Urbanization contributes little to global warming but substantially intensifies local and regional land surface warming
1
2022
... 全球变暖对陆地生态系统结构和功能产生深刻影响,从而降低生态系统服务功能[1 ] .截至2022年,全球平均气温比工业革命前升高了1.09 ℃,预估到2100年或升温1.8~4.0 ℃[2 -3 ] .陆地生态系统的群落结构和功能深受全球气候变暖的影响,因而备受关注[4 -5 ] .温度升高通过影响植物光合、呼吸等生理过程及土壤环境和营养物质的可利用性,进而影响植物的正常生长、生物量生产与分配[6 -7 ] . ...
A review of atmospheric changes amidst COVID19 pandemic in the light of IPCC special report 2018 and COP24 conference
1
2020
... 全球变暖对陆地生态系统结构和功能产生深刻影响,从而降低生态系统服务功能[1 ] .截至2022年,全球平均气温比工业革命前升高了1.09 ℃,预估到2100年或升温1.8~4.0 ℃[2 -3 ] .陆地生态系统的群落结构和功能深受全球气候变暖的影响,因而备受关注[4 -5 ] .温度升高通过影响植物光合、呼吸等生理过程及土壤环境和营养物质的可利用性,进而影响植物的正常生长、生物量生产与分配[6 -7 ] . ...
Transient response of the global mean warming rate and its spatial variation
1
2017
... 全球变暖对陆地生态系统结构和功能产生深刻影响,从而降低生态系统服务功能[1 ] .截至2022年,全球平均气温比工业革命前升高了1.09 ℃,预估到2100年或升温1.8~4.0 ℃[2 -3 ] .陆地生态系统的群落结构和功能深受全球气候变暖的影响,因而备受关注[4 -5 ] .温度升高通过影响植物光合、呼吸等生理过程及土壤环境和营养物质的可利用性,进而影响植物的正常生长、生物量生产与分配[6 -7 ] . ...
The links between ecosystem multifunctionality and above-and belowground biodiversity are mediated by climate
1
2015
... 全球变暖对陆地生态系统结构和功能产生深刻影响,从而降低生态系统服务功能[1 ] .截至2022年,全球平均气温比工业革命前升高了1.09 ℃,预估到2100年或升温1.8~4.0 ℃[2 -3 ] .陆地生态系统的群落结构和功能深受全球气候变暖的影响,因而备受关注[4 -5 ] .温度升高通过影响植物光合、呼吸等生理过程及土壤环境和营养物质的可利用性,进而影响植物的正常生长、生物量生产与分配[6 -7 ] . ...
Beyond the ivory tower:the scientific consensus on climate change
1
2004
... 全球变暖对陆地生态系统结构和功能产生深刻影响,从而降低生态系统服务功能[1 ] .截至2022年,全球平均气温比工业革命前升高了1.09 ℃,预估到2100年或升温1.8~4.0 ℃[2 -3 ] .陆地生态系统的群落结构和功能深受全球气候变暖的影响,因而备受关注[4 -5 ] .温度升高通过影响植物光合、呼吸等生理过程及土壤环境和营养物质的可利用性,进而影响植物的正常生长、生物量生产与分配[6 -7 ] . ...
Differential response of alpine steppe and alpine meadow to climate warming in the central Qinghai-Tibetan Plateau
2
2016
... 全球变暖对陆地生态系统结构和功能产生深刻影响,从而降低生态系统服务功能[1 ] .截至2022年,全球平均气温比工业革命前升高了1.09 ℃,预估到2100年或升温1.8~4.0 ℃[2 -3 ] .陆地生态系统的群落结构和功能深受全球气候变暖的影响,因而备受关注[4 -5 ] .温度升高通过影响植物光合、呼吸等生理过程及土壤环境和营养物质的可利用性,进而影响植物的正常生长、生物量生产与分配[6 -7 ] . ...
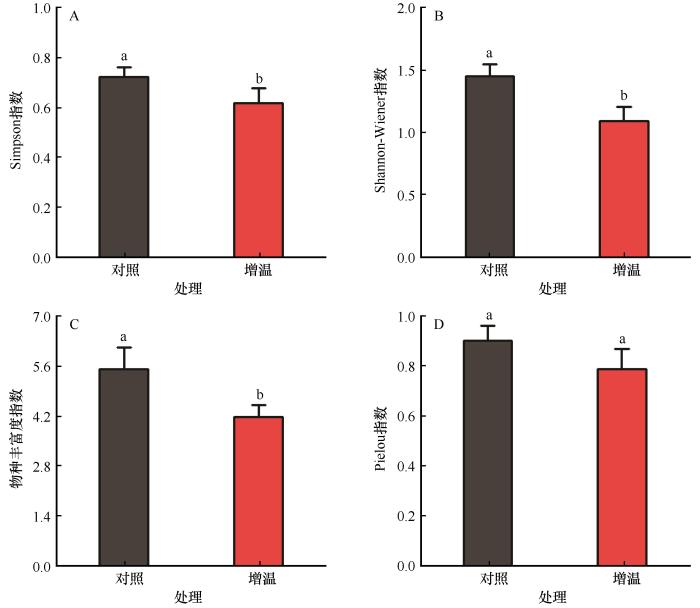
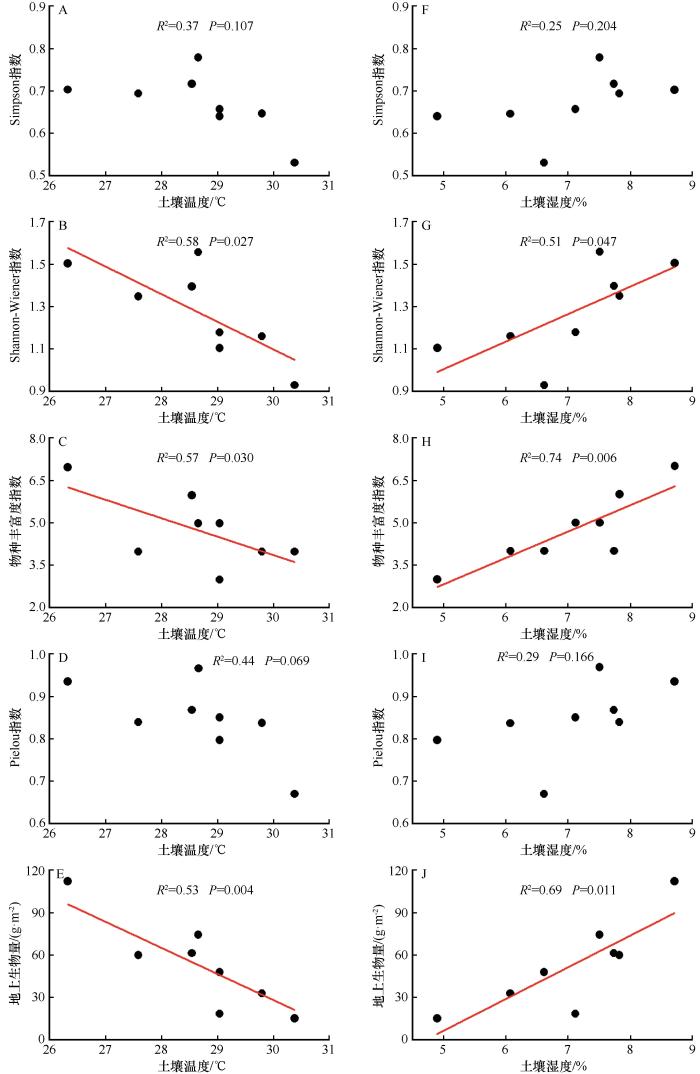
... 科尔沁沙质草地生态系统脆弱,长期放牧及土壤水分匮缺导致沙质草地不断退化.通过围封禁牧及人工修复会逐步恢复沙质草地,但气候的异常变化会影响沙质草地生态系统结构及其功能.降水的增加或减少会引起草地生态系统植物群落结构和功能发生明显改变[18 ,30 -31 ] .温度也是影响植物正常生长的重要因子,尤其在干旱半干旱区,植物除了对养分吸收的竞争加剧外,还存在对水分吸收的竞争[32 ] .本研究结果表明,沙质草地在模拟增温后草本群落组成发生明显变化,群落物种(科)数低于沙质草地(表2 ).以禾本科为主的优势种占比明显减少,这与Ganjurjav等[6 ] 的研究结果相似.禾本科植物占比减少,为杂草类植物提供了更大的生存空间.此外,狗尾草、糙隐子草、黄蒿和大籽蒿解释了模拟增温导致群落组成差异的63%,且以一年生草本狗尾草为主.吕朋等[33 ] 在研究沙质草地群落特征中发现,一年生草本的禾本科狗尾草是优势物种,这与本研究结果一致,但模拟增温后的狗尾草重要值显著降低,表明狗尾草等一年生草本对模拟增温后的土壤环境较敏感.在增温和水分胁迫的环境中,土壤养分因此受到的负面影响导致群落组成发生改变[34 ] .模拟增温后沙质草地群落多样性呈下降趋势,其中Simpson指数、Shannon-Wiener指数和物种丰富度指数显著下降,而Pielou指数下降趋势不显著(图3 ),这与之前的大部分研究结果一致[35 -37 ] .也有学者认为低幅增温对多样性产生促进作用,但当增温超过一定幅度时,促进作用会减弱甚至变成抑制作用[12 ] ,造成这种不一致的原因可能是物种组成以及研究区间的差异.郭轶瑞等[38 ] 的研究发现耐旱植物对增温的抵抗力较强,而喜湿植物在受水分限制时竞争能力减弱导致其比重降低.增温导致土壤含水量降低,同时,物种丰富度和多样性指数与土壤湿度呈显著正相关关系,表明在干旱半干旱区,多样性随水分胁迫强度的增加更加敏感. ...
荒漠草原物种多样性、生产力与气候因子和土壤养分之间关系的研究
6
2015
... 全球变暖对陆地生态系统结构和功能产生深刻影响,从而降低生态系统服务功能[1 ] .截至2022年,全球平均气温比工业革命前升高了1.09 ℃,预估到2100年或升温1.8~4.0 ℃[2 -3 ] .陆地生态系统的群落结构和功能深受全球气候变暖的影响,因而备受关注[4 -5 ] .温度升高通过影响植物光合、呼吸等生理过程及土壤环境和营养物质的可利用性,进而影响植物的正常生长、生物量生产与分配[6 -7 ] . ...
... 草地是全球分布面积最大的陆地生态系统,具有重要的生态和社会功能[8 -9 ] .草地生态系统的植物群落对变暖响应极为敏感[10 ] .增温对群落物种多样性具有促进或抑制或无明显作用[7 ,11 -12 ] ,对群落生物量积累的作用因增幅而异,低幅增温具有促进作用,高幅增温抑制积累[12 ] .除了物种组成、优势种、多样性、生产力等指标可以描述植物群落结构及功能之外,群落连接性、组织程度和关键种也是常见的有效指标[13 ] .目前,有关草地植物群落网络结构复杂性和关键种变化的研究多在人为干扰方面[14 ] ,很少涉及气候变化. ...
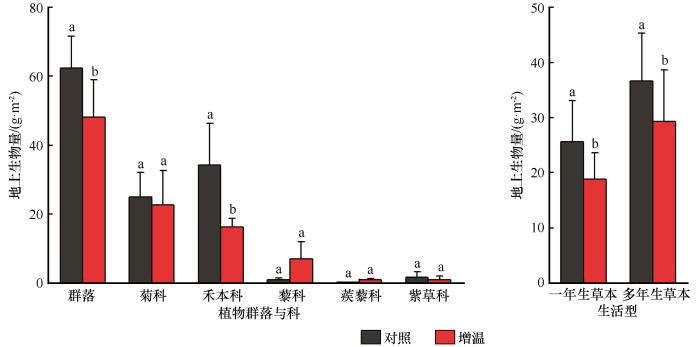
... 多样性与生产力的关系是反映群落结构及生态功能的关键指标[39 -40 ] .许多研究证实物种多样性与生产力主要存在正相关、负相关、单峰曲线和不相关4种关系[7 ] ,且正相关关系的结论居多.就科尔沁沙质草地而言,物种丰富度指数和生物量之间呈显著线性正相关,这与孙小丽等[7 ] 对内蒙古中温型荒漠草原区群落生物量随物种丰富度增加而增加的结论相一致,但与郭轶瑞等[41 ] 在科尔沁沙质草地的物种多样性与生产力呈单峰关系这一结论不同.极有可能是因为独特的生境特征差异.研究区处于干旱半干旱区,水热因子对干旱半干旱区植物群落生产力方面作用占主导地位[7 ,11 ] ,因此,推测科尔沁沙质草地生产力受到土壤水热因子的调控.本研究中,增温模拟后的群落和禾本科地上生物量与对照组相比显著下降(图4 ).Arnda等[42 ] 发现,禾本科植物地上生物量减少可能是因为更多杂草类根系在土壤剖面上的分配.也有研究表明,在干旱半干旱区,生产力深受水分限制[11 ] .Grime等[43 ] 发现,在增温和水分胁迫的环境中,植物通常以减少生物量生产来提高水分利用效率以此适应环境,解释了本研究在模拟增温后土壤湿度的降低导致了植物地上生产力的降低.由于水分限制,模拟增温后植物无法利用土壤温度升高的能量进行光合作用,从而呈随着能量增加群落多样性和生产力降低的关系.也有研究表明,增温对生物量的潜在影响不仅与土壤水热条件有关,土壤养分与多样性和生物量也存在显著相关的关系[34 ] .孙小丽等[7 ] 对草地生态系统的研究表明生物量与速效氮、有机碳等土壤养分呈显著的线性正相关.生物量分配还受到群落组成的影响,禾本科地上生物量减少为杂草类地下生物量提供了更多的水分和养分资源[42 ] . ...
... [7 ]对内蒙古中温型荒漠草原区群落生物量随物种丰富度增加而增加的结论相一致,但与郭轶瑞等[41 ] 在科尔沁沙质草地的物种多样性与生产力呈单峰关系这一结论不同.极有可能是因为独特的生境特征差异.研究区处于干旱半干旱区,水热因子对干旱半干旱区植物群落生产力方面作用占主导地位[7 ,11 ] ,因此,推测科尔沁沙质草地生产力受到土壤水热因子的调控.本研究中,增温模拟后的群落和禾本科地上生物量与对照组相比显著下降(图4 ).Arnda等[42 ] 发现,禾本科植物地上生物量减少可能是因为更多杂草类根系在土壤剖面上的分配.也有研究表明,在干旱半干旱区,生产力深受水分限制[11 ] .Grime等[43 ] 发现,在增温和水分胁迫的环境中,植物通常以减少生物量生产来提高水分利用效率以此适应环境,解释了本研究在模拟增温后土壤湿度的降低导致了植物地上生产力的降低.由于水分限制,模拟增温后植物无法利用土壤温度升高的能量进行光合作用,从而呈随着能量增加群落多样性和生产力降低的关系.也有研究表明,增温对生物量的潜在影响不仅与土壤水热条件有关,土壤养分与多样性和生物量也存在显著相关的关系[34 ] .孙小丽等[7 ] 对草地生态系统的研究表明生物量与速效氮、有机碳等土壤养分呈显著的线性正相关.生物量分配还受到群落组成的影响,禾本科地上生物量减少为杂草类地下生物量提供了更多的水分和养分资源[42 ] . ...
... [7 ,11 ],因此,推测科尔沁沙质草地生产力受到土壤水热因子的调控.本研究中,增温模拟后的群落和禾本科地上生物量与对照组相比显著下降(图4 ).Arnda等[42 ] 发现,禾本科植物地上生物量减少可能是因为更多杂草类根系在土壤剖面上的分配.也有研究表明,在干旱半干旱区,生产力深受水分限制[11 ] .Grime等[43 ] 发现,在增温和水分胁迫的环境中,植物通常以减少生物量生产来提高水分利用效率以此适应环境,解释了本研究在模拟增温后土壤湿度的降低导致了植物地上生产力的降低.由于水分限制,模拟增温后植物无法利用土壤温度升高的能量进行光合作用,从而呈随着能量增加群落多样性和生产力降低的关系.也有研究表明,增温对生物量的潜在影响不仅与土壤水热条件有关,土壤养分与多样性和生物量也存在显著相关的关系[34 ] .孙小丽等[7 ] 对草地生态系统的研究表明生物量与速效氮、有机碳等土壤养分呈显著的线性正相关.生物量分配还受到群落组成的影响,禾本科地上生物量减少为杂草类地下生物量提供了更多的水分和养分资源[42 ] . ...
... [7 ]对草地生态系统的研究表明生物量与速效氮、有机碳等土壤养分呈显著的线性正相关.生物量分配还受到群落组成的影响,禾本科地上生物量减少为杂草类地下生物量提供了更多的水分和养分资源[42 ] . ...
中国北方草地生态系统服务评估
1
2023
... 草地是全球分布面积最大的陆地生态系统,具有重要的生态和社会功能[8 -9 ] .草地生态系统的植物群落对变暖响应极为敏感[10 ] .增温对群落物种多样性具有促进或抑制或无明显作用[7 ,11 -12 ] ,对群落生物量积累的作用因增幅而异,低幅增温具有促进作用,高幅增温抑制积累[12 ] .除了物种组成、优势种、多样性、生产力等指标可以描述植物群落结构及功能之外,群落连接性、组织程度和关键种也是常见的有效指标[13 ] .目前,有关草地植物群落网络结构复杂性和关键种变化的研究多在人为干扰方面[14 ] ,很少涉及气候变化. ...
非生长季放牧对黄河上游草甸种子产量的影响
1
2022
... 草地是全球分布面积最大的陆地生态系统,具有重要的生态和社会功能[8 -9 ] .草地生态系统的植物群落对变暖响应极为敏感[10 ] .增温对群落物种多样性具有促进或抑制或无明显作用[7 ,11 -12 ] ,对群落生物量积累的作用因增幅而异,低幅增温具有促进作用,高幅增温抑制积累[12 ] .除了物种组成、优势种、多样性、生产力等指标可以描述植物群落结构及功能之外,群落连接性、组织程度和关键种也是常见的有效指标[13 ] .目前,有关草地植物群落网络结构复杂性和关键种变化的研究多在人为干扰方面[14 ] ,很少涉及气候变化. ...
Climate warming reduces the temporal stability of plant community biomass production
1
2017
... 草地是全球分布面积最大的陆地生态系统,具有重要的生态和社会功能[8 -9 ] .草地生态系统的植物群落对变暖响应极为敏感[10 ] .增温对群落物种多样性具有促进或抑制或无明显作用[7 ,11 -12 ] ,对群落生物量积累的作用因增幅而异,低幅增温具有促进作用,高幅增温抑制积累[12 ] .除了物种组成、优势种、多样性、生产力等指标可以描述植物群落结构及功能之外,群落连接性、组织程度和关键种也是常见的有效指标[13 ] .目前,有关草地植物群落网络结构复杂性和关键种变化的研究多在人为干扰方面[14 ] ,很少涉及气候变化. ...
短期增温对青藏高原高寒草甸植物群落结构和生物量的影响
3
2011
... 草地是全球分布面积最大的陆地生态系统,具有重要的生态和社会功能[8 -9 ] .草地生态系统的植物群落对变暖响应极为敏感[10 ] .增温对群落物种多样性具有促进或抑制或无明显作用[7 ,11 -12 ] ,对群落生物量积累的作用因增幅而异,低幅增温具有促进作用,高幅增温抑制积累[12 ] .除了物种组成、优势种、多样性、生产力等指标可以描述植物群落结构及功能之外,群落连接性、组织程度和关键种也是常见的有效指标[13 ] .目前,有关草地植物群落网络结构复杂性和关键种变化的研究多在人为干扰方面[14 ] ,很少涉及气候变化. ...
... 多样性与生产力的关系是反映群落结构及生态功能的关键指标[39 -40 ] .许多研究证实物种多样性与生产力主要存在正相关、负相关、单峰曲线和不相关4种关系[7 ] ,且正相关关系的结论居多.就科尔沁沙质草地而言,物种丰富度指数和生物量之间呈显著线性正相关,这与孙小丽等[7 ] 对内蒙古中温型荒漠草原区群落生物量随物种丰富度增加而增加的结论相一致,但与郭轶瑞等[41 ] 在科尔沁沙质草地的物种多样性与生产力呈单峰关系这一结论不同.极有可能是因为独特的生境特征差异.研究区处于干旱半干旱区,水热因子对干旱半干旱区植物群落生产力方面作用占主导地位[7 ,11 ] ,因此,推测科尔沁沙质草地生产力受到土壤水热因子的调控.本研究中,增温模拟后的群落和禾本科地上生物量与对照组相比显著下降(图4 ).Arnda等[42 ] 发现,禾本科植物地上生物量减少可能是因为更多杂草类根系在土壤剖面上的分配.也有研究表明,在干旱半干旱区,生产力深受水分限制[11 ] .Grime等[43 ] 发现,在增温和水分胁迫的环境中,植物通常以减少生物量生产来提高水分利用效率以此适应环境,解释了本研究在模拟增温后土壤湿度的降低导致了植物地上生产力的降低.由于水分限制,模拟增温后植物无法利用土壤温度升高的能量进行光合作用,从而呈随着能量增加群落多样性和生产力降低的关系.也有研究表明,增温对生物量的潜在影响不仅与土壤水热条件有关,土壤养分与多样性和生物量也存在显著相关的关系[34 ] .孙小丽等[7 ] 对草地生态系统的研究表明生物量与速效氮、有机碳等土壤养分呈显著的线性正相关.生物量分配还受到群落组成的影响,禾本科地上生物量减少为杂草类地下生物量提供了更多的水分和养分资源[42 ] . ...
... [11 ].Grime等[43 ] 发现,在增温和水分胁迫的环境中,植物通常以减少生物量生产来提高水分利用效率以此适应环境,解释了本研究在模拟增温后土壤湿度的降低导致了植物地上生产力的降低.由于水分限制,模拟增温后植物无法利用土壤温度升高的能量进行光合作用,从而呈随着能量增加群落多样性和生产力降低的关系.也有研究表明,增温对生物量的潜在影响不仅与土壤水热条件有关,土壤养分与多样性和生物量也存在显著相关的关系[34 ] .孙小丽等[7 ] 对草地生态系统的研究表明生物量与速效氮、有机碳等土壤养分呈显著的线性正相关.生物量分配还受到群落组成的影响,禾本科地上生物量减少为杂草类地下生物量提供了更多的水分和养分资源[42 ] . ...
短期增温对藏北高寒草甸植物群落特征的影响
3
2017
... 草地是全球分布面积最大的陆地生态系统,具有重要的生态和社会功能[8 -9 ] .草地生态系统的植物群落对变暖响应极为敏感[10 ] .增温对群落物种多样性具有促进或抑制或无明显作用[7 ,11 -12 ] ,对群落生物量积累的作用因增幅而异,低幅增温具有促进作用,高幅增温抑制积累[12 ] .除了物种组成、优势种、多样性、生产力等指标可以描述植物群落结构及功能之外,群落连接性、组织程度和关键种也是常见的有效指标[13 ] .目前,有关草地植物群落网络结构复杂性和关键种变化的研究多在人为干扰方面[14 ] ,很少涉及气候变化. ...
... [12 ].除了物种组成、优势种、多样性、生产力等指标可以描述植物群落结构及功能之外,群落连接性、组织程度和关键种也是常见的有效指标[13 ] .目前,有关草地植物群落网络结构复杂性和关键种变化的研究多在人为干扰方面[14 ] ,很少涉及气候变化. ...
... 科尔沁沙质草地生态系统脆弱,长期放牧及土壤水分匮缺导致沙质草地不断退化.通过围封禁牧及人工修复会逐步恢复沙质草地,但气候的异常变化会影响沙质草地生态系统结构及其功能.降水的增加或减少会引起草地生态系统植物群落结构和功能发生明显改变[18 ,30 -31 ] .温度也是影响植物正常生长的重要因子,尤其在干旱半干旱区,植物除了对养分吸收的竞争加剧外,还存在对水分吸收的竞争[32 ] .本研究结果表明,沙质草地在模拟增温后草本群落组成发生明显变化,群落物种(科)数低于沙质草地(表2 ).以禾本科为主的优势种占比明显减少,这与Ganjurjav等[6 ] 的研究结果相似.禾本科植物占比减少,为杂草类植物提供了更大的生存空间.此外,狗尾草、糙隐子草、黄蒿和大籽蒿解释了模拟增温导致群落组成差异的63%,且以一年生草本狗尾草为主.吕朋等[33 ] 在研究沙质草地群落特征中发现,一年生草本的禾本科狗尾草是优势物种,这与本研究结果一致,但模拟增温后的狗尾草重要值显著降低,表明狗尾草等一年生草本对模拟增温后的土壤环境较敏感.在增温和水分胁迫的环境中,土壤养分因此受到的负面影响导致群落组成发生改变[34 ] .模拟增温后沙质草地群落多样性呈下降趋势,其中Simpson指数、Shannon-Wiener指数和物种丰富度指数显著下降,而Pielou指数下降趋势不显著(图3 ),这与之前的大部分研究结果一致[35 -37 ] .也有学者认为低幅增温对多样性产生促进作用,但当增温超过一定幅度时,促进作用会减弱甚至变成抑制作用[12 ] ,造成这种不一致的原因可能是物种组成以及研究区间的差异.郭轶瑞等[38 ] 的研究发现耐旱植物对增温的抵抗力较强,而喜湿植物在受水分限制时竞争能力减弱导致其比重降低.增温导致土壤含水量降低,同时,物种丰富度和多样性指数与土壤湿度呈显著正相关关系,表明在干旱半干旱区,多样性随水分胁迫强度的增加更加敏感. ...
Responses of plant community structure to mowing in the degraded alpine meadows,Northwestern Yunnan Province
4
2022
... 草地是全球分布面积最大的陆地生态系统,具有重要的生态和社会功能[8 -9 ] .草地生态系统的植物群落对变暖响应极为敏感[10 ] .增温对群落物种多样性具有促进或抑制或无明显作用[7 ,11 -12 ] ,对群落生物量积累的作用因增幅而异,低幅增温具有促进作用,高幅增温抑制积累[12 ] .除了物种组成、优势种、多样性、生产力等指标可以描述植物群落结构及功能之外,群落连接性、组织程度和关键种也是常见的有效指标[13 ] .目前,有关草地植物群落网络结构复杂性和关键种变化的研究多在人为干扰方面[14 ] ,很少涉及气候变化. ...
... 基于物种间的Pearson相关系数进行共生网络分析,分析过程中仅保留相关性系数达到显著性(P <0.05)的物种[14 ] ,以此探究沙地植物群落在增温处理后的共现模式.利用Gephi 0.10.1进行网络分析可视化绘图,Statistics工具计算代表网络拓扑性质的参数,包括节点数、模块化属性、网络连通性等.在植物群落生态网络中,节点代表群落中的物种,连接代表物种之间的关系[26 -27 ] ;网络节点数、连接数及平均连通度用来指示网络的规模和复杂程度,其中,平均连通度越大表明网络节点之间具有越强的相互作用,群落越复杂[28 ] ;平均聚类系数表明了网络的组织水平,平均聚类系数越高,组织水平越高[14 ] ;连通性指数用来表示群落复杂度,值越大代表群落复杂度越高;模块化指数衡量了网络图结构的模块化程度[13 ] .从群落结构维持的角度出发,通常根据网络中Hub指标的最大值确定关键物种[29 ] . ...
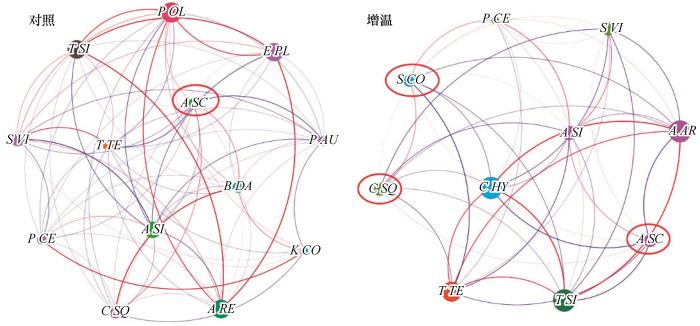
... 增温会导致沙质草地群落种间关系发生变化.除了多样性和生产力可以衡量群落特征以外,群落网络结构的复杂性以及关键类群也可以评价其结构和功能[13 ,44 ] .共生网络分析表明,沙质草地群落共现模式在增温模拟后明显改变(图5 ).相比于沙质草地,模拟增温后群落中物种之间的互作网络的节点数、相互作用强度和平均连通度明显降低,表明群落复杂性降低;而聚类系数增加,表明群落的组织水平增强;网络连通性指数增加而模块化指数降低,表明群落复杂度增加而组织程度降低(表3 ).有研究表明,植物群落的组织程度和复杂度还受到放牧和退化梯度的影响:草甸退化越严重,组织程度越低、复杂度越高;模拟放牧使组织程度呈上升趋势、网络连通性呈降低趋势[13 ] .亦有研究表明,群落稳定性与物种丰富度指数有很高的相关性[45 ] .沙质草地在模拟增温后,群落的丰富度指数与网络结构稳定性均降低,与袁梓裕等[46 ] 的研究中群落稳定性和物种丰富度呈显著正相关这一结果相近,说明植物在受到增温和水分胁迫的共同作用下,植物物种之间的竞争加剧导致群落组成减少以及各物种之间的相互作用强度和模块化水平均有降低趋势[43 ,32 ] .关键类群是群落中高度连接的物种类群,强烈影响着群落中的其他物种及其结构和功能[47 ] .模拟增温后的沙质草地关键物种除旱生植物菊科的黄蒿外,还有藜科的猪毛菜和禾本科的糙隐子草.研究区本就水分匮乏,模拟增温又使土壤含水量减少,因此,关键物种的变化也可用与土壤水分相关的原因来解释:被确定为关键物种的植物可能在保持土壤水分方面发挥关键作用[14 ] . ...
... [13 ].亦有研究表明,群落稳定性与物种丰富度指数有很高的相关性[45 ] .沙质草地在模拟增温后,群落的丰富度指数与网络结构稳定性均降低,与袁梓裕等[46 ] 的研究中群落稳定性和物种丰富度呈显著正相关这一结果相近,说明植物在受到增温和水分胁迫的共同作用下,植物物种之间的竞争加剧导致群落组成减少以及各物种之间的相互作用强度和模块化水平均有降低趋势[43 ,32 ] .关键类群是群落中高度连接的物种类群,强烈影响着群落中的其他物种及其结构和功能[47 ] .模拟增温后的沙质草地关键物种除旱生植物菊科的黄蒿外,还有藜科的猪毛菜和禾本科的糙隐子草.研究区本就水分匮乏,模拟增温又使土壤含水量减少,因此,关键物种的变化也可用与土壤水分相关的原因来解释:被确定为关键物种的植物可能在保持土壤水分方面发挥关键作用[14 ] . ...
Grazing exclusion changed the complexity and keystone species of alpine meadows on the Qinghai-Tibetan Plateau
4
2021
... 草地是全球分布面积最大的陆地生态系统,具有重要的生态和社会功能[8 -9 ] .草地生态系统的植物群落对变暖响应极为敏感[10 ] .增温对群落物种多样性具有促进或抑制或无明显作用[7 ,11 -12 ] ,对群落生物量积累的作用因增幅而异,低幅增温具有促进作用,高幅增温抑制积累[12 ] .除了物种组成、优势种、多样性、生产力等指标可以描述植物群落结构及功能之外,群落连接性、组织程度和关键种也是常见的有效指标[13 ] .目前,有关草地植物群落网络结构复杂性和关键种变化的研究多在人为干扰方面[14 ] ,很少涉及气候变化. ...
... 基于物种间的Pearson相关系数进行共生网络分析,分析过程中仅保留相关性系数达到显著性(P <0.05)的物种[14 ] ,以此探究沙地植物群落在增温处理后的共现模式.利用Gephi 0.10.1进行网络分析可视化绘图,Statistics工具计算代表网络拓扑性质的参数,包括节点数、模块化属性、网络连通性等.在植物群落生态网络中,节点代表群落中的物种,连接代表物种之间的关系[26 -27 ] ;网络节点数、连接数及平均连通度用来指示网络的规模和复杂程度,其中,平均连通度越大表明网络节点之间具有越强的相互作用,群落越复杂[28 ] ;平均聚类系数表明了网络的组织水平,平均聚类系数越高,组织水平越高[14 ] ;连通性指数用来表示群落复杂度,值越大代表群落复杂度越高;模块化指数衡量了网络图结构的模块化程度[13 ] .从群落结构维持的角度出发,通常根据网络中Hub指标的最大值确定关键物种[29 ] . ...
... [14 ];连通性指数用来表示群落复杂度,值越大代表群落复杂度越高;模块化指数衡量了网络图结构的模块化程度[13 ] .从群落结构维持的角度出发,通常根据网络中Hub指标的最大值确定关键物种[29 ] . ...
... 增温会导致沙质草地群落种间关系发生变化.除了多样性和生产力可以衡量群落特征以外,群落网络结构的复杂性以及关键类群也可以评价其结构和功能[13 ,44 ] .共生网络分析表明,沙质草地群落共现模式在增温模拟后明显改变(图5 ).相比于沙质草地,模拟增温后群落中物种之间的互作网络的节点数、相互作用强度和平均连通度明显降低,表明群落复杂性降低;而聚类系数增加,表明群落的组织水平增强;网络连通性指数增加而模块化指数降低,表明群落复杂度增加而组织程度降低(表3 ).有研究表明,植物群落的组织程度和复杂度还受到放牧和退化梯度的影响:草甸退化越严重,组织程度越低、复杂度越高;模拟放牧使组织程度呈上升趋势、网络连通性呈降低趋势[13 ] .亦有研究表明,群落稳定性与物种丰富度指数有很高的相关性[45 ] .沙质草地在模拟增温后,群落的丰富度指数与网络结构稳定性均降低,与袁梓裕等[46 ] 的研究中群落稳定性和物种丰富度呈显著正相关这一结果相近,说明植物在受到增温和水分胁迫的共同作用下,植物物种之间的竞争加剧导致群落组成减少以及各物种之间的相互作用强度和模块化水平均有降低趋势[43 ,32 ] .关键类群是群落中高度连接的物种类群,强烈影响着群落中的其他物种及其结构和功能[47 ] .模拟增温后的沙质草地关键物种除旱生植物菊科的黄蒿外,还有藜科的猪毛菜和禾本科的糙隐子草.研究区本就水分匮乏,模拟增温又使土壤含水量减少,因此,关键物种的变化也可用与土壤水分相关的原因来解释:被确定为关键物种的植物可能在保持土壤水分方面发挥关键作用[14 ] . ...
干旱、半干旱环境降水脉动对生态系统的影响
1
2011
... 科尔沁沙地是中国北方农牧交错带典型的生态脆弱区,对气候变化极为敏感[15 -16 ] .近年来,关于科尔沁沙质草地植物群落结构和功能的响应研究多在降水和人类干扰(氮添加、放牧)等方面[16 -18 ] ,而对气候变暖的关注较少.该地区近年来变暖趋势明显,2018年平均气温达到8 ℃[19 ] .所以,探究科尔沁沙质草地植物群落结构和功能对增温的响应显得尤为迫切.本研究以科尔沁沙质草地为研究对象,通过采用开顶式气室法(Open top chamber,OTC)模拟增温,分析优势种、物种多样性、生物量、复杂度、关键物种等植物群落结构和功能指标的变化规律,为该区域在全球气候变暖背景下草地植物群落结构及功能变化的研究提供理论依据和数据支撑. ...
极端干旱与氮添加对半干旱沙质草地物种多样性、叶性状和生产力的影响
2
2020
... 科尔沁沙地是中国北方农牧交错带典型的生态脆弱区,对气候变化极为敏感[15 -16 ] .近年来,关于科尔沁沙质草地植物群落结构和功能的响应研究多在降水和人类干扰(氮添加、放牧)等方面[16 -18 ] ,而对气候变暖的关注较少.该地区近年来变暖趋势明显,2018年平均气温达到8 ℃[19 ] .所以,探究科尔沁沙质草地植物群落结构和功能对增温的响应显得尤为迫切.本研究以科尔沁沙质草地为研究对象,通过采用开顶式气室法(Open top chamber,OTC)模拟增温,分析优势种、物种多样性、生物量、复杂度、关键物种等植物群落结构和功能指标的变化规律,为该区域在全球气候变暖背景下草地植物群落结构及功能变化的研究提供理论依据和数据支撑. ...
... [16 -18 ],而对气候变暖的关注较少.该地区近年来变暖趋势明显,2018年平均气温达到8 ℃[19 ] .所以,探究科尔沁沙质草地植物群落结构和功能对增温的响应显得尤为迫切.本研究以科尔沁沙质草地为研究对象,通过采用开顶式气室法(Open top chamber,OTC)模拟增温,分析优势种、物种多样性、生物量、复杂度、关键物种等植物群落结构和功能指标的变化规律,为该区域在全球气候变暖背景下草地植物群落结构及功能变化的研究提供理论依据和数据支撑. ...
人类放牧活动与气候变化对科尔沁沙质草地植物多样性的影响
0
2008
科尔沁沙质草地生物量积累过程对降水变化的响应模拟
2
2020
... 科尔沁沙地是中国北方农牧交错带典型的生态脆弱区,对气候变化极为敏感[15 -16 ] .近年来,关于科尔沁沙质草地植物群落结构和功能的响应研究多在降水和人类干扰(氮添加、放牧)等方面[16 -18 ] ,而对气候变暖的关注较少.该地区近年来变暖趋势明显,2018年平均气温达到8 ℃[19 ] .所以,探究科尔沁沙质草地植物群落结构和功能对增温的响应显得尤为迫切.本研究以科尔沁沙质草地为研究对象,通过采用开顶式气室法(Open top chamber,OTC)模拟增温,分析优势种、物种多样性、生物量、复杂度、关键物种等植物群落结构和功能指标的变化规律,为该区域在全球气候变暖背景下草地植物群落结构及功能变化的研究提供理论依据和数据支撑. ...
... 科尔沁沙质草地生态系统脆弱,长期放牧及土壤水分匮缺导致沙质草地不断退化.通过围封禁牧及人工修复会逐步恢复沙质草地,但气候的异常变化会影响沙质草地生态系统结构及其功能.降水的增加或减少会引起草地生态系统植物群落结构和功能发生明显改变[18 ,30 -31 ] .温度也是影响植物正常生长的重要因子,尤其在干旱半干旱区,植物除了对养分吸收的竞争加剧外,还存在对水分吸收的竞争[32 ] .本研究结果表明,沙质草地在模拟增温后草本群落组成发生明显变化,群落物种(科)数低于沙质草地(表2 ).以禾本科为主的优势种占比明显减少,这与Ganjurjav等[6 ] 的研究结果相似.禾本科植物占比减少,为杂草类植物提供了更大的生存空间.此外,狗尾草、糙隐子草、黄蒿和大籽蒿解释了模拟增温导致群落组成差异的63%,且以一年生草本狗尾草为主.吕朋等[33 ] 在研究沙质草地群落特征中发现,一年生草本的禾本科狗尾草是优势物种,这与本研究结果一致,但模拟增温后的狗尾草重要值显著降低,表明狗尾草等一年生草本对模拟增温后的土壤环境较敏感.在增温和水分胁迫的环境中,土壤养分因此受到的负面影响导致群落组成发生改变[34 ] .模拟增温后沙质草地群落多样性呈下降趋势,其中Simpson指数、Shannon-Wiener指数和物种丰富度指数显著下降,而Pielou指数下降趋势不显著(图3 ),这与之前的大部分研究结果一致[35 -37 ] .也有学者认为低幅增温对多样性产生促进作用,但当增温超过一定幅度时,促进作用会减弱甚至变成抑制作用[12 ] ,造成这种不一致的原因可能是物种组成以及研究区间的差异.郭轶瑞等[38 ] 的研究发现耐旱植物对增温的抵抗力较强,而喜湿植物在受水分限制时竞争能力减弱导致其比重降低.增温导致土壤含水量降低,同时,物种丰富度和多样性指数与土壤湿度呈显著正相关关系,表明在干旱半干旱区,多样性随水分胁迫强度的增加更加敏感. ...
1989-2018年奈曼旗气温变化特征分析
1
2019
... 科尔沁沙地是中国北方农牧交错带典型的生态脆弱区,对气候变化极为敏感[15 -16 ] .近年来,关于科尔沁沙质草地植物群落结构和功能的响应研究多在降水和人类干扰(氮添加、放牧)等方面[16 -18 ] ,而对气候变暖的关注较少.该地区近年来变暖趋势明显,2018年平均气温达到8 ℃[19 ] .所以,探究科尔沁沙质草地植物群落结构和功能对增温的响应显得尤为迫切.本研究以科尔沁沙质草地为研究对象,通过采用开顶式气室法(Open top chamber,OTC)模拟增温,分析优势种、物种多样性、生物量、复杂度、关键物种等植物群落结构和功能指标的变化规律,为该区域在全球气候变暖背景下草地植物群落结构及功能变化的研究提供理论依据和数据支撑. ...
基于多源数据的科尔沁沙地生态环境变化研究
1
2017
... 试验样地位于内蒙古自治区通辽市奈曼旗境内的中国科学院奈曼沙漠化研究站(42°41′N、120°55′E,海拔358 m),处于科尔沁沙地腹地.试验地属温带半干旱大陆性气候[20 ] ,年降水量356.9 mm,主要发生在6—8月,年蒸发量1 972.8 mm;年平均气温6.5 ℃,≥10 ℃的年积温大于3 000 ℃,无霜期151 d[21 ] .土壤以风沙土为主.植被类型为典型沙地榆树疏林草地,多为旱生和沙生植物,主要植物种有黄蒿(Artemisia scoparia )、狗尾草(Setaria viridis )、猪毛菜(Salsola collina )、虫实(Corispermum hyssopifolium )、糙隐子草(Cleistogenes squarrosa )等. ...
不同强度净风频繁吹袭对樟子松(Pinus sylvestris var.mongolica )幼苗光合蒸腾特征的影响
1
2017
... 试验样地位于内蒙古自治区通辽市奈曼旗境内的中国科学院奈曼沙漠化研究站(42°41′N、120°55′E,海拔358 m),处于科尔沁沙地腹地.试验地属温带半干旱大陆性气候[20 ] ,年降水量356.9 mm,主要发生在6—8月,年蒸发量1 972.8 mm;年平均气温6.5 ℃,≥10 ℃的年积温大于3 000 ℃,无霜期151 d[21 ] .土壤以风沙土为主.植被类型为典型沙地榆树疏林草地,多为旱生和沙生植物,主要植物种有黄蒿(Artemisia scoparia )、狗尾草(Setaria viridis )、猪毛菜(Salsola collina )、虫实(Corispermum hyssopifolium )、糙隐子草(Cleistogenes squarrosa )等. ...
科尔沁沙地植被恢复过程中群落组成及多样性演变特征
3
2022
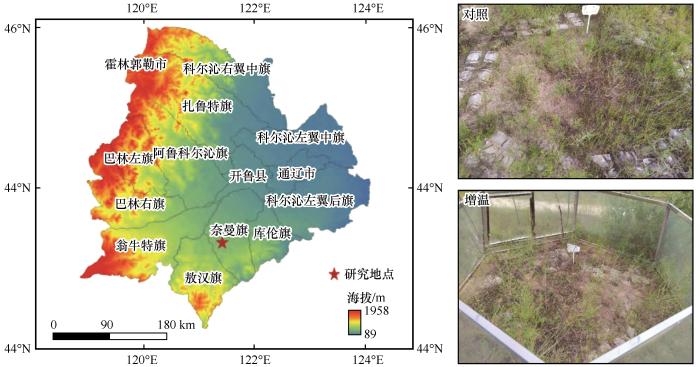
... 选择地势平坦、植物物种分布均匀的区域作为试验区.试验从2018年5月开始,采用增温模拟设计,分为增温和对照处理,每个处理重复8次,共计16个小区,小区面积为1.2 m×3 m(
图1 ).采用OTC全年增温.OTC为正六边形,高1 m,顶部直径2.4 m,底部直径3 m,面板材料采用聚碳酸酯板,透光率92%.
图1 试验样地<sup>[<xref ref-type="bibr" rid="R22">22</xref>]</sup>和增温装置图 Diagram of the experimental design<sup>[<xref ref-type="bibr" rid="R22">22</xref>]</sup> and passive open-top chamber (OTC) Fig.1 ![]()
每个试验小区内,使用iButton纽扣式温度记录仪(美国Maxim公司)测量土壤表层(0~10 cm)的温度.使用便携式湿度探针土壤湿度检测仪测量土壤表层10 cm深处的湿度,测量结果为体积含水量.本试验中2022年的OTC增温罩实现表层土壤(0~10 cm)平均温度增幅达到0.79 ℃,表层土壤(0~10 cm)含水量降幅达到0.13%(表1 ).同年7月下旬,在每个小区的中心区域设置1 m×1 m的样方框,进行植物群落特征调查,记录样方内各物种名、高度、盖度、密度[22 ] .植被调查结束后,齐地面剪除样方内所有植物,分物种装入袋中.所有植物样品先在烘箱中105 ℃条件下杀青0.5 h,然后在65 ℃下烘干48 h并称重,计算植物地上生物量.为了更好地研究科尔沁沙质草地在增温过程中草本植物物种组成的变化,本研究按照其生活型将草本植物划分为2个不同的功能群,即一年生草本和多年生草本. ...
... [
22 ] and passive open-top chamber (OTC)
Fig.1 ![]()
每个试验小区内,使用iButton纽扣式温度记录仪(美国Maxim公司)测量土壤表层(0~10 cm)的温度.使用便携式湿度探针土壤湿度检测仪测量土壤表层10 cm深处的湿度,测量结果为体积含水量.本试验中2022年的OTC增温罩实现表层土壤(0~10 cm)平均温度增幅达到0.79 ℃,表层土壤(0~10 cm)含水量降幅达到0.13%(表1 ).同年7月下旬,在每个小区的中心区域设置1 m×1 m的样方框,进行植物群落特征调查,记录样方内各物种名、高度、盖度、密度[22 ] .植被调查结束后,齐地面剪除样方内所有植物,分物种装入袋中.所有植物样品先在烘箱中105 ℃条件下杀青0.5 h,然后在65 ℃下烘干48 h并称重,计算植物地上生物量.为了更好地研究科尔沁沙质草地在增温过程中草本植物物种组成的变化,本研究按照其生活型将草本植物划分为2个不同的功能群,即一年生草本和多年生草本. ...
... 每个试验小区内,使用iButton纽扣式温度记录仪(美国Maxim公司)测量土壤表层(0~10 cm)的温度.使用便携式湿度探针土壤湿度检测仪测量土壤表层10 cm深处的湿度,测量结果为体积含水量.本试验中2022年的OTC增温罩实现表层土壤(0~10 cm)平均温度增幅达到0.79 ℃,表层土壤(0~10 cm)含水量降幅达到0.13%(表1 ).同年7月下旬,在每个小区的中心区域设置1 m×1 m的样方框,进行植物群落特征调查,记录样方内各物种名、高度、盖度、密度[22 ] .植被调查结束后,齐地面剪除样方内所有植物,分物种装入袋中.所有植物样品先在烘箱中105 ℃条件下杀青0.5 h,然后在65 ℃下烘干48 h并称重,计算植物地上生物量.为了更好地研究科尔沁沙质草地在增温过程中草本植物物种组成的变化,本研究按照其生活型将草本植物划分为2个不同的功能群,即一年生草本和多年生草本. ...
草地群落植物种等级划分及重要值方法改进
1
2017
... 基于植被调查的结果计算群落各物种的重要值和物种多样性指标(Simpson指数、Shannon-Wiener指数、Pielou指数和物种丰富度),其计算公式[23 -24 ] 如下: ...
放牧和模拟增温对藏北高寒草地植物群落特征及生产力的影响
1
2019
... 基于植被调查的结果计算群落各物种的重要值和物种多样性指标(Simpson指数、Shannon-Wiener指数、Pielou指数和物种丰富度),其计算公式[23 -24 ] 如下: ...
Nonparametric multivariate analyses of changes in community structure
1
1993
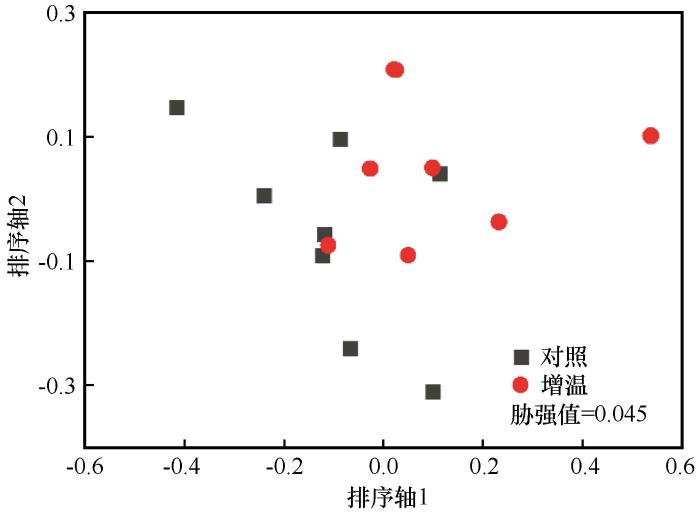
... 采用t 检验比较增温和对照处理下沙质草地群落多样性指数以及生物量的差异,统计分析使用SPSS 22.0软件包.在此基础上,利用线性拟合分析群落多样性与土壤温度和湿度以及生物量的关系,统计分析使用SPSS 22.0软件包.利用非度量多维尺度(Non-metric multidimensional scaling,NMDS)和多元方差(Permutational multivariate analysis of variance,PERMANOVA)分析增温和对照处理下的群落组成差异,然后利用相似性百分比(Similarity per⁃centage analysis,SIMPER)分析确定2种处理下群落的平均相异性和主要植物类群的贡献率,统计分析使用Past 4.01软件包.NMDS排序结果采用胁强系数(Stress)衡量排序分析结果的优劣,其中Stress<0.01,完全可信;0.01<Stress<0.05,可信;0.05<Stress<0.1,基本可信;0.1<Stress<0.2,部分信息不可信;0.2<Stress<0.3,不可信[25 ] . ...
Ecological networks and their fragility
1
2006
... 基于物种间的Pearson相关系数进行共生网络分析,分析过程中仅保留相关性系数达到显著性(P <0.05)的物种[14 ] ,以此探究沙地植物群落在增温处理后的共现模式.利用Gephi 0.10.1进行网络分析可视化绘图,Statistics工具计算代表网络拓扑性质的参数,包括节点数、模块化属性、网络连通性等.在植物群落生态网络中,节点代表群落中的物种,连接代表物种之间的关系[26 -27 ] ;网络节点数、连接数及平均连通度用来指示网络的规模和复杂程度,其中,平均连通度越大表明网络节点之间具有越强的相互作用,群落越复杂[28 ] ;平均聚类系数表明了网络的组织水平,平均聚类系数越高,组织水平越高[14 ] ;连通性指数用来表示群落复杂度,值越大代表群落复杂度越高;模块化指数衡量了网络图结构的模块化程度[13 ] .从群落结构维持的角度出发,通常根据网络中Hub指标的最大值确定关键物种[29 ] . ...
Networks in ecology
1
2007
... 基于物种间的Pearson相关系数进行共生网络分析,分析过程中仅保留相关性系数达到显著性(P <0.05)的物种[14 ] ,以此探究沙地植物群落在增温处理后的共现模式.利用Gephi 0.10.1进行网络分析可视化绘图,Statistics工具计算代表网络拓扑性质的参数,包括节点数、模块化属性、网络连通性等.在植物群落生态网络中,节点代表群落中的物种,连接代表物种之间的关系[26 -27 ] ;网络节点数、连接数及平均连通度用来指示网络的规模和复杂程度,其中,平均连通度越大表明网络节点之间具有越强的相互作用,群落越复杂[28 ] ;平均聚类系数表明了网络的组织水平,平均聚类系数越高,组织水平越高[14 ] ;连通性指数用来表示群落复杂度,值越大代表群落复杂度越高;模块化指数衡量了网络图结构的模块化程度[13 ] .从群落结构维持的角度出发,通常根据网络中Hub指标的最大值确定关键物种[29 ] . ...
太湖不同营养水平湖区沉积环境微生物分子生态网络特征及其环境响应分析
1
2018
... 基于物种间的Pearson相关系数进行共生网络分析,分析过程中仅保留相关性系数达到显著性(P <0.05)的物种[14 ] ,以此探究沙地植物群落在增温处理后的共现模式.利用Gephi 0.10.1进行网络分析可视化绘图,Statistics工具计算代表网络拓扑性质的参数,包括节点数、模块化属性、网络连通性等.在植物群落生态网络中,节点代表群落中的物种,连接代表物种之间的关系[26 -27 ] ;网络节点数、连接数及平均连通度用来指示网络的规模和复杂程度,其中,平均连通度越大表明网络节点之间具有越强的相互作用,群落越复杂[28 ] ;平均聚类系数表明了网络的组织水平,平均聚类系数越高,组织水平越高[14 ] ;连通性指数用来表示群落复杂度,值越大代表群落复杂度越高;模块化指数衡量了网络图结构的模块化程度[13 ] .从群落结构维持的角度出发,通常根据网络中Hub指标的最大值确定关键物种[29 ] . ...
Ecosystem-level effects of keystone species reintroduction:a literature review
1
2018
... 基于物种间的Pearson相关系数进行共生网络分析,分析过程中仅保留相关性系数达到显著性(P <0.05)的物种[14 ] ,以此探究沙地植物群落在增温处理后的共现模式.利用Gephi 0.10.1进行网络分析可视化绘图,Statistics工具计算代表网络拓扑性质的参数,包括节点数、模块化属性、网络连通性等.在植物群落生态网络中,节点代表群落中的物种,连接代表物种之间的关系[26 -27 ] ;网络节点数、连接数及平均连通度用来指示网络的规模和复杂程度,其中,平均连通度越大表明网络节点之间具有越强的相互作用,群落越复杂[28 ] ;平均聚类系数表明了网络的组织水平,平均聚类系数越高,组织水平越高[14 ] ;连通性指数用来表示群落复杂度,值越大代表群落复杂度越高;模块化指数衡量了网络图结构的模块化程度[13 ] .从群落结构维持的角度出发,通常根据网络中Hub指标的最大值确定关键物种[29 ] . ...
水分梯度下荒漠植物多样性与稳定性对土壤因子的响应
1
2021
... 科尔沁沙质草地生态系统脆弱,长期放牧及土壤水分匮缺导致沙质草地不断退化.通过围封禁牧及人工修复会逐步恢复沙质草地,但气候的异常变化会影响沙质草地生态系统结构及其功能.降水的增加或减少会引起草地生态系统植物群落结构和功能发生明显改变[18 ,30 -31 ] .温度也是影响植物正常生长的重要因子,尤其在干旱半干旱区,植物除了对养分吸收的竞争加剧外,还存在对水分吸收的竞争[32 ] .本研究结果表明,沙质草地在模拟增温后草本群落组成发生明显变化,群落物种(科)数低于沙质草地(表2 ).以禾本科为主的优势种占比明显减少,这与Ganjurjav等[6 ] 的研究结果相似.禾本科植物占比减少,为杂草类植物提供了更大的生存空间.此外,狗尾草、糙隐子草、黄蒿和大籽蒿解释了模拟增温导致群落组成差异的63%,且以一年生草本狗尾草为主.吕朋等[33 ] 在研究沙质草地群落特征中发现,一年生草本的禾本科狗尾草是优势物种,这与本研究结果一致,但模拟增温后的狗尾草重要值显著降低,表明狗尾草等一年生草本对模拟增温后的土壤环境较敏感.在增温和水分胁迫的环境中,土壤养分因此受到的负面影响导致群落组成发生改变[34 ] .模拟增温后沙质草地群落多样性呈下降趋势,其中Simpson指数、Shannon-Wiener指数和物种丰富度指数显著下降,而Pielou指数下降趋势不显著(图3 ),这与之前的大部分研究结果一致[35 -37 ] .也有学者认为低幅增温对多样性产生促进作用,但当增温超过一定幅度时,促进作用会减弱甚至变成抑制作用[12 ] ,造成这种不一致的原因可能是物种组成以及研究区间的差异.郭轶瑞等[38 ] 的研究发现耐旱植物对增温的抵抗力较强,而喜湿植物在受水分限制时竞争能力减弱导致其比重降低.增温导致土壤含水量降低,同时,物种丰富度和多样性指数与土壤湿度呈显著正相关关系,表明在干旱半干旱区,多样性随水分胁迫强度的增加更加敏感. ...
降水控制对荒漠植物群落物种多样性和生物量的影响
1
2018
... 科尔沁沙质草地生态系统脆弱,长期放牧及土壤水分匮缺导致沙质草地不断退化.通过围封禁牧及人工修复会逐步恢复沙质草地,但气候的异常变化会影响沙质草地生态系统结构及其功能.降水的增加或减少会引起草地生态系统植物群落结构和功能发生明显改变[18 ,30 -31 ] .温度也是影响植物正常生长的重要因子,尤其在干旱半干旱区,植物除了对养分吸收的竞争加剧外,还存在对水分吸收的竞争[32 ] .本研究结果表明,沙质草地在模拟增温后草本群落组成发生明显变化,群落物种(科)数低于沙质草地(表2 ).以禾本科为主的优势种占比明显减少,这与Ganjurjav等[6 ] 的研究结果相似.禾本科植物占比减少,为杂草类植物提供了更大的生存空间.此外,狗尾草、糙隐子草、黄蒿和大籽蒿解释了模拟增温导致群落组成差异的63%,且以一年生草本狗尾草为主.吕朋等[33 ] 在研究沙质草地群落特征中发现,一年生草本的禾本科狗尾草是优势物种,这与本研究结果一致,但模拟增温后的狗尾草重要值显著降低,表明狗尾草等一年生草本对模拟增温后的土壤环境较敏感.在增温和水分胁迫的环境中,土壤养分因此受到的负面影响导致群落组成发生改变[34 ] .模拟增温后沙质草地群落多样性呈下降趋势,其中Simpson指数、Shannon-Wiener指数和物种丰富度指数显著下降,而Pielou指数下降趋势不显著(图3 ),这与之前的大部分研究结果一致[35 -37 ] .也有学者认为低幅增温对多样性产生促进作用,但当增温超过一定幅度时,促进作用会减弱甚至变成抑制作用[12 ] ,造成这种不一致的原因可能是物种组成以及研究区间的差异.郭轶瑞等[38 ] 的研究发现耐旱植物对增温的抵抗力较强,而喜湿植物在受水分限制时竞争能力减弱导致其比重降低.增温导致土壤含水量降低,同时,物种丰富度和多样性指数与土壤湿度呈显著正相关关系,表明在干旱半干旱区,多样性随水分胁迫强度的增加更加敏感. ...
Precipitation alters temperature effects on ecosystem respiration in Tibetan alpine meadows
2
2018
... 科尔沁沙质草地生态系统脆弱,长期放牧及土壤水分匮缺导致沙质草地不断退化.通过围封禁牧及人工修复会逐步恢复沙质草地,但气候的异常变化会影响沙质草地生态系统结构及其功能.降水的增加或减少会引起草地生态系统植物群落结构和功能发生明显改变[18 ,30 -31 ] .温度也是影响植物正常生长的重要因子,尤其在干旱半干旱区,植物除了对养分吸收的竞争加剧外,还存在对水分吸收的竞争[32 ] .本研究结果表明,沙质草地在模拟增温后草本群落组成发生明显变化,群落物种(科)数低于沙质草地(表2 ).以禾本科为主的优势种占比明显减少,这与Ganjurjav等[6 ] 的研究结果相似.禾本科植物占比减少,为杂草类植物提供了更大的生存空间.此外,狗尾草、糙隐子草、黄蒿和大籽蒿解释了模拟增温导致群落组成差异的63%,且以一年生草本狗尾草为主.吕朋等[33 ] 在研究沙质草地群落特征中发现,一年生草本的禾本科狗尾草是优势物种,这与本研究结果一致,但模拟增温后的狗尾草重要值显著降低,表明狗尾草等一年生草本对模拟增温后的土壤环境较敏感.在增温和水分胁迫的环境中,土壤养分因此受到的负面影响导致群落组成发生改变[34 ] .模拟增温后沙质草地群落多样性呈下降趋势,其中Simpson指数、Shannon-Wiener指数和物种丰富度指数显著下降,而Pielou指数下降趋势不显著(图3 ),这与之前的大部分研究结果一致[35 -37 ] .也有学者认为低幅增温对多样性产生促进作用,但当增温超过一定幅度时,促进作用会减弱甚至变成抑制作用[12 ] ,造成这种不一致的原因可能是物种组成以及研究区间的差异.郭轶瑞等[38 ] 的研究发现耐旱植物对增温的抵抗力较强,而喜湿植物在受水分限制时竞争能力减弱导致其比重降低.增温导致土壤含水量降低,同时,物种丰富度和多样性指数与土壤湿度呈显著正相关关系,表明在干旱半干旱区,多样性随水分胁迫强度的增加更加敏感. ...
... 增温会导致沙质草地群落种间关系发生变化.除了多样性和生产力可以衡量群落特征以外,群落网络结构的复杂性以及关键类群也可以评价其结构和功能[13 ,44 ] .共生网络分析表明,沙质草地群落共现模式在增温模拟后明显改变(图5 ).相比于沙质草地,模拟增温后群落中物种之间的互作网络的节点数、相互作用强度和平均连通度明显降低,表明群落复杂性降低;而聚类系数增加,表明群落的组织水平增强;网络连通性指数增加而模块化指数降低,表明群落复杂度增加而组织程度降低(表3 ).有研究表明,植物群落的组织程度和复杂度还受到放牧和退化梯度的影响:草甸退化越严重,组织程度越低、复杂度越高;模拟放牧使组织程度呈上升趋势、网络连通性呈降低趋势[13 ] .亦有研究表明,群落稳定性与物种丰富度指数有很高的相关性[45 ] .沙质草地在模拟增温后,群落的丰富度指数与网络结构稳定性均降低,与袁梓裕等[46 ] 的研究中群落稳定性和物种丰富度呈显著正相关这一结果相近,说明植物在受到增温和水分胁迫的共同作用下,植物物种之间的竞争加剧导致群落组成减少以及各物种之间的相互作用强度和模块化水平均有降低趋势[43 ,32 ] .关键类群是群落中高度连接的物种类群,强烈影响着群落中的其他物种及其结构和功能[47 ] .模拟增温后的沙质草地关键物种除旱生植物菊科的黄蒿外,还有藜科的猪毛菜和禾本科的糙隐子草.研究区本就水分匮乏,模拟增温又使土壤含水量减少,因此,关键物种的变化也可用与土壤水分相关的原因来解释:被确定为关键物种的植物可能在保持土壤水分方面发挥关键作用[14 ] . ...
科尔沁沙地封育过程中植被特征的动态变化
1
2018
... 科尔沁沙质草地生态系统脆弱,长期放牧及土壤水分匮缺导致沙质草地不断退化.通过围封禁牧及人工修复会逐步恢复沙质草地,但气候的异常变化会影响沙质草地生态系统结构及其功能.降水的增加或减少会引起草地生态系统植物群落结构和功能发生明显改变[18 ,30 -31 ] .温度也是影响植物正常生长的重要因子,尤其在干旱半干旱区,植物除了对养分吸收的竞争加剧外,还存在对水分吸收的竞争[32 ] .本研究结果表明,沙质草地在模拟增温后草本群落组成发生明显变化,群落物种(科)数低于沙质草地(表2 ).以禾本科为主的优势种占比明显减少,这与Ganjurjav等[6 ] 的研究结果相似.禾本科植物占比减少,为杂草类植物提供了更大的生存空间.此外,狗尾草、糙隐子草、黄蒿和大籽蒿解释了模拟增温导致群落组成差异的63%,且以一年生草本狗尾草为主.吕朋等[33 ] 在研究沙质草地群落特征中发现,一年生草本的禾本科狗尾草是优势物种,这与本研究结果一致,但模拟增温后的狗尾草重要值显著降低,表明狗尾草等一年生草本对模拟增温后的土壤环境较敏感.在增温和水分胁迫的环境中,土壤养分因此受到的负面影响导致群落组成发生改变[34 ] .模拟增温后沙质草地群落多样性呈下降趋势,其中Simpson指数、Shannon-Wiener指数和物种丰富度指数显著下降,而Pielou指数下降趋势不显著(图3 ),这与之前的大部分研究结果一致[35 -37 ] .也有学者认为低幅增温对多样性产生促进作用,但当增温超过一定幅度时,促进作用会减弱甚至变成抑制作用[12 ] ,造成这种不一致的原因可能是物种组成以及研究区间的差异.郭轶瑞等[38 ] 的研究发现耐旱植物对增温的抵抗力较强,而喜湿植物在受水分限制时竞争能力减弱导致其比重降低.增温导致土壤含水量降低,同时,物种丰富度和多样性指数与土壤湿度呈显著正相关关系,表明在干旱半干旱区,多样性随水分胁迫强度的增加更加敏感. ...
Plant traits and soil fertility mediate productivity losses under extreme drought in C-3 grasslands
2
2021
... 科尔沁沙质草地生态系统脆弱,长期放牧及土壤水分匮缺导致沙质草地不断退化.通过围封禁牧及人工修复会逐步恢复沙质草地,但气候的异常变化会影响沙质草地生态系统结构及其功能.降水的增加或减少会引起草地生态系统植物群落结构和功能发生明显改变[18 ,30 -31 ] .温度也是影响植物正常生长的重要因子,尤其在干旱半干旱区,植物除了对养分吸收的竞争加剧外,还存在对水分吸收的竞争[32 ] .本研究结果表明,沙质草地在模拟增温后草本群落组成发生明显变化,群落物种(科)数低于沙质草地(表2 ).以禾本科为主的优势种占比明显减少,这与Ganjurjav等[6 ] 的研究结果相似.禾本科植物占比减少,为杂草类植物提供了更大的生存空间.此外,狗尾草、糙隐子草、黄蒿和大籽蒿解释了模拟增温导致群落组成差异的63%,且以一年生草本狗尾草为主.吕朋等[33 ] 在研究沙质草地群落特征中发现,一年生草本的禾本科狗尾草是优势物种,这与本研究结果一致,但模拟增温后的狗尾草重要值显著降低,表明狗尾草等一年生草本对模拟增温后的土壤环境较敏感.在增温和水分胁迫的环境中,土壤养分因此受到的负面影响导致群落组成发生改变[34 ] .模拟增温后沙质草地群落多样性呈下降趋势,其中Simpson指数、Shannon-Wiener指数和物种丰富度指数显著下降,而Pielou指数下降趋势不显著(图3 ),这与之前的大部分研究结果一致[35 -37 ] .也有学者认为低幅增温对多样性产生促进作用,但当增温超过一定幅度时,促进作用会减弱甚至变成抑制作用[12 ] ,造成这种不一致的原因可能是物种组成以及研究区间的差异.郭轶瑞等[38 ] 的研究发现耐旱植物对增温的抵抗力较强,而喜湿植物在受水分限制时竞争能力减弱导致其比重降低.增温导致土壤含水量降低,同时,物种丰富度和多样性指数与土壤湿度呈显著正相关关系,表明在干旱半干旱区,多样性随水分胁迫强度的增加更加敏感. ...
... 多样性与生产力的关系是反映群落结构及生态功能的关键指标[39 -40 ] .许多研究证实物种多样性与生产力主要存在正相关、负相关、单峰曲线和不相关4种关系[7 ] ,且正相关关系的结论居多.就科尔沁沙质草地而言,物种丰富度指数和生物量之间呈显著线性正相关,这与孙小丽等[7 ] 对内蒙古中温型荒漠草原区群落生物量随物种丰富度增加而增加的结论相一致,但与郭轶瑞等[41 ] 在科尔沁沙质草地的物种多样性与生产力呈单峰关系这一结论不同.极有可能是因为独特的生境特征差异.研究区处于干旱半干旱区,水热因子对干旱半干旱区植物群落生产力方面作用占主导地位[7 ,11 ] ,因此,推测科尔沁沙质草地生产力受到土壤水热因子的调控.本研究中,增温模拟后的群落和禾本科地上生物量与对照组相比显著下降(图4 ).Arnda等[42 ] 发现,禾本科植物地上生物量减少可能是因为更多杂草类根系在土壤剖面上的分配.也有研究表明,在干旱半干旱区,生产力深受水分限制[11 ] .Grime等[43 ] 发现,在增温和水分胁迫的环境中,植物通常以减少生物量生产来提高水分利用效率以此适应环境,解释了本研究在模拟增温后土壤湿度的降低导致了植物地上生产力的降低.由于水分限制,模拟增温后植物无法利用土壤温度升高的能量进行光合作用,从而呈随着能量增加群落多样性和生产力降低的关系.也有研究表明,增温对生物量的潜在影响不仅与土壤水热条件有关,土壤养分与多样性和生物量也存在显著相关的关系[34 ] .孙小丽等[7 ] 对草地生态系统的研究表明生物量与速效氮、有机碳等土壤养分呈显著的线性正相关.生物量分配还受到群落组成的影响,禾本科地上生物量减少为杂草类地下生物量提供了更多的水分和养分资源[42 ] . ...
短期增温对川西北高寒草甸植物群落结构和叶片性状的影响
1
2022
... 科尔沁沙质草地生态系统脆弱,长期放牧及土壤水分匮缺导致沙质草地不断退化.通过围封禁牧及人工修复会逐步恢复沙质草地,但气候的异常变化会影响沙质草地生态系统结构及其功能.降水的增加或减少会引起草地生态系统植物群落结构和功能发生明显改变[18 ,30 -31 ] .温度也是影响植物正常生长的重要因子,尤其在干旱半干旱区,植物除了对养分吸收的竞争加剧外,还存在对水分吸收的竞争[32 ] .本研究结果表明,沙质草地在模拟增温后草本群落组成发生明显变化,群落物种(科)数低于沙质草地(表2 ).以禾本科为主的优势种占比明显减少,这与Ganjurjav等[6 ] 的研究结果相似.禾本科植物占比减少,为杂草类植物提供了更大的生存空间.此外,狗尾草、糙隐子草、黄蒿和大籽蒿解释了模拟增温导致群落组成差异的63%,且以一年生草本狗尾草为主.吕朋等[33 ] 在研究沙质草地群落特征中发现,一年生草本的禾本科狗尾草是优势物种,这与本研究结果一致,但模拟增温后的狗尾草重要值显著降低,表明狗尾草等一年生草本对模拟增温后的土壤环境较敏感.在增温和水分胁迫的环境中,土壤养分因此受到的负面影响导致群落组成发生改变[34 ] .模拟增温后沙质草地群落多样性呈下降趋势,其中Simpson指数、Shannon-Wiener指数和物种丰富度指数显著下降,而Pielou指数下降趋势不显著(图3 ),这与之前的大部分研究结果一致[35 -37 ] .也有学者认为低幅增温对多样性产生促进作用,但当增温超过一定幅度时,促进作用会减弱甚至变成抑制作用[12 ] ,造成这种不一致的原因可能是物种组成以及研究区间的差异.郭轶瑞等[38 ] 的研究发现耐旱植物对增温的抵抗力较强,而喜湿植物在受水分限制时竞争能力减弱导致其比重降低.增温导致土壤含水量降低,同时,物种丰富度和多样性指数与土壤湿度呈显著正相关关系,表明在干旱半干旱区,多样性随水分胁迫强度的增加更加敏感. ...
Experimental warming causes rapid loss of plant diversity in New England salt marshes
0
2009
Experimental drought and warming decrease diversity and slow down post-fire succession in a Mediterranean shrubland
1
2009
... 科尔沁沙质草地生态系统脆弱,长期放牧及土壤水分匮缺导致沙质草地不断退化.通过围封禁牧及人工修复会逐步恢复沙质草地,但气候的异常变化会影响沙质草地生态系统结构及其功能.降水的增加或减少会引起草地生态系统植物群落结构和功能发生明显改变[18 ,30 -31 ] .温度也是影响植物正常生长的重要因子,尤其在干旱半干旱区,植物除了对养分吸收的竞争加剧外,还存在对水分吸收的竞争[32 ] .本研究结果表明,沙质草地在模拟增温后草本群落组成发生明显变化,群落物种(科)数低于沙质草地(表2 ).以禾本科为主的优势种占比明显减少,这与Ganjurjav等[6 ] 的研究结果相似.禾本科植物占比减少,为杂草类植物提供了更大的生存空间.此外,狗尾草、糙隐子草、黄蒿和大籽蒿解释了模拟增温导致群落组成差异的63%,且以一年生草本狗尾草为主.吕朋等[33 ] 在研究沙质草地群落特征中发现,一年生草本的禾本科狗尾草是优势物种,这与本研究结果一致,但模拟增温后的狗尾草重要值显著降低,表明狗尾草等一年生草本对模拟增温后的土壤环境较敏感.在增温和水分胁迫的环境中,土壤养分因此受到的负面影响导致群落组成发生改变[34 ] .模拟增温后沙质草地群落多样性呈下降趋势,其中Simpson指数、Shannon-Wiener指数和物种丰富度指数显著下降,而Pielou指数下降趋势不显著(图3 ),这与之前的大部分研究结果一致[35 -37 ] .也有学者认为低幅增温对多样性产生促进作用,但当增温超过一定幅度时,促进作用会减弱甚至变成抑制作用[12 ] ,造成这种不一致的原因可能是物种组成以及研究区间的差异.郭轶瑞等[38 ] 的研究发现耐旱植物对增温的抵抗力较强,而喜湿植物在受水分限制时竞争能力减弱导致其比重降低.增温导致土壤含水量降低,同时,物种丰富度和多样性指数与土壤湿度呈显著正相关关系,表明在干旱半干旱区,多样性随水分胁迫强度的增加更加敏感. ...
科尔沁沙地生物结皮的土壤种子库特征
1
2007
... 科尔沁沙质草地生态系统脆弱,长期放牧及土壤水分匮缺导致沙质草地不断退化.通过围封禁牧及人工修复会逐步恢复沙质草地,但气候的异常变化会影响沙质草地生态系统结构及其功能.降水的增加或减少会引起草地生态系统植物群落结构和功能发生明显改变[18 ,30 -31 ] .温度也是影响植物正常生长的重要因子,尤其在干旱半干旱区,植物除了对养分吸收的竞争加剧外,还存在对水分吸收的竞争[32 ] .本研究结果表明,沙质草地在模拟增温后草本群落组成发生明显变化,群落物种(科)数低于沙质草地(表2 ).以禾本科为主的优势种占比明显减少,这与Ganjurjav等[6 ] 的研究结果相似.禾本科植物占比减少,为杂草类植物提供了更大的生存空间.此外,狗尾草、糙隐子草、黄蒿和大籽蒿解释了模拟增温导致群落组成差异的63%,且以一年生草本狗尾草为主.吕朋等[33 ] 在研究沙质草地群落特征中发现,一年生草本的禾本科狗尾草是优势物种,这与本研究结果一致,但模拟增温后的狗尾草重要值显著降低,表明狗尾草等一年生草本对模拟增温后的土壤环境较敏感.在增温和水分胁迫的环境中,土壤养分因此受到的负面影响导致群落组成发生改变[34 ] .模拟增温后沙质草地群落多样性呈下降趋势,其中Simpson指数、Shannon-Wiener指数和物种丰富度指数显著下降,而Pielou指数下降趋势不显著(图3 ),这与之前的大部分研究结果一致[35 -37 ] .也有学者认为低幅增温对多样性产生促进作用,但当增温超过一定幅度时,促进作用会减弱甚至变成抑制作用[12 ] ,造成这种不一致的原因可能是物种组成以及研究区间的差异.郭轶瑞等[38 ] 的研究发现耐旱植物对增温的抵抗力较强,而喜湿植物在受水分限制时竞争能力减弱导致其比重降低.增温导致土壤含水量降低,同时,物种丰富度和多样性指数与土壤湿度呈显著正相关关系,表明在干旱半干旱区,多样性随水分胁迫强度的增加更加敏感. ...
科尔沁沙质草地不同恢复年限草本层群落结构及其与土壤理化性质的关系
1
2022
... 多样性与生产力的关系是反映群落结构及生态功能的关键指标[39 -40 ] .许多研究证实物种多样性与生产力主要存在正相关、负相关、单峰曲线和不相关4种关系[7 ] ,且正相关关系的结论居多.就科尔沁沙质草地而言,物种丰富度指数和生物量之间呈显著线性正相关,这与孙小丽等[7 ] 对内蒙古中温型荒漠草原区群落生物量随物种丰富度增加而增加的结论相一致,但与郭轶瑞等[41 ] 在科尔沁沙质草地的物种多样性与生产力呈单峰关系这一结论不同.极有可能是因为独特的生境特征差异.研究区处于干旱半干旱区,水热因子对干旱半干旱区植物群落生产力方面作用占主导地位[7 ,11 ] ,因此,推测科尔沁沙质草地生产力受到土壤水热因子的调控.本研究中,增温模拟后的群落和禾本科地上生物量与对照组相比显著下降(图4 ).Arnda等[42 ] 发现,禾本科植物地上生物量减少可能是因为更多杂草类根系在土壤剖面上的分配.也有研究表明,在干旱半干旱区,生产力深受水分限制[11 ] .Grime等[43 ] 发现,在增温和水分胁迫的环境中,植物通常以减少生物量生产来提高水分利用效率以此适应环境,解释了本研究在模拟增温后土壤湿度的降低导致了植物地上生产力的降低.由于水分限制,模拟增温后植物无法利用土壤温度升高的能量进行光合作用,从而呈随着能量增加群落多样性和生产力降低的关系.也有研究表明,增温对生物量的潜在影响不仅与土壤水热条件有关,土壤养分与多样性和生物量也存在显著相关的关系[34 ] .孙小丽等[7 ] 对草地生态系统的研究表明生物量与速效氮、有机碳等土壤养分呈显著的线性正相关.生物量分配还受到群落组成的影响,禾本科地上生物量减少为杂草类地下生物量提供了更多的水分和养分资源[42 ] . ...
气候变暖与管理方式对草地群落影响研究综述
1
2023
... 多样性与生产力的关系是反映群落结构及生态功能的关键指标[39 -40 ] .许多研究证实物种多样性与生产力主要存在正相关、负相关、单峰曲线和不相关4种关系[7 ] ,且正相关关系的结论居多.就科尔沁沙质草地而言,物种丰富度指数和生物量之间呈显著线性正相关,这与孙小丽等[7 ] 对内蒙古中温型荒漠草原区群落生物量随物种丰富度增加而增加的结论相一致,但与郭轶瑞等[41 ] 在科尔沁沙质草地的物种多样性与生产力呈单峰关系这一结论不同.极有可能是因为独特的生境特征差异.研究区处于干旱半干旱区,水热因子对干旱半干旱区植物群落生产力方面作用占主导地位[7 ,11 ] ,因此,推测科尔沁沙质草地生产力受到土壤水热因子的调控.本研究中,增温模拟后的群落和禾本科地上生物量与对照组相比显著下降(图4 ).Arnda等[42 ] 发现,禾本科植物地上生物量减少可能是因为更多杂草类根系在土壤剖面上的分配.也有研究表明,在干旱半干旱区,生产力深受水分限制[11 ] .Grime等[43 ] 发现,在增温和水分胁迫的环境中,植物通常以减少生物量生产来提高水分利用效率以此适应环境,解释了本研究在模拟增温后土壤湿度的降低导致了植物地上生产力的降低.由于水分限制,模拟增温后植物无法利用土壤温度升高的能量进行光合作用,从而呈随着能量增加群落多样性和生产力降低的关系.也有研究表明,增温对生物量的潜在影响不仅与土壤水热条件有关,土壤养分与多样性和生物量也存在显著相关的关系[34 ] .孙小丽等[7 ] 对草地生态系统的研究表明生物量与速效氮、有机碳等土壤养分呈显著的线性正相关.生物量分配还受到群落组成的影响,禾本科地上生物量减少为杂草类地下生物量提供了更多的水分和养分资源[42 ] . ...
科尔沁沙质草地物种多样性与生产力的关系
1
2007
... 多样性与生产力的关系是反映群落结构及生态功能的关键指标[39 -40 ] .许多研究证实物种多样性与生产力主要存在正相关、负相关、单峰曲线和不相关4种关系[7 ] ,且正相关关系的结论居多.就科尔沁沙质草地而言,物种丰富度指数和生物量之间呈显著线性正相关,这与孙小丽等[7 ] 对内蒙古中温型荒漠草原区群落生物量随物种丰富度增加而增加的结论相一致,但与郭轶瑞等[41 ] 在科尔沁沙质草地的物种多样性与生产力呈单峰关系这一结论不同.极有可能是因为独特的生境特征差异.研究区处于干旱半干旱区,水热因子对干旱半干旱区植物群落生产力方面作用占主导地位[7 ,11 ] ,因此,推测科尔沁沙质草地生产力受到土壤水热因子的调控.本研究中,增温模拟后的群落和禾本科地上生物量与对照组相比显著下降(图4 ).Arnda等[42 ] 发现,禾本科植物地上生物量减少可能是因为更多杂草类根系在土壤剖面上的分配.也有研究表明,在干旱半干旱区,生产力深受水分限制[11 ] .Grime等[43 ] 发现,在增温和水分胁迫的环境中,植物通常以减少生物量生产来提高水分利用效率以此适应环境,解释了本研究在模拟增温后土壤湿度的降低导致了植物地上生产力的降低.由于水分限制,模拟增温后植物无法利用土壤温度升高的能量进行光合作用,从而呈随着能量增加群落多样性和生产力降低的关系.也有研究表明,增温对生物量的潜在影响不仅与土壤水热条件有关,土壤养分与多样性和生物量也存在显著相关的关系[34 ] .孙小丽等[7 ] 对草地生态系统的研究表明生物量与速效氮、有机碳等土壤养分呈显著的线性正相关.生物量分配还受到群落组成的影响,禾本科地上生物量减少为杂草类地下生物量提供了更多的水分和养分资源[42 ] . ...
Fine root growth and vertical distribution in response to elevated CO2 ,warming and drought in a mixed heathland-grassland
2
2018
... 多样性与生产力的关系是反映群落结构及生态功能的关键指标[39 -40 ] .许多研究证实物种多样性与生产力主要存在正相关、负相关、单峰曲线和不相关4种关系[7 ] ,且正相关关系的结论居多.就科尔沁沙质草地而言,物种丰富度指数和生物量之间呈显著线性正相关,这与孙小丽等[7 ] 对内蒙古中温型荒漠草原区群落生物量随物种丰富度增加而增加的结论相一致,但与郭轶瑞等[41 ] 在科尔沁沙质草地的物种多样性与生产力呈单峰关系这一结论不同.极有可能是因为独特的生境特征差异.研究区处于干旱半干旱区,水热因子对干旱半干旱区植物群落生产力方面作用占主导地位[7 ,11 ] ,因此,推测科尔沁沙质草地生产力受到土壤水热因子的调控.本研究中,增温模拟后的群落和禾本科地上生物量与对照组相比显著下降(图4 ).Arnda等[42 ] 发现,禾本科植物地上生物量减少可能是因为更多杂草类根系在土壤剖面上的分配.也有研究表明,在干旱半干旱区,生产力深受水分限制[11 ] .Grime等[43 ] 发现,在增温和水分胁迫的环境中,植物通常以减少生物量生产来提高水分利用效率以此适应环境,解释了本研究在模拟增温后土壤湿度的降低导致了植物地上生产力的降低.由于水分限制,模拟增温后植物无法利用土壤温度升高的能量进行光合作用,从而呈随着能量增加群落多样性和生产力降低的关系.也有研究表明,增温对生物量的潜在影响不仅与土壤水热条件有关,土壤养分与多样性和生物量也存在显著相关的关系[34 ] .孙小丽等[7 ] 对草地生态系统的研究表明生物量与速效氮、有机碳等土壤养分呈显著的线性正相关.生物量分配还受到群落组成的影响,禾本科地上生物量减少为杂草类地下生物量提供了更多的水分和养分资源[42 ] . ...
... [42 ]. ...
2
2001
... 多样性与生产力的关系是反映群落结构及生态功能的关键指标[39 -40 ] .许多研究证实物种多样性与生产力主要存在正相关、负相关、单峰曲线和不相关4种关系[7 ] ,且正相关关系的结论居多.就科尔沁沙质草地而言,物种丰富度指数和生物量之间呈显著线性正相关,这与孙小丽等[7 ] 对内蒙古中温型荒漠草原区群落生物量随物种丰富度增加而增加的结论相一致,但与郭轶瑞等[41 ] 在科尔沁沙质草地的物种多样性与生产力呈单峰关系这一结论不同.极有可能是因为独特的生境特征差异.研究区处于干旱半干旱区,水热因子对干旱半干旱区植物群落生产力方面作用占主导地位[7 ,11 ] ,因此,推测科尔沁沙质草地生产力受到土壤水热因子的调控.本研究中,增温模拟后的群落和禾本科地上生物量与对照组相比显著下降(图4 ).Arnda等[42 ] 发现,禾本科植物地上生物量减少可能是因为更多杂草类根系在土壤剖面上的分配.也有研究表明,在干旱半干旱区,生产力深受水分限制[11 ] .Grime等[43 ] 发现,在增温和水分胁迫的环境中,植物通常以减少生物量生产来提高水分利用效率以此适应环境,解释了本研究在模拟增温后土壤湿度的降低导致了植物地上生产力的降低.由于水分限制,模拟增温后植物无法利用土壤温度升高的能量进行光合作用,从而呈随着能量增加群落多样性和生产力降低的关系.也有研究表明,增温对生物量的潜在影响不仅与土壤水热条件有关,土壤养分与多样性和生物量也存在显著相关的关系[34 ] .孙小丽等[7 ] 对草地生态系统的研究表明生物量与速效氮、有机碳等土壤养分呈显著的线性正相关.生物量分配还受到群落组成的影响,禾本科地上生物量减少为杂草类地下生物量提供了更多的水分和养分资源[42 ] . ...
... 增温会导致沙质草地群落种间关系发生变化.除了多样性和生产力可以衡量群落特征以外,群落网络结构的复杂性以及关键类群也可以评价其结构和功能[13 ,44 ] .共生网络分析表明,沙质草地群落共现模式在增温模拟后明显改变(图5 ).相比于沙质草地,模拟增温后群落中物种之间的互作网络的节点数、相互作用强度和平均连通度明显降低,表明群落复杂性降低;而聚类系数增加,表明群落的组织水平增强;网络连通性指数增加而模块化指数降低,表明群落复杂度增加而组织程度降低(表3 ).有研究表明,植物群落的组织程度和复杂度还受到放牧和退化梯度的影响:草甸退化越严重,组织程度越低、复杂度越高;模拟放牧使组织程度呈上升趋势、网络连通性呈降低趋势[13 ] .亦有研究表明,群落稳定性与物种丰富度指数有很高的相关性[45 ] .沙质草地在模拟增温后,群落的丰富度指数与网络结构稳定性均降低,与袁梓裕等[46 ] 的研究中群落稳定性和物种丰富度呈显著正相关这一结果相近,说明植物在受到增温和水分胁迫的共同作用下,植物物种之间的竞争加剧导致群落组成减少以及各物种之间的相互作用强度和模块化水平均有降低趋势[43 ,32 ] .关键类群是群落中高度连接的物种类群,强烈影响着群落中的其他物种及其结构和功能[47 ] .模拟增温后的沙质草地关键物种除旱生植物菊科的黄蒿外,还有藜科的猪毛菜和禾本科的糙隐子草.研究区本就水分匮乏,模拟增温又使土壤含水量减少,因此,关键物种的变化也可用与土壤水分相关的原因来解释:被确定为关键物种的植物可能在保持土壤水分方面发挥关键作用[14 ] . ...
The closing circle:nature,man,and technology
1
2015
... 增温会导致沙质草地群落种间关系发生变化.除了多样性和生产力可以衡量群落特征以外,群落网络结构的复杂性以及关键类群也可以评价其结构和功能[13 ,44 ] .共生网络分析表明,沙质草地群落共现模式在增温模拟后明显改变(图5 ).相比于沙质草地,模拟增温后群落中物种之间的互作网络的节点数、相互作用强度和平均连通度明显降低,表明群落复杂性降低;而聚类系数增加,表明群落的组织水平增强;网络连通性指数增加而模块化指数降低,表明群落复杂度增加而组织程度降低(表3 ).有研究表明,植物群落的组织程度和复杂度还受到放牧和退化梯度的影响:草甸退化越严重,组织程度越低、复杂度越高;模拟放牧使组织程度呈上升趋势、网络连通性呈降低趋势[13 ] .亦有研究表明,群落稳定性与物种丰富度指数有很高的相关性[45 ] .沙质草地在模拟增温后,群落的丰富度指数与网络结构稳定性均降低,与袁梓裕等[46 ] 的研究中群落稳定性和物种丰富度呈显著正相关这一结果相近,说明植物在受到增温和水分胁迫的共同作用下,植物物种之间的竞争加剧导致群落组成减少以及各物种之间的相互作用强度和模块化水平均有降低趋势[43 ,32 ] .关键类群是群落中高度连接的物种类群,强烈影响着群落中的其他物种及其结构和功能[47 ] .模拟增温后的沙质草地关键物种除旱生植物菊科的黄蒿外,还有藜科的猪毛菜和禾本科的糙隐子草.研究区本就水分匮乏,模拟增温又使土壤含水量减少,因此,关键物种的变化也可用与土壤水分相关的原因来解释:被确定为关键物种的植物可能在保持土壤水分方面发挥关键作用[14 ] . ...
半干旱区人工封育草场植物群落物种多样性与复杂性研究:以宁夏盐池为例
1
2014
... 增温会导致沙质草地群落种间关系发生变化.除了多样性和生产力可以衡量群落特征以外,群落网络结构的复杂性以及关键类群也可以评价其结构和功能[13 ,44 ] .共生网络分析表明,沙质草地群落共现模式在增温模拟后明显改变(图5 ).相比于沙质草地,模拟增温后群落中物种之间的互作网络的节点数、相互作用强度和平均连通度明显降低,表明群落复杂性降低;而聚类系数增加,表明群落的组织水平增强;网络连通性指数增加而模块化指数降低,表明群落复杂度增加而组织程度降低(表3 ).有研究表明,植物群落的组织程度和复杂度还受到放牧和退化梯度的影响:草甸退化越严重,组织程度越低、复杂度越高;模拟放牧使组织程度呈上升趋势、网络连通性呈降低趋势[13 ] .亦有研究表明,群落稳定性与物种丰富度指数有很高的相关性[45 ] .沙质草地在模拟增温后,群落的丰富度指数与网络结构稳定性均降低,与袁梓裕等[46 ] 的研究中群落稳定性和物种丰富度呈显著正相关这一结果相近,说明植物在受到增温和水分胁迫的共同作用下,植物物种之间的竞争加剧导致群落组成减少以及各物种之间的相互作用强度和模块化水平均有降低趋势[43 ,32 ] .关键类群是群落中高度连接的物种类群,强烈影响着群落中的其他物种及其结构和功能[47 ] .模拟增温后的沙质草地关键物种除旱生植物菊科的黄蒿外,还有藜科的猪毛菜和禾本科的糙隐子草.研究区本就水分匮乏,模拟增温又使土壤含水量减少,因此,关键物种的变化也可用与土壤水分相关的原因来解释:被确定为关键物种的植物可能在保持土壤水分方面发挥关键作用[14 ] . ...
黄土高原草地植物多样性与群落稳定性的关系及其驱动因素
1
2023
... 增温会导致沙质草地群落种间关系发生变化.除了多样性和生产力可以衡量群落特征以外,群落网络结构的复杂性以及关键类群也可以评价其结构和功能[13 ,44 ] .共生网络分析表明,沙质草地群落共现模式在增温模拟后明显改变(图5 ).相比于沙质草地,模拟增温后群落中物种之间的互作网络的节点数、相互作用强度和平均连通度明显降低,表明群落复杂性降低;而聚类系数增加,表明群落的组织水平增强;网络连通性指数增加而模块化指数降低,表明群落复杂度增加而组织程度降低(表3 ).有研究表明,植物群落的组织程度和复杂度还受到放牧和退化梯度的影响:草甸退化越严重,组织程度越低、复杂度越高;模拟放牧使组织程度呈上升趋势、网络连通性呈降低趋势[13 ] .亦有研究表明,群落稳定性与物种丰富度指数有很高的相关性[45 ] .沙质草地在模拟增温后,群落的丰富度指数与网络结构稳定性均降低,与袁梓裕等[46 ] 的研究中群落稳定性和物种丰富度呈显著正相关这一结果相近,说明植物在受到增温和水分胁迫的共同作用下,植物物种之间的竞争加剧导致群落组成减少以及各物种之间的相互作用强度和模块化水平均有降低趋势[43 ,32 ] .关键类群是群落中高度连接的物种类群,强烈影响着群落中的其他物种及其结构和功能[47 ] .模拟增温后的沙质草地关键物种除旱生植物菊科的黄蒿外,还有藜科的猪毛菜和禾本科的糙隐子草.研究区本就水分匮乏,模拟增温又使土壤含水量减少,因此,关键物种的变化也可用与土壤水分相关的原因来解释:被确定为关键物种的植物可能在保持土壤水分方面发挥关键作用[14 ] . ...
Molecular ecological network analyses:an effective conservation tool for the assessment of biodiversity,trophic interactions,and community structure
1
2020
... 增温会导致沙质草地群落种间关系发生变化.除了多样性和生产力可以衡量群落特征以外,群落网络结构的复杂性以及关键类群也可以评价其结构和功能[13 ,44 ] .共生网络分析表明,沙质草地群落共现模式在增温模拟后明显改变(图5 ).相比于沙质草地,模拟增温后群落中物种之间的互作网络的节点数、相互作用强度和平均连通度明显降低,表明群落复杂性降低;而聚类系数增加,表明群落的组织水平增强;网络连通性指数增加而模块化指数降低,表明群落复杂度增加而组织程度降低(表3 ).有研究表明,植物群落的组织程度和复杂度还受到放牧和退化梯度的影响:草甸退化越严重,组织程度越低、复杂度越高;模拟放牧使组织程度呈上升趋势、网络连通性呈降低趋势[13 ] .亦有研究表明,群落稳定性与物种丰富度指数有很高的相关性[45 ] .沙质草地在模拟增温后,群落的丰富度指数与网络结构稳定性均降低,与袁梓裕等[46 ] 的研究中群落稳定性和物种丰富度呈显著正相关这一结果相近,说明植物在受到增温和水分胁迫的共同作用下,植物物种之间的竞争加剧导致群落组成减少以及各物种之间的相互作用强度和模块化水平均有降低趋势[43 ,32 ] .关键类群是群落中高度连接的物种类群,强烈影响着群落中的其他物种及其结构和功能[47 ] .模拟增温后的沙质草地关键物种除旱生植物菊科的黄蒿外,还有藜科的猪毛菜和禾本科的糙隐子草.研究区本就水分匮乏,模拟增温又使土壤含水量减少,因此,关键物种的变化也可用与土壤水分相关的原因来解释:被确定为关键物种的植物可能在保持土壤水分方面发挥关键作用[14 ] . ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}