0 引言
植物通过叶片气孔与外界进行物质和能量交换,叶片气孔开合程度可以调节和能量交换过程[1 -2 ] 。气孔导度是气孔开合程度的度量,分析气孔导度与周围各环境因子的响应关系,对于理解环境因子对气孔的作用方式及气孔对环境因子的响应过程至关重要[3 -4 ] 。水分利用效率是植物重要的生理特征,是植物单位耗水生产出的同化量,用于估算环境因素对植物生理过程的影响,一定程度上可以反映植物的环境适应能力[5 -6 ] 。植物通过调节叶片气孔的开合程度来控制CO2 吸收和水分的散失水平,实现对光合作用、蒸腾作用的影响[7 ] ,气孔导度深刻影响植物的水分利用效率。要认识气孔的环境响应机理需要长时间序列的气孔导度数据,数值模拟是满足这一要求的有效方法[8 ] 。进行叶片尺度气孔行为特征、对水分利用的影响及与环境因子的响应研究,并进行叶片气孔导度的模拟,对于理解区域植被和生态系统对环境的适应机制和进行水资源管理具有重要意义。
柽柳(Tamarix chinensis )具有耐盐碱和抗旱的特性,主要分布于盐碱地区和干旱地区,如华北滨海盐碱地区和西北干旱半干旱地区[9 ] ,也是汾河流域中下游地区较为典型的河岸带植被。目前,国内外针对柽柳的生理生态研究集中在柽柳生理生态特征变化、影响因子以及适应机制等方面[10 -13 ] ,对其气孔导度对水分利用的影响及气孔导度的数值模拟方面的研究较少,或者气孔导度数值模拟限于完全经验模型和半经验模型[14 -15 ] ,机理模型较少。本文研究柽柳气孔导度特征与各环境因子的响应关系,选用经验模型(Jarvis)、半经验模型(BWB)和气孔导度机理模型(USO)对柽柳气孔导度进行数值模拟,能够为理解柽柳叶片气孔行为的环境响应机理和区域植被与大气环境物质通量交换过程提供参考。
1 研究区概况
研究区位于山西省应县境内(39°17′—39°45′N,112°58′—113°37′E),地处桑干河中游,地貌类型主要是盆地、平川以及丘陵。河岸植物种类以及分布主要受到河岸水分条件和地貌类型的影响,柽柳群落主要分布在中游河漫滩及淤积河岸两侧。试验区属温带大陆性气候,气候受季风影响季节差异明显,降水集中在夏秋两季,年蒸发量大于1 800 mm,无霜期为100~140 d,年平均气温为7 ℃,平均年降水量360 mm。
2 材料与方法
2.1 材料及测定方法
于6—9月每月选择2 d晴朗日观测。选择3株柽柳作为观测对象,要求样本健康挺直且长势良好。在每棵树顶部选取3簇向阳的叶片,用Li-6400便携式光合作用系统(Li-cor,Lincoln,Nebraska,USA)测量,分别于06:00、10:00、14:00和18:00进行气体交换参数测定。测定参数包括光合生理参数和叶表面环境参数,其中光合生理参数有气孔导度(G s ,mol∙m-2 ∙s-1 ,以H2 O计量)、净光合速率(A ,μmol∙m-2 ∙s-1 ,以CO2 计量)和蒸腾速率(Tr ,mmol∙m-2 ∙s-1 ,以H2 O计量),根据测定值计算瞬时水分利用效率(WUE ,mmol∙mol-1 ),WUE=A /Tr 。叶表面环境参数有大气相对湿度(h s ,%)、光合有效辐射(PAR ,mmol∙m-2 ∙s-1 )、空气温度(T a ,℃)、大气CO2 浓度(C a ,mmol∙mol-1 )和水汽压亏缺(VPD ,KPa)。同时,在柽柳样地内安装自动气象站,对区域环境相对湿度和空气温度进行连续测定,使用Zeno-3200-A-D数据记录器每60 min进行记录。
2.2 模型描述
Jarvis模型是多元非线性模型,将最大的气孔导度参数值与数个环境限制因子相乘,表达式包含了PAR 、VPD 、T a 和C a 这几种特定环境因子对G s 的影响,是气孔导度经验模型[15 ] 。
G s =g P (PAR )g V (VPD )g C (C a )g T (T a )(1)
g P (PAR )是PAR 对G s 的响应函数,具体表达式有3种:
g p1 (PAR )=a 1 +a 2 PAR (2)
g p2 (PAR )=a 1 PAR /(a 2 +PAR )(3)
g p3 (PAR )=(a 1 +a 2 PAR /a 3 )/(1+PAR /a 3 )(4)
g V (VPD )是VPD 对G s 的响应函数,具体表达式有3种:
g V1 (VPD )=1-b 1 VPD (5)
g V2 (VPD )=1/(1+b 1 VPD )(6)
g V3 (VPD )=(1-b 1 VPD )/(1+b 2 VPD )(7)
g C (C a )和g T (T a )分别是C a 和T a 对G s 的响应函数,具体表达式如下:
g C (C a )=1-d 1 C a (8)
g T (T a )=C 1 +C 2 T a +C 3 T a 2 (9)
式中:拟合参数总共有9个,即a 1 ,a 2 ,a 3 ,b 1 ,b 2 ,c 1 ,c 2 ,c 3 和d 1 。根据Jarvis模型的表达式,将特定环境因子PAR 、VPD 、T a 和C a 对G s 的各自响应函数分别进行组合,总共组合成9种模型表达式,以下称该模型表达式为Jarvis模型。
研究者基于气孔导度和光合作用关系的研究,指出G s 与A 、h s 和C a 之间均存在相关关系,这主要是将植物叶片生理活动的影响考虑到模型函数中,并据此提出了半经验的BWB模型[15 ] 。气孔导度的表达式是以上3个因子的函数。
G s = g 0 + m A h s C a (10)
式中:A h s C a g 0 和m 是拟合参数。以下称该式为BWB模型。
USO模型是基于最优气孔行为理论并借鉴BWB模型提出的气孔导度机理模型,用来研究不同环境和生物因素对气孔行为的影响[14 ] 。气孔的最优化行为是指在某一时间段内,植物叶片实现最大化碳固定作用的同时产生最小化蒸腾作用,该模型表达式为:
G s = g 0 + 1.6 1 + g 1 V P D A C a (11)
式中:g 0 和g 1 是拟合参数。A C a V P D
2.3 模型评定
本文引用3个指标来评定模型的模拟精度,即决定系数(R 2 )、修正效率系数(MEC )和均方根误差(RMSE )[16 ] 。
M E C = 1 - ∑ i = 1 N ( | O i - M i | ) ∑ i = 1 N ( | O i - O i ¯ | ) (12)
R M S E = ∑ i = 1 N ( O i - M i ) 2 N (13)
式中:O i M i O i ¯ N 为样本总数。通过选定的模型评定指标对比叶片气孔导度观测值与各模型模拟值,用来评定各模型的模拟效果。根据选定的指标,当R 2 和MEC 的值较大而RMSE 的值较小时,该模型模拟效果较好,模拟精度相对较高。
2.4 数据分析
采用SPSS19.0、Excel2007和Origin8.0软件进行数据处理和统计分析。对于各模型表达式中拟合参数和各模型表达式的确定,利用生长季6—9月柽柳叶片G s 的实测数据采用SPSS19.0软件进行多元回归分析,用Origin8.0进行绘图。
3 结果与分析
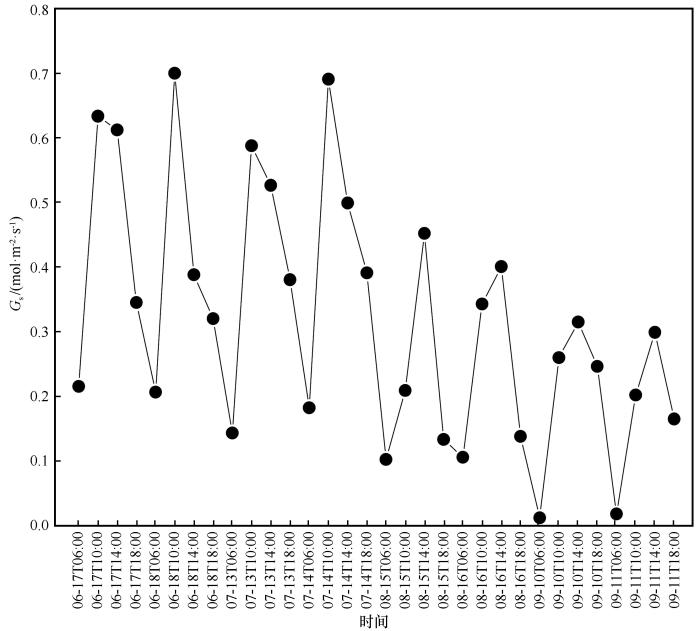
3.1 生长季期间气孔导度变化
柽柳气孔导度G s 日变化呈周期性,先升高后降低,整体呈倒“V”型(图1 )。柽柳G s 在06:00最小,然后升高,6、7月G s 在10:00左右达到峰值,8、9月G s 达到峰值的时间,在14:00左右。6—9月G s 平均峰值依次为0.67、0.64、0.43、0.31 mol∙m-2 ∙s-1 。6—9月G s 平均最小值依次为0.21、0.16、0.10、0.01 mol∙m-2 ∙s-1 。可见,整个生长季期间各月G s 均值整体呈现逐渐减小的趋势,G s 月变化大致呈降低趋势。
图1
图1
生长季气孔导度的日变化
Fig.1
Diurnal variation of stomatal conductance during the growing season
3.2 气孔导度影响因子
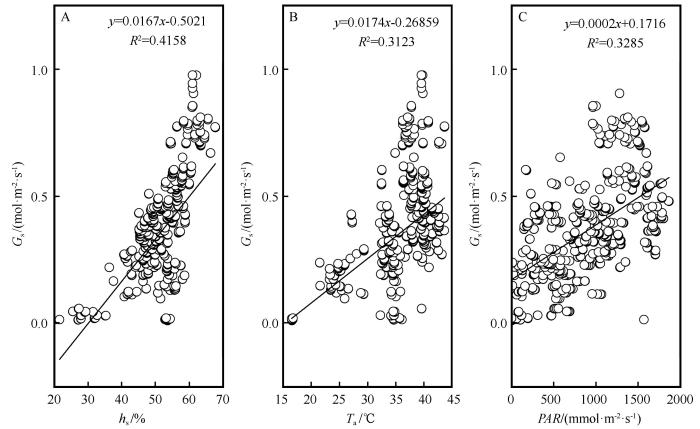
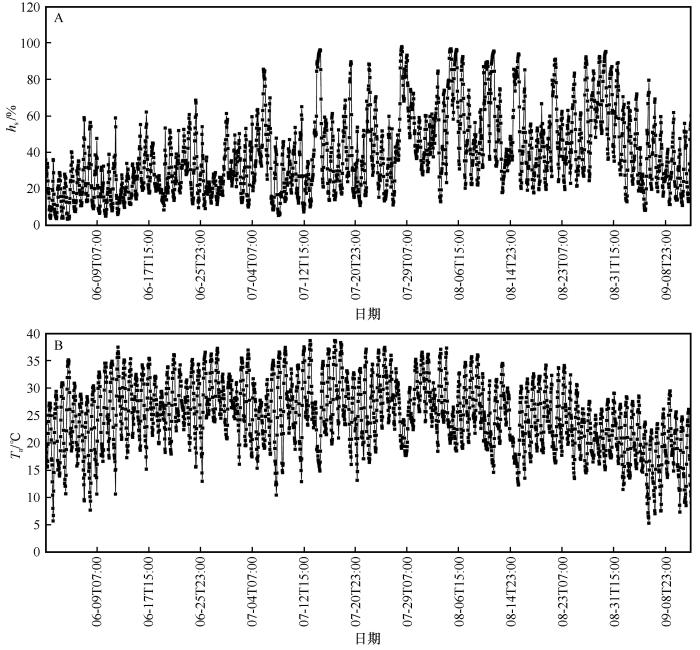
G s 与各环境因子的关联程度为h s (0.662)>PAR (0.613)>T a (0.601)>C a (0.223)>VPD (0.083,P <0.01)。h s 、PAR 和T a 是G s 的主要影响因子,其中h s 对G s 的影响最大,而C a 和VPD 对G s 的影响较小。对影响较大的3个主要环境因子h s 、PAR 、T a 与G s 进行了回归分析,决定系数R 2 分别是0.416、0.312和0.329(P <0.01)。随着h s 的增加G s 升高(图2 )。6—9月h s 均值依次分别为24.32%、39.25%、38.31%和24.15%(图3 )。G s 随T a 的增加而逐渐增大。当T a 超过某一界限时,G s 迅速降低。整个生长季期间T a 的平均值为24.9 ℃,范围为5.3~38.6 ℃。6—9月正午平均T a 依次分别为30.7、32.1、29.8 ℃和23.8 ℃(图3 )。6、7月正午平均T a 值较高导致柽柳出现光合午休现象,即气孔开合度减小,G s 降低,从而减少蒸腾和水分散失,致使14:00的G s 值下降,其值低于10:00的G s 值。可见G s 在T a 达到30 ℃后出现光合抑制现象从而G s 降低。光合有效辐射是影响植物叶片G s 的主要因子,随着PAR 的增加G s 升高。由于白天PAR 的增加,G s 逐渐增大。由于9月的h s 和T a 均值均最小,9月光照相对较弱,故9月G s 值最小。
图2
图2
主要环境因子对气孔导度的影响
Fig.2
Effects of major environmental factors on stomatal conductance
图3
图3
生长季期间大气相对湿度和气温的变化
Fig.3
The change in air humidity and temperature during the growing season
3.3 气孔导度对水分利用效率的影响
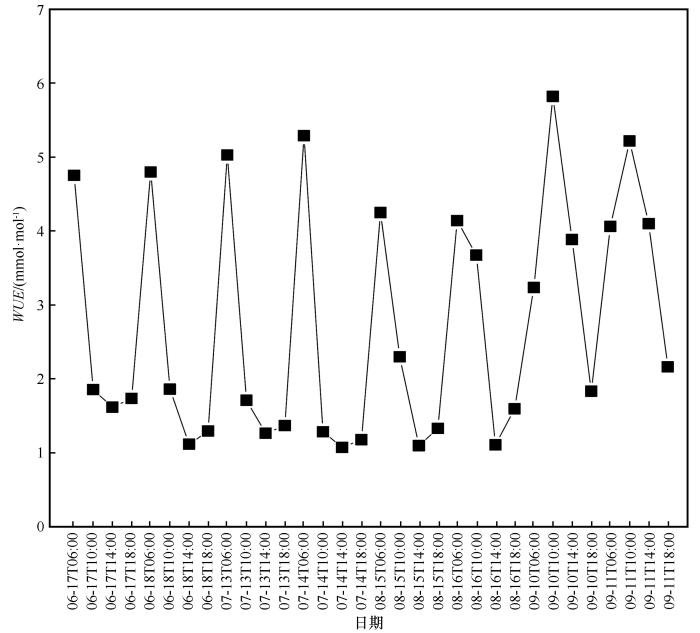
柽柳水分利用效率WUE 日变化呈周期性波动(图4 )。生长旺季(6—8月)柽柳WUE 值日变化为先减小后增加的趋势。06:00,WUE 达到最大值(平均值为4.71 mmol∙mol-1 ),从06:00开始降低,在14:00时WUE 达到最小值(平均值为1.21 mmol∙mol-1 )。在生长季末期(9月),WUE 日变化先增加后减小的趋势,整体呈倒“V”型。从06:00开始WUE 迅速增加,在10:00时达到最大值(平均值为5.52 mmol∙mol-1 ),继而降低。9月WUE 值高于其他月份。
图4
图4
生长季期间水分利用效率的日变化
Fig.4
Diurnal variation of WUE during the growing season
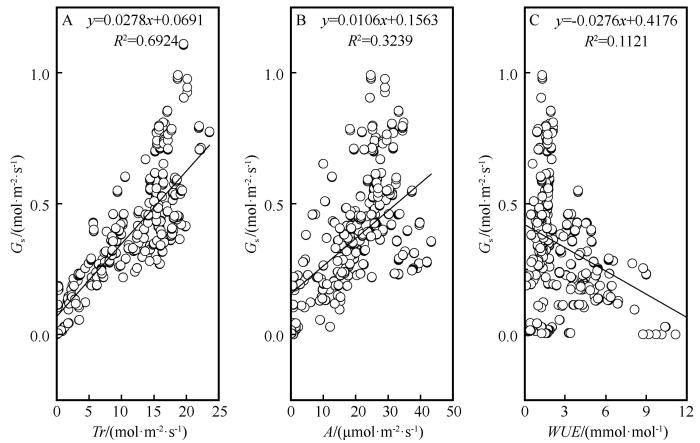
将净光合速率(A )、蒸腾速率(Tr )、水分利用效率(WUE )与G s 进行回归分析,决定系数R 2 分别是0.692、0.324、0.112(P <0.01)。G s 与Tr 和A 呈极显著正相关,两者变化同步,G s 的增加会引起Tr 和A 的增加(图5 )。G s 与WUE 呈极显著负相关,两者变化趋势相反。生长旺季(6—8月)和生长季末期(9月)WUE 具有不同的变化趋势,在生长旺季06:00—10:00 G s 急剧增加,对蒸腾作用的影响相对较大,从而导致WUE 的下降,在生长季末期06:00—10:00 G s 增加,对光合作用的影响相对较大,从而导致WUE 上升。
图5
图5
气孔导度和光合生理特征的关系
Fig.5
Relation between stomatal conductance and photosynthetic physiological characteristics
3.4 气孔导度模拟
3.4.1 基于Jarvis 模型的气孔导度模拟
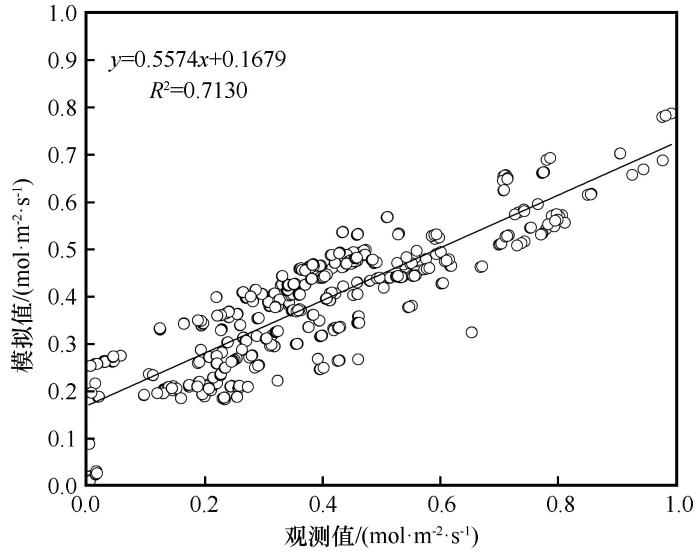
利用生长季6—9月柽柳叶片G s 的实测数据,对于各模型表达式中拟合参数和各模型表达式进行确定,进行多元回归分析。由于Jarvis模型中涉及的单一特定环境因子有多种表达式,在模型确定时需要选择决定系数相对较高的表达式,从而确定出对G s 影响最显著的拟合参数值。将确定的各拟合参数值代入,得到基于Jarvis模型进行G s 模拟的最优模型表达式为:
G s =0.081PAR /(2.796+PAR )×1/(1+58.659VPD )×
(5.353-0.492T a +0.013T a 2 )×(1+0.493C a )(14)
将生长季6—9月实测的T a 、VPD 、PAR 和C a 各值代入此最优模型表达式中,基于Jarvis模型从而模拟出柽柳叶片G s 的结果。图6 将G s 观测值与基于Jarvis模型计算的G s 模拟值进行了回归分析,决定系数R 2 =0.713(P <0.01)。
图6
图6
基于Jarvis模型气孔导度观测值与模拟值的回归分析
Fig.6
Regression analysis between the observed and the measured stomatal conductance based on Jarvis model
3.4.2 基于BWB 模型的气孔导度模拟
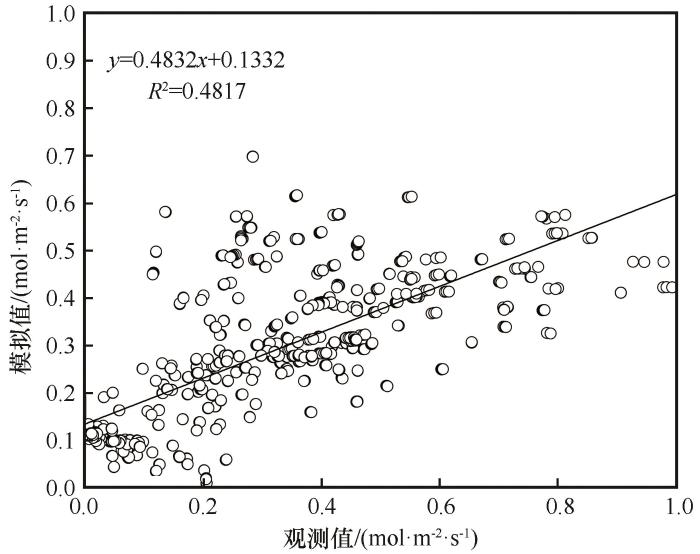
利用生长季6—9月柽柳叶片G s 的实测数据,对于模型表达式中拟合参数进行确定,进行多元回归分析,得到各拟合参数值。将确定的各拟合参数值代入,得到基于BWB模型进行G s 模拟的最优模型表达式为:
G s = 0.114 + 0.067 A h s C a (15)
将生长季6—9月实测的A 、h s 和C a 各值代入此最优模型表达式中,基于BWB模型从而模拟出柽柳叶片G s 的结果。图7 将G s 观测值与基于BWB模型计算的G s 模拟值进行了回归分析,决定系数R 2 =0.482(P <0.01)。
图7
图7
基于BWB模型气孔导度观测值与模拟值的回归分析
Fig.7
Regression analysis between the observed and the measured stomatal conductance based on the BWB model
3.4.3 基于USO 模型的气孔导度模拟
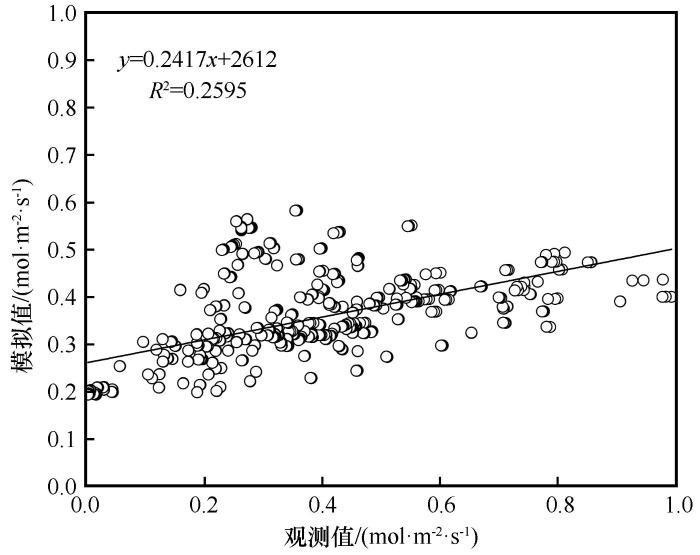
利用生长季6—9月柽柳叶片G s 的实测数据,对于模型表达式中拟合参数进行确定,进行多元回归分析,得到各拟合参数值。将确定的各拟合参数值代入,得到基于USO模型进行G s 模拟的最优模型表达式为:
G s = 0.194 + 1.6 1 + 1.015 V P D A C a (16)
将生长季6—9月实测的A 、VPD 和C a 各值代入此最优模型表达式中,基于USO模型从而模拟出柽柳叶片G s 的结果。图8 将G s 观测值与基于USO模型计算的G s 模拟值进行了回归分析,决定系数R 2 =0.260(P <0.01)。
图8
图8
基于USO模型气孔导度观测值与模拟值的回归分析
Fig.8
Regression analysis between the observed and the measured stomatal conductance based on the USO model
3.4.4 模型评定
从表1 可以看出,使用Jarvis模型进行G s 模拟后,修正效率系数(MEC )、均方根误差(RMSE )和决定系数值(R 2 )数值分别是0.409、0.024和0.713,使用Jarvis模型进行气孔导度模拟的MEC 和R 2 大于BWB模型和USO模型,且RMSE 最小,可知用Jarvis模型进行柽柳叶片G s 模拟的效果相对较好,准确性较高。相反,使用USO模型进行气孔导度模拟的MEC 和R 2 这两个指标均小于Jarvis模型和BWB模型,且RMSE 最大,可知用USO模型进行柽柳叶片G s 模拟的准确性较低,模拟效果较差。
4 讨论
本研究采用LI-6400光合作用测定系统,对生长季期间的柽柳气体交换参数进行测定,观测柽柳叶片G s 的日变化和月变化特征。结果显示在整个生长季G s 日变化呈现先增加后减少的周期性波动,G s 月变化整体呈现逐渐减小的趋势。先前研究表明,强光照引起的高温会导致叶片蒸腾速率加快,蒸腾作用过强使植物体内出现水分亏缺,会引起保卫细胞失水和气孔关闭,对气孔导度产生抑制作用[17 -18 ] 。自观测始点06:00开始,受PAR 和T a 迅速增加的影响,气孔开合增大,G s 迅速上升;6、7月午间T a 值较高(均值大于30 ℃),气孔关闭致使14:00 G s 值减小。这一现象与司建华等[19 ] 对黑河下游胡杨日变化研究结果一致。在8、9月午间柽柳G s 值在14:00左右达到最大值,而高冠龙等[15 ] 对黑河下游柽柳生长季G s 的研究表明其在10:00—12:00达到最大值,这与我们的研究结论并不相同,形成差异的原因可能在于研究期间环境条件的差异,即本研究中各月T a 差异明显,而黑河下游各月T a 变化基本不大。
研究发现环境因子h s 、PAR 、T a 与G s 的相关性较强,对柽柳G s 的影响较大,与G s 的相关性为h s (0.662)>PAR (0.613)>T a (0.601),C a 和VPD 对柽柳G s 的影响相对较小。司建华等[19 ] 发现黑河下游胡杨G s 与T a 、h s 有显著的相关关系,这与我们的研究结果一致,有学者认为C a 和VPD 是苹果树G s 主要影响因子[20 ] ,这与我们的研究结果形成差异,可能与研究区地理环境和物种有关。在与环境因子的响应关系中,叶片G s 和PAR 、h s 、T a 正相关,G s 随着PAR 、h s 、T a 这些影响因子的增加而增大,但超过影响因子阈值后G s 开始下降。本研究中G s 在T a 达到30 ℃后出现光合抑制现象从而G s 降低,我们认为影响柽柳G s 变化的T a 临界阈值可能是30 ℃左右。
G s 变化特征对植物光合作用、蒸腾作用等光合生理特性产生影响,进而能够影响植物的水分利用效率。水分利用效率是植物在单位水分消耗条件下进行碳固定的能力,能够反映植物有效的水分利用能力,也是衡量植物耐旱性和环境适应性的重要指标[21 ] 。生长旺季(6—8月)柽柳WUE 值日变化为先降低再升高。生长季末期(9月)WUE 日变化为先升高再降低。当G s 降低时,气孔关闭至几乎闭合时,水分可以继续通过角质层散失,所以蒸腾的降低比CO2 流入的降低程度小,使WUE 值降低。当G s 升高时,气孔张开至完全开放时,叶片内部存在扩散阻力,扩散阻力对CO2 流入的限制比对蒸腾的限制程度大,使WUE 值降低[18 ] 。可见,G s 升高或者降低都会引起WUE 的减小,只有当G s 变化使得水消耗和CO2 吸收实现最优协调时,WUE 才能达到最大值,因此WUE 出现波动性变化。研究发现,9月柽柳WUE 值相对较高,这是因为9月G s 整体较低,气孔部分关闭使得通过气孔进入叶片参与碳固定CO2 减少,从而导致光合速率降低,另一方面使得通过气孔蒸腾的水分减少,且蒸腾速率降低的幅度较大,所以植物的WUE 提高。降水集中在7、8月,9月h s 最低,则在整个生长季9月的干旱程度相对较高。我们的研究结果表明,柽柳具有较强的耐旱能力,能够通过G s 的调节提高WUE 值实现对水分的高效利用来适应干旱的环境。这与以往研究发现干旱地区植物有较高的水分利用效率,能通过提高水分利用效率来适应水分亏缺的生长环境一致[21 -24 ] 。可见,进行植物气孔特征及其对植物水分利用的影响研究,能够预测气候变化下气孔导度调节水分利用的动态变化,对于区域水资源管理具有重要的指导作用。
目前关于气孔导度的模拟研究,大多采用的是经验模型和半经验模型,本文运用Jarvis模型(经验模型)、BWB模型(半经验模型)和USO模型(机理模型)对柽柳气孔导度进行模拟,结果表明Jarvis模型比其他两类模型的模拟精度高。有学者对黑河下游胡杨和柽柳叶片气孔导度基于(半)经验模型模拟研究,同样得到Jarvis模型模拟精度较高的结论,研究认为在荒漠干旱地区Jarvis模型具有相对较好的适用性[15 ,17 ] 。本研究区域地处黄土高原,该区域干旱气候具有频次高、持续时间长且影响范围大的特点[25 ] ,也属于相对干旱的地区。因此,我们认为与半经验模型和气孔导度机理模型相比较而言,Jarvis模型对于干旱区的乔木、灌木等植物叶片气孔导度模拟效果较好。此外,Jarvis模型进行气孔导度模拟时,只需考虑单一环境因子作为气孔行为的影响因素,因此在现有的植物叶片气孔导度模拟中Jarvis模型得到广泛应用[26 -30 ] 。Jarvis模型为完全经验模型,在干旱半干旱地区,需要进一步探讨其对于不同植物种类叶片气孔导度模拟方面的适用性。值得一提的是,气孔导度机理模型用于研究不同环境和生物因素对气孔行为的影响,不仅考虑了叶片生理特性和气孔导度之间的关系,而且具有反馈机制,未来可以考虑将其和植被模型、生态系统模型等进行耦合,从而提高该模型模拟及预测的准确性。在干旱半干旱地区对叶片尺度气孔行为和影响因素进行深入研究,以及进行叶片气孔导度的模拟,不仅能够进行尺度扩展用于生态系统蒸散发估算和生态系统建模[31 -32 ] ,同时对于理解该区域植被和生态系统对环境的适应机制具有重要意义。
5 结论
大气相对湿度(h s )、光合有效辐射(PAR )和空气温度(T a )对柽柳叶片G s 影响较大,3个环境因子与柽柳叶片G s 的相关性为h s (0.662)>PAR (0.613)>T a (0.601)。柽柳叶片G s 日变化趋势为先升高后降低。06:00开始,随着PAR 增强和T a 升高,G s 升高,随后PAR 减弱和T a 降低,G s 降低。由于6、7月正午T a 过高引起叶片气孔闭合,致使6、7月G s 最大值出现在10:00左右,8、9月G s 最大值出现在14:00左右。在生长季末期,柽柳能够通过G s 的调节提高WUE 值,实现对水分的高效利用来适应干旱的环境。用经验模型(Jarvis)、半经验模型(BWB)和气孔导度机理模型(USO模拟G s ,模拟结果的决定系数(R 2 )、修正效率系数(MEC )和均方根误差(RMSE )分别是(0.713、0.409、0.024)、(0.482、0.309、0.158)和(0.260、0.135、0.255)。在3个模型中Jarvis模拟R 2 和MEC 值最大且RMSE 值最小,说明其模拟精度最高。
参考文献
View Option
[1]
石建红 ,周锁铨 ,余华 ,等 鄱阳湖流域典型树种夏季气孔导度模型及影响因素比较
[J].环境科学研究 ,2010 ,23 (1 ):33 -40 .
[本文引用: 1]
[2]
杜澜 ,夏捷 ,李海花 ,等 逐步失水过程中绿竹光响应进程及其拟合
[J].应用生态学报 ,2019 ,6 :2011 -2020 .
[本文引用: 1]
[3]
张中典 ,张大龙 ,李建明 ,等 黄瓜气孔导度、水力导度的环境响应及其调控蒸腾效应
[J].农业机械学报 ,2016 ,47 (6 ):139 -147 .
[本文引用: 1]
[4]
鱼腾飞 ,冯起 ,司建华 极端干旱区多枝柽柳叶片气孔导度的环境响应模拟
[J].植物生态学报 ,2012 ,36 (6 ):483 -491 .
[本文引用: 1]
[5]
Gago J Douthe C Florez-Sarasaet I et al Opportunities for improving leaf water use efficiency under climate change conditions
[J].Plant Science ,2014 ,226 :108 -119 .
[本文引用: 1]
[6]
Schwinning S Ehleringer J R Water use trade‐offs and optimal adaptations to pulse‐driven arid ecosystems
[J].Journal of Ecology ,2001 ,89 (3 ):464 -480 .
[本文引用: 1]
[7]
苏文华 ,张光飞 土壤温度与气温对紫花雪山报春光合作用和蒸腾作用的影响
[J].西北植物学报 ,2002 ,22 (4 ):824 -830 .
[本文引用: 1]
[8]
Hetherington A M Woodward F I The role of stomata in sensing and driving environmental change
[J].Nature ,2003 ,424 (6951 ):901 -908 .
[本文引用: 1]
[9]
陈亚宁 ,李卫红 ,周洪华 ,等 黑河下游荒漠河岸林植物水分传输观测试验研究
[J].北京师范大学学报(自然科学版) ,2016 ,52 (3 ):271 -276 .
[本文引用: 1]
[10]
刘冰 ,赵文智 荒漠绿洲过渡带柽柳和泡泡刺光合作用及水分代谢的生态适应性
[J].中国沙漠 ,2009 ,29 (1 ):101 -107 .
[本文引用: 1]
[11]
常宗强 ,冯起 ,张秀凤 ,等 CO2 浓度升高对极端干旱区多枝柽柳叶片水分利用效率的影响
[J].干旱区地理 ,2011 ,3 :499 -504 .
[12]
邓雄 ,李小明 ,张希明 ,等 4种荒漠植物气体交换特征的研究
[J].植物生态学报 ,2002 ,26 (5 ):605 -612 .
[13]
陈亚宁 ,李卫红 ,陈亚鹏 ,等 新疆塔里木河下游柽柳、芦苇对生态输水的响应
[J].冰川冻土 ,2004 ,26 (5 ):595 -601 .
[本文引用: 1]
[14]
范嘉智 ,王丹 ,胡亚林 ,等 最优气孔行为理论和气孔导度模拟
[J].植物生态学报 ,2016 ,40 (6 ):631 -642 .
[本文引用: 2]
[15]
高冠龙 ,冯起 ,刘贤德 ,等 三种经验模型模拟荒漠河岸柽柳叶片气孔导度
[J].生态学报 ,2020 ,40 (10 ):3486 -3494 .
[本文引用: 5]
[16]
Legates D R Mccabe G J Evaluating the use of “goodness‐of‐fit” measures in hydrologic and hydroclimatic model validation
[J].Water Resources Research ,1999 ,35 (1 ):233 -241 .
[本文引用: 1]
[17]
高冠龙 ,张小由 ,鱼腾飞 ,等 极端干旱条件下胡杨叶片气孔导度模拟
[J].干旱区地理 ,2016 ,39 (3 ):607 -612 .
[本文引用: 2]
[18]
邓雄 ,李小明 ,张希明 ,等 多枝柽柳气体交换特性研究
[J].生态学报 ,2003 ,23 (1 ):180 -187 .
[本文引用: 2]
[19]
司建华 ,常宗强 ,苏永红 ,等 胡杨叶片气孔导度特征及其对环境因子的响应
[J].西北植物学报 ,2008 ,28 (1 ):125 -130 .
[本文引用: 2]
[20]
苗玉 ,高冠龙 ,李伟 黄土高原苹果树叶片气孔导度的环境响应与模拟
[J].干旱区地理 ,2021 ,44 (2 ):525 -533 .
[本文引用: 1]
[21]
孟阳阳 ,刘冰 ,刘婵 水盐梯度下湿地柽柳(Tamarix ramosissima )光合响应特征和水分利用效率
[J].中国沙漠 ,2018 ,38 (3 ):568 -577 .
[本文引用: 2]
[22]
邢鹏 ,臧永新 ,闵孝君 ,等 塔克拉玛干沙漠南缘防护林和自然群落中多枝柽柳的光合特征及水分利用效率
[J].应用生态学报 ,2019 ,30 (3 ):61 -69 .
[23]
张佩 ,袁国富 ,庄伟 ,等 黑河中游荒漠绿洲过渡带多枝柽柳对地下水位变化的生理生态响应与适应
[J].生态学报 ,2011 ,31 (22 ):6677 -6687 .
[24]
解婷婷 ,苏培玺 ,丁松爽 黑河中游边缘绿洲不同水分条件对青贮玉米叶片光合特性及产量的影响
[J].西北农业学报 ,2009 ,6 :127 -133 .
[本文引用: 1]
[25]
张晨 ,王翠红 ,严俊霞 ,等 1971-2011年汾河流域降水时空特征
[J].干旱区研究 ,2018 ,35 (1 ):57 -67 .
[本文引用: 1]
[26]
Li G Lin L Dong Y Y et al Testing two models for the estimation of leaf stomatal conductance in four greenhouse crops cucumber,chrysanthemum,tulip and lilium
[J].Agricultural and Forest Meteorology ,2012 ,165 :92 -103 .
[本文引用: 1]
[27]
王秋玲 ,周广胜 春玉米持续干旱过程中常用气孔导度模型的比较研究
[J].生态学报 ,2018 ,38 (19 ):6846 -6856 .
[28]
王海珍 ,韩路 ,徐雅丽 ,等 灰胡杨叶片气孔导度特征及数值模拟
[J].林业科学 ,2016 ,52 (1 ):136 -142 .
[29]
罗紫东 ,关华德 ,章新平 ,等 桂花树冠层气孔导度模型的优化及其参数分析
[J].生态学报 ,2016 ,36 (13 ):3995 -4005 .
[30]
Bauerle W L Daniels A B Barnard D M Carbon and water flux responses to physiology by environment interactions:a sensitivity analysis of variation in climate on photosynthetic and stomatal parameters
[J].Climate Dynamics ,2014 ,42 (9 ):2539 -2554 .
[本文引用: 1]
[31]
高冠龙 ,张小由 ,常宗强 ,等 植物气孔导度的环境响应模拟及其尺度扩展
[J].生态学报 ,2016 ,36 (6 ):1491 -1500 .
[本文引用: 1]
[32]
苏里坦 ,阿迪力·吐拉尔别克 ,王兴勇 ,等 地下变水位条件下塔里木河下游河岸胡杨林蒸腾模型
[J].干旱区地理 ,2014 ,37 (5 ):916 -921 .
[本文引用: 1]
鄱阳湖流域典型树种夏季气孔导度模型及影响因素比较
1
2010
... 植物通过叶片气孔与外界进行物质和能量交换,叶片气孔开合程度可以调节和能量交换过程[1 -2 ] .气孔导度是气孔开合程度的度量,分析气孔导度与周围各环境因子的响应关系,对于理解环境因子对气孔的作用方式及气孔对环境因子的响应过程至关重要[3 -4 ] .水分利用效率是植物重要的生理特征,是植物单位耗水生产出的同化量,用于估算环境因素对植物生理过程的影响,一定程度上可以反映植物的环境适应能力[5 -6 ] .植物通过调节叶片气孔的开合程度来控制CO2 吸收和水分的散失水平,实现对光合作用、蒸腾作用的影响[7 ] ,气孔导度深刻影响植物的水分利用效率.要认识气孔的环境响应机理需要长时间序列的气孔导度数据,数值模拟是满足这一要求的有效方法[8 ] .进行叶片尺度气孔行为特征、对水分利用的影响及与环境因子的响应研究,并进行叶片气孔导度的模拟,对于理解区域植被和生态系统对环境的适应机制和进行水资源管理具有重要意义. ...
逐步失水过程中绿竹光响应进程及其拟合
1
2019
... 植物通过叶片气孔与外界进行物质和能量交换,叶片气孔开合程度可以调节和能量交换过程[1 -2 ] .气孔导度是气孔开合程度的度量,分析气孔导度与周围各环境因子的响应关系,对于理解环境因子对气孔的作用方式及气孔对环境因子的响应过程至关重要[3 -4 ] .水分利用效率是植物重要的生理特征,是植物单位耗水生产出的同化量,用于估算环境因素对植物生理过程的影响,一定程度上可以反映植物的环境适应能力[5 -6 ] .植物通过调节叶片气孔的开合程度来控制CO2 吸收和水分的散失水平,实现对光合作用、蒸腾作用的影响[7 ] ,气孔导度深刻影响植物的水分利用效率.要认识气孔的环境响应机理需要长时间序列的气孔导度数据,数值模拟是满足这一要求的有效方法[8 ] .进行叶片尺度气孔行为特征、对水分利用的影响及与环境因子的响应研究,并进行叶片气孔导度的模拟,对于理解区域植被和生态系统对环境的适应机制和进行水资源管理具有重要意义. ...
黄瓜气孔导度、水力导度的环境响应及其调控蒸腾效应
1
2016
... 植物通过叶片气孔与外界进行物质和能量交换,叶片气孔开合程度可以调节和能量交换过程[1 -2 ] .气孔导度是气孔开合程度的度量,分析气孔导度与周围各环境因子的响应关系,对于理解环境因子对气孔的作用方式及气孔对环境因子的响应过程至关重要[3 -4 ] .水分利用效率是植物重要的生理特征,是植物单位耗水生产出的同化量,用于估算环境因素对植物生理过程的影响,一定程度上可以反映植物的环境适应能力[5 -6 ] .植物通过调节叶片气孔的开合程度来控制CO2 吸收和水分的散失水平,实现对光合作用、蒸腾作用的影响[7 ] ,气孔导度深刻影响植物的水分利用效率.要认识气孔的环境响应机理需要长时间序列的气孔导度数据,数值模拟是满足这一要求的有效方法[8 ] .进行叶片尺度气孔行为特征、对水分利用的影响及与环境因子的响应研究,并进行叶片气孔导度的模拟,对于理解区域植被和生态系统对环境的适应机制和进行水资源管理具有重要意义. ...
极端干旱区多枝柽柳叶片气孔导度的环境响应模拟
1
2012
... 植物通过叶片气孔与外界进行物质和能量交换,叶片气孔开合程度可以调节和能量交换过程[1 -2 ] .气孔导度是气孔开合程度的度量,分析气孔导度与周围各环境因子的响应关系,对于理解环境因子对气孔的作用方式及气孔对环境因子的响应过程至关重要[3 -4 ] .水分利用效率是植物重要的生理特征,是植物单位耗水生产出的同化量,用于估算环境因素对植物生理过程的影响,一定程度上可以反映植物的环境适应能力[5 -6 ] .植物通过调节叶片气孔的开合程度来控制CO2 吸收和水分的散失水平,实现对光合作用、蒸腾作用的影响[7 ] ,气孔导度深刻影响植物的水分利用效率.要认识气孔的环境响应机理需要长时间序列的气孔导度数据,数值模拟是满足这一要求的有效方法[8 ] .进行叶片尺度气孔行为特征、对水分利用的影响及与环境因子的响应研究,并进行叶片气孔导度的模拟,对于理解区域植被和生态系统对环境的适应机制和进行水资源管理具有重要意义. ...
Opportunities for improving leaf water use efficiency under climate change conditions
1
2014
... 植物通过叶片气孔与外界进行物质和能量交换,叶片气孔开合程度可以调节和能量交换过程[1 -2 ] .气孔导度是气孔开合程度的度量,分析气孔导度与周围各环境因子的响应关系,对于理解环境因子对气孔的作用方式及气孔对环境因子的响应过程至关重要[3 -4 ] .水分利用效率是植物重要的生理特征,是植物单位耗水生产出的同化量,用于估算环境因素对植物生理过程的影响,一定程度上可以反映植物的环境适应能力[5 -6 ] .植物通过调节叶片气孔的开合程度来控制CO2 吸收和水分的散失水平,实现对光合作用、蒸腾作用的影响[7 ] ,气孔导度深刻影响植物的水分利用效率.要认识气孔的环境响应机理需要长时间序列的气孔导度数据,数值模拟是满足这一要求的有效方法[8 ] .进行叶片尺度气孔行为特征、对水分利用的影响及与环境因子的响应研究,并进行叶片气孔导度的模拟,对于理解区域植被和生态系统对环境的适应机制和进行水资源管理具有重要意义. ...
Water use trade‐offs and optimal adaptations to pulse‐driven arid ecosystems
1
2001
... 植物通过叶片气孔与外界进行物质和能量交换,叶片气孔开合程度可以调节和能量交换过程[1 -2 ] .气孔导度是气孔开合程度的度量,分析气孔导度与周围各环境因子的响应关系,对于理解环境因子对气孔的作用方式及气孔对环境因子的响应过程至关重要[3 -4 ] .水分利用效率是植物重要的生理特征,是植物单位耗水生产出的同化量,用于估算环境因素对植物生理过程的影响,一定程度上可以反映植物的环境适应能力[5 -6 ] .植物通过调节叶片气孔的开合程度来控制CO2 吸收和水分的散失水平,实现对光合作用、蒸腾作用的影响[7 ] ,气孔导度深刻影响植物的水分利用效率.要认识气孔的环境响应机理需要长时间序列的气孔导度数据,数值模拟是满足这一要求的有效方法[8 ] .进行叶片尺度气孔行为特征、对水分利用的影响及与环境因子的响应研究,并进行叶片气孔导度的模拟,对于理解区域植被和生态系统对环境的适应机制和进行水资源管理具有重要意义. ...
土壤温度与气温对紫花雪山报春光合作用和蒸腾作用的影响
1
2002
... 植物通过叶片气孔与外界进行物质和能量交换,叶片气孔开合程度可以调节和能量交换过程[1 -2 ] .气孔导度是气孔开合程度的度量,分析气孔导度与周围各环境因子的响应关系,对于理解环境因子对气孔的作用方式及气孔对环境因子的响应过程至关重要[3 -4 ] .水分利用效率是植物重要的生理特征,是植物单位耗水生产出的同化量,用于估算环境因素对植物生理过程的影响,一定程度上可以反映植物的环境适应能力[5 -6 ] .植物通过调节叶片气孔的开合程度来控制CO2 吸收和水分的散失水平,实现对光合作用、蒸腾作用的影响[7 ] ,气孔导度深刻影响植物的水分利用效率.要认识气孔的环境响应机理需要长时间序列的气孔导度数据,数值模拟是满足这一要求的有效方法[8 ] .进行叶片尺度气孔行为特征、对水分利用的影响及与环境因子的响应研究,并进行叶片气孔导度的模拟,对于理解区域植被和生态系统对环境的适应机制和进行水资源管理具有重要意义. ...
The role of stomata in sensing and driving environmental change
1
2003
... 植物通过叶片气孔与外界进行物质和能量交换,叶片气孔开合程度可以调节和能量交换过程[1 -2 ] .气孔导度是气孔开合程度的度量,分析气孔导度与周围各环境因子的响应关系,对于理解环境因子对气孔的作用方式及气孔对环境因子的响应过程至关重要[3 -4 ] .水分利用效率是植物重要的生理特征,是植物单位耗水生产出的同化量,用于估算环境因素对植物生理过程的影响,一定程度上可以反映植物的环境适应能力[5 -6 ] .植物通过调节叶片气孔的开合程度来控制CO2 吸收和水分的散失水平,实现对光合作用、蒸腾作用的影响[7 ] ,气孔导度深刻影响植物的水分利用效率.要认识气孔的环境响应机理需要长时间序列的气孔导度数据,数值模拟是满足这一要求的有效方法[8 ] .进行叶片尺度气孔行为特征、对水分利用的影响及与环境因子的响应研究,并进行叶片气孔导度的模拟,对于理解区域植被和生态系统对环境的适应机制和进行水资源管理具有重要意义. ...
黑河下游荒漠河岸林植物水分传输观测试验研究
1
2016
... 柽柳(Tamarix chinensis )具有耐盐碱和抗旱的特性,主要分布于盐碱地区和干旱地区,如华北滨海盐碱地区和西北干旱半干旱地区[9 ] ,也是汾河流域中下游地区较为典型的河岸带植被.目前,国内外针对柽柳的生理生态研究集中在柽柳生理生态特征变化、影响因子以及适应机制等方面[10 -13 ] ,对其气孔导度对水分利用的影响及气孔导度的数值模拟方面的研究较少,或者气孔导度数值模拟限于完全经验模型和半经验模型[14 -15 ] ,机理模型较少.本文研究柽柳气孔导度特征与各环境因子的响应关系,选用经验模型(Jarvis)、半经验模型(BWB)和气孔导度机理模型(USO)对柽柳气孔导度进行数值模拟,能够为理解柽柳叶片气孔行为的环境响应机理和区域植被与大气环境物质通量交换过程提供参考. ...
荒漠绿洲过渡带柽柳和泡泡刺光合作用及水分代谢的生态适应性
1
2009
... 柽柳(Tamarix chinensis )具有耐盐碱和抗旱的特性,主要分布于盐碱地区和干旱地区,如华北滨海盐碱地区和西北干旱半干旱地区[9 ] ,也是汾河流域中下游地区较为典型的河岸带植被.目前,国内外针对柽柳的生理生态研究集中在柽柳生理生态特征变化、影响因子以及适应机制等方面[10 -13 ] ,对其气孔导度对水分利用的影响及气孔导度的数值模拟方面的研究较少,或者气孔导度数值模拟限于完全经验模型和半经验模型[14 -15 ] ,机理模型较少.本文研究柽柳气孔导度特征与各环境因子的响应关系,选用经验模型(Jarvis)、半经验模型(BWB)和气孔导度机理模型(USO)对柽柳气孔导度进行数值模拟,能够为理解柽柳叶片气孔行为的环境响应机理和区域植被与大气环境物质通量交换过程提供参考. ...
CO2 浓度升高对极端干旱区多枝柽柳叶片水分利用效率的影响
0
2011
新疆塔里木河下游柽柳、芦苇对生态输水的响应
1
2004
... 柽柳(Tamarix chinensis )具有耐盐碱和抗旱的特性,主要分布于盐碱地区和干旱地区,如华北滨海盐碱地区和西北干旱半干旱地区[9 ] ,也是汾河流域中下游地区较为典型的河岸带植被.目前,国内外针对柽柳的生理生态研究集中在柽柳生理生态特征变化、影响因子以及适应机制等方面[10 -13 ] ,对其气孔导度对水分利用的影响及气孔导度的数值模拟方面的研究较少,或者气孔导度数值模拟限于完全经验模型和半经验模型[14 -15 ] ,机理模型较少.本文研究柽柳气孔导度特征与各环境因子的响应关系,选用经验模型(Jarvis)、半经验模型(BWB)和气孔导度机理模型(USO)对柽柳气孔导度进行数值模拟,能够为理解柽柳叶片气孔行为的环境响应机理和区域植被与大气环境物质通量交换过程提供参考. ...
最优气孔行为理论和气孔导度模拟
2
2016
... 柽柳(Tamarix chinensis )具有耐盐碱和抗旱的特性,主要分布于盐碱地区和干旱地区,如华北滨海盐碱地区和西北干旱半干旱地区[9 ] ,也是汾河流域中下游地区较为典型的河岸带植被.目前,国内外针对柽柳的生理生态研究集中在柽柳生理生态特征变化、影响因子以及适应机制等方面[10 -13 ] ,对其气孔导度对水分利用的影响及气孔导度的数值模拟方面的研究较少,或者气孔导度数值模拟限于完全经验模型和半经验模型[14 -15 ] ,机理模型较少.本文研究柽柳气孔导度特征与各环境因子的响应关系,选用经验模型(Jarvis)、半经验模型(BWB)和气孔导度机理模型(USO)对柽柳气孔导度进行数值模拟,能够为理解柽柳叶片气孔行为的环境响应机理和区域植被与大气环境物质通量交换过程提供参考. ...
... USO模型是基于最优气孔行为理论并借鉴BWB模型提出的气孔导度机理模型,用来研究不同环境和生物因素对气孔行为的影响[14 ] .气孔的最优化行为是指在某一时间段内,植物叶片实现最大化碳固定作用的同时产生最小化蒸腾作用,该模型表达式为: ...
三种经验模型模拟荒漠河岸柽柳叶片气孔导度
5
2020
... 柽柳(Tamarix chinensis )具有耐盐碱和抗旱的特性,主要分布于盐碱地区和干旱地区,如华北滨海盐碱地区和西北干旱半干旱地区[9 ] ,也是汾河流域中下游地区较为典型的河岸带植被.目前,国内外针对柽柳的生理生态研究集中在柽柳生理生态特征变化、影响因子以及适应机制等方面[10 -13 ] ,对其气孔导度对水分利用的影响及气孔导度的数值模拟方面的研究较少,或者气孔导度数值模拟限于完全经验模型和半经验模型[14 -15 ] ,机理模型较少.本文研究柽柳气孔导度特征与各环境因子的响应关系,选用经验模型(Jarvis)、半经验模型(BWB)和气孔导度机理模型(USO)对柽柳气孔导度进行数值模拟,能够为理解柽柳叶片气孔行为的环境响应机理和区域植被与大气环境物质通量交换过程提供参考. ...
... Jarvis模型是多元非线性模型,将最大的气孔导度参数值与数个环境限制因子相乘,表达式包含了PAR 、VPD 、T a 和C a 这几种特定环境因子对G s 的影响,是气孔导度经验模型[15 ] . ...
... 研究者基于气孔导度和光合作用关系的研究,指出G s 与A 、h s 和C a 之间均存在相关关系,这主要是将植物叶片生理活动的影响考虑到模型函数中,并据此提出了半经验的BWB模型[15 ] .气孔导度的表达式是以上3个因子的函数. ...
... 本研究采用LI-6400光合作用测定系统,对生长季期间的柽柳气体交换参数进行测定,观测柽柳叶片G s 的日变化和月变化特征.结果显示在整个生长季G s 日变化呈现先增加后减少的周期性波动,G s 月变化整体呈现逐渐减小的趋势.先前研究表明,强光照引起的高温会导致叶片蒸腾速率加快,蒸腾作用过强使植物体内出现水分亏缺,会引起保卫细胞失水和气孔关闭,对气孔导度产生抑制作用[17 -18 ] .自观测始点06:00开始,受PAR 和T a 迅速增加的影响,气孔开合增大,G s 迅速上升;6、7月午间T a 值较高(均值大于30 ℃),气孔关闭致使14:00 G s 值减小.这一现象与司建华等[19 ] 对黑河下游胡杨日变化研究结果一致.在8、9月午间柽柳G s 值在14:00左右达到最大值,而高冠龙等[15 ] 对黑河下游柽柳生长季G s 的研究表明其在10:00—12:00达到最大值,这与我们的研究结论并不相同,形成差异的原因可能在于研究期间环境条件的差异,即本研究中各月T a 差异明显,而黑河下游各月T a 变化基本不大. ...
... 目前关于气孔导度的模拟研究,大多采用的是经验模型和半经验模型,本文运用Jarvis模型(经验模型)、BWB模型(半经验模型)和USO模型(机理模型)对柽柳气孔导度进行模拟,结果表明Jarvis模型比其他两类模型的模拟精度高.有学者对黑河下游胡杨和柽柳叶片气孔导度基于(半)经验模型模拟研究,同样得到Jarvis模型模拟精度较高的结论,研究认为在荒漠干旱地区Jarvis模型具有相对较好的适用性[15 ,17 ] .本研究区域地处黄土高原,该区域干旱气候具有频次高、持续时间长且影响范围大的特点[25 ] ,也属于相对干旱的地区.因此,我们认为与半经验模型和气孔导度机理模型相比较而言,Jarvis模型对于干旱区的乔木、灌木等植物叶片气孔导度模拟效果较好.此外,Jarvis模型进行气孔导度模拟时,只需考虑单一环境因子作为气孔行为的影响因素,因此在现有的植物叶片气孔导度模拟中Jarvis模型得到广泛应用[26 -30 ] .Jarvis模型为完全经验模型,在干旱半干旱地区,需要进一步探讨其对于不同植物种类叶片气孔导度模拟方面的适用性.值得一提的是,气孔导度机理模型用于研究不同环境和生物因素对气孔行为的影响,不仅考虑了叶片生理特性和气孔导度之间的关系,而且具有反馈机制,未来可以考虑将其和植被模型、生态系统模型等进行耦合,从而提高该模型模拟及预测的准确性.在干旱半干旱地区对叶片尺度气孔行为和影响因素进行深入研究,以及进行叶片气孔导度的模拟,不仅能够进行尺度扩展用于生态系统蒸散发估算和生态系统建模[31 -32 ] ,同时对于理解该区域植被和生态系统对环境的适应机制具有重要意义. ...
Evaluating the use of “goodness‐of‐fit” measures in hydrologic and hydroclimatic model validation
1
1999
... 本文引用3个指标来评定模型的模拟精度,即决定系数(R 2 )、修正效率系数(MEC )和均方根误差(RMSE )[16 ] . ...
极端干旱条件下胡杨叶片气孔导度模拟
2
2016
... 本研究采用LI-6400光合作用测定系统,对生长季期间的柽柳气体交换参数进行测定,观测柽柳叶片G s 的日变化和月变化特征.结果显示在整个生长季G s 日变化呈现先增加后减少的周期性波动,G s 月变化整体呈现逐渐减小的趋势.先前研究表明,强光照引起的高温会导致叶片蒸腾速率加快,蒸腾作用过强使植物体内出现水分亏缺,会引起保卫细胞失水和气孔关闭,对气孔导度产生抑制作用[17 -18 ] .自观测始点06:00开始,受PAR 和T a 迅速增加的影响,气孔开合增大,G s 迅速上升;6、7月午间T a 值较高(均值大于30 ℃),气孔关闭致使14:00 G s 值减小.这一现象与司建华等[19 ] 对黑河下游胡杨日变化研究结果一致.在8、9月午间柽柳G s 值在14:00左右达到最大值,而高冠龙等[15 ] 对黑河下游柽柳生长季G s 的研究表明其在10:00—12:00达到最大值,这与我们的研究结论并不相同,形成差异的原因可能在于研究期间环境条件的差异,即本研究中各月T a 差异明显,而黑河下游各月T a 变化基本不大. ...
... 目前关于气孔导度的模拟研究,大多采用的是经验模型和半经验模型,本文运用Jarvis模型(经验模型)、BWB模型(半经验模型)和USO模型(机理模型)对柽柳气孔导度进行模拟,结果表明Jarvis模型比其他两类模型的模拟精度高.有学者对黑河下游胡杨和柽柳叶片气孔导度基于(半)经验模型模拟研究,同样得到Jarvis模型模拟精度较高的结论,研究认为在荒漠干旱地区Jarvis模型具有相对较好的适用性[15 ,17 ] .本研究区域地处黄土高原,该区域干旱气候具有频次高、持续时间长且影响范围大的特点[25 ] ,也属于相对干旱的地区.因此,我们认为与半经验模型和气孔导度机理模型相比较而言,Jarvis模型对于干旱区的乔木、灌木等植物叶片气孔导度模拟效果较好.此外,Jarvis模型进行气孔导度模拟时,只需考虑单一环境因子作为气孔行为的影响因素,因此在现有的植物叶片气孔导度模拟中Jarvis模型得到广泛应用[26 -30 ] .Jarvis模型为完全经验模型,在干旱半干旱地区,需要进一步探讨其对于不同植物种类叶片气孔导度模拟方面的适用性.值得一提的是,气孔导度机理模型用于研究不同环境和生物因素对气孔行为的影响,不仅考虑了叶片生理特性和气孔导度之间的关系,而且具有反馈机制,未来可以考虑将其和植被模型、生态系统模型等进行耦合,从而提高该模型模拟及预测的准确性.在干旱半干旱地区对叶片尺度气孔行为和影响因素进行深入研究,以及进行叶片气孔导度的模拟,不仅能够进行尺度扩展用于生态系统蒸散发估算和生态系统建模[31 -32 ] ,同时对于理解该区域植被和生态系统对环境的适应机制具有重要意义. ...
多枝柽柳气体交换特性研究
2
2003
... 本研究采用LI-6400光合作用测定系统,对生长季期间的柽柳气体交换参数进行测定,观测柽柳叶片G s 的日变化和月变化特征.结果显示在整个生长季G s 日变化呈现先增加后减少的周期性波动,G s 月变化整体呈现逐渐减小的趋势.先前研究表明,强光照引起的高温会导致叶片蒸腾速率加快,蒸腾作用过强使植物体内出现水分亏缺,会引起保卫细胞失水和气孔关闭,对气孔导度产生抑制作用[17 -18 ] .自观测始点06:00开始,受PAR 和T a 迅速增加的影响,气孔开合增大,G s 迅速上升;6、7月午间T a 值较高(均值大于30 ℃),气孔关闭致使14:00 G s 值减小.这一现象与司建华等[19 ] 对黑河下游胡杨日变化研究结果一致.在8、9月午间柽柳G s 值在14:00左右达到最大值,而高冠龙等[15 ] 对黑河下游柽柳生长季G s 的研究表明其在10:00—12:00达到最大值,这与我们的研究结论并不相同,形成差异的原因可能在于研究期间环境条件的差异,即本研究中各月T a 差异明显,而黑河下游各月T a 变化基本不大. ...
... G s 变化特征对植物光合作用、蒸腾作用等光合生理特性产生影响,进而能够影响植物的水分利用效率.水分利用效率是植物在单位水分消耗条件下进行碳固定的能力,能够反映植物有效的水分利用能力,也是衡量植物耐旱性和环境适应性的重要指标[21 ] .生长旺季(6—8月)柽柳WUE 值日变化为先降低再升高.生长季末期(9月)WUE 日变化为先升高再降低.当G s 降低时,气孔关闭至几乎闭合时,水分可以继续通过角质层散失,所以蒸腾的降低比CO2 流入的降低程度小,使WUE 值降低.当G s 升高时,气孔张开至完全开放时,叶片内部存在扩散阻力,扩散阻力对CO2 流入的限制比对蒸腾的限制程度大,使WUE 值降低[18 ] .可见,G s 升高或者降低都会引起WUE 的减小,只有当G s 变化使得水消耗和CO2 吸收实现最优协调时,WUE 才能达到最大值,因此WUE 出现波动性变化.研究发现,9月柽柳WUE 值相对较高,这是因为9月G s 整体较低,气孔部分关闭使得通过气孔进入叶片参与碳固定CO2 减少,从而导致光合速率降低,另一方面使得通过气孔蒸腾的水分减少,且蒸腾速率降低的幅度较大,所以植物的WUE 提高.降水集中在7、8月,9月h s 最低,则在整个生长季9月的干旱程度相对较高.我们的研究结果表明,柽柳具有较强的耐旱能力,能够通过G s 的调节提高WUE 值实现对水分的高效利用来适应干旱的环境.这与以往研究发现干旱地区植物有较高的水分利用效率,能通过提高水分利用效率来适应水分亏缺的生长环境一致[21 -24 ] .可见,进行植物气孔特征及其对植物水分利用的影响研究,能够预测气候变化下气孔导度调节水分利用的动态变化,对于区域水资源管理具有重要的指导作用. ...
胡杨叶片气孔导度特征及其对环境因子的响应
2
2008
... 本研究采用LI-6400光合作用测定系统,对生长季期间的柽柳气体交换参数进行测定,观测柽柳叶片G s 的日变化和月变化特征.结果显示在整个生长季G s 日变化呈现先增加后减少的周期性波动,G s 月变化整体呈现逐渐减小的趋势.先前研究表明,强光照引起的高温会导致叶片蒸腾速率加快,蒸腾作用过强使植物体内出现水分亏缺,会引起保卫细胞失水和气孔关闭,对气孔导度产生抑制作用[17 -18 ] .自观测始点06:00开始,受PAR 和T a 迅速增加的影响,气孔开合增大,G s 迅速上升;6、7月午间T a 值较高(均值大于30 ℃),气孔关闭致使14:00 G s 值减小.这一现象与司建华等[19 ] 对黑河下游胡杨日变化研究结果一致.在8、9月午间柽柳G s 值在14:00左右达到最大值,而高冠龙等[15 ] 对黑河下游柽柳生长季G s 的研究表明其在10:00—12:00达到最大值,这与我们的研究结论并不相同,形成差异的原因可能在于研究期间环境条件的差异,即本研究中各月T a 差异明显,而黑河下游各月T a 变化基本不大. ...
... 研究发现环境因子h s 、PAR 、T a 与G s 的相关性较强,对柽柳G s 的影响较大,与G s 的相关性为h s (0.662)>PAR (0.613)>T a (0.601),C a 和VPD 对柽柳G s 的影响相对较小.司建华等[19 ] 发现黑河下游胡杨G s 与T a 、h s 有显著的相关关系,这与我们的研究结果一致,有学者认为C a 和VPD 是苹果树G s 主要影响因子[20 ] ,这与我们的研究结果形成差异,可能与研究区地理环境和物种有关.在与环境因子的响应关系中,叶片G s 和PAR 、h s 、T a 正相关,G s 随着PAR 、h s 、T a 这些影响因子的增加而增大,但超过影响因子阈值后G s 开始下降.本研究中G s 在T a 达到30 ℃后出现光合抑制现象从而G s 降低,我们认为影响柽柳G s 变化的T a 临界阈值可能是30 ℃左右. ...
黄土高原苹果树叶片气孔导度的环境响应与模拟
1
2021
... 研究发现环境因子h s 、PAR 、T a 与G s 的相关性较强,对柽柳G s 的影响较大,与G s 的相关性为h s (0.662)>PAR (0.613)>T a (0.601),C a 和VPD 对柽柳G s 的影响相对较小.司建华等[19 ] 发现黑河下游胡杨G s 与T a 、h s 有显著的相关关系,这与我们的研究结果一致,有学者认为C a 和VPD 是苹果树G s 主要影响因子[20 ] ,这与我们的研究结果形成差异,可能与研究区地理环境和物种有关.在与环境因子的响应关系中,叶片G s 和PAR 、h s 、T a 正相关,G s 随着PAR 、h s 、T a 这些影响因子的增加而增大,但超过影响因子阈值后G s 开始下降.本研究中G s 在T a 达到30 ℃后出现光合抑制现象从而G s 降低,我们认为影响柽柳G s 变化的T a 临界阈值可能是30 ℃左右. ...
水盐梯度下湿地柽柳(Tamarix ramosissima )光合响应特征和水分利用效率
2
2018
... G s 变化特征对植物光合作用、蒸腾作用等光合生理特性产生影响,进而能够影响植物的水分利用效率.水分利用效率是植物在单位水分消耗条件下进行碳固定的能力,能够反映植物有效的水分利用能力,也是衡量植物耐旱性和环境适应性的重要指标[21 ] .生长旺季(6—8月)柽柳WUE 值日变化为先降低再升高.生长季末期(9月)WUE 日变化为先升高再降低.当G s 降低时,气孔关闭至几乎闭合时,水分可以继续通过角质层散失,所以蒸腾的降低比CO2 流入的降低程度小,使WUE 值降低.当G s 升高时,气孔张开至完全开放时,叶片内部存在扩散阻力,扩散阻力对CO2 流入的限制比对蒸腾的限制程度大,使WUE 值降低[18 ] .可见,G s 升高或者降低都会引起WUE 的减小,只有当G s 变化使得水消耗和CO2 吸收实现最优协调时,WUE 才能达到最大值,因此WUE 出现波动性变化.研究发现,9月柽柳WUE 值相对较高,这是因为9月G s 整体较低,气孔部分关闭使得通过气孔进入叶片参与碳固定CO2 减少,从而导致光合速率降低,另一方面使得通过气孔蒸腾的水分减少,且蒸腾速率降低的幅度较大,所以植物的WUE 提高.降水集中在7、8月,9月h s 最低,则在整个生长季9月的干旱程度相对较高.我们的研究结果表明,柽柳具有较强的耐旱能力,能够通过G s 的调节提高WUE 值实现对水分的高效利用来适应干旱的环境.这与以往研究发现干旱地区植物有较高的水分利用效率,能通过提高水分利用效率来适应水分亏缺的生长环境一致[21 -24 ] .可见,进行植物气孔特征及其对植物水分利用的影响研究,能够预测气候变化下气孔导度调节水分利用的动态变化,对于区域水资源管理具有重要的指导作用. ...
... [21 -24 ].可见,进行植物气孔特征及其对植物水分利用的影响研究,能够预测气候变化下气孔导度调节水分利用的动态变化,对于区域水资源管理具有重要的指导作用. ...
塔克拉玛干沙漠南缘防护林和自然群落中多枝柽柳的光合特征及水分利用效率
0
2019
黑河中游荒漠绿洲过渡带多枝柽柳对地下水位变化的生理生态响应与适应
0
2011
黑河中游边缘绿洲不同水分条件对青贮玉米叶片光合特性及产量的影响
1
2009
... G s 变化特征对植物光合作用、蒸腾作用等光合生理特性产生影响,进而能够影响植物的水分利用效率.水分利用效率是植物在单位水分消耗条件下进行碳固定的能力,能够反映植物有效的水分利用能力,也是衡量植物耐旱性和环境适应性的重要指标[21 ] .生长旺季(6—8月)柽柳WUE 值日变化为先降低再升高.生长季末期(9月)WUE 日变化为先升高再降低.当G s 降低时,气孔关闭至几乎闭合时,水分可以继续通过角质层散失,所以蒸腾的降低比CO2 流入的降低程度小,使WUE 值降低.当G s 升高时,气孔张开至完全开放时,叶片内部存在扩散阻力,扩散阻力对CO2 流入的限制比对蒸腾的限制程度大,使WUE 值降低[18 ] .可见,G s 升高或者降低都会引起WUE 的减小,只有当G s 变化使得水消耗和CO2 吸收实现最优协调时,WUE 才能达到最大值,因此WUE 出现波动性变化.研究发现,9月柽柳WUE 值相对较高,这是因为9月G s 整体较低,气孔部分关闭使得通过气孔进入叶片参与碳固定CO2 减少,从而导致光合速率降低,另一方面使得通过气孔蒸腾的水分减少,且蒸腾速率降低的幅度较大,所以植物的WUE 提高.降水集中在7、8月,9月h s 最低,则在整个生长季9月的干旱程度相对较高.我们的研究结果表明,柽柳具有较强的耐旱能力,能够通过G s 的调节提高WUE 值实现对水分的高效利用来适应干旱的环境.这与以往研究发现干旱地区植物有较高的水分利用效率,能通过提高水分利用效率来适应水分亏缺的生长环境一致[21 -24 ] .可见,进行植物气孔特征及其对植物水分利用的影响研究,能够预测气候变化下气孔导度调节水分利用的动态变化,对于区域水资源管理具有重要的指导作用. ...
1971-2011年汾河流域降水时空特征
1
2018
... 目前关于气孔导度的模拟研究,大多采用的是经验模型和半经验模型,本文运用Jarvis模型(经验模型)、BWB模型(半经验模型)和USO模型(机理模型)对柽柳气孔导度进行模拟,结果表明Jarvis模型比其他两类模型的模拟精度高.有学者对黑河下游胡杨和柽柳叶片气孔导度基于(半)经验模型模拟研究,同样得到Jarvis模型模拟精度较高的结论,研究认为在荒漠干旱地区Jarvis模型具有相对较好的适用性[15 ,17 ] .本研究区域地处黄土高原,该区域干旱气候具有频次高、持续时间长且影响范围大的特点[25 ] ,也属于相对干旱的地区.因此,我们认为与半经验模型和气孔导度机理模型相比较而言,Jarvis模型对于干旱区的乔木、灌木等植物叶片气孔导度模拟效果较好.此外,Jarvis模型进行气孔导度模拟时,只需考虑单一环境因子作为气孔行为的影响因素,因此在现有的植物叶片气孔导度模拟中Jarvis模型得到广泛应用[26 -30 ] .Jarvis模型为完全经验模型,在干旱半干旱地区,需要进一步探讨其对于不同植物种类叶片气孔导度模拟方面的适用性.值得一提的是,气孔导度机理模型用于研究不同环境和生物因素对气孔行为的影响,不仅考虑了叶片生理特性和气孔导度之间的关系,而且具有反馈机制,未来可以考虑将其和植被模型、生态系统模型等进行耦合,从而提高该模型模拟及预测的准确性.在干旱半干旱地区对叶片尺度气孔行为和影响因素进行深入研究,以及进行叶片气孔导度的模拟,不仅能够进行尺度扩展用于生态系统蒸散发估算和生态系统建模[31 -32 ] ,同时对于理解该区域植被和生态系统对环境的适应机制具有重要意义. ...
Testing two models for the estimation of leaf stomatal conductance in four greenhouse crops cucumber,chrysanthemum,tulip and lilium
1
2012
... 目前关于气孔导度的模拟研究,大多采用的是经验模型和半经验模型,本文运用Jarvis模型(经验模型)、BWB模型(半经验模型)和USO模型(机理模型)对柽柳气孔导度进行模拟,结果表明Jarvis模型比其他两类模型的模拟精度高.有学者对黑河下游胡杨和柽柳叶片气孔导度基于(半)经验模型模拟研究,同样得到Jarvis模型模拟精度较高的结论,研究认为在荒漠干旱地区Jarvis模型具有相对较好的适用性[15 ,17 ] .本研究区域地处黄土高原,该区域干旱气候具有频次高、持续时间长且影响范围大的特点[25 ] ,也属于相对干旱的地区.因此,我们认为与半经验模型和气孔导度机理模型相比较而言,Jarvis模型对于干旱区的乔木、灌木等植物叶片气孔导度模拟效果较好.此外,Jarvis模型进行气孔导度模拟时,只需考虑单一环境因子作为气孔行为的影响因素,因此在现有的植物叶片气孔导度模拟中Jarvis模型得到广泛应用[26 -30 ] .Jarvis模型为完全经验模型,在干旱半干旱地区,需要进一步探讨其对于不同植物种类叶片气孔导度模拟方面的适用性.值得一提的是,气孔导度机理模型用于研究不同环境和生物因素对气孔行为的影响,不仅考虑了叶片生理特性和气孔导度之间的关系,而且具有反馈机制,未来可以考虑将其和植被模型、生态系统模型等进行耦合,从而提高该模型模拟及预测的准确性.在干旱半干旱地区对叶片尺度气孔行为和影响因素进行深入研究,以及进行叶片气孔导度的模拟,不仅能够进行尺度扩展用于生态系统蒸散发估算和生态系统建模[31 -32 ] ,同时对于理解该区域植被和生态系统对环境的适应机制具有重要意义. ...
春玉米持续干旱过程中常用气孔导度模型的比较研究
0
2018
桂花树冠层气孔导度模型的优化及其参数分析
0
2016
Carbon and water flux responses to physiology by environment interactions:a sensitivity analysis of variation in climate on photosynthetic and stomatal parameters
1
2014
... 目前关于气孔导度的模拟研究,大多采用的是经验模型和半经验模型,本文运用Jarvis模型(经验模型)、BWB模型(半经验模型)和USO模型(机理模型)对柽柳气孔导度进行模拟,结果表明Jarvis模型比其他两类模型的模拟精度高.有学者对黑河下游胡杨和柽柳叶片气孔导度基于(半)经验模型模拟研究,同样得到Jarvis模型模拟精度较高的结论,研究认为在荒漠干旱地区Jarvis模型具有相对较好的适用性[15 ,17 ] .本研究区域地处黄土高原,该区域干旱气候具有频次高、持续时间长且影响范围大的特点[25 ] ,也属于相对干旱的地区.因此,我们认为与半经验模型和气孔导度机理模型相比较而言,Jarvis模型对于干旱区的乔木、灌木等植物叶片气孔导度模拟效果较好.此外,Jarvis模型进行气孔导度模拟时,只需考虑单一环境因子作为气孔行为的影响因素,因此在现有的植物叶片气孔导度模拟中Jarvis模型得到广泛应用[26 -30 ] .Jarvis模型为完全经验模型,在干旱半干旱地区,需要进一步探讨其对于不同植物种类叶片气孔导度模拟方面的适用性.值得一提的是,气孔导度机理模型用于研究不同环境和生物因素对气孔行为的影响,不仅考虑了叶片生理特性和气孔导度之间的关系,而且具有反馈机制,未来可以考虑将其和植被模型、生态系统模型等进行耦合,从而提高该模型模拟及预测的准确性.在干旱半干旱地区对叶片尺度气孔行为和影响因素进行深入研究,以及进行叶片气孔导度的模拟,不仅能够进行尺度扩展用于生态系统蒸散发估算和生态系统建模[31 -32 ] ,同时对于理解该区域植被和生态系统对环境的适应机制具有重要意义. ...
植物气孔导度的环境响应模拟及其尺度扩展
1
2016
... 目前关于气孔导度的模拟研究,大多采用的是经验模型和半经验模型,本文运用Jarvis模型(经验模型)、BWB模型(半经验模型)和USO模型(机理模型)对柽柳气孔导度进行模拟,结果表明Jarvis模型比其他两类模型的模拟精度高.有学者对黑河下游胡杨和柽柳叶片气孔导度基于(半)经验模型模拟研究,同样得到Jarvis模型模拟精度较高的结论,研究认为在荒漠干旱地区Jarvis模型具有相对较好的适用性[15 ,17 ] .本研究区域地处黄土高原,该区域干旱气候具有频次高、持续时间长且影响范围大的特点[25 ] ,也属于相对干旱的地区.因此,我们认为与半经验模型和气孔导度机理模型相比较而言,Jarvis模型对于干旱区的乔木、灌木等植物叶片气孔导度模拟效果较好.此外,Jarvis模型进行气孔导度模拟时,只需考虑单一环境因子作为气孔行为的影响因素,因此在现有的植物叶片气孔导度模拟中Jarvis模型得到广泛应用[26 -30 ] .Jarvis模型为完全经验模型,在干旱半干旱地区,需要进一步探讨其对于不同植物种类叶片气孔导度模拟方面的适用性.值得一提的是,气孔导度机理模型用于研究不同环境和生物因素对气孔行为的影响,不仅考虑了叶片生理特性和气孔导度之间的关系,而且具有反馈机制,未来可以考虑将其和植被模型、生态系统模型等进行耦合,从而提高该模型模拟及预测的准确性.在干旱半干旱地区对叶片尺度气孔行为和影响因素进行深入研究,以及进行叶片气孔导度的模拟,不仅能够进行尺度扩展用于生态系统蒸散发估算和生态系统建模[31 -32 ] ,同时对于理解该区域植被和生态系统对环境的适应机制具有重要意义. ...
地下变水位条件下塔里木河下游河岸胡杨林蒸腾模型
1
2014
... 目前关于气孔导度的模拟研究,大多采用的是经验模型和半经验模型,本文运用Jarvis模型(经验模型)、BWB模型(半经验模型)和USO模型(机理模型)对柽柳气孔导度进行模拟,结果表明Jarvis模型比其他两类模型的模拟精度高.有学者对黑河下游胡杨和柽柳叶片气孔导度基于(半)经验模型模拟研究,同样得到Jarvis模型模拟精度较高的结论,研究认为在荒漠干旱地区Jarvis模型具有相对较好的适用性[15 ,17 ] .本研究区域地处黄土高原,该区域干旱气候具有频次高、持续时间长且影响范围大的特点[25 ] ,也属于相对干旱的地区.因此,我们认为与半经验模型和气孔导度机理模型相比较而言,Jarvis模型对于干旱区的乔木、灌木等植物叶片气孔导度模拟效果较好.此外,Jarvis模型进行气孔导度模拟时,只需考虑单一环境因子作为气孔行为的影响因素,因此在现有的植物叶片气孔导度模拟中Jarvis模型得到广泛应用[26 -30 ] .Jarvis模型为完全经验模型,在干旱半干旱地区,需要进一步探讨其对于不同植物种类叶片气孔导度模拟方面的适用性.值得一提的是,气孔导度机理模型用于研究不同环境和生物因素对气孔行为的影响,不仅考虑了叶片生理特性和气孔导度之间的关系,而且具有反馈机制,未来可以考虑将其和植被模型、生态系统模型等进行耦合,从而提高该模型模拟及预测的准确性.在干旱半干旱地区对叶片尺度气孔行为和影响因素进行深入研究,以及进行叶片气孔导度的模拟,不仅能够进行尺度扩展用于生态系统蒸散发估算和生态系统建模[31 -32 ] ,同时对于理解该区域植被和生态系统对环境的适应机制具有重要意义. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}