改变植物体内能量代谢过程[54],PPi作为植物细胞代谢能源为盐胁迫下植物提供的选择性代谢系统是植物适应土壤盐环境的策略[56]. ...
... [54],PPi作为植物细胞代谢能源为盐胁迫下植物提供的选择性代谢系统是植物适应土壤盐环境的策略[56]. ...
... 光合作用是植物生长的基础和新陈代谢的重要途径[54],也是植物吸收、固定和积累能量的重要过程[27,70].在正常条件下,植物将光合作用获得的能量大部分用于植物生长和维持各种生命活动的进行[11].在逆境胁迫下,当植物光合作用积累的能量高于生长和抵御胁迫所消耗的能量时,植物才能成功生长与繁殖[71]. ...
... 盐胁迫下,①水分亏缺致使植物气孔关闭,限制了CO2向叶绿体的扩散,导致CO2同化减少,植物正常物质积累过程受阻,光合产物输出缓慢;②叶绿体中离子失衡导致光反应和暗反应速率降低,光合作用基因和核心能量代谢过程下调,降低了光合作用中能量的产生;③ROS产生过多,对光合系统和膜系统造成氧化损伤,致使光系统I(PSI)和光系统II(PSII)核心复合物降解,最终导致光合速率降低和能量代谢过程受到抑制[13,54,72-73].对沙枣光合作用相关基因进行转录组分析发现,盐胁迫下8个定位于光捕获蛋白的基因表达和8个定位于光合途径的基因表达均被下调,可能会抑制光能的吸收、转移、分布与电子传递,最终导致沙枣体内能量减少[74].可见,盐胁迫下的植物需要更多的周转蛋白对光合系统进行修复和保持叶绿体内离子平衡,使得植物体内能量供应增加[13].然而,在干旱区,当植物处于干旱胁迫时添加一定的盐分反而能够有效减弱干旱对植物光合系统造成的损伤和降低生物量建成成本[39].因此,旱盐胁迫下,盐分对光合系统的影响以及盐分在植物能量代谢过程所起的作用值得今后深入研究. ...
3
2012
... 磷(Phosphorus,P)是核酸、磷脂和ATP等生命大分子的重要组成部分,也是植物体内能量代谢的关键底物,对植物的生长代谢至关重要[50].根系对P的吸收和P在植物细胞和组织间的运输是与H+共运输(H+/Pi)为主要方式,需要能量介导且受质子载体效率限制[51].土壤中Na+与H2PO竞争使PO活性降低,土壤吸附作用也会影响植物对P的有效吸收利用[52-53],导致CO2同化受抑制、光合作用相关基因下调和核心能量代谢过程受阻,抑制了ATP的合成[54],导致植株生长变缓和发育延迟,进而影响植物耐盐性能[55].当植物处于缺P状态时,细胞无法通过正常途径提供充足的能量,细胞质中的PPi可以作为潜在的第二能量供体,来部分替代ATP的功用参与能量代谢过程,为细胞提供能量[56].因此,盐胁迫对植物造成的P饥饿会改变植物体内能量代谢过程[54],PPi作为植物细胞代谢能源为盐胁迫下植物提供的选择性代谢系统是植物适应土壤盐环境的策略[56]. ...
... 盐胁迫下,叶片同化物输出下降,导致植物用于营养生长和生殖生长的能量供应减少,用于抵御盐胁迫的能量消耗增多,为同化物周转提供的能量不足,致使同化物周转速率降低[55,77].这不仅抑制植物对营养物质的吸收,而且影响植物生殖生理和种子成熟过程,最终影响作物产量和品质[55,78].土壤盐分的升高通过抑制小孢子发生、延长雄蕊花丝、加速细胞程序性死亡、促进受精胚胎衰老和胚珠败育等各个方面影响生殖发育[79],最终导致作物产量降低、营养含量不足和籽粒品质不佳[80]. ...
... [55,78].土壤盐分的升高通过抑制小孢子发生、延长雄蕊花丝、加速细胞程序性死亡、促进受精胚胎衰老和胚珠败育等各个方面影响生殖发育[79],最终导致作物产量降低、营养含量不足和籽粒品质不佳[80]. ...
焦磷酸在植物细胞能量代谢中的作用(综述)
2
1997
... 磷(Phosphorus,P)是核酸、磷脂和ATP等生命大分子的重要组成部分,也是植物体内能量代谢的关键底物,对植物的生长代谢至关重要[50].根系对P的吸收和P在植物细胞和组织间的运输是与H+共运输(H+/Pi)为主要方式,需要能量介导且受质子载体效率限制[51].土壤中Na+与H2PO竞争使PO活性降低,土壤吸附作用也会影响植物对P的有效吸收利用[52-53],导致CO2同化受抑制、光合作用相关基因下调和核心能量代谢过程受阻,抑制了ATP的合成[54],导致植株生长变缓和发育延迟,进而影响植物耐盐性能[55].当植物处于缺P状态时,细胞无法通过正常途径提供充足的能量,细胞质中的PPi可以作为潜在的第二能量供体,来部分替代ATP的功用参与能量代谢过程,为细胞提供能量[56].因此,盐胁迫对植物造成的P饥饿会改变植物体内能量代谢过程[54],PPi作为植物细胞代谢能源为盐胁迫下植物提供的选择性代谢系统是植物适应土壤盐环境的策略[56]. ...
... [56]. ...
Doing 'business as usual' comes with a cost:evaluating energy cost of maintaining plant intracellular K+ homeostasis under saline conditions
5
2019
... K+是植物生长发育所必需的营养元素、“抗逆元素”和“品质元素”[32].在盐胁迫下,Na+会通过抑制高亲和性K+转运系统和内向整流型K+通道来抑制植物对K+的吸收[57],从而导致植物体内K+亏缺[58].减少K+流失和保持细胞内K+浓度是植物耐盐的重要机制[57,59].一般,植物利用单向转运通道和K+/H+同向转运体吸收1 mol的K+需要分别消耗1 mol和2 mol的ATP[57].然而,在盐胁迫下,由于Na+使质膜电位去极化,显著降低了K+的电化学梯度,这不仅影响了植物对K+的吸收,而且K+吸收更多依赖于K+/H+同向转运体,导致植物吸收K+所要消耗的能量增多[57].因此,Na+的增加会导致植物吸收K+所要消耗的能量增多且能量投入增大.K+的外排已成为植物能量由维持生长转为抵御盐胁迫的“开关”[57]. ...
... [57,59].一般,植物利用单向转运通道和K+/H+同向转运体吸收1 mol的K+需要分别消耗1 mol和2 mol的ATP[57].然而,在盐胁迫下,由于Na+使质膜电位去极化,显著降低了K+的电化学梯度,这不仅影响了植物对K+的吸收,而且K+吸收更多依赖于K+/H+同向转运体,导致植物吸收K+所要消耗的能量增多[57].因此,Na+的增加会导致植物吸收K+所要消耗的能量增多且能量投入增大.K+的外排已成为植物能量由维持生长转为抵御盐胁迫的“开关”[57]. ...
... [57].然而,在盐胁迫下,由于Na+使质膜电位去极化,显著降低了K+的电化学梯度,这不仅影响了植物对K+的吸收,而且K+吸收更多依赖于K+/H+同向转运体,导致植物吸收K+所要消耗的能量增多[57].因此,Na+的增加会导致植物吸收K+所要消耗的能量增多且能量投入增大.K+的外排已成为植物能量由维持生长转为抵御盐胁迫的“开关”[57]. ...
... [57].因此,Na+的增加会导致植物吸收K+所要消耗的能量增多且能量投入增大.K+的外排已成为植物能量由维持生长转为抵御盐胁迫的“开关”[57]. ...
... [57]. ...
Durum and bread wheat differ in their ability to retain potassium in leaf mesophyll:implications for salinity stress tolerance
1
2014
... K+是植物生长发育所必需的营养元素、“抗逆元素”和“品质元素”[32].在盐胁迫下,Na+会通过抑制高亲和性K+转运系统和内向整流型K+通道来抑制植物对K+的吸收[57],从而导致植物体内K+亏缺[58].减少K+流失和保持细胞内K+浓度是植物耐盐的重要机制[57,59].一般,植物利用单向转运通道和K+/H+同向转运体吸收1 mol的K+需要分别消耗1 mol和2 mol的ATP[57].然而,在盐胁迫下,由于Na+使质膜电位去极化,显著降低了K+的电化学梯度,这不仅影响了植物对K+的吸收,而且K+吸收更多依赖于K+/H+同向转运体,导致植物吸收K+所要消耗的能量增多[57].因此,Na+的增加会导致植物吸收K+所要消耗的能量增多且能量投入增大.K+的外排已成为植物能量由维持生长转为抵御盐胁迫的“开关”[57]. ...
盐胁迫对燕麦幼苗Na+、K+吸收和离子积累的影响
1
2019
... K+是植物生长发育所必需的营养元素、“抗逆元素”和“品质元素”[32].在盐胁迫下,Na+会通过抑制高亲和性K+转运系统和内向整流型K+通道来抑制植物对K+的吸收[57],从而导致植物体内K+亏缺[58].减少K+流失和保持细胞内K+浓度是植物耐盐的重要机制[57,59].一般,植物利用单向转运通道和K+/H+同向转运体吸收1 mol的K+需要分别消耗1 mol和2 mol的ATP[57].然而,在盐胁迫下,由于Na+使质膜电位去极化,显著降低了K+的电化学梯度,这不仅影响了植物对K+的吸收,而且K+吸收更多依赖于K+/H+同向转运体,导致植物吸收K+所要消耗的能量增多[57].因此,Na+的增加会导致植物吸收K+所要消耗的能量增多且能量投入增大.K+的外排已成为植物能量由维持生长转为抵御盐胁迫的“开关”[57]. ...
干旱胁迫对小麦幼苗抗氰呼吸和活性氧代谢的影响
3
2006
... 正常生境下,植物依靠细胞器的ROS清除系统维持自身ROS的动态平衡.线粒体是参与植物能量代谢的细胞器和植物根部产生ROS的主要部位[60],也是细胞呼吸链(Respiration chain,RC)和氧化磷酸化的中心[61-62].线粒体电子输运链(Mitochondrial electron transport chain,mETC)是由镶嵌在线粒体内囊体膜上的蛋白复合体组成,是ROS产生的主要位点,其正常运行对维持细胞膜结构的完整性和稳定性至关重要.盐胁迫下,植物细胞中的ROS过多会抑制mETC[63].mETC包含两种途径.第一种途径是以细胞色素氧化酶(Cytochrome oxidase, COX)作为末端氧化酶的细胞色素途径(Cytochrome pathway,CP).CP是产生跨膜质子梯度的主要电子传递途径[64],制造了植物体中95%的能量;CP降低会导致mETC的过分还原,造成ROS产生和ATP降低[65].第二种途径是以交替氧化酶(Alternative oxidase,AOX)作为末端氧化酶的交替途径(Alternative pathway,AP)[13].AP是植物特有的耗能电子传递途径,能加强底物水平磷酸化,部分弥补CP下降造成的ATP不足,可缓解ROS对植物的伤害[65]. ...
... 在盐逆境中产生的渗透胁迫和离子胁迫的相互作用下,植物细胞器中ROS产生与清除间的动态平衡被破坏,过量ROS对植物造成氧化胁迫[66],破坏了细胞膜结构的完整性和稳定性,导致RC的主要途径——CP受抑制和COX活性降低,ATP合成迅速减少[13,67].同时,ROS作为触发植物胁迫防御途径和适应性反应的关键信号[62],激活AP和AOX相关基因[60].AP的激活加强了底物水平磷酸化,调节了细胞能量代谢平衡,部分弥补了CP受抑制后造成的ATP供应不足状况,有利于降低线粒体中ROS含量,使TCA循环能够继续为新陈代谢提高碳价和能量[65,68].因此,AP的激活有利于维持细胞线粒体与叶绿体间、植物生长与抵抗盐胁迫间的能量代谢平衡,建立起光合作用和呼吸作用间的联系[13,67,69].然而,当胁迫程度不断增加后,ROS含量持续增多也会对AOX带来不可逆损伤,破坏ROS对AP的诱导与AP降低ROS生成之间的平衡状态,不利于胁迫条件下的能量供应[60]. ...
... [60]. ...
光照强度对紫花地丁物质和能量代谢的影响
2
2020
... 正常生境下,植物依靠细胞器的ROS清除系统维持自身ROS的动态平衡.线粒体是参与植物能量代谢的细胞器和植物根部产生ROS的主要部位[60],也是细胞呼吸链(Respiration chain,RC)和氧化磷酸化的中心[61-62].线粒体电子输运链(Mitochondrial electron transport chain,mETC)是由镶嵌在线粒体内囊体膜上的蛋白复合体组成,是ROS产生的主要位点,其正常运行对维持细胞膜结构的完整性和稳定性至关重要.盐胁迫下,植物细胞中的ROS过多会抑制mETC[63].mETC包含两种途径.第一种途径是以细胞色素氧化酶(Cytochrome oxidase, COX)作为末端氧化酶的细胞色素途径(Cytochrome pathway,CP).CP是产生跨膜质子梯度的主要电子传递途径[64],制造了植物体中95%的能量;CP降低会导致mETC的过分还原,造成ROS产生和ATP降低[65].第二种途径是以交替氧化酶(Alternative oxidase,AOX)作为末端氧化酶的交替途径(Alternative pathway,AP)[13].AP是植物特有的耗能电子传递途径,能加强底物水平磷酸化,部分弥补CP下降造成的ATP不足,可缓解ROS对植物的伤害[65]. ...
... 叶绿体是植物进行光合作用的主要场所,糖、脂肪和蛋白质是光合作用产生的主要有机物质,它们最终通过mETC转运电子生成ATP[61,75].mETC的两种途径在调节植物光合作用的过程中发挥着重要作用[72].正常生长条件下,AP途径和AOX对光合效率无显著影响.然而,盐胁迫导致线粒体AOX通路被上调,可以防止叶绿体中过量还原等价物的积累,避免PSII过还原,保护光合机构免受过量能量的伤害,维持光合机构功能的正常运行[72,76].尽管AOX对逆境条件下植物的光合作用很重要,但目前对盐胁迫下CP和AP途径的变化以及AOX参与调控植物抗逆的相关机制探索仍然较少. ...
干旱胁迫下ROS的产生、清除及信号转导研究进展
2
2021
... 正常生境下,植物依靠细胞器的ROS清除系统维持自身ROS的动态平衡.线粒体是参与植物能量代谢的细胞器和植物根部产生ROS的主要部位[60],也是细胞呼吸链(Respiration chain,RC)和氧化磷酸化的中心[61-62].线粒体电子输运链(Mitochondrial electron transport chain,mETC)是由镶嵌在线粒体内囊体膜上的蛋白复合体组成,是ROS产生的主要位点,其正常运行对维持细胞膜结构的完整性和稳定性至关重要.盐胁迫下,植物细胞中的ROS过多会抑制mETC[63].mETC包含两种途径.第一种途径是以细胞色素氧化酶(Cytochrome oxidase, COX)作为末端氧化酶的细胞色素途径(Cytochrome pathway,CP).CP是产生跨膜质子梯度的主要电子传递途径[64],制造了植物体中95%的能量;CP降低会导致mETC的过分还原,造成ROS产生和ATP降低[65].第二种途径是以交替氧化酶(Alternative oxidase,AOX)作为末端氧化酶的交替途径(Alternative pathway,AP)[13].AP是植物特有的耗能电子传递途径,能加强底物水平磷酸化,部分弥补CP下降造成的ATP不足,可缓解ROS对植物的伤害[65]. ...
... 在盐逆境中产生的渗透胁迫和离子胁迫的相互作用下,植物细胞器中ROS产生与清除间的动态平衡被破坏,过量ROS对植物造成氧化胁迫[66],破坏了细胞膜结构的完整性和稳定性,导致RC的主要途径——CP受抑制和COX活性降低,ATP合成迅速减少[13,67].同时,ROS作为触发植物胁迫防御途径和适应性反应的关键信号[62],激活AP和AOX相关基因[60].AP的激活加强了底物水平磷酸化,调节了细胞能量代谢平衡,部分弥补了CP受抑制后造成的ATP供应不足状况,有利于降低线粒体中ROS含量,使TCA循环能够继续为新陈代谢提高碳价和能量[65,68].因此,AP的激活有利于维持细胞线粒体与叶绿体间、植物生长与抵抗盐胁迫间的能量代谢平衡,建立起光合作用和呼吸作用间的联系[13,67,69].然而,当胁迫程度不断增加后,ROS含量持续增多也会对AOX带来不可逆损伤,破坏ROS对AP的诱导与AP降低ROS生成之间的平衡状态,不利于胁迫条件下的能量供应[60]. ...
低温胁迫对紫花苜蓿根系呼吸作用的影响
1
2020
... 正常生境下,植物依靠细胞器的ROS清除系统维持自身ROS的动态平衡.线粒体是参与植物能量代谢的细胞器和植物根部产生ROS的主要部位[60],也是细胞呼吸链(Respiration chain,RC)和氧化磷酸化的中心[61-62].线粒体电子输运链(Mitochondrial electron transport chain,mETC)是由镶嵌在线粒体内囊体膜上的蛋白复合体组成,是ROS产生的主要位点,其正常运行对维持细胞膜结构的完整性和稳定性至关重要.盐胁迫下,植物细胞中的ROS过多会抑制mETC[63].mETC包含两种途径.第一种途径是以细胞色素氧化酶(Cytochrome oxidase, COX)作为末端氧化酶的细胞色素途径(Cytochrome pathway,CP).CP是产生跨膜质子梯度的主要电子传递途径[64],制造了植物体中95%的能量;CP降低会导致mETC的过分还原,造成ROS产生和ATP降低[65].第二种途径是以交替氧化酶(Alternative oxidase,AOX)作为末端氧化酶的交替途径(Alternative pathway,AP)[13].AP是植物特有的耗能电子传递途径,能加强底物水平磷酸化,部分弥补CP下降造成的ATP不足,可缓解ROS对植物的伤害[65]. ...
植物根系呼吸代谢及影响根系呼吸的环境因子研究进展
1
2011
... 正常生境下,植物依靠细胞器的ROS清除系统维持自身ROS的动态平衡.线粒体是参与植物能量代谢的细胞器和植物根部产生ROS的主要部位[60],也是细胞呼吸链(Respiration chain,RC)和氧化磷酸化的中心[61-62].线粒体电子输运链(Mitochondrial electron transport chain,mETC)是由镶嵌在线粒体内囊体膜上的蛋白复合体组成,是ROS产生的主要位点,其正常运行对维持细胞膜结构的完整性和稳定性至关重要.盐胁迫下,植物细胞中的ROS过多会抑制mETC[63].mETC包含两种途径.第一种途径是以细胞色素氧化酶(Cytochrome oxidase, COX)作为末端氧化酶的细胞色素途径(Cytochrome pathway,CP).CP是产生跨膜质子梯度的主要电子传递途径[64],制造了植物体中95%的能量;CP降低会导致mETC的过分还原,造成ROS产生和ATP降低[65].第二种途径是以交替氧化酶(Alternative oxidase,AOX)作为末端氧化酶的交替途径(Alternative pathway,AP)[13].AP是植物特有的耗能电子传递途径,能加强底物水平磷酸化,部分弥补CP下降造成的ATP不足,可缓解ROS对植物的伤害[65]. ...
等渗的Ca(NO3)2和NaCl胁迫对番茄根系呼吸和活性氧代谢的影响
3
2013
... 正常生境下,植物依靠细胞器的ROS清除系统维持自身ROS的动态平衡.线粒体是参与植物能量代谢的细胞器和植物根部产生ROS的主要部位[60],也是细胞呼吸链(Respiration chain,RC)和氧化磷酸化的中心[61-62].线粒体电子输运链(Mitochondrial electron transport chain,mETC)是由镶嵌在线粒体内囊体膜上的蛋白复合体组成,是ROS产生的主要位点,其正常运行对维持细胞膜结构的完整性和稳定性至关重要.盐胁迫下,植物细胞中的ROS过多会抑制mETC[63].mETC包含两种途径.第一种途径是以细胞色素氧化酶(Cytochrome oxidase, COX)作为末端氧化酶的细胞色素途径(Cytochrome pathway,CP).CP是产生跨膜质子梯度的主要电子传递途径[64],制造了植物体中95%的能量;CP降低会导致mETC的过分还原,造成ROS产生和ATP降低[65].第二种途径是以交替氧化酶(Alternative oxidase,AOX)作为末端氧化酶的交替途径(Alternative pathway,AP)[13].AP是植物特有的耗能电子传递途径,能加强底物水平磷酸化,部分弥补CP下降造成的ATP不足,可缓解ROS对植物的伤害[65]. ...
... [65]. ...
... 在盐逆境中产生的渗透胁迫和离子胁迫的相互作用下,植物细胞器中ROS产生与清除间的动态平衡被破坏,过量ROS对植物造成氧化胁迫[66],破坏了细胞膜结构的完整性和稳定性,导致RC的主要途径——CP受抑制和COX活性降低,ATP合成迅速减少[13,67].同时,ROS作为触发植物胁迫防御途径和适应性反应的关键信号[62],激活AP和AOX相关基因[60].AP的激活加强了底物水平磷酸化,调节了细胞能量代谢平衡,部分弥补了CP受抑制后造成的ATP供应不足状况,有利于降低线粒体中ROS含量,使TCA循环能够继续为新陈代谢提高碳价和能量[65,68].因此,AP的激活有利于维持细胞线粒体与叶绿体间、植物生长与抵抗盐胁迫间的能量代谢平衡,建立起光合作用和呼吸作用间的联系[13,67,69].然而,当胁迫程度不断增加后,ROS含量持续增多也会对AOX带来不可逆损伤,破坏ROS对AP的诱导与AP降低ROS生成之间的平衡状态,不利于胁迫条件下的能量供应[60]. ...
Plant uncoupling mitochondrial protein and alternative oxidase:energy metabolism and stress
1
2005
... 在盐逆境中产生的渗透胁迫和离子胁迫的相互作用下,植物细胞器中ROS产生与清除间的动态平衡被破坏,过量ROS对植物造成氧化胁迫[66],破坏了细胞膜结构的完整性和稳定性,导致RC的主要途径——CP受抑制和COX活性降低,ATP合成迅速减少[13,67].同时,ROS作为触发植物胁迫防御途径和适应性反应的关键信号[62],激活AP和AOX相关基因[60].AP的激活加强了底物水平磷酸化,调节了细胞能量代谢平衡,部分弥补了CP受抑制后造成的ATP供应不足状况,有利于降低线粒体中ROS含量,使TCA循环能够继续为新陈代谢提高碳价和能量[65,68].因此,AP的激活有利于维持细胞线粒体与叶绿体间、植物生长与抵抗盐胁迫间的能量代谢平衡,建立起光合作用和呼吸作用间的联系[13,67,69].然而,当胁迫程度不断增加后,ROS含量持续增多也会对AOX带来不可逆损伤,破坏ROS对AP的诱导与AP降低ROS生成之间的平衡状态,不利于胁迫条件下的能量供应[60]. ...
环境胁迫与植物抗氰呼吸
2
2003
... 在盐逆境中产生的渗透胁迫和离子胁迫的相互作用下,植物细胞器中ROS产生与清除间的动态平衡被破坏,过量ROS对植物造成氧化胁迫[66],破坏了细胞膜结构的完整性和稳定性,导致RC的主要途径——CP受抑制和COX活性降低,ATP合成迅速减少[13,67].同时,ROS作为触发植物胁迫防御途径和适应性反应的关键信号[62],激活AP和AOX相关基因[60].AP的激活加强了底物水平磷酸化,调节了细胞能量代谢平衡,部分弥补了CP受抑制后造成的ATP供应不足状况,有利于降低线粒体中ROS含量,使TCA循环能够继续为新陈代谢提高碳价和能量[65,68].因此,AP的激活有利于维持细胞线粒体与叶绿体间、植物生长与抵抗盐胁迫间的能量代谢平衡,建立起光合作用和呼吸作用间的联系[13,67,69].然而,当胁迫程度不断增加后,ROS含量持续增多也会对AOX带来不可逆损伤,破坏ROS对AP的诱导与AP降低ROS生成之间的平衡状态,不利于胁迫条件下的能量供应[60]. ...
... ,67,69].然而,当胁迫程度不断增加后,ROS含量持续增多也会对AOX带来不可逆损伤,破坏ROS对AP的诱导与AP降低ROS生成之间的平衡状态,不利于胁迫条件下的能量供应[60]. ...
Involvement of ethylene and hydrogen peroxide in induction of alternative respiratory pathway in salt-treated Arabidopsis Calluses
1
2010
... 在盐逆境中产生的渗透胁迫和离子胁迫的相互作用下,植物细胞器中ROS产生与清除间的动态平衡被破坏,过量ROS对植物造成氧化胁迫[66],破坏了细胞膜结构的完整性和稳定性,导致RC的主要途径——CP受抑制和COX活性降低,ATP合成迅速减少[13,67].同时,ROS作为触发植物胁迫防御途径和适应性反应的关键信号[62],激活AP和AOX相关基因[60].AP的激活加强了底物水平磷酸化,调节了细胞能量代谢平衡,部分弥补了CP受抑制后造成的ATP供应不足状况,有利于降低线粒体中ROS含量,使TCA循环能够继续为新陈代谢提高碳价和能量[65,68].因此,AP的激活有利于维持细胞线粒体与叶绿体间、植物生长与抵抗盐胁迫间的能量代谢平衡,建立起光合作用和呼吸作用间的联系[13,67,69].然而,当胁迫程度不断增加后,ROS含量持续增多也会对AOX带来不可逆损伤,破坏ROS对AP的诱导与AP降低ROS生成之间的平衡状态,不利于胁迫条件下的能量供应[60]. ...
外源钙对巴蜀报春抗氰呼吸调节机理的研究
1
2019
... 在盐逆境中产生的渗透胁迫和离子胁迫的相互作用下,植物细胞器中ROS产生与清除间的动态平衡被破坏,过量ROS对植物造成氧化胁迫[66],破坏了细胞膜结构的完整性和稳定性,导致RC的主要途径——CP受抑制和COX活性降低,ATP合成迅速减少[13,67].同时,ROS作为触发植物胁迫防御途径和适应性反应的关键信号[62],激活AP和AOX相关基因[60].AP的激活加强了底物水平磷酸化,调节了细胞能量代谢平衡,部分弥补了CP受抑制后造成的ATP供应不足状况,有利于降低线粒体中ROS含量,使TCA循环能够继续为新陈代谢提高碳价和能量[65,68].因此,AP的激活有利于维持细胞线粒体与叶绿体间、植物生长与抵抗盐胁迫间的能量代谢平衡,建立起光合作用和呼吸作用间的联系[13,67,69].然而,当胁迫程度不断增加后,ROS含量持续增多也会对AOX带来不可逆损伤,破坏ROS对AP的诱导与AP降低ROS生成之间的平衡状态,不利于胁迫条件下的能量供应[60]. ...
iTRAQ-based quantitative proteomic analysis of salt stress in Spica Prunellae
1
2019
... 光合作用是植物生长的基础和新陈代谢的重要途径[54],也是植物吸收、固定和积累能量的重要过程[27,70].在正常条件下,植物将光合作用获得的能量大部分用于植物生长和维持各种生命活动的进行[11].在逆境胁迫下,当植物光合作用积累的能量高于生长和抵御胁迫所消耗的能量时,植物才能成功生长与繁殖[71]. ...
红树植物生活史过程的能量利用策略
1
2021
... 光合作用是植物生长的基础和新陈代谢的重要途径[54],也是植物吸收、固定和积累能量的重要过程[27,70].在正常条件下,植物将光合作用获得的能量大部分用于植物生长和维持各种生命活动的进行[11].在逆境胁迫下,当植物光合作用积累的能量高于生长和抵御胁迫所消耗的能量时,植物才能成功生长与繁殖[71]. ...
Up-regulation of the mitochondrial alternative oxidase pathway enhances photosynthetic electron transport under drought conditions
3
2005
... 盐胁迫下,①水分亏缺致使植物气孔关闭,限制了CO2向叶绿体的扩散,导致CO2同化减少,植物正常物质积累过程受阻,光合产物输出缓慢;②叶绿体中离子失衡导致光反应和暗反应速率降低,光合作用基因和核心能量代谢过程下调,降低了光合作用中能量的产生;③ROS产生过多,对光合系统和膜系统造成氧化损伤,致使光系统I(PSI)和光系统II(PSII)核心复合物降解,最终导致光合速率降低和能量代谢过程受到抑制[13,54,72-73].对沙枣光合作用相关基因进行转录组分析发现,盐胁迫下8个定位于光捕获蛋白的基因表达和8个定位于光合途径的基因表达均被下调,可能会抑制光能的吸收、转移、分布与电子传递,最终导致沙枣体内能量减少[74].可见,盐胁迫下的植物需要更多的周转蛋白对光合系统进行修复和保持叶绿体内离子平衡,使得植物体内能量供应增加[13].然而,在干旱区,当植物处于干旱胁迫时添加一定的盐分反而能够有效减弱干旱对植物光合系统造成的损伤和降低生物量建成成本[39].因此,旱盐胁迫下,盐分对光合系统的影响以及盐分在植物能量代谢过程所起的作用值得今后深入研究. ...
... 叶绿体是植物进行光合作用的主要场所,糖、脂肪和蛋白质是光合作用产生的主要有机物质,它们最终通过mETC转运电子生成ATP[61,75].mETC的两种途径在调节植物光合作用的过程中发挥着重要作用[72].正常生长条件下,AP途径和AOX对光合效率无显著影响.然而,盐胁迫导致线粒体AOX通路被上调,可以防止叶绿体中过量还原等价物的积累,避免PSII过还原,保护光合机构免受过量能量的伤害,维持光合机构功能的正常运行[72,76].尽管AOX对逆境条件下植物的光合作用很重要,但目前对盐胁迫下CP和AP途径的变化以及AOX参与调控植物抗逆的相关机制探索仍然较少. ...
... [72,76].尽管AOX对逆境条件下植物的光合作用很重要,但目前对盐胁迫下CP和AP途径的变化以及AOX参与调控植物抗逆的相关机制探索仍然较少. ...
Strategies to mitigate the adverse effect of drought stress on crop plants-influences of soil bacteria:a review
1
2021
... 盐胁迫下,①水分亏缺致使植物气孔关闭,限制了CO2向叶绿体的扩散,导致CO2同化减少,植物正常物质积累过程受阻,光合产物输出缓慢;②叶绿体中离子失衡导致光反应和暗反应速率降低,光合作用基因和核心能量代谢过程下调,降低了光合作用中能量的产生;③ROS产生过多,对光合系统和膜系统造成氧化损伤,致使光系统I(PSI)和光系统II(PSII)核心复合物降解,最终导致光合速率降低和能量代谢过程受到抑制[13,54,72-73].对沙枣光合作用相关基因进行转录组分析发现,盐胁迫下8个定位于光捕获蛋白的基因表达和8个定位于光合途径的基因表达均被下调,可能会抑制光能的吸收、转移、分布与电子传递,最终导致沙枣体内能量减少[74].可见,盐胁迫下的植物需要更多的周转蛋白对光合系统进行修复和保持叶绿体内离子平衡,使得植物体内能量供应增加[13].然而,在干旱区,当植物处于干旱胁迫时添加一定的盐分反而能够有效减弱干旱对植物光合系统造成的损伤和降低生物量建成成本[39].因此,旱盐胁迫下,盐分对光合系统的影响以及盐分在植物能量代谢过程所起的作用值得今后深入研究. ...
Transcriptome profiling of genes involved in photosynthesis in Elaeagnus angustifolia L. under salt stress
1
2018
... 盐胁迫下,①水分亏缺致使植物气孔关闭,限制了CO2向叶绿体的扩散,导致CO2同化减少,植物正常物质积累过程受阻,光合产物输出缓慢;②叶绿体中离子失衡导致光反应和暗反应速率降低,光合作用基因和核心能量代谢过程下调,降低了光合作用中能量的产生;③ROS产生过多,对光合系统和膜系统造成氧化损伤,致使光系统I(PSI)和光系统II(PSII)核心复合物降解,最终导致光合速率降低和能量代谢过程受到抑制[13,54,72-73].对沙枣光合作用相关基因进行转录组分析发现,盐胁迫下8个定位于光捕获蛋白的基因表达和8个定位于光合途径的基因表达均被下调,可能会抑制光能的吸收、转移、分布与电子传递,最终导致沙枣体内能量减少[74].可见,盐胁迫下的植物需要更多的周转蛋白对光合系统进行修复和保持叶绿体内离子平衡,使得植物体内能量供应增加[13].然而,在干旱区,当植物处于干旱胁迫时添加一定的盐分反而能够有效减弱干旱对植物光合系统造成的损伤和降低生物量建成成本[39].因此,旱盐胁迫下,盐分对光合系统的影响以及盐分在植物能量代谢过程所起的作用值得今后深入研究. ...
交替呼吸途径在油菜素内酯调控本氏烟响应高温胁迫中的作用研究
1
2020
... 叶绿体是植物进行光合作用的主要场所,糖、脂肪和蛋白质是光合作用产生的主要有机物质,它们最终通过mETC转运电子生成ATP[61,75].mETC的两种途径在调节植物光合作用的过程中发挥着重要作用[72].正常生长条件下,AP途径和AOX对光合效率无显著影响.然而,盐胁迫导致线粒体AOX通路被上调,可以防止叶绿体中过量还原等价物的积累,避免PSII过还原,保护光合机构免受过量能量的伤害,维持光合机构功能的正常运行[72,76].尽管AOX对逆境条件下植物的光合作用很重要,但目前对盐胁迫下CP和AP途径的变化以及AOX参与调控植物抗逆的相关机制探索仍然较少. ...
交替途径及外源过氧化氢在唐古特白刺耐盐性中的作用
1
2018
... 叶绿体是植物进行光合作用的主要场所,糖、脂肪和蛋白质是光合作用产生的主要有机物质,它们最终通过mETC转运电子生成ATP[61,75].mETC的两种途径在调节植物光合作用的过程中发挥着重要作用[72].正常生长条件下,AP途径和AOX对光合效率无显著影响.然而,盐胁迫导致线粒体AOX通路被上调,可以防止叶绿体中过量还原等价物的积累,避免PSII过还原,保护光合机构免受过量能量的伤害,维持光合机构功能的正常运行[72,76].尽管AOX对逆境条件下植物的光合作用很重要,但目前对盐胁迫下CP和AP途径的变化以及AOX参与调控植物抗逆的相关机制探索仍然较少. ...
Salinity tolerance of crops-what is the cost?
2
2015
... 盐胁迫下,叶片同化物输出下降,导致植物用于营养生长和生殖生长的能量供应减少,用于抵御盐胁迫的能量消耗增多,为同化物周转提供的能量不足,致使同化物周转速率降低[55,77].这不仅抑制植物对营养物质的吸收,而且影响植物生殖生理和种子成熟过程,最终影响作物产量和品质[55,78].土壤盐分的升高通过抑制小孢子发生、延长雄蕊花丝、加速细胞程序性死亡、促进受精胚胎衰老和胚珠败育等各个方面影响生殖发育[79],最终导致作物产量降低、营养含量不足和籽粒品质不佳[80]. ...
... 然而,也有研究发现,在植物特定生长阶段施加适度胁迫有利于促进作物生长、提高作物产量和改善籽粒品质[80-81].碳水化合物是植物进行光合作用的主要产物,可分为参与植物结构形态构建的结构性碳水化合物和影响植物生长代谢过程中能量供应的非结构性碳水化合物(Non-structural carbohydrate,NSC)[82].适度干旱胁迫诱导NSC合成与转运相关基因的表达,促进NSC的积累,提高NSC转运量、转运率和表现贡献率,从而为植物生长提供能量,进而提高植物抗性、产量和品质[83-85].因此,充分认识干旱区植物能量需求和植物抵御胁迫的能量消耗,提高胁迫条件下植物代谢和生理过程中能量利用效率、增加外界能量物质的输入和促进生殖生长阶段物质能量从“源”到“库”的运输,是提高产量和品质的重要途径[12,77,86]. ...
植物根际促生菌诱导植物耐盐促生作用机制
1
2020
... 盐胁迫下,叶片同化物输出下降,导致植物用于营养生长和生殖生长的能量供应减少,用于抵御盐胁迫的能量消耗增多,为同化物周转提供的能量不足,致使同化物周转速率降低[55,77].这不仅抑制植物对营养物质的吸收,而且影响植物生殖生理和种子成熟过程,最终影响作物产量和品质[55,78].土壤盐分的升高通过抑制小孢子发生、延长雄蕊花丝、加速细胞程序性死亡、促进受精胚胎衰老和胚珠败育等各个方面影响生殖发育[79],最终导致作物产量降低、营养含量不足和籽粒品质不佳[80]. ...
植物对盐碱胁迫的适应机理研究进展
1
2020
... 盐胁迫下,叶片同化物输出下降,导致植物用于营养生长和生殖生长的能量供应减少,用于抵御盐胁迫的能量消耗增多,为同化物周转提供的能量不足,致使同化物周转速率降低[55,77].这不仅抑制植物对营养物质的吸收,而且影响植物生殖生理和种子成熟过程,最终影响作物产量和品质[55,78].土壤盐分的升高通过抑制小孢子发生、延长雄蕊花丝、加速细胞程序性死亡、促进受精胚胎衰老和胚珠败育等各个方面影响生殖发育[79],最终导致作物产量降低、营养含量不足和籽粒品质不佳[80]. ...
不同耐盐高粱品种全生育期对盐胁迫的响应
2
2020
... 盐胁迫下,叶片同化物输出下降,导致植物用于营养生长和生殖生长的能量供应减少,用于抵御盐胁迫的能量消耗增多,为同化物周转提供的能量不足,致使同化物周转速率降低[55,77].这不仅抑制植物对营养物质的吸收,而且影响植物生殖生理和种子成熟过程,最终影响作物产量和品质[55,78].土壤盐分的升高通过抑制小孢子发生、延长雄蕊花丝、加速细胞程序性死亡、促进受精胚胎衰老和胚珠败育等各个方面影响生殖发育[79],最终导致作物产量降低、营养含量不足和籽粒品质不佳[80]. ...
... 然而,也有研究发现,在植物特定生长阶段施加适度胁迫有利于促进作物生长、提高作物产量和改善籽粒品质[80-81].碳水化合物是植物进行光合作用的主要产物,可分为参与植物结构形态构建的结构性碳水化合物和影响植物生长代谢过程中能量供应的非结构性碳水化合物(Non-structural carbohydrate,NSC)[82].适度干旱胁迫诱导NSC合成与转运相关基因的表达,促进NSC的积累,提高NSC转运量、转运率和表现贡献率,从而为植物生长提供能量,进而提高植物抗性、产量和品质[83-85].因此,充分认识干旱区植物能量需求和植物抵御胁迫的能量消耗,提高胁迫条件下植物代谢和生理过程中能量利用效率、增加外界能量物质的输入和促进生殖生长阶段物质能量从“源”到“库”的运输,是提高产量和品质的重要途径[12,77,86]. ...
粮食作物对高温干旱胁迫的响应及其阈值研究进展与展望
1
2016
... 然而,也有研究发现,在植物特定生长阶段施加适度胁迫有利于促进作物生长、提高作物产量和改善籽粒品质[80-81].碳水化合物是植物进行光合作用的主要产物,可分为参与植物结构形态构建的结构性碳水化合物和影响植物生长代谢过程中能量供应的非结构性碳水化合物(Non-structural carbohydrate,NSC)[82].适度干旱胁迫诱导NSC合成与转运相关基因的表达,促进NSC的积累,提高NSC转运量、转运率和表现贡献率,从而为植物生长提供能量,进而提高植物抗性、产量和品质[83-85].因此,充分认识干旱区植物能量需求和植物抵御胁迫的能量消耗,提高胁迫条件下植物代谢和生理过程中能量利用效率、增加外界能量物质的输入和促进生殖生长阶段物质能量从“源”到“库”的运输,是提高产量和品质的重要途径[12,77,86]. ...
氮磷添加对黑果枸杞(Lycium ruthenicum)营养器官非结构性碳水化合物特征的影响
1
2021
... 然而,也有研究发现,在植物特定生长阶段施加适度胁迫有利于促进作物生长、提高作物产量和改善籽粒品质[80-81].碳水化合物是植物进行光合作用的主要产物,可分为参与植物结构形态构建的结构性碳水化合物和影响植物生长代谢过程中能量供应的非结构性碳水化合物(Non-structural carbohydrate,NSC)[82].适度干旱胁迫诱导NSC合成与转运相关基因的表达,促进NSC的积累,提高NSC转运量、转运率和表现贡献率,从而为植物生长提供能量,进而提高植物抗性、产量和品质[83-85].因此,充分认识干旱区植物能量需求和植物抵御胁迫的能量消耗,提高胁迫条件下植物代谢和生理过程中能量利用效率、增加外界能量物质的输入和促进生殖生长阶段物质能量从“源”到“库”的运输,是提高产量和品质的重要途径[12,77,86]. ...
果树中糖类代谢和调控研究
1
2020
... 然而,也有研究发现,在植物特定生长阶段施加适度胁迫有利于促进作物生长、提高作物产量和改善籽粒品质[80-81].碳水化合物是植物进行光合作用的主要产物,可分为参与植物结构形态构建的结构性碳水化合物和影响植物生长代谢过程中能量供应的非结构性碳水化合物(Non-structural carbohydrate,NSC)[82].适度干旱胁迫诱导NSC合成与转运相关基因的表达,促进NSC的积累,提高NSC转运量、转运率和表现贡献率,从而为植物生长提供能量,进而提高植物抗性、产量和品质[83-85].因此,充分认识干旱区植物能量需求和植物抵御胁迫的能量消耗,提高胁迫条件下植物代谢和生理过程中能量利用效率、增加外界能量物质的输入和促进生殖生长阶段物质能量从“源”到“库”的运输,是提高产量和品质的重要途径[12,77,86]. ...
促进稻麦同化物转运和籽粒灌浆的途径与机制
0
2018
水稻茎鞘非结构性碳水化合物转运机理及栽培调控研究进展
1
2021
... 然而,也有研究发现,在植物特定生长阶段施加适度胁迫有利于促进作物生长、提高作物产量和改善籽粒品质[80-81].碳水化合物是植物进行光合作用的主要产物,可分为参与植物结构形态构建的结构性碳水化合物和影响植物生长代谢过程中能量供应的非结构性碳水化合物(Non-structural carbohydrate,NSC)[82].适度干旱胁迫诱导NSC合成与转运相关基因的表达,促进NSC的积累,提高NSC转运量、转运率和表现贡献率,从而为植物生长提供能量,进而提高植物抗性、产量和品质[83-85].因此,充分认识干旱区植物能量需求和植物抵御胁迫的能量消耗,提高胁迫条件下植物代谢和生理过程中能量利用效率、增加外界能量物质的输入和促进生殖生长阶段物质能量从“源”到“库”的运输,是提高产量和品质的重要途径[12,77,86]. ...
Metabolic energy cost for plant cells exposed to salinity
1
1984
... 然而,也有研究发现,在植物特定生长阶段施加适度胁迫有利于促进作物生长、提高作物产量和改善籽粒品质[80-81].碳水化合物是植物进行光合作用的主要产物,可分为参与植物结构形态构建的结构性碳水化合物和影响植物生长代谢过程中能量供应的非结构性碳水化合物(Non-structural carbohydrate,NSC)[82].适度干旱胁迫诱导NSC合成与转运相关基因的表达,促进NSC的积累,提高NSC转运量、转运率和表现贡献率,从而为植物生长提供能量,进而提高植物抗性、产量和品质[83-85].因此,充分认识干旱区植物能量需求和植物抵御胁迫的能量消耗,提高胁迫条件下植物代谢和生理过程中能量利用效率、增加外界能量物质的输入和促进生殖生长阶段物质能量从“源”到“库”的运输,是提高产量和品质的重要途径[12,77,86]. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}