羽毛针禾( Stipagrostis pennata 6 -磷酸-1 -差向异构酶基因 SpG6P1E
唐蕊 , 1 , 尹珊 1 , 陈凯露 1 , 李悦涛 1 , 程淋渊 1 , 王景儒 1 , 王斐 , 1 , 2 , 3 , 李榕 , 1 , 2 , 3 , 李鸿彬 1 , 2 , 3
1.石河子大学,生命科学学院,新疆 石河子 832003
2.石河子大学,绿洲城镇与山盆系统生态兵团重点实验室,新疆 石河子 832003
3.石河子大学,新疆植物药资源利用教育部重点实验室,新疆 石河子 832003
Cloning and expression analysis of Glucose -6 -phospho 1 epimerase SpG6P1E in Stipagrostis pennata
Tang Rui , 1 , Yin Shan 1 , Chen Kailu 1 , Li Yuetao 1 , Cheng Linyuan 1 , Wang Jingru 1 , Wang Fei , 1 , 2 , 3 , Li Rong , 1 , 2 , 3 , Li Hongbin 1 , 2 , 3
1.College of Life Science /, Shihezi University,Shihezi 832003,Xinjiang,China
2.Xinjiang Production and Construction Corps Key Laboratory of Oasis Town and Mountain-basin System Ecology /, Shihezi University,Shihezi 832003,Xinjiang,China
3.Key Laboratory of Xinjiang Phytomedicine Resource and Utilization of Ministry of Education, Shihezi University,Shihezi 832003,Xinjiang,China
通讯作者: 李榕(E-mail: lirong@shzu.edu.cn ) 王斐(E-mail: feiw@shzu.edu.cn )
收稿日期: 2023-12-04
修回日期: 2024-01-05
基金资助:
国家自然科学基金项目 . 32060082 新疆生产建设兵团科技计划项目 . 2020BC002 石河子大学科技计划项目 . KX008505 促进与加拿大、澳大利亚、新西兰及拉美地区科研合作与高层次人才培养项目
Received: 2023-12-04
Revised: 2024-01-05
作者简介 About authors
唐蕊(1997—),女,河北张家口人,硕士研究生,研究方向为生物与医药E-mail:2604791944@qq.com
, E-mail:2604791944@qq.com
摘要
葡萄糖-6-磷酸-1-差向异构酶(G6P1E)在植物生长发育和逆境胁迫响应中发挥着重要作用。本研究旨在通过克隆和分析沙漠植物羽毛针禾(Stipagrostis pennata )葡萄糖-6-磷酸-1-差向异构酶基因(SpG6P1E ),为进一步探究其影响羽毛针禾根部沙套发育的机制及功能奠定基础。利用分子克隆技术从羽毛针禾中克隆获得一个SpG6P1E 基因,该基因编码一个含有325个氨基酸的蛋白质,定位于细胞质,为亲水性稳定蛋白。序列及进化分析表明该基因含有一个保守性很高的醛糖异构酶(Aldose_epim)结构域,且与单子叶植物的直系同源基因亲缘性更高。亚细胞定位分析显示该基因定位于细胞质,参与胞质糖代谢过程。qRT-PCR结果显示SpG6P1E 基因的表达与羽毛针禾沙套发育过程及可溶性糖含量具有较高的相关性,表明其对于沙套发育的重要作用;SpG6P1E 基因的表达显著受到干旱、高温、盐等多种非生物胁迫的诱导,表明其对于胁迫响应的重要作用。蛋白互作网络及注释分析显示SpG6P1E 可能通过参与糖合成和糖代谢等一系列过程进而影响羽毛针禾可溶性糖的含量。本研究结果为深入研究SpG6P1E 基因的功能及其调控植物组织发育和适应逆境的机制奠定了基础。
关键词:
羽毛针禾(Stipagrostis pennata ) 葡萄糖-6-磷酸-1-差向异构酶 SpG6P1E 生物信息学分析 亚细胞定位 沙套发育 非生物胁迫
Abstract
Glucose-6-phospho 1 epimerase (G6P1E) plays an important role in plant growth and development and stress response. The purpose of this study is to clone and analyze the expression characteristics of Stipagrosis penata SpG6P1E gene, and lay a foundation for further exploring the mechanism and function of its influence on the development of S. Pennata root sand trap. The full-length open reading frame sequence of S. pennata SpG6P1E gene was cloned by molecular cloning technology. SpG6P1E was cloned from the desert plant S. pennata , which includes a 325 amino acids. SpG6P1E was localized in the cytoplasm. Sequence and evolutionary analysis showed that this gene contains a aldose_epim domain, which is closely related to orthologous gene of monocotyledon plants. Subcellular localization analysis showed that the gene was located in cytoplasm and involved in cytoplasmic glucose metabolism. The results of qRT-PCR showed that SpG6P1E expression had a close link with rhizosheath development and soluble sugar content. The expression of SpG6P1E gene was significantly induced by abiotic stress. Protein interaction and annotation analysis indicated that SpG6P1E might affect the content of soluble sugar by participating in a series of processes such as sugar synthesis and sugar metabolism. The results of this study provide a foundation for in-depth investigation of the functions of G6P1E genes and the mechanisms by which they regulate plant tissue development and adaptation to stress.
Keywords:
Stipagrostis pennata Glucose-6-phospho 1 epimerase SpG6P1E bioinformatics analysis subcellular localization rhizosheath abiotic stress
本文引用格式
唐蕊, 尹珊, 陈凯露, 李悦涛, 程淋渊, 王景儒, 王斐, 李榕, 李鸿彬. 羽毛针禾( Stipagrostis pennata 6 -磷酸-1 -差向异构酶基因 SpG6P1E . 中国沙漠 [J], 2024, 44(4): 126-136 doi:10.7522/j.issn.1000-694X.2024.00011
Tang Rui, Yin Shan, Chen Kailu, Li Yuetao, Cheng Linyuan, Wang Jingru, Wang Fei, Li Rong, Li Hongbin. Cloning and expression analysis of Glucose -6 -phospho 1 epimerase SpG6P1E in Stipagrostis pennata Journal of Desert Research
0 引言
由于全球变暖,气候变化可能导致极端天气的增加和加剧[1 ] 。极端天气和气候会对生态环境和人类社会造成毁灭性影响。干旱是极端气候的直接产物,是影响植物生长和作物产量的重要环境因子。新疆是典型的干旱半干旱区,中国三大沙漠位于新疆,它们构成了新疆独特的自然景观。长期干旱的气候、沙质丰富的自然条件容易形成和发展沙漠[2 ] 。新疆沙漠中活动沙丘分布广泛,约占沙漠面积的85%[3 ] 。土地风蚀导致沙漠化,地表出现风沙流、流沙堆积和沙丘前移,对人类生产、生活等方面造成严重危害。沙漠植物羽毛针禾(Stipagrostis pennata )是古尔班通古特沙漠中的固沙先锋植物[4 ] ,羽毛针禾的根具有特殊的沙套结构,对于其发挥防风固沙、适应沙漠干旱环境等功能具有关键作用。
植物的生长发育和逆境胁迫响应与可溶性糖含量紧密相关。植物通过对体内可溶性糖进行再分配,维持细胞渗透势的平衡,可在逆境胁迫下维持正常生长[5 ] 。植物沙套的形成需要沙粒和根表分泌物的共同作用[6 ] 。沙套代谢组学研究显示植物根部遭受干旱胁迫后,积累了大多数糖类物质和部分氨基酸[7 ] ,尤其是糖类物质含量与干旱条件下的沙套生成密切相关[8 ] 。异构酶是一类催化底物生成同分异构体的酶,根据反应类型和特征可被分为消旋酶、差向异构酶(或表异构酶)、顺反异构酶、互变异构酶和变位酶等[9 ] 。葡萄糖代谢途径中涉及到的异构酶种类丰富,主要包括醛糖差向异构酶(EC 5.1.3.3)、磷酸葡萄糖异构酶(PGI,EC 5.3.1.9)、磷酸葡萄糖变位酶(PGM,EC 5.4.2.2)和葡萄糖-6-磷酸-1-差向异构酶(G6P1E,EC 5.1.3.15)等。G6P1E处在葡萄糖糖代谢途径的分支点,催化葡萄糖-6-磷酸两种差向异构体的平衡。平衡状态下葡萄糖-6-磷酸的组成为37%的α-D-葡萄糖-6-磷酸和63%的 β-D-葡萄糖-6-磷酸[10 ] ,G6P1E与植物中可溶性糖含量显著相关[11 ] 。
目前对于羽毛针禾的研究主要在种群特性、生态适应性及根部微生物等方面,还未有从植物自身发育的角度解析沙套发育机制的报道。我们在种植羽毛针禾的过程中,通过收集材料比对发现,羽毛针禾根部沙套发育30、60、90 d及90 d无沙套根(90d-f)发育具有明显的阶段性。我们前期的研究表明SpG6P1E 基因的表达量在沙套发育过程中持续上升,且在90d-f表达量很低,表明其与沙套发育的紧密联系。本研究对SpG6P1E 所在的全长开放读码框序列进行克隆,并对其组织表达特异性和响应非生物胁迫的表达特征进行分析,为进一步解析SpG6P1E 在羽毛针禾沙套发育的功能及适应极端干旱的机制提供参考。
1 材料与方法
1.1 材料
1.1.1 供试材料
供试材料的种子收集自生长在沙漠中的羽毛针禾(图1 ),采集地点在新疆维吾尔自治区石河子市莫索湾水库附近。种子通过无菌组培发育而成的植株作为供试材料。
图1
图1
羽毛针禾及其根部沙套结构
Fig.1
Plant and rhizosheaths of Stipagrostis pennata
1.1.2 试剂材料
大肠杆菌感受态菌株DH5α购买自北京全式金生物公司,农杆菌菌株为石河子大学生命科学学院农业生物技术重点实验室保存。pMD-19T克隆载体、LA taq酶、Real time PCR试剂、Xba Ⅰ、Xma Ⅰ 等相关酶购自TaKaRa生物公司;RNA提取试剂盒购自Omega;可溶性糖含量测定试剂盒购自索莱宝公司;cDNA第一链反转录试剂盒、RNA反转录试剂盒、2×Taq PCR Master Mix,DNA凝胶回收试剂盒、质粒提取试剂盒、同源重组酶等试剂均购自TIANGEN公司;卡那霉素、青霉素氨苄、庆大霉素、MES、乙酰丁香酮、MgCl2 及培养基配制等化学试剂均为国产分析纯,购自上海生工生物工程公司。PCR所用引物的合成和DNA测序均由新疆有康生物科技有限公司完成。
1.2 方法
1.2.1 羽毛针禾组培及种植
将从莫索湾采集的羽毛针禾种子放置在37 ℃烘箱,烘干后选择饱满的籽粒,去除颖果果皮后,用75%酒精和0.1%次氯酸钠各清洗90 s和10 min。在1/2MS培养基里发芽生根后转至MS培养基。45 d后将幼苗分别用500 mmol·L-1 的甘露醇溶液、250 mmol·L-1 NaCl溶液及50 ℃条件下分别处理0、3、6、12、24 h,收取处理后的样本备用。
1.2.2 羽毛针禾总RNA 的提取及cDNA 的合成
羽毛针禾RNA的提取参照TIANGEN公司的植物总RNA提取试剂盒的说明书进行,经1.0%的琼脂糖凝胶电泳检测其完整性后,用配套的反转录试剂盒合成cDNA。
1.2.3 SpG6P1E
在前期研究基础上,参照转录组测序的转录本拼接结果,设计引物SpG6P1E-F和SpG6P1E-R 及含有Xba I和Xma I酶切位点的同源臂SpG6P1E-tong-F 和SpG6P1E-tong-R (表1 ),以羽毛针禾根组织的cDNA为模板,进行PCR扩增,反应体系如下:cDNA (50 ng·µL-1 ) 1 µL,2×Taq PCR Master Mix 10 µL,引物SpG6P1E-F 0.5 µL,引物SpG6P1E-R 0.5 µL,ddH2O 8 µL,共20 µL。扩增程序为:95 ℃下5 min,95 ℃下30 s,55 ℃下30 s,72 ℃下1 min,32个循环;72 ℃下10 min;4 ℃下保存。使用浓度为1%的琼脂糖凝胶电泳检测并回收目的条带,送至新疆有康生物科技有限公司进行测序。测序和验证正确的核酸序列提交至GenBank数据库(编号:OR797614)。
1.2.4 SpG6P1E
序列比对分析使用序列分析软件DNAMAN(Version9)完成。羽毛针禾SpG6P1E的保守结构域分析使用NCBI网站在线工具ConservedDomains完成。使用ExPSAy在线服务器中的PortParam (https://web.expasy.org/protparam/ ) 在线软件分析计算SpG6P1E编码蛋白质的理化参数[12 ] ;使用PortParam在线软件对该蛋白质进行疏水性分析[12 ] ;利用phyre2 (http://www.sbg.bio.ic.ac.uk/phyre2/ )在线软件分析预测蛋白质二级结构[13 ] ;使用alphafold (https://alphafold.ebi.ac.uk/ ) 在线预测蛋白质三级结构[14 -15 ] ;使用Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/ )在线软件对该蛋白质的亚细胞定位进行分析[16 ] ;使用MEGA(Version11)进行蛋白的进化分析[17 ] ,用MEME在线网站进行motif分析[18 ] 。使用TBtools对进化、保守域和motif进行可视化[19 ] 。
1.2.5 可溶性糖含量的测定
收取羽毛针禾沙套发育各个时期的样本,以10 mg·mL-1 葡萄糖溶液为标准品,参照索莱宝公司植物可溶性糖含量检测试剂盒说明书(BC0030)进行可溶性糖含量测定,每时期进行3次生物学重复。
1.2.6 SpG6P1E
收取羽毛针禾根、节、叶、花、种子,GA3处理0、3、6、12、24 h的根,不同PEG6000浓度处理过的根等组织。提取这些组织的RNA,反转录得到cDNA,以反转录cDNA为模板,分别以羽毛针禾ELF 、GAPDH 、APR6 基因为内参基因[20 ] 。利用Premier5.0软件设计内参基因Actin 以及SpG6P1E 引物(表1 ),参照荧光定量试剂盒说明书 (加拿大,ABM) 进行qRT-PCR (Quantitative real-time polymerase chain reaction) 反应,每个样本进行3个技术重复。反应结束后,根据目的基因和内参基因Ct 值,使用2-ΔΔ Ct
1.2.7 SpG6P1E
把SpG6P1E 基因加上pCAMBIA1300-GFP 的同源臂 (引物SpG6P1E-tong-F 和SpG6P1E-tong-R )连接到pMD19-T 载体上,再转入DH5α大肠杆菌的感受态细胞;将其中的阳性单菌落送新疆有康生物科技有限公司进行检测,将测序正确的单菌落摇菌并提取质粒得到含有SpG6P1E 基因的阳性克隆。将得到的阳性质粒使用Takara公司的Xba I和Xma I进行双酶切验证,用1%的琼脂糖凝胶进行分离,回收纯化后的线性SpG6P1E 基因。再将包含有35S启动子的pCAMBIA1300-GFP 克隆载体用Xba I和Xma I进行双酶切。将得到的SpG6P1E 基因和线性载体进行同源重组。得到重组质粒后转化DH5α大肠杆菌感受态,将培养饱和的大肠杆菌进行质粒提取,对重组质粒进行验证无误后,将35S ::SpG6P1E-GFP 重组质粒进行农杆菌GV3101的转化。转化后,对其阳性单菌落进行PCR验证,将结果正确的菌落进行保存,待用。
1.2.8 SpG6P1E
将生长4周的本氏烟草暗培养过夜,将转化成功的35S ::SpG6P1E-GFP 重组质粒根癌农杆菌在含有硫酸卡那霉素、利福平、庆大霉素的 LB 液体培养基中200 r·min-1 、28 ℃震荡培养8 h;5 000 r·min-1 进行10 min菌体离心,收集菌体沉淀;而后用含有100 mmol·L-1 MES (pH=5.7) 、10 mmol·L-1 MgCl2 、100 μmol·L-1 AS的MS液体重悬液重悬菌体至OD600=0.8。在叶片背面的每个部分打上1个或2个孔,用1 mL无针注射器从烟草叶片背面渗入打孔部位。用塑料罩盖住植物,让渗透的植物在室温和昏暗的光照条件下过夜,之后转移到光照培养室进行正常培养。
1.2.9 SpG6P1E 蛋白质相互作用网络构建及注释分析
将上述得到的羽毛针禾SpG6P1E蛋白序列提交至STRING数据库(https://string-db.org/ ),预测与其存在潜在互作关系的蛋白质及所有相关蛋白的GO和KEGG注释分析。注释结果用R语言进行可视化。
2 结果
2.1 SpG6P1E
以实验室已有的羽毛针禾转录组所组装的转录本序列来设计引物(表1 ),用叶片的cDNA为模版扩增SpG6P1E 基因。经PCR电泳检测和序列测定,成功克隆G6P1E 基因(图2 )。该基因包含975个核苷酸,编码一个由325个氨基酸组成的蛋白质。
图2
图2
SpG6P1E 序列特征分析
Fig.2
Sequence feature analysis of SpG6P1E
对该蛋白质的理化性质进行分析,结果显示其等电点(pI)为5.88,分子量为36.39 kDa;亲水性总平均值 (Grandaverageofhydropathicity, GRAVY)为-0.466,脂溶指数(Aliphaticindex, AI)为76.14。这表明SpG6P1E蛋白具有强亲水性。亚细胞定位预测结果显示其定位于细胞质。序列结构的分析表明,SpG6P1E蛋白的二级结构主要是β折叠和无规卷曲,其中β折叠的组成占比达到了45%;其无序(Disorder)蛋白区域占比较低且集中于N端。蛋白的磷酸化分析结果显示,SpG6P1E 含有15个磷酸化位点,包含5个丝氨酸位点、6个苏氨酸位点和4个酪氨酸位点,其中无序蛋白区域含有3个磷酸化位点,包括1个丝氨酸、2个酪氨酸。该蛋白质不稳定指数 (Theinstabilityindex, II)为33.28,低于40,为稳定蛋白。
2.2 SpG6P1E
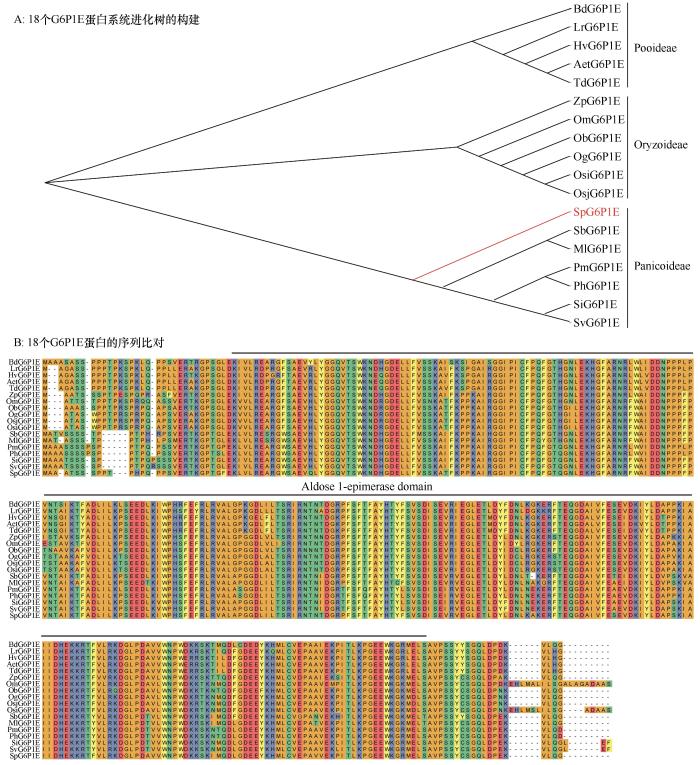
将SpG6P1E蛋白在ncbi上进行blastp比对,取序列相似性最高的30个直系同源基因进行分析。结果显示,这些基因集中在17种单子叶植物,分别归属于早熟禾亚科,稻亚科和黍亚科。其中早熟禾亚科包含硬直黑麦草(Lolium rigidum )、二粒小麦(Triticum dicoccoides )、节节麦(Aegilopst auschii )、大麦(Hordeum vulgare )和二穗短柄草(Brachypo diumdistachyon ),黍亚科包含狗尾草(Setaria viridis )、南荻(Miscanthus lutarioriparius ),谷子(Setaria italica )、高粱(Sorghum bicolor )、黍属野草(Panicum hallii )、糜子(Panicum miliaceum ),稻亚科包含沼生菰(Zizania palustris )、疣粒稻(Oryza meyeriana )、粳稻(Oryzasativa japonica )、非洲稻(Oryza glaberrima )、短花稻(Oryza brachyantha )、籼稻(Oryzasativa indica )。每个物种取一个序列同源性最高的G6P1E 基因,用MEGA和DNAman分别进行进化和序列比对分析,其结果表明该类基因在进化上的亲缘性关系,与其所在物种的植物学分类结果完全一致(图3 );18个G6P1E 基因的序列比对相似性达到了85.44%。这说明,SpG6P1E 及其直系同源基因具有很高的保守性,在单子叶植物中可能具有不可替代的作用。
图3
图3
SpG6P1E序列保守性分析
注: SvG6P1E:XP_034604257.1;MlG6P1E:CAD6259717.1;SiG6P1E:XP_004976707.1;SbG6P1E:XP_021318977.1;ZpG6P1E:KAG8043371.1;OmG6P1E:KAF0892331.1;PhG6P1E:XP_025825614.1;PmG6P1E:RLM64691.1;OgG6P1E:XP_052151522.1;ObG6P1E:XP_006652766.1;OsiG6P1E:EEC77935.1;BdG6P1E:XP_003580451.1;LrG6P1E:XP_047070035.1;TdG6P1E:XP_037479152.1;AetG6P1E:XP_020196260.1;HvG6P1E:XP_044967751.1
Fig.3
Sequence conservative analysis of SpG6P1E
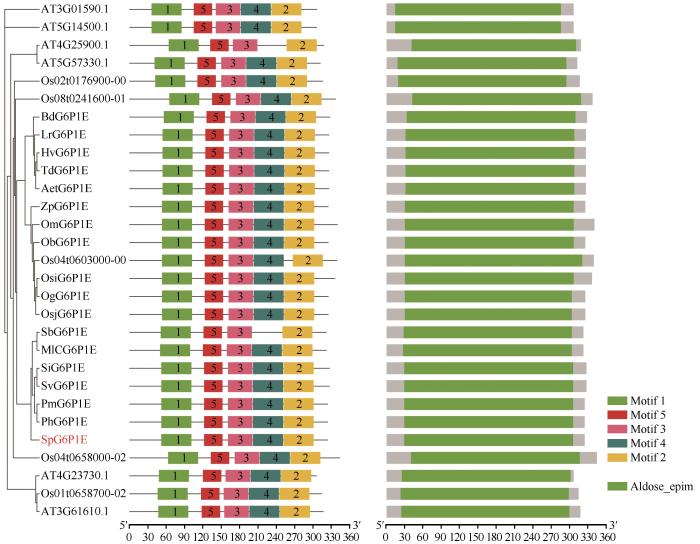
将SpG6P1E 的蛋白序列为输入序列,寻找单子叶模式植物水稻和双子叶模式植物拟南芥中G6P1E 的基因家族。结果显示,水稻中G6P1E 还有5个家族成员,拟南芥中有6个。结构域分析显示,所有的G6P1E 都含有一个共同的保守结构域Aldose_epim (PF01263),除了AT4G25900.1和SbG6P1E 含有4个保守的motif之外,其余所有的基因都含有5个motif(图4 )。该结果进一步验证了G6P1E 的高保守性。进化分析显示SpG6P1E 的单子叶直系同源基因与拟南芥G6P1E 家族成员不在一个聚类组,其亲缘关系相对较远。该结果说明本研究中的SpG6P1E 可能出现在单双子叶植物分化之后,由更原始的G6P1E 基因分化而来。
图4
图4
G6P1E 家族结构域与motif分析
Fig.4
G6P1E family domain and motif analysis
2.3 SpG6P1E
实验室前期将沙套发育的阶段初步界定在3个阶段,即播种后30、60、90 d。同时采集到了90 d未出现沙套的根组织样品(90d-f)。荧光定量PCR的结果表明,SpG6P1E 基因在沙套形成的3个阶段表达量持续升高,且在90d-f样品中表达量很低(图5 )。非生物胁迫的结果表明SpG6P1E 基因在盐胁迫、干旱胁迫和高温胁迫处理后,都表现出了显著的上调趋势。以上结果表明SpG6P1E 在沙套发育的过程及非生物胁迫响应过程中发挥重要作用。
图5
图5
SpG6P1E 的表达分析
Fig.5
The qRT-PCR expression analysis of SpG6P1E
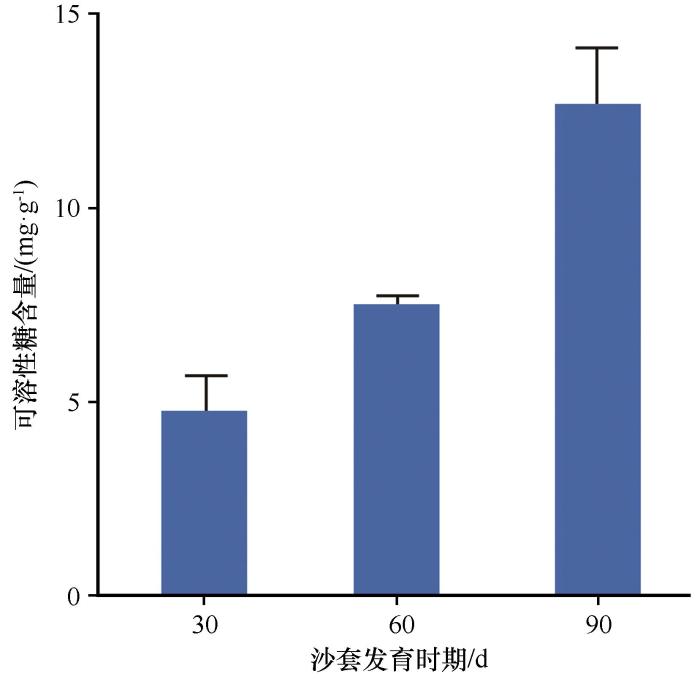
2.4 沙套发育的可溶性糖含量
SpG6P1E 表达分析结果显示,该基因在羽毛针禾沙套发育阶段持续高表达,为了进一步研究该基因对沙套发育的影响,对不同发育阶段沙套的可溶性糖含量进行检测,结果显示,羽毛针禾根部的可溶性糖含量随着沙套的发育逐渐上升(图6 )。结果表明SpG6P1E 基因的表达与沙套发育过程中的可溶性糖含量紧密相关。
图6
图6
沙套发育过程中可溶性糖含量
Fig.6
Contents of soluble sugar in rhizosheath development
2.5 SpG6P1E
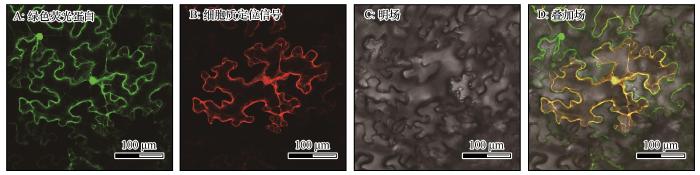
将SpG6P1E 基因与pCAMBIA1300-GFP 载体连接通过同源重组的方法成功构建35S ::SpG6P1E-GFP 载体。将重组质粒转化烟草,利用激光共聚焦显微镜进行亚细胞定位观察,结果显示羽毛针禾SpG6P1E在烟草叶片细胞的细胞质中表达(图7 )。
图7
图7
SpG6P1E 亚细胞定位
Fig.7
Subcellular localization of SpG6P1E
2.6 SpG6P1E 的蛋白互作网络及注释
利用STRING数据库(https://cn.string-db.org/ )对羽毛针禾SpG6P1E进行蛋白互作网络分析。结果显示最可能与SpG6P1E存在相互作用的蛋白质分别为2个葡萄糖磷酸变位酶 (Phosphoglucomutase),3个6-磷酸葡萄糖异构酶(Glucose-6-phosphate isomerase),2个转酮醇酶(Transketolase),1个转醛酮酶(Transaldolase),1个醛脱氢酶 (Aldehyde dehydrogenase)和1个己糖激酶(hexokinase-3,图8 )。这些互作蛋白在GO(Gene Ontology)注释中富集在葡萄糖-6-磷酸代谢过程(GO:0051156)、戊糖磷酸旁路(GO:0006098)、糖异生(GO:0006094)、葡萄糖代谢(GO:0006006)等通路;在KEGG(Kyoto Encyclopedia of Genes and Genomes)通路中显著富集在戊糖磷酸途径(map00030)、糖酵解与糖异生途径(map00010)、半乳糖代谢 (map00052)、光合生物中的碳固定(map00710) 、氨基糖和核苷酸糖(map00520)、淀粉和蔗糖代谢(map00500)等过程。以上结果显示了SpG6P1E蛋白及其互作蛋白广泛参与糖代谢过程,该基因可能通过调整羽毛针禾根部组织中可溶性糖的含量来参与沙套的形成。
图8
图8
蛋白互作网络及注释分析
Fig.8
Protein interaction network and annotation analysis
3 讨论
干旱是影响植物生长和作物产量的重要环境因素,近年来干旱胁迫对环境、作物生产的影响愈发严重[1 ] 。新疆沙漠化问题突出,生态环境十分脆弱,严重影响着人们的生产生活。新疆的沙漠主要为流动沙丘,羽毛针禾主要生长在流动半流动沙地,是沙漠中的固沙先锋植物[21 ] 。羽毛针禾具有抗旱、耐风蚀、耐沙埋等特点,尤其是根部具有特殊的沙套结构,对于其适应沙漠极端干旱环境具有重要作用。研究表明,在干旱胁迫处理后,根部沙套组织显著积累大多数糖类物质和部分氨基酸[7 ] ,尤其是糖类物质,表明其与干旱条件下的沙套生成密切相关。
异构酶广泛存在于植物中,可以通过催化底物生成同分异构体。葡萄糖代谢途径中包含多种异构酶类,其中G6P1E处在葡萄糖代谢途径的分支点,其功能是催化α-D-葡萄糖-6-磷酸和β-D-葡萄糖-6-磷酸间的转化和平衡[22 ] 。该相互转化过程可为丝状真菌的生长和侵染寄主植物提供能量[23 -24 ] 。G6P1E的底物范围较广,能催化葡萄糖-6-磷酸、半乳糖-6-磷酸和甘露糖-6-磷酸等己糖-6-磷酸从α到β的相互转化[22 ] ,表明其可能广泛参与了生物体内的糖代谢过程。
G6P1E最初是在酵母中被分离鉴定的,此后在拟南芥、水稻、烟草、狗尾草、石竹等植物中都发现了该蛋白[11 ] ,但关于羽毛针禾中SpG6P1E 基因的研究尚未被报道。本研究克隆获得一个羽毛针禾G6P1E 基因,该基因编码一个含有325个氨基酸的蛋白质,定位于细胞质,为亲水性稳定蛋白(图2 、7 ) 。SpG6P1E 基因具有保守的基因序列和结构域(图3 、4 ),其序列相似度较高的直系同源基因主要分布在单子叶植物中的黍亚科、早熟禾亚科和稻亚科中,尤其与黍亚科中狗尾草的G6P1E 同源性最高。本研究中,qRT-PCR分析表明该基因的表达与沙套发育过程呈现出紧密的正相关关系(图5 ),暗示其与根部沙套发育之间的密切联系。在干旱、高温和盐胁迫等非生物胁迫下,SpG6P1E 基因的表达受到显著的诱导(图5 ),表明SpG6P1E 基因与沙漠极端干旱环境适应之间具有紧密联系。
SpG6P1E 基因与植物体内可溶性糖的含量显著相关。可溶性糖含量与花色在石竹属开花植物中显著相关,通过VIGS沉默掉DchG6P1E 基因后,酶活性与对照相比显著下降,葡萄糖含量显著降低,且花色出现明显变化,较对照整体变浅。这表明Dch6GP1E 通过参与糖的代谢过程,影响了植物体内可溶性糖的含量,进而影响中国石竹的花色[11 ] 。烟草G6P1E 同源基因NbAELP 被敲除后,叶片中的葡萄糖含量显著降低,植物生长受到影响,抵御非生物胁迫和生物胁迫的能力降低,该研究表明G6P1E 通过影响烟草中的葡萄糖含量,进而影响其生长和抵御胁迫的能力[25 ] 。本研究结果也表明羽毛针禾沙套发育过程中,SpG6P1E 基因的表达量与可溶性糖含量呈现显著正相关关系(图5 、6 )。此外,基因的亚细胞定位与功能发挥之间具有紧密的联系[26 ] 。SpG6P1E 亚细胞定位于细胞质,可能参与了细胞质中糖的相关代谢过程。蛋白互作网络预测也显示SpG6P1E 和其互作蛋白参与了糖的合成与代谢等一系列过程(图7 、8 )。
以上结果初步揭示了SpG6P1E 基因与羽毛针禾沙套发育、抵御逆境胁迫的紧密联系,其可能通过影响羽毛针禾可溶性糖的含量在沙套形成过程中发挥了重要作用,进而帮助羽毛针禾适应沙漠的极端干旱环境。因此,对于SpG6P1E 基因参与羽毛针禾响应非生物胁迫的具体作用机制,值得进一步研究。
4 结论
本研究首次克隆获得羽毛针禾的一个葡萄糖-6-磷酸-1-差向异构酶SpG6P1E ,该基因定位在细胞质,与羽毛针禾沙套发育显著相关并显著受到干旱、高温、盐等非生物胁迫的诱导,可能与羽毛针禾可溶性糖含量紧密相关。本研究为进一步解析羽毛针禾G6P1E 基因影响沙套发育的功能和调控机制解析提供了理论基础。
参考文献
View Option
[1]
张晓芹 西北旱区典型生态经济树种地理分布与气候适宜性研究
[D].北京 :中国科学院大学 ,2018 .
[本文引用: 2]
[2]
Bengough A G Water dynamics of the root zone:rhizosphere biophysics and its control on soil hydrology
[J].Vadose Zone ,2012 ,11 (2 ):1 -12 .
[本文引用: 1]
[3]
Pang J Ryan M H Kadambot H M et al Unwrapping the rhizosheath
[J].Plant and Soil ,2017 ,418 :129 -139 .
[本文引用: 1]
[4]
龙丽红 ,王慧 ,马晓丽 ,等 古尔班通古特沙漠羽毛针禾 (Stipagrostis pennata ) 种群种子雨特征
[J].干旱区研究 ,2014 ,31 (3 ):516 -522 .
[本文引用: 1]
[5]
Yamada K Osakabe Y Mizoi J et al Functional analysis of an Arabidopsis thaliana abiotic stress-inducible facilitated diffusion transporter for monosaccharides
[J].Biochemistry & Molecular Biology ,2010 ,285 (2 ):1138 -1146 .
[本文引用: 1]
[6]
Mahmood T Mehnaz S Fleischmann F et al Soil sterilization effects on root growth and formation of rhizosheaths in wheat seedlings
[J].Pedobiologia ,2014 ,57 (3 ):123 -130 .
[本文引用: 1]
[7]
Nambiar E Uptake of Zn65from dry soil by plants
[J].Plant & Soil ,1976 ,44 (1 ):267 -271 .
[本文引用: 2]
[8]
Liu T Y Chen M X Zhang Y et al Comparative metabolite profiling of two switchgrass ecotypes reveals differences in drought stress responses and rhizosheath weight
[J].Planta ,2019 ,250 (4 ):1355 -1369 .
[本文引用: 1]
[9]
陈尚武 课堂教学随笔:异构酶、变位酶、消旋酶、差向异构酶与互变异构酶
[J].生命的化学 ,2020 ,40 (3 ):433 -435 .
[本文引用: 1]
[10]
Wurster B Hess B Glucose-6-phosphate-1-epimerase from baker's yeast.a new enzyme
[J].FEBS Letters ,1972 ,23 (3 ):341 -344 .
[本文引用: 1]
[11]
高亚南 中国石竹中葡萄糖-6-磷酸-1-差向异构酶基因的功能鉴定及表达分析
[D].呼和浩特 :内蒙古农业大学 ,2021 .
[本文引用: 3]
[12]
Ison J Kalas M Jonassen I et al EDAM:an ontology of bioinformatics operations, types of data and identifiers, topics and formats
[J].Bioinformatics ,2013 ,29 (10 ):1325 -1332 .
[本文引用: 2]
[13]
Kelley L A Mezulis S Yates C M et al The Phyre2 web portal for protein modeling, prediction and analysis
[J].Nature Protocols ,2015 ,10 (6 ):845 -858 .
[本文引用: 1]
[14]
Jumper J Evans R Pritzel A et al Highly accurate protein structure prediction with AlphaFold
[J].Nature ,2021 ,596 (7873 ):583 -589 .
[本文引用: 1]
[15]
Varadi M Anyango S Deshpande M et al AlphaFold protein structure database:massively expanding the structural coverage of protein-sequence space with high-accuracy models
[J].Nucleic Acids Research ,2022 ,50 (D1 ):D439 -D444 .
[本文引用: 1]
[16]
Chou K C Shen H B Cell-PLoc 2.0:an improved package of web-servers for predicting subcellular localization of proteins in various organisms
[J].Natural Science ,2010 ,2 (10 ):1090 .
[本文引用: 1]
[17]
Tamura K Stecher G Kumar S MEGA11:molecular evolutionary genetics analysis version 11
[J].Molecular Biology and Evolution ,2021 ,38 (7 ):3022 -3027 .
[本文引用: 1]
[18]
Bailey T L Johnson J Grant C E et al The MEME suite
[J].Nucleic Acids Research ,2015 ,43 (W1 ):W39 -W49 .
[本文引用: 1]
[19]
Chen C Wu Y Li J et al TBtools-II:A" One for All, All for One" bioinformatics platform for biological big-data mining
[J].Molecular Plant ,2023 ,16 (11 ):1733 -1742 .
[本文引用: 1]
[20]
Li R Cui K Xie Q et al Selection of the reference genes for quantitative gene expression by RT-qPCR in the desert plant Stipagrostis pennata
[J].Scientific Reports ,2021 (1 ):21711 .
[本文引用: 1]
[21]
张丽霞 ,张霞 ,王绍明 ,等 公路阻碍作用对古尔班通古特沙漠南缘羽毛针禾种群基因流的影响
[J].江苏农业科学 ,2019 ,47 (9 ):104 -109 .
[本文引用: 1]
[22]
Graille M Baltaze J P Leulliot N et al Structure-based functional annotation:yeast ymr099c codes for a D-hexose-6-phosphate mutarotase
[J].Journal of Biological Chemistry ,2006 ,281 (40 ):30175 -30185 .
[本文引用: 2]
[23]
Lv B Guo Y Zhao X et al Glucose-6-phosphate 1-Epimerase CrGlu6 contributes to development and biocontrol efficiency in clonostachys chloroleuca
[J].Journal of Fungi ,2023 ,9 (7 ):764 .
[本文引用: 1]
[24]
Samolski I de Luis A Vizcaíno J A et al Gene expression analysis of the biocontrol fungus Trichoderma harzianum in the presence of tomato plants,chitin,or glucose using a high-density oligonucleotide microarray
[J].BMC Microbiology ,2009 ,9 (1 ):1 -14 .
[本文引用: 1]
[25]
Sheshukova E V Komarova T V Pozdyshev D V et al The intergenic interplay between aldose 1-epimerase-like protein and pectin methylesterase in abiotic and biotic stress control
[J].Frontiers in Plant Science ,2017 ,8 :1646 .
[本文引用: 1]
[26]
申龙斌 ,李瑞梅 ,段瑞军 ,等 海马齿 Spmet 基因表达产物的亚细胞定位及其组织表达特异性分析
[J].中国农学通报 ,2012 ,28 (27 ):213 -217 .
[本文引用: 1]
西北旱区典型生态经济树种地理分布与气候适宜性研究
2
2018
... 由于全球变暖,气候变化可能导致极端天气的增加和加剧[1 ] .极端天气和气候会对生态环境和人类社会造成毁灭性影响.干旱是极端气候的直接产物,是影响植物生长和作物产量的重要环境因子.新疆是典型的干旱半干旱区,中国三大沙漠位于新疆,它们构成了新疆独特的自然景观.长期干旱的气候、沙质丰富的自然条件容易形成和发展沙漠[2 ] .新疆沙漠中活动沙丘分布广泛,约占沙漠面积的85%[3 ] .土地风蚀导致沙漠化,地表出现风沙流、流沙堆积和沙丘前移,对人类生产、生活等方面造成严重危害.沙漠植物羽毛针禾(Stipagrostis pennata )是古尔班通古特沙漠中的固沙先锋植物[4 ] ,羽毛针禾的根具有特殊的沙套结构,对于其发挥防风固沙、适应沙漠干旱环境等功能具有关键作用. ...
... 干旱是影响植物生长和作物产量的重要环境因素,近年来干旱胁迫对环境、作物生产的影响愈发严重[1 ] .新疆沙漠化问题突出,生态环境十分脆弱,严重影响着人们的生产生活.新疆的沙漠主要为流动沙丘,羽毛针禾主要生长在流动半流动沙地,是沙漠中的固沙先锋植物[21 ] .羽毛针禾具有抗旱、耐风蚀、耐沙埋等特点,尤其是根部具有特殊的沙套结构,对于其适应沙漠极端干旱环境具有重要作用.研究表明,在干旱胁迫处理后,根部沙套组织显著积累大多数糖类物质和部分氨基酸[7 ] ,尤其是糖类物质,表明其与干旱条件下的沙套生成密切相关. ...
Water dynamics of the root zone:rhizosphere biophysics and its control on soil hydrology
1
2012
... 由于全球变暖,气候变化可能导致极端天气的增加和加剧[1 ] .极端天气和气候会对生态环境和人类社会造成毁灭性影响.干旱是极端气候的直接产物,是影响植物生长和作物产量的重要环境因子.新疆是典型的干旱半干旱区,中国三大沙漠位于新疆,它们构成了新疆独特的自然景观.长期干旱的气候、沙质丰富的自然条件容易形成和发展沙漠[2 ] .新疆沙漠中活动沙丘分布广泛,约占沙漠面积的85%[3 ] .土地风蚀导致沙漠化,地表出现风沙流、流沙堆积和沙丘前移,对人类生产、生活等方面造成严重危害.沙漠植物羽毛针禾(Stipagrostis pennata )是古尔班通古特沙漠中的固沙先锋植物[4 ] ,羽毛针禾的根具有特殊的沙套结构,对于其发挥防风固沙、适应沙漠干旱环境等功能具有关键作用. ...
Unwrapping the rhizosheath
1
2017
... 由于全球变暖,气候变化可能导致极端天气的增加和加剧[1 ] .极端天气和气候会对生态环境和人类社会造成毁灭性影响.干旱是极端气候的直接产物,是影响植物生长和作物产量的重要环境因子.新疆是典型的干旱半干旱区,中国三大沙漠位于新疆,它们构成了新疆独特的自然景观.长期干旱的气候、沙质丰富的自然条件容易形成和发展沙漠[2 ] .新疆沙漠中活动沙丘分布广泛,约占沙漠面积的85%[3 ] .土地风蚀导致沙漠化,地表出现风沙流、流沙堆积和沙丘前移,对人类生产、生活等方面造成严重危害.沙漠植物羽毛针禾(Stipagrostis pennata )是古尔班通古特沙漠中的固沙先锋植物[4 ] ,羽毛针禾的根具有特殊的沙套结构,对于其发挥防风固沙、适应沙漠干旱环境等功能具有关键作用. ...
古尔班通古特沙漠羽毛针禾 (Stipagrostis pennata ) 种群种子雨特征
1
2014
... 由于全球变暖,气候变化可能导致极端天气的增加和加剧[1 ] .极端天气和气候会对生态环境和人类社会造成毁灭性影响.干旱是极端气候的直接产物,是影响植物生长和作物产量的重要环境因子.新疆是典型的干旱半干旱区,中国三大沙漠位于新疆,它们构成了新疆独特的自然景观.长期干旱的气候、沙质丰富的自然条件容易形成和发展沙漠[2 ] .新疆沙漠中活动沙丘分布广泛,约占沙漠面积的85%[3 ] .土地风蚀导致沙漠化,地表出现风沙流、流沙堆积和沙丘前移,对人类生产、生活等方面造成严重危害.沙漠植物羽毛针禾(Stipagrostis pennata )是古尔班通古特沙漠中的固沙先锋植物[4 ] ,羽毛针禾的根具有特殊的沙套结构,对于其发挥防风固沙、适应沙漠干旱环境等功能具有关键作用. ...
Functional analysis of an Arabidopsis thaliana abiotic stress-inducible facilitated diffusion transporter for monosaccharides
1
2010
... 植物的生长发育和逆境胁迫响应与可溶性糖含量紧密相关.植物通过对体内可溶性糖进行再分配,维持细胞渗透势的平衡,可在逆境胁迫下维持正常生长[5 ] .植物沙套的形成需要沙粒和根表分泌物的共同作用[6 ] .沙套代谢组学研究显示植物根部遭受干旱胁迫后,积累了大多数糖类物质和部分氨基酸[7 ] ,尤其是糖类物质含量与干旱条件下的沙套生成密切相关[8 ] .异构酶是一类催化底物生成同分异构体的酶,根据反应类型和特征可被分为消旋酶、差向异构酶(或表异构酶)、顺反异构酶、互变异构酶和变位酶等[9 ] .葡萄糖代谢途径中涉及到的异构酶种类丰富,主要包括醛糖差向异构酶(EC 5.1.3.3)、磷酸葡萄糖异构酶(PGI,EC 5.3.1.9)、磷酸葡萄糖变位酶(PGM,EC 5.4.2.2)和葡萄糖-6-磷酸-1-差向异构酶(G6P1E,EC 5.1.3.15)等.G6P1E处在葡萄糖糖代谢途径的分支点,催化葡萄糖-6-磷酸两种差向异构体的平衡.平衡状态下葡萄糖-6-磷酸的组成为37%的α-D-葡萄糖-6-磷酸和63%的 β-D-葡萄糖-6-磷酸[10 ] ,G6P1E与植物中可溶性糖含量显著相关[11 ] . ...
Soil sterilization effects on root growth and formation of rhizosheaths in wheat seedlings
1
2014
... 植物的生长发育和逆境胁迫响应与可溶性糖含量紧密相关.植物通过对体内可溶性糖进行再分配,维持细胞渗透势的平衡,可在逆境胁迫下维持正常生长[5 ] .植物沙套的形成需要沙粒和根表分泌物的共同作用[6 ] .沙套代谢组学研究显示植物根部遭受干旱胁迫后,积累了大多数糖类物质和部分氨基酸[7 ] ,尤其是糖类物质含量与干旱条件下的沙套生成密切相关[8 ] .异构酶是一类催化底物生成同分异构体的酶,根据反应类型和特征可被分为消旋酶、差向异构酶(或表异构酶)、顺反异构酶、互变异构酶和变位酶等[9 ] .葡萄糖代谢途径中涉及到的异构酶种类丰富,主要包括醛糖差向异构酶(EC 5.1.3.3)、磷酸葡萄糖异构酶(PGI,EC 5.3.1.9)、磷酸葡萄糖变位酶(PGM,EC 5.4.2.2)和葡萄糖-6-磷酸-1-差向异构酶(G6P1E,EC 5.1.3.15)等.G6P1E处在葡萄糖糖代谢途径的分支点,催化葡萄糖-6-磷酸两种差向异构体的平衡.平衡状态下葡萄糖-6-磷酸的组成为37%的α-D-葡萄糖-6-磷酸和63%的 β-D-葡萄糖-6-磷酸[10 ] ,G6P1E与植物中可溶性糖含量显著相关[11 ] . ...
Uptake of Zn65from dry soil by plants
2
1976
... 植物的生长发育和逆境胁迫响应与可溶性糖含量紧密相关.植物通过对体内可溶性糖进行再分配,维持细胞渗透势的平衡,可在逆境胁迫下维持正常生长[5 ] .植物沙套的形成需要沙粒和根表分泌物的共同作用[6 ] .沙套代谢组学研究显示植物根部遭受干旱胁迫后,积累了大多数糖类物质和部分氨基酸[7 ] ,尤其是糖类物质含量与干旱条件下的沙套生成密切相关[8 ] .异构酶是一类催化底物生成同分异构体的酶,根据反应类型和特征可被分为消旋酶、差向异构酶(或表异构酶)、顺反异构酶、互变异构酶和变位酶等[9 ] .葡萄糖代谢途径中涉及到的异构酶种类丰富,主要包括醛糖差向异构酶(EC 5.1.3.3)、磷酸葡萄糖异构酶(PGI,EC 5.3.1.9)、磷酸葡萄糖变位酶(PGM,EC 5.4.2.2)和葡萄糖-6-磷酸-1-差向异构酶(G6P1E,EC 5.1.3.15)等.G6P1E处在葡萄糖糖代谢途径的分支点,催化葡萄糖-6-磷酸两种差向异构体的平衡.平衡状态下葡萄糖-6-磷酸的组成为37%的α-D-葡萄糖-6-磷酸和63%的 β-D-葡萄糖-6-磷酸[10 ] ,G6P1E与植物中可溶性糖含量显著相关[11 ] . ...
... 干旱是影响植物生长和作物产量的重要环境因素,近年来干旱胁迫对环境、作物生产的影响愈发严重[1 ] .新疆沙漠化问题突出,生态环境十分脆弱,严重影响着人们的生产生活.新疆的沙漠主要为流动沙丘,羽毛针禾主要生长在流动半流动沙地,是沙漠中的固沙先锋植物[21 ] .羽毛针禾具有抗旱、耐风蚀、耐沙埋等特点,尤其是根部具有特殊的沙套结构,对于其适应沙漠极端干旱环境具有重要作用.研究表明,在干旱胁迫处理后,根部沙套组织显著积累大多数糖类物质和部分氨基酸[7 ] ,尤其是糖类物质,表明其与干旱条件下的沙套生成密切相关. ...
Comparative metabolite profiling of two switchgrass ecotypes reveals differences in drought stress responses and rhizosheath weight
1
2019
... 植物的生长发育和逆境胁迫响应与可溶性糖含量紧密相关.植物通过对体内可溶性糖进行再分配,维持细胞渗透势的平衡,可在逆境胁迫下维持正常生长[5 ] .植物沙套的形成需要沙粒和根表分泌物的共同作用[6 ] .沙套代谢组学研究显示植物根部遭受干旱胁迫后,积累了大多数糖类物质和部分氨基酸[7 ] ,尤其是糖类物质含量与干旱条件下的沙套生成密切相关[8 ] .异构酶是一类催化底物生成同分异构体的酶,根据反应类型和特征可被分为消旋酶、差向异构酶(或表异构酶)、顺反异构酶、互变异构酶和变位酶等[9 ] .葡萄糖代谢途径中涉及到的异构酶种类丰富,主要包括醛糖差向异构酶(EC 5.1.3.3)、磷酸葡萄糖异构酶(PGI,EC 5.3.1.9)、磷酸葡萄糖变位酶(PGM,EC 5.4.2.2)和葡萄糖-6-磷酸-1-差向异构酶(G6P1E,EC 5.1.3.15)等.G6P1E处在葡萄糖糖代谢途径的分支点,催化葡萄糖-6-磷酸两种差向异构体的平衡.平衡状态下葡萄糖-6-磷酸的组成为37%的α-D-葡萄糖-6-磷酸和63%的 β-D-葡萄糖-6-磷酸[10 ] ,G6P1E与植物中可溶性糖含量显著相关[11 ] . ...
课堂教学随笔:异构酶、变位酶、消旋酶、差向异构酶与互变异构酶
1
2020
... 植物的生长发育和逆境胁迫响应与可溶性糖含量紧密相关.植物通过对体内可溶性糖进行再分配,维持细胞渗透势的平衡,可在逆境胁迫下维持正常生长[5 ] .植物沙套的形成需要沙粒和根表分泌物的共同作用[6 ] .沙套代谢组学研究显示植物根部遭受干旱胁迫后,积累了大多数糖类物质和部分氨基酸[7 ] ,尤其是糖类物质含量与干旱条件下的沙套生成密切相关[8 ] .异构酶是一类催化底物生成同分异构体的酶,根据反应类型和特征可被分为消旋酶、差向异构酶(或表异构酶)、顺反异构酶、互变异构酶和变位酶等[9 ] .葡萄糖代谢途径中涉及到的异构酶种类丰富,主要包括醛糖差向异构酶(EC 5.1.3.3)、磷酸葡萄糖异构酶(PGI,EC 5.3.1.9)、磷酸葡萄糖变位酶(PGM,EC 5.4.2.2)和葡萄糖-6-磷酸-1-差向异构酶(G6P1E,EC 5.1.3.15)等.G6P1E处在葡萄糖糖代谢途径的分支点,催化葡萄糖-6-磷酸两种差向异构体的平衡.平衡状态下葡萄糖-6-磷酸的组成为37%的α-D-葡萄糖-6-磷酸和63%的 β-D-葡萄糖-6-磷酸[10 ] ,G6P1E与植物中可溶性糖含量显著相关[11 ] . ...
Glucose-6-phosphate-1-epimerase from baker's yeast.a new enzyme
1
1972
... 植物的生长发育和逆境胁迫响应与可溶性糖含量紧密相关.植物通过对体内可溶性糖进行再分配,维持细胞渗透势的平衡,可在逆境胁迫下维持正常生长[5 ] .植物沙套的形成需要沙粒和根表分泌物的共同作用[6 ] .沙套代谢组学研究显示植物根部遭受干旱胁迫后,积累了大多数糖类物质和部分氨基酸[7 ] ,尤其是糖类物质含量与干旱条件下的沙套生成密切相关[8 ] .异构酶是一类催化底物生成同分异构体的酶,根据反应类型和特征可被分为消旋酶、差向异构酶(或表异构酶)、顺反异构酶、互变异构酶和变位酶等[9 ] .葡萄糖代谢途径中涉及到的异构酶种类丰富,主要包括醛糖差向异构酶(EC 5.1.3.3)、磷酸葡萄糖异构酶(PGI,EC 5.3.1.9)、磷酸葡萄糖变位酶(PGM,EC 5.4.2.2)和葡萄糖-6-磷酸-1-差向异构酶(G6P1E,EC 5.1.3.15)等.G6P1E处在葡萄糖糖代谢途径的分支点,催化葡萄糖-6-磷酸两种差向异构体的平衡.平衡状态下葡萄糖-6-磷酸的组成为37%的α-D-葡萄糖-6-磷酸和63%的 β-D-葡萄糖-6-磷酸[10 ] ,G6P1E与植物中可溶性糖含量显著相关[11 ] . ...
中国石竹中葡萄糖-6-磷酸-1-差向异构酶基因的功能鉴定及表达分析
3
2021
... 植物的生长发育和逆境胁迫响应与可溶性糖含量紧密相关.植物通过对体内可溶性糖进行再分配,维持细胞渗透势的平衡,可在逆境胁迫下维持正常生长[5 ] .植物沙套的形成需要沙粒和根表分泌物的共同作用[6 ] .沙套代谢组学研究显示植物根部遭受干旱胁迫后,积累了大多数糖类物质和部分氨基酸[7 ] ,尤其是糖类物质含量与干旱条件下的沙套生成密切相关[8 ] .异构酶是一类催化底物生成同分异构体的酶,根据反应类型和特征可被分为消旋酶、差向异构酶(或表异构酶)、顺反异构酶、互变异构酶和变位酶等[9 ] .葡萄糖代谢途径中涉及到的异构酶种类丰富,主要包括醛糖差向异构酶(EC 5.1.3.3)、磷酸葡萄糖异构酶(PGI,EC 5.3.1.9)、磷酸葡萄糖变位酶(PGM,EC 5.4.2.2)和葡萄糖-6-磷酸-1-差向异构酶(G6P1E,EC 5.1.3.15)等.G6P1E处在葡萄糖糖代谢途径的分支点,催化葡萄糖-6-磷酸两种差向异构体的平衡.平衡状态下葡萄糖-6-磷酸的组成为37%的α-D-葡萄糖-6-磷酸和63%的 β-D-葡萄糖-6-磷酸[10 ] ,G6P1E与植物中可溶性糖含量显著相关[11 ] . ...
... G6P1E最初是在酵母中被分离鉴定的,此后在拟南芥、水稻、烟草、狗尾草、石竹等植物中都发现了该蛋白[11 ] ,但关于羽毛针禾中SpG6P1E 基因的研究尚未被报道.本研究克隆获得一个羽毛针禾G6P1E 基因,该基因编码一个含有325个氨基酸的蛋白质,定位于细胞质,为亲水性稳定蛋白(图2 、7 ) .SpG6P1E 基因具有保守的基因序列和结构域(图3 、4 ),其序列相似度较高的直系同源基因主要分布在单子叶植物中的黍亚科、早熟禾亚科和稻亚科中,尤其与黍亚科中狗尾草的G6P1E 同源性最高.本研究中,qRT-PCR分析表明该基因的表达与沙套发育过程呈现出紧密的正相关关系(图5 ),暗示其与根部沙套发育之间的密切联系.在干旱、高温和盐胁迫等非生物胁迫下,SpG6P1E 基因的表达受到显著的诱导(图5 ),表明SpG6P1E 基因与沙漠极端干旱环境适应之间具有紧密联系. ...
... SpG6P1E 基因与植物体内可溶性糖的含量显著相关.可溶性糖含量与花色在石竹属开花植物中显著相关,通过VIGS沉默掉DchG6P1E 基因后,酶活性与对照相比显著下降,葡萄糖含量显著降低,且花色出现明显变化,较对照整体变浅.这表明Dch6GP1E 通过参与糖的代谢过程,影响了植物体内可溶性糖的含量,进而影响中国石竹的花色[11 ] .烟草G6P1E 同源基因NbAELP 被敲除后,叶片中的葡萄糖含量显著降低,植物生长受到影响,抵御非生物胁迫和生物胁迫的能力降低,该研究表明G6P1E 通过影响烟草中的葡萄糖含量,进而影响其生长和抵御胁迫的能力[25 ] .本研究结果也表明羽毛针禾沙套发育过程中,SpG6P1E 基因的表达量与可溶性糖含量呈现显著正相关关系(图5 、6 ).此外,基因的亚细胞定位与功能发挥之间具有紧密的联系[26 ] .SpG6P1E 亚细胞定位于细胞质,可能参与了细胞质中糖的相关代谢过程.蛋白互作网络预测也显示SpG6P1E 和其互作蛋白参与了糖的合成与代谢等一系列过程(图7 、8 ). ...
EDAM:an ontology of bioinformatics operations, types of data and identifiers, topics and formats
2
2013
... 序列比对分析使用序列分析软件DNAMAN(Version9)完成.羽毛针禾SpG6P1E的保守结构域分析使用NCBI网站在线工具ConservedDomains完成.使用ExPSAy在线服务器中的PortParam (https://web.expasy.org/protparam/ ) 在线软件分析计算SpG6P1E编码蛋白质的理化参数[12 ] ;使用PortParam在线软件对该蛋白质进行疏水性分析[12 ] ;利用phyre2 (http://www.sbg.bio.ic.ac.uk/phyre2/ )在线软件分析预测蛋白质二级结构[13 ] ;使用alphafold (https://alphafold.ebi.ac.uk/ ) 在线预测蛋白质三级结构[14 -15 ] ;使用Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/ )在线软件对该蛋白质的亚细胞定位进行分析[16 ] ;使用MEGA(Version11)进行蛋白的进化分析[17 ] ,用MEME在线网站进行motif分析[18 ] .使用TBtools对进化、保守域和motif进行可视化[19 ] . ...
... [12 ];利用phyre2 (http://www.sbg.bio.ic.ac.uk/phyre2/ )在线软件分析预测蛋白质二级结构[13 ] ;使用alphafold (https://alphafold.ebi.ac.uk/ ) 在线预测蛋白质三级结构[14 -15 ] ;使用Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/ )在线软件对该蛋白质的亚细胞定位进行分析[16 ] ;使用MEGA(Version11)进行蛋白的进化分析[17 ] ,用MEME在线网站进行motif分析[18 ] .使用TBtools对进化、保守域和motif进行可视化[19 ] . ...
The Phyre2 web portal for protein modeling, prediction and analysis
1
2015
... 序列比对分析使用序列分析软件DNAMAN(Version9)完成.羽毛针禾SpG6P1E的保守结构域分析使用NCBI网站在线工具ConservedDomains完成.使用ExPSAy在线服务器中的PortParam (https://web.expasy.org/protparam/ ) 在线软件分析计算SpG6P1E编码蛋白质的理化参数[12 ] ;使用PortParam在线软件对该蛋白质进行疏水性分析[12 ] ;利用phyre2 (http://www.sbg.bio.ic.ac.uk/phyre2/ )在线软件分析预测蛋白质二级结构[13 ] ;使用alphafold (https://alphafold.ebi.ac.uk/ ) 在线预测蛋白质三级结构[14 -15 ] ;使用Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/ )在线软件对该蛋白质的亚细胞定位进行分析[16 ] ;使用MEGA(Version11)进行蛋白的进化分析[17 ] ,用MEME在线网站进行motif分析[18 ] .使用TBtools对进化、保守域和motif进行可视化[19 ] . ...
Highly accurate protein structure prediction with AlphaFold
1
2021
... 序列比对分析使用序列分析软件DNAMAN(Version9)完成.羽毛针禾SpG6P1E的保守结构域分析使用NCBI网站在线工具ConservedDomains完成.使用ExPSAy在线服务器中的PortParam (https://web.expasy.org/protparam/ ) 在线软件分析计算SpG6P1E编码蛋白质的理化参数[12 ] ;使用PortParam在线软件对该蛋白质进行疏水性分析[12 ] ;利用phyre2 (http://www.sbg.bio.ic.ac.uk/phyre2/ )在线软件分析预测蛋白质二级结构[13 ] ;使用alphafold (https://alphafold.ebi.ac.uk/ ) 在线预测蛋白质三级结构[14 -15 ] ;使用Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/ )在线软件对该蛋白质的亚细胞定位进行分析[16 ] ;使用MEGA(Version11)进行蛋白的进化分析[17 ] ,用MEME在线网站进行motif分析[18 ] .使用TBtools对进化、保守域和motif进行可视化[19 ] . ...
AlphaFold protein structure database:massively expanding the structural coverage of protein-sequence space with high-accuracy models
1
2022
... 序列比对分析使用序列分析软件DNAMAN(Version9)完成.羽毛针禾SpG6P1E的保守结构域分析使用NCBI网站在线工具ConservedDomains完成.使用ExPSAy在线服务器中的PortParam (https://web.expasy.org/protparam/ ) 在线软件分析计算SpG6P1E编码蛋白质的理化参数[12 ] ;使用PortParam在线软件对该蛋白质进行疏水性分析[12 ] ;利用phyre2 (http://www.sbg.bio.ic.ac.uk/phyre2/ )在线软件分析预测蛋白质二级结构[13 ] ;使用alphafold (https://alphafold.ebi.ac.uk/ ) 在线预测蛋白质三级结构[14 -15 ] ;使用Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/ )在线软件对该蛋白质的亚细胞定位进行分析[16 ] ;使用MEGA(Version11)进行蛋白的进化分析[17 ] ,用MEME在线网站进行motif分析[18 ] .使用TBtools对进化、保守域和motif进行可视化[19 ] . ...
Cell-PLoc 2.0:an improved package of web-servers for predicting subcellular localization of proteins in various organisms
1
2010
... 序列比对分析使用序列分析软件DNAMAN(Version9)完成.羽毛针禾SpG6P1E的保守结构域分析使用NCBI网站在线工具ConservedDomains完成.使用ExPSAy在线服务器中的PortParam (https://web.expasy.org/protparam/ ) 在线软件分析计算SpG6P1E编码蛋白质的理化参数[12 ] ;使用PortParam在线软件对该蛋白质进行疏水性分析[12 ] ;利用phyre2 (http://www.sbg.bio.ic.ac.uk/phyre2/ )在线软件分析预测蛋白质二级结构[13 ] ;使用alphafold (https://alphafold.ebi.ac.uk/ ) 在线预测蛋白质三级结构[14 -15 ] ;使用Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/ )在线软件对该蛋白质的亚细胞定位进行分析[16 ] ;使用MEGA(Version11)进行蛋白的进化分析[17 ] ,用MEME在线网站进行motif分析[18 ] .使用TBtools对进化、保守域和motif进行可视化[19 ] . ...
MEGA11:molecular evolutionary genetics analysis version 11
1
2021
... 序列比对分析使用序列分析软件DNAMAN(Version9)完成.羽毛针禾SpG6P1E的保守结构域分析使用NCBI网站在线工具ConservedDomains完成.使用ExPSAy在线服务器中的PortParam (https://web.expasy.org/protparam/ ) 在线软件分析计算SpG6P1E编码蛋白质的理化参数[12 ] ;使用PortParam在线软件对该蛋白质进行疏水性分析[12 ] ;利用phyre2 (http://www.sbg.bio.ic.ac.uk/phyre2/ )在线软件分析预测蛋白质二级结构[13 ] ;使用alphafold (https://alphafold.ebi.ac.uk/ ) 在线预测蛋白质三级结构[14 -15 ] ;使用Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/ )在线软件对该蛋白质的亚细胞定位进行分析[16 ] ;使用MEGA(Version11)进行蛋白的进化分析[17 ] ,用MEME在线网站进行motif分析[18 ] .使用TBtools对进化、保守域和motif进行可视化[19 ] . ...
The MEME suite
1
2015
... 序列比对分析使用序列分析软件DNAMAN(Version9)完成.羽毛针禾SpG6P1E的保守结构域分析使用NCBI网站在线工具ConservedDomains完成.使用ExPSAy在线服务器中的PortParam (https://web.expasy.org/protparam/ ) 在线软件分析计算SpG6P1E编码蛋白质的理化参数[12 ] ;使用PortParam在线软件对该蛋白质进行疏水性分析[12 ] ;利用phyre2 (http://www.sbg.bio.ic.ac.uk/phyre2/ )在线软件分析预测蛋白质二级结构[13 ] ;使用alphafold (https://alphafold.ebi.ac.uk/ ) 在线预测蛋白质三级结构[14 -15 ] ;使用Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/ )在线软件对该蛋白质的亚细胞定位进行分析[16 ] ;使用MEGA(Version11)进行蛋白的进化分析[17 ] ,用MEME在线网站进行motif分析[18 ] .使用TBtools对进化、保守域和motif进行可视化[19 ] . ...
TBtools-II:A" One for All, All for One" bioinformatics platform for biological big-data mining
1
2023
... 序列比对分析使用序列分析软件DNAMAN(Version9)完成.羽毛针禾SpG6P1E的保守结构域分析使用NCBI网站在线工具ConservedDomains完成.使用ExPSAy在线服务器中的PortParam (https://web.expasy.org/protparam/ ) 在线软件分析计算SpG6P1E编码蛋白质的理化参数[12 ] ;使用PortParam在线软件对该蛋白质进行疏水性分析[12 ] ;利用phyre2 (http://www.sbg.bio.ic.ac.uk/phyre2/ )在线软件分析预测蛋白质二级结构[13 ] ;使用alphafold (https://alphafold.ebi.ac.uk/ ) 在线预测蛋白质三级结构[14 -15 ] ;使用Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/ )在线软件对该蛋白质的亚细胞定位进行分析[16 ] ;使用MEGA(Version11)进行蛋白的进化分析[17 ] ,用MEME在线网站进行motif分析[18 ] .使用TBtools对进化、保守域和motif进行可视化[19 ] . ...
Selection of the reference genes for quantitative gene expression by RT-qPCR in the desert plant Stipagrostis pennata
1
2021
... 收取羽毛针禾根、节、叶、花、种子,GA3处理0、3、6、12、24 h的根,不同PEG6000浓度处理过的根等组织.提取这些组织的RNA,反转录得到cDNA,以反转录cDNA为模板,分别以羽毛针禾ELF 、GAPDH 、APR6 基因为内参基因[20 ] .利用Premier5.0软件设计内参基因Actin 以及SpG6P1E 引物(表1 ),参照荧光定量试剂盒说明书 (加拿大,ABM) 进行qRT-PCR (Quantitative real-time polymerase chain reaction) 反应,每个样本进行3个技术重复.反应结束后,根据目的基因和内参基因Ct 值,使用2-ΔΔ Ct
公路阻碍作用对古尔班通古特沙漠南缘羽毛针禾种群基因流的影响
1
2019
... 干旱是影响植物生长和作物产量的重要环境因素,近年来干旱胁迫对环境、作物生产的影响愈发严重[1 ] .新疆沙漠化问题突出,生态环境十分脆弱,严重影响着人们的生产生活.新疆的沙漠主要为流动沙丘,羽毛针禾主要生长在流动半流动沙地,是沙漠中的固沙先锋植物[21 ] .羽毛针禾具有抗旱、耐风蚀、耐沙埋等特点,尤其是根部具有特殊的沙套结构,对于其适应沙漠极端干旱环境具有重要作用.研究表明,在干旱胁迫处理后,根部沙套组织显著积累大多数糖类物质和部分氨基酸[7 ] ,尤其是糖类物质,表明其与干旱条件下的沙套生成密切相关. ...
Structure-based functional annotation:yeast ymr099c codes for a D-hexose-6-phosphate mutarotase
2
2006
... 异构酶广泛存在于植物中,可以通过催化底物生成同分异构体.葡萄糖代谢途径中包含多种异构酶类,其中G6P1E处在葡萄糖代谢途径的分支点,其功能是催化α-D-葡萄糖-6-磷酸和β-D-葡萄糖-6-磷酸间的转化和平衡[22 ] .该相互转化过程可为丝状真菌的生长和侵染寄主植物提供能量[23 -24 ] .G6P1E的底物范围较广,能催化葡萄糖-6-磷酸、半乳糖-6-磷酸和甘露糖-6-磷酸等己糖-6-磷酸从α到β的相互转化[22 ] ,表明其可能广泛参与了生物体内的糖代谢过程. ...
... [22 ],表明其可能广泛参与了生物体内的糖代谢过程. ...
Glucose-6-phosphate 1-Epimerase CrGlu6 contributes to development and biocontrol efficiency in clonostachys chloroleuca
1
2023
... 异构酶广泛存在于植物中,可以通过催化底物生成同分异构体.葡萄糖代谢途径中包含多种异构酶类,其中G6P1E处在葡萄糖代谢途径的分支点,其功能是催化α-D-葡萄糖-6-磷酸和β-D-葡萄糖-6-磷酸间的转化和平衡[22 ] .该相互转化过程可为丝状真菌的生长和侵染寄主植物提供能量[23 -24 ] .G6P1E的底物范围较广,能催化葡萄糖-6-磷酸、半乳糖-6-磷酸和甘露糖-6-磷酸等己糖-6-磷酸从α到β的相互转化[22 ] ,表明其可能广泛参与了生物体内的糖代谢过程. ...
Gene expression analysis of the biocontrol fungus Trichoderma harzianum in the presence of tomato plants,chitin,or glucose using a high-density oligonucleotide microarray
1
2009
... 异构酶广泛存在于植物中,可以通过催化底物生成同分异构体.葡萄糖代谢途径中包含多种异构酶类,其中G6P1E处在葡萄糖代谢途径的分支点,其功能是催化α-D-葡萄糖-6-磷酸和β-D-葡萄糖-6-磷酸间的转化和平衡[22 ] .该相互转化过程可为丝状真菌的生长和侵染寄主植物提供能量[23 -24 ] .G6P1E的底物范围较广,能催化葡萄糖-6-磷酸、半乳糖-6-磷酸和甘露糖-6-磷酸等己糖-6-磷酸从α到β的相互转化[22 ] ,表明其可能广泛参与了生物体内的糖代谢过程. ...
The intergenic interplay between aldose 1-epimerase-like protein and pectin methylesterase in abiotic and biotic stress control
1
2017
... SpG6P1E 基因与植物体内可溶性糖的含量显著相关.可溶性糖含量与花色在石竹属开花植物中显著相关,通过VIGS沉默掉DchG6P1E 基因后,酶活性与对照相比显著下降,葡萄糖含量显著降低,且花色出现明显变化,较对照整体变浅.这表明Dch6GP1E 通过参与糖的代谢过程,影响了植物体内可溶性糖的含量,进而影响中国石竹的花色[11 ] .烟草G6P1E 同源基因NbAELP 被敲除后,叶片中的葡萄糖含量显著降低,植物生长受到影响,抵御非生物胁迫和生物胁迫的能力降低,该研究表明G6P1E 通过影响烟草中的葡萄糖含量,进而影响其生长和抵御胁迫的能力[25 ] .本研究结果也表明羽毛针禾沙套发育过程中,SpG6P1E 基因的表达量与可溶性糖含量呈现显著正相关关系(图5 、6 ).此外,基因的亚细胞定位与功能发挥之间具有紧密的联系[26 ] .SpG6P1E 亚细胞定位于细胞质,可能参与了细胞质中糖的相关代谢过程.蛋白互作网络预测也显示SpG6P1E 和其互作蛋白参与了糖的合成与代谢等一系列过程(图7 、8 ). ...
海马齿 Spmet 基因表达产物的亚细胞定位及其组织表达特异性分析
1
2012
... SpG6P1E 基因与植物体内可溶性糖的含量显著相关.可溶性糖含量与花色在石竹属开花植物中显著相关,通过VIGS沉默掉DchG6P1E 基因后,酶活性与对照相比显著下降,葡萄糖含量显著降低,且花色出现明显变化,较对照整体变浅.这表明Dch6GP1E 通过参与糖的代谢过程,影响了植物体内可溶性糖的含量,进而影响中国石竹的花色[11 ] .烟草G6P1E 同源基因NbAELP 被敲除后,叶片中的葡萄糖含量显著降低,植物生长受到影响,抵御非生物胁迫和生物胁迫的能力降低,该研究表明G6P1E 通过影响烟草中的葡萄糖含量,进而影响其生长和抵御胁迫的能力[25 ] .本研究结果也表明羽毛针禾沙套发育过程中,SpG6P1E 基因的表达量与可溶性糖含量呈现显著正相关关系(图5 、6 ).此外,基因的亚细胞定位与功能发挥之间具有紧密的联系[26 ] .SpG6P1E 亚细胞定位于细胞质,可能参与了细胞质中糖的相关代谢过程.蛋白互作网络预测也显示SpG6P1E 和其互作蛋白参与了糖的合成与代谢等一系列过程(图7 、8 ). ...








 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}