Growth rate-stoichiometry couplings in diverse biota
1
2003
... 土壤碳(C)、氮(N)、磷(P)是陆地生态系统最基本的营养元素,也是植物赖以生存的必需条件,进而影响着植物功能性状、功能多样性与生态系统功能.生态化学计量学以元素组成来揭示生物化学和生态学之间的联系,可以更加深入探究生物之间元素循环的内在机理以及生态系统组分之间的联系[1-4].因此,研究土壤系统C、N、P含量及其生态化学计量特征规律,可以认识C、N、P在生态系统中的循环、平衡机制及其对植物群落结构和功能的影响.土壤微生物量是指土壤中体积小于5.0×103 μm3活的微生物总量,但不包括活的植物体,它是活的土壤有机质部分[5].土壤微生物量可以反映土壤矿化能力和土壤活力[6],直接影响微生物驱动的物质循环过程,并调控土壤肥力形成、植物生长以及群落演替[7-9].山地生态系统是全球范围内对气候变化最敏感的陆地生态系统,约占全球陆地面积的24%[10],面临着生态环境破坏、生物多样性减少等一系列生态环境问题[11].山地生态系统中的气候、植被和土壤性质等环境因子沿海拔梯度在短距离内变化显著[12],土壤性质和气候条件的空间异质性也导致土壤生态化学计量和土壤微生物生物量及其垂直分布发生巨大变化[13].海拔导致相对短的地理距离内气候、土壤和生物等环境因素剧烈变化,从而影响生物地球化学循环过程.因此,在山地生态系统中研究土壤-微生物碳氮磷生态化学计量特征沿海拔梯度的变化规律及其影响因素,对于揭示山地生态系统土壤养分循环对环境变化的响应机制具有重要意义. ...
Biological stoichiometry from genes to ecosystems
0
2000
The ratios of life
0
2003
生态系统碳氮磷元素的生态化学计量学特征
1
2008
... 土壤碳(C)、氮(N)、磷(P)是陆地生态系统最基本的营养元素,也是植物赖以生存的必需条件,进而影响着植物功能性状、功能多样性与生态系统功能.生态化学计量学以元素组成来揭示生物化学和生态学之间的联系,可以更加深入探究生物之间元素循环的内在机理以及生态系统组分之间的联系[1-4].因此,研究土壤系统C、N、P含量及其生态化学计量特征规律,可以认识C、N、P在生态系统中的循环、平衡机制及其对植物群落结构和功能的影响.土壤微生物量是指土壤中体积小于5.0×103 μm3活的微生物总量,但不包括活的植物体,它是活的土壤有机质部分[5].土壤微生物量可以反映土壤矿化能力和土壤活力[6],直接影响微生物驱动的物质循环过程,并调控土壤肥力形成、植物生长以及群落演替[7-9].山地生态系统是全球范围内对气候变化最敏感的陆地生态系统,约占全球陆地面积的24%[10],面临着生态环境破坏、生物多样性减少等一系列生态环境问题[11].山地生态系统中的气候、植被和土壤性质等环境因子沿海拔梯度在短距离内变化显著[12],土壤性质和气候条件的空间异质性也导致土壤生态化学计量和土壤微生物生物量及其垂直分布发生巨大变化[13].海拔导致相对短的地理距离内气候、土壤和生物等环境因素剧烈变化,从而影响生物地球化学循环过程.因此,在山地生态系统中研究土壤-微生物碳氮磷生态化学计量特征沿海拔梯度的变化规律及其影响因素,对于揭示山地生态系统土壤养分循环对环境变化的响应机制具有重要意义. ...
Temporal analysis of the soil microbial community along a top sequence in pineland soils
1
2001
... 土壤碳(C)、氮(N)、磷(P)是陆地生态系统最基本的营养元素,也是植物赖以生存的必需条件,进而影响着植物功能性状、功能多样性与生态系统功能.生态化学计量学以元素组成来揭示生物化学和生态学之间的联系,可以更加深入探究生物之间元素循环的内在机理以及生态系统组分之间的联系[1-4].因此,研究土壤系统C、N、P含量及其生态化学计量特征规律,可以认识C、N、P在生态系统中的循环、平衡机制及其对植物群落结构和功能的影响.土壤微生物量是指土壤中体积小于5.0×103 μm3活的微生物总量,但不包括活的植物体,它是活的土壤有机质部分[5].土壤微生物量可以反映土壤矿化能力和土壤活力[6],直接影响微生物驱动的物质循环过程,并调控土壤肥力形成、植物生长以及群落演替[7-9].山地生态系统是全球范围内对气候变化最敏感的陆地生态系统,约占全球陆地面积的24%[10],面临着生态环境破坏、生物多样性减少等一系列生态环境问题[11].山地生态系统中的气候、植被和土壤性质等环境因子沿海拔梯度在短距离内变化显著[12],土壤性质和气候条件的空间异质性也导致土壤生态化学计量和土壤微生物生物量及其垂直分布发生巨大变化[13].海拔导致相对短的地理距离内气候、土壤和生物等环境因素剧烈变化,从而影响生物地球化学循环过程.因此,在山地生态系统中研究土壤-微生物碳氮磷生态化学计量特征沿海拔梯度的变化规律及其影响因素,对于揭示山地生态系统土壤养分循环对环境变化的响应机制具有重要意义. ...
退化喀斯特植被恢复过程中土壤微生物量碳的变化
1
2008
... 土壤碳(C)、氮(N)、磷(P)是陆地生态系统最基本的营养元素,也是植物赖以生存的必需条件,进而影响着植物功能性状、功能多样性与生态系统功能.生态化学计量学以元素组成来揭示生物化学和生态学之间的联系,可以更加深入探究生物之间元素循环的内在机理以及生态系统组分之间的联系[1-4].因此,研究土壤系统C、N、P含量及其生态化学计量特征规律,可以认识C、N、P在生态系统中的循环、平衡机制及其对植物群落结构和功能的影响.土壤微生物量是指土壤中体积小于5.0×103 μm3活的微生物总量,但不包括活的植物体,它是活的土壤有机质部分[5].土壤微生物量可以反映土壤矿化能力和土壤活力[6],直接影响微生物驱动的物质循环过程,并调控土壤肥力形成、植物生长以及群落演替[7-9].山地生态系统是全球范围内对气候变化最敏感的陆地生态系统,约占全球陆地面积的24%[10],面临着生态环境破坏、生物多样性减少等一系列生态环境问题[11].山地生态系统中的气候、植被和土壤性质等环境因子沿海拔梯度在短距离内变化显著[12],土壤性质和气候条件的空间异质性也导致土壤生态化学计量和土壤微生物生物量及其垂直分布发生巨大变化[13].海拔导致相对短的地理距离内气候、土壤和生物等环境因素剧烈变化,从而影响生物地球化学循环过程.因此,在山地生态系统中研究土壤-微生物碳氮磷生态化学计量特征沿海拔梯度的变化规律及其影响因素,对于揭示山地生态系统土壤养分循环对环境变化的响应机制具有重要意义. ...
长期施肥下我国农田土壤微生物及氨氧化菌研究进展
1
2018
... 土壤碳(C)、氮(N)、磷(P)是陆地生态系统最基本的营养元素,也是植物赖以生存的必需条件,进而影响着植物功能性状、功能多样性与生态系统功能.生态化学计量学以元素组成来揭示生物化学和生态学之间的联系,可以更加深入探究生物之间元素循环的内在机理以及生态系统组分之间的联系[1-4].因此,研究土壤系统C、N、P含量及其生态化学计量特征规律,可以认识C、N、P在生态系统中的循环、平衡机制及其对植物群落结构和功能的影响.土壤微生物量是指土壤中体积小于5.0×103 μm3活的微生物总量,但不包括活的植物体,它是活的土壤有机质部分[5].土壤微生物量可以反映土壤矿化能力和土壤活力[6],直接影响微生物驱动的物质循环过程,并调控土壤肥力形成、植物生长以及群落演替[7-9].山地生态系统是全球范围内对气候变化最敏感的陆地生态系统,约占全球陆地面积的24%[10],面临着生态环境破坏、生物多样性减少等一系列生态环境问题[11].山地生态系统中的气候、植被和土壤性质等环境因子沿海拔梯度在短距离内变化显著[12],土壤性质和气候条件的空间异质性也导致土壤生态化学计量和土壤微生物生物量及其垂直分布发生巨大变化[13].海拔导致相对短的地理距离内气候、土壤和生物等环境因素剧烈变化,从而影响生物地球化学循环过程.因此,在山地生态系统中研究土壤-微生物碳氮磷生态化学计量特征沿海拔梯度的变化规律及其影响因素,对于揭示山地生态系统土壤养分循环对环境变化的响应机制具有重要意义. ...
不同海拔红松混交林土壤微生物量碳、氮的生长季动态
0
2016
文山国家级自然保护区不同海拔地带性植被的土壤微生物生物量碳氮分布特征
2
2022
... 土壤碳(C)、氮(N)、磷(P)是陆地生态系统最基本的营养元素,也是植物赖以生存的必需条件,进而影响着植物功能性状、功能多样性与生态系统功能.生态化学计量学以元素组成来揭示生物化学和生态学之间的联系,可以更加深入探究生物之间元素循环的内在机理以及生态系统组分之间的联系[1-4].因此,研究土壤系统C、N、P含量及其生态化学计量特征规律,可以认识C、N、P在生态系统中的循环、平衡机制及其对植物群落结构和功能的影响.土壤微生物量是指土壤中体积小于5.0×103 μm3活的微生物总量,但不包括活的植物体,它是活的土壤有机质部分[5].土壤微生物量可以反映土壤矿化能力和土壤活力[6],直接影响微生物驱动的物质循环过程,并调控土壤肥力形成、植物生长以及群落演替[7-9].山地生态系统是全球范围内对气候变化最敏感的陆地生态系统,约占全球陆地面积的24%[10],面临着生态环境破坏、生物多样性减少等一系列生态环境问题[11].山地生态系统中的气候、植被和土壤性质等环境因子沿海拔梯度在短距离内变化显著[12],土壤性质和气候条件的空间异质性也导致土壤生态化学计量和土壤微生物生物量及其垂直分布发生巨大变化[13].海拔导致相对短的地理距离内气候、土壤和生物等环境因素剧烈变化,从而影响生物地球化学循环过程.因此,在山地生态系统中研究土壤-微生物碳氮磷生态化学计量特征沿海拔梯度的变化规律及其影响因素,对于揭示山地生态系统土壤养分循环对环境变化的响应机制具有重要意义. ...
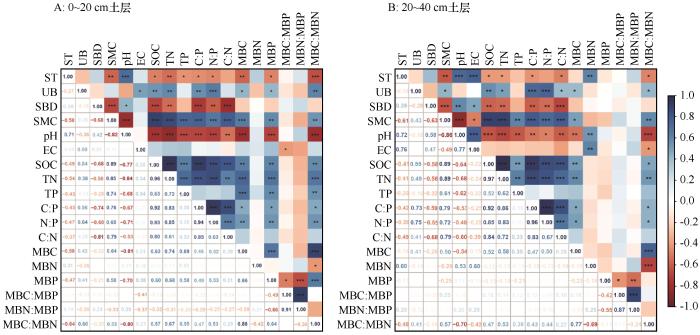
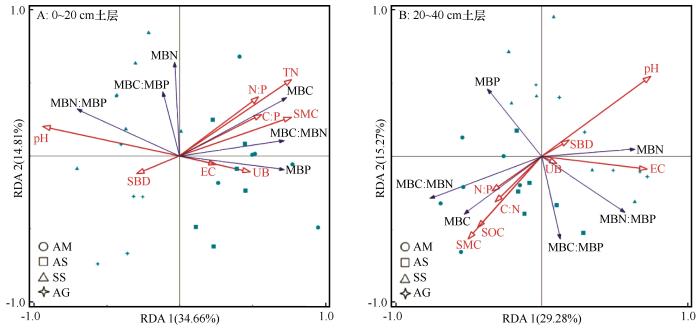
... 土壤理化性质与水热条件等环境因子显著影响植物的生长发育状况,继而影响植被凋落物质量及养分的归还,导致不同海拔土壤微生物生物量差异显著.本研究的冗余分析结果表明,0~20 cm土层中对MBC、MBN、MBP及其化学计量比解释率最高的3个因素为pH、TN、EC,20~40 cm土层中解释率最高的3个因素为pH、SOC、EC.随着海拔的增加pH值和EC减小,MBC、MBP、MBC∶MBN增加,MBC、MBP、MBC∶MBN与pH、EC均负相关.李聪等[9]在文山国家级自然保护区的研究也表明MBC含量与土壤pH值呈极显著负相关(P<0.01),土壤pH和EC可以改变土壤养分有效性及植物、微生物对养分的利用,直接或间接影响了微生物生长发育及群落结构,从而对微生物生物量也产生影响[44].表层土壤MBC、MBP和MBC∶MBN海拔分布特征与SOC、TN大致相同,这与Chen等[27]在青藏高原高寒草地以及曾全超等[45]在黄土高原不同乔木林的研究结果一致,同时He等[46]利用全世界73个海拔样带的59项研究进行对比也证明了这一结果.因为土壤有机质对微生物的生长发育有着至关重要的作用. ...
The responses of ammonia-oxidizing microorganisms to different environmental factors determine their elevational distribution and assembly patterns
1
2022
... 土壤碳(C)、氮(N)、磷(P)是陆地生态系统最基本的营养元素,也是植物赖以生存的必需条件,进而影响着植物功能性状、功能多样性与生态系统功能.生态化学计量学以元素组成来揭示生物化学和生态学之间的联系,可以更加深入探究生物之间元素循环的内在机理以及生态系统组分之间的联系[1-4].因此,研究土壤系统C、N、P含量及其生态化学计量特征规律,可以认识C、N、P在生态系统中的循环、平衡机制及其对植物群落结构和功能的影响.土壤微生物量是指土壤中体积小于5.0×103 μm3活的微生物总量,但不包括活的植物体,它是活的土壤有机质部分[5].土壤微生物量可以反映土壤矿化能力和土壤活力[6],直接影响微生物驱动的物质循环过程,并调控土壤肥力形成、植物生长以及群落演替[7-9].山地生态系统是全球范围内对气候变化最敏感的陆地生态系统,约占全球陆地面积的24%[10],面临着生态环境破坏、生物多样性减少等一系列生态环境问题[11].山地生态系统中的气候、植被和土壤性质等环境因子沿海拔梯度在短距离内变化显著[12],土壤性质和气候条件的空间异质性也导致土壤生态化学计量和土壤微生物生物量及其垂直分布发生巨大变化[13].海拔导致相对短的地理距离内气候、土壤和生物等环境因素剧烈变化,从而影响生物地球化学循环过程.因此,在山地生态系统中研究土壤-微生物碳氮磷生态化学计量特征沿海拔梯度的变化规律及其影响因素,对于揭示山地生态系统土壤养分循环对环境变化的响应机制具有重要意义. ...
Geographical,climatic,and soil factors control the altitudinal pattern of rhizosphere microbial diversity and its driving effect on root zone soil multifunctionality in mountain ecosystems
1
2023
... 土壤碳(C)、氮(N)、磷(P)是陆地生态系统最基本的营养元素,也是植物赖以生存的必需条件,进而影响着植物功能性状、功能多样性与生态系统功能.生态化学计量学以元素组成来揭示生物化学和生态学之间的联系,可以更加深入探究生物之间元素循环的内在机理以及生态系统组分之间的联系[1-4].因此,研究土壤系统C、N、P含量及其生态化学计量特征规律,可以认识C、N、P在生态系统中的循环、平衡机制及其对植物群落结构和功能的影响.土壤微生物量是指土壤中体积小于5.0×103 μm3活的微生物总量,但不包括活的植物体,它是活的土壤有机质部分[5].土壤微生物量可以反映土壤矿化能力和土壤活力[6],直接影响微生物驱动的物质循环过程,并调控土壤肥力形成、植物生长以及群落演替[7-9].山地生态系统是全球范围内对气候变化最敏感的陆地生态系统,约占全球陆地面积的24%[10],面临着生态环境破坏、生物多样性减少等一系列生态环境问题[11].山地生态系统中的气候、植被和土壤性质等环境因子沿海拔梯度在短距离内变化显著[12],土壤性质和气候条件的空间异质性也导致土壤生态化学计量和土壤微生物生物量及其垂直分布发生巨大变化[13].海拔导致相对短的地理距离内气候、土壤和生物等环境因素剧烈变化,从而影响生物地球化学循环过程.因此,在山地生态系统中研究土壤-微生物碳氮磷生态化学计量特征沿海拔梯度的变化规律及其影响因素,对于揭示山地生态系统土壤养分循环对环境变化的响应机制具有重要意义. ...
Could temperature and water availability drive elevational species richness patterns?a global case study for bats
1
2007
... 土壤碳(C)、氮(N)、磷(P)是陆地生态系统最基本的营养元素,也是植物赖以生存的必需条件,进而影响着植物功能性状、功能多样性与生态系统功能.生态化学计量学以元素组成来揭示生物化学和生态学之间的联系,可以更加深入探究生物之间元素循环的内在机理以及生态系统组分之间的联系[1-4].因此,研究土壤系统C、N、P含量及其生态化学计量特征规律,可以认识C、N、P在生态系统中的循环、平衡机制及其对植物群落结构和功能的影响.土壤微生物量是指土壤中体积小于5.0×103 μm3活的微生物总量,但不包括活的植物体,它是活的土壤有机质部分[5].土壤微生物量可以反映土壤矿化能力和土壤活力[6],直接影响微生物驱动的物质循环过程,并调控土壤肥力形成、植物生长以及群落演替[7-9].山地生态系统是全球范围内对气候变化最敏感的陆地生态系统,约占全球陆地面积的24%[10],面临着生态环境破坏、生物多样性减少等一系列生态环境问题[11].山地生态系统中的气候、植被和土壤性质等环境因子沿海拔梯度在短距离内变化显著[12],土壤性质和气候条件的空间异质性也导致土壤生态化学计量和土壤微生物生物量及其垂直分布发生巨大变化[13].海拔导致相对短的地理距离内气候、土壤和生物等环境因素剧烈变化,从而影响生物地球化学循环过程.因此,在山地生态系统中研究土壤-微生物碳氮磷生态化学计量特征沿海拔梯度的变化规律及其影响因素,对于揭示山地生态系统土壤养分循环对环境变化的响应机制具有重要意义. ...
A global analysis of soil microbial biomass carbon,nitrogen and phosphorus in terrestrial ecosystems
1
2013
... 土壤碳(C)、氮(N)、磷(P)是陆地生态系统最基本的营养元素,也是植物赖以生存的必需条件,进而影响着植物功能性状、功能多样性与生态系统功能.生态化学计量学以元素组成来揭示生物化学和生态学之间的联系,可以更加深入探究生物之间元素循环的内在机理以及生态系统组分之间的联系[1-4].因此,研究土壤系统C、N、P含量及其生态化学计量特征规律,可以认识C、N、P在生态系统中的循环、平衡机制及其对植物群落结构和功能的影响.土壤微生物量是指土壤中体积小于5.0×103 μm3活的微生物总量,但不包括活的植物体,它是活的土壤有机质部分[5].土壤微生物量可以反映土壤矿化能力和土壤活力[6],直接影响微生物驱动的物质循环过程,并调控土壤肥力形成、植物生长以及群落演替[7-9].山地生态系统是全球范围内对气候变化最敏感的陆地生态系统,约占全球陆地面积的24%[10],面临着生态环境破坏、生物多样性减少等一系列生态环境问题[11].山地生态系统中的气候、植被和土壤性质等环境因子沿海拔梯度在短距离内变化显著[12],土壤性质和气候条件的空间异质性也导致土壤生态化学计量和土壤微生物生物量及其垂直分布发生巨大变化[13].海拔导致相对短的地理距离内气候、土壤和生物等环境因素剧烈变化,从而影响生物地球化学循环过程.因此,在山地生态系统中研究土壤-微生物碳氮磷生态化学计量特征沿海拔梯度的变化规律及其影响因素,对于揭示山地生态系统土壤养分循环对环境变化的响应机制具有重要意义. ...
关帝山华北落叶松林下植物多样性和土壤碳氮磷的海拔梯度格局
1
2016
... 国内外学者就土壤理化性质沿海拔梯度的变化进行了深入、广泛的研究.陈顺治[14]发现,关帝山不同土层土壤总碳(TC)、全氮(TN)和全磷(TP)含量的分布特征均表现为沿着海拔上升而不断增加,显著正相关;刘倩等[15]研究表明,武功山山地草甸土壤C、N、P含量随海拔升高先增加后降低,土壤TP含量变异较小,土壤C∶N随海拔升高没有显著变化,土壤C∶P、N∶P随海拔升高先升高后下降;宁朋等[16]对贡嘎山的研究表明,贡嘎山各海拔土壤腐殖化能力和固碳能力相对偏弱,最低海拔2 800 m处土壤养分含量和含水量最为丰富,然而相较于其他海拔P元素更为缺乏;秦岭太白山地区土层深度和总土壤有机碳密度(SOCD)随着海拔梯度的增加呈现出减少的趋势,但是单位土层深度上的土壤有机碳密度(SOCD)却呈现出增加的趋势.随着海拔增加,土壤整体的TC、TN含量呈上升趋势,而TP含量无明显变化,随海拔的增加C∶N与C∶P表现出相似的变化趋势,N∶P随海拔增加先升高后下降[17-19];此外,Nottingham等[20]对不同海拔热带土壤的研究表明,C∶P与N∶P具有较高的空间变异特征,C∶N、C∶P及N∶P均随海拔的升高呈现逐渐上升的趋势.近年来,关于土壤微生物生物量沿海拔梯度的分布格局的研究也不断增多.贾培龙等[21]以黄土高原东西森林带作为研究区域,发现土壤微生物生物量C、N、P含量随经度整体呈先减少后增加的趋势,土壤微生物生物量C∶P呈先增后减的变化趋势;贺兰山土壤微生物量碳、氮随海拔高度升高而增加[22],Arunachalam等[23]在印度东北部丘陵地区的研究以及从静等[24]在神农架自然保护区的研究也取得了同样的结果.陈婕妮等[25]在武夷山进行研究不仅发现土壤微生物量碳、氮随海拔高度升高而增加,还得出武夷山土壤微生物生物量及其生态化学计量特征的主要影响因素是土壤有机碳的含量.Manral等[26]以印度喜马拉雅中部温带混交林为研究对象,发现微生物生物量C、N、P随海拔升高而显著降低(P<0.01).虽然围绕不同陆地生态系统及不同地理尺度土壤生态化学计量的研究日益增加,但相比之下,目前关于土壤微生物和土壤生态化学计量沿环境梯度的分布格局及影响因素尚无定论[27].因此,土壤-微生物化学计量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究. ...
武功山山地草甸不同海拔凋落物-土壤碳、氮、磷含量及其生态化学计量特征
1
2018
... 国内外学者就土壤理化性质沿海拔梯度的变化进行了深入、广泛的研究.陈顺治[14]发现,关帝山不同土层土壤总碳(TC)、全氮(TN)和全磷(TP)含量的分布特征均表现为沿着海拔上升而不断增加,显著正相关;刘倩等[15]研究表明,武功山山地草甸土壤C、N、P含量随海拔升高先增加后降低,土壤TP含量变异较小,土壤C∶N随海拔升高没有显著变化,土壤C∶P、N∶P随海拔升高先升高后下降;宁朋等[16]对贡嘎山的研究表明,贡嘎山各海拔土壤腐殖化能力和固碳能力相对偏弱,最低海拔2 800 m处土壤养分含量和含水量最为丰富,然而相较于其他海拔P元素更为缺乏;秦岭太白山地区土层深度和总土壤有机碳密度(SOCD)随着海拔梯度的增加呈现出减少的趋势,但是单位土层深度上的土壤有机碳密度(SOCD)却呈现出增加的趋势.随着海拔增加,土壤整体的TC、TN含量呈上升趋势,而TP含量无明显变化,随海拔的增加C∶N与C∶P表现出相似的变化趋势,N∶P随海拔增加先升高后下降[17-19];此外,Nottingham等[20]对不同海拔热带土壤的研究表明,C∶P与N∶P具有较高的空间变异特征,C∶N、C∶P及N∶P均随海拔的升高呈现逐渐上升的趋势.近年来,关于土壤微生物生物量沿海拔梯度的分布格局的研究也不断增多.贾培龙等[21]以黄土高原东西森林带作为研究区域,发现土壤微生物生物量C、N、P含量随经度整体呈先减少后增加的趋势,土壤微生物生物量C∶P呈先增后减的变化趋势;贺兰山土壤微生物量碳、氮随海拔高度升高而增加[22],Arunachalam等[23]在印度东北部丘陵地区的研究以及从静等[24]在神农架自然保护区的研究也取得了同样的结果.陈婕妮等[25]在武夷山进行研究不仅发现土壤微生物量碳、氮随海拔高度升高而增加,还得出武夷山土壤微生物生物量及其生态化学计量特征的主要影响因素是土壤有机碳的含量.Manral等[26]以印度喜马拉雅中部温带混交林为研究对象,发现微生物生物量C、N、P随海拔升高而显著降低(P<0.01).虽然围绕不同陆地生态系统及不同地理尺度土壤生态化学计量的研究日益增加,但相比之下,目前关于土壤微生物和土壤生态化学计量沿环境梯度的分布格局及影响因素尚无定论[27].因此,土壤-微生物化学计量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究. ...
贡嘎山不同海拔峨眉冷杉根际土壤化学性质及胞外酶活性
1
2021
... 国内外学者就土壤理化性质沿海拔梯度的变化进行了深入、广泛的研究.陈顺治[14]发现,关帝山不同土层土壤总碳(TC)、全氮(TN)和全磷(TP)含量的分布特征均表现为沿着海拔上升而不断增加,显著正相关;刘倩等[15]研究表明,武功山山地草甸土壤C、N、P含量随海拔升高先增加后降低,土壤TP含量变异较小,土壤C∶N随海拔升高没有显著变化,土壤C∶P、N∶P随海拔升高先升高后下降;宁朋等[16]对贡嘎山的研究表明,贡嘎山各海拔土壤腐殖化能力和固碳能力相对偏弱,最低海拔2 800 m处土壤养分含量和含水量最为丰富,然而相较于其他海拔P元素更为缺乏;秦岭太白山地区土层深度和总土壤有机碳密度(SOCD)随着海拔梯度的增加呈现出减少的趋势,但是单位土层深度上的土壤有机碳密度(SOCD)却呈现出增加的趋势.随着海拔增加,土壤整体的TC、TN含量呈上升趋势,而TP含量无明显变化,随海拔的增加C∶N与C∶P表现出相似的变化趋势,N∶P随海拔增加先升高后下降[17-19];此外,Nottingham等[20]对不同海拔热带土壤的研究表明,C∶P与N∶P具有较高的空间变异特征,C∶N、C∶P及N∶P均随海拔的升高呈现逐渐上升的趋势.近年来,关于土壤微生物生物量沿海拔梯度的分布格局的研究也不断增多.贾培龙等[21]以黄土高原东西森林带作为研究区域,发现土壤微生物生物量C、N、P含量随经度整体呈先减少后增加的趋势,土壤微生物生物量C∶P呈先增后减的变化趋势;贺兰山土壤微生物量碳、氮随海拔高度升高而增加[22],Arunachalam等[23]在印度东北部丘陵地区的研究以及从静等[24]在神农架自然保护区的研究也取得了同样的结果.陈婕妮等[25]在武夷山进行研究不仅发现土壤微生物量碳、氮随海拔高度升高而增加,还得出武夷山土壤微生物生物量及其生态化学计量特征的主要影响因素是土壤有机碳的含量.Manral等[26]以印度喜马拉雅中部温带混交林为研究对象,发现微生物生物量C、N、P随海拔升高而显著降低(P<0.01).虽然围绕不同陆地生态系统及不同地理尺度土壤生态化学计量的研究日益增加,但相比之下,目前关于土壤微生物和土壤生态化学计量沿环境梯度的分布格局及影响因素尚无定论[27].因此,土壤-微生物化学计量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究. ...
秦岭太白山北坡土壤有机碳储量的海拔梯度格局
1
2020
... 国内外学者就土壤理化性质沿海拔梯度的变化进行了深入、广泛的研究.陈顺治[14]发现,关帝山不同土层土壤总碳(TC)、全氮(TN)和全磷(TP)含量的分布特征均表现为沿着海拔上升而不断增加,显著正相关;刘倩等[15]研究表明,武功山山地草甸土壤C、N、P含量随海拔升高先增加后降低,土壤TP含量变异较小,土壤C∶N随海拔升高没有显著变化,土壤C∶P、N∶P随海拔升高先升高后下降;宁朋等[16]对贡嘎山的研究表明,贡嘎山各海拔土壤腐殖化能力和固碳能力相对偏弱,最低海拔2 800 m处土壤养分含量和含水量最为丰富,然而相较于其他海拔P元素更为缺乏;秦岭太白山地区土层深度和总土壤有机碳密度(SOCD)随着海拔梯度的增加呈现出减少的趋势,但是单位土层深度上的土壤有机碳密度(SOCD)却呈现出增加的趋势.随着海拔增加,土壤整体的TC、TN含量呈上升趋势,而TP含量无明显变化,随海拔的增加C∶N与C∶P表现出相似的变化趋势,N∶P随海拔增加先升高后下降[17-19];此外,Nottingham等[20]对不同海拔热带土壤的研究表明,C∶P与N∶P具有较高的空间变异特征,C∶N、C∶P及N∶P均随海拔的升高呈现逐渐上升的趋势.近年来,关于土壤微生物生物量沿海拔梯度的分布格局的研究也不断增多.贾培龙等[21]以黄土高原东西森林带作为研究区域,发现土壤微生物生物量C、N、P含量随经度整体呈先减少后增加的趋势,土壤微生物生物量C∶P呈先增后减的变化趋势;贺兰山土壤微生物量碳、氮随海拔高度升高而增加[22],Arunachalam等[23]在印度东北部丘陵地区的研究以及从静等[24]在神农架自然保护区的研究也取得了同样的结果.陈婕妮等[25]在武夷山进行研究不仅发现土壤微生物量碳、氮随海拔高度升高而增加,还得出武夷山土壤微生物生物量及其生态化学计量特征的主要影响因素是土壤有机碳的含量.Manral等[26]以印度喜马拉雅中部温带混交林为研究对象,发现微生物生物量C、N、P随海拔升高而显著降低(P<0.01).虽然围绕不同陆地生态系统及不同地理尺度土壤生态化学计量的研究日益增加,但相比之下,目前关于土壤微生物和土壤生态化学计量沿环境梯度的分布格局及影响因素尚无定论[27].因此,土壤-微生物化学计量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究. ...
秦岭松栎林土壤生态化学计量特征及其对海拔梯度的响应
0
2019
太白山不同海拔土壤碳、氮、磷含量及生态化学计量特征
2
2017
... 国内外学者就土壤理化性质沿海拔梯度的变化进行了深入、广泛的研究.陈顺治[14]发现,关帝山不同土层土壤总碳(TC)、全氮(TN)和全磷(TP)含量的分布特征均表现为沿着海拔上升而不断增加,显著正相关;刘倩等[15]研究表明,武功山山地草甸土壤C、N、P含量随海拔升高先增加后降低,土壤TP含量变异较小,土壤C∶N随海拔升高没有显著变化,土壤C∶P、N∶P随海拔升高先升高后下降;宁朋等[16]对贡嘎山的研究表明,贡嘎山各海拔土壤腐殖化能力和固碳能力相对偏弱,最低海拔2 800 m处土壤养分含量和含水量最为丰富,然而相较于其他海拔P元素更为缺乏;秦岭太白山地区土层深度和总土壤有机碳密度(SOCD)随着海拔梯度的增加呈现出减少的趋势,但是单位土层深度上的土壤有机碳密度(SOCD)却呈现出增加的趋势.随着海拔增加,土壤整体的TC、TN含量呈上升趋势,而TP含量无明显变化,随海拔的增加C∶N与C∶P表现出相似的变化趋势,N∶P随海拔增加先升高后下降[17-19];此外,Nottingham等[20]对不同海拔热带土壤的研究表明,C∶P与N∶P具有较高的空间变异特征,C∶N、C∶P及N∶P均随海拔的升高呈现逐渐上升的趋势.近年来,关于土壤微生物生物量沿海拔梯度的分布格局的研究也不断增多.贾培龙等[21]以黄土高原东西森林带作为研究区域,发现土壤微生物生物量C、N、P含量随经度整体呈先减少后增加的趋势,土壤微生物生物量C∶P呈先增后减的变化趋势;贺兰山土壤微生物量碳、氮随海拔高度升高而增加[22],Arunachalam等[23]在印度东北部丘陵地区的研究以及从静等[24]在神农架自然保护区的研究也取得了同样的结果.陈婕妮等[25]在武夷山进行研究不仅发现土壤微生物量碳、氮随海拔高度升高而增加,还得出武夷山土壤微生物生物量及其生态化学计量特征的主要影响因素是土壤有机碳的含量.Manral等[26]以印度喜马拉雅中部温带混交林为研究对象,发现微生物生物量C、N、P随海拔升高而显著降低(P<0.01).虽然围绕不同陆地生态系统及不同地理尺度土壤生态化学计量的研究日益增加,但相比之下,目前关于土壤微生物和土壤生态化学计量沿环境梯度的分布格局及影响因素尚无定论[27].因此,土壤-微生物化学计量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究. ...
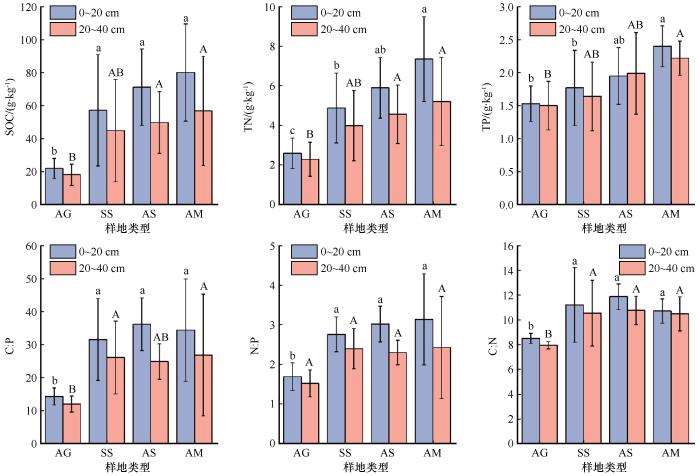
... 生态系统中植物营养主要来源于土壤养分,土壤养分又受到土壤性质和水热条件的影响,而各种环境因子沿海拔存在一定的空间变异性[19,33].本研究表明,马衔山不同土层土壤SOC和TN的含量均沿海拔升高呈现上升趋势,这与Bhople等[34]在阿尔卑斯山脉的研究结果相同,可能是由于温度随海拔升高而降低,高海拔有机物的循环受到低温限制[35].同时,温度降低也会导致土壤微生物活性下降,有机质分解变慢,增加土壤养分的积累[36].土壤TP主要受到土壤母质影响,来源相对固定,因而变异相对较小[37].本研究区域的土壤养分均随土层深度的增加而降低,与朱秋莲等[37]的研究结果一致.这是由于植被凋落物及有机质分解的影响,不同土层存在较大空间变异性.本研究所得C∶P、C∶N均沿海拔呈现先增加后减小的趋势,且最大值均出现在AS,与黄伟佳等[33]在南岭山地的研究结果相同,这是因为高山灌丛植被凋落物较难分解,向土壤归还的磷元素较少.同时,这也表明在中等海拔(3 100~3 500 m)N和P可能会限制土壤有机质分解和营养物质循环. ...
Soil microbial nutrient constraints along a tropical forest elevation gradient:a belowground test of a biogeochemical paradigm
1
2015
... 国内外学者就土壤理化性质沿海拔梯度的变化进行了深入、广泛的研究.陈顺治[14]发现,关帝山不同土层土壤总碳(TC)、全氮(TN)和全磷(TP)含量的分布特征均表现为沿着海拔上升而不断增加,显著正相关;刘倩等[15]研究表明,武功山山地草甸土壤C、N、P含量随海拔升高先增加后降低,土壤TP含量变异较小,土壤C∶N随海拔升高没有显著变化,土壤C∶P、N∶P随海拔升高先升高后下降;宁朋等[16]对贡嘎山的研究表明,贡嘎山各海拔土壤腐殖化能力和固碳能力相对偏弱,最低海拔2 800 m处土壤养分含量和含水量最为丰富,然而相较于其他海拔P元素更为缺乏;秦岭太白山地区土层深度和总土壤有机碳密度(SOCD)随着海拔梯度的增加呈现出减少的趋势,但是单位土层深度上的土壤有机碳密度(SOCD)却呈现出增加的趋势.随着海拔增加,土壤整体的TC、TN含量呈上升趋势,而TP含量无明显变化,随海拔的增加C∶N与C∶P表现出相似的变化趋势,N∶P随海拔增加先升高后下降[17-19];此外,Nottingham等[20]对不同海拔热带土壤的研究表明,C∶P与N∶P具有较高的空间变异特征,C∶N、C∶P及N∶P均随海拔的升高呈现逐渐上升的趋势.近年来,关于土壤微生物生物量沿海拔梯度的分布格局的研究也不断增多.贾培龙等[21]以黄土高原东西森林带作为研究区域,发现土壤微生物生物量C、N、P含量随经度整体呈先减少后增加的趋势,土壤微生物生物量C∶P呈先增后减的变化趋势;贺兰山土壤微生物量碳、氮随海拔高度升高而增加[22],Arunachalam等[23]在印度东北部丘陵地区的研究以及从静等[24]在神农架自然保护区的研究也取得了同样的结果.陈婕妮等[25]在武夷山进行研究不仅发现土壤微生物量碳、氮随海拔高度升高而增加,还得出武夷山土壤微生物生物量及其生态化学计量特征的主要影响因素是土壤有机碳的含量.Manral等[26]以印度喜马拉雅中部温带混交林为研究对象,发现微生物生物量C、N、P随海拔升高而显著降低(P<0.01).虽然围绕不同陆地生态系统及不同地理尺度土壤生态化学计量的研究日益增加,但相比之下,目前关于土壤微生物和土壤生态化学计量沿环境梯度的分布格局及影响因素尚无定论[27].因此,土壤-微生物化学计量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究. ...
黄土高原森林带土壤养分和微生物量及其生态化学计量变化特征
1
2020
... 国内外学者就土壤理化性质沿海拔梯度的变化进行了深入、广泛的研究.陈顺治[14]发现,关帝山不同土层土壤总碳(TC)、全氮(TN)和全磷(TP)含量的分布特征均表现为沿着海拔上升而不断增加,显著正相关;刘倩等[15]研究表明,武功山山地草甸土壤C、N、P含量随海拔升高先增加后降低,土壤TP含量变异较小,土壤C∶N随海拔升高没有显著变化,土壤C∶P、N∶P随海拔升高先升高后下降;宁朋等[16]对贡嘎山的研究表明,贡嘎山各海拔土壤腐殖化能力和固碳能力相对偏弱,最低海拔2 800 m处土壤养分含量和含水量最为丰富,然而相较于其他海拔P元素更为缺乏;秦岭太白山地区土层深度和总土壤有机碳密度(SOCD)随着海拔梯度的增加呈现出减少的趋势,但是单位土层深度上的土壤有机碳密度(SOCD)却呈现出增加的趋势.随着海拔增加,土壤整体的TC、TN含量呈上升趋势,而TP含量无明显变化,随海拔的增加C∶N与C∶P表现出相似的变化趋势,N∶P随海拔增加先升高后下降[17-19];此外,Nottingham等[20]对不同海拔热带土壤的研究表明,C∶P与N∶P具有较高的空间变异特征,C∶N、C∶P及N∶P均随海拔的升高呈现逐渐上升的趋势.近年来,关于土壤微生物生物量沿海拔梯度的分布格局的研究也不断增多.贾培龙等[21]以黄土高原东西森林带作为研究区域,发现土壤微生物生物量C、N、P含量随经度整体呈先减少后增加的趋势,土壤微生物生物量C∶P呈先增后减的变化趋势;贺兰山土壤微生物量碳、氮随海拔高度升高而增加[22],Arunachalam等[23]在印度东北部丘陵地区的研究以及从静等[24]在神农架自然保护区的研究也取得了同样的结果.陈婕妮等[25]在武夷山进行研究不仅发现土壤微生物量碳、氮随海拔高度升高而增加,还得出武夷山土壤微生物生物量及其生态化学计量特征的主要影响因素是土壤有机碳的含量.Manral等[26]以印度喜马拉雅中部温带混交林为研究对象,发现微生物生物量C、N、P随海拔升高而显著降低(P<0.01).虽然围绕不同陆地生态系统及不同地理尺度土壤生态化学计量的研究日益增加,但相比之下,目前关于土壤微生物和土壤生态化学计量沿环境梯度的分布格局及影响因素尚无定论[27].因此,土壤-微生物化学计量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究. ...
贺兰山东坡典型植物群落土壤微生物量碳、氮沿海拔梯度的变化特征
1
2010
... 国内外学者就土壤理化性质沿海拔梯度的变化进行了深入、广泛的研究.陈顺治[14]发现,关帝山不同土层土壤总碳(TC)、全氮(TN)和全磷(TP)含量的分布特征均表现为沿着海拔上升而不断增加,显著正相关;刘倩等[15]研究表明,武功山山地草甸土壤C、N、P含量随海拔升高先增加后降低,土壤TP含量变异较小,土壤C∶N随海拔升高没有显著变化,土壤C∶P、N∶P随海拔升高先升高后下降;宁朋等[16]对贡嘎山的研究表明,贡嘎山各海拔土壤腐殖化能力和固碳能力相对偏弱,最低海拔2 800 m处土壤养分含量和含水量最为丰富,然而相较于其他海拔P元素更为缺乏;秦岭太白山地区土层深度和总土壤有机碳密度(SOCD)随着海拔梯度的增加呈现出减少的趋势,但是单位土层深度上的土壤有机碳密度(SOCD)却呈现出增加的趋势.随着海拔增加,土壤整体的TC、TN含量呈上升趋势,而TP含量无明显变化,随海拔的增加C∶N与C∶P表现出相似的变化趋势,N∶P随海拔增加先升高后下降[17-19];此外,Nottingham等[20]对不同海拔热带土壤的研究表明,C∶P与N∶P具有较高的空间变异特征,C∶N、C∶P及N∶P均随海拔的升高呈现逐渐上升的趋势.近年来,关于土壤微生物生物量沿海拔梯度的分布格局的研究也不断增多.贾培龙等[21]以黄土高原东西森林带作为研究区域,发现土壤微生物生物量C、N、P含量随经度整体呈先减少后增加的趋势,土壤微生物生物量C∶P呈先增后减的变化趋势;贺兰山土壤微生物量碳、氮随海拔高度升高而增加[22],Arunachalam等[23]在印度东北部丘陵地区的研究以及从静等[24]在神农架自然保护区的研究也取得了同样的结果.陈婕妮等[25]在武夷山进行研究不仅发现土壤微生物量碳、氮随海拔高度升高而增加,还得出武夷山土壤微生物生物量及其生态化学计量特征的主要影响因素是土壤有机碳的含量.Manral等[26]以印度喜马拉雅中部温带混交林为研究对象,发现微生物生物量C、N、P随海拔升高而显著降低(P<0.01).虽然围绕不同陆地生态系统及不同地理尺度土壤生态化学计量的研究日益增加,但相比之下,目前关于土壤微生物和土壤生态化学计量沿环境梯度的分布格局及影响因素尚无定论[27].因此,土壤-微生物化学计量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究. ...
Ecosystem restoration of jhum fallows in northeast India:microbial C and N along altitudinal and successional gradients
1
2003
... 国内外学者就土壤理化性质沿海拔梯度的变化进行了深入、广泛的研究.陈顺治[14]发现,关帝山不同土层土壤总碳(TC)、全氮(TN)和全磷(TP)含量的分布特征均表现为沿着海拔上升而不断增加,显著正相关;刘倩等[15]研究表明,武功山山地草甸土壤C、N、P含量随海拔升高先增加后降低,土壤TP含量变异较小,土壤C∶N随海拔升高没有显著变化,土壤C∶P、N∶P随海拔升高先升高后下降;宁朋等[16]对贡嘎山的研究表明,贡嘎山各海拔土壤腐殖化能力和固碳能力相对偏弱,最低海拔2 800 m处土壤养分含量和含水量最为丰富,然而相较于其他海拔P元素更为缺乏;秦岭太白山地区土层深度和总土壤有机碳密度(SOCD)随着海拔梯度的增加呈现出减少的趋势,但是单位土层深度上的土壤有机碳密度(SOCD)却呈现出增加的趋势.随着海拔增加,土壤整体的TC、TN含量呈上升趋势,而TP含量无明显变化,随海拔的增加C∶N与C∶P表现出相似的变化趋势,N∶P随海拔增加先升高后下降[17-19];此外,Nottingham等[20]对不同海拔热带土壤的研究表明,C∶P与N∶P具有较高的空间变异特征,C∶N、C∶P及N∶P均随海拔的升高呈现逐渐上升的趋势.近年来,关于土壤微生物生物量沿海拔梯度的分布格局的研究也不断增多.贾培龙等[21]以黄土高原东西森林带作为研究区域,发现土壤微生物生物量C、N、P含量随经度整体呈先减少后增加的趋势,土壤微生物生物量C∶P呈先增后减的变化趋势;贺兰山土壤微生物量碳、氮随海拔高度升高而增加[22],Arunachalam等[23]在印度东北部丘陵地区的研究以及从静等[24]在神农架自然保护区的研究也取得了同样的结果.陈婕妮等[25]在武夷山进行研究不仅发现土壤微生物量碳、氮随海拔高度升高而增加,还得出武夷山土壤微生物生物量及其生态化学计量特征的主要影响因素是土壤有机碳的含量.Manral等[26]以印度喜马拉雅中部温带混交林为研究对象,发现微生物生物量C、N、P随海拔升高而显著降低(P<0.01).虽然围绕不同陆地生态系统及不同地理尺度土壤生态化学计量的研究日益增加,但相比之下,目前关于土壤微生物和土壤生态化学计量沿环境梯度的分布格局及影响因素尚无定论[27].因此,土壤-微生物化学计量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究. ...
神农架自然保护区土壤微生物生物量碳、氮沿海拔梯度的变化及其影响因素
1
2014
... 国内外学者就土壤理化性质沿海拔梯度的变化进行了深入、广泛的研究.陈顺治[14]发现,关帝山不同土层土壤总碳(TC)、全氮(TN)和全磷(TP)含量的分布特征均表现为沿着海拔上升而不断增加,显著正相关;刘倩等[15]研究表明,武功山山地草甸土壤C、N、P含量随海拔升高先增加后降低,土壤TP含量变异较小,土壤C∶N随海拔升高没有显著变化,土壤C∶P、N∶P随海拔升高先升高后下降;宁朋等[16]对贡嘎山的研究表明,贡嘎山各海拔土壤腐殖化能力和固碳能力相对偏弱,最低海拔2 800 m处土壤养分含量和含水量最为丰富,然而相较于其他海拔P元素更为缺乏;秦岭太白山地区土层深度和总土壤有机碳密度(SOCD)随着海拔梯度的增加呈现出减少的趋势,但是单位土层深度上的土壤有机碳密度(SOCD)却呈现出增加的趋势.随着海拔增加,土壤整体的TC、TN含量呈上升趋势,而TP含量无明显变化,随海拔的增加C∶N与C∶P表现出相似的变化趋势,N∶P随海拔增加先升高后下降[17-19];此外,Nottingham等[20]对不同海拔热带土壤的研究表明,C∶P与N∶P具有较高的空间变异特征,C∶N、C∶P及N∶P均随海拔的升高呈现逐渐上升的趋势.近年来,关于土壤微生物生物量沿海拔梯度的分布格局的研究也不断增多.贾培龙等[21]以黄土高原东西森林带作为研究区域,发现土壤微生物生物量C、N、P含量随经度整体呈先减少后增加的趋势,土壤微生物生物量C∶P呈先增后减的变化趋势;贺兰山土壤微生物量碳、氮随海拔高度升高而增加[22],Arunachalam等[23]在印度东北部丘陵地区的研究以及从静等[24]在神农架自然保护区的研究也取得了同样的结果.陈婕妮等[25]在武夷山进行研究不仅发现土壤微生物量碳、氮随海拔高度升高而增加,还得出武夷山土壤微生物生物量及其生态化学计量特征的主要影响因素是土壤有机碳的含量.Manral等[26]以印度喜马拉雅中部温带混交林为研究对象,发现微生物生物量C、N、P随海拔升高而显著降低(P<0.01).虽然围绕不同陆地生态系统及不同地理尺度土壤生态化学计量的研究日益增加,但相比之下,目前关于土壤微生物和土壤生态化学计量沿环境梯度的分布格局及影响因素尚无定论[27].因此,土壤-微生物化学计量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究. ...
武夷山不同海拔土壤微生物生物量碳、氮、磷含量及其生态化学计量特征
1
2023
... 国内外学者就土壤理化性质沿海拔梯度的变化进行了深入、广泛的研究.陈顺治[14]发现,关帝山不同土层土壤总碳(TC)、全氮(TN)和全磷(TP)含量的分布特征均表现为沿着海拔上升而不断增加,显著正相关;刘倩等[15]研究表明,武功山山地草甸土壤C、N、P含量随海拔升高先增加后降低,土壤TP含量变异较小,土壤C∶N随海拔升高没有显著变化,土壤C∶P、N∶P随海拔升高先升高后下降;宁朋等[16]对贡嘎山的研究表明,贡嘎山各海拔土壤腐殖化能力和固碳能力相对偏弱,最低海拔2 800 m处土壤养分含量和含水量最为丰富,然而相较于其他海拔P元素更为缺乏;秦岭太白山地区土层深度和总土壤有机碳密度(SOCD)随着海拔梯度的增加呈现出减少的趋势,但是单位土层深度上的土壤有机碳密度(SOCD)却呈现出增加的趋势.随着海拔增加,土壤整体的TC、TN含量呈上升趋势,而TP含量无明显变化,随海拔的增加C∶N与C∶P表现出相似的变化趋势,N∶P随海拔增加先升高后下降[17-19];此外,Nottingham等[20]对不同海拔热带土壤的研究表明,C∶P与N∶P具有较高的空间变异特征,C∶N、C∶P及N∶P均随海拔的升高呈现逐渐上升的趋势.近年来,关于土壤微生物生物量沿海拔梯度的分布格局的研究也不断增多.贾培龙等[21]以黄土高原东西森林带作为研究区域,发现土壤微生物生物量C、N、P含量随经度整体呈先减少后增加的趋势,土壤微生物生物量C∶P呈先增后减的变化趋势;贺兰山土壤微生物量碳、氮随海拔高度升高而增加[22],Arunachalam等[23]在印度东北部丘陵地区的研究以及从静等[24]在神农架自然保护区的研究也取得了同样的结果.陈婕妮等[25]在武夷山进行研究不仅发现土壤微生物量碳、氮随海拔高度升高而增加,还得出武夷山土壤微生物生物量及其生态化学计量特征的主要影响因素是土壤有机碳的含量.Manral等[26]以印度喜马拉雅中部温带混交林为研究对象,发现微生物生物量C、N、P随海拔升高而显著降低(P<0.01).虽然围绕不同陆地生态系统及不同地理尺度土壤生态化学计量的研究日益增加,但相比之下,目前关于土壤微生物和土壤生态化学计量沿环境梯度的分布格局及影响因素尚无定论[27].因此,土壤-微生物化学计量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究. ...
Seasonal dynamics of soil microbial biomass C,N and P along an altitudinal gradient in Central Himalaya,India
1
2023
... 国内外学者就土壤理化性质沿海拔梯度的变化进行了深入、广泛的研究.陈顺治[14]发现,关帝山不同土层土壤总碳(TC)、全氮(TN)和全磷(TP)含量的分布特征均表现为沿着海拔上升而不断增加,显著正相关;刘倩等[15]研究表明,武功山山地草甸土壤C、N、P含量随海拔升高先增加后降低,土壤TP含量变异较小,土壤C∶N随海拔升高没有显著变化,土壤C∶P、N∶P随海拔升高先升高后下降;宁朋等[16]对贡嘎山的研究表明,贡嘎山各海拔土壤腐殖化能力和固碳能力相对偏弱,最低海拔2 800 m处土壤养分含量和含水量最为丰富,然而相较于其他海拔P元素更为缺乏;秦岭太白山地区土层深度和总土壤有机碳密度(SOCD)随着海拔梯度的增加呈现出减少的趋势,但是单位土层深度上的土壤有机碳密度(SOCD)却呈现出增加的趋势.随着海拔增加,土壤整体的TC、TN含量呈上升趋势,而TP含量无明显变化,随海拔的增加C∶N与C∶P表现出相似的变化趋势,N∶P随海拔增加先升高后下降[17-19];此外,Nottingham等[20]对不同海拔热带土壤的研究表明,C∶P与N∶P具有较高的空间变异特征,C∶N、C∶P及N∶P均随海拔的升高呈现逐渐上升的趋势.近年来,关于土壤微生物生物量沿海拔梯度的分布格局的研究也不断增多.贾培龙等[21]以黄土高原东西森林带作为研究区域,发现土壤微生物生物量C、N、P含量随经度整体呈先减少后增加的趋势,土壤微生物生物量C∶P呈先增后减的变化趋势;贺兰山土壤微生物量碳、氮随海拔高度升高而增加[22],Arunachalam等[23]在印度东北部丘陵地区的研究以及从静等[24]在神农架自然保护区的研究也取得了同样的结果.陈婕妮等[25]在武夷山进行研究不仅发现土壤微生物量碳、氮随海拔高度升高而增加,还得出武夷山土壤微生物生物量及其生态化学计量特征的主要影响因素是土壤有机碳的含量.Manral等[26]以印度喜马拉雅中部温带混交林为研究对象,发现微生物生物量C、N、P随海拔升高而显著降低(P<0.01).虽然围绕不同陆地生态系统及不同地理尺度土壤生态化学计量的研究日益增加,但相比之下,目前关于土壤微生物和土壤生态化学计量沿环境梯度的分布格局及影响因素尚无定论[27].因此,土壤-微生物化学计量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究. ...
Linking microbial C∶N∶P stoichiometry to microbial community and abiotic factors along a 3500‐km grassland transect on the Tibetan Plateau
3
2016
... 国内外学者就土壤理化性质沿海拔梯度的变化进行了深入、广泛的研究.陈顺治[14]发现,关帝山不同土层土壤总碳(TC)、全氮(TN)和全磷(TP)含量的分布特征均表现为沿着海拔上升而不断增加,显著正相关;刘倩等[15]研究表明,武功山山地草甸土壤C、N、P含量随海拔升高先增加后降低,土壤TP含量变异较小,土壤C∶N随海拔升高没有显著变化,土壤C∶P、N∶P随海拔升高先升高后下降;宁朋等[16]对贡嘎山的研究表明,贡嘎山各海拔土壤腐殖化能力和固碳能力相对偏弱,最低海拔2 800 m处土壤养分含量和含水量最为丰富,然而相较于其他海拔P元素更为缺乏;秦岭太白山地区土层深度和总土壤有机碳密度(SOCD)随着海拔梯度的增加呈现出减少的趋势,但是单位土层深度上的土壤有机碳密度(SOCD)却呈现出增加的趋势.随着海拔增加,土壤整体的TC、TN含量呈上升趋势,而TP含量无明显变化,随海拔的增加C∶N与C∶P表现出相似的变化趋势,N∶P随海拔增加先升高后下降[17-19];此外,Nottingham等[20]对不同海拔热带土壤的研究表明,C∶P与N∶P具有较高的空间变异特征,C∶N、C∶P及N∶P均随海拔的升高呈现逐渐上升的趋势.近年来,关于土壤微生物生物量沿海拔梯度的分布格局的研究也不断增多.贾培龙等[21]以黄土高原东西森林带作为研究区域,发现土壤微生物生物量C、N、P含量随经度整体呈先减少后增加的趋势,土壤微生物生物量C∶P呈先增后减的变化趋势;贺兰山土壤微生物量碳、氮随海拔高度升高而增加[22],Arunachalam等[23]在印度东北部丘陵地区的研究以及从静等[24]在神农架自然保护区的研究也取得了同样的结果.陈婕妮等[25]在武夷山进行研究不仅发现土壤微生物量碳、氮随海拔高度升高而增加,还得出武夷山土壤微生物生物量及其生态化学计量特征的主要影响因素是土壤有机碳的含量.Manral等[26]以印度喜马拉雅中部温带混交林为研究对象,发现微生物生物量C、N、P随海拔升高而显著降低(P<0.01).虽然围绕不同陆地生态系统及不同地理尺度土壤生态化学计量的研究日益增加,但相比之下,目前关于土壤微生物和土壤生态化学计量沿环境梯度的分布格局及影响因素尚无定论[27].因此,土壤-微生物化学计量沿着海拔的垂直分布格局及其潜在驱动机制还有待进一步深入研究. ...
... MBC和MBN采用氯仿(CHCl3)熏蒸浸提(FE)法测定.对于每个样品,在0.5 mol·L-1 K2SO4(土壤∶溶液=1∶4)中提取一个子样品.第二个子样品在真空中用氯仿熏蒸24小时后用0.5 mol·L-1 K2SO4浸提.用multi N/C 3000分析仪测定提取液中的C和N浓度.使用MBC和MBN的相应转换系数0.45和0.54,乘以熏蒸和非熏蒸提取液元素浓度之间的差异,计算土壤微生物C、N含量.土壤微生物生物量磷(MBP)的测定方法中熏蒸程序与MBC和MBN相同,因此熏蒸和未熏蒸的样品都用0.5 mol·L-1 NaHCO3(土壤∶溶液=1∶20)浸提,并使用钼蓝比色法分析提取液以确定磷浓度.使用MBP的相应转换系数0.40,乘以熏蒸和非熏蒸提取液元素浓度之间的差异,以确定MBP[27]. ...
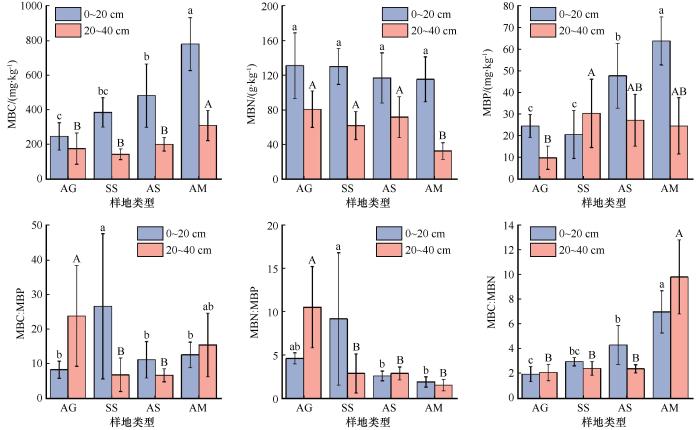
... 土壤理化性质与水热条件等环境因子显著影响植物的生长发育状况,继而影响植被凋落物质量及养分的归还,导致不同海拔土壤微生物生物量差异显著.本研究的冗余分析结果表明,0~20 cm土层中对MBC、MBN、MBP及其化学计量比解释率最高的3个因素为pH、TN、EC,20~40 cm土层中解释率最高的3个因素为pH、SOC、EC.随着海拔的增加pH值和EC减小,MBC、MBP、MBC∶MBN增加,MBC、MBP、MBC∶MBN与pH、EC均负相关.李聪等[9]在文山国家级自然保护区的研究也表明MBC含量与土壤pH值呈极显著负相关(P<0.01),土壤pH和EC可以改变土壤养分有效性及植物、微生物对养分的利用,直接或间接影响了微生物生长发育及群落结构,从而对微生物生物量也产生影响[44].表层土壤MBC、MBP和MBC∶MBN海拔分布特征与SOC、TN大致相同,这与Chen等[27]在青藏高原高寒草地以及曾全超等[45]在黄土高原不同乔木林的研究结果一致,同时He等[46]利用全世界73个海拔样带的59项研究进行对比也证明了这一结果.因为土壤有机质对微生物的生长发育有着至关重要的作用. ...
兰州马衔山发现多年冻土
1
1986
... 马衔山位于兰州市区以南约40 km处,主峰海拔3 670.3 m[28],是陇右黄土高原最高峰.马衔山属于东部季风区和西部干旱区之间,具有高原和季风气候的双重特点[29],马衔山山顶平均气温为-2.0~-2.2 ℃,多年平均降水量为494 mm[30].研究区内植被类型多样,垂直分布明显[31],依次为高山草原(2 600~2 900 m)、亚高山灌丛(2 900~3 100 m)、高山灌丛(3 300 m)、高山草甸(3 500 m)、荒漠草原和多年冻土区冻胀草丘(3 600 m). ...
兰州马衔山多年冻土区地表能量平衡特征分析
1
2013
... 马衔山位于兰州市区以南约40 km处,主峰海拔3 670.3 m[28],是陇右黄土高原最高峰.马衔山属于东部季风区和西部干旱区之间,具有高原和季风气候的双重特点[29],马衔山山顶平均气温为-2.0~-2.2 ℃,多年平均降水量为494 mm[30].研究区内植被类型多样,垂直分布明显[31],依次为高山草原(2 600~2 900 m)、亚高山灌丛(2 900~3 100 m)、高山灌丛(3 300 m)、高山草甸(3 500 m)、荒漠草原和多年冻土区冻胀草丘(3 600 m). ...
兰州马衔山多年冻土特征及变化趋势分析
1
2010
... 马衔山位于兰州市区以南约40 km处,主峰海拔3 670.3 m[28],是陇右黄土高原最高峰.马衔山属于东部季风区和西部干旱区之间,具有高原和季风气候的双重特点[29],马衔山山顶平均气温为-2.0~-2.2 ℃,多年平均降水量为494 mm[30].研究区内植被类型多样,垂直分布明显[31],依次为高山草原(2 600~2 900 m)、亚高山灌丛(2 900~3 100 m)、高山灌丛(3 300 m)、高山草甸(3 500 m)、荒漠草原和多年冻土区冻胀草丘(3 600 m). ...
甘肃省马衔山地区植被特征分析
1
1995
... 马衔山位于兰州市区以南约40 km处,主峰海拔3 670.3 m[28],是陇右黄土高原最高峰.马衔山属于东部季风区和西部干旱区之间,具有高原和季风气候的双重特点[29],马衔山山顶平均气温为-2.0~-2.2 ℃,多年平均降水量为494 mm[30].研究区内植被类型多样,垂直分布明显[31],依次为高山草原(2 600~2 900 m)、亚高山灌丛(2 900~3 100 m)、高山灌丛(3 300 m)、高山草甸(3 500 m)、荒漠草原和多年冻土区冻胀草丘(3 600 m). ...
1
2000
... 统一将TDR 350便携式土壤水分温度电导率速测仪(美国)的探针(20 cm)插入采样点0~20 cm和20~40 cm土层,待读数稳定之后读取土壤温度;用铝盒烘干法测定土壤含水量;在烘箱105 ℃下烘干24小时后测定EMB;土壤pH值采用pH计测定(土∶水=1∶2.5);使用电导率仪测定土壤EC(土∶水=1∶5);土壤有机碳含量(SOC)采用重铬酸钾硫酸氧化-外加热法测定;土壤TN含量用凯氏定氮法测定;土壤TP含量采用钼锑抗比色法测定[32]. ...
南岭山地不同海拔土壤生态化学计量特征及影响因素
3
2023
... 生态系统中植物营养主要来源于土壤养分,土壤养分又受到土壤性质和水热条件的影响,而各种环境因子沿海拔存在一定的空间变异性[19,33].本研究表明,马衔山不同土层土壤SOC和TN的含量均沿海拔升高呈现上升趋势,这与Bhople等[34]在阿尔卑斯山脉的研究结果相同,可能是由于温度随海拔升高而降低,高海拔有机物的循环受到低温限制[35].同时,温度降低也会导致土壤微生物活性下降,有机质分解变慢,增加土壤养分的积累[36].土壤TP主要受到土壤母质影响,来源相对固定,因而变异相对较小[37].本研究区域的土壤养分均随土层深度的增加而降低,与朱秋莲等[37]的研究结果一致.这是由于植被凋落物及有机质分解的影响,不同土层存在较大空间变异性.本研究所得C∶P、C∶N均沿海拔呈现先增加后减小的趋势,且最大值均出现在AS,与黄伟佳等[33]在南岭山地的研究结果相同,这是因为高山灌丛植被凋落物较难分解,向土壤归还的磷元素较少.同时,这也表明在中等海拔(3 100~3 500 m)N和P可能会限制土壤有机质分解和营养物质循环. ...
... [33]在南岭山地的研究结果相同,这是因为高山灌丛植被凋落物较难分解,向土壤归还的磷元素较少.同时,这也表明在中等海拔(3 100~3 500 m)N和P可能会限制土壤有机质分解和营养物质循环. ...
... 本研究区域,马衔山沿山体垂直带4种典型植被类型的表层土壤SOC、TN、MBC、MBP含量分别为21.95~80.11 g·kg-1、2.59~7.35 g·kg-1、246.26~780.26 mg·kg-1、20.56~63.86 mg·kg-1,空间变异性较大,海拔的变化引起区域内水热条件的变化,导致植被类型和微生物群落构成发生改变,并对土壤-微生物碳氮磷的空间分布格局产生影响,这与很多研究结果一致[42-43].本研究结果还表明,随着海拔的升高和土层深度的增加,土壤温度均呈现下降趋势,土壤含水率沿海拔升高逐渐增加,随深度的增加而减少.由于表层土壤通气性和水热条件良好,并且植被凋落物和动植物残体等为表层土壤微生物提供了丰富的底物来源[33],所以土壤养分及微生物生物量主要聚集在土壤表层. ...
Variations in soil and microbial biomass C,N and fungal biomass ergosterol along elevation and depth gradients in Alpine ecosystems
1
2019
... 生态系统中植物营养主要来源于土壤养分,土壤养分又受到土壤性质和水热条件的影响,而各种环境因子沿海拔存在一定的空间变异性[19,33].本研究表明,马衔山不同土层土壤SOC和TN的含量均沿海拔升高呈现上升趋势,这与Bhople等[34]在阿尔卑斯山脉的研究结果相同,可能是由于温度随海拔升高而降低,高海拔有机物的循环受到低温限制[35].同时,温度降低也会导致土壤微生物活性下降,有机质分解变慢,增加土壤养分的积累[36].土壤TP主要受到土壤母质影响,来源相对固定,因而变异相对较小[37].本研究区域的土壤养分均随土层深度的增加而降低,与朱秋莲等[37]的研究结果一致.这是由于植被凋落物及有机质分解的影响,不同土层存在较大空间变异性.本研究所得C∶P、C∶N均沿海拔呈现先增加后减小的趋势,且最大值均出现在AS,与黄伟佳等[33]在南岭山地的研究结果相同,这是因为高山灌丛植被凋落物较难分解,向土壤归还的磷元素较少.同时,这也表明在中等海拔(3 100~3 500 m)N和P可能会限制土壤有机质分解和营养物质循环. ...
Altitudinal patterns and controls of plant and soil nutrient concentrations and stoichiometry in subtropical China
1
2016
... 生态系统中植物营养主要来源于土壤养分,土壤养分又受到土壤性质和水热条件的影响,而各种环境因子沿海拔存在一定的空间变异性[19,33].本研究表明,马衔山不同土层土壤SOC和TN的含量均沿海拔升高呈现上升趋势,这与Bhople等[34]在阿尔卑斯山脉的研究结果相同,可能是由于温度随海拔升高而降低,高海拔有机物的循环受到低温限制[35].同时,温度降低也会导致土壤微生物活性下降,有机质分解变慢,增加土壤养分的积累[36].土壤TP主要受到土壤母质影响,来源相对固定,因而变异相对较小[37].本研究区域的土壤养分均随土层深度的增加而降低,与朱秋莲等[37]的研究结果一致.这是由于植被凋落物及有机质分解的影响,不同土层存在较大空间变异性.本研究所得C∶P、C∶N均沿海拔呈现先增加后减小的趋势,且最大值均出现在AS,与黄伟佳等[33]在南岭山地的研究结果相同,这是因为高山灌丛植被凋落物较难分解,向土壤归还的磷元素较少.同时,这也表明在中等海拔(3 100~3 500 m)N和P可能会限制土壤有机质分解和营养物质循环. ...
Spatial pattern of C∶N∶P stoichiometry characteristics of alpine grassland in the Altunshan Nature Reserve at North Qinghai-Tibet Plateau
1
2021
... 生态系统中植物营养主要来源于土壤养分,土壤养分又受到土壤性质和水热条件的影响,而各种环境因子沿海拔存在一定的空间变异性[19,33].本研究表明,马衔山不同土层土壤SOC和TN的含量均沿海拔升高呈现上升趋势,这与Bhople等[34]在阿尔卑斯山脉的研究结果相同,可能是由于温度随海拔升高而降低,高海拔有机物的循环受到低温限制[35].同时,温度降低也会导致土壤微生物活性下降,有机质分解变慢,增加土壤养分的积累[36].土壤TP主要受到土壤母质影响,来源相对固定,因而变异相对较小[37].本研究区域的土壤养分均随土层深度的增加而降低,与朱秋莲等[37]的研究结果一致.这是由于植被凋落物及有机质分解的影响,不同土层存在较大空间变异性.本研究所得C∶P、C∶N均沿海拔呈现先增加后减小的趋势,且最大值均出现在AS,与黄伟佳等[33]在南岭山地的研究结果相同,这是因为高山灌丛植被凋落物较难分解,向土壤归还的磷元素较少.同时,这也表明在中等海拔(3 100~3 500 m)N和P可能会限制土壤有机质分解和营养物质循环. ...
黄土丘陵沟壑区不同植被区土壤生态化学计量特征
2
2013
... 生态系统中植物营养主要来源于土壤养分,土壤养分又受到土壤性质和水热条件的影响,而各种环境因子沿海拔存在一定的空间变异性[19,33].本研究表明,马衔山不同土层土壤SOC和TN的含量均沿海拔升高呈现上升趋势,这与Bhople等[34]在阿尔卑斯山脉的研究结果相同,可能是由于温度随海拔升高而降低,高海拔有机物的循环受到低温限制[35].同时,温度降低也会导致土壤微生物活性下降,有机质分解变慢,增加土壤养分的积累[36].土壤TP主要受到土壤母质影响,来源相对固定,因而变异相对较小[37].本研究区域的土壤养分均随土层深度的增加而降低,与朱秋莲等[37]的研究结果一致.这是由于植被凋落物及有机质分解的影响,不同土层存在较大空间变异性.本研究所得C∶P、C∶N均沿海拔呈现先增加后减小的趋势,且最大值均出现在AS,与黄伟佳等[33]在南岭山地的研究结果相同,这是因为高山灌丛植被凋落物较难分解,向土壤归还的磷元素较少.同时,这也表明在中等海拔(3 100~3 500 m)N和P可能会限制土壤有机质分解和营养物质循环. ...
... [37]的研究结果一致.这是由于植被凋落物及有机质分解的影响,不同土层存在较大空间变异性.本研究所得C∶P、C∶N均沿海拔呈现先增加后减小的趋势,且最大值均出现在AS,与黄伟佳等[33]在南岭山地的研究结果相同,这是因为高山灌丛植被凋落物较难分解,向土壤归还的磷元素较少.同时,这也表明在中等海拔(3 100~3 500 m)N和P可能会限制土壤有机质分解和营养物质循环. ...
贺兰山不同海拔典型植被带土壤微生物多样性
1
2013
... 土壤微生物生物量受到土壤养分和植被类型等因素影响,其中SOC是关键因素[38].本研究的结果也显示表层土壤MBC、MBP海拔分布特征与SOC、TN基本一致,即沿海拔呈上升趋势.这也进一步说明了土壤有机质对微生物生长发育的重要作用.柳杨等[39]在鼎湖山的研究表明,土壤MBC和MBN随海拔升高逐渐降低,与本研究结果相反,可能与海拔跨度和研究区地理位置有关.本研究区域属于中国东部季风区和西部干旱区之间,而鼎湖山地处热带和亚热带的过渡区,并且本研究选取海拔梯度为2 600~3 670 m,而柳杨等[39]选取的海拔梯度为100~600 m,跨度较小可能无法产生显著性差异.除土壤有机质含量外,土壤温度、水分及pH等环境因子也可以影响土壤微生物群落组成、微生物活性等因素,从而改变微生物量化学计量比.马衔山表层土壤MBN∶MBP沿海拔呈先增后减趋势,在SS取得最大值,说明海拔超过3 100 m后微生物对P的限制作用减弱;MBC∶MBN沿海拔逐渐增加,表明在海拔2 600~2 900 m土壤氮素利用率较高,随海拔增加利用率减小,与赵盼盼等[40]在戴云山黄山松林的研究结果一致.本研究中,不同海拔表层土壤MBC∶MBN、MBN∶MBP的平均值分别为4.04、4.55,远低于中国陆地生态系统土壤MBC∶MBN、MBN∶MBP平均值(13.7和9.23),说明该研究区土壤有效氮和有效磷含量低[41]. ...
鼎湖山森林演替和海拔梯度上的土壤微生物生物量碳氮变化
2
2017
... 土壤微生物生物量受到土壤养分和植被类型等因素影响,其中SOC是关键因素[38].本研究的结果也显示表层土壤MBC、MBP海拔分布特征与SOC、TN基本一致,即沿海拔呈上升趋势.这也进一步说明了土壤有机质对微生物生长发育的重要作用.柳杨等[39]在鼎湖山的研究表明,土壤MBC和MBN随海拔升高逐渐降低,与本研究结果相反,可能与海拔跨度和研究区地理位置有关.本研究区域属于中国东部季风区和西部干旱区之间,而鼎湖山地处热带和亚热带的过渡区,并且本研究选取海拔梯度为2 600~3 670 m,而柳杨等[39]选取的海拔梯度为100~600 m,跨度较小可能无法产生显著性差异.除土壤有机质含量外,土壤温度、水分及pH等环境因子也可以影响土壤微生物群落组成、微生物活性等因素,从而改变微生物量化学计量比.马衔山表层土壤MBN∶MBP沿海拔呈先增后减趋势,在SS取得最大值,说明海拔超过3 100 m后微生物对P的限制作用减弱;MBC∶MBN沿海拔逐渐增加,表明在海拔2 600~2 900 m土壤氮素利用率较高,随海拔增加利用率减小,与赵盼盼等[40]在戴云山黄山松林的研究结果一致.本研究中,不同海拔表层土壤MBC∶MBN、MBN∶MBP的平均值分别为4.04、4.55,远低于中国陆地生态系统土壤MBC∶MBN、MBN∶MBP平均值(13.7和9.23),说明该研究区土壤有效氮和有效磷含量低[41]. ...
... [39]选取的海拔梯度为100~600 m,跨度较小可能无法产生显著性差异.除土壤有机质含量外,土壤温度、水分及pH等环境因子也可以影响土壤微生物群落组成、微生物活性等因素,从而改变微生物量化学计量比.马衔山表层土壤MBN∶MBP沿海拔呈先增后减趋势,在SS取得最大值,说明海拔超过3 100 m后微生物对P的限制作用减弱;MBC∶MBN沿海拔逐渐增加,表明在海拔2 600~2 900 m土壤氮素利用率较高,随海拔增加利用率减小,与赵盼盼等[40]在戴云山黄山松林的研究结果一致.本研究中,不同海拔表层土壤MBC∶MBN、MBN∶MBP的平均值分别为4.04、4.55,远低于中国陆地生态系统土壤MBC∶MBN、MBN∶MBP平均值(13.7和9.23),说明该研究区土壤有效氮和有效磷含量低[41]. ...
海拔梯度变化对中亚热带黄山松土壤微生物生物量和群落结构的影响
1
2019
... 土壤微生物生物量受到土壤养分和植被类型等因素影响,其中SOC是关键因素[38].本研究的结果也显示表层土壤MBC、MBP海拔分布特征与SOC、TN基本一致,即沿海拔呈上升趋势.这也进一步说明了土壤有机质对微生物生长发育的重要作用.柳杨等[39]在鼎湖山的研究表明,土壤MBC和MBN随海拔升高逐渐降低,与本研究结果相反,可能与海拔跨度和研究区地理位置有关.本研究区域属于中国东部季风区和西部干旱区之间,而鼎湖山地处热带和亚热带的过渡区,并且本研究选取海拔梯度为2 600~3 670 m,而柳杨等[39]选取的海拔梯度为100~600 m,跨度较小可能无法产生显著性差异.除土壤有机质含量外,土壤温度、水分及pH等环境因子也可以影响土壤微生物群落组成、微生物活性等因素,从而改变微生物量化学计量比.马衔山表层土壤MBN∶MBP沿海拔呈先增后减趋势,在SS取得最大值,说明海拔超过3 100 m后微生物对P的限制作用减弱;MBC∶MBN沿海拔逐渐增加,表明在海拔2 600~2 900 m土壤氮素利用率较高,随海拔增加利用率减小,与赵盼盼等[40]在戴云山黄山松林的研究结果一致.本研究中,不同海拔表层土壤MBC∶MBN、MBN∶MBP的平均值分别为4.04、4.55,远低于中国陆地生态系统土壤MBC∶MBN、MBN∶MBP平均值(13.7和9.23),说明该研究区土壤有效氮和有效磷含量低[41]. ...
贺兰山不同海拔凋落物-土壤-微生物生态化学计量特征及其关系研究
1
2022
... 土壤微生物生物量受到土壤养分和植被类型等因素影响,其中SOC是关键因素[38].本研究的结果也显示表层土壤MBC、MBP海拔分布特征与SOC、TN基本一致,即沿海拔呈上升趋势.这也进一步说明了土壤有机质对微生物生长发育的重要作用.柳杨等[39]在鼎湖山的研究表明,土壤MBC和MBN随海拔升高逐渐降低,与本研究结果相反,可能与海拔跨度和研究区地理位置有关.本研究区域属于中国东部季风区和西部干旱区之间,而鼎湖山地处热带和亚热带的过渡区,并且本研究选取海拔梯度为2 600~3 670 m,而柳杨等[39]选取的海拔梯度为100~600 m,跨度较小可能无法产生显著性差异.除土壤有机质含量外,土壤温度、水分及pH等环境因子也可以影响土壤微生物群落组成、微生物活性等因素,从而改变微生物量化学计量比.马衔山表层土壤MBN∶MBP沿海拔呈先增后减趋势,在SS取得最大值,说明海拔超过3 100 m后微生物对P的限制作用减弱;MBC∶MBN沿海拔逐渐增加,表明在海拔2 600~2 900 m土壤氮素利用率较高,随海拔增加利用率减小,与赵盼盼等[40]在戴云山黄山松林的研究结果一致.本研究中,不同海拔表层土壤MBC∶MBN、MBN∶MBP的平均值分别为4.04、4.55,远低于中国陆地生态系统土壤MBC∶MBN、MBN∶MBP平均值(13.7和9.23),说明该研究区土壤有效氮和有效磷含量低[41]. ...
祁连山典型植被土壤碳、氮、磷含量及生态化学计量特征的垂直变化
1
2024
... 本研究区域,马衔山沿山体垂直带4种典型植被类型的表层土壤SOC、TN、MBC、MBP含量分别为21.95~80.11 g·kg-1、2.59~7.35 g·kg-1、246.26~780.26 mg·kg-1、20.56~63.86 mg·kg-1,空间变异性较大,海拔的变化引起区域内水热条件的变化,导致植被类型和微生物群落构成发生改变,并对土壤-微生物碳氮磷的空间分布格局产生影响,这与很多研究结果一致[42-43].本研究结果还表明,随着海拔的升高和土层深度的增加,土壤温度均呈现下降趋势,土壤含水率沿海拔升高逐渐增加,随深度的增加而减少.由于表层土壤通气性和水热条件良好,并且植被凋落物和动植物残体等为表层土壤微生物提供了丰富的底物来源[33],所以土壤养分及微生物生物量主要聚集在土壤表层. ...
亚高山草地土壤原生生物群落结构和多样性海拔分布格局
1
2021
... 本研究区域,马衔山沿山体垂直带4种典型植被类型的表层土壤SOC、TN、MBC、MBP含量分别为21.95~80.11 g·kg-1、2.59~7.35 g·kg-1、246.26~780.26 mg·kg-1、20.56~63.86 mg·kg-1,空间变异性较大,海拔的变化引起区域内水热条件的变化,导致植被类型和微生物群落构成发生改变,并对土壤-微生物碳氮磷的空间分布格局产生影响,这与很多研究结果一致[42-43].本研究结果还表明,随着海拔的升高和土层深度的增加,土壤温度均呈现下降趋势,土壤含水率沿海拔升高逐渐增加,随深度的增加而减少.由于表层土壤通气性和水热条件良好,并且植被凋落物和动植物残体等为表层土壤微生物提供了丰富的底物来源[33],所以土壤养分及微生物生物量主要聚集在土壤表层. ...
土壤微生物生物量碳氮磷与土壤酶化学计量对气候变化的响应机制
1
2018
... 土壤理化性质与水热条件等环境因子显著影响植物的生长发育状况,继而影响植被凋落物质量及养分的归还,导致不同海拔土壤微生物生物量差异显著.本研究的冗余分析结果表明,0~20 cm土层中对MBC、MBN、MBP及其化学计量比解释率最高的3个因素为pH、TN、EC,20~40 cm土层中解释率最高的3个因素为pH、SOC、EC.随着海拔的增加pH值和EC减小,MBC、MBP、MBC∶MBN增加,MBC、MBP、MBC∶MBN与pH、EC均负相关.李聪等[9]在文山国家级自然保护区的研究也表明MBC含量与土壤pH值呈极显著负相关(P<0.01),土壤pH和EC可以改变土壤养分有效性及植物、微生物对养分的利用,直接或间接影响了微生物生长发育及群落结构,从而对微生物生物量也产生影响[44].表层土壤MBC、MBP和MBC∶MBN海拔分布特征与SOC、TN大致相同,这与Chen等[27]在青藏高原高寒草地以及曾全超等[45]在黄土高原不同乔木林的研究结果一致,同时He等[46]利用全世界73个海拔样带的59项研究进行对比也证明了这一结果.因为土壤有机质对微生物的生长发育有着至关重要的作用. ...
黄土高原不同乔木林土壤微生物量碳氮和溶解性碳氮的特征
1
2015
... 土壤理化性质与水热条件等环境因子显著影响植物的生长发育状况,继而影响植被凋落物质量及养分的归还,导致不同海拔土壤微生物生物量差异显著.本研究的冗余分析结果表明,0~20 cm土层中对MBC、MBN、MBP及其化学计量比解释率最高的3个因素为pH、TN、EC,20~40 cm土层中解释率最高的3个因素为pH、SOC、EC.随着海拔的增加pH值和EC减小,MBC、MBP、MBC∶MBN增加,MBC、MBP、MBC∶MBN与pH、EC均负相关.李聪等[9]在文山国家级自然保护区的研究也表明MBC含量与土壤pH值呈极显著负相关(P<0.01),土壤pH和EC可以改变土壤养分有效性及植物、微生物对养分的利用,直接或间接影响了微生物生长发育及群落结构,从而对微生物生物量也产生影响[44].表层土壤MBC、MBP和MBC∶MBN海拔分布特征与SOC、TN大致相同,这与Chen等[27]在青藏高原高寒草地以及曾全超等[45]在黄土高原不同乔木林的研究结果一致,同时He等[46]利用全世界73个海拔样带的59项研究进行对比也证明了这一结果.因为土壤有机质对微生物的生长发育有着至关重要的作用. ...
Soil microbial biomass increases along elevational gradients in the tropics and subtropics but not elsewhere
1
2020
... 土壤理化性质与水热条件等环境因子显著影响植物的生长发育状况,继而影响植被凋落物质量及养分的归还,导致不同海拔土壤微生物生物量差异显著.本研究的冗余分析结果表明,0~20 cm土层中对MBC、MBN、MBP及其化学计量比解释率最高的3个因素为pH、TN、EC,20~40 cm土层中解释率最高的3个因素为pH、SOC、EC.随着海拔的增加pH值和EC减小,MBC、MBP、MBC∶MBN增加,MBC、MBP、MBC∶MBN与pH、EC均负相关.李聪等[9]在文山国家级自然保护区的研究也表明MBC含量与土壤pH值呈极显著负相关(P<0.01),土壤pH和EC可以改变土壤养分有效性及植物、微生物对养分的利用,直接或间接影响了微生物生长发育及群落结构,从而对微生物生物量也产生影响[44].表层土壤MBC、MBP和MBC∶MBN海拔分布特征与SOC、TN大致相同,这与Chen等[27]在青藏高原高寒草地以及曾全超等[45]在黄土高原不同乔木林的研究结果一致,同时He等[46]利用全世界73个海拔样带的59项研究进行对比也证明了这一结果.因为土壤有机质对微生物的生长发育有着至关重要的作用. ...
青藏高原高寒草地土壤微生物群落结构特征及影响因素
1
2022
... 植物地下生物量可用于表征区域土壤养分状况,所以除土壤理化性质外,地下生物量的大小也会影响土壤-微生物碳氮磷生态化学计量,本研究的相关性分析结果表明表层土壤MBC、MBP、MBC∶MBN与地下生物量显著正相关.此外,由于深层土壤地下生物量较少,所以生物量与生态化学计量没有很强的相关性.张路[47]对青藏高原高寒草地土壤微生物群落结构的影响因素的研究结果也表明地下生物量对微生物多样性有显著影响,从而改变微生物碳氮磷生态化学计量.产生以上结果的原因可能是由于植物根系分泌物可为微生物提供生长发育和繁殖所需能源,因此根系分泌物可以通过改变微生物多样性、调节微生物代谢过程等影响土壤-微生物碳氮磷含量[48]. ...
森林根系分泌物生态学研究:问题与展望
1
2018
... 植物地下生物量可用于表征区域土壤养分状况,所以除土壤理化性质外,地下生物量的大小也会影响土壤-微生物碳氮磷生态化学计量,本研究的相关性分析结果表明表层土壤MBC、MBP、MBC∶MBN与地下生物量显著正相关.此外,由于深层土壤地下生物量较少,所以生物量与生态化学计量没有很强的相关性.张路[47]对青藏高原高寒草地土壤微生物群落结构的影响因素的研究结果也表明地下生物量对微生物多样性有显著影响,从而改变微生物碳氮磷生态化学计量.产生以上结果的原因可能是由于植物根系分泌物可为微生物提供生长发育和繁殖所需能源,因此根系分泌物可以通过改变微生物多样性、调节微生物代谢过程等影响土壤-微生物碳氮磷含量[48]. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}