中国干旱区恢复生态学研究进展及趋势评述
3
2014
... 人工固沙植被修复是干旱区沙区退化生态系统稳定性维持的有效途径[1].宁夏沙坡头作为人工植被固沙的典型样板,自1956年人工植被防护体系建立以来,后又分别于不同年份(1964、1981、1987年)进行了逐步扩建.固沙植被的建立和演替促进了隐花植物和浅根系草本植物的繁殖,增加了该区生态系统的物种多样性[2].藓类植物作为其中的拓荒者,对维持和促进生态系统功能具有重要作用.它们的根系分泌物与土壤微生物和土壤颗粒胶结形成的藓结皮,是干旱荒漠区生物结皮发育成熟的类群,具有较高的水分拦截效率和光合作用能力,对干旱区土壤、水文和生物过程具有重要促进作用[1,3].全球大部分干旱土地正处于日益严重的压力下,土壤正在逐渐退化或已经退化,生物土壤结皮是旱地恢复和可持续性发展的新希望[4].因此对干旱区藓类植物“拓荒过程”的适应性研究可以帮助我们更全面和深入地认识藓类植物适应环境变化的途径和机制,同时可为未来干旱区生态系统修复的藓类物种选择提供一定的科学依据. ...
... [1,3].全球大部分干旱土地正处于日益严重的压力下,土壤正在逐渐退化或已经退化,生物土壤结皮是旱地恢复和可持续性发展的新希望[4].因此对干旱区藓类植物“拓荒过程”的适应性研究可以帮助我们更全面和深入地认识藓类植物适应环境变化的途径和机制,同时可为未来干旱区生态系统修复的藓类物种选择提供一定的科学依据. ...
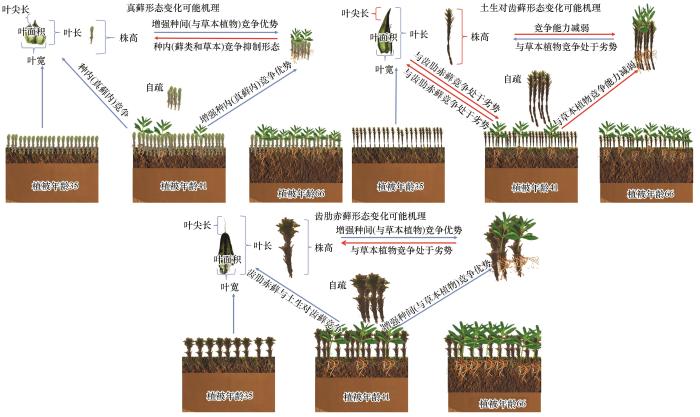
... 腾格里沙漠沙坡头人工固沙植被区经常面临高温、强辐射、干旱等胁迫因子的影响,人工固沙植被的建立为微生物、隐花植物和草本植物的繁育提供了基础[2],改善了该区浅层土壤养分状况[26],有利于浅根系植物的生长发育.其中藓类植物与土壤微生物以及其他生物体通过假根和分泌物等与土壤颗粒胶结形成生物土壤结皮[1],而生物结皮可以改变土壤表面粗糙度、土壤温度、湿度和养分含量等土壤性质,进而影响维管植物种子的萌发、定植和存活.此外,生物结皮还会影响浅层土壤含水量,从而增加浅根系草本植物物种的丰富度和生物量[34-35].前人对沙坡头人工固沙植被区的群落演替特征做了大量的研究和分析,发现生物结皮改变了土壤水分的时空格局和再分配特征,导致固沙植被区植被从高盖度的旱生灌木群落向以低盖度的浅根草本植物为主的复杂群落转变[34].与深根系灌木相比,一年生草本植物与生物结皮生态位更为接近[34],当样地内草本植物十分发达时,会与藓类植物产生种间竞争,此时,即便藓类植物不断调节自身资源获取能力以适应环境,但仍然处于劣势.过多的草本植物抑制了藓类植物的生长,恶劣的环境条件和低养分获取导致藓类植物的叶面积、叶宽、叶长、叶尖长、生物量及密度下降 [36-37].因此,我们推测该区藓类植物在演替后一阶段同样被样地内草本植物抑制了生长,从而导致各形态指标普遍降低.其中叶尖长和叶长与最大光合速率、最大叶绿素荧光相关,这也表明在演替后一阶段藓类植物光合竞争能力有所减弱[14]. ...
腾格里沙漠人工固沙植被演替过程中生物土壤结皮盖度对沙埋的响应
2
2017
... 人工固沙植被修复是干旱区沙区退化生态系统稳定性维持的有效途径[1].宁夏沙坡头作为人工植被固沙的典型样板,自1956年人工植被防护体系建立以来,后又分别于不同年份(1964、1981、1987年)进行了逐步扩建.固沙植被的建立和演替促进了隐花植物和浅根系草本植物的繁殖,增加了该区生态系统的物种多样性[2].藓类植物作为其中的拓荒者,对维持和促进生态系统功能具有重要作用.它们的根系分泌物与土壤微生物和土壤颗粒胶结形成的藓结皮,是干旱荒漠区生物结皮发育成熟的类群,具有较高的水分拦截效率和光合作用能力,对干旱区土壤、水文和生物过程具有重要促进作用[1,3].全球大部分干旱土地正处于日益严重的压力下,土壤正在逐渐退化或已经退化,生物土壤结皮是旱地恢复和可持续性发展的新希望[4].因此对干旱区藓类植物“拓荒过程”的适应性研究可以帮助我们更全面和深入地认识藓类植物适应环境变化的途径和机制,同时可为未来干旱区生态系统修复的藓类物种选择提供一定的科学依据. ...
... 腾格里沙漠沙坡头人工固沙植被区经常面临高温、强辐射、干旱等胁迫因子的影响,人工固沙植被的建立为微生物、隐花植物和草本植物的繁育提供了基础[2],改善了该区浅层土壤养分状况[26],有利于浅根系植物的生长发育.其中藓类植物与土壤微生物以及其他生物体通过假根和分泌物等与土壤颗粒胶结形成生物土壤结皮[1],而生物结皮可以改变土壤表面粗糙度、土壤温度、湿度和养分含量等土壤性质,进而影响维管植物种子的萌发、定植和存活.此外,生物结皮还会影响浅层土壤含水量,从而增加浅根系草本植物物种的丰富度和生物量[34-35].前人对沙坡头人工固沙植被区的群落演替特征做了大量的研究和分析,发现生物结皮改变了土壤水分的时空格局和再分配特征,导致固沙植被区植被从高盖度的旱生灌木群落向以低盖度的浅根草本植物为主的复杂群落转变[34].与深根系灌木相比,一年生草本植物与生物结皮生态位更为接近[34],当样地内草本植物十分发达时,会与藓类植物产生种间竞争,此时,即便藓类植物不断调节自身资源获取能力以适应环境,但仍然处于劣势.过多的草本植物抑制了藓类植物的生长,恶劣的环境条件和低养分获取导致藓类植物的叶面积、叶宽、叶长、叶尖长、生物量及密度下降 [36-37].因此,我们推测该区藓类植物在演替后一阶段同样被样地内草本植物抑制了生长,从而导致各形态指标普遍降低.其中叶尖长和叶长与最大光合速率、最大叶绿素荧光相关,这也表明在演替后一阶段藓类植物光合竞争能力有所减弱[14]. ...
中国荒漠与沙地生物土壤结皮研究
1
2018
... 人工固沙植被修复是干旱区沙区退化生态系统稳定性维持的有效途径[1].宁夏沙坡头作为人工植被固沙的典型样板,自1956年人工植被防护体系建立以来,后又分别于不同年份(1964、1981、1987年)进行了逐步扩建.固沙植被的建立和演替促进了隐花植物和浅根系草本植物的繁殖,增加了该区生态系统的物种多样性[2].藓类植物作为其中的拓荒者,对维持和促进生态系统功能具有重要作用.它们的根系分泌物与土壤微生物和土壤颗粒胶结形成的藓结皮,是干旱荒漠区生物结皮发育成熟的类群,具有较高的水分拦截效率和光合作用能力,对干旱区土壤、水文和生物过程具有重要促进作用[1,3].全球大部分干旱土地正处于日益严重的压力下,土壤正在逐渐退化或已经退化,生物土壤结皮是旱地恢复和可持续性发展的新希望[4].因此对干旱区藓类植物“拓荒过程”的适应性研究可以帮助我们更全面和深入地认识藓类植物适应环境变化的途径和机制,同时可为未来干旱区生态系统修复的藓类物种选择提供一定的科学依据. ...
Dual use of solar power plants as biocrust nurseries for large-scale arid soil restoration
1
2023
... 人工固沙植被修复是干旱区沙区退化生态系统稳定性维持的有效途径[1].宁夏沙坡头作为人工植被固沙的典型样板,自1956年人工植被防护体系建立以来,后又分别于不同年份(1964、1981、1987年)进行了逐步扩建.固沙植被的建立和演替促进了隐花植物和浅根系草本植物的繁殖,增加了该区生态系统的物种多样性[2].藓类植物作为其中的拓荒者,对维持和促进生态系统功能具有重要作用.它们的根系分泌物与土壤微生物和土壤颗粒胶结形成的藓结皮,是干旱荒漠区生物结皮发育成熟的类群,具有较高的水分拦截效率和光合作用能力,对干旱区土壤、水文和生物过程具有重要促进作用[1,3].全球大部分干旱土地正处于日益严重的压力下,土壤正在逐渐退化或已经退化,生物土壤结皮是旱地恢复和可持续性发展的新希望[4].因此对干旱区藓类植物“拓荒过程”的适应性研究可以帮助我们更全面和深入地认识藓类植物适应环境变化的途径和机制,同时可为未来干旱区生态系统修复的藓类物种选择提供一定的科学依据. ...
Cold adaptation drives variability in needle structure and anatomy in Pinus sylvestris L.along a 1,900 km temperate-boreal transect
1
2017
... 植物的形态结构是功能的基础,植物形态的多样性反映了资源利用和抵抗外部胁迫的多元化适应策略[5-6],在解析植物生活史对策方面具有重要作用.藓类植物是典型的变水性植物,具有快速平衡组织含水量和适应间歇性代谢的特性[7],他们的形态结构表现出较强的可塑性[8].亚热带山地生长的扇形藓类植物的扇形生命形态(茎叶体扁平)有助于调节水分平衡和进行气体交换[9],而干旱区藓类植物则可以通过调节不同水平上的形态特征来增强抗逆性.前期对干旱区藓类植物的研究发现,在茎叶水平上,透明的叶尖端具有减缓水分散失、反射部分紫外辐射以及运输水分等功能[10-11];在个体水平上,较矮小的植株个体在强光下可减缓水分散失[12];在种群和群落水平上,藓类植物的形态特征以集丛型为主,有利于减少空气在叶片缝隙中的流动[13].此外,Coe等[14]认为旱生藓类植物可以通过调整叶长、叶宽和叶面积等形态特征从而调整个体碳收支.这些形态适应都有助于增强藓类植物的抗逆性.但是,前人大部分的研究基于特定年份的静态的属性描述,而对于形态特征在时间演替方面的塑造规律尚缺乏相关认知. ...
The worldwide leaf economics spectrum
1
2004
... 植物的形态结构是功能的基础,植物形态的多样性反映了资源利用和抵抗外部胁迫的多元化适应策略[5-6],在解析植物生活史对策方面具有重要作用.藓类植物是典型的变水性植物,具有快速平衡组织含水量和适应间歇性代谢的特性[7],他们的形态结构表现出较强的可塑性[8].亚热带山地生长的扇形藓类植物的扇形生命形态(茎叶体扁平)有助于调节水分平衡和进行气体交换[9],而干旱区藓类植物则可以通过调节不同水平上的形态特征来增强抗逆性.前期对干旱区藓类植物的研究发现,在茎叶水平上,透明的叶尖端具有减缓水分散失、反射部分紫外辐射以及运输水分等功能[10-11];在个体水平上,较矮小的植株个体在强光下可减缓水分散失[12];在种群和群落水平上,藓类植物的形态特征以集丛型为主,有利于减少空气在叶片缝隙中的流动[13].此外,Coe等[14]认为旱生藓类植物可以通过调整叶长、叶宽和叶面积等形态特征从而调整个体碳收支.这些形态适应都有助于增强藓类植物的抗逆性.但是,前人大部分的研究基于特定年份的静态的属性描述,而对于形态特征在时间演替方面的塑造规律尚缺乏相关认知. ...
Patterns of desiccation tolerance and recovery in bryophytes
1
2001
... 植物的形态结构是功能的基础,植物形态的多样性反映了资源利用和抵抗外部胁迫的多元化适应策略[5-6],在解析植物生活史对策方面具有重要作用.藓类植物是典型的变水性植物,具有快速平衡组织含水量和适应间歇性代谢的特性[7],他们的形态结构表现出较强的可塑性[8].亚热带山地生长的扇形藓类植物的扇形生命形态(茎叶体扁平)有助于调节水分平衡和进行气体交换[9],而干旱区藓类植物则可以通过调节不同水平上的形态特征来增强抗逆性.前期对干旱区藓类植物的研究发现,在茎叶水平上,透明的叶尖端具有减缓水分散失、反射部分紫外辐射以及运输水分等功能[10-11];在个体水平上,较矮小的植株个体在强光下可减缓水分散失[12];在种群和群落水平上,藓类植物的形态特征以集丛型为主,有利于减少空气在叶片缝隙中的流动[13].此外,Coe等[14]认为旱生藓类植物可以通过调整叶长、叶宽和叶面积等形态特征从而调整个体碳收支.这些形态适应都有助于增强藓类植物的抗逆性.但是,前人大部分的研究基于特定年份的静态的属性描述,而对于形态特征在时间演替方面的塑造规律尚缺乏相关认知. ...
Biological soil crusts as an integral component of desert environments
1
2013
... 植物的形态结构是功能的基础,植物形态的多样性反映了资源利用和抵抗外部胁迫的多元化适应策略[5-6],在解析植物生活史对策方面具有重要作用.藓类植物是典型的变水性植物,具有快速平衡组织含水量和适应间歇性代谢的特性[7],他们的形态结构表现出较强的可塑性[8].亚热带山地生长的扇形藓类植物的扇形生命形态(茎叶体扁平)有助于调节水分平衡和进行气体交换[9],而干旱区藓类植物则可以通过调节不同水平上的形态特征来增强抗逆性.前期对干旱区藓类植物的研究发现,在茎叶水平上,透明的叶尖端具有减缓水分散失、反射部分紫外辐射以及运输水分等功能[10-11];在个体水平上,较矮小的植株个体在强光下可减缓水分散失[12];在种群和群落水平上,藓类植物的形态特征以集丛型为主,有利于减少空气在叶片缝隙中的流动[13].此外,Coe等[14]认为旱生藓类植物可以通过调整叶长、叶宽和叶面积等形态特征从而调整个体碳收支.这些形态适应都有助于增强藓类植物的抗逆性.但是,前人大部分的研究基于特定年份的静态的属性描述,而对于形态特征在时间演替方面的塑造规律尚缺乏相关认知. ...
Water relations and gas exchange of fan bryophytes and their adaptations to microhabitats in an Asian subtropical montane cloud forest
1
2015
... 植物的形态结构是功能的基础,植物形态的多样性反映了资源利用和抵抗外部胁迫的多元化适应策略[5-6],在解析植物生活史对策方面具有重要作用.藓类植物是典型的变水性植物,具有快速平衡组织含水量和适应间歇性代谢的特性[7],他们的形态结构表现出较强的可塑性[8].亚热带山地生长的扇形藓类植物的扇形生命形态(茎叶体扁平)有助于调节水分平衡和进行气体交换[9],而干旱区藓类植物则可以通过调节不同水平上的形态特征来增强抗逆性.前期对干旱区藓类植物的研究发现,在茎叶水平上,透明的叶尖端具有减缓水分散失、反射部分紫外辐射以及运输水分等功能[10-11];在个体水平上,较矮小的植株个体在强光下可减缓水分散失[12];在种群和群落水平上,藓类植物的形态特征以集丛型为主,有利于减少空气在叶片缝隙中的流动[13].此外,Coe等[14]认为旱生藓类植物可以通过调整叶长、叶宽和叶面积等形态特征从而调整个体碳收支.这些形态适应都有助于增强藓类植物的抗逆性.但是,前人大部分的研究基于特定年份的静态的属性描述,而对于形态特征在时间演替方面的塑造规律尚缺乏相关认知. ...
Effects of leaf hair points of a desert moss on water retention and dew formation:implications for desiccation tolerance
1
2012
... 植物的形态结构是功能的基础,植物形态的多样性反映了资源利用和抵抗外部胁迫的多元化适应策略[5-6],在解析植物生活史对策方面具有重要作用.藓类植物是典型的变水性植物,具有快速平衡组织含水量和适应间歇性代谢的特性[7],他们的形态结构表现出较强的可塑性[8].亚热带山地生长的扇形藓类植物的扇形生命形态(茎叶体扁平)有助于调节水分平衡和进行气体交换[9],而干旱区藓类植物则可以通过调节不同水平上的形态特征来增强抗逆性.前期对干旱区藓类植物的研究发现,在茎叶水平上,透明的叶尖端具有减缓水分散失、反射部分紫外辐射以及运输水分等功能[10-11];在个体水平上,较矮小的植株个体在强光下可减缓水分散失[12];在种群和群落水平上,藓类植物的形态特征以集丛型为主,有利于减少空气在叶片缝隙中的流动[13].此外,Coe等[14]认为旱生藓类植物可以通过调整叶长、叶宽和叶面积等形态特征从而调整个体碳收支.这些形态适应都有助于增强藓类植物的抗逆性.但是,前人大部分的研究基于特定年份的静态的属性描述,而对于形态特征在时间演替方面的塑造规律尚缺乏相关认知. ...
The upside-down water collection system of Syntrichia caninervis
1
2016
... 植物的形态结构是功能的基础,植物形态的多样性反映了资源利用和抵抗外部胁迫的多元化适应策略[5-6],在解析植物生活史对策方面具有重要作用.藓类植物是典型的变水性植物,具有快速平衡组织含水量和适应间歇性代谢的特性[7],他们的形态结构表现出较强的可塑性[8].亚热带山地生长的扇形藓类植物的扇形生命形态(茎叶体扁平)有助于调节水分平衡和进行气体交换[9],而干旱区藓类植物则可以通过调节不同水平上的形态特征来增强抗逆性.前期对干旱区藓类植物的研究发现,在茎叶水平上,透明的叶尖端具有减缓水分散失、反射部分紫外辐射以及运输水分等功能[10-11];在个体水平上,较矮小的植株个体在强光下可减缓水分散失[12];在种群和群落水平上,藓类植物的形态特征以集丛型为主,有利于减少空气在叶片缝隙中的流动[13].此外,Coe等[14]认为旱生藓类植物可以通过调整叶长、叶宽和叶面积等形态特征从而调整个体碳收支.这些形态适应都有助于增强藓类植物的抗逆性.但是,前人大部分的研究基于特定年份的静态的属性描述,而对于形态特征在时间演替方面的塑造规律尚缺乏相关认知. ...
石漠化干旱环境中石生藓类水分吸收特征及其结构适应性
1
2012
... 植物的形态结构是功能的基础,植物形态的多样性反映了资源利用和抵抗外部胁迫的多元化适应策略[5-6],在解析植物生活史对策方面具有重要作用.藓类植物是典型的变水性植物,具有快速平衡组织含水量和适应间歇性代谢的特性[7],他们的形态结构表现出较强的可塑性[8].亚热带山地生长的扇形藓类植物的扇形生命形态(茎叶体扁平)有助于调节水分平衡和进行气体交换[9],而干旱区藓类植物则可以通过调节不同水平上的形态特征来增强抗逆性.前期对干旱区藓类植物的研究发现,在茎叶水平上,透明的叶尖端具有减缓水分散失、反射部分紫外辐射以及运输水分等功能[10-11];在个体水平上,较矮小的植株个体在强光下可减缓水分散失[12];在种群和群落水平上,藓类植物的形态特征以集丛型为主,有利于减少空气在叶片缝隙中的流动[13].此外,Coe等[14]认为旱生藓类植物可以通过调整叶长、叶宽和叶面积等形态特征从而调整个体碳收支.这些形态适应都有助于增强藓类植物的抗逆性.但是,前人大部分的研究基于特定年份的静态的属性描述,而对于形态特征在时间演替方面的塑造规律尚缺乏相关认知. ...
西藏高寒草原藓类植物群丛特征及其与生态因子的关系
1
2023
... 植物的形态结构是功能的基础,植物形态的多样性反映了资源利用和抵抗外部胁迫的多元化适应策略[5-6],在解析植物生活史对策方面具有重要作用.藓类植物是典型的变水性植物,具有快速平衡组织含水量和适应间歇性代谢的特性[7],他们的形态结构表现出较强的可塑性[8].亚热带山地生长的扇形藓类植物的扇形生命形态(茎叶体扁平)有助于调节水分平衡和进行气体交换[9],而干旱区藓类植物则可以通过调节不同水平上的形态特征来增强抗逆性.前期对干旱区藓类植物的研究发现,在茎叶水平上,透明的叶尖端具有减缓水分散失、反射部分紫外辐射以及运输水分等功能[10-11];在个体水平上,较矮小的植株个体在强光下可减缓水分散失[12];在种群和群落水平上,藓类植物的形态特征以集丛型为主,有利于减少空气在叶片缝隙中的流动[13].此外,Coe等[14]认为旱生藓类植物可以通过调整叶长、叶宽和叶面积等形态特征从而调整个体碳收支.这些形态适应都有助于增强藓类植物的抗逆性.但是,前人大部分的研究基于特定年份的静态的属性描述,而对于形态特征在时间演替方面的塑造规律尚缺乏相关认知. ...
Morphological and physiological traits in relation to carbon balance in a diverse clade of dryland mosses
2
2019
... 植物的形态结构是功能的基础,植物形态的多样性反映了资源利用和抵抗外部胁迫的多元化适应策略[5-6],在解析植物生活史对策方面具有重要作用.藓类植物是典型的变水性植物,具有快速平衡组织含水量和适应间歇性代谢的特性[7],他们的形态结构表现出较强的可塑性[8].亚热带山地生长的扇形藓类植物的扇形生命形态(茎叶体扁平)有助于调节水分平衡和进行气体交换[9],而干旱区藓类植物则可以通过调节不同水平上的形态特征来增强抗逆性.前期对干旱区藓类植物的研究发现,在茎叶水平上,透明的叶尖端具有减缓水分散失、反射部分紫外辐射以及运输水分等功能[10-11];在个体水平上,较矮小的植株个体在强光下可减缓水分散失[12];在种群和群落水平上,藓类植物的形态特征以集丛型为主,有利于减少空气在叶片缝隙中的流动[13].此外,Coe等[14]认为旱生藓类植物可以通过调整叶长、叶宽和叶面积等形态特征从而调整个体碳收支.这些形态适应都有助于增强藓类植物的抗逆性.但是,前人大部分的研究基于特定年份的静态的属性描述,而对于形态特征在时间演替方面的塑造规律尚缺乏相关认知. ...
... 腾格里沙漠沙坡头人工固沙植被区经常面临高温、强辐射、干旱等胁迫因子的影响,人工固沙植被的建立为微生物、隐花植物和草本植物的繁育提供了基础[2],改善了该区浅层土壤养分状况[26],有利于浅根系植物的生长发育.其中藓类植物与土壤微生物以及其他生物体通过假根和分泌物等与土壤颗粒胶结形成生物土壤结皮[1],而生物结皮可以改变土壤表面粗糙度、土壤温度、湿度和养分含量等土壤性质,进而影响维管植物种子的萌发、定植和存活.此外,生物结皮还会影响浅层土壤含水量,从而增加浅根系草本植物物种的丰富度和生物量[34-35].前人对沙坡头人工固沙植被区的群落演替特征做了大量的研究和分析,发现生物结皮改变了土壤水分的时空格局和再分配特征,导致固沙植被区植被从高盖度的旱生灌木群落向以低盖度的浅根草本植物为主的复杂群落转变[34].与深根系灌木相比,一年生草本植物与生物结皮生态位更为接近[34],当样地内草本植物十分发达时,会与藓类植物产生种间竞争,此时,即便藓类植物不断调节自身资源获取能力以适应环境,但仍然处于劣势.过多的草本植物抑制了藓类植物的生长,恶劣的环境条件和低养分获取导致藓类植物的叶面积、叶宽、叶长、叶尖长、生物量及密度下降 [36-37].因此,我们推测该区藓类植物在演替后一阶段同样被样地内草本植物抑制了生长,从而导致各形态指标普遍降低.其中叶尖长和叶长与最大光合速率、最大叶绿素荧光相关,这也表明在演替后一阶段藓类植物光合竞争能力有所减弱[14]. ...
Responses of biological soil crusts to sand burial in a revegetated area of the Tengger Desert,Northern China
2
2008
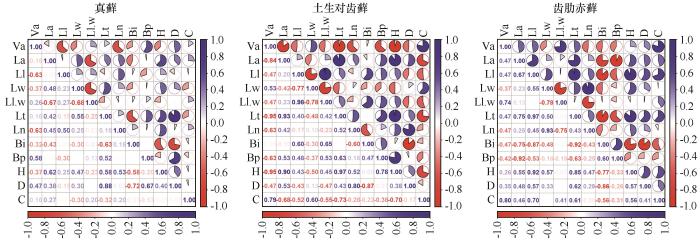
... 研究地点位于腾格里沙漠东南缘(37.45°N,104.95°E,海拔1 330 m),属于温带荒漠化草原向草原化荒漠的过渡区.自1956年起,该区域初步建立了人工植被防护体系,以保护包兰铁路线免受危害[15].后又分别于1964、1981、1987年逐步扩建,有效遏制了沙埋对铁路的威胁.该区域植被以人工种植的花棒(Hedysarum scoparium)、油蒿(Artemisia ordosic)、柠条锦鸡儿(Caragana korshinskii)等灌木为主,且伴有沙米(Agriophyllum squarrosum)和画眉草(Eragrostis pilosa)等沙区草本植物[16].经过近70年的演替,隐花植物(藻类、地衣和藓类)逐渐形成并发育,总盖度已超过70%.藓类以真藓、土生对齿藓、齿肋赤藓为主.腾格里沙漠沙坡头人工固沙植被区是人类逆转沙漠化的成功典范,流动沙丘已成功转化为多产、稳定的生态系统[17]. ...
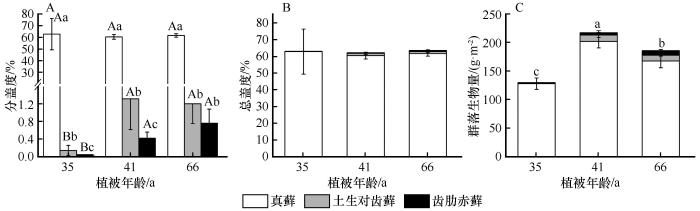
... 藓类植物是陆地植物的第二大多样性类群,虽然和维管植物有着显著的差别,但是在演替进程中所表现出来的对环境适应的特征却与维管植物呈现了相似的规律.比如,密度制约现象.此外,齿肋赤藓的生长环境优于真藓,其株高、生物量、叶面积等指标值大于真藓,表现出更强的资源竞争能力.而真藓分布在地势相对平缓的区域,受风扰、沙埋等因素的干扰较大,受到的干旱、风蚀胁迫更强[15],且真藓个体、生物量和株高远小于齿肋赤藓,即表现出较弱资源竞争能力,但是其密度却是齿肋赤藓的数倍,即反映出真藓的繁殖能力更强,以“量”应对逆境胁迫.因此,我们认为真藓倾向于“r”对策者(抗逆性较强),而齿肋赤藓倾向于“K”对策者(资源竞争能力更强).从整体特征来看,腾格里沙漠人工固沙植被区藓类植物在生态系统修复过程中普遍经历了由茎叶体小、密度大向茎叶体大、密度小的演变趋势. ...
Antagonistic effects of drought and sand burial enable the survival of the biocrust moss Bryum argenteum in an arid sandy desert
1
2018
... 研究地点位于腾格里沙漠东南缘(37.45°N,104.95°E,海拔1 330 m),属于温带荒漠化草原向草原化荒漠的过渡区.自1956年起,该区域初步建立了人工植被防护体系,以保护包兰铁路线免受危害[15].后又分别于1964、1981、1987年逐步扩建,有效遏制了沙埋对铁路的威胁.该区域植被以人工种植的花棒(Hedysarum scoparium)、油蒿(Artemisia ordosic)、柠条锦鸡儿(Caragana korshinskii)等灌木为主,且伴有沙米(Agriophyllum squarrosum)和画眉草(Eragrostis pilosa)等沙区草本植物[16].经过近70年的演替,隐花植物(藻类、地衣和藓类)逐渐形成并发育,总盖度已超过70%.藓类以真藓、土生对齿藓、齿肋赤藓为主.腾格里沙漠沙坡头人工固沙植被区是人类逆转沙漠化的成功典范,流动沙丘已成功转化为多产、稳定的生态系统[17]. ...
Cryptogam diversity and formation of soil crusts in temperate desert
1
2004
... 研究地点位于腾格里沙漠东南缘(37.45°N,104.95°E,海拔1 330 m),属于温带荒漠化草原向草原化荒漠的过渡区.自1956年起,该区域初步建立了人工植被防护体系,以保护包兰铁路线免受危害[15].后又分别于1964、1981、1987年逐步扩建,有效遏制了沙埋对铁路的威胁.该区域植被以人工种植的花棒(Hedysarum scoparium)、油蒿(Artemisia ordosic)、柠条锦鸡儿(Caragana korshinskii)等灌木为主,且伴有沙米(Agriophyllum squarrosum)和画眉草(Eragrostis pilosa)等沙区草本植物[16].经过近70年的演替,隐花植物(藻类、地衣和藓类)逐渐形成并发育,总盖度已超过70%.藓类以真藓、土生对齿藓、齿肋赤藓为主.腾格里沙漠沙坡头人工固沙植被区是人类逆转沙漠化的成功典范,流动沙丘已成功转化为多产、稳定的生态系统[17]. ...
腾格里沙漠人工固沙植被演替过程中生物土壤结皮归一化植被指数的变化特征
1
2017
... 参照赵芸等[18]的针点样方法测量真藓、土生对齿藓以及齿肋赤藓的分盖度.以不同演替阶段的藓类植物的盖度为权重进行加权计算,得到群落水平指标. ...
Variation in leaf traits at different altitudes reflects the adaptive strategy of plants to environmental changes
1
2020
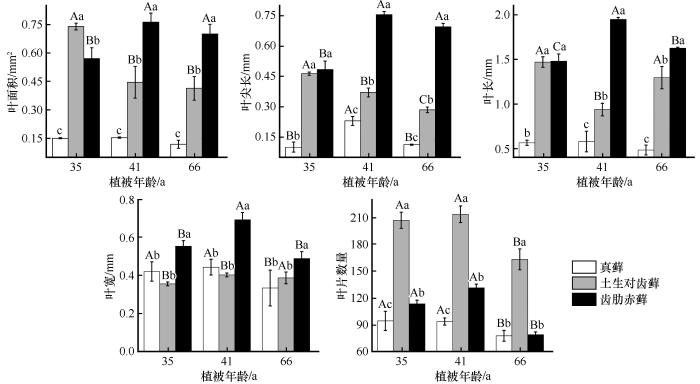
... 叶片的形态特征是植物体功能性状的重要体现,能够直观地反映植物对外界环境的响应与适应[19].本研究分析了腾格里沙漠东南缘不同年代人工固沙植被区(1956、1981、1987年)内3种藓叶片形态的变化特征.我们发现随着植被年龄的增加,3种藓的叶片形态均发生了变化,这可能与土壤养分状况的改变有关.沙坡头人工固沙植被区浅层土壤(0~5 cm)环境状况随着植被年龄的增加而逐渐改善[20].在土壤环境改善后,土壤中N、P含量的增加会导致藓类植物的叶面积、叶尖长以及叶长和叶宽等逐渐增大[21-22].而本研究发现,随着植被年龄的增加,3种藓的叶面积、叶尖长、叶片数量等形态指标在演替后期(66年)时减少了(图1).我们推测可能有两个方面的原因;第一,虽然随着植被年龄的增加,浅层土壤养分状况逐渐改善,但是藓类植物实际可获取的养分并没有相应地增加;第二,较小的叶片形态能够提高水分的利用效率进而增加了植株的耐旱性[23].维管植物在生长发育过程中叶形态变化的相关研究表明,即使在环境条件改善时,植物仍然会通过减小叶面积的方式来增强防御能力[24].叶片大小体现了植物光合产能与逆境适应间的权衡[25],因此我们的研究结果中3种藓的叶面积、叶尖长等指标值的减小可能也是为了增强防御能力. ...
沙坡头人工植被年龄为过程的土壤呼吸特征
2
2012
... 叶片的形态特征是植物体功能性状的重要体现,能够直观地反映植物对外界环境的响应与适应[19].本研究分析了腾格里沙漠东南缘不同年代人工固沙植被区(1956、1981、1987年)内3种藓叶片形态的变化特征.我们发现随着植被年龄的增加,3种藓的叶片形态均发生了变化,这可能与土壤养分状况的改变有关.沙坡头人工固沙植被区浅层土壤(0~5 cm)环境状况随着植被年龄的增加而逐渐改善[20].在土壤环境改善后,土壤中N、P含量的增加会导致藓类植物的叶面积、叶尖长以及叶长和叶宽等逐渐增大[21-22].而本研究发现,随着植被年龄的增加,3种藓的叶面积、叶尖长、叶片数量等形态指标在演替后期(66年)时减少了(图1).我们推测可能有两个方面的原因;第一,虽然随着植被年龄的增加,浅层土壤养分状况逐渐改善,但是藓类植物实际可获取的养分并没有相应地增加;第二,较小的叶片形态能够提高水分的利用效率进而增加了植株的耐旱性[23].维管植物在生长发育过程中叶形态变化的相关研究表明,即使在环境条件改善时,植物仍然会通过减小叶面积的方式来增强防御能力[24].叶片大小体现了植物光合产能与逆境适应间的权衡[25],因此我们的研究结果中3种藓的叶面积、叶尖长等指标值的减小可能也是为了增强防御能力. ...
... 群落性状演替在空间上的差异可以反映种群性状在时间上的差异[32],因此将3种藓的种群性状和群落演替各阶段结合起来分析,有助于理解群落的性状及其发展趋势.相关分析的结果表明,3种藓的盖度与植被年龄具有正相关性,其中土生对齿藓和齿肋赤藓相关性系数大于真藓(图5).真藓在固沙植被区属于藓类植物中的先锋种,对土壤稳定和改善土壤养分状况有重要作用,在固沙植被区内总盖度超过了70%,显著大于土生对齿藓和齿肋赤藓[20],在3种藓中占主导地位,决定了群落生物量和总盖度的变化(图4).齿肋赤藓属于该区藓类植物中演替等级较高的物种,其盖度在演替早中期显著小于土生对齿藓,在演替后期和土生对齿藓不存在显著性差异(图3),这是因为齿肋赤藓资源获取能力较强,随着土壤环境的逐渐改善,齿肋赤藓的拓殖速度不断加快. ...
Plant height as a simple predictor of the root to shoot ratio:evidence from alpine grasslands on the Tibetan Plateau
1
2008
... 叶片的形态特征是植物体功能性状的重要体现,能够直观地反映植物对外界环境的响应与适应[19].本研究分析了腾格里沙漠东南缘不同年代人工固沙植被区(1956、1981、1987年)内3种藓叶片形态的变化特征.我们发现随着植被年龄的增加,3种藓的叶片形态均发生了变化,这可能与土壤养分状况的改变有关.沙坡头人工固沙植被区浅层土壤(0~5 cm)环境状况随着植被年龄的增加而逐渐改善[20].在土壤环境改善后,土壤中N、P含量的增加会导致藓类植物的叶面积、叶尖长以及叶长和叶宽等逐渐增大[21-22].而本研究发现,随着植被年龄的增加,3种藓的叶面积、叶尖长、叶片数量等形态指标在演替后期(66年)时减少了(图1).我们推测可能有两个方面的原因;第一,虽然随着植被年龄的增加,浅层土壤养分状况逐渐改善,但是藓类植物实际可获取的养分并没有相应地增加;第二,较小的叶片形态能够提高水分的利用效率进而增加了植株的耐旱性[23].维管植物在生长发育过程中叶形态变化的相关研究表明,即使在环境条件改善时,植物仍然会通过减小叶面积的方式来增强防御能力[24].叶片大小体现了植物光合产能与逆境适应间的权衡[25],因此我们的研究结果中3种藓的叶面积、叶尖长等指标值的减小可能也是为了增强防御能力. ...
Morphological traits of Bryum argenteum and its response to environmental variation in arid and semi-arid areas of Tibet
2
2019
... 叶片的形态特征是植物体功能性状的重要体现,能够直观地反映植物对外界环境的响应与适应[19].本研究分析了腾格里沙漠东南缘不同年代人工固沙植被区(1956、1981、1987年)内3种藓叶片形态的变化特征.我们发现随着植被年龄的增加,3种藓的叶片形态均发生了变化,这可能与土壤养分状况的改变有关.沙坡头人工固沙植被区浅层土壤(0~5 cm)环境状况随着植被年龄的增加而逐渐改善[20].在土壤环境改善后,土壤中N、P含量的增加会导致藓类植物的叶面积、叶尖长以及叶长和叶宽等逐渐增大[21-22].而本研究发现,随着植被年龄的增加,3种藓的叶面积、叶尖长、叶片数量等形态指标在演替后期(66年)时减少了(图1).我们推测可能有两个方面的原因;第一,虽然随着植被年龄的增加,浅层土壤养分状况逐渐改善,但是藓类植物实际可获取的养分并没有相应地增加;第二,较小的叶片形态能够提高水分的利用效率进而增加了植株的耐旱性[23].维管植物在生长发育过程中叶形态变化的相关研究表明,即使在环境条件改善时,植物仍然会通过减小叶面积的方式来增强防御能力[24].叶片大小体现了植物光合产能与逆境适应间的权衡[25],因此我们的研究结果中3种藓的叶面积、叶尖长等指标值的减小可能也是为了增强防御能力. ...
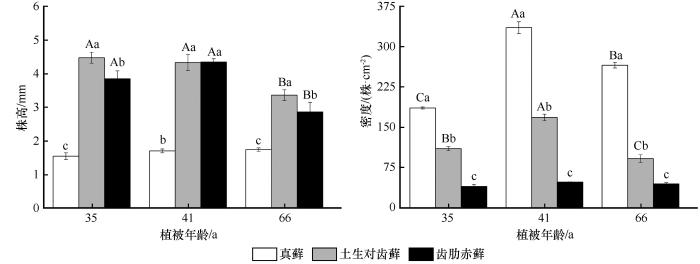
... 干旱区藓类植物属于典型的变水性植物,在野外干燥环境中以“suntan (可塑性深色色素积累)”形态来适应强紫外辐射,从而提升抗旱性[28],而湿润后叶片则快速展开,植株膨胀恢复成黄绿色.这种变水特性是藓类植物强耐旱性特征.仝治国[29]发现旱藓(Indusiella thianschanica)和墙藓(Tortula subulate)也呈现相同的特征.这说明“suntan”形态和变水特征是干旱区藓类植物共有的抗逆策略.株高是藓类植物个体环境适应策略最直接的表型特征.在本研究中我们发现真藓的平均株高为2.02 mm,显著低于Wang等[22]2019年在西藏测量的真藓的平均株高(5.01 mm).株高在藓类植物间变化很大,这种差异可能是由于地理位置造成的气候不同引起的[30].而真藓的株高和生物量普遍小于土生对齿藓和齿肋赤藓,这可能是由于土生对齿藓和齿肋赤藓出现于真藓之后,生长环境相对较好[31]. ...
1
2008
... 叶片的形态特征是植物体功能性状的重要体现,能够直观地反映植物对外界环境的响应与适应[19].本研究分析了腾格里沙漠东南缘不同年代人工固沙植被区(1956、1981、1987年)内3种藓叶片形态的变化特征.我们发现随着植被年龄的增加,3种藓的叶片形态均发生了变化,这可能与土壤养分状况的改变有关.沙坡头人工固沙植被区浅层土壤(0~5 cm)环境状况随着植被年龄的增加而逐渐改善[20].在土壤环境改善后,土壤中N、P含量的增加会导致藓类植物的叶面积、叶尖长以及叶长和叶宽等逐渐增大[21-22].而本研究发现,随着植被年龄的增加,3种藓的叶面积、叶尖长、叶片数量等形态指标在演替后期(66年)时减少了(图1).我们推测可能有两个方面的原因;第一,虽然随着植被年龄的增加,浅层土壤养分状况逐渐改善,但是藓类植物实际可获取的养分并没有相应地增加;第二,较小的叶片形态能够提高水分的利用效率进而增加了植株的耐旱性[23].维管植物在生长发育过程中叶形态变化的相关研究表明,即使在环境条件改善时,植物仍然会通过减小叶面积的方式来增强防御能力[24].叶片大小体现了植物光合产能与逆境适应间的权衡[25],因此我们的研究结果中3种藓的叶面积、叶尖长等指标值的减小可能也是为了增强防御能力. ...
毛乌素沙地沙丘不同固定阶段黑沙蒿叶性状特征
1
2022
... 叶片的形态特征是植物体功能性状的重要体现,能够直观地反映植物对外界环境的响应与适应[19].本研究分析了腾格里沙漠东南缘不同年代人工固沙植被区(1956、1981、1987年)内3种藓叶片形态的变化特征.我们发现随着植被年龄的增加,3种藓的叶片形态均发生了变化,这可能与土壤养分状况的改变有关.沙坡头人工固沙植被区浅层土壤(0~5 cm)环境状况随着植被年龄的增加而逐渐改善[20].在土壤环境改善后,土壤中N、P含量的增加会导致藓类植物的叶面积、叶尖长以及叶长和叶宽等逐渐增大[21-22].而本研究发现,随着植被年龄的增加,3种藓的叶面积、叶尖长、叶片数量等形态指标在演替后期(66年)时减少了(图1).我们推测可能有两个方面的原因;第一,虽然随着植被年龄的增加,浅层土壤养分状况逐渐改善,但是藓类植物实际可获取的养分并没有相应地增加;第二,较小的叶片形态能够提高水分的利用效率进而增加了植株的耐旱性[23].维管植物在生长发育过程中叶形态变化的相关研究表明,即使在环境条件改善时,植物仍然会通过减小叶面积的方式来增强防御能力[24].叶片大小体现了植物光合产能与逆境适应间的权衡[25],因此我们的研究结果中3种藓的叶面积、叶尖长等指标值的减小可能也是为了增强防御能力. ...
植物叶片形态的生态功能、地理分布与成因
1
2021
... 叶片的形态特征是植物体功能性状的重要体现,能够直观地反映植物对外界环境的响应与适应[19].本研究分析了腾格里沙漠东南缘不同年代人工固沙植被区(1956、1981、1987年)内3种藓叶片形态的变化特征.我们发现随着植被年龄的增加,3种藓的叶片形态均发生了变化,这可能与土壤养分状况的改变有关.沙坡头人工固沙植被区浅层土壤(0~5 cm)环境状况随着植被年龄的增加而逐渐改善[20].在土壤环境改善后,土壤中N、P含量的增加会导致藓类植物的叶面积、叶尖长以及叶长和叶宽等逐渐增大[21-22].而本研究发现,随着植被年龄的增加,3种藓的叶面积、叶尖长、叶片数量等形态指标在演替后期(66年)时减少了(图1).我们推测可能有两个方面的原因;第一,虽然随着植被年龄的增加,浅层土壤养分状况逐渐改善,但是藓类植物实际可获取的养分并没有相应地增加;第二,较小的叶片形态能够提高水分的利用效率进而增加了植株的耐旱性[23].维管植物在生长发育过程中叶形态变化的相关研究表明,即使在环境条件改善时,植物仍然会通过减小叶面积的方式来增强防御能力[24].叶片大小体现了植物光合产能与逆境适应间的权衡[25],因此我们的研究结果中3种藓的叶面积、叶尖长等指标值的减小可能也是为了增强防御能力. ...
Effects of sand burial on dew deposition on moss soil crust in a revegetated area of the Tengger Desert,Northern China
2
2014
... 固沙植被建立后,植被区内依次出现了真藓、土生对齿藓和齿肋赤藓[26],且这3种藓的叶面积、叶尖长、叶长和叶宽等逐渐增大,这与前人的研究结果一致.在不同建殖阶段的对比方面,齿肋赤藓的叶面积、叶尖长等形态指标在建殖第35年时小于土生对齿藓,这可能是因为齿肋赤藓距今为止最长发育年龄只有2年,可能还未处于幼龄期,故而叶片形态较小[27]. ...
... 腾格里沙漠沙坡头人工固沙植被区经常面临高温、强辐射、干旱等胁迫因子的影响,人工固沙植被的建立为微生物、隐花植物和草本植物的繁育提供了基础[2],改善了该区浅层土壤养分状况[26],有利于浅根系植物的生长发育.其中藓类植物与土壤微生物以及其他生物体通过假根和分泌物等与土壤颗粒胶结形成生物土壤结皮[1],而生物结皮可以改变土壤表面粗糙度、土壤温度、湿度和养分含量等土壤性质,进而影响维管植物种子的萌发、定植和存活.此外,生物结皮还会影响浅层土壤含水量,从而增加浅根系草本植物物种的丰富度和生物量[34-35].前人对沙坡头人工固沙植被区的群落演替特征做了大量的研究和分析,发现生物结皮改变了土壤水分的时空格局和再分配特征,导致固沙植被区植被从高盖度的旱生灌木群落向以低盖度的浅根草本植物为主的复杂群落转变[34].与深根系灌木相比,一年生草本植物与生物结皮生态位更为接近[34],当样地内草本植物十分发达时,会与藓类植物产生种间竞争,此时,即便藓类植物不断调节自身资源获取能力以适应环境,但仍然处于劣势.过多的草本植物抑制了藓类植物的生长,恶劣的环境条件和低养分获取导致藓类植物的叶面积、叶宽、叶长、叶尖长、生物量及密度下降 [36-37].因此,我们推测该区藓类植物在演替后一阶段同样被样地内草本植物抑制了生长,从而导致各形态指标普遍降低.其中叶尖长和叶长与最大光合速率、最大叶绿素荧光相关,这也表明在演替后一阶段藓类植物光合竞争能力有所减弱[14]. ...
沙坡头人工固沙植被演替过程中主要结皮生物生态位和种间关联变化特征
1
2021
... 固沙植被建立后,植被区内依次出现了真藓、土生对齿藓和齿肋赤藓[26],且这3种藓的叶面积、叶尖长、叶长和叶宽等逐渐增大,这与前人的研究结果一致.在不同建殖阶段的对比方面,齿肋赤藓的叶面积、叶尖长等形态指标在建殖第35年时小于土生对齿藓,这可能是因为齿肋赤藓距今为止最长发育年龄只有2年,可能还未处于幼龄期,故而叶片形态较小[27]. ...
Natural ultraviolet radiation exposure alters photosynthetic biology and improves recovery from desiccation in a desert moss
1
2018
... 干旱区藓类植物属于典型的变水性植物,在野外干燥环境中以“suntan (可塑性深色色素积累)”形态来适应强紫外辐射,从而提升抗旱性[28],而湿润后叶片则快速展开,植株膨胀恢复成黄绿色.这种变水特性是藓类植物强耐旱性特征.仝治国[29]发现旱藓(Indusiella thianschanica)和墙藓(Tortula subulate)也呈现相同的特征.这说明“suntan”形态和变水特征是干旱区藓类植物共有的抗逆策略.株高是藓类植物个体环境适应策略最直接的表型特征.在本研究中我们发现真藓的平均株高为2.02 mm,显著低于Wang等[22]2019年在西藏测量的真藓的平均株高(5.01 mm).株高在藓类植物间变化很大,这种差异可能是由于地理位置造成的气候不同引起的[30].而真藓的株高和生物量普遍小于土生对齿藓和齿肋赤藓,这可能是由于土生对齿藓和齿肋赤藓出现于真藓之后,生长环境相对较好[31]. ...
中国几种旱生藓类的新分布
1
1963
... 干旱区藓类植物属于典型的变水性植物,在野外干燥环境中以“suntan (可塑性深色色素积累)”形态来适应强紫外辐射,从而提升抗旱性[28],而湿润后叶片则快速展开,植株膨胀恢复成黄绿色.这种变水特性是藓类植物强耐旱性特征.仝治国[29]发现旱藓(Indusiella thianschanica)和墙藓(Tortula subulate)也呈现相同的特征.这说明“suntan”形态和变水特征是干旱区藓类植物共有的抗逆策略.株高是藓类植物个体环境适应策略最直接的表型特征.在本研究中我们发现真藓的平均株高为2.02 mm,显著低于Wang等[22]2019年在西藏测量的真藓的平均株高(5.01 mm).株高在藓类植物间变化很大,这种差异可能是由于地理位置造成的气候不同引起的[30].而真藓的株高和生物量普遍小于土生对齿藓和齿肋赤藓,这可能是由于土生对齿藓和齿肋赤藓出现于真藓之后,生长环境相对较好[31]. ...
Temperature,not precipitation,drives the morphological traits of Didymodon rigidulus in Tibet
1
2021
... 干旱区藓类植物属于典型的变水性植物,在野外干燥环境中以“suntan (可塑性深色色素积累)”形态来适应强紫外辐射,从而提升抗旱性[28],而湿润后叶片则快速展开,植株膨胀恢复成黄绿色.这种变水特性是藓类植物强耐旱性特征.仝治国[29]发现旱藓(Indusiella thianschanica)和墙藓(Tortula subulate)也呈现相同的特征.这说明“suntan”形态和变水特征是干旱区藓类植物共有的抗逆策略.株高是藓类植物个体环境适应策略最直接的表型特征.在本研究中我们发现真藓的平均株高为2.02 mm,显著低于Wang等[22]2019年在西藏测量的真藓的平均株高(5.01 mm).株高在藓类植物间变化很大,这种差异可能是由于地理位置造成的气候不同引起的[30].而真藓的株高和生物量普遍小于土生对齿藓和齿肋赤藓,这可能是由于土生对齿藓和齿肋赤藓出现于真藓之后,生长环境相对较好[31]. ...
腾格里沙漠固定沙丘结皮层藓类植物的生态功能及与土壤环境因子的关系
2
2005
... 干旱区藓类植物属于典型的变水性植物,在野外干燥环境中以“suntan (可塑性深色色素积累)”形态来适应强紫外辐射,从而提升抗旱性[28],而湿润后叶片则快速展开,植株膨胀恢复成黄绿色.这种变水特性是藓类植物强耐旱性特征.仝治国[29]发现旱藓(Indusiella thianschanica)和墙藓(Tortula subulate)也呈现相同的特征.这说明“suntan”形态和变水特征是干旱区藓类植物共有的抗逆策略.株高是藓类植物个体环境适应策略最直接的表型特征.在本研究中我们发现真藓的平均株高为2.02 mm,显著低于Wang等[22]2019年在西藏测量的真藓的平均株高(5.01 mm).株高在藓类植物间变化很大,这种差异可能是由于地理位置造成的气候不同引起的[30].而真藓的株高和生物量普遍小于土生对齿藓和齿肋赤藓,这可能是由于土生对齿藓和齿肋赤藓出现于真藓之后,生长环境相对较好[31]. ...
... 随着植被年龄的增加,3种藓生物量和株高并未表现出单调增加的趋势,这与徐杰等[31]对该区藓类植物的研究结果有所不同,他们发现随着植被年龄的增加藓类植物生物量逐渐增大.而目前已有关于沙坡头人工固沙植被区的研究并未表述过类似现象,造成这一变化的原因是什么呢?我们推测藓类植物间(内)可能发生了竞争现象. ...
浙江省松阳县黄山松种群的密度与生物量动态
3
1991
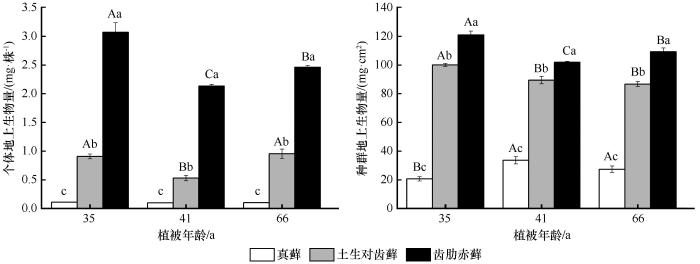
... 种群的密度也是藓类植物环境适应性的重要功能性状.干旱区藓类植物属于耐旱型藓类,它们通常聚集丛生呈垫状分布,从而减缓水分丧失.这与高寒区植物体垫状丛生现象相似.本研究中,随着植被年龄的增加,3种藓在种群密度上发生了显著变化,在建殖初期阶段到中期阶段密度较高,而后随着演替的进程却不断降低(图2).同样的现象也在森林演替的先锋种——黄山松种群中被发现[32].黄山松种群密度在演替中期达到饱和,在演替后期由于自疏现象以及阔叶树种的入侵,种群密度减少.此外,张利权[32]对黄山松种群生物量进行了统计,发现生物量的变化呈现和密度相同的特征,而本研究中不同藓类种群密度与种群生物量呈现不同的变化特征.但是,3种藓个体地上生物量与密度均呈现负相关性(图5),表现为种群密度越大,植株个体生物量越小.株高与种群密度存在较强的正相关性(图2、6),这一现象与维管植物一致.孙强等[33]2005年对泥炭藓(Sphagnum palustre)的研究时也曾发现了同样的现象,他们分析在种群拥挤的情况下,植株间除了竞争养分之外还要竞争生存空间,在垂直空间上加速生长以获得更多的光照.本研究中真藓和齿肋赤藓均表现出上述类似现象. ...
... [32]对黄山松种群生物量进行了统计,发现生物量的变化呈现和密度相同的特征,而本研究中不同藓类种群密度与种群生物量呈现不同的变化特征.但是,3种藓个体地上生物量与密度均呈现负相关性(图5),表现为种群密度越大,植株个体生物量越小.株高与种群密度存在较强的正相关性(图2、6),这一现象与维管植物一致.孙强等[33]2005年对泥炭藓(Sphagnum palustre)的研究时也曾发现了同样的现象,他们分析在种群拥挤的情况下,植株间除了竞争养分之外还要竞争生存空间,在垂直空间上加速生长以获得更多的光照.本研究中真藓和齿肋赤藓均表现出上述类似现象. ...
... 群落性状演替在空间上的差异可以反映种群性状在时间上的差异[32],因此将3种藓的种群性状和群落演替各阶段结合起来分析,有助于理解群落的性状及其发展趋势.相关分析的结果表明,3种藓的盖度与植被年龄具有正相关性,其中土生对齿藓和齿肋赤藓相关性系数大于真藓(图5).真藓在固沙植被区属于藓类植物中的先锋种,对土壤稳定和改善土壤养分状况有重要作用,在固沙植被区内总盖度超过了70%,显著大于土生对齿藓和齿肋赤藓[20],在3种藓中占主导地位,决定了群落生物量和总盖度的变化(图4).齿肋赤藓属于该区藓类植物中演替等级较高的物种,其盖度在演替早中期显著小于土生对齿藓,在演替后期和土生对齿藓不存在显著性差异(图3),这是因为齿肋赤藓资源获取能力较强,随着土壤环境的逐渐改善,齿肋赤藓的拓殖速度不断加快. ...
哈泥贫营养泥炭沼泽毛壁泥炭藓种群密度制约初探
2
2005
... 种群的密度也是藓类植物环境适应性的重要功能性状.干旱区藓类植物属于耐旱型藓类,它们通常聚集丛生呈垫状分布,从而减缓水分丧失.这与高寒区植物体垫状丛生现象相似.本研究中,随着植被年龄的增加,3种藓在种群密度上发生了显著变化,在建殖初期阶段到中期阶段密度较高,而后随着演替的进程却不断降低(图2).同样的现象也在森林演替的先锋种——黄山松种群中被发现[32].黄山松种群密度在演替中期达到饱和,在演替后期由于自疏现象以及阔叶树种的入侵,种群密度减少.此外,张利权[32]对黄山松种群生物量进行了统计,发现生物量的变化呈现和密度相同的特征,而本研究中不同藓类种群密度与种群生物量呈现不同的变化特征.但是,3种藓个体地上生物量与密度均呈现负相关性(图5),表现为种群密度越大,植株个体生物量越小.株高与种群密度存在较强的正相关性(图2、6),这一现象与维管植物一致.孙强等[33]2005年对泥炭藓(Sphagnum palustre)的研究时也曾发现了同样的现象,他们分析在种群拥挤的情况下,植株间除了竞争养分之外还要竞争生存空间,在垂直空间上加速生长以获得更多的光照.本研究中真藓和齿肋赤藓均表现出上述类似现象. ...
... 密度制约是发生在相邻植株间的调节机制,反映了个体与种群数量的变化[38-39].本研究中3种藓密度随建殖进程呈现出先增加后减少的变化特征(图2).对3种藓个体和种群性状分析表明,种群密度越大个体生物量越小,3种藓间发生了类似于维管植物的由种内竞争导致的自疏现象(图6).孙强等[33]发现生长于哈尼贫营养沼泽的毛壁泥炭藓(Sphagnum imbricatum)间存在明显的自疏现象,每株泥炭藓的生物量随密度增加而减少,而株高随密度增加而增加,这与本研究的结果基本一致;zamfir等[40]对7种藓类植物种群之间的相对竞争强度进行了分析,明确了竞争等级.这说明密度制约可能是生长在各个地区的藓类植物间普遍存在的现象. ...
Do biological soil crusts determine vegetation changes in sandy deserts?Implications for managing artificial vegetation
3
2010
... 腾格里沙漠沙坡头人工固沙植被区经常面临高温、强辐射、干旱等胁迫因子的影响,人工固沙植被的建立为微生物、隐花植物和草本植物的繁育提供了基础[2],改善了该区浅层土壤养分状况[26],有利于浅根系植物的生长发育.其中藓类植物与土壤微生物以及其他生物体通过假根和分泌物等与土壤颗粒胶结形成生物土壤结皮[1],而生物结皮可以改变土壤表面粗糙度、土壤温度、湿度和养分含量等土壤性质,进而影响维管植物种子的萌发、定植和存活.此外,生物结皮还会影响浅层土壤含水量,从而增加浅根系草本植物物种的丰富度和生物量[34-35].前人对沙坡头人工固沙植被区的群落演替特征做了大量的研究和分析,发现生物结皮改变了土壤水分的时空格局和再分配特征,导致固沙植被区植被从高盖度的旱生灌木群落向以低盖度的浅根草本植物为主的复杂群落转变[34].与深根系灌木相比,一年生草本植物与生物结皮生态位更为接近[34],当样地内草本植物十分发达时,会与藓类植物产生种间竞争,此时,即便藓类植物不断调节自身资源获取能力以适应环境,但仍然处于劣势.过多的草本植物抑制了藓类植物的生长,恶劣的环境条件和低养分获取导致藓类植物的叶面积、叶宽、叶长、叶尖长、生物量及密度下降 [36-37].因此,我们推测该区藓类植物在演替后一阶段同样被样地内草本植物抑制了生长,从而导致各形态指标普遍降低.其中叶尖长和叶长与最大光合速率、最大叶绿素荧光相关,这也表明在演替后一阶段藓类植物光合竞争能力有所减弱[14]. ...
... [34].与深根系灌木相比,一年生草本植物与生物结皮生态位更为接近[34],当样地内草本植物十分发达时,会与藓类植物产生种间竞争,此时,即便藓类植物不断调节自身资源获取能力以适应环境,但仍然处于劣势.过多的草本植物抑制了藓类植物的生长,恶劣的环境条件和低养分获取导致藓类植物的叶面积、叶宽、叶长、叶尖长、生物量及密度下降 [36-37].因此,我们推测该区藓类植物在演替后一阶段同样被样地内草本植物抑制了生长,从而导致各形态指标普遍降低.其中叶尖长和叶长与最大光合速率、最大叶绿素荧光相关,这也表明在演替后一阶段藓类植物光合竞争能力有所减弱[14]. ...
... [34],当样地内草本植物十分发达时,会与藓类植物产生种间竞争,此时,即便藓类植物不断调节自身资源获取能力以适应环境,但仍然处于劣势.过多的草本植物抑制了藓类植物的生长,恶劣的环境条件和低养分获取导致藓类植物的叶面积、叶宽、叶长、叶尖长、生物量及密度下降 [36-37].因此,我们推测该区藓类植物在演替后一阶段同样被样地内草本植物抑制了生长,从而导致各形态指标普遍降低.其中叶尖长和叶长与最大光合速率、最大叶绿素荧光相关,这也表明在演替后一阶段藓类植物光合竞争能力有所减弱[14]. ...
Degradation and reorganization of thylakoid protein complexes of Bryum argenteum in response to dehydration and rehydration
1
2014
... 腾格里沙漠沙坡头人工固沙植被区经常面临高温、强辐射、干旱等胁迫因子的影响,人工固沙植被的建立为微生物、隐花植物和草本植物的繁育提供了基础[2],改善了该区浅层土壤养分状况[26],有利于浅根系植物的生长发育.其中藓类植物与土壤微生物以及其他生物体通过假根和分泌物等与土壤颗粒胶结形成生物土壤结皮[1],而生物结皮可以改变土壤表面粗糙度、土壤温度、湿度和养分含量等土壤性质,进而影响维管植物种子的萌发、定植和存活.此外,生物结皮还会影响浅层土壤含水量,从而增加浅根系草本植物物种的丰富度和生物量[34-35].前人对沙坡头人工固沙植被区的群落演替特征做了大量的研究和分析,发现生物结皮改变了土壤水分的时空格局和再分配特征,导致固沙植被区植被从高盖度的旱生灌木群落向以低盖度的浅根草本植物为主的复杂群落转变[34].与深根系灌木相比,一年生草本植物与生物结皮生态位更为接近[34],当样地内草本植物十分发达时,会与藓类植物产生种间竞争,此时,即便藓类植物不断调节自身资源获取能力以适应环境,但仍然处于劣势.过多的草本植物抑制了藓类植物的生长,恶劣的环境条件和低养分获取导致藓类植物的叶面积、叶宽、叶长、叶尖长、生物量及密度下降 [36-37].因此,我们推测该区藓类植物在演替后一阶段同样被样地内草本植物抑制了生长,从而导致各形态指标普遍降低.其中叶尖长和叶长与最大光合速率、最大叶绿素荧光相关,这也表明在演替后一阶段藓类植物光合竞争能力有所减弱[14]. ...
Positive interactions between an exotic invader and moss biocrusts vary across life stage and correspond with the effect of water pulses on soil nitrogen
1
2021
... 腾格里沙漠沙坡头人工固沙植被区经常面临高温、强辐射、干旱等胁迫因子的影响,人工固沙植被的建立为微生物、隐花植物和草本植物的繁育提供了基础[2],改善了该区浅层土壤养分状况[26],有利于浅根系植物的生长发育.其中藓类植物与土壤微生物以及其他生物体通过假根和分泌物等与土壤颗粒胶结形成生物土壤结皮[1],而生物结皮可以改变土壤表面粗糙度、土壤温度、湿度和养分含量等土壤性质,进而影响维管植物种子的萌发、定植和存活.此外,生物结皮还会影响浅层土壤含水量,从而增加浅根系草本植物物种的丰富度和生物量[34-35].前人对沙坡头人工固沙植被区的群落演替特征做了大量的研究和分析,发现生物结皮改变了土壤水分的时空格局和再分配特征,导致固沙植被区植被从高盖度的旱生灌木群落向以低盖度的浅根草本植物为主的复杂群落转变[34].与深根系灌木相比,一年生草本植物与生物结皮生态位更为接近[34],当样地内草本植物十分发达时,会与藓类植物产生种间竞争,此时,即便藓类植物不断调节自身资源获取能力以适应环境,但仍然处于劣势.过多的草本植物抑制了藓类植物的生长,恶劣的环境条件和低养分获取导致藓类植物的叶面积、叶宽、叶长、叶尖长、生物量及密度下降 [36-37].因此,我们推测该区藓类植物在演替后一阶段同样被样地内草本植物抑制了生长,从而导致各形态指标普遍降低.其中叶尖长和叶长与最大光合速率、最大叶绿素荧光相关,这也表明在演替后一阶段藓类植物光合竞争能力有所减弱[14]. ...
不同生境对藏东南地面生苔藓多样性和生物量的影响
1
2022
... 腾格里沙漠沙坡头人工固沙植被区经常面临高温、强辐射、干旱等胁迫因子的影响,人工固沙植被的建立为微生物、隐花植物和草本植物的繁育提供了基础[2],改善了该区浅层土壤养分状况[26],有利于浅根系植物的生长发育.其中藓类植物与土壤微生物以及其他生物体通过假根和分泌物等与土壤颗粒胶结形成生物土壤结皮[1],而生物结皮可以改变土壤表面粗糙度、土壤温度、湿度和养分含量等土壤性质,进而影响维管植物种子的萌发、定植和存活.此外,生物结皮还会影响浅层土壤含水量,从而增加浅根系草本植物物种的丰富度和生物量[34-35].前人对沙坡头人工固沙植被区的群落演替特征做了大量的研究和分析,发现生物结皮改变了土壤水分的时空格局和再分配特征,导致固沙植被区植被从高盖度的旱生灌木群落向以低盖度的浅根草本植物为主的复杂群落转变[34].与深根系灌木相比,一年生草本植物与生物结皮生态位更为接近[34],当样地内草本植物十分发达时,会与藓类植物产生种间竞争,此时,即便藓类植物不断调节自身资源获取能力以适应环境,但仍然处于劣势.过多的草本植物抑制了藓类植物的生长,恶劣的环境条件和低养分获取导致藓类植物的叶面积、叶宽、叶长、叶尖长、生物量及密度下降 [36-37].因此,我们推测该区藓类植物在演替后一阶段同样被样地内草本植物抑制了生长,从而导致各形态指标普遍降低.其中叶尖长和叶长与最大光合速率、最大叶绿素荧光相关,这也表明在演替后一阶段藓类植物光合竞争能力有所减弱[14]. ...
喜旱莲子草在增温背景下的密度制约与生长权衡
1
2022
... 密度制约是发生在相邻植株间的调节机制,反映了个体与种群数量的变化[38-39].本研究中3种藓密度随建殖进程呈现出先增加后减少的变化特征(图2).对3种藓个体和种群性状分析表明,种群密度越大个体生物量越小,3种藓间发生了类似于维管植物的由种内竞争导致的自疏现象(图6).孙强等[33]发现生长于哈尼贫营养沼泽的毛壁泥炭藓(Sphagnum imbricatum)间存在明显的自疏现象,每株泥炭藓的生物量随密度增加而减少,而株高随密度增加而增加,这与本研究的结果基本一致;zamfir等[40]对7种藓类植物种群之间的相对竞争强度进行了分析,明确了竞争等级.这说明密度制约可能是生长在各个地区的藓类植物间普遍存在的现象. ...
长白山次生杨桦林种群空间点格局及密度制约效应
1
2021
... 密度制约是发生在相邻植株间的调节机制,反映了个体与种群数量的变化[38-39].本研究中3种藓密度随建殖进程呈现出先增加后减少的变化特征(图2).对3种藓个体和种群性状分析表明,种群密度越大个体生物量越小,3种藓间发生了类似于维管植物的由种内竞争导致的自疏现象(图6).孙强等[33]发现生长于哈尼贫营养沼泽的毛壁泥炭藓(Sphagnum imbricatum)间存在明显的自疏现象,每株泥炭藓的生物量随密度增加而减少,而株高随密度增加而增加,这与本研究的结果基本一致;zamfir等[40]对7种藓类植物种群之间的相对竞争强度进行了分析,明确了竞争等级.这说明密度制约可能是生长在各个地区的藓类植物间普遍存在的现象. ...
The effect of initial density on interactions between bryophytes at individual and community levels
1
2000
... 密度制约是发生在相邻植株间的调节机制,反映了个体与种群数量的变化[38-39].本研究中3种藓密度随建殖进程呈现出先增加后减少的变化特征(图2).对3种藓个体和种群性状分析表明,种群密度越大个体生物量越小,3种藓间发生了类似于维管植物的由种内竞争导致的自疏现象(图6).孙强等[33]发现生长于哈尼贫营养沼泽的毛壁泥炭藓(Sphagnum imbricatum)间存在明显的自疏现象,每株泥炭藓的生物量随密度增加而减少,而株高随密度增加而增加,这与本研究的结果基本一致;zamfir等[40]对7种藓类植物种群之间的相对竞争强度进行了分析,明确了竞争等级.这说明密度制约可能是生长在各个地区的藓类植物间普遍存在的现象. ...
藓类植物相互作用的研究进展
1
2009
... 虽然在严酷的环境中藓类植物间往往以正相互作用为主,但是也不乏竞争实例[41].从演替早期到演替后期,真藓的生物量增加了,土生对齿藓的生物量减少了,齿肋赤藓的生物量先减少后增加(图3);齿肋赤藓的盖度在演替早中期显著小于土生对齿藓,而在后期两者无显著性差异,且齿肋赤藓的盖度单调增加.因此,我们推测藓类植物间的竞争主要发生在土生对齿藓和齿肋赤藓之间,并且随着植被年龄的增加,土壤浅层含水量逐渐增多,由此说明演替后期植物可获取的资源大于演替中期,即藓类植物间在演替中期种内竞争激烈强度大于演替后期,这可能也是藓类植物个体生物量在演替中期最小的原因. ...
Moss functional trait ecology:trends,gaps,and biases in the current literature
1
2024
... 藓类植物的形态可塑性受生物胁迫和非生物胁迫等多种因素的干扰,形态性状需要权衡资源竞争,生长繁殖,即生长策略和适应策略之间的关系[42].形态变化规律可以帮助我们更全面和深入地理解藓类植物适应环境变化的途径和机制.本文较详细地分析了沙坡头人工固沙植被区藓类植物演替过程中的形态变化特征,为揭示藓类植物在演替过程中的适应性奠定了基础,为评估和判断藓类植物及其结皮在变化环境下的稳定性提供了参考,为生态修复过程中藓类植物的物种筛选提供科学依据. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}