


0 引言
植被分布格局是树木对气候长期适应的结果,气候变化会影响树木的生长,并导致其适宜分布区发生改变[10]。因此,研究树木生长的适宜气候条件对生态系统的稳定具有重要意义。目前,已有众多学者针对气候变化对油松分布区的影响进行了广泛研究[11-15]。徐文铎等[12]研究发现降水与气温是影响中国油松生长的主要气候因素,且年降水量500~600 mm、年均气温5~7 ℃的区域为油松天然分布区。Li等[13]利用MaxEnt模型预测表明,年降水量431~1 122 mm、年均气温2.9~14.7 ℃,为油松的适宜分布区,且在气候变暖的背景下,未来油松人工林的分布边界存在向中国东北和西南移动的趋势。Wang等[14]和Chen等[15]对西北干旱半旱区油松径向生长与气候因子关系的研究结果表明,高海拔地区充足的降水与适宜的气温可能更有助于油松的径向生长。这些研究极大地提高了气候变化对油松分布区影响的认识。树木生长不仅受到降水、气温等气候因素的影响,还受到海拔、坡向等地形因素的影响。同时,油松作为中国干旱半干旱区主要的造林树种,也被广泛种植,其造林面积已达到天然林面积的2倍以上[16-17];然而,由于在引种过程中未充分考虑气候变化、海拔差异等因素对油松径向生长的影响,部分区域油松生长出现衰退或死亡等现象[18]。因此,迫切需要从不同角度分析油松径向生长对气候变化的响应特征,探讨油松适宜的分布区,为油松天然林和人工林管理提供理论依据。
荟萃分析(Meta-analysis)是通过对若干独立研究的结果进行统计分析,最终得到目标事物综合定量结论的分析方法[19]。本文选择了53个采样点,通过Meta分析方法,分析了中国北方地区油松径向生长与气候响应的区域分异特征,以期深入理解气候变暖背景下不同气候带油松径向生长及其适宜分布区的变化趋势。
1 树种与研究区域
1.1 油松生理特征
1.2 油松天然林和人工林主要分布区域
图1
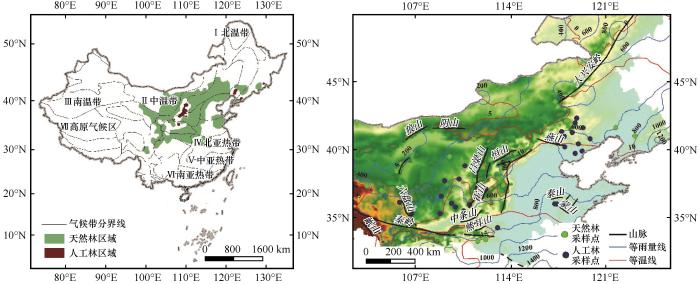
图1
研究区油松天然林与人工林采样点分布区
Fig.1
Main distribution and tree-ring sampling points of the natural and planted Pinus tabulaeformis forests in China
2 研究内容与方法
2.1 数据来源
本研究检索了3个电子数据库( Pubmed、Web of Science和中国知网)。搜索关键词包含:“油松”(Pinus tabuliformis)、“径向生长”(Radial growth)、 “气候响应”(Climate response)、“天然林”(natural forests)和“人工林”(plantation forests)。详细检索的方法如表1所列(以Pubmed为例)
表1 PubMed搜索策略
Table 1
| #1 | (Pinus tabuliformis [Title/Abstract]) OR (Chinese pine [Title/Abstract]) |
|---|---|
| #2 | (plantation[Title/Abstract]) OR (natural forests[Title/Abstract]) |
| #3 | (#1) AND (#2) |
| #4 | ((((Precipitation[Title/Abstract]) OR (Precipitation[Title/Abstract])) OR (PDSI[Title/Abstract])) OR (scPDSI[Title/Abstract])) OR (SPEI[Title/Abstract]) |
| #5 | (radial growth) OR (climate response) |
| #6 | (#4) AND (#5) |
| #7 | (#3) AND (#6) |
筛选出2000年1月至2022年12月符合以下基本要求的文献:①油松径向生长与气候因子的相关性分析中,至少包含对单月气候因子的相关性分析;②油松径向生长与气候因子的相关性分析中,选择分析的气候因子(降水、气温、PDSI、scPDSI和SPEI)能够基本涵盖油松整个分布区。
基于以上标准,从23篇文献(21篇天然林,2篇人工林)中选取了53个采样点(天然林34个,人工林19个),具体见图1。
2.2 数据整理
对每篇文献中的以下内容进行了统计:①每个采样点油松树芯的数量。②油松天然林径向生长与单月降水量、平均气温以及帕默尔干旱指数(PDSI)的相关系数。③油松人工林径向生长与PDSI的相关系数。以文字和表格形式的数据直接提取,柱形图数据用 Getdata Graph Digitizer 软件获取;热图数据则用Snipaste1.16软件提取。
2.3 数据分组
2.4 数据分析
本文采用合并相关系数(总体平均效应值R)来描述气候因子对油松径向生长的影响。
式中:r指单样点树木径向生长与气候因子间相关系数;n指样本量。Fisher's Z为转换后的相关系数;Vz 为Fisher's Z的方差;SE 为Fisher's Z的标准差;Wi 为每项独立研究的权重;R为总体平均效应值。
采用R语言中“MetaCor”包,进行Fisher's Z转换,获取Z值和SE 值,用Stata16.0软件进行合并相关系数的计算。若合并相关系数(R)的95%置信区间与0重叠,则表示油松生长与气候因子的相关性未达到显著水平;反之则达到显著水平。
3 结果
3.1 油松天然林与人工林对气候因子的响应特征
油松天然林径向生长主要与降水呈正相关,与气温呈负相关关系;其中上年9—10月和当年2—7月降水量,上年9月、当年1—3月和5—8月平均气温均达到显著相关水平(图2A、B)。
图2
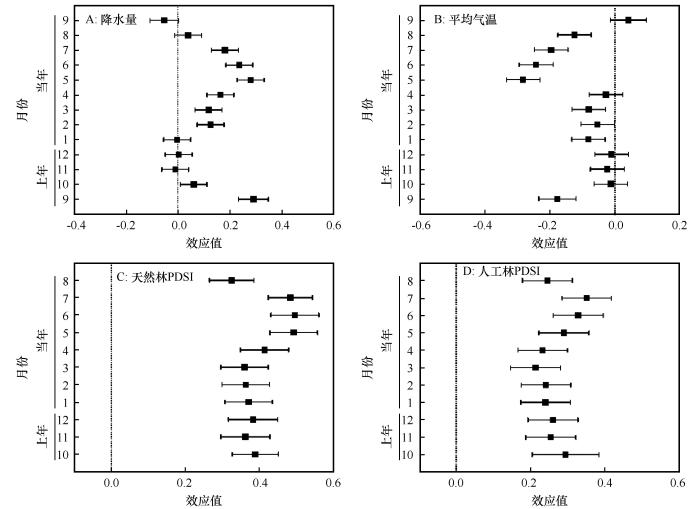
图2
油松天然林(A~C)与人工林(D)径向生长与气候因子的效应值
Fig.2
Effect size (R) between the growth of Pinus tabulaeformis in natural (A-C) and plantation(D) forests and climate factors
同时,无论天然林还是人工林,油松径向生长与上年10月至当年8月PDSI均呈正相关,且均达到显著水平。这说明中国整个北方地区的油松径向生长均受干旱胁迫的影响(图2C、D)。
3.2 油松径向生长对降水与气温的响应特征
基于图2的分析结果,选择了上年9月、当年3月和5—7月的降水量,以及当年2月和5—7月的平均气温,分析降水与气温梯度下油松天然林径向生长对上述气候因子的响应。
图3
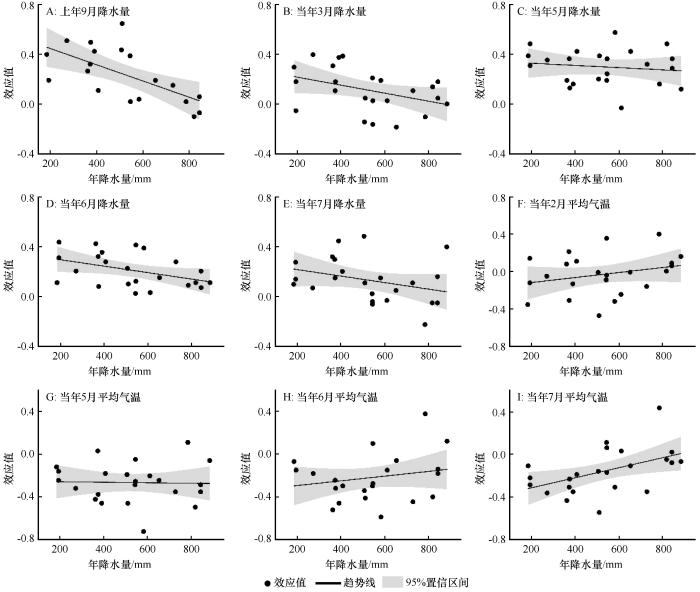
图3
降水梯度下油松径向生长与气候因子之间的效应值
Fig.3
Effect size (R) between radial growth of Pinus tabulaeformis and climatic factors under precipitation gradients
表2 降水梯度下油松径向生长与气候因子效应值及统计参数
Table 2
| 气候因子 | 月份 | 效应值的系数 | 标准差 | t | P>|t| |
|---|---|---|---|---|---|
| 降水 | 上年9月 | -0.0006 | 0.0002 | -2.93 | 0.009 |
| 当年3月 | -0.0003 | 0.0001 | -1.64 | 0.116 | |
| 当年5月 | -0.0001 | 0.0002 | -0.68 | 0.505 | |
| 当年6月 | -0.0003 | 0.0001 | -1.95 | 0.064 | |
| 当年7月 | -0.0003 | 0.0002 | -1.45 | 0.161 | |
| 气温 | 当年2月 | 0.0003 | 0.0002 | 1.21 | 0.240 |
| 当年5月 | 8.39×10-6 | 0.0002 | 0.04 | 0.965 | |
| 当年6月 | 0.0002 | 0.0002 | 1.06 | 0.300 | |
| 当年7月 | 0.0005 | 0.0950 | -4.33 | 0.000 |
图4
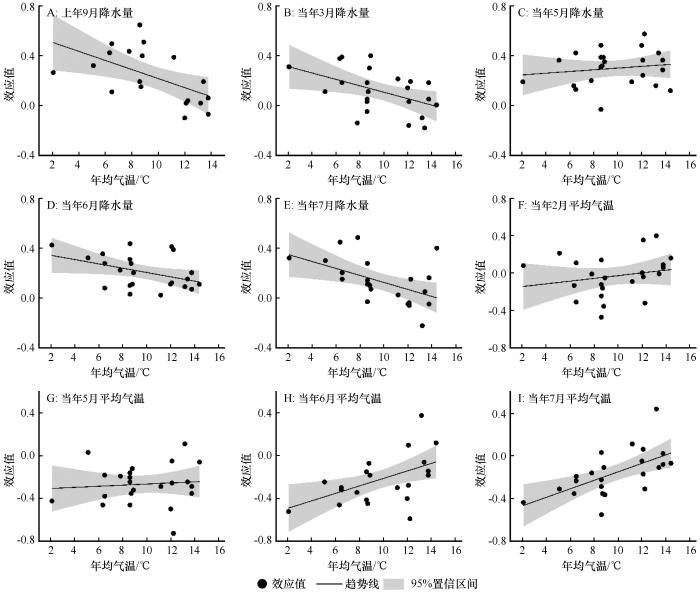
图4
气温梯度下油松径向生长与气候因子之间的效应值
Fig.4
Effect size (R) between radial growth of Pinus tabulaeformis and climatic factors under temperature gradients
表3 气温梯度下油松径向生长与气候因子之间的效应值及统计参数
Table 3
| 气候因子 | 月份 | 效应值的系数 | 标准差 | t | P>|t| |
|---|---|---|---|---|---|
| 降水 | 上年9月 | -0.035 | 0.012 | -2.82 | 0.011 |
| 当年3月 | -0.025 | 0.010 | -0.29 | 0.032 | |
| 当年5月 | 0.01 | 0.01 | 1.03 | 0.315 | |
| 当年6月 | -0.015 | 0.01 | -1.50 | 0.147 | |
| 当年7月 | -0.028 | 0.01 | -2.54 | 0.018 | |
| 气温 | 当年2月 | 0.015 | 0.015 | 1.04 | 0.312 |
| 当年5月 | 0.009 | 0.012 | 0.72 | 0.478 | |
| 当年6月 | 0.037 | 0.012 | 2.97 | 0.007 | |
| 当年7月 | 0.039 | 0.01 | 3.73 | 0.001 |
分组分析结果表明,油松径向生长与上年9月降水量的效应值在<400 mm和400~600 mm年降水量组,以及<7 ℃和7~9 ℃年均气温组的区域达到显著水平,效应值分别为0.370、0.319、0.393和0.356;上年9月降水量对年降水量> 600 mm 和年均气温> 9 ℃区域油松的生长没有显著影响(图5)。油松径向生长与当年3月降水量的效应值在年降水量< 400 mm以及年均气温< 7 ℃的区域达到显著水平,效应值分别为0.200和0.263;与年降水量在400~600 mm和> 600 mm 以及年均气温在7~9 ℃和>9 ℃的区域未达到显著水平。油松径向生长与5月降水量的效应值在不同的降水区域分别为0.272、0.328和0.242,在不同的气温区域分别为0.19、0.319和0.319,且均达到显著水平。油松径向生长与6月降水量的效应值在不同的降水区域分别为0.280、0.244和0.127,在不同的气温区域分别为0.251、0.255和0.194,均达到显著水平。油松径向生长与7月降水量的效应值在年降水量< 400 mm和400~600 mm以及年均气温<7 ℃和7~9 ℃的区域达到显著水平,而在年降水量> 600 mm和年均气温> 9 ℃的区域未达到显著水平(图5)。
图5
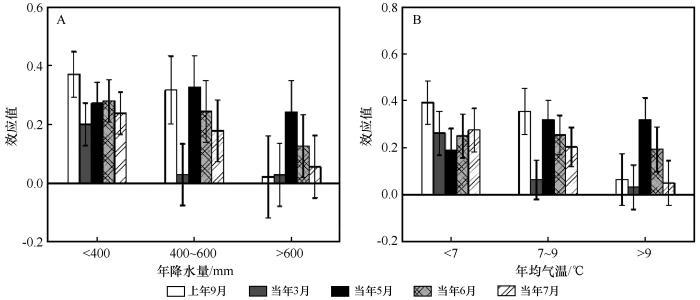
图5
降水(A)和气温(B)梯度下油松径向生长与降水之间的效应值
Fig.5
Effect size (R) between radial growth of Pinus tabulaeformis and precipitation under precipitation (A) and temperature (B) gradients
油松径向生长与当年2月平均气温的效应值在年降水量<400 mm以及年均气温<7 ℃和7~9 ℃的区域达到显著水平,效应值分别为-0.089、-0.114和-0.099;在年降水量400~600 mm和> 600 mm以及年均气温> 9 ℃的区域未达到显著水平(图6)。油松径向生长与当年5月平均气温的效应值在不同的气温与降水区域均达到显著水平。油松径向生长与当年6月平均气温在年降水量< 400 mm和400~600 mm以及年均气温<7 ℃、7~9 ℃ 和> 9 ℃的区域达到显著水平,效应值分别为-0.285、-0.308、-0.363、-0.252和-0.121;而在年降水量>600 mm的区域未达到显著水平。油松径向生长与当年7月平均气温的效应值在年降水量<400 mm和400~600 mm以及年均气温< 7 ℃和7~9 ℃的区域达到显著水平,在年降水量> 600 mm和年均气温>9 ℃的区域未达到显著水平(图6)。
图6
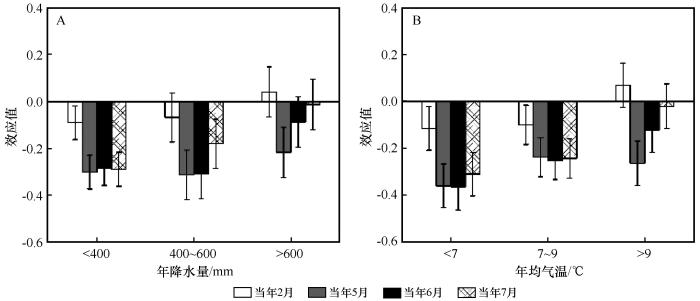
图6
降水(A)和气温(B)梯度下油松径向生长与气温之间的效应值
Fig.6
Effect size (R) between radial growth of Pinus tabulaeformis and temperature under precipitation (A) and temperature (B) gradients
上述结果表明,上一年降水对下一年油松径向生长的促进作用主要体现在年降水量<600 mm和年均气温<9 ℃的区域,5月的降水量和平均气温影响整个北方地区油松的生长,7月的降水量和平均气温则主要影响年降水量<600 mm和年均气温<9 ℃区域油松的生长。此外,2月平均气温对年降水量>600 mm和年均气温>9 ℃区域油松的生长起到促进作用,对年降水量<600 mm和年均气温<9 ℃区域油松的生长起到抑制作用。
3.3 降水与气温梯度下油松天然林与人工林径向生长对干旱的响应特征
基于图2的分析结果,我们选择当年5月、6月、7月(生长季5—7月)的PDSI进一步分析降水与气温梯度下,油松天然林与人工林生长对干旱胁迫响应的差异。
图7
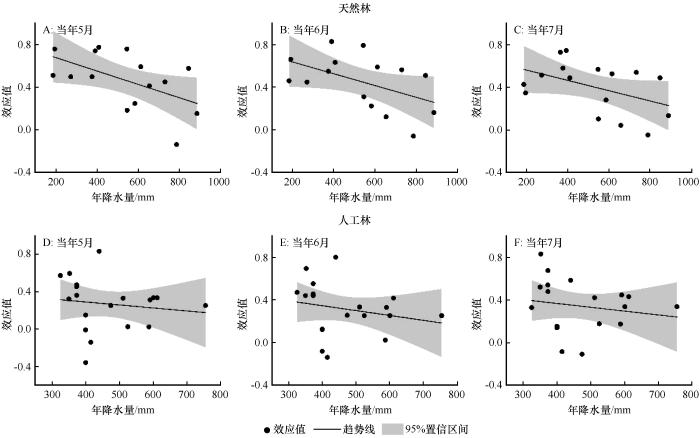
图7
降水梯度下油松天然林(A~C)和人工林(D~F)径向生长与PDSI的效应值
Fig.7
Effect size (R) between radial growth of Pinus tabulaeformis in natural (A-C) and plantation (D-F) forests and PDSI under precipitation gradients
图8
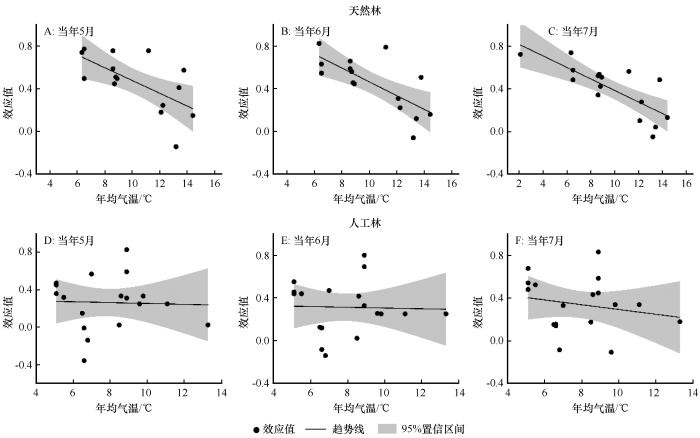
图8
气温梯度下油松天然林(A~C)和人工林(D~F)径向生长与PDSI的效应值
Fig.8
Effect size (R) between radial growth of Pinus tabulaeformis in natural (A-C) and plantation (D-F) forests and PDSI under temperature gradients
表4 降水梯度下油松径向生长与PDSI之间的效应值及统计参数
Table 4
| 林分 来源 | 月份 | 效应值的 系数 | 标准差 | t | P>|t| |
|---|---|---|---|---|---|
| 天然林 | 当年5月 | -0.0006 | 0.0003 | -2.43 | 0.028 |
| 当年6月 | -0.0006 | 0.0003 | -2.18 | 0.046 | |
| 当年7月 | -0.0005 | 0.0003 | -1.54 | 0.145 | |
| 人工林 | 当年5月 | -0.0004 | 0.0005 | -0.69 | 0.500 |
| 当年6月 | -0.0005 | 0.0005 | -1.06 | 0.306 | |
| 当年7月 | -0.0004 | 0.0005 | -0.77 | 0.453 |
表5 气温梯度下油松径向生长与PDSI之间的效应值及统计参数
Table 5
| 林分 来源 | 月份 | 效应值的 系数 | 标准差 | t | P>|t| |
|---|---|---|---|---|---|
| 天然林 | 当年5月 | -0.06 | 0.020 | -3.03 | 0.008 |
| 当年6月 | -0.06 | 0.020 | -3.98 | 0.001 | |
| 当年7月 | -0.06 | 0.016 | -3.55 | 0.003 | |
| 人工林 | 当年5月 | -0.007 | 0.029 | -0.23 | 0.069 |
| 当年6月 | -0.005 | 0.026 | -0.20 | 0.845 | |
| 当年7月 | -0.023 | 0.026 | -0.86 | 0.403 |
分组分析结果表明,油松天然林径向生长与当年5月PDSI的效应值在年降水量<400 mm、400~600 mm和>600 mm的区域分别为0.613、0.402和0.341,在年均气温<7 ℃、7~9 ℃和>9 ℃的区域分别为0.608、0.608和0.290,且均达到显著水平;与当年6月PDSI在不同的降水区域分别为0.623、0.422和0.320,在不同的气温区域分别为0.656、0.572和0.282,且均达到显著水平;与当年7月PDSI的效应值在不同的降水区域分别为0.572、0.498和0.284,在不同的气温区域分别为0.649、0.553和0.215,也且均达到显著水平。
油松人工林径向生长与当年5月PDSI的效应值在年降水量<400 mm和400~600 mm的区域分别为0.462和0.175,在年均气温< 7 ℃和7~9 ℃的区域分别为0.194和0.43,且均达到显著水平;与当年6月PDSI的效应值在不同的降水区域分别为0.507和0.221,在不同的气温区域分别为0.269和0.433,且均达到显著水平;与当年7月PDSI的效应值在不同的降水区域分别为0.562和0.211,在不同的气温区域分别为0.263和0.439,也均达到显著水平(图9~10)。由于年降水量>600 mm和年均气温>9 ℃区域的样本量太少,本文不做进一步分析。这说明中国北方地区油松的生长受干旱胁迫的影响显著。
图9
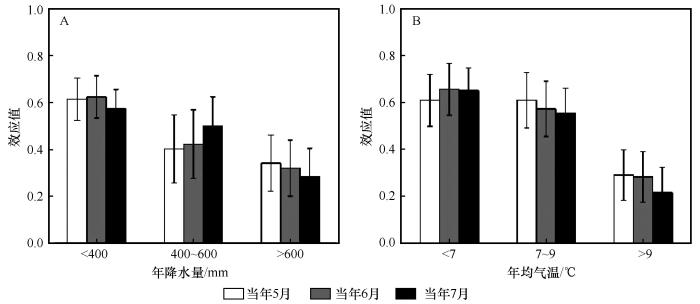
图9
降水(A)和气温(B)梯度下油松天然林径向生长与PDSI间的效应值
Fig.9
Effect size (R) between radial growth of natural Pinus tabulaeformis and PDSI under precipitation (A) and temperature (B) gradients
图10
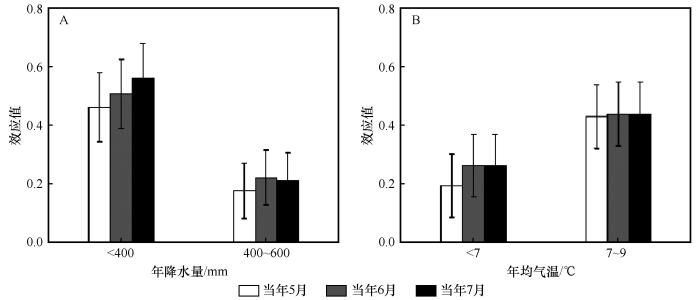
图10
降水(A)和气温(B)梯度下油松人工林径向生长与PDSI间的效应值
Fig.10
Effect size (R) between radial growth of planted Pinus tabulaeformis and PDSI under precipitation (A) and temperature (B) gradients
4 讨论
4.1 气候变暖背景下的油松径向生长对气候响应的区域分异
大尺度区域研究发现,600 mm降水量等值线是中国树木生长对气候响应的重要边界:这条线以北,树木生长主要与降水呈正相关;这条线以南,树木生长主要与气温负相关[23]。本文研究结果与大尺度的研究基本一致。然而,不同的树种生长与气候响应仍然具有部分异质性。图5和图6的结果表明,上年9月降水量对油松生长的促进作用体现在年降水量<600 mm和年均气温<9 ℃的区域。主要原因为上年9月降水充沛,有效提高了有机物质的积累与土壤水分的补给,为下一年树木形成层的活动提供充足的营养物质[1];而随着降水和气温的升高,树木形成层活动期延长,过多的降水反而会使温度的降低加快。尤其对于冬季,不仅降低了上一年降水对下一年油松生长的促进作用,甚至会抑制油松的生长[24]。
5月的降水量和平均气温对整个北方地区油松生长的影响具有一致性(图3~6)。主要原因为从5月开始,树木进入耗水高峰期和干物质快速增加期,树木生理代谢活动旺盛。较高的温度加快了树木的蒸腾作用与土壤水分的蒸发,导致树木根系无法获取光合作用所需的水分,干旱胁迫加剧,间接限制了树木的生长[29-30]。因此,充足的降水成为维持树木生长的关键因素。然而,由于中国北方地区降水主要受季风气候控制,从干旱半干旱区到半湿润区,5月雨季还未到来;此时有效的降水能够增加土壤含水量,补充生理活动所必需的水分,进而促进树木的生长[25]。此外,由图5和6的结果可知,7月平均气温显著影响年均气温<9 ℃区域油松的生长,对年均气温>9 ℃区域油松生长的影响较小,生长季前期低温会推迟油松的萌发与生长[18]。进入7月,油松依然处于木质部形成层活动的旺盛时期,较高的温度加快了土壤水分的散失,树木根系无法获得供给光合作用所需的水分,树木受到干旱胁迫的影响加剧,从而间接限制了油松的径向生长[30],这也与Mäkinen等[31]对欧洲Picea abies的研究结果基本一致。
图7和图8的结果还表明,北方地区油松天然林和人工林生长与生长季(5—7月)PDSI的相关性均达到显著水平。这说明中国北方地区油松的生长均易受生长季干旱胁迫的影响,尤其对干旱半干旱地区,生长季的干旱胁迫对油松生长的影响更高(图7~10)。这也与Camarero等[32]对P. lentiscus研究结果一致。相对于人工林,随着降水量与气温的不断增加,干旱胁迫对油松天然林生长影响降低的趋势更加显著(表4和表5)。部分研究认为,人工林由于进行了抚育经营,降低了对气候变化的敏感性。因此,与人工林相比,天然林对气候变化的响应更加敏感[33];也有部分研究表明,与天然林相比,人工林受到干旱胁迫后树木生长量的下降更加严重,导致其生长对干旱胁迫的敏感性更强[34]。研究表明,人工林的生长不仅受气候因素的影响,还受造林密度、间伐抚育等人工管理的影响[35-36]。本文中未考虑这些因素对人工林生长的影响,因而目前的结果是否具有代表性,还有待进一步研究。
4.2 气候变暖背景下中国北方地区油松生长状态与分布格局
本文研究结果表明,油松的生长对水热条件的响应随气温与降水的变化,呈现出明显的区域分异特征。即降水与气温较低时,干旱胁迫对油松生长的影响较高;随着降水与气温不断升高,干旱胁迫对油松生长的影响不断降低。尤其在秦岭南坡的木王山,油松径向生长与PDSI的相关性在部分月份呈负相关 (图7)。此外,研究发现从年降水量<400 mm的贺兰山和昌岭山到年降水量>500 mm的北京松山,气候因子对低海拔区域油松生长的影响高于高海拔区域[14-15,41],树木林线有不断上升趋势[42],气候变化对低海拔区域油松生长的影响将会更加显著,高海拔区域可能更适宜油松的径向生长。因此,水平尺度上,未来中国北方地区油松分布区面临从干旱半干旱区域向更加湿润区域移动的趋势;垂直尺度上,油松分布区面临从低海拔区域向更高海拔区域移动的趋势。
5 结论
在年降水量<400 mm的区域,限制油松径向生长的主要因素是生长季降水;在年降水量400~600 mm的区域,生长季的水热因素共同影响油松的径向生长;在年降水量>600 mm区域,油松的径向生长主要受5月降水与平均气温及其共同作用的干旱胁迫影响。此外,降水对油松径向生长的“滞后效应”(如上年9月的降水量)主要体现降水量<600 mm和气温<9 ℃区域油松的生长;冬季气温对不同气候区油松生长的影响具有差异:冬季气温对年降水量>600 mm和年均气温>9 ℃区域油松的生长起促进作用,对年降水量<600 mm和年均气温>9 ℃的区域起抑制作用。在全球气候变暖背景下,未来油松的分布格局随着气温的上升发生变化:水平尺度上,油松分布区面临从干旱半干旱区域向相对湿润区域移动的趋势;垂直尺度上,油松分布区面临从低海拔区域向高海拔区域移动的趋势。
参考文献
Evaluation of ecosystem services of Chinese pine forests in China
[J].
腾格里沙漠南缘青海云杉(Picea crassifolia)和油松(Pinus tabulaeformis)年轮记录的气候变化
[J].
Reconstruction of spring temperature on the southern edge of the Gobi Desert,Asia,reveals recent climatic warming
[J].
Tree-ring based reconstruction of drought variability (1615-2009) in the Kongtong Mountain area,northern China
[J].
Tree-ring based runoff reconstruction of the upper Fenhe River basin,North China,since 1799 AD
[J].
Reconstructed precipitation for the north-central China over the past 380 years and its linkages to East Asian summer monsoon variability
[J].
Geographical boundary and climatic analysis of Pinus tabulaeformis in China:insights on its afforestation
[J].
Divergent tree radial growth at alpine coniferous forest ecotone and corresponding responses to climate change in northwestern China
[J].
Evaluation of the response stability of two dominant conifer species to climate change in the southern margin of the Tengger Desert
[J].
Variations in the growth response of Pinus tabulaeformis to a warming climate at the northern limits of its natural range
[J].
Mixed-species plantations can alleviate water stress on the Loess Plateau
[J].
The 600-mm precipitation isoline distinguishes tree-ring-width responses to climate in China
[J].
Widespread increase of tree mortality rates in the western United States
[J].
Temperature variations recorded in Pinus tabulaeformis tree rings from the southern and northern slopes of the central Qinling Mountains,central China
[J].
A scPDSI-based global data set of dry and wet spells for 1901-2009
[J].
Rapid tree growth with respect to the last 400 years in response to climate warming,northeastern Tibetan Plateau
[J].
Climatic response of three tree species growing at different elevations in the Lüliang Mountains of northern China
[J].
Interannual variability of PDSI from tree-ring widths for the past 278 years in Baotou,China
[J].
Large-scale climatic variability and radial increment variation of Picea abies (L.) in central and northern Europe
[J].
Relating climate,drought and radial growth in broadleaf mediterranean tree and shrub species:a new approach to quantify climate-growth relationships
[J].
Different responses of the radial growth of the planted and natural forests to climate change in humid subtropical China
[J].
Conversion of natural forests to managed forest plantations decreases tree resistance to prolonged droughts
[J].
Growth response of plantations Hippophae rhamnoides Linn.on different slope aspects and natural Caragana opulens Kom.to climate and implications for plantations management
[J].
Radial growth of Korshinsk peashrub and its response to drought in different sub-arid climate regions of northwest China
[J].
A reconstruction of areasal and global temperature for the past 11,300 years
[J].
The origin of remnant forest stands of Pinus tabulaeformis in southeastern Inner Mongolia
[J].
Warming-induced upward migration of the alpine treeline in the Changbai Mountains,northeast China
[J].

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}