The mineral nutrition of wild plants revisited:a re-evaluation of processes and patterns
1
1999
... 氮素回收是植物将氮元素通过输导系统从衰败组织向活体贮存结构或新组织转移的生理过程,该过程有助于减缓植物在衰败过程中引起的氮损失,降低植物对环境中氮的依赖从而提高对于环境的适应性,对草地生态系统养分循环具有重要意义[1].在资源相对贫瘠的生态系统中较长时间的氮驻留有利于植物在资源获取和生存的过程中具备更强的竞争性,而较短时间氮驻留的植物则具有较低的生态适应性[2]. ...
Nutrient resorption from senescing leaves in two Stipa species native to central Argentina
1
2003
... 氮素回收是植物将氮元素通过输导系统从衰败组织向活体贮存结构或新组织转移的生理过程,该过程有助于减缓植物在衰败过程中引起的氮损失,降低植物对环境中氮的依赖从而提高对于环境的适应性,对草地生态系统养分循环具有重要意义[1].在资源相对贫瘠的生态系统中较长时间的氮驻留有利于植物在资源获取和生存的过程中具备更强的竞争性,而较短时间氮驻留的植物则具有较低的生态适应性[2]. ...
Climate Change 2013:The Physical Science Basis.Contribution of Working Group I to the Fifth Assessment Report of the Inter-governmental Panel on Climate Change
1
2013
... 全球气候变暖导致极端天气气候事件频繁发生,主要表现为中低纬度降水量不断增加、降水分布和格局发生了重大变化,近年来以降水间隔时间增长、单次降水量增大为特征的极端降水事件频率持续增加[3-4],降水变化可以在较短时间尺度上改变植物生理代谢反应从而影响氮素的可利用性[5-6].其主要原因一方面是水分能够影响植物气孔导度,影响光合作用的同时改变氮利用效率,另一方面是降水通过促进土壤微生物活性,刺激土壤氮矿化作用并间接改善土壤氮可利用性,从而提高植物叶片氮含量,降低养分回收能力[7-8].降水梯度变化对养分回收影响的研究表明,随着降水湿润端向干旱端变化,氮回收效率(NRE)逐渐升高[9-10].然而也有研究发现增加降水降低了土壤氮的有效性,其主要原因是增加降水提高了地上生产力、土壤有机质和微生物总量,导致氮固持能力增加,从而降低土壤中的可利用氮[11-12].因此植物叶片NRE对水分胁迫的响应在不同生态系统中可能表现出不一致性. ...
Enhanced precipitation variability effects on water losses and ecosystem functioning: differential response of arid and mesic regions
1
2015
... 全球气候变暖导致极端天气气候事件频繁发生,主要表现为中低纬度降水量不断增加、降水分布和格局发生了重大变化,近年来以降水间隔时间增长、单次降水量增大为特征的极端降水事件频率持续增加[3-4],降水变化可以在较短时间尺度上改变植物生理代谢反应从而影响氮素的可利用性[5-6].其主要原因一方面是水分能够影响植物气孔导度,影响光合作用的同时改变氮利用效率,另一方面是降水通过促进土壤微生物活性,刺激土壤氮矿化作用并间接改善土壤氮可利用性,从而提高植物叶片氮含量,降低养分回收能力[7-8].降水梯度变化对养分回收影响的研究表明,随着降水湿润端向干旱端变化,氮回收效率(NRE)逐渐升高[9-10].然而也有研究发现增加降水降低了土壤氮的有效性,其主要原因是增加降水提高了地上生产力、土壤有机质和微生物总量,导致氮固持能力增加,从而降低土壤中的可利用氮[11-12].因此植物叶片NRE对水分胁迫的响应在不同生态系统中可能表现出不一致性. ...
Water,nitrogen and phosphorus use efficiencies of four tree species in response to variable water and nutrient supply
1
2016
... 全球气候变暖导致极端天气气候事件频繁发生,主要表现为中低纬度降水量不断增加、降水分布和格局发生了重大变化,近年来以降水间隔时间增长、单次降水量增大为特征的极端降水事件频率持续增加[3-4],降水变化可以在较短时间尺度上改变植物生理代谢反应从而影响氮素的可利用性[5-6].其主要原因一方面是水分能够影响植物气孔导度,影响光合作用的同时改变氮利用效率,另一方面是降水通过促进土壤微生物活性,刺激土壤氮矿化作用并间接改善土壤氮可利用性,从而提高植物叶片氮含量,降低养分回收能力[7-8].降水梯度变化对养分回收影响的研究表明,随着降水湿润端向干旱端变化,氮回收效率(NRE)逐渐升高[9-10].然而也有研究发现增加降水降低了土壤氮的有效性,其主要原因是增加降水提高了地上生产力、土壤有机质和微生物总量,导致氮固持能力增加,从而降低土壤中的可利用氮[11-12].因此植物叶片NRE对水分胁迫的响应在不同生态系统中可能表现出不一致性. ...
Higher precipitation strengthens the microbial interactions in semi-arid grassland soils
1
2018
... 全球气候变暖导致极端天气气候事件频繁发生,主要表现为中低纬度降水量不断增加、降水分布和格局发生了重大变化,近年来以降水间隔时间增长、单次降水量增大为特征的极端降水事件频率持续增加[3-4],降水变化可以在较短时间尺度上改变植物生理代谢反应从而影响氮素的可利用性[5-6].其主要原因一方面是水分能够影响植物气孔导度,影响光合作用的同时改变氮利用效率,另一方面是降水通过促进土壤微生物活性,刺激土壤氮矿化作用并间接改善土壤氮可利用性,从而提高植物叶片氮含量,降低养分回收能力[7-8].降水梯度变化对养分回收影响的研究表明,随着降水湿润端向干旱端变化,氮回收效率(NRE)逐渐升高[9-10].然而也有研究发现增加降水降低了土壤氮的有效性,其主要原因是增加降水提高了地上生产力、土壤有机质和微生物总量,导致氮固持能力增加,从而降低土壤中的可利用氮[11-12].因此植物叶片NRE对水分胁迫的响应在不同生态系统中可能表现出不一致性. ...
Effects of plant functional types,climate and soil nitrogen on leaf nitrogen along the north-south transect of eastern China
1
2013
... 全球气候变暖导致极端天气气候事件频繁发生,主要表现为中低纬度降水量不断增加、降水分布和格局发生了重大变化,近年来以降水间隔时间增长、单次降水量增大为特征的极端降水事件频率持续增加[3-4],降水变化可以在较短时间尺度上改变植物生理代谢反应从而影响氮素的可利用性[5-6].其主要原因一方面是水分能够影响植物气孔导度,影响光合作用的同时改变氮利用效率,另一方面是降水通过促进土壤微生物活性,刺激土壤氮矿化作用并间接改善土壤氮可利用性,从而提高植物叶片氮含量,降低养分回收能力[7-8].降水梯度变化对养分回收影响的研究表明,随着降水湿润端向干旱端变化,氮回收效率(NRE)逐渐升高[9-10].然而也有研究发现增加降水降低了土壤氮的有效性,其主要原因是增加降水提高了地上生产力、土壤有机质和微生物总量,导致氮固持能力增加,从而降低土壤中的可利用氮[11-12].因此植物叶片NRE对水分胁迫的响应在不同生态系统中可能表现出不一致性. ...
Convergent responses of nitrogen and phosphorus resorption to nitrogen inputs in a semi-arid grassland
1
2013
... 全球气候变暖导致极端天气气候事件频繁发生,主要表现为中低纬度降水量不断增加、降水分布和格局发生了重大变化,近年来以降水间隔时间增长、单次降水量增大为特征的极端降水事件频率持续增加[3-4],降水变化可以在较短时间尺度上改变植物生理代谢反应从而影响氮素的可利用性[5-6].其主要原因一方面是水分能够影响植物气孔导度,影响光合作用的同时改变氮利用效率,另一方面是降水通过促进土壤微生物活性,刺激土壤氮矿化作用并间接改善土壤氮可利用性,从而提高植物叶片氮含量,降低养分回收能力[7-8].降水梯度变化对养分回收影响的研究表明,随着降水湿润端向干旱端变化,氮回收效率(NRE)逐渐升高[9-10].然而也有研究发现增加降水降低了土壤氮的有效性,其主要原因是增加降水提高了地上生产力、土壤有机质和微生物总量,导致氮固持能力增加,从而降低土壤中的可利用氮[11-12].因此植物叶片NRE对水分胁迫的响应在不同生态系统中可能表现出不一致性. ...
Nutrient dynamics along a precipitation gradient in European beech forests
1
2014
... 全球气候变暖导致极端天气气候事件频繁发生,主要表现为中低纬度降水量不断增加、降水分布和格局发生了重大变化,近年来以降水间隔时间增长、单次降水量增大为特征的极端降水事件频率持续增加[3-4],降水变化可以在较短时间尺度上改变植物生理代谢反应从而影响氮素的可利用性[5-6].其主要原因一方面是水分能够影响植物气孔导度,影响光合作用的同时改变氮利用效率,另一方面是降水通过促进土壤微生物活性,刺激土壤氮矿化作用并间接改善土壤氮可利用性,从而提高植物叶片氮含量,降低养分回收能力[7-8].降水梯度变化对养分回收影响的研究表明,随着降水湿润端向干旱端变化,氮回收效率(NRE)逐渐升高[9-10].然而也有研究发现增加降水降低了土壤氮的有效性,其主要原因是增加降水提高了地上生产力、土壤有机质和微生物总量,导致氮固持能力增加,从而降低土壤中的可利用氮[11-12].因此植物叶片NRE对水分胁迫的响应在不同生态系统中可能表现出不一致性. ...
羌塘高原降水梯度带紫花针茅叶片氮回收特征及影响因素
1
2016
... 全球气候变暖导致极端天气气候事件频繁发生,主要表现为中低纬度降水量不断增加、降水分布和格局发生了重大变化,近年来以降水间隔时间增长、单次降水量增大为特征的极端降水事件频率持续增加[3-4],降水变化可以在较短时间尺度上改变植物生理代谢反应从而影响氮素的可利用性[5-6].其主要原因一方面是水分能够影响植物气孔导度,影响光合作用的同时改变氮利用效率,另一方面是降水通过促进土壤微生物活性,刺激土壤氮矿化作用并间接改善土壤氮可利用性,从而提高植物叶片氮含量,降低养分回收能力[7-8].降水梯度变化对养分回收影响的研究表明,随着降水湿润端向干旱端变化,氮回收效率(NRE)逐渐升高[9-10].然而也有研究发现增加降水降低了土壤氮的有效性,其主要原因是增加降水提高了地上生产力、土壤有机质和微生物总量,导致氮固持能力增加,从而降低土壤中的可利用氮[11-12].因此植物叶片NRE对水分胁迫的响应在不同生态系统中可能表现出不一致性. ...
降雨、温度和氮沉降增加对新疆温带荒漠生态系统氮素去向和植物化学计量特征的影响
1
2018
... 全球气候变暖导致极端天气气候事件频繁发生,主要表现为中低纬度降水量不断增加、降水分布和格局发生了重大变化,近年来以降水间隔时间增长、单次降水量增大为特征的极端降水事件频率持续增加[3-4],降水变化可以在较短时间尺度上改变植物生理代谢反应从而影响氮素的可利用性[5-6].其主要原因一方面是水分能够影响植物气孔导度,影响光合作用的同时改变氮利用效率,另一方面是降水通过促进土壤微生物活性,刺激土壤氮矿化作用并间接改善土壤氮可利用性,从而提高植物叶片氮含量,降低养分回收能力[7-8].降水梯度变化对养分回收影响的研究表明,随着降水湿润端向干旱端变化,氮回收效率(NRE)逐渐升高[9-10].然而也有研究发现增加降水降低了土壤氮的有效性,其主要原因是增加降水提高了地上生产力、土壤有机质和微生物总量,导致氮固持能力增加,从而降低土壤中的可利用氮[11-12].因此植物叶片NRE对水分胁迫的响应在不同生态系统中可能表现出不一致性. ...
Plant functional groups mediate drought resistance and recovery in a multisite grassland experiment
1
2019
... 全球气候变暖导致极端天气气候事件频繁发生,主要表现为中低纬度降水量不断增加、降水分布和格局发生了重大变化,近年来以降水间隔时间增长、单次降水量增大为特征的极端降水事件频率持续增加[3-4],降水变化可以在较短时间尺度上改变植物生理代谢反应从而影响氮素的可利用性[5-6].其主要原因一方面是水分能够影响植物气孔导度,影响光合作用的同时改变氮利用效率,另一方面是降水通过促进土壤微生物活性,刺激土壤氮矿化作用并间接改善土壤氮可利用性,从而提高植物叶片氮含量,降低养分回收能力[7-8].降水梯度变化对养分回收影响的研究表明,随着降水湿润端向干旱端变化,氮回收效率(NRE)逐渐升高[9-10].然而也有研究发现增加降水降低了土壤氮的有效性,其主要原因是增加降水提高了地上生产力、土壤有机质和微生物总量,导致氮固持能力增加,从而降低土壤中的可利用氮[11-12].因此植物叶片NRE对水分胁迫的响应在不同生态系统中可能表现出不一致性. ...
Nitrogen cycles: past,present,and future
1
2005
... 化石燃料燃烧和人类活动导致地球表面的氮沉降量在过去100多年间急剧增长[13],并且在未来几十年内会持续增加,这将极大改变植物-土壤间的互馈关系以及养分循环特征[14].养分回收作为植物内部养分循环机制,常用来指示植物养分限制状况[15].全球456种陆地植物对氮添加响应整合分析表明氮添加导致土壤中可利用氮增加,缓解植物生长过程中氮限制,可以使植物绿叶期叶片平均氮含量提高28.5%,从而提高NRE[16],然而也有研究表明氮添加能够有效提高枯叶的叶片氮含量,从而降低NRE[17],由此说明外源养分供应对生态系统养分循环具有调节作用且NRE与土壤养分之间的关系存在不一致性[18].此外不同物种或者不同生活型的植物在叶片凋落阶段NRE差异明显[19-20],这是因为植物叶片中未被回收的养分以凋落物的形式转移到土壤中继续分解和矿化,被植物再次利用,因此NRE也会通过调控不同生活型植物凋落物的氮含量来调节植被-土壤系统的养分循环[21]. ...
The C∶N∶P stoichiometry of organisms and ecosystems in a changing world:a review and perspectives
1
2012
... 化石燃料燃烧和人类活动导致地球表面的氮沉降量在过去100多年间急剧增长[13],并且在未来几十年内会持续增加,这将极大改变植物-土壤间的互馈关系以及养分循环特征[14].养分回收作为植物内部养分循环机制,常用来指示植物养分限制状况[15].全球456种陆地植物对氮添加响应整合分析表明氮添加导致土壤中可利用氮增加,缓解植物生长过程中氮限制,可以使植物绿叶期叶片平均氮含量提高28.5%,从而提高NRE[16],然而也有研究表明氮添加能够有效提高枯叶的叶片氮含量,从而降低NRE[17],由此说明外源养分供应对生态系统养分循环具有调节作用且NRE与土壤养分之间的关系存在不一致性[18].此外不同物种或者不同生活型的植物在叶片凋落阶段NRE差异明显[19-20],这是因为植物叶片中未被回收的养分以凋落物的形式转移到土壤中继续分解和矿化,被植物再次利用,因此NRE也会通过调控不同生活型植物凋落物的氮含量来调节植被-土壤系统的养分循环[21]. ...
增温和氮素添加降低荒漠草原多年生植物氮素回收效率
1
2015
... 化石燃料燃烧和人类活动导致地球表面的氮沉降量在过去100多年间急剧增长[13],并且在未来几十年内会持续增加,这将极大改变植物-土壤间的互馈关系以及养分循环特征[14].养分回收作为植物内部养分循环机制,常用来指示植物养分限制状况[15].全球456种陆地植物对氮添加响应整合分析表明氮添加导致土壤中可利用氮增加,缓解植物生长过程中氮限制,可以使植物绿叶期叶片平均氮含量提高28.5%,从而提高NRE[16],然而也有研究表明氮添加能够有效提高枯叶的叶片氮含量,从而降低NRE[17],由此说明外源养分供应对生态系统养分循环具有调节作用且NRE与土壤养分之间的关系存在不一致性[18].此外不同物种或者不同生活型的植物在叶片凋落阶段NRE差异明显[19-20],这是因为植物叶片中未被回收的养分以凋落物的形式转移到土壤中继续分解和矿化,被植物再次利用,因此NRE也会通过调控不同生活型植物凋落物的氮含量来调节植被-土壤系统的养分循环[21]. ...
Negative effects of fertilization on plant nutrient resorption
1
2015
... 化石燃料燃烧和人类活动导致地球表面的氮沉降量在过去100多年间急剧增长[13],并且在未来几十年内会持续增加,这将极大改变植物-土壤间的互馈关系以及养分循环特征[14].养分回收作为植物内部养分循环机制,常用来指示植物养分限制状况[15].全球456种陆地植物对氮添加响应整合分析表明氮添加导致土壤中可利用氮增加,缓解植物生长过程中氮限制,可以使植物绿叶期叶片平均氮含量提高28.5%,从而提高NRE[16],然而也有研究表明氮添加能够有效提高枯叶的叶片氮含量,从而降低NRE[17],由此说明外源养分供应对生态系统养分循环具有调节作用且NRE与土壤养分之间的关系存在不一致性[18].此外不同物种或者不同生活型的植物在叶片凋落阶段NRE差异明显[19-20],这是因为植物叶片中未被回收的养分以凋落物的形式转移到土壤中继续分解和矿化,被植物再次利用,因此NRE也会通过调控不同生活型植物凋落物的氮含量来调节植被-土壤系统的养分循环[21]. ...
Nutrient resorption responses to water and nitrogen amendment in semi-arid grassland of Inner Mongolia,China
1
2010
... 化石燃料燃烧和人类活动导致地球表面的氮沉降量在过去100多年间急剧增长[13],并且在未来几十年内会持续增加,这将极大改变植物-土壤间的互馈关系以及养分循环特征[14].养分回收作为植物内部养分循环机制,常用来指示植物养分限制状况[15].全球456种陆地植物对氮添加响应整合分析表明氮添加导致土壤中可利用氮增加,缓解植物生长过程中氮限制,可以使植物绿叶期叶片平均氮含量提高28.5%,从而提高NRE[16],然而也有研究表明氮添加能够有效提高枯叶的叶片氮含量,从而降低NRE[17],由此说明外源养分供应对生态系统养分循环具有调节作用且NRE与土壤养分之间的关系存在不一致性[18].此外不同物种或者不同生活型的植物在叶片凋落阶段NRE差异明显[19-20],这是因为植物叶片中未被回收的养分以凋落物的形式转移到土壤中继续分解和矿化,被植物再次利用,因此NRE也会通过调控不同生活型植物凋落物的氮含量来调节植被-土壤系统的养分循环[21]. ...
Global response patterns of terrestrial plant species to nitrogen addition
1
2008
... 化石燃料燃烧和人类活动导致地球表面的氮沉降量在过去100多年间急剧增长[13],并且在未来几十年内会持续增加,这将极大改变植物-土壤间的互馈关系以及养分循环特征[14].养分回收作为植物内部养分循环机制,常用来指示植物养分限制状况[15].全球456种陆地植物对氮添加响应整合分析表明氮添加导致土壤中可利用氮增加,缓解植物生长过程中氮限制,可以使植物绿叶期叶片平均氮含量提高28.5%,从而提高NRE[16],然而也有研究表明氮添加能够有效提高枯叶的叶片氮含量,从而降低NRE[17],由此说明外源养分供应对生态系统养分循环具有调节作用且NRE与土壤养分之间的关系存在不一致性[18].此外不同物种或者不同生活型的植物在叶片凋落阶段NRE差异明显[19-20],这是因为植物叶片中未被回收的养分以凋落物的形式转移到土壤中继续分解和矿化,被植物再次利用,因此NRE也会通过调控不同生活型植物凋落物的氮含量来调节植被-土壤系统的养分循环[21]. ...
Nutrient resorption patterns of plant functional groups in a tropical Savanna: variation and functional significance
1
2008
... 化石燃料燃烧和人类活动导致地球表面的氮沉降量在过去100多年间急剧增长[13],并且在未来几十年内会持续增加,这将极大改变植物-土壤间的互馈关系以及养分循环特征[14].养分回收作为植物内部养分循环机制,常用来指示植物养分限制状况[15].全球456种陆地植物对氮添加响应整合分析表明氮添加导致土壤中可利用氮增加,缓解植物生长过程中氮限制,可以使植物绿叶期叶片平均氮含量提高28.5%,从而提高NRE[16],然而也有研究表明氮添加能够有效提高枯叶的叶片氮含量,从而降低NRE[17],由此说明外源养分供应对生态系统养分循环具有调节作用且NRE与土壤养分之间的关系存在不一致性[18].此外不同物种或者不同生活型的植物在叶片凋落阶段NRE差异明显[19-20],这是因为植物叶片中未被回收的养分以凋落物的形式转移到土壤中继续分解和矿化,被植物再次利用,因此NRE也会通过调控不同生活型植物凋落物的氮含量来调节植被-土壤系统的养分循环[21]. ...
Nitrogen resorption from senescing leaves in 28 plant species in a semi-arid region of northern China
1
2005
... 化石燃料燃烧和人类活动导致地球表面的氮沉降量在过去100多年间急剧增长[13],并且在未来几十年内会持续增加,这将极大改变植物-土壤间的互馈关系以及养分循环特征[14].养分回收作为植物内部养分循环机制,常用来指示植物养分限制状况[15].全球456种陆地植物对氮添加响应整合分析表明氮添加导致土壤中可利用氮增加,缓解植物生长过程中氮限制,可以使植物绿叶期叶片平均氮含量提高28.5%,从而提高NRE[16],然而也有研究表明氮添加能够有效提高枯叶的叶片氮含量,从而降低NRE[17],由此说明外源养分供应对生态系统养分循环具有调节作用且NRE与土壤养分之间的关系存在不一致性[18].此外不同物种或者不同生活型的植物在叶片凋落阶段NRE差异明显[19-20],这是因为植物叶片中未被回收的养分以凋落物的形式转移到土壤中继续分解和矿化,被植物再次利用,因此NRE也会通过调控不同生活型植物凋落物的氮含量来调节植被-土壤系统的养分循环[21]. ...
Differential responses of litter decomposition to increased soil nutrients and water between two contrasting grassland plant species of Inner Mongolia,China
1
2006
... 化石燃料燃烧和人类活动导致地球表面的氮沉降量在过去100多年间急剧增长[13],并且在未来几十年内会持续增加,这将极大改变植物-土壤间的互馈关系以及养分循环特征[14].养分回收作为植物内部养分循环机制,常用来指示植物养分限制状况[15].全球456种陆地植物对氮添加响应整合分析表明氮添加导致土壤中可利用氮增加,缓解植物生长过程中氮限制,可以使植物绿叶期叶片平均氮含量提高28.5%,从而提高NRE[16],然而也有研究表明氮添加能够有效提高枯叶的叶片氮含量,从而降低NRE[17],由此说明外源养分供应对生态系统养分循环具有调节作用且NRE与土壤养分之间的关系存在不一致性[18].此外不同物种或者不同生活型的植物在叶片凋落阶段NRE差异明显[19-20],这是因为植物叶片中未被回收的养分以凋落物的形式转移到土壤中继续分解和矿化,被植物再次利用,因此NRE也会通过调控不同生活型植物凋落物的氮含量来调节植被-土壤系统的养分循环[21]. ...
Species richness mediates within species nutrient resorption: implications for the biodiversity-productivity relationship
1
2019
... 内蒙古半干旱草原区对气候变化和人类活动的干扰十分敏感,水分和氮素是该区域影响植物生长最为重要的限制性环境影响因子[22],在全球气候变化和人类活动加剧背景下,分析该区域草地优势植物成熟绿叶和枯叶氮含量变化特征及影响因素,对于预测中国半干旱草原区生态系统养分循环规律及对未来全球变化的响应与反馈具有重要的生态学意义[23].近50年来,位于内蒙古东部的科尔沁沙地在剧烈气候变化和人类活动的影响下,草地生态环境不断恶化且生产力锐减,成为沙漠化典型区域[24].目前有关该区域沙质草地植物叶片养分回收状况在水分和养分变化下如何响应的研究较少.水分和养分变化如何通过影响土壤水分和养分有效性,从而影响沙质草地优势植物成熟绿叶和枯叶氮含量,进而影响NRE?基于以上问题,本研究通过调查科尔沁沙地沙质草地土壤水分和土壤全氮及4种优势植物的生物量、成熟叶和枯叶的叶片氮含量及植物叶片NRE,解析降水变化和氮沉降下土壤水分和养分对植物叶片NRE的调控机制,为阐释中国半干旱区植物适应环境变化的养分利用策略提供理论依据. ...
Plant nitrogen concentration,use efficiency,and contents in a tallgrass prairie ecosystem under experimental warming
1
2005
... 内蒙古半干旱草原区对气候变化和人类活动的干扰十分敏感,水分和氮素是该区域影响植物生长最为重要的限制性环境影响因子[22],在全球气候变化和人类活动加剧背景下,分析该区域草地优势植物成熟绿叶和枯叶氮含量变化特征及影响因素,对于预测中国半干旱草原区生态系统养分循环规律及对未来全球变化的响应与反馈具有重要的生态学意义[23].近50年来,位于内蒙古东部的科尔沁沙地在剧烈气候变化和人类活动的影响下,草地生态环境不断恶化且生产力锐减,成为沙漠化典型区域[24].目前有关该区域沙质草地植物叶片养分回收状况在水分和养分变化下如何响应的研究较少.水分和养分变化如何通过影响土壤水分和养分有效性,从而影响沙质草地优势植物成熟绿叶和枯叶氮含量,进而影响NRE?基于以上问题,本研究通过调查科尔沁沙地沙质草地土壤水分和土壤全氮及4种优势植物的生物量、成熟叶和枯叶的叶片氮含量及植物叶片NRE,解析降水变化和氮沉降下土壤水分和养分对植物叶片NRE的调控机制,为阐释中国半干旱区植物适应环境变化的养分利用策略提供理论依据. ...
科尔沁地区现代沙漠化过程的驱动因素分析
1
2004
... 内蒙古半干旱草原区对气候变化和人类活动的干扰十分敏感,水分和氮素是该区域影响植物生长最为重要的限制性环境影响因子[22],在全球气候变化和人类活动加剧背景下,分析该区域草地优势植物成熟绿叶和枯叶氮含量变化特征及影响因素,对于预测中国半干旱草原区生态系统养分循环规律及对未来全球变化的响应与反馈具有重要的生态学意义[23].近50年来,位于内蒙古东部的科尔沁沙地在剧烈气候变化和人类活动的影响下,草地生态环境不断恶化且生产力锐减,成为沙漠化典型区域[24].目前有关该区域沙质草地植物叶片养分回收状况在水分和养分变化下如何响应的研究较少.水分和养分变化如何通过影响土壤水分和养分有效性,从而影响沙质草地优势植物成熟绿叶和枯叶氮含量,进而影响NRE?基于以上问题,本研究通过调查科尔沁沙地沙质草地土壤水分和土壤全氮及4种优势植物的生物量、成熟叶和枯叶的叶片氮含量及植物叶片NRE,解析降水变化和氮沉降下土壤水分和养分对植物叶片NRE的调控机制,为阐释中国半干旱区植物适应环境变化的养分利用策略提供理论依据. ...
Spatial and temporal variability of drought during 1960-2012 in Inner Mongolia,north China
1
2015
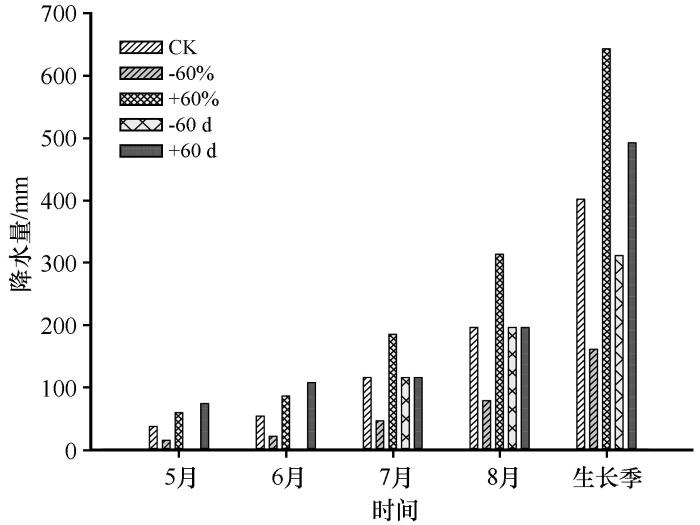
... 2017年8月,选取相对均匀、退化程度不高且长期围封的沙质草地,该样地中土壤全碳含量平均值为3.96 g·kg-1,土壤全氮平均值为0.42 g·kg-1.基于该地区生长季年总降水量、极端干旱和极端降水事件的长期观测数据[25],设置4个降水处理:整个生长季(5—8月)增加或减少降水60%(±60%),生长季前期(5—6月)增加或减少降水100%(±60 d).于2017年4月20日布设增减水装置,2017年10月20日拆除,该装置具有成本低及对气候变化影响甚微的特点,广泛应用于气候变化的研究[26-27].科尔沁沙地大规模的人类活动(农田施肥和畜牧业)加剧了氮输入,导致该地区氮负荷过高[28],参考世界上其他国家(美国、欧洲)的氮沉降水平,添加20 g·m-2N是中国区域较高的氮沉降水平[29].因此分别在每年5月和7月各添加10 g·m-2N,总计20 g·m-2N.本试验拟采取双因素随机区组设计,选取24个8 m×8 m的大样方,再从中随机选取12个样方开展增减水试验,每个增减水样方的面积为2.5 m×2.5 m,10个处理,6个重复,总计60个样方. ...
一种野外增减雨试验装置
1
... 2017年8月,选取相对均匀、退化程度不高且长期围封的沙质草地,该样地中土壤全碳含量平均值为3.96 g·kg-1,土壤全氮平均值为0.42 g·kg-1.基于该地区生长季年总降水量、极端干旱和极端降水事件的长期观测数据[25],设置4个降水处理:整个生长季(5—8月)增加或减少降水60%(±60%),生长季前期(5—6月)增加或减少降水100%(±60 d).于2017年4月20日布设增减水装置,2017年10月20日拆除,该装置具有成本低及对气候变化影响甚微的特点,广泛应用于气候变化的研究[26-27].科尔沁沙地大规模的人类活动(农田施肥和畜牧业)加剧了氮输入,导致该地区氮负荷过高[28],参考世界上其他国家(美国、欧洲)的氮沉降水平,添加20 g·m-2N是中国区域较高的氮沉降水平[29].因此分别在每年5月和7月各添加10 g·m-2N,总计20 g·m-2N.本试验拟采取双因素随机区组设计,选取24个8 m×8 m的大样方,再从中随机选取12个样方开展增减水试验,每个增减水样方的面积为2.5 m×2.5 m,10个处理,6个重复,总计60个样方. ...
Effects of Grazing,Extreme drought,extreme rainfall and nitrogen addition on vegetation characteristics and productivity of semiarid grassland
1
2023
... 2017年8月,选取相对均匀、退化程度不高且长期围封的沙质草地,该样地中土壤全碳含量平均值为3.96 g·kg-1,土壤全氮平均值为0.42 g·kg-1.基于该地区生长季年总降水量、极端干旱和极端降水事件的长期观测数据[25],设置4个降水处理:整个生长季(5—8月)增加或减少降水60%(±60%),生长季前期(5—6月)增加或减少降水100%(±60 d).于2017年4月20日布设增减水装置,2017年10月20日拆除,该装置具有成本低及对气候变化影响甚微的特点,广泛应用于气候变化的研究[26-27].科尔沁沙地大规模的人类活动(农田施肥和畜牧业)加剧了氮输入,导致该地区氮负荷过高[28],参考世界上其他国家(美国、欧洲)的氮沉降水平,添加20 g·m-2N是中国区域较高的氮沉降水平[29].因此分别在每年5月和7月各添加10 g·m-2N,总计20 g·m-2N.本试验拟采取双因素随机区组设计,选取24个8 m×8 m的大样方,再从中随机选取12个样方开展增减水试验,每个增减水样方的面积为2.5 m×2.5 m,10个处理,6个重复,总计60个样方. ...
氮素添加对科尔沁沙质草地物种多样性和生产力的影响
1
2009
... 2017年8月,选取相对均匀、退化程度不高且长期围封的沙质草地,该样地中土壤全碳含量平均值为3.96 g·kg-1,土壤全氮平均值为0.42 g·kg-1.基于该地区生长季年总降水量、极端干旱和极端降水事件的长期观测数据[25],设置4个降水处理:整个生长季(5—8月)增加或减少降水60%(±60%),生长季前期(5—6月)增加或减少降水100%(±60 d).于2017年4月20日布设增减水装置,2017年10月20日拆除,该装置具有成本低及对气候变化影响甚微的特点,广泛应用于气候变化的研究[26-27].科尔沁沙地大规模的人类活动(农田施肥和畜牧业)加剧了氮输入,导致该地区氮负荷过高[28],参考世界上其他国家(美国、欧洲)的氮沉降水平,添加20 g·m-2N是中国区域较高的氮沉降水平[29].因此分别在每年5月和7月各添加10 g·m-2N,总计20 g·m-2N.本试验拟采取双因素随机区组设计,选取24个8 m×8 m的大样方,再从中随机选取12个样方开展增减水试验,每个增减水样方的面积为2.5 m×2.5 m,10个处理,6个重复,总计60个样方. ...
Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands
1
2008
... 2017年8月,选取相对均匀、退化程度不高且长期围封的沙质草地,该样地中土壤全碳含量平均值为3.96 g·kg-1,土壤全氮平均值为0.42 g·kg-1.基于该地区生长季年总降水量、极端干旱和极端降水事件的长期观测数据[25],设置4个降水处理:整个生长季(5—8月)增加或减少降水60%(±60%),生长季前期(5—6月)增加或减少降水100%(±60 d).于2017年4月20日布设增减水装置,2017年10月20日拆除,该装置具有成本低及对气候变化影响甚微的特点,广泛应用于气候变化的研究[26-27].科尔沁沙地大规模的人类活动(农田施肥和畜牧业)加剧了氮输入,导致该地区氮负荷过高[28],参考世界上其他国家(美国、欧洲)的氮沉降水平,添加20 g·m-2N是中国区域较高的氮沉降水平[29].因此分别在每年5月和7月各添加10 g·m-2N,总计20 g·m-2N.本试验拟采取双因素随机区组设计,选取24个8 m×8 m的大样方,再从中随机选取12个样方开展增减水试验,每个增减水样方的面积为2.5 m×2.5 m,10个处理,6个重复,总计60个样方. ...
全球降水格局变化下土壤氮循环研究进展
1
2020
... 降水变化会影响土壤的物理、化学性质和微生物特性,从而影响土壤持水性和养分含量[30-32].本研究中,生长季减水60%显著降低了土壤水分,其主要原因可能是沙质土壤颗粒较大,土壤饱和导水率高且蒸发量大,较低的持水力在长期干旱条件下土壤水分长效匮乏[31],这表明土壤水分除了受到降水的影响,还受到温度、地形和水文条件等多因素的影响.研究表明,降水强度变化会对植物功能群多样性和生产力产生积极或消极的影响[33].本研究中降水变化对糙隐子草和狗尾草的生物量产生了波动式影响,而对黄蒿和白草的生物量无显著影响,其原因主要是不同的降水强度与不同氮添加组合引起了物种组成和结构的变化,当沙质草地的水分和养分状况发生改变时,竞争力强的物种会打破种间平衡,各个物种的优势地位发生变化引起了相应生物量的改变[34-35]. ...
Plant responses to warming and increased precipitation in three categories of dune stabilization in northeastern China
1
2017
... 降水变化会影响土壤的物理、化学性质和微生物特性,从而影响土壤持水性和养分含量[30-32].本研究中,生长季减水60%显著降低了土壤水分,其主要原因可能是沙质土壤颗粒较大,土壤饱和导水率高且蒸发量大,较低的持水力在长期干旱条件下土壤水分长效匮乏[31],这表明土壤水分除了受到降水的影响,还受到温度、地形和水文条件等多因素的影响.研究表明,降水强度变化会对植物功能群多样性和生产力产生积极或消极的影响[33].本研究中降水变化对糙隐子草和狗尾草的生物量产生了波动式影响,而对黄蒿和白草的生物量无显著影响,其原因主要是不同的降水强度与不同氮添加组合引起了物种组成和结构的变化,当沙质草地的水分和养分状况发生改变时,竞争力强的物种会打破种间平衡,各个物种的优势地位发生变化引起了相应生物量的改变[34-35]. ...
氮添加对亚高山针叶林土壤结构及水分入渗性能的影响
1
2023
... 降水变化会影响土壤的物理、化学性质和微生物特性,从而影响土壤持水性和养分含量[30-32].本研究中,生长季减水60%显著降低了土壤水分,其主要原因可能是沙质土壤颗粒较大,土壤饱和导水率高且蒸发量大,较低的持水力在长期干旱条件下土壤水分长效匮乏[31],这表明土壤水分除了受到降水的影响,还受到温度、地形和水文条件等多因素的影响.研究表明,降水强度变化会对植物功能群多样性和生产力产生积极或消极的影响[33].本研究中降水变化对糙隐子草和狗尾草的生物量产生了波动式影响,而对黄蒿和白草的生物量无显著影响,其原因主要是不同的降水强度与不同氮添加组合引起了物种组成和结构的变化,当沙质草地的水分和养分状况发生改变时,竞争力强的物种会打破种间平衡,各个物种的优势地位发生变化引起了相应生物量的改变[34-35]. ...
封育对草地植被功能群多样性及土壤养分特征的影响
1
2018
... 降水变化会影响土壤的物理、化学性质和微生物特性,从而影响土壤持水性和养分含量[30-32].本研究中,生长季减水60%显著降低了土壤水分,其主要原因可能是沙质土壤颗粒较大,土壤饱和导水率高且蒸发量大,较低的持水力在长期干旱条件下土壤水分长效匮乏[31],这表明土壤水分除了受到降水的影响,还受到温度、地形和水文条件等多因素的影响.研究表明,降水强度变化会对植物功能群多样性和生产力产生积极或消极的影响[33].本研究中降水变化对糙隐子草和狗尾草的生物量产生了波动式影响,而对黄蒿和白草的生物量无显著影响,其原因主要是不同的降水强度与不同氮添加组合引起了物种组成和结构的变化,当沙质草地的水分和养分状况发生改变时,竞争力强的物种会打破种间平衡,各个物种的优势地位发生变化引起了相应生物量的改变[34-35]. ...
放牧和模拟降水对短花针茅荒漠草原植物功能群多样性的影响
1
2023
... 降水变化会影响土壤的物理、化学性质和微生物特性,从而影响土壤持水性和养分含量[30-32].本研究中,生长季减水60%显著降低了土壤水分,其主要原因可能是沙质土壤颗粒较大,土壤饱和导水率高且蒸发量大,较低的持水力在长期干旱条件下土壤水分长效匮乏[31],这表明土壤水分除了受到降水的影响,还受到温度、地形和水文条件等多因素的影响.研究表明,降水强度变化会对植物功能群多样性和生产力产生积极或消极的影响[33].本研究中降水变化对糙隐子草和狗尾草的生物量产生了波动式影响,而对黄蒿和白草的生物量无显著影响,其原因主要是不同的降水强度与不同氮添加组合引起了物种组成和结构的变化,当沙质草地的水分和养分状况发生改变时,竞争力强的物种会打破种间平衡,各个物种的优势地位发生变化引起了相应生物量的改变[34-35]. ...
Nutrient enrichment,biodiversity loss,and consequent declines in eco-system productivity
2
2013
... 降水变化会影响土壤的物理、化学性质和微生物特性,从而影响土壤持水性和养分含量[30-32].本研究中,生长季减水60%显著降低了土壤水分,其主要原因可能是沙质土壤颗粒较大,土壤饱和导水率高且蒸发量大,较低的持水力在长期干旱条件下土壤水分长效匮乏[31],这表明土壤水分除了受到降水的影响,还受到温度、地形和水文条件等多因素的影响.研究表明,降水强度变化会对植物功能群多样性和生产力产生积极或消极的影响[33].本研究中降水变化对糙隐子草和狗尾草的生物量产生了波动式影响,而对黄蒿和白草的生物量无显著影响,其原因主要是不同的降水强度与不同氮添加组合引起了物种组成和结构的变化,当沙质草地的水分和养分状况发生改变时,竞争力强的物种会打破种间平衡,各个物种的优势地位发生变化引起了相应生物量的改变[34-35]. ...
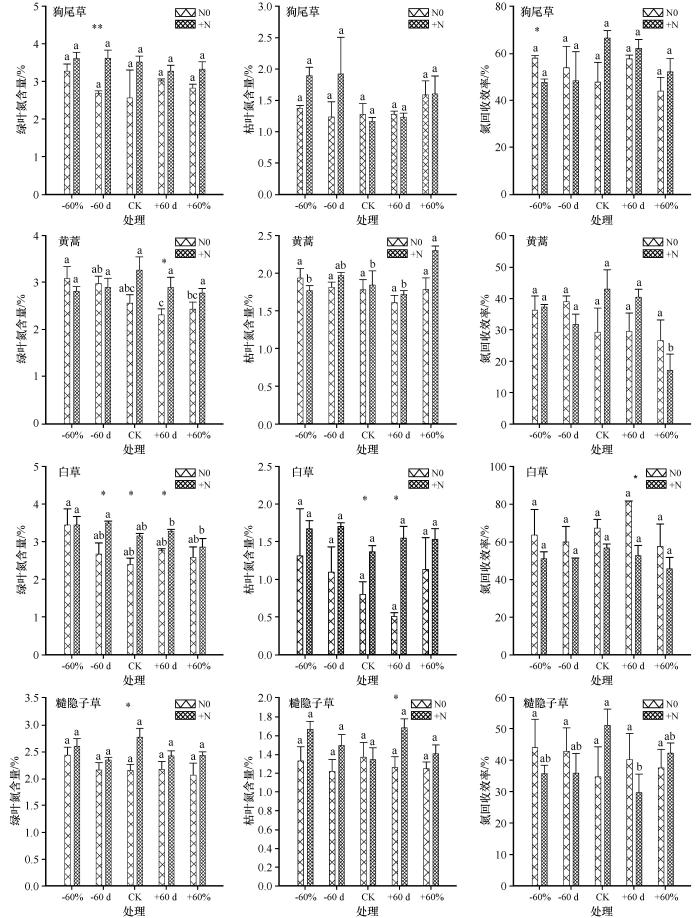
... 不同植物在绿叶期和枯叶期的NRE有显著差异,生活型也是影响NRE的主要因素[36-37].相较于多年生植物,一年生植物具有较强的光合生产能力和资源利用效率,更能适应相对干旱和贫瘠的环境[38].本研究中一年生狗尾草在干旱和氮添加的耦合效应下绿叶氮含量显著提高而NRE显著降低,说明狗尾草等一年生植物能够在干旱情况下利用有限的养分资源迅速完成生长和繁殖,从而提高对干旱环境的适应性[39],一年生黄蒿增加降水和氮添加的耦合效应下绿叶氮含量显著提高,表明黄蒿等一年生植物在水分和养分状况理想的情况下,也可以快速吸收可溶性有机氮源从而提高绿叶氮含量[40].多年生的白草和糙隐子草,氮添加能够同时提高绿叶和枯叶的叶片氮含量,在生长季前期增水和氮添加耦合效应下白草的NRE显著降低,这与NRE随着水分和养分条件改善情况下显著降低的研究结果一致[41],其主要原因是多年生植物在枯叶期回收氮是克服氮限制的适应对策,在氮匮乏的环境中,多年生植物能够利用有效养分且不断延长养分在叶片内的停留周期,从而最大限度地保证叶片中氮再次被回收和利用[35],然而在水分和养分条件改善时,多年生植物的生长繁殖会更加依赖土壤中的水分和养分,减缓对衰败叶片组织中氮的依赖性,从而降低NRE[42].因此,科尔沁沙地一年生植物和多年生植物氮回收策略的差异可能是该区域生态系统中不同生活型植物得以共存的重要解释机理,不同生活型植物的氮回收策略也是科尔沁沙质草地生态系统植物主要的养分驻留策略之一. ...
Nitrogen enrichment reduces nitrogen and phosphorus resorption through changes to species resorption and plant community composition
1
2021
... 不同植物在绿叶期和枯叶期的NRE有显著差异,生活型也是影响NRE的主要因素[36-37].相较于多年生植物,一年生植物具有较强的光合生产能力和资源利用效率,更能适应相对干旱和贫瘠的环境[38].本研究中一年生狗尾草在干旱和氮添加的耦合效应下绿叶氮含量显著提高而NRE显著降低,说明狗尾草等一年生植物能够在干旱情况下利用有限的养分资源迅速完成生长和繁殖,从而提高对干旱环境的适应性[39],一年生黄蒿增加降水和氮添加的耦合效应下绿叶氮含量显著提高,表明黄蒿等一年生植物在水分和养分状况理想的情况下,也可以快速吸收可溶性有机氮源从而提高绿叶氮含量[40].多年生的白草和糙隐子草,氮添加能够同时提高绿叶和枯叶的叶片氮含量,在生长季前期增水和氮添加耦合效应下白草的NRE显著降低,这与NRE随着水分和养分条件改善情况下显著降低的研究结果一致[41],其主要原因是多年生植物在枯叶期回收氮是克服氮限制的适应对策,在氮匮乏的环境中,多年生植物能够利用有效养分且不断延长养分在叶片内的停留周期,从而最大限度地保证叶片中氮再次被回收和利用[35],然而在水分和养分条件改善时,多年生植物的生长繁殖会更加依赖土壤中的水分和养分,减缓对衰败叶片组织中氮的依赖性,从而降低NRE[42].因此,科尔沁沙地一年生植物和多年生植物氮回收策略的差异可能是该区域生态系统中不同生活型植物得以共存的重要解释机理,不同生活型植物的氮回收策略也是科尔沁沙质草地生态系统植物主要的养分驻留策略之一. ...
科尔沁沙质草地优势多年生植物氮素回收效率的分异特征
2
2013
... 不同植物在绿叶期和枯叶期的NRE有显著差异,生活型也是影响NRE的主要因素[36-37].相较于多年生植物,一年生植物具有较强的光合生产能力和资源利用效率,更能适应相对干旱和贫瘠的环境[38].本研究中一年生狗尾草在干旱和氮添加的耦合效应下绿叶氮含量显著提高而NRE显著降低,说明狗尾草等一年生植物能够在干旱情况下利用有限的养分资源迅速完成生长和繁殖,从而提高对干旱环境的适应性[39],一年生黄蒿增加降水和氮添加的耦合效应下绿叶氮含量显著提高,表明黄蒿等一年生植物在水分和养分状况理想的情况下,也可以快速吸收可溶性有机氮源从而提高绿叶氮含量[40].多年生的白草和糙隐子草,氮添加能够同时提高绿叶和枯叶的叶片氮含量,在生长季前期增水和氮添加耦合效应下白草的NRE显著降低,这与NRE随着水分和养分条件改善情况下显著降低的研究结果一致[41],其主要原因是多年生植物在枯叶期回收氮是克服氮限制的适应对策,在氮匮乏的环境中,多年生植物能够利用有效养分且不断延长养分在叶片内的停留周期,从而最大限度地保证叶片中氮再次被回收和利用[35],然而在水分和养分条件改善时,多年生植物的生长繁殖会更加依赖土壤中的水分和养分,减缓对衰败叶片组织中氮的依赖性,从而降低NRE[42].因此,科尔沁沙地一年生植物和多年生植物氮回收策略的差异可能是该区域生态系统中不同生活型植物得以共存的重要解释机理,不同生活型植物的氮回收策略也是科尔沁沙质草地生态系统植物主要的养分驻留策略之一. ...
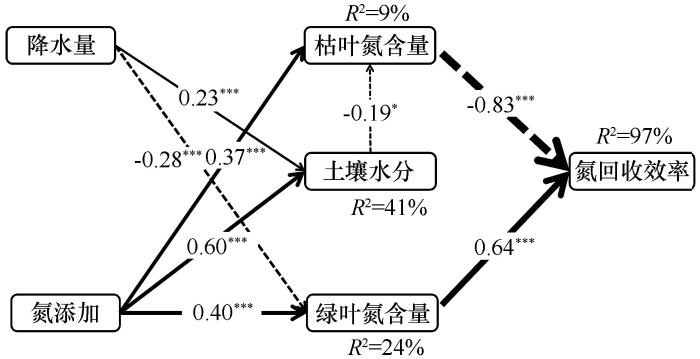
... 研究区4种优势植物绿叶氮含量、枯叶氮含量和NRE影响因素的SEM表明,降水量对土壤水分具有正效应,对绿叶氮含量具有负效应,土壤水分对枯叶氮含量具有负效应,这与增加降水能够通过降低绿叶氮含量和枯叶氮含量从而影响NRE的研究相一致[43].氮添加量对土壤水分、绿叶和枯叶氮含量都具有正效应,这主要是因为氮添加提高了土壤氮矿化速率,导致土壤有机碳含量增加,相应植物的生存力和凋落物数量同时增加导致水分贮存力提高而蒸散率降低[44],这与氮添加能显著提高土壤水分和叶片氮含量最终影响NRE的研究结果相一致[45-46].SEM表明绿叶氮含量对NRE具有负效应,而枯叶氮对NRE具有正效应,这表明除了绿叶氮含量之外,枯叶氮含量也是影响氮回收效率的决定性因素,尤其是多年生植物叶片在衰败过程中尽可能贮存更多氮,延长氮在植物体内的驻留时间,为第二年的返青做准备[37].然而土壤全氮在SEM中未被选入,表明在该区域,相较于土壤全氮,土壤水分在调控植物叶片NRE的过程中所发挥的作用可能更为重要. ...
科尔沁沙地草地植物群落功能性状对封育和放牧的响应
1
2017
... 不同植物在绿叶期和枯叶期的NRE有显著差异,生活型也是影响NRE的主要因素[36-37].相较于多年生植物,一年生植物具有较强的光合生产能力和资源利用效率,更能适应相对干旱和贫瘠的环境[38].本研究中一年生狗尾草在干旱和氮添加的耦合效应下绿叶氮含量显著提高而NRE显著降低,说明狗尾草等一年生植物能够在干旱情况下利用有限的养分资源迅速完成生长和繁殖,从而提高对干旱环境的适应性[39],一年生黄蒿增加降水和氮添加的耦合效应下绿叶氮含量显著提高,表明黄蒿等一年生植物在水分和养分状况理想的情况下,也可以快速吸收可溶性有机氮源从而提高绿叶氮含量[40].多年生的白草和糙隐子草,氮添加能够同时提高绿叶和枯叶的叶片氮含量,在生长季前期增水和氮添加耦合效应下白草的NRE显著降低,这与NRE随着水分和养分条件改善情况下显著降低的研究结果一致[41],其主要原因是多年生植物在枯叶期回收氮是克服氮限制的适应对策,在氮匮乏的环境中,多年生植物能够利用有效养分且不断延长养分在叶片内的停留周期,从而最大限度地保证叶片中氮再次被回收和利用[35],然而在水分和养分条件改善时,多年生植物的生长繁殖会更加依赖土壤中的水分和养分,减缓对衰败叶片组织中氮的依赖性,从而降低NRE[42].因此,科尔沁沙地一年生植物和多年生植物氮回收策略的差异可能是该区域生态系统中不同生活型植物得以共存的重要解释机理,不同生活型植物的氮回收策略也是科尔沁沙质草地生态系统植物主要的养分驻留策略之一. ...
Experimental investigation of fire ecology in the C3 and C4 subspecies of Alloteropsis semialata
1
2010
... 不同植物在绿叶期和枯叶期的NRE有显著差异,生活型也是影响NRE的主要因素[36-37].相较于多年生植物,一年生植物具有较强的光合生产能力和资源利用效率,更能适应相对干旱和贫瘠的环境[38].本研究中一年生狗尾草在干旱和氮添加的耦合效应下绿叶氮含量显著提高而NRE显著降低,说明狗尾草等一年生植物能够在干旱情况下利用有限的养分资源迅速完成生长和繁殖,从而提高对干旱环境的适应性[39],一年生黄蒿增加降水和氮添加的耦合效应下绿叶氮含量显著提高,表明黄蒿等一年生植物在水分和养分状况理想的情况下,也可以快速吸收可溶性有机氮源从而提高绿叶氮含量[40].多年生的白草和糙隐子草,氮添加能够同时提高绿叶和枯叶的叶片氮含量,在生长季前期增水和氮添加耦合效应下白草的NRE显著降低,这与NRE随着水分和养分条件改善情况下显著降低的研究结果一致[41],其主要原因是多年生植物在枯叶期回收氮是克服氮限制的适应对策,在氮匮乏的环境中,多年生植物能够利用有效养分且不断延长养分在叶片内的停留周期,从而最大限度地保证叶片中氮再次被回收和利用[35],然而在水分和养分条件改善时,多年生植物的生长繁殖会更加依赖土壤中的水分和养分,减缓对衰败叶片组织中氮的依赖性,从而降低NRE[42].因此,科尔沁沙地一年生植物和多年生植物氮回收策略的差异可能是该区域生态系统中不同生活型植物得以共存的重要解释机理,不同生活型植物的氮回收策略也是科尔沁沙质草地生态系统植物主要的养分驻留策略之一. ...
古尔班通古特沙漠不同生活型草本植物对氮素吸收偏好性研究
1
2022
... 不同植物在绿叶期和枯叶期的NRE有显著差异,生活型也是影响NRE的主要因素[36-37].相较于多年生植物,一年生植物具有较强的光合生产能力和资源利用效率,更能适应相对干旱和贫瘠的环境[38].本研究中一年生狗尾草在干旱和氮添加的耦合效应下绿叶氮含量显著提高而NRE显著降低,说明狗尾草等一年生植物能够在干旱情况下利用有限的养分资源迅速完成生长和繁殖,从而提高对干旱环境的适应性[39],一年生黄蒿增加降水和氮添加的耦合效应下绿叶氮含量显著提高,表明黄蒿等一年生植物在水分和养分状况理想的情况下,也可以快速吸收可溶性有机氮源从而提高绿叶氮含量[40].多年生的白草和糙隐子草,氮添加能够同时提高绿叶和枯叶的叶片氮含量,在生长季前期增水和氮添加耦合效应下白草的NRE显著降低,这与NRE随着水分和养分条件改善情况下显著降低的研究结果一致[41],其主要原因是多年生植物在枯叶期回收氮是克服氮限制的适应对策,在氮匮乏的环境中,多年生植物能够利用有效养分且不断延长养分在叶片内的停留周期,从而最大限度地保证叶片中氮再次被回收和利用[35],然而在水分和养分条件改善时,多年生植物的生长繁殖会更加依赖土壤中的水分和养分,减缓对衰败叶片组织中氮的依赖性,从而降低NRE[42].因此,科尔沁沙地一年生植物和多年生植物氮回收策略的差异可能是该区域生态系统中不同生活型植物得以共存的重要解释机理,不同生活型植物的氮回收策略也是科尔沁沙质草地生态系统植物主要的养分驻留策略之一. ...
氮素添加对黄土高原典型草原长芒草氮磷重吸收率及C∶N∶P化学计量特征的影响
1
2011
... 不同植物在绿叶期和枯叶期的NRE有显著差异,生活型也是影响NRE的主要因素[36-37].相较于多年生植物,一年生植物具有较强的光合生产能力和资源利用效率,更能适应相对干旱和贫瘠的环境[38].本研究中一年生狗尾草在干旱和氮添加的耦合效应下绿叶氮含量显著提高而NRE显著降低,说明狗尾草等一年生植物能够在干旱情况下利用有限的养分资源迅速完成生长和繁殖,从而提高对干旱环境的适应性[39],一年生黄蒿增加降水和氮添加的耦合效应下绿叶氮含量显著提高,表明黄蒿等一年生植物在水分和养分状况理想的情况下,也可以快速吸收可溶性有机氮源从而提高绿叶氮含量[40].多年生的白草和糙隐子草,氮添加能够同时提高绿叶和枯叶的叶片氮含量,在生长季前期增水和氮添加耦合效应下白草的NRE显著降低,这与NRE随着水分和养分条件改善情况下显著降低的研究结果一致[41],其主要原因是多年生植物在枯叶期回收氮是克服氮限制的适应对策,在氮匮乏的环境中,多年生植物能够利用有效养分且不断延长养分在叶片内的停留周期,从而最大限度地保证叶片中氮再次被回收和利用[35],然而在水分和养分条件改善时,多年生植物的生长繁殖会更加依赖土壤中的水分和养分,减缓对衰败叶片组织中氮的依赖性,从而降低NRE[42].因此,科尔沁沙地一年生植物和多年生植物氮回收策略的差异可能是该区域生态系统中不同生活型植物得以共存的重要解释机理,不同生活型植物的氮回收策略也是科尔沁沙质草地生态系统植物主要的养分驻留策略之一. ...
Convergent responses of nitrogen and phosphorus resorption to nitrogen inputs in a semiarid grassland
1
2013
... 不同植物在绿叶期和枯叶期的NRE有显著差异,生活型也是影响NRE的主要因素[36-37].相较于多年生植物,一年生植物具有较强的光合生产能力和资源利用效率,更能适应相对干旱和贫瘠的环境[38].本研究中一年生狗尾草在干旱和氮添加的耦合效应下绿叶氮含量显著提高而NRE显著降低,说明狗尾草等一年生植物能够在干旱情况下利用有限的养分资源迅速完成生长和繁殖,从而提高对干旱环境的适应性[39],一年生黄蒿增加降水和氮添加的耦合效应下绿叶氮含量显著提高,表明黄蒿等一年生植物在水分和养分状况理想的情况下,也可以快速吸收可溶性有机氮源从而提高绿叶氮含量[40].多年生的白草和糙隐子草,氮添加能够同时提高绿叶和枯叶的叶片氮含量,在生长季前期增水和氮添加耦合效应下白草的NRE显著降低,这与NRE随着水分和养分条件改善情况下显著降低的研究结果一致[41],其主要原因是多年生植物在枯叶期回收氮是克服氮限制的适应对策,在氮匮乏的环境中,多年生植物能够利用有效养分且不断延长养分在叶片内的停留周期,从而最大限度地保证叶片中氮再次被回收和利用[35],然而在水分和养分条件改善时,多年生植物的生长繁殖会更加依赖土壤中的水分和养分,减缓对衰败叶片组织中氮的依赖性,从而降低NRE[42].因此,科尔沁沙地一年生植物和多年生植物氮回收策略的差异可能是该区域生态系统中不同生活型植物得以共存的重要解释机理,不同生活型植物的氮回收策略也是科尔沁沙质草地生态系统植物主要的养分驻留策略之一. ...
Nutrient resorption responses to water and nitrogen amendment in semi-arid grassland of Inner Mongolia China
1
2010
... 研究区4种优势植物绿叶氮含量、枯叶氮含量和NRE影响因素的SEM表明,降水量对土壤水分具有正效应,对绿叶氮含量具有负效应,土壤水分对枯叶氮含量具有负效应,这与增加降水能够通过降低绿叶氮含量和枯叶氮含量从而影响NRE的研究相一致[43].氮添加量对土壤水分、绿叶和枯叶氮含量都具有正效应,这主要是因为氮添加提高了土壤氮矿化速率,导致土壤有机碳含量增加,相应植物的生存力和凋落物数量同时增加导致水分贮存力提高而蒸散率降低[44],这与氮添加能显著提高土壤水分和叶片氮含量最终影响NRE的研究结果相一致[45-46].SEM表明绿叶氮含量对NRE具有负效应,而枯叶氮对NRE具有正效应,这表明除了绿叶氮含量之外,枯叶氮含量也是影响氮回收效率的决定性因素,尤其是多年生植物叶片在衰败过程中尽可能贮存更多氮,延长氮在植物体内的驻留时间,为第二年的返青做准备[37].然而土壤全氮在SEM中未被选入,表明在该区域,相较于土壤全氮,土壤水分在调控植物叶片NRE的过程中所发挥的作用可能更为重要. ...
氮添加对巴音布鲁克高寒湿地土壤无机磷形态的影响
1
2022
... 研究区4种优势植物绿叶氮含量、枯叶氮含量和NRE影响因素的SEM表明,降水量对土壤水分具有正效应,对绿叶氮含量具有负效应,土壤水分对枯叶氮含量具有负效应,这与增加降水能够通过降低绿叶氮含量和枯叶氮含量从而影响NRE的研究相一致[43].氮添加量对土壤水分、绿叶和枯叶氮含量都具有正效应,这主要是因为氮添加提高了土壤氮矿化速率,导致土壤有机碳含量增加,相应植物的生存力和凋落物数量同时增加导致水分贮存力提高而蒸散率降低[44],这与氮添加能显著提高土壤水分和叶片氮含量最终影响NRE的研究结果相一致[45-46].SEM表明绿叶氮含量对NRE具有负效应,而枯叶氮对NRE具有正效应,这表明除了绿叶氮含量之外,枯叶氮含量也是影响氮回收效率的决定性因素,尤其是多年生植物叶片在衰败过程中尽可能贮存更多氮,延长氮在植物体内的驻留时间,为第二年的返青做准备[37].然而土壤全氮在SEM中未被选入,表明在该区域,相较于土壤全氮,土壤水分在调控植物叶片NRE的过程中所发挥的作用可能更为重要. ...
不同降水条件下荒漠草原植物的养分含量及回收对增温和氮素添加的响应
1
2019
... 研究区4种优势植物绿叶氮含量、枯叶氮含量和NRE影响因素的SEM表明,降水量对土壤水分具有正效应,对绿叶氮含量具有负效应,土壤水分对枯叶氮含量具有负效应,这与增加降水能够通过降低绿叶氮含量和枯叶氮含量从而影响NRE的研究相一致[43].氮添加量对土壤水分、绿叶和枯叶氮含量都具有正效应,这主要是因为氮添加提高了土壤氮矿化速率,导致土壤有机碳含量增加,相应植物的生存力和凋落物数量同时增加导致水分贮存力提高而蒸散率降低[44],这与氮添加能显著提高土壤水分和叶片氮含量最终影响NRE的研究结果相一致[45-46].SEM表明绿叶氮含量对NRE具有负效应,而枯叶氮对NRE具有正效应,这表明除了绿叶氮含量之外,枯叶氮含量也是影响氮回收效率的决定性因素,尤其是多年生植物叶片在衰败过程中尽可能贮存更多氮,延长氮在植物体内的驻留时间,为第二年的返青做准备[37].然而土壤全氮在SEM中未被选入,表明在该区域,相较于土壤全氮,土壤水分在调控植物叶片NRE的过程中所发挥的作用可能更为重要. ...
Resorption efficiency decreases with increasing green leaf nutrients in a global data set
1
2005
... 研究区4种优势植物绿叶氮含量、枯叶氮含量和NRE影响因素的SEM表明,降水量对土壤水分具有正效应,对绿叶氮含量具有负效应,土壤水分对枯叶氮含量具有负效应,这与增加降水能够通过降低绿叶氮含量和枯叶氮含量从而影响NRE的研究相一致[43].氮添加量对土壤水分、绿叶和枯叶氮含量都具有正效应,这主要是因为氮添加提高了土壤氮矿化速率,导致土壤有机碳含量增加,相应植物的生存力和凋落物数量同时增加导致水分贮存力提高而蒸散率降低[44],这与氮添加能显著提高土壤水分和叶片氮含量最终影响NRE的研究结果相一致[45-46].SEM表明绿叶氮含量对NRE具有负效应,而枯叶氮对NRE具有正效应,这表明除了绿叶氮含量之外,枯叶氮含量也是影响氮回收效率的决定性因素,尤其是多年生植物叶片在衰败过程中尽可能贮存更多氮,延长氮在植物体内的驻留时间,为第二年的返青做准备[37].然而土壤全氮在SEM中未被选入,表明在该区域,相较于土壤全氮,土壤水分在调控植物叶片NRE的过程中所发挥的作用可能更为重要. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}