IPCC最新报告:全球温升预计在2021年至2040年内达到1.5 ℃
1
2023
... IPCC报告显示,21世纪末全球平均气温与1850—1900年的水平相比,可能升高1~1.8 ℃(最佳估算1.4 ℃),并且随着气温持续升高,部分地区最高升温幅度可能达到5.7 ℃[1].随着全球变暖,干旱发生频率和强度增加、影响范围扩大[2],对全球生态系统造成不同程度的影响,荒漠植物的生存将面临严峻挑战[3-4],高温干旱可引发荒漠植物死亡[5].梭梭(Haloxylon ammodendron)具有顽强的生态适应能力,能在极端高温、土壤贫瘠、水分匮乏且高盐碱的地区生存[6-9],防风固沙效果好,广泛分布于中国西北荒漠地区,对维系荒漠-绿洲共生关系作用重大,关乎中国西北干旱区乃至中亚地区生态安全和经济社会的可持续发展[10].作为温度敏感型的荒漠灌木[11],不同年龄的梭梭生理过程对温度的适应策略不同,而这种差异将会对环境呈现出不同的抗逆能力.水分是荒漠区植物生长最主要限制因子,控制着荒漠植物的生长与分布[12].对于荒漠植物而言,为了强化吸水能力,同化枝水势(Ψ)随干旱胁迫加剧而降低[13].另一方面,为了降低水分损耗,梭梭在高温时叶片响应敏感,关闭气孔降低气孔导度(Gs)[14].为应对高温缺水环境,梭梭会优先投入于根系生长,攫取水分和营养[15-16].梭梭水分利用效率(WUE)也体现其对干旱高温环境胁迫的应变机制[17].但不同林龄梭梭植株大小、根系、组织发育、组织生理特性和光合面积的差异[18],预示着对极端高温的气体交换和水分利用等策略会有较大差异.在全球温度持续升高的背景下,探究梭梭气体交换对极端高温适应策略,有助于预测未来不同林龄梭梭在极端高温下的生存境况[19]. ...
对IPCC第六次评估报告中有关干旱变化的解读
1
2021
... IPCC报告显示,21世纪末全球平均气温与1850—1900年的水平相比,可能升高1~1.8 ℃(最佳估算1.4 ℃),并且随着气温持续升高,部分地区最高升温幅度可能达到5.7 ℃[1].随着全球变暖,干旱发生频率和强度增加、影响范围扩大[2],对全球生态系统造成不同程度的影响,荒漠植物的生存将面临严峻挑战[3-4],高温干旱可引发荒漠植物死亡[5].梭梭(Haloxylon ammodendron)具有顽强的生态适应能力,能在极端高温、土壤贫瘠、水分匮乏且高盐碱的地区生存[6-9],防风固沙效果好,广泛分布于中国西北荒漠地区,对维系荒漠-绿洲共生关系作用重大,关乎中国西北干旱区乃至中亚地区生态安全和经济社会的可持续发展[10].作为温度敏感型的荒漠灌木[11],不同年龄的梭梭生理过程对温度的适应策略不同,而这种差异将会对环境呈现出不同的抗逆能力.水分是荒漠区植物生长最主要限制因子,控制着荒漠植物的生长与分布[12].对于荒漠植物而言,为了强化吸水能力,同化枝水势(Ψ)随干旱胁迫加剧而降低[13].另一方面,为了降低水分损耗,梭梭在高温时叶片响应敏感,关闭气孔降低气孔导度(Gs)[14].为应对高温缺水环境,梭梭会优先投入于根系生长,攫取水分和营养[15-16].梭梭水分利用效率(WUE)也体现其对干旱高温环境胁迫的应变机制[17].但不同林龄梭梭植株大小、根系、组织发育、组织生理特性和光合面积的差异[18],预示着对极端高温的气体交换和水分利用等策略会有较大差异.在全球温度持续升高的背景下,探究梭梭气体交换对极端高温适应策略,有助于预测未来不同林龄梭梭在极端高温下的生存境况[19]. ...
全球变暖背景下土壤微生物呼吸的热适应性:证据、机理和争议
1
2018
... IPCC报告显示,21世纪末全球平均气温与1850—1900年的水平相比,可能升高1~1.8 ℃(最佳估算1.4 ℃),并且随着气温持续升高,部分地区最高升温幅度可能达到5.7 ℃[1].随着全球变暖,干旱发生频率和强度增加、影响范围扩大[2],对全球生态系统造成不同程度的影响,荒漠植物的生存将面临严峻挑战[3-4],高温干旱可引发荒漠植物死亡[5].梭梭(Haloxylon ammodendron)具有顽强的生态适应能力,能在极端高温、土壤贫瘠、水分匮乏且高盐碱的地区生存[6-9],防风固沙效果好,广泛分布于中国西北荒漠地区,对维系荒漠-绿洲共生关系作用重大,关乎中国西北干旱区乃至中亚地区生态安全和经济社会的可持续发展[10].作为温度敏感型的荒漠灌木[11],不同年龄的梭梭生理过程对温度的适应策略不同,而这种差异将会对环境呈现出不同的抗逆能力.水分是荒漠区植物生长最主要限制因子,控制着荒漠植物的生长与分布[12].对于荒漠植物而言,为了强化吸水能力,同化枝水势(Ψ)随干旱胁迫加剧而降低[13].另一方面,为了降低水分损耗,梭梭在高温时叶片响应敏感,关闭气孔降低气孔导度(Gs)[14].为应对高温缺水环境,梭梭会优先投入于根系生长,攫取水分和营养[15-16].梭梭水分利用效率(WUE)也体现其对干旱高温环境胁迫的应变机制[17].但不同林龄梭梭植株大小、根系、组织发育、组织生理特性和光合面积的差异[18],预示着对极端高温的气体交换和水分利用等策略会有较大差异.在全球温度持续升高的背景下,探究梭梭气体交换对极端高温适应策略,有助于预测未来不同林龄梭梭在极端高温下的生存境况[19]. ...
生长季末期干旱胁迫对刺槐幼苗非结构性碳水化合物的影响
1
2016
... IPCC报告显示,21世纪末全球平均气温与1850—1900年的水平相比,可能升高1~1.8 ℃(最佳估算1.4 ℃),并且随着气温持续升高,部分地区最高升温幅度可能达到5.7 ℃[1].随着全球变暖,干旱发生频率和强度增加、影响范围扩大[2],对全球生态系统造成不同程度的影响,荒漠植物的生存将面临严峻挑战[3-4],高温干旱可引发荒漠植物死亡[5].梭梭(Haloxylon ammodendron)具有顽强的生态适应能力,能在极端高温、土壤贫瘠、水分匮乏且高盐碱的地区生存[6-9],防风固沙效果好,广泛分布于中国西北荒漠地区,对维系荒漠-绿洲共生关系作用重大,关乎中国西北干旱区乃至中亚地区生态安全和经济社会的可持续发展[10].作为温度敏感型的荒漠灌木[11],不同年龄的梭梭生理过程对温度的适应策略不同,而这种差异将会对环境呈现出不同的抗逆能力.水分是荒漠区植物生长最主要限制因子,控制着荒漠植物的生长与分布[12].对于荒漠植物而言,为了强化吸水能力,同化枝水势(Ψ)随干旱胁迫加剧而降低[13].另一方面,为了降低水分损耗,梭梭在高温时叶片响应敏感,关闭气孔降低气孔导度(Gs)[14].为应对高温缺水环境,梭梭会优先投入于根系生长,攫取水分和营养[15-16].梭梭水分利用效率(WUE)也体现其对干旱高温环境胁迫的应变机制[17].但不同林龄梭梭植株大小、根系、组织发育、组织生理特性和光合面积的差异[18],预示着对极端高温的气体交换和水分利用等策略会有较大差异.在全球温度持续升高的背景下,探究梭梭气体交换对极端高温适应策略,有助于预测未来不同林龄梭梭在极端高温下的生存境况[19]. ...
Hydraulic dysfunction due to root-exposure-initiated water stress is responsible for the mortality of.Salix gordejevii.shrubs on the windward slopes of active sand dunes
1
2021
... IPCC报告显示,21世纪末全球平均气温与1850—1900年的水平相比,可能升高1~1.8 ℃(最佳估算1.4 ℃),并且随着气温持续升高,部分地区最高升温幅度可能达到5.7 ℃[1].随着全球变暖,干旱发生频率和强度增加、影响范围扩大[2],对全球生态系统造成不同程度的影响,荒漠植物的生存将面临严峻挑战[3-4],高温干旱可引发荒漠植物死亡[5].梭梭(Haloxylon ammodendron)具有顽强的生态适应能力,能在极端高温、土壤贫瘠、水分匮乏且高盐碱的地区生存[6-9],防风固沙效果好,广泛分布于中国西北荒漠地区,对维系荒漠-绿洲共生关系作用重大,关乎中国西北干旱区乃至中亚地区生态安全和经济社会的可持续发展[10].作为温度敏感型的荒漠灌木[11],不同年龄的梭梭生理过程对温度的适应策略不同,而这种差异将会对环境呈现出不同的抗逆能力.水分是荒漠区植物生长最主要限制因子,控制着荒漠植物的生长与分布[12].对于荒漠植物而言,为了强化吸水能力,同化枝水势(Ψ)随干旱胁迫加剧而降低[13].另一方面,为了降低水分损耗,梭梭在高温时叶片响应敏感,关闭气孔降低气孔导度(Gs)[14].为应对高温缺水环境,梭梭会优先投入于根系生长,攫取水分和营养[15-16].梭梭水分利用效率(WUE)也体现其对干旱高温环境胁迫的应变机制[17].但不同林龄梭梭植株大小、根系、组织发育、组织生理特性和光合面积的差异[18],预示着对极端高温的气体交换和水分利用等策略会有较大差异.在全球温度持续升高的背景下,探究梭梭气体交换对极端高温适应策略,有助于预测未来不同林龄梭梭在极端高温下的生存境况[19]. ...
近60年间巴丹吉林沙漠气温和降水变化及其对湖泊的影响
1
2023
... IPCC报告显示,21世纪末全球平均气温与1850—1900年的水平相比,可能升高1~1.8 ℃(最佳估算1.4 ℃),并且随着气温持续升高,部分地区最高升温幅度可能达到5.7 ℃[1].随着全球变暖,干旱发生频率和强度增加、影响范围扩大[2],对全球生态系统造成不同程度的影响,荒漠植物的生存将面临严峻挑战[3-4],高温干旱可引发荒漠植物死亡[5].梭梭(Haloxylon ammodendron)具有顽强的生态适应能力,能在极端高温、土壤贫瘠、水分匮乏且高盐碱的地区生存[6-9],防风固沙效果好,广泛分布于中国西北荒漠地区,对维系荒漠-绿洲共生关系作用重大,关乎中国西北干旱区乃至中亚地区生态安全和经济社会的可持续发展[10].作为温度敏感型的荒漠灌木[11],不同年龄的梭梭生理过程对温度的适应策略不同,而这种差异将会对环境呈现出不同的抗逆能力.水分是荒漠区植物生长最主要限制因子,控制着荒漠植物的生长与分布[12].对于荒漠植物而言,为了强化吸水能力,同化枝水势(Ψ)随干旱胁迫加剧而降低[13].另一方面,为了降低水分损耗,梭梭在高温时叶片响应敏感,关闭气孔降低气孔导度(Gs)[14].为应对高温缺水环境,梭梭会优先投入于根系生长,攫取水分和营养[15-16].梭梭水分利用效率(WUE)也体现其对干旱高温环境胁迫的应变机制[17].但不同林龄梭梭植株大小、根系、组织发育、组织生理特性和光合面积的差异[18],预示着对极端高温的气体交换和水分利用等策略会有较大差异.在全球温度持续升高的背景下,探究梭梭气体交换对极端高温适应策略,有助于预测未来不同林龄梭梭在极端高温下的生存境况[19]. ...
Hydraulic characteristics and carbon metabolism of Haloxylon ammodendron under different water-salt content
0
2022
Regional signatures of plant response to drought and elevated temperature across a desert ecosystem
0
2013
Response of morphological characters and photosynthetic characteristics of Haloxylon ammodendron to water and salt stress
3
2021
... IPCC报告显示,21世纪末全球平均气温与1850—1900年的水平相比,可能升高1~1.8 ℃(最佳估算1.4 ℃),并且随着气温持续升高,部分地区最高升温幅度可能达到5.7 ℃[1].随着全球变暖,干旱发生频率和强度增加、影响范围扩大[2],对全球生态系统造成不同程度的影响,荒漠植物的生存将面临严峻挑战[3-4],高温干旱可引发荒漠植物死亡[5].梭梭(Haloxylon ammodendron)具有顽强的生态适应能力,能在极端高温、土壤贫瘠、水分匮乏且高盐碱的地区生存[6-9],防风固沙效果好,广泛分布于中国西北荒漠地区,对维系荒漠-绿洲共生关系作用重大,关乎中国西北干旱区乃至中亚地区生态安全和经济社会的可持续发展[10].作为温度敏感型的荒漠灌木[11],不同年龄的梭梭生理过程对温度的适应策略不同,而这种差异将会对环境呈现出不同的抗逆能力.水分是荒漠区植物生长最主要限制因子,控制着荒漠植物的生长与分布[12].对于荒漠植物而言,为了强化吸水能力,同化枝水势(Ψ)随干旱胁迫加剧而降低[13].另一方面,为了降低水分损耗,梭梭在高温时叶片响应敏感,关闭气孔降低气孔导度(Gs)[14].为应对高温缺水环境,梭梭会优先投入于根系生长,攫取水分和营养[15-16].梭梭水分利用效率(WUE)也体现其对干旱高温环境胁迫的应变机制[17].但不同林龄梭梭植株大小、根系、组织发育、组织生理特性和光合面积的差异[18],预示着对极端高温的气体交换和水分利用等策略会有较大差异.在全球温度持续升高的背景下,探究梭梭气体交换对极端高温适应策略,有助于预测未来不同林龄梭梭在极端高温下的生存境况[19]. ...
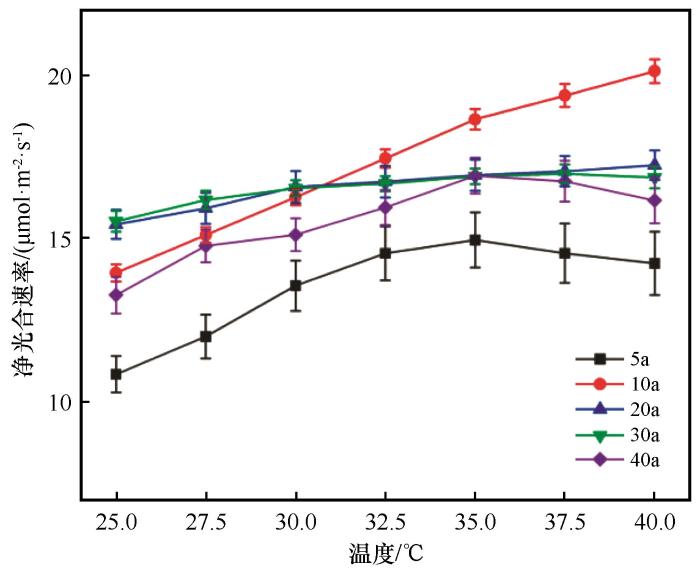
... Pn不仅受温度[21-22]和湿度等外界环境变化影响,也受蒸腾作用和气孔开放等生理过程影响[23](表2).不同林龄梭梭Pn随温度升高有较大差异,5 a梭梭整体最小(图2),对应的同化枝水势最低(图4),受到的水分胁迫最严重,研究结果表明环境可利用有效水分较少会限制植物光合作用[9,24].5 a梭梭Gs和Tr(图3、4)均小于其余林龄梭梭,虽然较小Gs和Tr能有效降低水分散失,但是也阻碍了二氧化碳的获取,导致5 a梭梭Pn整体最小.此外各个林龄梭梭在25~35 ℃下Pn波动幅度比35~40 ℃大,说明水分胁迫在极端高温时对梭梭的影响较大. ...
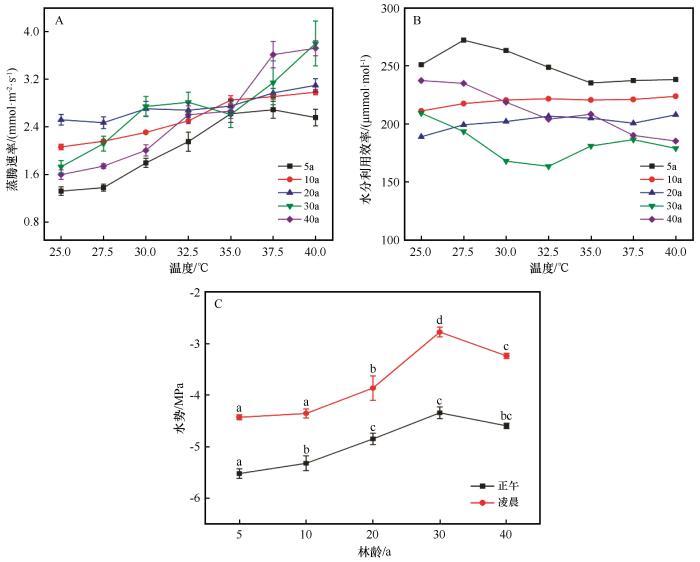
... 水分是限制荒漠植物梭梭生长发育分布的关键因素[24,27-28],梭梭同化枝水势是显示植物体内外水分状态的重要指标,通过测量正午和凌晨水势可以判断植物水分亏缺情况以及受到干旱胁迫程度的大小[29].有研究表明柠条干旱月份水势有明显降低,水势大小和环境水分状况相关联[30].在本研究中,5~30 a梭梭正午、凌晨同化枝水势依次增加,10~30 a梭梭增势明显.随着梭梭林龄增加,其受到的水分胁迫程度越小.根形态结构大小和分布在干旱环境中是影响植物个体水分可用性的关键因素,随着梭梭不断生长,其地下根系网络愈加发达,能够更有效地利用土壤水和地下水,缓解水分胁迫[18,31-33].但是40 a梭梭水势降低,说明其受到的水分胁迫程度增加.水势降低的原因可能为40 a梭梭新生叶片与老化叶片比例远远低于幼龄梭梭,老叶、中叶绿素a、b比例出现异常,致使梭梭叶片生理活性下降明显[34],进一步导致光合作用时对水分的利用效率下降,加剧水分胁迫.植物叶片功能性状活性是植物工厂运行的源动力[35],在极端干旱条件下会改变叶片形态结构和面积[9],本研究结果显示30 a和40 a梭梭叶片面积大幅缩减[36-37](表1).光合作用面积减少一方面有利于减少对水分和能量的需求,协调机体水分平衡,这是对水分胁迫加剧的应对策略.此外光合面积的减少也降低了光合产物的积累量,植物将有限的光合产物优先对根的投入导致地下部分呼吸作用消耗更多光合产物,可能会导致碳饥饿从而导致梭梭树死亡[15,38]. ...
绿洲-荒漠共生关系实验模拟平台(绿洲-荒漠平台)
1
2021
... IPCC报告显示,21世纪末全球平均气温与1850—1900年的水平相比,可能升高1~1.8 ℃(最佳估算1.4 ℃),并且随着气温持续升高,部分地区最高升温幅度可能达到5.7 ℃[1].随着全球变暖,干旱发生频率和强度增加、影响范围扩大[2],对全球生态系统造成不同程度的影响,荒漠植物的生存将面临严峻挑战[3-4],高温干旱可引发荒漠植物死亡[5].梭梭(Haloxylon ammodendron)具有顽强的生态适应能力,能在极端高温、土壤贫瘠、水分匮乏且高盐碱的地区生存[6-9],防风固沙效果好,广泛分布于中国西北荒漠地区,对维系荒漠-绿洲共生关系作用重大,关乎中国西北干旱区乃至中亚地区生态安全和经济社会的可持续发展[10].作为温度敏感型的荒漠灌木[11],不同年龄的梭梭生理过程对温度的适应策略不同,而这种差异将会对环境呈现出不同的抗逆能力.水分是荒漠区植物生长最主要限制因子,控制着荒漠植物的生长与分布[12].对于荒漠植物而言,为了强化吸水能力,同化枝水势(Ψ)随干旱胁迫加剧而降低[13].另一方面,为了降低水分损耗,梭梭在高温时叶片响应敏感,关闭气孔降低气孔导度(Gs)[14].为应对高温缺水环境,梭梭会优先投入于根系生长,攫取水分和营养[15-16].梭梭水分利用效率(WUE)也体现其对干旱高温环境胁迫的应变机制[17].但不同林龄梭梭植株大小、根系、组织发育、组织生理特性和光合面积的差异[18],预示着对极端高温的气体交换和水分利用等策略会有较大差异.在全球温度持续升高的背景下,探究梭梭气体交换对极端高温适应策略,有助于预测未来不同林龄梭梭在极端高温下的生存境况[19]. ...
民勤绿洲-荒漠过渡带梭梭人工林净碳交换及其影响因子
1
2019
... IPCC报告显示,21世纪末全球平均气温与1850—1900年的水平相比,可能升高1~1.8 ℃(最佳估算1.4 ℃),并且随着气温持续升高,部分地区最高升温幅度可能达到5.7 ℃[1].随着全球变暖,干旱发生频率和强度增加、影响范围扩大[2],对全球生态系统造成不同程度的影响,荒漠植物的生存将面临严峻挑战[3-4],高温干旱可引发荒漠植物死亡[5].梭梭(Haloxylon ammodendron)具有顽强的生态适应能力,能在极端高温、土壤贫瘠、水分匮乏且高盐碱的地区生存[6-9],防风固沙效果好,广泛分布于中国西北荒漠地区,对维系荒漠-绿洲共生关系作用重大,关乎中国西北干旱区乃至中亚地区生态安全和经济社会的可持续发展[10].作为温度敏感型的荒漠灌木[11],不同年龄的梭梭生理过程对温度的适应策略不同,而这种差异将会对环境呈现出不同的抗逆能力.水分是荒漠区植物生长最主要限制因子,控制着荒漠植物的生长与分布[12].对于荒漠植物而言,为了强化吸水能力,同化枝水势(Ψ)随干旱胁迫加剧而降低[13].另一方面,为了降低水分损耗,梭梭在高温时叶片响应敏感,关闭气孔降低气孔导度(Gs)[14].为应对高温缺水环境,梭梭会优先投入于根系生长,攫取水分和营养[15-16].梭梭水分利用效率(WUE)也体现其对干旱高温环境胁迫的应变机制[17].但不同林龄梭梭植株大小、根系、组织发育、组织生理特性和光合面积的差异[18],预示着对极端高温的气体交换和水分利用等策略会有较大差异.在全球温度持续升高的背景下,探究梭梭气体交换对极端高温适应策略,有助于预测未来不同林龄梭梭在极端高温下的生存境况[19]. ...
Desert ecosystems:environment and producers
1
1973
... IPCC报告显示,21世纪末全球平均气温与1850—1900年的水平相比,可能升高1~1.8 ℃(最佳估算1.4 ℃),并且随着气温持续升高,部分地区最高升温幅度可能达到5.7 ℃[1].随着全球变暖,干旱发生频率和强度增加、影响范围扩大[2],对全球生态系统造成不同程度的影响,荒漠植物的生存将面临严峻挑战[3-4],高温干旱可引发荒漠植物死亡[5].梭梭(Haloxylon ammodendron)具有顽强的生态适应能力,能在极端高温、土壤贫瘠、水分匮乏且高盐碱的地区生存[6-9],防风固沙效果好,广泛分布于中国西北荒漠地区,对维系荒漠-绿洲共生关系作用重大,关乎中国西北干旱区乃至中亚地区生态安全和经济社会的可持续发展[10].作为温度敏感型的荒漠灌木[11],不同年龄的梭梭生理过程对温度的适应策略不同,而这种差异将会对环境呈现出不同的抗逆能力.水分是荒漠区植物生长最主要限制因子,控制着荒漠植物的生长与分布[12].对于荒漠植物而言,为了强化吸水能力,同化枝水势(Ψ)随干旱胁迫加剧而降低[13].另一方面,为了降低水分损耗,梭梭在高温时叶片响应敏感,关闭气孔降低气孔导度(Gs)[14].为应对高温缺水环境,梭梭会优先投入于根系生长,攫取水分和营养[15-16].梭梭水分利用效率(WUE)也体现其对干旱高温环境胁迫的应变机制[17].但不同林龄梭梭植株大小、根系、组织发育、组织生理特性和光合面积的差异[18],预示着对极端高温的气体交换和水分利用等策略会有较大差异.在全球温度持续升高的背景下,探究梭梭气体交换对极端高温适应策略,有助于预测未来不同林龄梭梭在极端高温下的生存境况[19]. ...
Variation characteristics of water content in two typical eremophytes under drought stress in the drift desert hinterland
1
2013
... IPCC报告显示,21世纪末全球平均气温与1850—1900年的水平相比,可能升高1~1.8 ℃(最佳估算1.4 ℃),并且随着气温持续升高,部分地区最高升温幅度可能达到5.7 ℃[1].随着全球变暖,干旱发生频率和强度增加、影响范围扩大[2],对全球生态系统造成不同程度的影响,荒漠植物的生存将面临严峻挑战[3-4],高温干旱可引发荒漠植物死亡[5].梭梭(Haloxylon ammodendron)具有顽强的生态适应能力,能在极端高温、土壤贫瘠、水分匮乏且高盐碱的地区生存[6-9],防风固沙效果好,广泛分布于中国西北荒漠地区,对维系荒漠-绿洲共生关系作用重大,关乎中国西北干旱区乃至中亚地区生态安全和经济社会的可持续发展[10].作为温度敏感型的荒漠灌木[11],不同年龄的梭梭生理过程对温度的适应策略不同,而这种差异将会对环境呈现出不同的抗逆能力.水分是荒漠区植物生长最主要限制因子,控制着荒漠植物的生长与分布[12].对于荒漠植物而言,为了强化吸水能力,同化枝水势(Ψ)随干旱胁迫加剧而降低[13].另一方面,为了降低水分损耗,梭梭在高温时叶片响应敏感,关闭气孔降低气孔导度(Gs)[14].为应对高温缺水环境,梭梭会优先投入于根系生长,攫取水分和营养[15-16].梭梭水分利用效率(WUE)也体现其对干旱高温环境胁迫的应变机制[17].但不同林龄梭梭植株大小、根系、组织发育、组织生理特性和光合面积的差异[18],预示着对极端高温的气体交换和水分利用等策略会有较大差异.在全球温度持续升高的背景下,探究梭梭气体交换对极端高温适应策略,有助于预测未来不同林龄梭梭在极端高温下的生存境况[19]. ...
梭梭和多枝柽柳的枝干光合及其主要影响因子
1
2022
... IPCC报告显示,21世纪末全球平均气温与1850—1900年的水平相比,可能升高1~1.8 ℃(最佳估算1.4 ℃),并且随着气温持续升高,部分地区最高升温幅度可能达到5.7 ℃[1].随着全球变暖,干旱发生频率和强度增加、影响范围扩大[2],对全球生态系统造成不同程度的影响,荒漠植物的生存将面临严峻挑战[3-4],高温干旱可引发荒漠植物死亡[5].梭梭(Haloxylon ammodendron)具有顽强的生态适应能力,能在极端高温、土壤贫瘠、水分匮乏且高盐碱的地区生存[6-9],防风固沙效果好,广泛分布于中国西北荒漠地区,对维系荒漠-绿洲共生关系作用重大,关乎中国西北干旱区乃至中亚地区生态安全和经济社会的可持续发展[10].作为温度敏感型的荒漠灌木[11],不同年龄的梭梭生理过程对温度的适应策略不同,而这种差异将会对环境呈现出不同的抗逆能力.水分是荒漠区植物生长最主要限制因子,控制着荒漠植物的生长与分布[12].对于荒漠植物而言,为了强化吸水能力,同化枝水势(Ψ)随干旱胁迫加剧而降低[13].另一方面,为了降低水分损耗,梭梭在高温时叶片响应敏感,关闭气孔降低气孔导度(Gs)[14].为应对高温缺水环境,梭梭会优先投入于根系生长,攫取水分和营养[15-16].梭梭水分利用效率(WUE)也体现其对干旱高温环境胁迫的应变机制[17].但不同林龄梭梭植株大小、根系、组织发育、组织生理特性和光合面积的差异[18],预示着对极端高温的气体交换和水分利用等策略会有较大差异.在全球温度持续升高的背景下,探究梭梭气体交换对极端高温适应策略,有助于预测未来不同林龄梭梭在极端高温下的生存境况[19]. ...
A possible link between life and death of a xeric tree in desert
3
2016
... IPCC报告显示,21世纪末全球平均气温与1850—1900年的水平相比,可能升高1~1.8 ℃(最佳估算1.4 ℃),并且随着气温持续升高,部分地区最高升温幅度可能达到5.7 ℃[1].随着全球变暖,干旱发生频率和强度增加、影响范围扩大[2],对全球生态系统造成不同程度的影响,荒漠植物的生存将面临严峻挑战[3-4],高温干旱可引发荒漠植物死亡[5].梭梭(Haloxylon ammodendron)具有顽强的生态适应能力,能在极端高温、土壤贫瘠、水分匮乏且高盐碱的地区生存[6-9],防风固沙效果好,广泛分布于中国西北荒漠地区,对维系荒漠-绿洲共生关系作用重大,关乎中国西北干旱区乃至中亚地区生态安全和经济社会的可持续发展[10].作为温度敏感型的荒漠灌木[11],不同年龄的梭梭生理过程对温度的适应策略不同,而这种差异将会对环境呈现出不同的抗逆能力.水分是荒漠区植物生长最主要限制因子,控制着荒漠植物的生长与分布[12].对于荒漠植物而言,为了强化吸水能力,同化枝水势(Ψ)随干旱胁迫加剧而降低[13].另一方面,为了降低水分损耗,梭梭在高温时叶片响应敏感,关闭气孔降低气孔导度(Gs)[14].为应对高温缺水环境,梭梭会优先投入于根系生长,攫取水分和营养[15-16].梭梭水分利用效率(WUE)也体现其对干旱高温环境胁迫的应变机制[17].但不同林龄梭梭植株大小、根系、组织发育、组织生理特性和光合面积的差异[18],预示着对极端高温的气体交换和水分利用等策略会有较大差异.在全球温度持续升高的背景下,探究梭梭气体交换对极端高温适应策略,有助于预测未来不同林龄梭梭在极端高温下的生存境况[19]. ...
... 水分是限制荒漠植物梭梭生长发育分布的关键因素[24,27-28],梭梭同化枝水势是显示植物体内外水分状态的重要指标,通过测量正午和凌晨水势可以判断植物水分亏缺情况以及受到干旱胁迫程度的大小[29].有研究表明柠条干旱月份水势有明显降低,水势大小和环境水分状况相关联[30].在本研究中,5~30 a梭梭正午、凌晨同化枝水势依次增加,10~30 a梭梭增势明显.随着梭梭林龄增加,其受到的水分胁迫程度越小.根形态结构大小和分布在干旱环境中是影响植物个体水分可用性的关键因素,随着梭梭不断生长,其地下根系网络愈加发达,能够更有效地利用土壤水和地下水,缓解水分胁迫[18,31-33].但是40 a梭梭水势降低,说明其受到的水分胁迫程度增加.水势降低的原因可能为40 a梭梭新生叶片与老化叶片比例远远低于幼龄梭梭,老叶、中叶绿素a、b比例出现异常,致使梭梭叶片生理活性下降明显[34],进一步导致光合作用时对水分的利用效率下降,加剧水分胁迫.植物叶片功能性状活性是植物工厂运行的源动力[35],在极端干旱条件下会改变叶片形态结构和面积[9],本研究结果显示30 a和40 a梭梭叶片面积大幅缩减[36-37](表1).光合作用面积减少一方面有利于减少对水分和能量的需求,协调机体水分平衡,这是对水分胁迫加剧的应对策略.此外光合面积的减少也降低了光合产物的积累量,植物将有限的光合产物优先对根的投入导致地下部分呼吸作用消耗更多光合产物,可能会导致碳饥饿从而导致梭梭树死亡[15,38]. ...
... WUE是衡量植物吸收水分利用效率的指标,与光合作用、Gs、Tr和温度等因素相关[43-44],Gs、Tr增加在增强水分运输能力的同时也会致使水分损耗增大,而温度会影响气孔开放继而影响Tr(表2).植物调整WUE是关键的水分调节策略,特别是在极端高温的荒漠环境中,这对植物的生存至关重要.本研究中除40 a梭梭外,不同林龄梭梭WUE随林龄增加而降低(图4),这与已有学者研究结果不完全一致[45],原因可能为本研究区气候环境条件和其研究区域差异较大.除了10 a梭梭外,其余不同生长林龄梭梭WUE和Gs随温度升高变化趋势相反,且在Gs最低时WUE达到最大值,说明只有Gs降低,Tr和梭梭用于交换二氧化碳进行光合作用过程受到轻微限制时,WUE才能达到最大值,这与邓雄等[46]对怪柳气体水分交换研究得出的结论一致[47].5 a梭梭WUE最高,10 a次之,30 a最低,与它们受到水分胁迫程度(图4)刚好相反,即受到水分胁迫的梭梭会采取提高WUE的适应策略,侧面表明在干旱缺水的荒漠地区,不同林龄梭梭在受到不同的水分胁迫程度下会严格地做出相应的水分调节策略.较高的WUE有利于5 a和10 a梭梭在极端高温缺水的荒漠环境中降低对水分的损耗,提高光合产物产量,将有机物优先投入地下根系[15]以适应极端高温环境.5 a梭梭WUE随温度升高先增后减,在35 ℃后趋于平缓(图4),这与梭梭Gs、Tr变化趋势刚好相反.5 a梭梭Gs和Tr严格控制着WUE大小,在应对极端高温环境时对水分调节的管控能力强,气体交换各部分水分生理变化联系紧密,光合系统反应灵敏.除40 a梭梭外,其余林龄梭梭WUE在高温时波动较小,10、20 a梭梭响应曲线都较为平稳,表明它们在极端高温时能保持稳定的水分状态,减少不必要的水分损耗同时还能维持较好的光合作用状态.由于40 a梭梭Tr随温度不断升高且在极端高温时增势明显,导致其WUE不断减小.水分利用效率降低会导致梭梭在水资源限制较大的极端高温荒漠环境中生长和发育受到抑制,对极端高温逆境的适应力下降,致使植物的生物量减少,生长速度变慢,甚至可能无法正常完成生命周期. ...
Deepening rooting depths improve plant water and carbon status of a xeric tree during Summer drought
1
2019
... IPCC报告显示,21世纪末全球平均气温与1850—1900年的水平相比,可能升高1~1.8 ℃(最佳估算1.4 ℃),并且随着气温持续升高,部分地区最高升温幅度可能达到5.7 ℃[1].随着全球变暖,干旱发生频率和强度增加、影响范围扩大[2],对全球生态系统造成不同程度的影响,荒漠植物的生存将面临严峻挑战[3-4],高温干旱可引发荒漠植物死亡[5].梭梭(Haloxylon ammodendron)具有顽强的生态适应能力,能在极端高温、土壤贫瘠、水分匮乏且高盐碱的地区生存[6-9],防风固沙效果好,广泛分布于中国西北荒漠地区,对维系荒漠-绿洲共生关系作用重大,关乎中国西北干旱区乃至中亚地区生态安全和经济社会的可持续发展[10].作为温度敏感型的荒漠灌木[11],不同年龄的梭梭生理过程对温度的适应策略不同,而这种差异将会对环境呈现出不同的抗逆能力.水分是荒漠区植物生长最主要限制因子,控制着荒漠植物的生长与分布[12].对于荒漠植物而言,为了强化吸水能力,同化枝水势(Ψ)随干旱胁迫加剧而降低[13].另一方面,为了降低水分损耗,梭梭在高温时叶片响应敏感,关闭气孔降低气孔导度(Gs)[14].为应对高温缺水环境,梭梭会优先投入于根系生长,攫取水分和营养[15-16].梭梭水分利用效率(WUE)也体现其对干旱高温环境胁迫的应变机制[17].但不同林龄梭梭植株大小、根系、组织发育、组织生理特性和光合面积的差异[18],预示着对极端高温的气体交换和水分利用等策略会有较大差异.在全球温度持续升高的背景下,探究梭梭气体交换对极端高温适应策略,有助于预测未来不同林龄梭梭在极端高温下的生存境况[19]. ...
不同光环境下辣椒光合特性和瞬时水分利用效率
1
2019
... IPCC报告显示,21世纪末全球平均气温与1850—1900年的水平相比,可能升高1~1.8 ℃(最佳估算1.4 ℃),并且随着气温持续升高,部分地区最高升温幅度可能达到5.7 ℃[1].随着全球变暖,干旱发生频率和强度增加、影响范围扩大[2],对全球生态系统造成不同程度的影响,荒漠植物的生存将面临严峻挑战[3-4],高温干旱可引发荒漠植物死亡[5].梭梭(Haloxylon ammodendron)具有顽强的生态适应能力,能在极端高温、土壤贫瘠、水分匮乏且高盐碱的地区生存[6-9],防风固沙效果好,广泛分布于中国西北荒漠地区,对维系荒漠-绿洲共生关系作用重大,关乎中国西北干旱区乃至中亚地区生态安全和经济社会的可持续发展[10].作为温度敏感型的荒漠灌木[11],不同年龄的梭梭生理过程对温度的适应策略不同,而这种差异将会对环境呈现出不同的抗逆能力.水分是荒漠区植物生长最主要限制因子,控制着荒漠植物的生长与分布[12].对于荒漠植物而言,为了强化吸水能力,同化枝水势(Ψ)随干旱胁迫加剧而降低[13].另一方面,为了降低水分损耗,梭梭在高温时叶片响应敏感,关闭气孔降低气孔导度(Gs)[14].为应对高温缺水环境,梭梭会优先投入于根系生长,攫取水分和营养[15-16].梭梭水分利用效率(WUE)也体现其对干旱高温环境胁迫的应变机制[17].但不同林龄梭梭植株大小、根系、组织发育、组织生理特性和光合面积的差异[18],预示着对极端高温的气体交换和水分利用等策略会有较大差异.在全球温度持续升高的背景下,探究梭梭气体交换对极端高温适应策略,有助于预测未来不同林龄梭梭在极端高温下的生存境况[19]. ...
Differential drought responses between saplings and adult trees in four co-occurring species of New England
2
2005
... IPCC报告显示,21世纪末全球平均气温与1850—1900年的水平相比,可能升高1~1.8 ℃(最佳估算1.4 ℃),并且随着气温持续升高,部分地区最高升温幅度可能达到5.7 ℃[1].随着全球变暖,干旱发生频率和强度增加、影响范围扩大[2],对全球生态系统造成不同程度的影响,荒漠植物的生存将面临严峻挑战[3-4],高温干旱可引发荒漠植物死亡[5].梭梭(Haloxylon ammodendron)具有顽强的生态适应能力,能在极端高温、土壤贫瘠、水分匮乏且高盐碱的地区生存[6-9],防风固沙效果好,广泛分布于中国西北荒漠地区,对维系荒漠-绿洲共生关系作用重大,关乎中国西北干旱区乃至中亚地区生态安全和经济社会的可持续发展[10].作为温度敏感型的荒漠灌木[11],不同年龄的梭梭生理过程对温度的适应策略不同,而这种差异将会对环境呈现出不同的抗逆能力.水分是荒漠区植物生长最主要限制因子,控制着荒漠植物的生长与分布[12].对于荒漠植物而言,为了强化吸水能力,同化枝水势(Ψ)随干旱胁迫加剧而降低[13].另一方面,为了降低水分损耗,梭梭在高温时叶片响应敏感,关闭气孔降低气孔导度(Gs)[14].为应对高温缺水环境,梭梭会优先投入于根系生长,攫取水分和营养[15-16].梭梭水分利用效率(WUE)也体现其对干旱高温环境胁迫的应变机制[17].但不同林龄梭梭植株大小、根系、组织发育、组织生理特性和光合面积的差异[18],预示着对极端高温的气体交换和水分利用等策略会有较大差异.在全球温度持续升高的背景下,探究梭梭气体交换对极端高温适应策略,有助于预测未来不同林龄梭梭在极端高温下的生存境况[19]. ...
... 水分是限制荒漠植物梭梭生长发育分布的关键因素[24,27-28],梭梭同化枝水势是显示植物体内外水分状态的重要指标,通过测量正午和凌晨水势可以判断植物水分亏缺情况以及受到干旱胁迫程度的大小[29].有研究表明柠条干旱月份水势有明显降低,水势大小和环境水分状况相关联[30].在本研究中,5~30 a梭梭正午、凌晨同化枝水势依次增加,10~30 a梭梭增势明显.随着梭梭林龄增加,其受到的水分胁迫程度越小.根形态结构大小和分布在干旱环境中是影响植物个体水分可用性的关键因素,随着梭梭不断生长,其地下根系网络愈加发达,能够更有效地利用土壤水和地下水,缓解水分胁迫[18,31-33].但是40 a梭梭水势降低,说明其受到的水分胁迫程度增加.水势降低的原因可能为40 a梭梭新生叶片与老化叶片比例远远低于幼龄梭梭,老叶、中叶绿素a、b比例出现异常,致使梭梭叶片生理活性下降明显[34],进一步导致光合作用时对水分的利用效率下降,加剧水分胁迫.植物叶片功能性状活性是植物工厂运行的源动力[35],在极端干旱条件下会改变叶片形态结构和面积[9],本研究结果显示30 a和40 a梭梭叶片面积大幅缩减[36-37](表1).光合作用面积减少一方面有利于减少对水分和能量的需求,协调机体水分平衡,这是对水分胁迫加剧的应对策略.此外光合面积的减少也降低了光合产物的积累量,植物将有限的光合产物优先对根的投入导致地下部分呼吸作用消耗更多光合产物,可能会导致碳饥饿从而导致梭梭树死亡[15,38]. ...
Forecasting climate change impacts to plant community composition in the Sonoran Desert region
1
2011
... IPCC报告显示,21世纪末全球平均气温与1850—1900年的水平相比,可能升高1~1.8 ℃(最佳估算1.4 ℃),并且随着气温持续升高,部分地区最高升温幅度可能达到5.7 ℃[1].随着全球变暖,干旱发生频率和强度增加、影响范围扩大[2],对全球生态系统造成不同程度的影响,荒漠植物的生存将面临严峻挑战[3-4],高温干旱可引发荒漠植物死亡[5].梭梭(Haloxylon ammodendron)具有顽强的生态适应能力,能在极端高温、土壤贫瘠、水分匮乏且高盐碱的地区生存[6-9],防风固沙效果好,广泛分布于中国西北荒漠地区,对维系荒漠-绿洲共生关系作用重大,关乎中国西北干旱区乃至中亚地区生态安全和经济社会的可持续发展[10].作为温度敏感型的荒漠灌木[11],不同年龄的梭梭生理过程对温度的适应策略不同,而这种差异将会对环境呈现出不同的抗逆能力.水分是荒漠区植物生长最主要限制因子,控制着荒漠植物的生长与分布[12].对于荒漠植物而言,为了强化吸水能力,同化枝水势(Ψ)随干旱胁迫加剧而降低[13].另一方面,为了降低水分损耗,梭梭在高温时叶片响应敏感,关闭气孔降低气孔导度(Gs)[14].为应对高温缺水环境,梭梭会优先投入于根系生长,攫取水分和营养[15-16].梭梭水分利用效率(WUE)也体现其对干旱高温环境胁迫的应变机制[17].但不同林龄梭梭植株大小、根系、组织发育、组织生理特性和光合面积的差异[18],预示着对极端高温的气体交换和水分利用等策略会有较大差异.在全球温度持续升高的背景下,探究梭梭气体交换对极端高温适应策略,有助于预测未来不同林龄梭梭在极端高温下的生存境况[19]. ...
Physiological responses of Haloxylon ammodendron to rainfall pulses in temperate desert regions,Northwestern China
1
2014
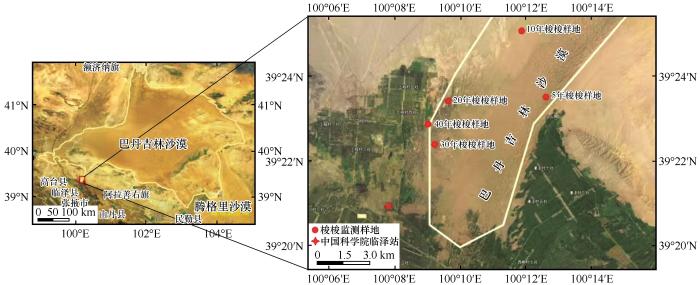
... 本研究于2023年7—9月在中国科学院临泽内陆河流域研究站(39°21′N、100°07′E,海拔1 381 m,图1)东北方向的固定半固定沙区进行.该区域属于巴丹吉林沙漠南缘,广泛种植有1~50年林龄的梭梭,本研究选取5、10、20、30、40 a林龄梭梭样地,调查梭梭的高度、基茎、枯死率和冠幅(表1).该区域属于典型的内陆性中温带干旱气候,干燥、多大风、太阳辐射强烈、昼夜温差大,年平均气温7.6 ℃,最高气温出现在7月(39.1 ℃),最低气温出现在1月(-27.3 ℃).年降水量为110.4 mm,降雨集中在7—9月,占全年降水量的56.7%~62.6%.年蒸发量为1 900~2 088 mm,年日照时数3 051 h,年无霜期165 d,相对湿度46%,常年以西北风为主,风沙活动强烈,年平均风速为3.2 m·s-1,最大风速21 m·s-1[20]. ...
模拟增温对梭梭光合生理生态特征的影响
1
2017
... Pn不仅受温度[21-22]和湿度等外界环境变化影响,也受蒸腾作用和气孔开放等生理过程影响[23](表2).不同林龄梭梭Pn随温度升高有较大差异,5 a梭梭整体最小(图2),对应的同化枝水势最低(图4),受到的水分胁迫最严重,研究结果表明环境可利用有效水分较少会限制植物光合作用[9,24].5 a梭梭Gs和Tr(图3、4)均小于其余林龄梭梭,虽然较小Gs和Tr能有效降低水分散失,但是也阻碍了二氧化碳的获取,导致5 a梭梭Pn整体最小.此外各个林龄梭梭在25~35 ℃下Pn波动幅度比35~40 ℃大,说明水分胁迫在极端高温时对梭梭的影响较大. ...
民勤绿洲荒漠过渡带植物白刺和梭梭光合特性
1
2005
... Pn不仅受温度[21-22]和湿度等外界环境变化影响,也受蒸腾作用和气孔开放等生理过程影响[23](表2).不同林龄梭梭Pn随温度升高有较大差异,5 a梭梭整体最小(图2),对应的同化枝水势最低(图4),受到的水分胁迫最严重,研究结果表明环境可利用有效水分较少会限制植物光合作用[9,24].5 a梭梭Gs和Tr(图3、4)均小于其余林龄梭梭,虽然较小Gs和Tr能有效降低水分散失,但是也阻碍了二氧化碳的获取,导致5 a梭梭Pn整体最小.此外各个林龄梭梭在25~35 ℃下Pn波动幅度比35~40 ℃大,说明水分胁迫在极端高温时对梭梭的影响较大. ...
干旱区3种典型灌木光合特性及其对环境因子的响应
1
2021
... Pn不仅受温度[21-22]和湿度等外界环境变化影响,也受蒸腾作用和气孔开放等生理过程影响[23](表2).不同林龄梭梭Pn随温度升高有较大差异,5 a梭梭整体最小(图2),对应的同化枝水势最低(图4),受到的水分胁迫最严重,研究结果表明环境可利用有效水分较少会限制植物光合作用[9,24].5 a梭梭Gs和Tr(图3、4)均小于其余林龄梭梭,虽然较小Gs和Tr能有效降低水分散失,但是也阻碍了二氧化碳的获取,导致5 a梭梭Pn整体最小.此外各个林龄梭梭在25~35 ℃下Pn波动幅度比35~40 ℃大,说明水分胁迫在极端高温时对梭梭的影响较大. ...
Variations in water use strategies of sand-binding vegetation along a precipitation gradient in sandy regions,northern China
2
2021
... Pn不仅受温度[21-22]和湿度等外界环境变化影响,也受蒸腾作用和气孔开放等生理过程影响[23](表2).不同林龄梭梭Pn随温度升高有较大差异,5 a梭梭整体最小(图2),对应的同化枝水势最低(图4),受到的水分胁迫最严重,研究结果表明环境可利用有效水分较少会限制植物光合作用[9,24].5 a梭梭Gs和Tr(图3、4)均小于其余林龄梭梭,虽然较小Gs和Tr能有效降低水分散失,但是也阻碍了二氧化碳的获取,导致5 a梭梭Pn整体最小.此外各个林龄梭梭在25~35 ℃下Pn波动幅度比35~40 ℃大,说明水分胁迫在极端高温时对梭梭的影响较大. ...
... 水分是限制荒漠植物梭梭生长发育分布的关键因素[24,27-28],梭梭同化枝水势是显示植物体内外水分状态的重要指标,通过测量正午和凌晨水势可以判断植物水分亏缺情况以及受到干旱胁迫程度的大小[29].有研究表明柠条干旱月份水势有明显降低,水势大小和环境水分状况相关联[30].在本研究中,5~30 a梭梭正午、凌晨同化枝水势依次增加,10~30 a梭梭增势明显.随着梭梭林龄增加,其受到的水分胁迫程度越小.根形态结构大小和分布在干旱环境中是影响植物个体水分可用性的关键因素,随着梭梭不断生长,其地下根系网络愈加发达,能够更有效地利用土壤水和地下水,缓解水分胁迫[18,31-33].但是40 a梭梭水势降低,说明其受到的水分胁迫程度增加.水势降低的原因可能为40 a梭梭新生叶片与老化叶片比例远远低于幼龄梭梭,老叶、中叶绿素a、b比例出现异常,致使梭梭叶片生理活性下降明显[34],进一步导致光合作用时对水分的利用效率下降,加剧水分胁迫.植物叶片功能性状活性是植物工厂运行的源动力[35],在极端干旱条件下会改变叶片形态结构和面积[9],本研究结果显示30 a和40 a梭梭叶片面积大幅缩减[36-37](表1).光合作用面积减少一方面有利于减少对水分和能量的需求,协调机体水分平衡,这是对水分胁迫加剧的应对策略.此外光合面积的减少也降低了光合产物的积累量,植物将有限的光合产物优先对根的投入导致地下部分呼吸作用消耗更多光合产物,可能会导致碳饥饿从而导致梭梭树死亡[15,38]. ...
干旱和高温对油菜叶片光合作用和叶绿素荧光特性的影响
1
2017
... 5 a 和40 a梭梭Pn温度响应曲线变化趋势相似,Pn随温度增加先上升后下降且都在35 ℃时达到峰值,说明两个林龄梭梭应对极端高温的策略比较保守,在极端高温时通过降低Pn维持机体系统机能保持稳定.当温度过高时会影响与光合作用有关的核酮糖1,5-二磷酸核酮糖羧化酶/加氧酶Rubisco、磷酸烯醇式丙酮酸羧化酶(PEPC)和苹果酸脱氢酶(NADP-MDH)等相关酶活性,减弱Pn.还可能破坏光系统Ⅱ生理结构(PSⅡ),这种破坏是不可逆的[25-26].5 a梭梭WUE最大(图4),较高的WUE可以略微缓解水分胁迫对Pn的影响.降低水势和Pn,并且保持较高WUE是5 a梭梭应对极端高温和水分胁迫环境的适应策略,即保守型资源策略.10 a梭梭Pn随温度升高不断升高.可能原因为10 a梭梭根系逐渐发达和器官组织生理功能进一步发展完善,同时植株较小,对水分的需求也较小,受到水分胁迫程度相较于5 a梭梭有所降低(图3),对Pn的限制减弱.其次也表明10 a梭梭对极端高温环境适应能力较好,对资源的利用效率高,生产有机物的能力强,在极端高温下能保持极高Pn且不断升高,能有效积累有机物用于生长发育,属于获取型资源策略.20、30 a梭梭Pn温度响应曲线类似,都随温度升高且增幅较小,在30 ℃后细微波动(图1).20、30 a梭梭光合系统经过对极端高温环境长期的适应,形成了应对极端高温环境的应对机制,即在外界温度波动甚至在极端高温时也能保持较高的Pn且变化不敏感,对极端高温环境有很强的适应能力,属于保守型资源策略. ...
黄瓜幼苗光合作用对高温胁迫的响应与适应
1
2017
... 5 a 和40 a梭梭Pn温度响应曲线变化趋势相似,Pn随温度增加先上升后下降且都在35 ℃时达到峰值,说明两个林龄梭梭应对极端高温的策略比较保守,在极端高温时通过降低Pn维持机体系统机能保持稳定.当温度过高时会影响与光合作用有关的核酮糖1,5-二磷酸核酮糖羧化酶/加氧酶Rubisco、磷酸烯醇式丙酮酸羧化酶(PEPC)和苹果酸脱氢酶(NADP-MDH)等相关酶活性,减弱Pn.还可能破坏光系统Ⅱ生理结构(PSⅡ),这种破坏是不可逆的[25-26].5 a梭梭WUE最大(图4),较高的WUE可以略微缓解水分胁迫对Pn的影响.降低水势和Pn,并且保持较高WUE是5 a梭梭应对极端高温和水分胁迫环境的适应策略,即保守型资源策略.10 a梭梭Pn随温度升高不断升高.可能原因为10 a梭梭根系逐渐发达和器官组织生理功能进一步发展完善,同时植株较小,对水分的需求也较小,受到水分胁迫程度相较于5 a梭梭有所降低(图3),对Pn的限制减弱.其次也表明10 a梭梭对极端高温环境适应能力较好,对资源的利用效率高,生产有机物的能力强,在极端高温下能保持极高Pn且不断升高,能有效积累有机物用于生长发育,属于获取型资源策略.20、30 a梭梭Pn温度响应曲线类似,都随温度升高且增幅较小,在30 ℃后细微波动(图1).20、30 a梭梭光合系统经过对极端高温环境长期的适应,形成了应对极端高温环境的应对机制,即在外界温度波动甚至在极端高温时也能保持较高的Pn且变化不敏感,对极端高温环境有很强的适应能力,属于保守型资源策略. ...
Canopy and leaf gas exchange of Haloxylon ammodendron under different soil moisture regimes
1
2010
... 水分是限制荒漠植物梭梭生长发育分布的关键因素[24,27-28],梭梭同化枝水势是显示植物体内外水分状态的重要指标,通过测量正午和凌晨水势可以判断植物水分亏缺情况以及受到干旱胁迫程度的大小[29].有研究表明柠条干旱月份水势有明显降低,水势大小和环境水分状况相关联[30].在本研究中,5~30 a梭梭正午、凌晨同化枝水势依次增加,10~30 a梭梭增势明显.随着梭梭林龄增加,其受到的水分胁迫程度越小.根形态结构大小和分布在干旱环境中是影响植物个体水分可用性的关键因素,随着梭梭不断生长,其地下根系网络愈加发达,能够更有效地利用土壤水和地下水,缓解水分胁迫[18,31-33].但是40 a梭梭水势降低,说明其受到的水分胁迫程度增加.水势降低的原因可能为40 a梭梭新生叶片与老化叶片比例远远低于幼龄梭梭,老叶、中叶绿素a、b比例出现异常,致使梭梭叶片生理活性下降明显[34],进一步导致光合作用时对水分的利用效率下降,加剧水分胁迫.植物叶片功能性状活性是植物工厂运行的源动力[35],在极端干旱条件下会改变叶片形态结构和面积[9],本研究结果显示30 a和40 a梭梭叶片面积大幅缩减[36-37](表1).光合作用面积减少一方面有利于减少对水分和能量的需求,协调机体水分平衡,这是对水分胁迫加剧的应对策略.此外光合面积的减少也降低了光合产物的积累量,植物将有限的光合产物优先对根的投入导致地下部分呼吸作用消耗更多光合产物,可能会导致碳饥饿从而导致梭梭树死亡[15,38]. ...
The relationships between grasslands and soil moisture on the Loess Plateau of China:a review
1
2016
... 水分是限制荒漠植物梭梭生长发育分布的关键因素[24,27-28],梭梭同化枝水势是显示植物体内外水分状态的重要指标,通过测量正午和凌晨水势可以判断植物水分亏缺情况以及受到干旱胁迫程度的大小[29].有研究表明柠条干旱月份水势有明显降低,水势大小和环境水分状况相关联[30].在本研究中,5~30 a梭梭正午、凌晨同化枝水势依次增加,10~30 a梭梭增势明显.随着梭梭林龄增加,其受到的水分胁迫程度越小.根形态结构大小和分布在干旱环境中是影响植物个体水分可用性的关键因素,随着梭梭不断生长,其地下根系网络愈加发达,能够更有效地利用土壤水和地下水,缓解水分胁迫[18,31-33].但是40 a梭梭水势降低,说明其受到的水分胁迫程度增加.水势降低的原因可能为40 a梭梭新生叶片与老化叶片比例远远低于幼龄梭梭,老叶、中叶绿素a、b比例出现异常,致使梭梭叶片生理活性下降明显[34],进一步导致光合作用时对水分的利用效率下降,加剧水分胁迫.植物叶片功能性状活性是植物工厂运行的源动力[35],在极端干旱条件下会改变叶片形态结构和面积[9],本研究结果显示30 a和40 a梭梭叶片面积大幅缩减[36-37](表1).光合作用面积减少一方面有利于减少对水分和能量的需求,协调机体水分平衡,这是对水分胁迫加剧的应对策略.此外光合面积的减少也降低了光合产物的积累量,植物将有限的光合产物优先对根的投入导致地下部分呼吸作用消耗更多光合产物,可能会导致碳饥饿从而导致梭梭树死亡[15,38]. ...
地下水埋深对幼龄梭梭功能性状的影响
1
2022
... 水分是限制荒漠植物梭梭生长发育分布的关键因素[24,27-28],梭梭同化枝水势是显示植物体内外水分状态的重要指标,通过测量正午和凌晨水势可以判断植物水分亏缺情况以及受到干旱胁迫程度的大小[29].有研究表明柠条干旱月份水势有明显降低,水势大小和环境水分状况相关联[30].在本研究中,5~30 a梭梭正午、凌晨同化枝水势依次增加,10~30 a梭梭增势明显.随着梭梭林龄增加,其受到的水分胁迫程度越小.根形态结构大小和分布在干旱环境中是影响植物个体水分可用性的关键因素,随着梭梭不断生长,其地下根系网络愈加发达,能够更有效地利用土壤水和地下水,缓解水分胁迫[18,31-33].但是40 a梭梭水势降低,说明其受到的水分胁迫程度增加.水势降低的原因可能为40 a梭梭新生叶片与老化叶片比例远远低于幼龄梭梭,老叶、中叶绿素a、b比例出现异常,致使梭梭叶片生理活性下降明显[34],进一步导致光合作用时对水分的利用效率下降,加剧水分胁迫.植物叶片功能性状活性是植物工厂运行的源动力[35],在极端干旱条件下会改变叶片形态结构和面积[9],本研究结果显示30 a和40 a梭梭叶片面积大幅缩减[36-37](表1).光合作用面积减少一方面有利于减少对水分和能量的需求,协调机体水分平衡,这是对水分胁迫加剧的应对策略.此外光合面积的减少也降低了光合产物的积累量,植物将有限的光合产物优先对根的投入导致地下部分呼吸作用消耗更多光合产物,可能会导致碳饥饿从而导致梭梭树死亡[15,38]. ...
不同林龄柠条(Caragana korshinskii)的光合特性和水分利用特征
1
2016
... 水分是限制荒漠植物梭梭生长发育分布的关键因素[24,27-28],梭梭同化枝水势是显示植物体内外水分状态的重要指标,通过测量正午和凌晨水势可以判断植物水分亏缺情况以及受到干旱胁迫程度的大小[29].有研究表明柠条干旱月份水势有明显降低,水势大小和环境水分状况相关联[30].在本研究中,5~30 a梭梭正午、凌晨同化枝水势依次增加,10~30 a梭梭增势明显.随着梭梭林龄增加,其受到的水分胁迫程度越小.根形态结构大小和分布在干旱环境中是影响植物个体水分可用性的关键因素,随着梭梭不断生长,其地下根系网络愈加发达,能够更有效地利用土壤水和地下水,缓解水分胁迫[18,31-33].但是40 a梭梭水势降低,说明其受到的水分胁迫程度增加.水势降低的原因可能为40 a梭梭新生叶片与老化叶片比例远远低于幼龄梭梭,老叶、中叶绿素a、b比例出现异常,致使梭梭叶片生理活性下降明显[34],进一步导致光合作用时对水分的利用效率下降,加剧水分胁迫.植物叶片功能性状活性是植物工厂运行的源动力[35],在极端干旱条件下会改变叶片形态结构和面积[9],本研究结果显示30 a和40 a梭梭叶片面积大幅缩减[36-37](表1).光合作用面积减少一方面有利于减少对水分和能量的需求,协调机体水分平衡,这是对水分胁迫加剧的应对策略.此外光合面积的减少也降低了光合产物的积累量,植物将有限的光合产物优先对根的投入导致地下部分呼吸作用消耗更多光合产物,可能会导致碳饥饿从而导致梭梭树死亡[15,38]. ...
三种荒漠灌木根系的构形特征与叶性因子对干旱生境的适应性比较
1
2008
... 水分是限制荒漠植物梭梭生长发育分布的关键因素[24,27-28],梭梭同化枝水势是显示植物体内外水分状态的重要指标,通过测量正午和凌晨水势可以判断植物水分亏缺情况以及受到干旱胁迫程度的大小[29].有研究表明柠条干旱月份水势有明显降低,水势大小和环境水分状况相关联[30].在本研究中,5~30 a梭梭正午、凌晨同化枝水势依次增加,10~30 a梭梭增势明显.随着梭梭林龄增加,其受到的水分胁迫程度越小.根形态结构大小和分布在干旱环境中是影响植物个体水分可用性的关键因素,随着梭梭不断生长,其地下根系网络愈加发达,能够更有效地利用土壤水和地下水,缓解水分胁迫[18,31-33].但是40 a梭梭水势降低,说明其受到的水分胁迫程度增加.水势降低的原因可能为40 a梭梭新生叶片与老化叶片比例远远低于幼龄梭梭,老叶、中叶绿素a、b比例出现异常,致使梭梭叶片生理活性下降明显[34],进一步导致光合作用时对水分的利用效率下降,加剧水分胁迫.植物叶片功能性状活性是植物工厂运行的源动力[35],在极端干旱条件下会改变叶片形态结构和面积[9],本研究结果显示30 a和40 a梭梭叶片面积大幅缩减[36-37](表1).光合作用面积减少一方面有利于减少对水分和能量的需求,协调机体水分平衡,这是对水分胁迫加剧的应对策略.此外光合面积的减少也降低了光合产物的积累量,植物将有限的光合产物优先对根的投入导致地下部分呼吸作用消耗更多光合产物,可能会导致碳饥饿从而导致梭梭树死亡[15,38]. ...
Hydrologic regulation of plant rooting depth
0
2017
The eco-hydrological threshold for evaluating the stability of sand-binding vegetation in different climatic zones
1
2017
... 水分是限制荒漠植物梭梭生长发育分布的关键因素[24,27-28],梭梭同化枝水势是显示植物体内外水分状态的重要指标,通过测量正午和凌晨水势可以判断植物水分亏缺情况以及受到干旱胁迫程度的大小[29].有研究表明柠条干旱月份水势有明显降低,水势大小和环境水分状况相关联[30].在本研究中,5~30 a梭梭正午、凌晨同化枝水势依次增加,10~30 a梭梭增势明显.随着梭梭林龄增加,其受到的水分胁迫程度越小.根形态结构大小和分布在干旱环境中是影响植物个体水分可用性的关键因素,随着梭梭不断生长,其地下根系网络愈加发达,能够更有效地利用土壤水和地下水,缓解水分胁迫[18,31-33].但是40 a梭梭水势降低,说明其受到的水分胁迫程度增加.水势降低的原因可能为40 a梭梭新生叶片与老化叶片比例远远低于幼龄梭梭,老叶、中叶绿素a、b比例出现异常,致使梭梭叶片生理活性下降明显[34],进一步导致光合作用时对水分的利用效率下降,加剧水分胁迫.植物叶片功能性状活性是植物工厂运行的源动力[35],在极端干旱条件下会改变叶片形态结构和面积[9],本研究结果显示30 a和40 a梭梭叶片面积大幅缩减[36-37](表1).光合作用面积减少一方面有利于减少对水分和能量的需求,协调机体水分平衡,这是对水分胁迫加剧的应对策略.此外光合面积的减少也降低了光合产物的积累量,植物将有限的光合产物优先对根的投入导致地下部分呼吸作用消耗更多光合产物,可能会导致碳饥饿从而导致梭梭树死亡[15,38]. ...
荒漠绿洲过渡带不同年限雨养梭梭(Haloxylon ammodendron)对土壤水分变化的响应
1
2021
... 水分是限制荒漠植物梭梭生长发育分布的关键因素[24,27-28],梭梭同化枝水势是显示植物体内外水分状态的重要指标,通过测量正午和凌晨水势可以判断植物水分亏缺情况以及受到干旱胁迫程度的大小[29].有研究表明柠条干旱月份水势有明显降低,水势大小和环境水分状况相关联[30].在本研究中,5~30 a梭梭正午、凌晨同化枝水势依次增加,10~30 a梭梭增势明显.随着梭梭林龄增加,其受到的水分胁迫程度越小.根形态结构大小和分布在干旱环境中是影响植物个体水分可用性的关键因素,随着梭梭不断生长,其地下根系网络愈加发达,能够更有效地利用土壤水和地下水,缓解水分胁迫[18,31-33].但是40 a梭梭水势降低,说明其受到的水分胁迫程度增加.水势降低的原因可能为40 a梭梭新生叶片与老化叶片比例远远低于幼龄梭梭,老叶、中叶绿素a、b比例出现异常,致使梭梭叶片生理活性下降明显[34],进一步导致光合作用时对水分的利用效率下降,加剧水分胁迫.植物叶片功能性状活性是植物工厂运行的源动力[35],在极端干旱条件下会改变叶片形态结构和面积[9],本研究结果显示30 a和40 a梭梭叶片面积大幅缩减[36-37](表1).光合作用面积减少一方面有利于减少对水分和能量的需求,协调机体水分平衡,这是对水分胁迫加剧的应对策略.此外光合面积的减少也降低了光合产物的积累量,植物将有限的光合产物优先对根的投入导致地下部分呼吸作用消耗更多光合产物,可能会导致碳饥饿从而导致梭梭树死亡[15,38]. ...
植物叶片水力与经济性状权衡关系的研究进展
1
2015
... 水分是限制荒漠植物梭梭生长发育分布的关键因素[24,27-28],梭梭同化枝水势是显示植物体内外水分状态的重要指标,通过测量正午和凌晨水势可以判断植物水分亏缺情况以及受到干旱胁迫程度的大小[29].有研究表明柠条干旱月份水势有明显降低,水势大小和环境水分状况相关联[30].在本研究中,5~30 a梭梭正午、凌晨同化枝水势依次增加,10~30 a梭梭增势明显.随着梭梭林龄增加,其受到的水分胁迫程度越小.根形态结构大小和分布在干旱环境中是影响植物个体水分可用性的关键因素,随着梭梭不断生长,其地下根系网络愈加发达,能够更有效地利用土壤水和地下水,缓解水分胁迫[18,31-33].但是40 a梭梭水势降低,说明其受到的水分胁迫程度增加.水势降低的原因可能为40 a梭梭新生叶片与老化叶片比例远远低于幼龄梭梭,老叶、中叶绿素a、b比例出现异常,致使梭梭叶片生理活性下降明显[34],进一步导致光合作用时对水分的利用效率下降,加剧水分胁迫.植物叶片功能性状活性是植物工厂运行的源动力[35],在极端干旱条件下会改变叶片形态结构和面积[9],本研究结果显示30 a和40 a梭梭叶片面积大幅缩减[36-37](表1).光合作用面积减少一方面有利于减少对水分和能量的需求,协调机体水分平衡,这是对水分胁迫加剧的应对策略.此外光合面积的减少也降低了光合产物的积累量,植物将有限的光合产物优先对根的投入导致地下部分呼吸作用消耗更多光合产物,可能会导致碳饥饿从而导致梭梭树死亡[15,38]. ...
Within-crown plasticity of hydraulic properties influence branch dieback patterns of two woody plants under experimental drought conditions
1
2023
... 水分是限制荒漠植物梭梭生长发育分布的关键因素[24,27-28],梭梭同化枝水势是显示植物体内外水分状态的重要指标,通过测量正午和凌晨水势可以判断植物水分亏缺情况以及受到干旱胁迫程度的大小[29].有研究表明柠条干旱月份水势有明显降低,水势大小和环境水分状况相关联[30].在本研究中,5~30 a梭梭正午、凌晨同化枝水势依次增加,10~30 a梭梭增势明显.随着梭梭林龄增加,其受到的水分胁迫程度越小.根形态结构大小和分布在干旱环境中是影响植物个体水分可用性的关键因素,随着梭梭不断生长,其地下根系网络愈加发达,能够更有效地利用土壤水和地下水,缓解水分胁迫[18,31-33].但是40 a梭梭水势降低,说明其受到的水分胁迫程度增加.水势降低的原因可能为40 a梭梭新生叶片与老化叶片比例远远低于幼龄梭梭,老叶、中叶绿素a、b比例出现异常,致使梭梭叶片生理活性下降明显[34],进一步导致光合作用时对水分的利用效率下降,加剧水分胁迫.植物叶片功能性状活性是植物工厂运行的源动力[35],在极端干旱条件下会改变叶片形态结构和面积[9],本研究结果显示30 a和40 a梭梭叶片面积大幅缩减[36-37](表1).光合作用面积减少一方面有利于减少对水分和能量的需求,协调机体水分平衡,这是对水分胁迫加剧的应对策略.此外光合面积的减少也降低了光合产物的积累量,植物将有限的光合产物优先对根的投入导致地下部分呼吸作用消耗更多光合产物,可能会导致碳饥饿从而导致梭梭树死亡[15,38]. ...
Leaf chlorophyll parameters and photosynthetic characteristic variations with stand age in a typical desert species (Haloxylon ammodendron)
1
2022
... 水分是限制荒漠植物梭梭生长发育分布的关键因素[24,27-28],梭梭同化枝水势是显示植物体内外水分状态的重要指标,通过测量正午和凌晨水势可以判断植物水分亏缺情况以及受到干旱胁迫程度的大小[29].有研究表明柠条干旱月份水势有明显降低,水势大小和环境水分状况相关联[30].在本研究中,5~30 a梭梭正午、凌晨同化枝水势依次增加,10~30 a梭梭增势明显.随着梭梭林龄增加,其受到的水分胁迫程度越小.根形态结构大小和分布在干旱环境中是影响植物个体水分可用性的关键因素,随着梭梭不断生长,其地下根系网络愈加发达,能够更有效地利用土壤水和地下水,缓解水分胁迫[18,31-33].但是40 a梭梭水势降低,说明其受到的水分胁迫程度增加.水势降低的原因可能为40 a梭梭新生叶片与老化叶片比例远远低于幼龄梭梭,老叶、中叶绿素a、b比例出现异常,致使梭梭叶片生理活性下降明显[34],进一步导致光合作用时对水分的利用效率下降,加剧水分胁迫.植物叶片功能性状活性是植物工厂运行的源动力[35],在极端干旱条件下会改变叶片形态结构和面积[9],本研究结果显示30 a和40 a梭梭叶片面积大幅缩减[36-37](表1).光合作用面积减少一方面有利于减少对水分和能量的需求,协调机体水分平衡,这是对水分胁迫加剧的应对策略.此外光合面积的减少也降低了光合产物的积累量,植物将有限的光合产物优先对根的投入导致地下部分呼吸作用消耗更多光合产物,可能会导致碳饥饿从而导致梭梭树死亡[15,38]. ...
Varying responses of two Haloxylon species to extreme drought and groundwater depth
1
2019
... 水分是限制荒漠植物梭梭生长发育分布的关键因素[24,27-28],梭梭同化枝水势是显示植物体内外水分状态的重要指标,通过测量正午和凌晨水势可以判断植物水分亏缺情况以及受到干旱胁迫程度的大小[29].有研究表明柠条干旱月份水势有明显降低,水势大小和环境水分状况相关联[30].在本研究中,5~30 a梭梭正午、凌晨同化枝水势依次增加,10~30 a梭梭增势明显.随着梭梭林龄增加,其受到的水分胁迫程度越小.根形态结构大小和分布在干旱环境中是影响植物个体水分可用性的关键因素,随着梭梭不断生长,其地下根系网络愈加发达,能够更有效地利用土壤水和地下水,缓解水分胁迫[18,31-33].但是40 a梭梭水势降低,说明其受到的水分胁迫程度增加.水势降低的原因可能为40 a梭梭新生叶片与老化叶片比例远远低于幼龄梭梭,老叶、中叶绿素a、b比例出现异常,致使梭梭叶片生理活性下降明显[34],进一步导致光合作用时对水分的利用效率下降,加剧水分胁迫.植物叶片功能性状活性是植物工厂运行的源动力[35],在极端干旱条件下会改变叶片形态结构和面积[9],本研究结果显示30 a和40 a梭梭叶片面积大幅缩减[36-37](表1).光合作用面积减少一方面有利于减少对水分和能量的需求,协调机体水分平衡,这是对水分胁迫加剧的应对策略.此外光合面积的减少也降低了光合产物的积累量,植物将有限的光合产物优先对根的投入导致地下部分呼吸作用消耗更多光合产物,可能会导致碳饥饿从而导致梭梭树死亡[15,38]. ...
Does homeostasis or disturbance of homeostasis in minimum leaf water potential explain the isohydric versus anisohydric behavior of Vitis vinifera L.cultivars?
1
2012
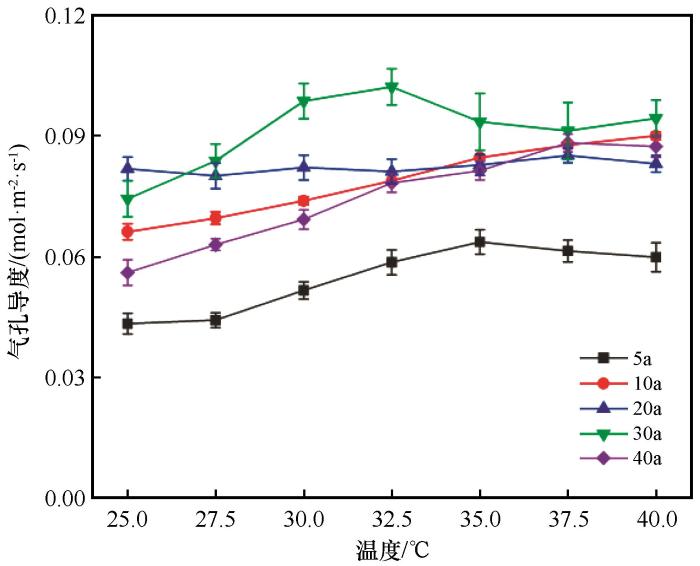
... 植物对不同气候与土壤水分条件的长期适应会形成由一整套相关联的性状组成的水分调节策略.本研究中,不同林龄梭梭Gs与Tr温度响应曲线有着较大差异,Gs表示植物气孔的开放程度,是显示植物对环境适应能力大小的重要生理参数,其大小波动受到环境温度、湿度、VPD[39]和不同植物适应机制等因素影响.它通过影响植物与外界气体和水分交换[40]等过程直接或间接对植物光合、呼吸作用、Tr和WUE等造成影响.在本研究结果中(图3),随着温度升高,不同林龄梭梭Gs整体都有增加,这与已有研究结果一致[41].气孔开放程度与蒸腾作用紧密关联(表2),当Gs随温度上升增加时,Tr下降(图4),这与前人研究结果一致[42].各个林龄Gs在35~40 ℃(极端高温)波动较小,但是30 a和40 a梭梭Tr相比于温度较低时却有着较大增幅,同时Pn波动却不大,表明梭梭在极端高温时Tr增加的原因不是为了加快水分运输速率从而增强Pn,而是通过蒸腾作用从而降低叶片温度避免叶片受到高温侵害以保持原有的Pn;其次在极端高温时两个林龄梭梭WUE都有降低(图4),Tr的提高是为了补偿因WUE降低所导致供应光合作用而减少的水分和养分.20 a梭梭Gs和Tr温度响应曲线波动在不同林龄梭梭中最小,甚至在极端高温时波动也小,对极端高温和水分胁迫环境形成了稳定的适应机制,对环境变化不敏感.5 a和10 a梭梭Gs和Tr温度响应曲线在25~35 ℃波动幅度比35~40 ℃时大且是上升趋势,当温度在适宜温度范围内升高时能促进光合作用,在极端高温时则较为保守,这样可以有效防止水分散失维持机体水分平衡.此外10 a梭梭在极端高温时也保持着Pn的快速增长,极端高温能促进蒸腾效率从而维持Pn增长. ...
最优气孔行为理论和气孔导度模拟
1
2016
... 植物对不同气候与土壤水分条件的长期适应会形成由一整套相关联的性状组成的水分调节策略.本研究中,不同林龄梭梭Gs与Tr温度响应曲线有着较大差异,Gs表示植物气孔的开放程度,是显示植物对环境适应能力大小的重要生理参数,其大小波动受到环境温度、湿度、VPD[39]和不同植物适应机制等因素影响.它通过影响植物与外界气体和水分交换[40]等过程直接或间接对植物光合、呼吸作用、Tr和WUE等造成影响.在本研究结果中(图3),随着温度升高,不同林龄梭梭Gs整体都有增加,这与已有研究结果一致[41].气孔开放程度与蒸腾作用紧密关联(表2),当Gs随温度上升增加时,Tr下降(图4),这与前人研究结果一致[42].各个林龄Gs在35~40 ℃(极端高温)波动较小,但是30 a和40 a梭梭Tr相比于温度较低时却有着较大增幅,同时Pn波动却不大,表明梭梭在极端高温时Tr增加的原因不是为了加快水分运输速率从而增强Pn,而是通过蒸腾作用从而降低叶片温度避免叶片受到高温侵害以保持原有的Pn;其次在极端高温时两个林龄梭梭WUE都有降低(图4),Tr的提高是为了补偿因WUE降低所导致供应光合作用而减少的水分和养分.20 a梭梭Gs和Tr温度响应曲线波动在不同林龄梭梭中最小,甚至在极端高温时波动也小,对极端高温和水分胁迫环境形成了稳定的适应机制,对环境变化不敏感.5 a和10 a梭梭Gs和Tr温度响应曲线在25~35 ℃波动幅度比35~40 ℃时大且是上升趋势,当温度在适宜温度范围内升高时能促进光合作用,在极端高温时则较为保守,这样可以有效防止水分散失维持机体水分平衡.此外10 a梭梭在极端高温时也保持着Pn的快速增长,极端高温能促进蒸腾效率从而维持Pn增长. ...
干旱区荒漠-绿洲过渡带3种典型灌木气孔导度对环境变化的响应及其对蒸腾的调控
1
2021
... 植物对不同气候与土壤水分条件的长期适应会形成由一整套相关联的性状组成的水分调节策略.本研究中,不同林龄梭梭Gs与Tr温度响应曲线有着较大差异,Gs表示植物气孔的开放程度,是显示植物对环境适应能力大小的重要生理参数,其大小波动受到环境温度、湿度、VPD[39]和不同植物适应机制等因素影响.它通过影响植物与外界气体和水分交换[40]等过程直接或间接对植物光合、呼吸作用、Tr和WUE等造成影响.在本研究结果中(图3),随着温度升高,不同林龄梭梭Gs整体都有增加,这与已有研究结果一致[41].气孔开放程度与蒸腾作用紧密关联(表2),当Gs随温度上升增加时,Tr下降(图4),这与前人研究结果一致[42].各个林龄Gs在35~40 ℃(极端高温)波动较小,但是30 a和40 a梭梭Tr相比于温度较低时却有着较大增幅,同时Pn波动却不大,表明梭梭在极端高温时Tr增加的原因不是为了加快水分运输速率从而增强Pn,而是通过蒸腾作用从而降低叶片温度避免叶片受到高温侵害以保持原有的Pn;其次在极端高温时两个林龄梭梭WUE都有降低(图4),Tr的提高是为了补偿因WUE降低所导致供应光合作用而减少的水分和养分.20 a梭梭Gs和Tr温度响应曲线波动在不同林龄梭梭中最小,甚至在极端高温时波动也小,对极端高温和水分胁迫环境形成了稳定的适应机制,对环境变化不敏感.5 a和10 a梭梭Gs和Tr温度响应曲线在25~35 ℃波动幅度比35~40 ℃时大且是上升趋势,当温度在适宜温度范围内升高时能促进光合作用,在极端高温时则较为保守,这样可以有效防止水分散失维持机体水分平衡.此外10 a梭梭在极端高温时也保持着Pn的快速增长,极端高温能促进蒸腾效率从而维持Pn增长. ...
三种经验模型模拟荒漠河岸柽柳叶片气孔导度
1
2020
... 植物对不同气候与土壤水分条件的长期适应会形成由一整套相关联的性状组成的水分调节策略.本研究中,不同林龄梭梭Gs与Tr温度响应曲线有着较大差异,Gs表示植物气孔的开放程度,是显示植物对环境适应能力大小的重要生理参数,其大小波动受到环境温度、湿度、VPD[39]和不同植物适应机制等因素影响.它通过影响植物与外界气体和水分交换[40]等过程直接或间接对植物光合、呼吸作用、Tr和WUE等造成影响.在本研究结果中(图3),随着温度升高,不同林龄梭梭Gs整体都有增加,这与已有研究结果一致[41].气孔开放程度与蒸腾作用紧密关联(表2),当Gs随温度上升增加时,Tr下降(图4),这与前人研究结果一致[42].各个林龄Gs在35~40 ℃(极端高温)波动较小,但是30 a和40 a梭梭Tr相比于温度较低时却有着较大增幅,同时Pn波动却不大,表明梭梭在极端高温时Tr增加的原因不是为了加快水分运输速率从而增强Pn,而是通过蒸腾作用从而降低叶片温度避免叶片受到高温侵害以保持原有的Pn;其次在极端高温时两个林龄梭梭WUE都有降低(图4),Tr的提高是为了补偿因WUE降低所导致供应光合作用而减少的水分和养分.20 a梭梭Gs和Tr温度响应曲线波动在不同林龄梭梭中最小,甚至在极端高温时波动也小,对极端高温和水分胁迫环境形成了稳定的适应机制,对环境变化不敏感.5 a和10 a梭梭Gs和Tr温度响应曲线在25~35 ℃波动幅度比35~40 ℃时大且是上升趋势,当温度在适宜温度范围内升高时能促进光合作用,在极端高温时则较为保守,这样可以有效防止水分散失维持机体水分平衡.此外10 a梭梭在极端高温时也保持着Pn的快速增长,极端高温能促进蒸腾效率从而维持Pn增长. ...
不同生育期玉米叶片光合特性及水分利用效率对水分胁迫的响应
1
2015
... WUE是衡量植物吸收水分利用效率的指标,与光合作用、Gs、Tr和温度等因素相关[43-44],Gs、Tr增加在增强水分运输能力的同时也会致使水分损耗增大,而温度会影响气孔开放继而影响Tr(表2).植物调整WUE是关键的水分调节策略,特别是在极端高温的荒漠环境中,这对植物的生存至关重要.本研究中除40 a梭梭外,不同林龄梭梭WUE随林龄增加而降低(图4),这与已有学者研究结果不完全一致[45],原因可能为本研究区气候环境条件和其研究区域差异较大.除了10 a梭梭外,其余不同生长林龄梭梭WUE和Gs随温度升高变化趋势相反,且在Gs最低时WUE达到最大值,说明只有Gs降低,Tr和梭梭用于交换二氧化碳进行光合作用过程受到轻微限制时,WUE才能达到最大值,这与邓雄等[46]对怪柳气体水分交换研究得出的结论一致[47].5 a梭梭WUE最高,10 a次之,30 a最低,与它们受到水分胁迫程度(图4)刚好相反,即受到水分胁迫的梭梭会采取提高WUE的适应策略,侧面表明在干旱缺水的荒漠地区,不同林龄梭梭在受到不同的水分胁迫程度下会严格地做出相应的水分调节策略.较高的WUE有利于5 a和10 a梭梭在极端高温缺水的荒漠环境中降低对水分的损耗,提高光合产物产量,将有机物优先投入地下根系[15]以适应极端高温环境.5 a梭梭WUE随温度升高先增后减,在35 ℃后趋于平缓(图4),这与梭梭Gs、Tr变化趋势刚好相反.5 a梭梭Gs和Tr严格控制着WUE大小,在应对极端高温环境时对水分调节的管控能力强,气体交换各部分水分生理变化联系紧密,光合系统反应灵敏.除40 a梭梭外,其余林龄梭梭WUE在高温时波动较小,10、20 a梭梭响应曲线都较为平稳,表明它们在极端高温时能保持稳定的水分状态,减少不必要的水分损耗同时还能维持较好的光合作用状态.由于40 a梭梭Tr随温度不断升高且在极端高温时增势明显,导致其WUE不断减小.水分利用效率降低会导致梭梭在水资源限制较大的极端高温荒漠环境中生长和发育受到抑制,对极端高温逆境的适应力下降,致使植物的生物量减少,生长速度变慢,甚至可能无法正常完成生命周期. ...
Effects of Ssalinity on stomatal conductance,photosynthetic capacity,and carbon isotope discrimination of salt-tolerant (Gossypium hirsutum L.) and salt-sensitive (Phaseolus vulgaris L.) C(3) Non-halophytes
1
1991
... WUE是衡量植物吸收水分利用效率的指标,与光合作用、Gs、Tr和温度等因素相关[43-44],Gs、Tr增加在增强水分运输能力的同时也会致使水分损耗增大,而温度会影响气孔开放继而影响Tr(表2).植物调整WUE是关键的水分调节策略,特别是在极端高温的荒漠环境中,这对植物的生存至关重要.本研究中除40 a梭梭外,不同林龄梭梭WUE随林龄增加而降低(图4),这与已有学者研究结果不完全一致[45],原因可能为本研究区气候环境条件和其研究区域差异较大.除了10 a梭梭外,其余不同生长林龄梭梭WUE和Gs随温度升高变化趋势相反,且在Gs最低时WUE达到最大值,说明只有Gs降低,Tr和梭梭用于交换二氧化碳进行光合作用过程受到轻微限制时,WUE才能达到最大值,这与邓雄等[46]对怪柳气体水分交换研究得出的结论一致[47].5 a梭梭WUE最高,10 a次之,30 a最低,与它们受到水分胁迫程度(图4)刚好相反,即受到水分胁迫的梭梭会采取提高WUE的适应策略,侧面表明在干旱缺水的荒漠地区,不同林龄梭梭在受到不同的水分胁迫程度下会严格地做出相应的水分调节策略.较高的WUE有利于5 a和10 a梭梭在极端高温缺水的荒漠环境中降低对水分的损耗,提高光合产物产量,将有机物优先投入地下根系[15]以适应极端高温环境.5 a梭梭WUE随温度升高先增后减,在35 ℃后趋于平缓(图4),这与梭梭Gs、Tr变化趋势刚好相反.5 a梭梭Gs和Tr严格控制着WUE大小,在应对极端高温环境时对水分调节的管控能力强,气体交换各部分水分生理变化联系紧密,光合系统反应灵敏.除40 a梭梭外,其余林龄梭梭WUE在高温时波动较小,10、20 a梭梭响应曲线都较为平稳,表明它们在极端高温时能保持稳定的水分状态,减少不必要的水分损耗同时还能维持较好的光合作用状态.由于40 a梭梭Tr随温度不断升高且在极端高温时增势明显,导致其WUE不断减小.水分利用效率降低会导致梭梭在水资源限制较大的极端高温荒漠环境中生长和发育受到抑制,对极端高温逆境的适应力下降,致使植物的生物量减少,生长速度变慢,甚至可能无法正常完成生命周期. ...
太行山南麓不同龄级荆条光合特性及光响应研究
1
2020
... WUE是衡量植物吸收水分利用效率的指标,与光合作用、Gs、Tr和温度等因素相关[43-44],Gs、Tr增加在增强水分运输能力的同时也会致使水分损耗增大,而温度会影响气孔开放继而影响Tr(表2).植物调整WUE是关键的水分调节策略,特别是在极端高温的荒漠环境中,这对植物的生存至关重要.本研究中除40 a梭梭外,不同林龄梭梭WUE随林龄增加而降低(图4),这与已有学者研究结果不完全一致[45],原因可能为本研究区气候环境条件和其研究区域差异较大.除了10 a梭梭外,其余不同生长林龄梭梭WUE和Gs随温度升高变化趋势相反,且在Gs最低时WUE达到最大值,说明只有Gs降低,Tr和梭梭用于交换二氧化碳进行光合作用过程受到轻微限制时,WUE才能达到最大值,这与邓雄等[46]对怪柳气体水分交换研究得出的结论一致[47].5 a梭梭WUE最高,10 a次之,30 a最低,与它们受到水分胁迫程度(图4)刚好相反,即受到水分胁迫的梭梭会采取提高WUE的适应策略,侧面表明在干旱缺水的荒漠地区,不同林龄梭梭在受到不同的水分胁迫程度下会严格地做出相应的水分调节策略.较高的WUE有利于5 a和10 a梭梭在极端高温缺水的荒漠环境中降低对水分的损耗,提高光合产物产量,将有机物优先投入地下根系[15]以适应极端高温环境.5 a梭梭WUE随温度升高先增后减,在35 ℃后趋于平缓(图4),这与梭梭Gs、Tr变化趋势刚好相反.5 a梭梭Gs和Tr严格控制着WUE大小,在应对极端高温环境时对水分调节的管控能力强,气体交换各部分水分生理变化联系紧密,光合系统反应灵敏.除40 a梭梭外,其余林龄梭梭WUE在高温时波动较小,10、20 a梭梭响应曲线都较为平稳,表明它们在极端高温时能保持稳定的水分状态,减少不必要的水分损耗同时还能维持较好的光合作用状态.由于40 a梭梭Tr随温度不断升高且在极端高温时增势明显,导致其WUE不断减小.水分利用效率降低会导致梭梭在水资源限制较大的极端高温荒漠环境中生长和发育受到抑制,对极端高温逆境的适应力下降,致使植物的生物量减少,生长速度变慢,甚至可能无法正常完成生命周期. ...
多枝柽柳气体交换特性研究
1
2003
... WUE是衡量植物吸收水分利用效率的指标,与光合作用、Gs、Tr和温度等因素相关[43-44],Gs、Tr增加在增强水分运输能力的同时也会致使水分损耗增大,而温度会影响气孔开放继而影响Tr(表2).植物调整WUE是关键的水分调节策略,特别是在极端高温的荒漠环境中,这对植物的生存至关重要.本研究中除40 a梭梭外,不同林龄梭梭WUE随林龄增加而降低(图4),这与已有学者研究结果不完全一致[45],原因可能为本研究区气候环境条件和其研究区域差异较大.除了10 a梭梭外,其余不同生长林龄梭梭WUE和Gs随温度升高变化趋势相反,且在Gs最低时WUE达到最大值,说明只有Gs降低,Tr和梭梭用于交换二氧化碳进行光合作用过程受到轻微限制时,WUE才能达到最大值,这与邓雄等[46]对怪柳气体水分交换研究得出的结论一致[47].5 a梭梭WUE最高,10 a次之,30 a最低,与它们受到水分胁迫程度(图4)刚好相反,即受到水分胁迫的梭梭会采取提高WUE的适应策略,侧面表明在干旱缺水的荒漠地区,不同林龄梭梭在受到不同的水分胁迫程度下会严格地做出相应的水分调节策略.较高的WUE有利于5 a和10 a梭梭在极端高温缺水的荒漠环境中降低对水分的损耗,提高光合产物产量,将有机物优先投入地下根系[15]以适应极端高温环境.5 a梭梭WUE随温度升高先增后减,在35 ℃后趋于平缓(图4),这与梭梭Gs、Tr变化趋势刚好相反.5 a梭梭Gs和Tr严格控制着WUE大小,在应对极端高温环境时对水分调节的管控能力强,气体交换各部分水分生理变化联系紧密,光合系统反应灵敏.除40 a梭梭外,其余林龄梭梭WUE在高温时波动较小,10、20 a梭梭响应曲线都较为平稳,表明它们在极端高温时能保持稳定的水分状态,减少不必要的水分损耗同时还能维持较好的光合作用状态.由于40 a梭梭Tr随温度不断升高且在极端高温时增势明显,导致其WUE不断减小.水分利用效率降低会导致梭梭在水资源限制较大的极端高温荒漠环境中生长和发育受到抑制,对极端高温逆境的适应力下降,致使植物的生物量减少,生长速度变慢,甚至可能无法正常完成生命周期. ...
毛乌素沙地黑沙蒿水分利用效率环境调控:从叶片到生态系统
1
2024
... WUE是衡量植物吸收水分利用效率的指标,与光合作用、Gs、Tr和温度等因素相关[43-44],Gs、Tr增加在增强水分运输能力的同时也会致使水分损耗增大,而温度会影响气孔开放继而影响Tr(表2).植物调整WUE是关键的水分调节策略,特别是在极端高温的荒漠环境中,这对植物的生存至关重要.本研究中除40 a梭梭外,不同林龄梭梭WUE随林龄增加而降低(图4),这与已有学者研究结果不完全一致[45],原因可能为本研究区气候环境条件和其研究区域差异较大.除了10 a梭梭外,其余不同生长林龄梭梭WUE和Gs随温度升高变化趋势相反,且在Gs最低时WUE达到最大值,说明只有Gs降低,Tr和梭梭用于交换二氧化碳进行光合作用过程受到轻微限制时,WUE才能达到最大值,这与邓雄等[46]对怪柳气体水分交换研究得出的结论一致[47].5 a梭梭WUE最高,10 a次之,30 a最低,与它们受到水分胁迫程度(图4)刚好相反,即受到水分胁迫的梭梭会采取提高WUE的适应策略,侧面表明在干旱缺水的荒漠地区,不同林龄梭梭在受到不同的水分胁迫程度下会严格地做出相应的水分调节策略.较高的WUE有利于5 a和10 a梭梭在极端高温缺水的荒漠环境中降低对水分的损耗,提高光合产物产量,将有机物优先投入地下根系[15]以适应极端高温环境.5 a梭梭WUE随温度升高先增后减,在35 ℃后趋于平缓(图4),这与梭梭Gs、Tr变化趋势刚好相反.5 a梭梭Gs和Tr严格控制着WUE大小,在应对极端高温环境时对水分调节的管控能力强,气体交换各部分水分生理变化联系紧密,光合系统反应灵敏.除40 a梭梭外,其余林龄梭梭WUE在高温时波动较小,10、20 a梭梭响应曲线都较为平稳,表明它们在极端高温时能保持稳定的水分状态,减少不必要的水分损耗同时还能维持较好的光合作用状态.由于40 a梭梭Tr随温度不断升高且在极端高温时增势明显,导致其WUE不断减小.水分利用效率降低会导致梭梭在水资源限制较大的极端高温荒漠环境中生长和发育受到抑制,对极端高温逆境的适应力下降,致使植物的生物量减少,生长速度变慢,甚至可能无法正常完成生命周期. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}