


0 引言
宁夏荒漠草原是西北干旱区重要的生态屏障,总面积达155.16万 hm²(占全区草原面积的74.27%)[9]。该区域东接毛乌素沙地,西邻腾格里沙漠,南临黄土高原,具有典型的生态过渡带特征——年降水量仅200~300 mm,蒸发量却高达2 200 mm,植被稳定性直接影响黄河中上游水土保持和防风固沙效能。该生态系统对气候变化响应敏感,近20年人类活动叠加气候暖干化趋势已导致草原退化率超过60%,亟需探明关键物种的生态适应机制以指导修复实践。乡土植物被认为对于当地环境具有更强的适应能力,普遍抗逆适应性更强,植被群落稳定性更高[10],成为众多学者的热点研究对象,因此开展乡土植物研究更能反映宁夏荒漠草原的生态特征。
基于上述背景,本研究选取3种具有典型生态功能的宁夏荒漠草原优势种——甘肃蒿(Artemisia gansuensis,深根系半灌木)、胡枝子(Lespedeza bicolor,豆科固氮灌木)、针茅(Stipa capillata,丛生禾草)为研究对象。通过设置梯度水氮控制实验,分析荒漠草原植物光合作用与蒸腾特性的生理变化,系统比较不同功能群植物在水氮作用下的适应策略差异,不仅为干旱区植被恢复的物种选择提供生理生态学依据,更能揭示资源添加可能引发的种间关系重构风险,对完善荒漠草原适应性管理具有重要意义。
1 研究方法
1.1 研究区概况
本研究位于宁夏农林科学院大水坑野外试验基地(37°24′N,106°58′E)。研究区位于宁夏盐池县,地处宁夏东部,海拔1 295~1 951 m[16],地势呈南高北低、中部高东西低的特点。该地区位于毛乌素沙地和黄土高原的过渡带,也是干旱与半干旱气候的过渡区,属于典型的大陆性荒漠干旱气候。年降水量250~350 mm,80%发生在5—9月,从南向北呈递减趋势。年平均气温7.8 ℃,日照时数2 712 h,无霜期188 d。研究区主要优势物种包括甘肃蒿、胡枝子、针茅、赖草(Leymus secalinus)、油蒿(Artemisia ordosica)、白草(Pennisetum flaccidum)等[17]。
1.2 实验设计
实验观测对象为甘肃蒿、胡枝子和针茅等3种重要的荒漠草原植物。本研究于2024年8月在植物生长季之前设置控水、加氮处理。
1.2.1 加氮处理
控氮处理组包括对照组(CK)、N1(加氮10 kg·hm-2)和N2(加氮20 kg·hm-2),每组处理设置3个2 m×2 m的实验样方。对照组不做任何处理,保证实验样方内研究对象生长环境与当地自然条件一致。
1.2.2 控水处理
控水处理组包括加水50%组和减水50%组,减水组在实验样方上方分别设置带集水器的挡雨板,挡雨板间距均匀,保证能够均匀阻挡并收集减水处理实验样方接受约50%的降水,并将收集到的降水通入加水处理组,每组处理设置3个2 m×2 m的实验样方。整个控水实验装置在雨季之前完成,以确保各组处理的实际效果。
1.3 测定方法
植物光合日变化测定选择晴朗无云天气,采用LI-6800便携式光合仪(LI-COR,Inc., Lincoln,美国)在野外自然条件下测定植物的净光合速率(A,µmol·m-2·s-1)、蒸腾速率(E,mol·m-2·s-1)、胞间二氧化碳浓度(Ci,µmol·mol-1)和气孔导度(gsw,mol· m-2·s-1)等数据。测定时选取目标植物长势健康的完整、新鲜叶片,每组测量15片叶片,观测时间为08:00—18:00,每2 h观测一次。
通过LI-6800便携式光合仪测量自然光强,当自然光强达到1 200±300 µmol·m-2·s-1时,开始观测并记录植物光合生理响应机制研究的相关数据。不同处理组每组测量20片叶片。
1.4 数据处理
使用Excel进行数据整理和初步处理,将整理后的数据在R软件中利用tidyr、Hmisc等包进行数据处理及t检验,并利用ggplot2包进行数据可视化输出。
2 结果与分析
2.1 不同处理下植物的光合-蒸腾速率日变化
2.1.1 甘肃蒿光合-蒸腾速率日变化
在对照处理下,甘肃蒿的净光合速率(A)日变化表现为“双峰型”曲线,具有明显的“午休”现象;最大值出现在14:00,高达63.8 µmol·m-2·s-1,最小值出现在08:00,仅为9.29 µmol·m-2·s-1。甘肃蒿的蒸腾速率(E)日变化也表现为“双峰型”曲线,最大值出现在14:00,为0.0416 mol·m-2·s-1,最小值为0.0113 mol·m-2·s-1(图1)。
图1
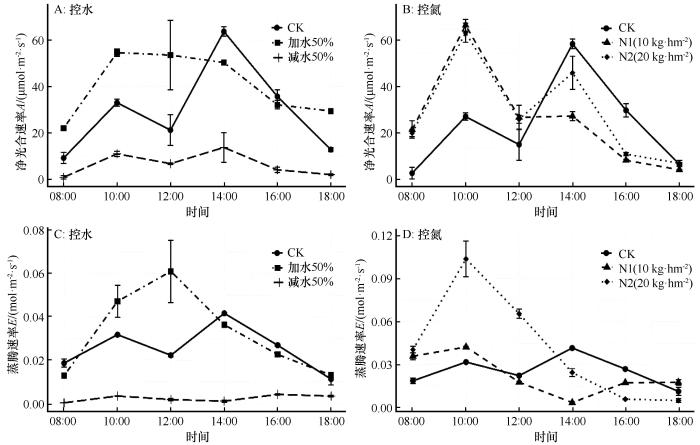
图1
不同处理下甘肃蒿光合-蒸腾速率日变化
Fig.1
Diurnal variations in photosynthesis and transpiration of Artemisia gansuensis under different treatment conditions
在减水处理下,甘肃蒿的净光合速率和蒸腾速率水平均显著降低,这或许与减水处理后,干旱胁迫导致甘肃蒿气孔关闭有关。在干旱条件下,植物会通过关闭气孔减少蒸腾作用,从而保护自身水分平衡。而在加水处理下,甘肃蒿的净光合速率和蒸腾速率均呈现出“单峰型”曲线,“午休”现象消失。对比减水处理,意味着甘肃蒿的水分限制得到缓解,植物无需通过降低光合作用来减少水分消耗,能够在白天维持较高的光合效率,表明水分是调节甘肃蒿植物净光合速率日变化的重要因子。
2.1.2 胡枝子光合-蒸腾速率日变化
图2
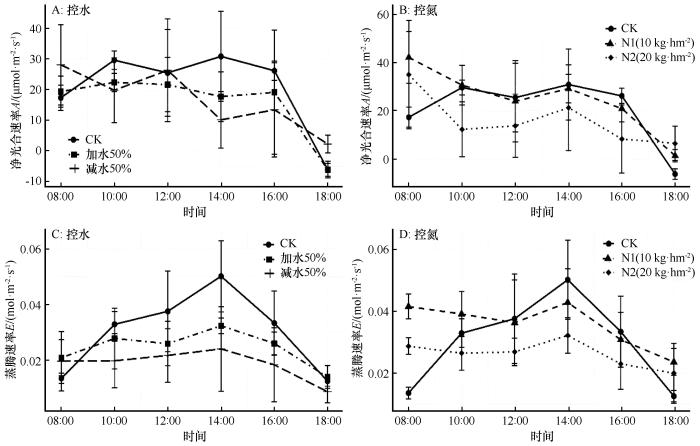
图2
不同处理下胡枝子光合-蒸腾速率日变化
Fig.2
Diurnal variations in photosynthesis and transpiration of Lespedezabicolor under different treatment conditions
2.1.3 针茅光合-蒸腾速率日变化
对照处理下,针茅的净光合速率最大值为43.98 µmol·m-2·s-1,出现在10:00,不同于甘肃蒿和胡枝子;针茅的净光合速率最小值为3.34 µmol·m-2·s-1,出现在18:00(图3A)。针茅的蒸腾速率最大值为0.0438 mol·m-2·s-1,最小值为0.0068 mol·m-2·s-1。
图3
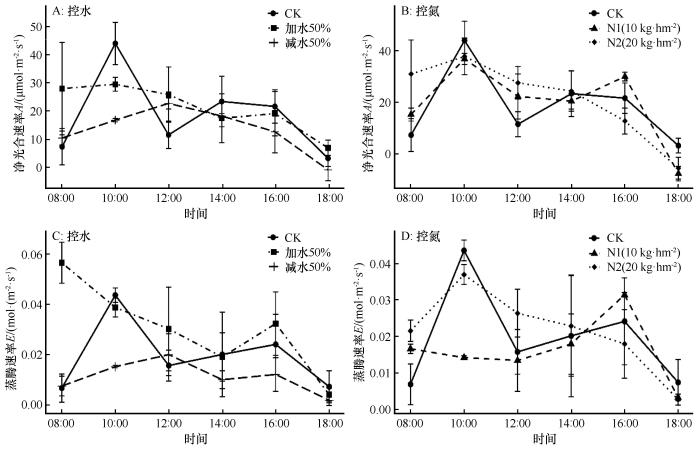
图3
不同处理下针茅光合-蒸腾速率日变化
Fig.3
Diurnal variations in photosynthesis and transpiration of Stipacapillata under different treatment conditions
在对照处理下,3种植物净光合速率的日变化均呈现为“双峰型”曲线,表示这3种植物在自然条件下均出现不同程度的“午休”现象,符合干旱区植物光合作用的一般规律。“午休”现象是植物光合作用对不利环境条件(如高温、干旱和强光)的自我保护机制,反映了植物在特定环境中的生理适应能力。
2.2 不同处理下植物的生理响应调控
2.2.1 不同处理下植物生理性状差异
图4
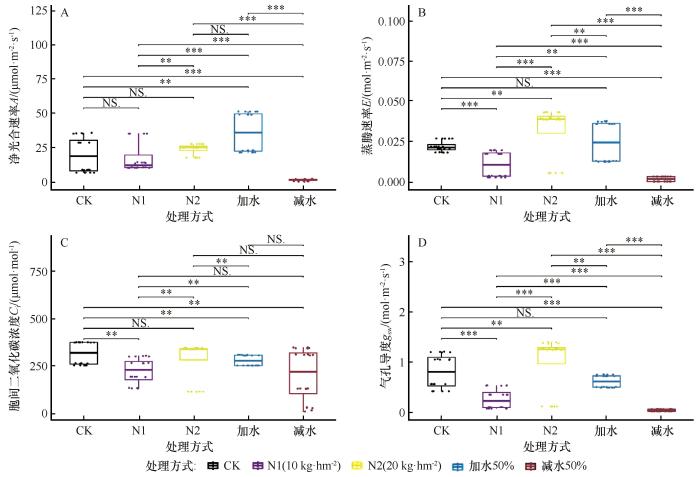
图4
不同处理下甘肃蒿生理性状数据箱线图
Fig.4
Box plots of physiological trait data for Artemisia gansuensis under different treatment conditions
图5
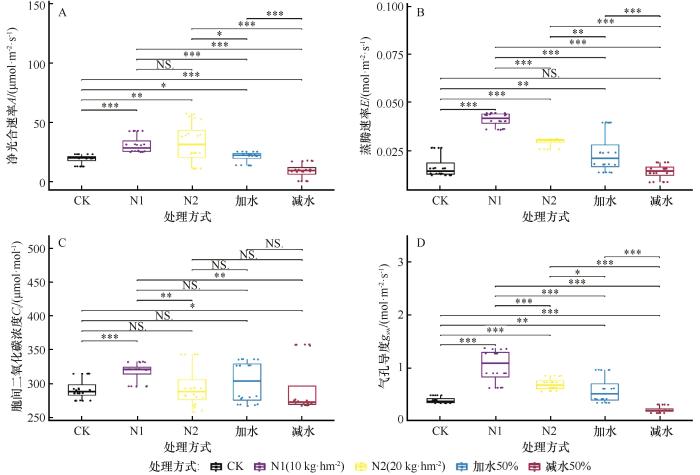
图5
不同处理下胡枝子生理性状数据箱线图
Fig.5
Box plots of physiological trait data for Lespedezabicolor under different treatment conditions
图6
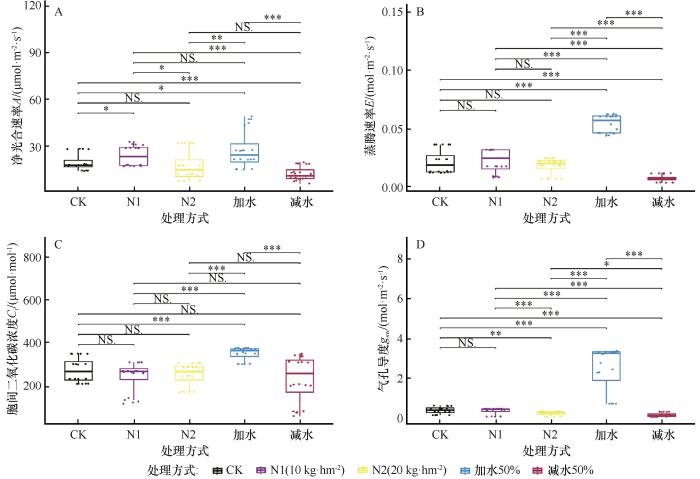
图6
不同处理下针茅生理性状数据箱线图
Fig.6
Box plots of physiological trait data for Stipacapillata under different treatment conditions
2.2.2 不同处理下植物生理性状相关性
对照组的甘肃蒿4项植物生理性状之间均具有显著的相关性。其中,Ci与gsw、A与E之间呈现显著正相关,其他性状之间则表现为显著负相关。氮添加处理时,甘肃蒿的4项生理性状之间均呈现显著正相关关系,且随着氮浓度增加,相关性更强。在控水处理下,甘肃蒿4项生理性状之间的相关性表现出一定差异。其中,加水处理时,A与gsw、E与gsw、A与E之间均呈现显著正相关,其他性状之间则表现为显著负相关。而减水处理时,A与gsw、A与Ci、A与E之间均呈现显著负相关,其他性状之间则表现为显著正相关(图7)。
图7
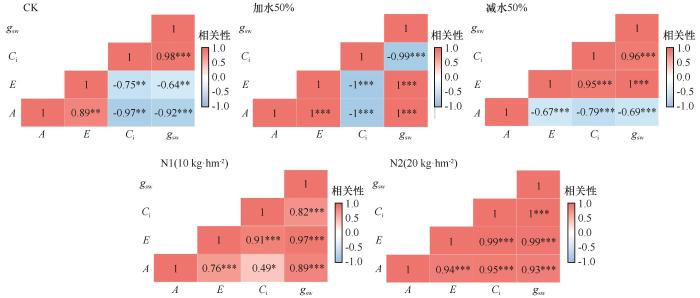
图7
不同处理下甘肃蒿生理性状相关性热图
Fig.7
Heatmap of correlations among physiological traits of Artemisia gansuensis under different treatment conditions
在充足水分和氮添加条件下,甘肃蒿的光合作用和蒸腾作用均与气孔导度具有显著正相关关系,说明三者之间具有密切耦合关系。在加水条件下,光合作用增强,植物叶片对胞间CO₂的吸收和利用速率加快,导致Ci下降,气孔导度显著增大,不仅提升CO₂的供应,提高光合作用,还加快水分通过气孔的蒸腾散失。而在水分胁迫时,甘肃蒿光合作用降低,与叶片气孔因素有关,受非气孔因素的影响可能更大。
由图8可知,3种处理下,胡枝子植物生理性状之间的关系均存在较大差异。对照下,胡枝子植物生理性状中Ci与gsw、Ci与E之间呈现一定负相关性,但差异均不显著。其他性状之间则表现为显著相关关系。加水处理时,胡枝子植物生理性状中A与gsw、Ci、E之间相关性均表现为差异不显著,而其他性状之间则表现为显著正相关;而减水处理时,胡枝子生理性状之间均表现出显著相关性,A与gsw、E与gsw、A与E之间均呈现显著正相关,其他性状之间则表现为显著负相关。N1处理下,胡枝子的生理性状之间均呈现正相关关系,除A与Ci之间差异不显著外,其他性状之间均表现为差异显著;N2处理下,E与gsw之间差异不显著,A与gsw、A与E之间呈现显著正相关,其他性状之间均表现为显著负相关。
图8
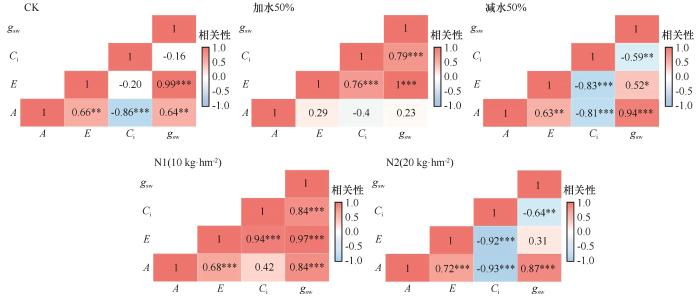
图8
不同处理下胡枝子生理性状相关性热图
Fig.8
Heatmap of correlations among physiological traits of Lespedezabicolor under different treatment conditions
低浓度的氮添加显著增加了胡枝子各项指标间的相关性,而较高浓度的氮添加却降低了蒸腾速率与气孔导度之间的相关性。减水处理下,胡枝子净光合速率与气孔导度之间的相关性显著增大,这或许表明在干旱条件下,气孔导度是胡枝子光合作用的主要限制因子。净光合速率与气孔导度和蒸腾速率的相关性较强,当水分供应充分时,胡枝子蒸腾速率与气孔导度的相关性较强,而净光合速率与气孔导度的相关性较弱,可能是气孔限制被解除,非气孔因素对光合作用的调控作用增强。
由图9可知,3种处理下,针茅植物生理性状之间的关系均存在较大差异。对照下,针茅各项指标间的相关性相对较弱,A与Ci、gsw之间呈现显著相关性,而E与A、Ci、gsw之间的相关性差异均不显著。这说明针茅光合作用主要受气孔行为等植物自身因素的影响,而蒸腾作用可能受环境因素的影响更大。加水处理时,针茅植物生理性状中A与gsw、Ci之间相关性均表现为差异不显著,而其他性状之间则表现为显著相关;而减水处理时,针茅生理性状之间均表现出显著相关性,A与gsw、Ci、E之间均呈现显著负相关,其他性状之间则表现为显著负相关。N1处理下,针茅的生理性状之间均呈现正相关关系,除A与Ci之间差异不显著外,其他性状之间均表现为差异显著;而在N2处理下,A与Ci、E与gsw、gsw与Ci之间差异显著,其他性状之间的关系不显著。
图9
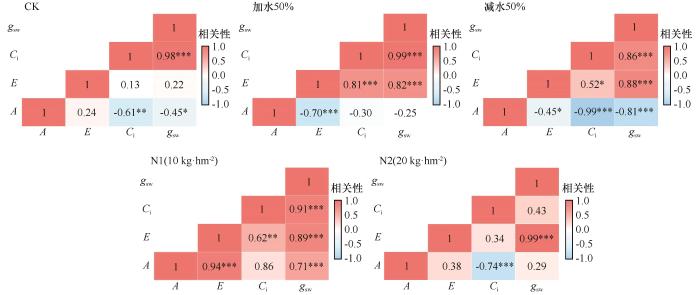
图9
不同处理下针茅生理性状相关性热图
Fig.9
Heatmap of correlations among physiological traits of Stipacapillata under different treatment conditions
干旱胁迫加剧时,针茅光合作用降低受非气孔因素的影响增大;适度改善水分条件时,针茅蒸腾作用受气孔调节作用显著增强。在低浓度氮添加条件下,针茅光合作用和蒸腾作用受气孔调节的影响明显增大;而在高浓度氮添加条件下,针茅蒸腾作用受气孔调节的影响更大,而光合作用受氮代谢负担或胁迫影响各指标间的联动性显著减弱,或许反映了针茅氮添加的“阈值效应”。
3 讨论
3.1 荒漠植物对水分条件的生理生态响应
干旱环境条件下,水分是影响植物生理生态特性的主要限制因子[18]。不同荒漠植物对水分条件改变的响应具有较大差异性。本研究中,甘肃蒿对水分条件的改变较为敏感。减水处理下,甘肃蒿的光合作用和蒸腾作用均显著降低;加水处理下,甘肃蒿的光合作用和蒸腾作用均显著提高,且光合作用日变化曲线由“双峰型”转变为“单峰型”,光合午休现象消失,水分条件是调控甘肃蒿气孔行为的重要环境条件。有学者指出,沙蒿等荒漠优势灌木,应对环境胁迫会采用叶侧重水力效率、茎强化安全机制等开发性策略[19]。Alhaithloul[20]指出,一些蒿属植物会对干旱和高温胁迫表现协同响应机制:双重胁迫通过诱导氧化损伤(过氧化氢积累和电解质渗漏)和抑制光合功能(降低叶绿素合成、气孔导度及光合效率),显著影响植物生长,甘肃蒿面对水分胁迫的敏感表现或许亦是如此。胡枝子在面对水分条件改变时,表现出明显的适应性和耐受性,在减水处理下,胡枝子的蒸腾速率明显降低,并且在下午高温条件下,减水处理下的胡枝子光合速率显著减小;加水处理下光合作用日变化曲线由“双峰型”转变为“单峰型”,但是整体光合水平较对照相差不大。Ahmed等[21]发现,豆科植物存在两种互补的抗旱策略,即依赖高效的光合-蒸腾耦合维持碳获取和侧重氧化防御与渗透调节,胡枝子采用该抗旱策略,通过快速生理调节缓冲胁迫,又保留资源捕获潜力,反映了豆科植物在固氮代谢约束下特有的生态适应智慧,使其成为干旱波动区植被恢复的理想过渡物种。本研究中针茅在面对水分条件的改变时,具有一定保守性,水分条件改变后,针茅的光合和蒸腾作用水平并未产生明显改变,Madouh[22]发现荒漠禾草通过整合气孔调节、资源分配和代谢调整(根冠协调-营养保存策略)来适应水分胁迫,与本研究结论相似。
3.2 荒漠植物对氮添加的生理生态响应
氮素作为植物生长发育过程中重要的营养元素,对植物的生理生态特性至关重要[23-24]。不同生态系统对氮素的响应模式亦有较大差异[25-26],已有学者指出荒漠生态系统对于氮素添加有独特的响应机制,荒漠物种之间对氮添加的反应差异较小,且水分胁迫对生长和生理反应的负面影响通过增加氮供应得到部分补偿[27]。也有学者指出,干旱可以加强叶片光合特性之间的相关性,水分条件在叶片光合作用对氮富集的响应中的起到关键作用[28]。本研究中,甘肃蒿对氮素条件改变较为敏感,低浓度的氮添加迅速改变了甘肃蒿的光合作用水平,高浓度的氮添加条件下,甘肃蒿的光合和蒸腾作用均在早上出现较大提升。胡枝子对低浓度的氮添加并不敏感,而高浓度的氮添加条件下,胡枝子的光合与蒸腾作用水平均有所下降,限制了胡枝子的生理响应。针茅在氮添加条件下,仍表现出一定的保守性,不同浓度氮添加条件下,针茅的光合作用与蒸腾作用日变化整体水平均未产生明显改变。
3.3 荒漠植物应对环境变化的生理生态响应
干旱胁迫会降低植物的光合作用,限制植物生长,从而减少产量[34],其影响植物光合作用的途径是通过影响气孔[35-36]以及代谢过程[37]。净光合速率的降低与气孔关闭和CO2扩散阻力增加之间的关系密不可分[38-40]。本研究中荒漠草原3种植物光合-蒸腾特性面对环境条件改变,表现出不同响应机制。甘肃蒿的光合、蒸腾速率日变化在控水实验中表现出敏感响应状态,水分是影响甘肃蒿气孔导度的关键因子,调节气孔是甘肃蒿响应干旱胁迫的主要手段,在氮处理中,净光合速率出现峰值前移现象,且在N2处理中,上午的蒸腾速率明显剧增,这或许是由于适量的氮素添加促进了叶绿素的合成以及叶片结构的优化,从而提升了甘肃蒿光合作用的表观量子效率,使得甘肃蒿在较低光强的条件下也能发挥更高的光合效率。环境条件改变对胡枝子净光合速率日变化的改变并不明显,而蒸腾速率日变化则出现不同程度的降低,随着外源施氮浓度升高,胡枝子蒸腾速率日变化水平降低。已有大量研究表明[41-42],高浓度施氮会影响豆科植物的共生固氮效率产生,长期施氮将抑制豆科植物固氮酶活性,减少根瘤菌共生,使得豆科植物在群落中竞争力下降,具体可能表现为根系细长化,叶片氮含量下降,光合速率受影响,蒸腾速率下降。不同于甘肃蒿和胡枝子,针茅在控水处理中气孔调节表现出高度的保守性,水分减少使得针茅的光合速率日变化出现明显“双峰型”向“单峰型”的转化,气孔导度显著降低,保守的气孔行为使得针茅在面对干旱胁迫时能极大地减少水分散失,维持植物的正常生长发育。在本研究中,不同浓度的氮素添加对针茅的影响不大,各项指标的变化均不如甘肃蒿和胡枝子显著,已有研究发现[43],短花针茅对氮素的添加反应甚微,即使在较高水平的氮浓度添加以及适宜的土壤湿度条件下仍是如此,这种对氮素的保守利用有助于针茅抵御大气氮沉降,允许其在氮素受限制的土壤中更好地生存。
4 结论
荒漠植物的光合-蒸腾特性是其适应环境胁迫的关键生理枢纽。①水分调控主导种间响应差异:干旱胁迫使3种植物光合-蒸腾速率显著下降,但响应策略呈现功能型分化,深根系灌木甘肃蒿通过消除光合午休(双峰向单峰转变)提升水分利用可塑性;豆科灌木胡枝子则增强气孔调节维持水分平衡;丛生禾草针茅则依赖非气孔限制的保守策略。②氮素效应存在剂量-物种互作:低氮促进豆科植物(胡枝子)气孔导度提升,却抑制非豆科植物(甘肃蒿、针茅)光合性能;高氮则逆转该模式,表明豆科植物固氮特性改变其氮响应阈值。③水氮互作重塑调控路径:干旱条件下氮添加加剧非气孔限制(RuBisCO活性下降),而适水环境则增强气孔主导调控,揭示植物碳-水权衡策略的环境依赖性。
参考文献
光伏电站建设对植物群落与土壤特征的影响
[J].
New-generation geostationary satellite reveals widespread midday depression in dryland photosynthesis during 2020 western U.S.heatwave
[J].
Adaptive strategies based on shrub leaf-stem anatomy and their environmental interpretations in the eastern Qaidam Basin
[J].
Impact of combined heat and drought stress on the potential growth responses of the desert grass artemisia sieberi alba:relation to biochemical and molecular adaptation
[J].
Eco-physiological responses of desert and riverain legume plant species to extreme environmental stress
[J].
Eco-physiological responses of native desert plant species to drought and nutritional levels:case of kuwait
[J].
Additive effects of simulated climate changes,elevated CO2,and nitrogen deposition on grassland diversity
[J].
Response of soil microbial biomass and community composition to chronic nitrogen additions at Harvard forest
[J].
effects of an eight-year nitrogen and phosphorous addition on leaf photosynthesis and chemistry of mature Castanopsis sclerophylla trees in subtropical China
[J].
Nitrogen saturation in northern forest ecosystems
[J].
Combined effects of nitrogen deposition and water stress on growth and physiological responses of two annual desert plants in Northwestern China
[J].
Water controls the divergent responses of terrestrial plant photosynthesis under nitrogen enrichment
[J].
Erodium oxyrhinchum sustains severe drought stress by maintaining stable photosynthetic electron transport under progressive drought conditions
[J].
Is photoprotection of PSII one of the key mechanisms for drought tolerance in maize?
[J].
Linking leaf water potential,photosynthesis and chlorophyll loss with mechanisms of photo and antioxidant protection in juvenile olive trees subjected to severe drought
[J].
Stomata coordinate with plant hydraulics to regulate transpiration response to vapour pressure deficit in wheat
[J].
Transpiration response to soil drying and vapor pressure deficit is soil texture specific
[J].
Plant drought stress:effects,mechanisms and management
[J].
Drought-inhibition of photosynthesis in C3 plants:stomatal and non-stomatal limitations revisited
[J].
Decreased Rubisco activity during water stress is not induced by decreased relative water content but related to conditions of low stomatal conductance and chloroplast CO2 concentration
[J].
Water stress inhibits plant photosynthesis by decreasing coupling factor and ATP
[J].
Plant responses to drought and rewatering
[J].
The response of photosynthesis to soil water stress
[M]//
Regulation of photosynthesis and stomatal and mesophyll conductance under water stress and recovery in olive trees:correlation with gene expression of carbonic anhydrase and aquaporins
[J].
Biological nitrogen fixation and its response to nitrogen input in two mature tropical plantations with and without legume trees
[J].
Nitrogen fertilization reduces nitrogen fixation activity of diverse diazotrophs in switchgrass roots
[J].
Growth of Stipa breviflora does not respond to nitrogen addition because of its conservative nitrogen utilization
[J].

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}