宁夏盐池人工柠条( Caragana korshinskii
石苗苗 , 1 , 2 , 霍建强 2 , 3 , 韩高玲 2 , 3 , 南益聪 2 , 3 , 朱小娟 1 , 虎瑞 , 2 , 苏雪 , 1
1.西北师范大学,甘肃 兰州 730070
2.中国科学院西北生态环境资源研究院 沙坡头沙漠研究试验站/干旱区生态安全与可持续发展全国重点实验室,甘肃 兰州 730000
3.中国科学院大学,北京 100049
Characteristics of herbaceous community in revegetation Caragana korshinskii shrublands in Yanchi , Ningxia , China
Shi Miaomiao , 1 , 2 , Huo Jianqiang 2 , 3 , Han Gaoling 2 , 3 , Nan Yicong 2 , 3 , Zhu Xiaojuan 1 , Hu Rui , 2 , Su Xue , 1
1.Northwest Normal University,Lanzhou 730070,China
2.Shapotou Desert Research and Experiment Station / State Key Laboratory of Ecological Safety and Sustainable Development in Arid Lands,Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
3.University of Chinese Academy of Sciences,Beijing 100049,China
Received: 2025-03-31
Revised: 2025-05-09
作者简介 About authors
石苗苗(2001—),女,甘肃陇南人,硕士研究生,主要从事干旱区生态学研究E-mail:3074007849@qq.com
, E-mail:3074007849@qq.com
摘要
本研究以宁夏盐池1984、1996、2005、2015年的人工柠条(Caragana korshinskii )林的草本群落为对象,采用时空替代法,调查草本植物种类、数量等群落特征,测定土壤理化性质,探讨不同恢复年限下草本群落结构、相似性及物种多样性的变化特征,并分析影响群落变化的关键因素。结果表明:该区域草本物种共19科39属。随着植被恢复年限增加,草本物种丰富度和多样性指数呈波动性变化,且由于放牧原因,1996、2005年样地丰富度低于其余2个样地,2015年Shannon多样性指数最高,其次是1984年;同时,各样地间草本层群落差异性较大。此外,随着植被恢复,土壤有机碳、全氮、微生物量碳氮的含量均呈增加趋势,同时,较高的土壤有机碳和微生物量碳氮也增加了草本植被盖度、多度、地上生物量,并且浅层土壤含水量(20~60 cm)决定了草本植被丰富度和多样性。
关键词:
沙区 人工柠条林 草本群落变化 不同年限 放牧 土壤理化性质
Abstract
This study focused on the herbaceous communities in revegetation Caragana korshinskii shrubland plots in Yanchi, Ningxia, from four different years:1984, 1996, 2005, and 2015. Using a space-for-time substitution method, the research investigated community characteristics such as species and abundance of herbaceous plants, measured soil physical and chemical properties, and explored the changes in community structure, similarity, and species diversity under different years. The study also analyzed key factors influencing community changes. Results showed a total of 39 genera belonging to 19 families of herbaceous species in the area. With the increase in vegetation restoration years, the richness and diversity index of herbaceous species generally showed fluctuating changes. And this was due to grazing activities, the richness in the 1996 and 2005 plots was lower than in the other two plots. The 2015 plot had the highest Shannon diversity index, followed by the 1984 plot, with significant differences in the herbaceous layer community structure among plots. In addition, with the restoration of vegetation, the contents of soil organic carbon, total nitrogen, microbial biomass carbon and nitrogen all showed an increasing trend. Meanwhile, higher soil organic carbon and microbial biomass carbon and nitrogen also increased the coverage, abundance and aboveground biomass of herbaceous vegetation. Moreover, the soil water content in the shallow layer (20-60 cm) determined the richness and diversity of herbaceous vegetation.
Keywords:
sandy area Caragana korshinskii shrublandherbaceous community changes different years grazing soil physical and chemical properties
本文引用格式
石苗苗, 霍建强, 韩高玲, 南益聪, 朱小娟, 虎瑞, 苏雪. 宁夏盐池人工柠条( Caragana korshinskii . 中国沙漠 [J], 2025, 45(3): 337-345 doi:10.7522/j.issn.1000-694X.2025.00059
Shi Miaomiao, Huo Jianqiang, Han Gaoling, Nan Yicong, Zhu Xiaojuan, Hu Rui, Su Xue. Characteristics of herbaceous community in revegetation Caragana korshinskii shrublands in Yanchi , Ningxia , China Journal of Desert Research
0 引言
宁夏盐池位处毛乌素沙地西端,因不合理土地利用,区域生态系统严重失衡,风沙区域有蔓延趋势[1 ] 。据全国沙化土地监测结果,盐池沙化土地面积达到486 932 hm2 ,占宁夏沙化土地面积的39.4%,占盐池面积的 68.3%[2 ] ,土地沙化已成为限制该地区经济发展的主要因素[3 ] 。植物重建与恢复作为沙化土地修复的有效手段,自20世纪60年代以来,该区域通过大规模人工柠条(Caragana korshinskii )灌木林的营造,提高了土壤有机质的输入,显著改善了土壤物理、化学及微生物属性,增加了生物多样性。
人工固沙植物群落演替呈现明显的时间依赖性。腾格里沙漠东南缘人工固沙植被区的研究显示,随着时间推移,植被物种丰富度和多样性指数呈上升趋势;土壤理化性质得到改善,风沙活动减少,为植物种子的传播和定居创造了有利条件,从而逐渐提升物种丰富度[4 ] 。类似地,高寒沙区的研究也显示,在人工治理初期以柠条和油蒿(Artemisia ordosica )为主的人工固沙植被区,植物群落的物种丰富度、Shannon多样性指数和Pielou均匀度指数均不断增加并逐渐达到峰值,随后赖草(Leymus racemosus )占优势地位[5 ] 。植被恢复过程中,土壤环境改善、种间竞争与生态位分化等因素共同作用,推动群落结构复杂化与功能完善化,进一步证实植被重建能够有效促进土壤环境改良和植被群落恢复。在干旱沙区,柠条作为建群种,能够降低地表温度、减少水分蒸发,有效固定沙土,减少风蚀和水蚀,从而提高土壤质量[6 ] 。对樟子松(Pinus sylvestris )人工林林下植物多样性和生物量动态变化的研究显示,随着林龄增加,林下草本植物科、属、种数呈先减少后增加的规律,林下草本层Shannon多样性指数、Simpson优势度指数均随着林龄增加呈先减少后增加的规律,Pielou均匀度指数随林龄增加而逐渐增加[7 ] 。这说明植物群落随植被的动态演替逐渐趋于稳定。那么在本研究区,随着柠条林建植年限的增加,其对草本植物群落结构、多样性指数以及土壤理化性质如何影响,在干旱区是否具有明显的演替特征等有待深入研究。
除了植被恢复时间外,放牧对植被演替过程的影响同样不可忽视。放牧改变土壤水分和养分状况,导致植被高度、盖度及地上生物量的降低[8 ] 。盐池自2002年开始采取水土保持生态修复以来,植被覆盖度从修复前的30%提升到目前的65%[9 ] 。尽管过度放牧会导致植被退化和土壤侵蚀,适度放牧却可能促进草本植物的生长[10 ] 。在本研究区域内,部分样地仍然存在放牧现象,需要了解放牧如何影响该地区草本群落的动态变化,以及植被恢复过程中群落结构和多样性。因此,本研究采用时空替代法,重点关注人工种植柠条灌木林,系统探究不同年限下草本群落结构、Shannon多样性指数、Simpson多样性指数、Pielou均匀度指数以及土壤理化性质的变化特征。旨在明晰人工固沙植物群落随时间的变化规律,揭示驱动群落变化的关键因子与影响因素,为沙化地区的生态修复实践提供科学依据与理论支持。
1 研究区概况
盐池县(37°86′N,107°54′E)位于宁夏回族自治区东部,地处鄂尔多斯台地向黄土丘陵区的过渡地带,地形地貌较为复杂,曾是中国沙化危害的严重区域。常年干旱少雨、多风沙,多为固定和半固定沙丘,属于典型的中温带大陆型气候。当地土壤以风沙土为主,植被类型主要为沙生植被,如柠条、茵陈蒿(Artemisia capillaris )、小画眉(Eragrostis pilosa )、地锦草(Euphorbia humifusa )、刺藜(Teloxys aristata )、苦豆子(Sophora alopecuroides )、尖头叶藜(Chenopodium acuminatum )、针茅(Stipa capillata )、刺蓬(Salsola tragus )、雾冰藜(Grubovia dasyphylla )等。年平均气温为7.8 ℃,多年平均降水量为258 mm,雨季为6—9月,多年平均蒸发量高达2 000 mm,土壤水分长期处于亏缺状态。
2 研究方法
2.1 样地设置及物种多样性调查
在毛乌素沙地西段的盐池地区选取4个年份(1984、1996、2005、2015年)种植的人工柠条林样地进行研究(表1 )。所有年份的植被恢复样地均已封育,但调查过程中,在1996年和2005年样地发现放牧痕迹,因此这2个样地被视为放牧样地。
在每个年份的样地中,从东到西分别设置3条100 m的样带,样带间隔10 m,包括丘顶、丘间低地、迎风坡、背风坡等多种地形,以减小地形差异造成的影响。在每条样带上设置5个大样方(10 m×10 m),样方间隔10 m。在每个大样方内,沿着对角线随机选5个点,去除表层凋落物及杂质后,采取0~5 cm和5~10 cm土壤。将同一深度土样混合为一个混合样,装入自封袋,并放入冰盒带回实验室,过2 mm筛子以去除根系和枯落物,研磨混匀用于后续分析。测量指标包括土壤微生物量碳(MBC)、微生物量氮(MBN)、有机碳(SOC)、全氮(TN)、全磷(TP)含量,以及土壤pH。
在每个大样方内,随机设置1个草本样方(0.5 m×0.5 m)。调查草本物种种类、数量、盖度,并剪取所有植株的地上部分,装入信封袋中,带回实验室烘干,以测量草本地上部分生物量。
随机选取6个大样方,在样方中心用土钻采集不同深度(0~20、20~60、60~200 cm)的土壤样品,装入带有编号的铝盒中,现场称重后,带回实验室在105 ℃下烘干24小时,利用烘干法计算土壤质量含水量。
2.2 土壤样品测定方法和植物群落多样性计算方法
土壤pH采用玻璃电极法测定,有机碳采用重铬酸钾容量法-外加热法测定,全氮采用凯氏定氮法测定,全磷采用酸消解-钼锑抗分光光度法测定[11 ] ;微生物量碳氮采用氯仿熏蒸法提取-碳氮分析仪测定[12 ] ;含水率采用烘干法测定[13 ] 。
根据样方调查的草本及木本物种的种类、盖度、多度计算出物种丰富度(R )、 Shannon多样性指数(H )、Simpson指数(D ),公式如下[14 -18 ] :
R = S (1)
H = - ∑ i = 1 S P i l n P i (2)
D = 1 - ∑ i = 1 S ( P i ) 2 (3)
Pi =(相对多度+相对盖度+相对频度)/3(4)
式中:S 代表样方内物种数目;Pi 代表样方内i 种的相对重要值。
基于Shannon多样性指数计算Pielou均匀度指数,公式如下[19 ] :
J = H / l n S (5)
群落相似性采用Jaccard相似性系数计算,公式如下[20 ] :
q = w / ( a + b - w ) (6)
式中:q 表示群落的相似性系数;w 表示2个植物群落中的共同物种数;a 和b 分别表示群落A、B的物种总数。
2.3 数据处理
使用Excel 2010和IBM SPSS Statistics 26软件对数据进行统计,用单因素方差分析法(ANOVA),分析不同植被恢复年限对植物群落的影响,用独立样本T检验分析法,分析放牧对植物群落的影响,使用Canoco进行冗余分析(RDA),并使用Origin Pro 2021作图表。
3 结果与分析
3.1 不同恢复年限草本群落结构特征
1984年的物种数(23种)明显高于1996年(18种)和2005年(16种),且1984年和2015年草本植物的地上生物量和凋落物含量均高于其余两个样地。1984年样地中草本植被优势种为茵陈蒿、刺藜、刺蓬;1996年为苦豆子、地锦草、小画眉;2005年样地中为茵陈蒿、雾冰藜、尖头叶藜;2015年样地中为刺蓬、针茅、茵陈蒿(表2 )。
3.2 不同恢复年限草本群落物种多样性特征
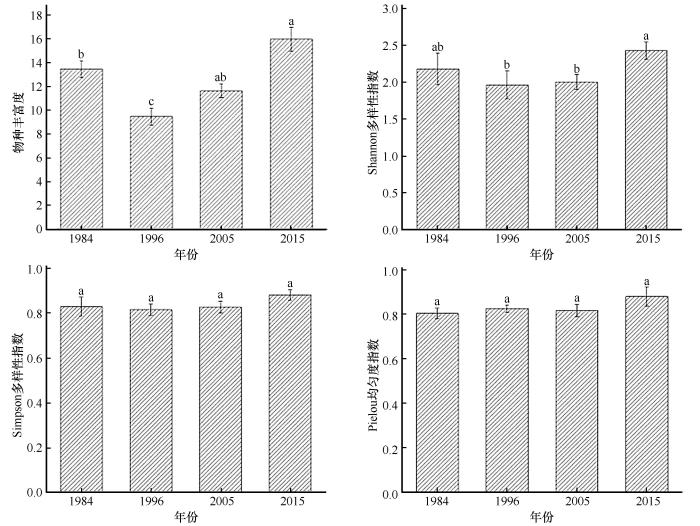
草本植被的物种丰富度(R )、Shannon多样性指数、Simpson多样性指数和Peilou均匀度指数随着植被恢复年限的增加呈波动性变化(图1 )。2015年样地的物种丰富度最高,其次为1984年样地。1984年和2015年样地的 Shannon多样性指数、Simpson多样性指数均高于其余2个样地。
图1
图1
不同样地草本植被多样性指数
注: 图中的值为平均值±标准误差(SE );不同小写字母表示不同植被恢复年限样地差异显著(P <0.05)
Fig.1
Herbaceous vegetation diversity indices in different sample plots
3.3 不同恢复年限草本群落相似性系数
草本层样地间的相似性系数为0.139~0.412,相似性较低(表3 )。1984年和1996年样地间相似性极低(0.139),1984年和2015年样地间相似性较高(0.412)。
3.4 放牧对草本群落的影响
放牧对草本盖度、地上生物量、凋落物、Shannon多样性指数、物种丰富度、Simpson多样性指数及Pielou均匀度指数造成影响,且对草本物种丰富度、Shannon多样性指数和凋落物影响显著(表4 )。
3.5 不同恢复年限样地植被-土壤关系
除1996年样地外,土壤有机碳、全氮、微生物量碳氮含量在植被恢复过程中波动变化(表5 )。表层土壤有机碳、全氮、微生物量碳氮的含量在1984年样地中最高,在1996年样地中最低。下层土壤中表现出相似的规律。
1984年样地的表层土壤(0~20 cm)含水量显著高于其余样地,而在1996年样地中,表层土壤含水量(0~20 cm)低于浅层土壤(20~60 cm,表6 )。
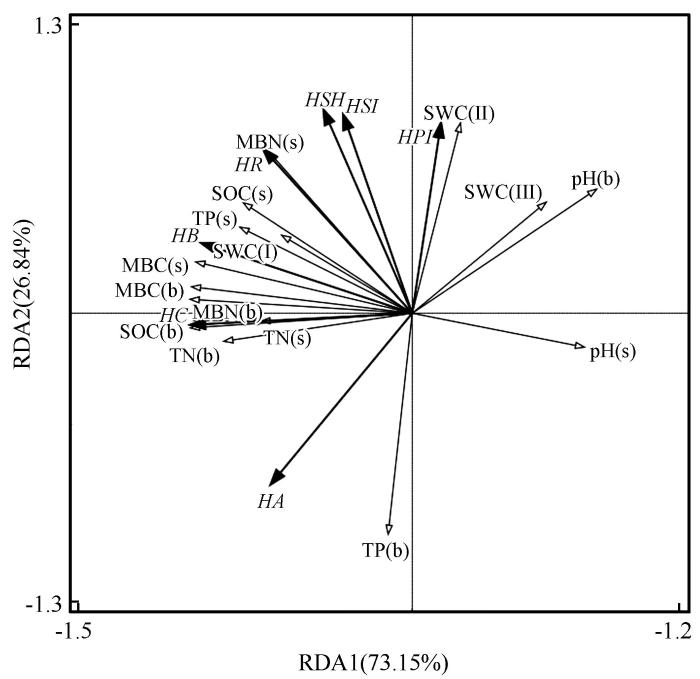
土壤有机碳、微生物量碳氮和浅层土壤(20~60 cm)含水量是影响草本丰富度和多样性的关键因素(图2 )。分析发现第一轴(RDA1)解释了土壤因子对草本植被特征73.15%的影响,第二轴(RDA2)解释了26.84%的影响;土壤有机碳、微生物量碳氮与草本物种丰富度、多度、盖度、Shannon多样性指数及Simpson多样性指数均呈正相关。表层土壤因子中,土壤有机碳和微生物量碳氮与草本盖度及地上生物量相关性较强;下层土壤因子中,土壤有机碳和微生物量碳氮与草本丰富度及多样性相关性较强。表层土壤(0~20 cm)含水量与草本多度、盖度、地上生物量、丰富度及多样性相关性较强,浅层土壤(20~60 cm)含水量与草本多样性和丰富度相关性较强。
图2
图2
不同样地植被特征与土壤因子的冗余分析
注: SOC(b):表层土壤有机碳(g·kg-1 );TN(b):表层土壤全氮(g·kg-1 );TP(b):表层土壤全磷(g·kg-1 ); MBC(b):表层土壤微生物量碳(mg·kg-1 );MBN(b):表层土壤微生物量氮(mg·kg-1 );SOC(s):下层土壤有机碳(g·kg-1 );TN(s):下层土壤全氮(g·kg-1 );TP(s):下层土壤全磷(g·kg-1 ); MBC(s):下层土壤微生物量碳(mg·kg-1 );MBN(s):下层土壤微生物量氮(mg·kg-1 );SWC(Ⅰ):0~20 cm土层土壤水分含量(g·g-1 );SWC(Ⅱ):20~60 cm土层土壤水分含量(g·g-1 );SWC(Ⅲ):60~200 cm土层土壤水分含量(g·g-1 );HA:草本多度;HR:草本丰富度;HSH:草本Shannon多样性指数;HSI:草本 Simpson多样性指数;HPI:草本均匀度指数;HB:草本生物量(g);HC:草本盖度(%)
Fig.2
Redundancy analysis of vegetation characteristics and soil factors in different plots
土壤养分随着植物生长、枯落物积累和根系分泌物等过程在土壤中富集,从而土壤性质得以改善,这种改善后的土壤环境促进了生物土壤结皮的形成和生长,而结皮拓殖后,使得表层土壤水分蒸发量减少,且土壤水分浅层化。而草本植物的根系大多在40~60 cm处,因此,养分富集后形成的富含水分且稳定的土壤环境为草本的定植、生长和发育创造了有利条件。
4 讨论
通过对4个年份的人工柠条灌木林的调查发现,研究区共记录草本植物43种,隶属于19科、39属,总体上物种组成较为简单。这主要是由于研究区位于鄂尔多斯台地向黄土丘陵区的过渡地带,该地区常年干旱少雨,水分匮乏,影响了沙化土地的恢复以及人工固沙植被的稳定性,限制了植物种类的丰富度[21 -22 ] 。随着植被恢复年限的增加,草本植被的盖度总体呈现上升趋势。其中,刺蓬、虫实、雾冰藜等一年生植物的比例逐渐减少,而诸如苦豆子和茵陈蒿这类多年生草本植物增多。多年生植物凭借发达的根系,能够高效吸收深层土壤水分和养分,其较长的生命周期有益于生物量的持续积累,从而在草本群落中逐渐占据主导地位[23 ] 。如茵陈蒿、小画眉、刺蓬、针茅、地锦草等在沙区植被演替过程中逐渐成为优势种(表2 ),这与其强大的生命力和耐旱特性有关。例如,小画眉草发育出较多的浅层须根系,能够在降雨季节快速吸收水分,这种资源利用策略使其在沙区植被演替中具有显著竞争优势[24 ] 。
物种多样性对维持生态平衡和人类生存至关重要,而多样性指数用于衡量生态系统中物种的多样性和异质性[25 ] 。本研究显示,草本物种丰富度随植被恢复年限的增加具有波动性,且1984年和2015年样地的物种丰富度、Shannon多样性指数和Simpson多样性指数均高于其余两个样地(图1 ),与王树力等[26 ] 关于植被物种丰富度随着植被恢复年限的增加而增大的结果不同。在柠条灌木林重建后,沙地中自然侵入的植物种数逐渐增多,尤其是一些先锋草本植物的拓殖,对沙丘表面的固定起到了关键作用,有效改善了土壤水分和养分状况,为植被的定居与繁殖提供了理想的环境,从而提升了物种的丰富度。但在本研究中,2015年样地的丰富度及多样性指数最高,1984年次之。1984年样地中生物土壤结皮盖度超过90%,且存在较多苔藓结皮,其致密结构阻碍降水入渗和种子着床,改变了土壤中降水的再分配过程,使水分浅层化[27 ] ,进而引起草本植物退化。此外,致密的结皮组织还会影响草本种子的定殖。然而,2015年样地中较高土壤水分和较少外界干扰,促进了草本植物的快速生长和扩散,表现为更高的物种丰富度、Shannon多样性指数和Simpson多样性指数。这表明,在植被恢复的过程中,虽然植被恢复年限增加有利于草本群落的发展,但生物土壤结皮在这个过程中扮演着不可忽视的角色,对植被群落的动态有一定的调节作用。
此外,值得注意的是,1996年和2005年样地的草本物种丰富度、多样性指数、地上生物量及凋落物均低于1984年和2015年的样地(表2 、图1 )。调查显示,1996年和2005年样地存在放牧现象。放牧显著影响了草本植被的物种丰富度、多样性及凋落物(表4 )。许多研究表明,放牧会破坏土壤结构,导致土壤保水保肥能力下降,并加剧土壤养分流失。这种变化使得植物无法获得生长所需的养分和水分,导致其生长和繁殖受到限制,从而降低了植被的丰富度和多样性[28 ] 。这与孙宗玖等[29 ] 的结论一致,他们发现放牧后草本群落盖度及地上生物量明显降低,并显著影响了Shannon 指数。尽管有研究指出,适度放牧可增加植被多样性,促进荒漠草原的可持续利用[30 ] 。但在本研究区域,由于环境恶劣和水分极度缺乏,放牧干扰使得原本脆弱的生态系统变得更加紧张,因此抑制作用更加突出。然而,本研究中放牧对草本植被的 Simpson多样性指数及Pielou均匀度指数的影响并不显著,这可能与放牧的时间、强度以及种源等因素有关[31 -32 ] 。
植物群落相似性指数用于量化植物群落之间的异质性,反映植物群落生境条件的异质性[33 ] 。本研究结果表明,草本层不同恢复年限的群落样地间相似系数为0.139~0.412(表3 ),样地间整体相似性较低,草本群落结构存在较大差异。此外,由于封育措施,2015年样地的草本群落特征与恢复时间较长的1984年样地表现出较高相似性。
土壤作为植物赖以生存的物质基础,其性质的变化能够反映植被的变化。本研究发现,除1996年样地外,土壤有机碳、全氮、微生物量碳氮的含量在植被恢复过程中不断富集,且随着土壤深度的增加而降低(表5 )。这一现象印证了植被重建对土壤的理化性质的改良。柠条灌木林通过凋落物输入和根系分泌物释放,促进微生物分解代谢,加速腐殖质形成,驱动碳氮养分的富集,从而更加有利于草本植物的生长[34 -35 ] 。土壤pH则呈相反的变化趋势,可能是由于表层土壤中的植物呼吸作用释放出的二氧化碳会降低土壤pH[36 ] 。而1996年样地因受放牧的影响,土壤pH升高而土壤养分含量均低于其余年限样地。这是因为放牧破坏土壤积盐与脱盐的平衡,使可溶性盐类迁移,并累积于土表,导致土壤pH值升高,从而引起土壤肥力下降[37 ] 。同时,牲畜的踩踏对表层土壤造成严重破坏,也加剧了土壤养分流失,被破坏后的土壤难以维持植被的生长,进而导致草本植被丰富度和多样性降低。
本研究通过冗余分析(RDA)研究不同植被恢复年限样地的植被-土壤关系,草本植被的盖度、地上生物量、丰富度、Shannon多样性指数和Simpson多样性指数与土壤有机碳、全氮、微生物量碳氮之间表现出较强的正相关性(图2 )。这表明土壤的有机碳、微生物量碳氮是驱动草本植被群落结构变化的关键环境因子。有研究指出,土壤微生物量碳氮在土壤养分的供应和转化过程中发挥着决定作用[38 ] ,他们不仅为植物提供必要的营养,还增强了植物的抗逆性,这对于植物的生长至关重要。同时,高含量的土壤全氮和有机碳能够促进植物生长,从而提高植物群落的多样性[39 ] 。在高寒草甸退化和沙化草地中,植物群落的地上生物量、盖度、物种丰富度、Shannon多样性指数与土壤有机碳和全氮含量呈极显著正相关[40 ] 。这与本研究中土壤全氮、有机碳含量与草本多度、盖度及生物量的正相关一致。在干旱半干旱地区,土壤含水量是植物生长和群落稳定性的关键限制因子[41 ] 。充足的土壤水分不仅满足植物的水分需求,提高植物的成活率和生长速度,还影响土壤中养分的有效性,从而间接影响植物的生长。本研究发现,草本植被多度、盖度、地上生物量和丰富度与表层土壤(0~20 cm)水分有较强的相关性,此外,草本植被的Shannon多样性指数和Simpson多样性指数也与浅层土壤(20~60 cm)水分呈较强的相关性。这可能是因为草本植物更容易获取表层土壤(0~20 cm)水分,有利于种子的萌发、幼苗的生长以及植株的发育,进而提高了草本植被的多度、盖度和地上生物量。而浅层土壤(20~60 cm)水分为具有不同根系深度的草本植物提供了多样化的生存空间,一些根系较深的多年生草本植物利用浅层土壤水分,与主要依赖表层土壤水分的植物形成生态位分化,减少种间竞争,使更多种类的草本植物能够共存,从而提高了草本植被的多样性。这与腾格里沙漠土壤水分含量与植被的研究结果相似[42 ] 。在民勤青土湖区退耕40年的研究中也发现草本植被的Shannon多样性指数与0~20 cm土层的土壤含水量呈极显著正相关,强调了表层土壤水分在植被自然演替中的关键性作用[43 ] 。
5 结论
随着植被恢复年限的增加,草本植被的物种丰富度、Shannon多样性指数、Simpson多样性指数和Pielou均匀度指数均呈波动性变化,草本群落结构变得更加复杂,并且样地间群落相似性表现出显著的差异。除1996年样地,土壤有机碳、全氮、微生物量碳氮的含量随植被恢复年限的增加而呈增大的趋势,且随着土壤深度的增加而减小。土壤有机碳和微生物量碳氮的增加促进了草本植物的生长,其多度、盖度、地上生物量均显著增加,并且浅层土壤(20~60 cm)含水量决定了草本植被丰富度和多样性。此外,放牧对植被恢复过程中草本植物的盖度、地上生物量、丰富度和多样性等特征有显著影响。然而,放牧强度对植被恢复的具体影响还需要进一步深入研究。
参考文献
View Option
[1]
张曦 宁夏盐池北部沙化土地综合治理决策支持系统设计与实现
[D].西安 :电子科技大学 ,2013 .
[本文引用: 1]
[2]
孙景梅 ,李谭宝 ,谷洁 ,等 宁夏盐池县沙区植被覆盖与植物多样性变化
[J].林业科技通讯 ,2021 ,581 (5 ):3 -7 .
[本文引用: 1]
[3]
文妙霞 ,李谭宝 ,吴海平 盐池县沙化土地及其利用动态变化分析
[J].水土保持通报 ,2004 ,24 (5 ):83 -85 .
[本文引用: 1]
[4]
刘任涛,腾格里沙漠固沙造林后植被-土壤系统演变特征及生态效应评价
[D].银川 :宁夏大学 ,2020 .
[本文引用: 1]
[5]
杨洪晓 ,卢琦 ,吴波 ,等 高寒沙区植被人工修复与种子植物物种多样性的变化
[J].林业科学 ,2004 ,40 (5 ):45 -49 .
[本文引用: 1]
[6]
杨钧 ,王瑞霞 ,王俊 ,等 毛乌素沙地不同恢复年限柠条固沙林微生物群落特征及其影响因素
[J].应用生态学报 ,2024 ,35 (7 ):1807 -1814 .
[本文引用: 1]
[7]
胡尔查 ,王铮 ,李梓豪 ,等 毛乌素沙地不同林龄樟子松人工林林下植物多样性和生物量的动态变化
[J].生态学杂志 ,2024 ,43 (11 ):3246 -3254 .
[本文引用: 1]
[8]
张伟华 ,关世英 ,李跃进 不同牧压强度对草原土壤水分、养分及其地上生物量的影响
[J].干旱区资源与环境 ,2000 ,14 (4 ):62 -65 .
[本文引用: 1]
[9]
张晓童 盐池县抗旱治沙生态修复的探索与实践
[J].草业与畜牧 ,2011 (10 ):27 -28 .
[本文引用: 1]
[10]
连仲民 ,徐文轩 ,杨维康 ,等 放牧对草地土壤种子库的影响
[J].草业科学 ,2014 ,31 (12 ):2301 -2307 .
[本文引用: 1]
[11]
贺郝钰 ,刘蔚 ,常宗强 ,等 沙区生物土壤结皮演替对表层土壤养分和微生物群落组成的影响
[J].干旱区地理 ,2024 ,47 (10 ):1724 -1734 .
[本文引用: 1]
[12]
Vance E D Brooks P C Jenkinson D S An extraction method for measuring soil microbial biomass
[J].Soil Biology Biochemistry ,1987 ,19 :703 -707 .
[本文引用: 1]
[13]
雷菲亚 ,李小双 ,陶冶 ,等 西北干旱区藓类结皮覆盖下土壤多功能性特征及影响因子
[J].干旱区研究 ,2024 ,41 (5 ):812 -820 .
[本文引用: 1]
[14]
马克平 ,刘玉明 生物群落多样性的测度方法Ⅰ:α多样性的测度方法(下)
[J].生物多样性 ,1994 (4 ):231 -239 .
[本文引用: 1]
[15]
马克平 生物群落多样性的测度方法Ⅰ:α多样性的测度方法(上)
[J].生物多样性 ,1994 (3 ):162 -168 .
[16]
白永飞 ,李凌浩 ,王其兵 ,等 锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究
[J].植物生态学报 , 2000 ,24 (6 ):667 -673 .
[17]
Bai Y Wu J Clark C M et al Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from Inner Mongolia grasslands
[J].Global Change Biology ,2010 ,16 (1 ):358 -372 .
[18]
胡一鸣 ,梁健超 ,金崑 ,等 喜马拉雅山哺乳动物物种多样性垂直分布格局
[J].生物多样性 ,2018 ,26 (2 ):191 -201 .
[本文引用: 1]
[19]
段遍红 ,滕玉风 ,占玉芳 ,等 黑河桥湿地植物群落组成与多样性研究
[J].林业科技通讯 ,2016 (12 ):27 -30 .
[本文引用: 1]
[20]
梅象信 ,徐正会 ,张继玲 ,等 昆明西山森林公园东坡蚂蚁物种多样性研究
[J].林业科学研究 ,2006 ,19 (2 ):170 -176 .
[本文引用: 1]
[21]
韩高玲 ,霍建强 ,赵燕翘 ,等 鄂尔多斯高原砒砂岩地区草本物种组成及多样性
[J].中国沙漠 ,2023 ,43 (3 ):243 -251 .
URL
[本文引用: 1]
[22]
刘丽丹 ,谢应忠 ,邱开阳 宁夏盐池沙地3种植物群落的数量特征
[J].中国沙漠 ,2013 ,33 (2 ):343 -348 .
URL
[本文引用: 1]
[23]
赵啸龙 ,谢玉鸿 ,马旭君 ,等 科尔沁沙质草地不同恢复年限草本层群落结构及其与土壤理化性质的关系
[J].中国沙漠 ,2022 ,42 (2 ):134 -141 .
URL
[本文引用: 1]
[24]
张景光 ,李新荣 ,王新平 ,等 人工固沙区一年生植物小画眉草种群异速生长动态研究
[J].中国沙漠 ,2002 ,22 (6 ):85 -89 .
[本文引用: 1]
[25]
陈芙蓉 ,程积民 ,刘伟 ,等 不同干扰对黄土区典型草原物种多样性和生物量的影响
[J].生态学报 ,2013 ,33 (9 ):2856 -2866 .
[本文引用: 1]
[26]
王树力 ,范宁宁 浑善达克沙地植被恢复特征研究
[J].安徽农业科学 ,2014 ,42 (2 ):546 -548 .
[本文引用: 1]
[27]
张军红 ,李永华 ,姚斌 干旱·半干旱地区生物结皮对植被演替的影响
[J].安徽农业科学 ,2013 ,41 (10 ):4549 -4550 .
[本文引用: 1]
[28]
Zhang M Delgado-Baquerizo M Li G et al Experimental impacts of grazing on grassland biodiversity and function are explained by aridity
[J].Nature Communications ,2023 ,14 (1 ):5040 .
[本文引用: 1]
[29]
孙宗玖 ,朱进忠 ,张鲜花 短期放牧强度对昭苏草甸草原植被特征及多样性影响
[J].新疆农业大学学报 ,2014 ,37 (1 ):35 -39 .
[本文引用: 1]
[30]
郭振宁 ,孙世贤 ,擅建国 ,等 放牧强度对荒漠草原土壤含水量及群落特征的影响
[J].中国草地学报 ,2022 ,44 (6 ):27 -35 .
[本文引用: 1]
[31]
吕朋 ,左小安 ,张婧 ,等 放牧强度对科尔沁沙地沙质草地植被的影响
[J].中国沙漠 ,2016 ,36 (1 ):34 -39 .
URL
[本文引用: 1]
[32]
殷国梅 ,王明盈 ,薛艳林 ,等 草甸草原区不同放牧方式对植被群落特征的影响
[J].中国草地学报 ,2013 ,35 (2 ):89 -93 .
[本文引用: 1]
[33]
马凯 ,李永宁 ,金辉 ,等 不同生境类型金莲花群落物种多样性比较
[J].草业科学 ,2011 ,28 (8 ):1467 -1472 .
[本文引用: 1]
[34]
Wang W J Qiu L Zu Y G et al Changes in soil organic carbon, nitrogen,pH and bulk density with the development of larch (Larix gmelinii ) plantations in China
[J].Global Change Biology ,2011 ,17 (8 ):2657 -2676 .
[本文引用: 1]
[35]
都军 ,李宜轩 ,杨晓霞 ,等 腾格里沙漠东南缘生物土壤结皮对土壤理化性质的影响
[J].中国沙漠 ,2018 ,38 (1 ):111 -116 .
URL
[本文引用: 1]
[36]
Lane R W Menon M McQuaid J B et al Laboratory analysis of the effects of elevated atmospheric carbon dioxide on respiration in biological soil crusts
[J].Journal of Arid Environments ,2013 ,98 (5 ):52 -59 .
[本文引用: 1]
[37]
赵文迪 ,秦富仓 ,杨振奇 ,等 放牧对荒漠草原区植被、土壤及根系的影响研究综述
[J].内蒙古林业调查设计 ,2025 ,48 (1 ):97 -101 .
[本文引用: 1]
[38]
刘宝 ,吴文峰 ,林思祖 ,等 中亚热带4种林分类型土壤微生物生物量碳氮特征及季节变化
[J].应用生态学报 ,2019 ,30 (6 ):1901 -1910 .
[本文引用: 1]
[39]
Yang H Yuan Y Zhang Q et al Changes in soil organic carbon, total nitrogen, and abundance of arbuscular mycorrhizal fungi along a large-scale aridity gradient
[J].Catena ,2011 ,87 (1 ):70 -77 .
[本文引用: 1]
[40]
伍晶 ,刘世章 ,陈兰英 ,等 围封对高寒草甸轻度退化和沙化草地植物群落及土壤理化性质的影响
[J/OL].草原与草坪 ,1 -16 [2025-06-11 ]..
[本文引用: 1]
[41]
赵文智 ,程国栋 干旱区生态水文过程研究若干问题评述
[J].科学通报 ,2001 ,46 (44 ):1851 -1857 .
[本文引用: 1]
[42]
宁婷 ,张定海 ,赵有益 ,等 腾格里沙漠土壤水分含量与地形、植被的关系
[J].中国沙漠 ,2024 ,44 (5 ):133 -142 .
URL
[本文引用: 1]
[43]
王佳 ,田青 ,王理德 ,等 民勤青土湖区不同年限退耕地对土壤水分与物种多样性的影响
[J].干旱区研究 ,2022 ,39 (2 ):605 -614 .
[本文引用: 1]
宁夏盐池北部沙化土地综合治理决策支持系统设计与实现
1
2013
... 宁夏盐池位处毛乌素沙地西端,因不合理土地利用,区域生态系统严重失衡,风沙区域有蔓延趋势[1 ] .据全国沙化土地监测结果,盐池沙化土地面积达到486 932 hm2 ,占宁夏沙化土地面积的39.4%,占盐池面积的 68.3%[2 ] ,土地沙化已成为限制该地区经济发展的主要因素[3 ] .植物重建与恢复作为沙化土地修复的有效手段,自20世纪60年代以来,该区域通过大规模人工柠条(Caragana korshinskii )灌木林的营造,提高了土壤有机质的输入,显著改善了土壤物理、化学及微生物属性,增加了生物多样性. ...
宁夏盐池县沙区植被覆盖与植物多样性变化
1
2021
... 宁夏盐池位处毛乌素沙地西端,因不合理土地利用,区域生态系统严重失衡,风沙区域有蔓延趋势[1 ] .据全国沙化土地监测结果,盐池沙化土地面积达到486 932 hm2 ,占宁夏沙化土地面积的39.4%,占盐池面积的 68.3%[2 ] ,土地沙化已成为限制该地区经济发展的主要因素[3 ] .植物重建与恢复作为沙化土地修复的有效手段,自20世纪60年代以来,该区域通过大规模人工柠条(Caragana korshinskii )灌木林的营造,提高了土壤有机质的输入,显著改善了土壤物理、化学及微生物属性,增加了生物多样性. ...
盐池县沙化土地及其利用动态变化分析
1
2004
... 宁夏盐池位处毛乌素沙地西端,因不合理土地利用,区域生态系统严重失衡,风沙区域有蔓延趋势[1 ] .据全国沙化土地监测结果,盐池沙化土地面积达到486 932 hm2 ,占宁夏沙化土地面积的39.4%,占盐池面积的 68.3%[2 ] ,土地沙化已成为限制该地区经济发展的主要因素[3 ] .植物重建与恢复作为沙化土地修复的有效手段,自20世纪60年代以来,该区域通过大规模人工柠条(Caragana korshinskii )灌木林的营造,提高了土壤有机质的输入,显著改善了土壤物理、化学及微生物属性,增加了生物多样性. ...
刘任涛,腾格里沙漠固沙造林后植被-土壤系统演变特征及生态效应评价
1
2020
... 人工固沙植物群落演替呈现明显的时间依赖性.腾格里沙漠东南缘人工固沙植被区的研究显示,随着时间推移,植被物种丰富度和多样性指数呈上升趋势;土壤理化性质得到改善,风沙活动减少,为植物种子的传播和定居创造了有利条件,从而逐渐提升物种丰富度[4 ] .类似地,高寒沙区的研究也显示,在人工治理初期以柠条和油蒿(Artemisia ordosica )为主的人工固沙植被区,植物群落的物种丰富度、Shannon多样性指数和Pielou均匀度指数均不断增加并逐渐达到峰值,随后赖草(Leymus racemosus )占优势地位[5 ] .植被恢复过程中,土壤环境改善、种间竞争与生态位分化等因素共同作用,推动群落结构复杂化与功能完善化,进一步证实植被重建能够有效促进土壤环境改良和植被群落恢复.在干旱沙区,柠条作为建群种,能够降低地表温度、减少水分蒸发,有效固定沙土,减少风蚀和水蚀,从而提高土壤质量[6 ] .对樟子松(Pinus sylvestris )人工林林下植物多样性和生物量动态变化的研究显示,随着林龄增加,林下草本植物科、属、种数呈先减少后增加的规律,林下草本层Shannon多样性指数、Simpson优势度指数均随着林龄增加呈先减少后增加的规律,Pielou均匀度指数随林龄增加而逐渐增加[7 ] .这说明植物群落随植被的动态演替逐渐趋于稳定.那么在本研究区,随着柠条林建植年限的增加,其对草本植物群落结构、多样性指数以及土壤理化性质如何影响,在干旱区是否具有明显的演替特征等有待深入研究. ...
高寒沙区植被人工修复与种子植物物种多样性的变化
1
2004
... 人工固沙植物群落演替呈现明显的时间依赖性.腾格里沙漠东南缘人工固沙植被区的研究显示,随着时间推移,植被物种丰富度和多样性指数呈上升趋势;土壤理化性质得到改善,风沙活动减少,为植物种子的传播和定居创造了有利条件,从而逐渐提升物种丰富度[4 ] .类似地,高寒沙区的研究也显示,在人工治理初期以柠条和油蒿(Artemisia ordosica )为主的人工固沙植被区,植物群落的物种丰富度、Shannon多样性指数和Pielou均匀度指数均不断增加并逐渐达到峰值,随后赖草(Leymus racemosus )占优势地位[5 ] .植被恢复过程中,土壤环境改善、种间竞争与生态位分化等因素共同作用,推动群落结构复杂化与功能完善化,进一步证实植被重建能够有效促进土壤环境改良和植被群落恢复.在干旱沙区,柠条作为建群种,能够降低地表温度、减少水分蒸发,有效固定沙土,减少风蚀和水蚀,从而提高土壤质量[6 ] .对樟子松(Pinus sylvestris )人工林林下植物多样性和生物量动态变化的研究显示,随着林龄增加,林下草本植物科、属、种数呈先减少后增加的规律,林下草本层Shannon多样性指数、Simpson优势度指数均随着林龄增加呈先减少后增加的规律,Pielou均匀度指数随林龄增加而逐渐增加[7 ] .这说明植物群落随植被的动态演替逐渐趋于稳定.那么在本研究区,随着柠条林建植年限的增加,其对草本植物群落结构、多样性指数以及土壤理化性质如何影响,在干旱区是否具有明显的演替特征等有待深入研究. ...
毛乌素沙地不同恢复年限柠条固沙林微生物群落特征及其影响因素
1
2024
... 人工固沙植物群落演替呈现明显的时间依赖性.腾格里沙漠东南缘人工固沙植被区的研究显示,随着时间推移,植被物种丰富度和多样性指数呈上升趋势;土壤理化性质得到改善,风沙活动减少,为植物种子的传播和定居创造了有利条件,从而逐渐提升物种丰富度[4 ] .类似地,高寒沙区的研究也显示,在人工治理初期以柠条和油蒿(Artemisia ordosica )为主的人工固沙植被区,植物群落的物种丰富度、Shannon多样性指数和Pielou均匀度指数均不断增加并逐渐达到峰值,随后赖草(Leymus racemosus )占优势地位[5 ] .植被恢复过程中,土壤环境改善、种间竞争与生态位分化等因素共同作用,推动群落结构复杂化与功能完善化,进一步证实植被重建能够有效促进土壤环境改良和植被群落恢复.在干旱沙区,柠条作为建群种,能够降低地表温度、减少水分蒸发,有效固定沙土,减少风蚀和水蚀,从而提高土壤质量[6 ] .对樟子松(Pinus sylvestris )人工林林下植物多样性和生物量动态变化的研究显示,随着林龄增加,林下草本植物科、属、种数呈先减少后增加的规律,林下草本层Shannon多样性指数、Simpson优势度指数均随着林龄增加呈先减少后增加的规律,Pielou均匀度指数随林龄增加而逐渐增加[7 ] .这说明植物群落随植被的动态演替逐渐趋于稳定.那么在本研究区,随着柠条林建植年限的增加,其对草本植物群落结构、多样性指数以及土壤理化性质如何影响,在干旱区是否具有明显的演替特征等有待深入研究. ...
毛乌素沙地不同林龄樟子松人工林林下植物多样性和生物量的动态变化
1
2024
... 人工固沙植物群落演替呈现明显的时间依赖性.腾格里沙漠东南缘人工固沙植被区的研究显示,随着时间推移,植被物种丰富度和多样性指数呈上升趋势;土壤理化性质得到改善,风沙活动减少,为植物种子的传播和定居创造了有利条件,从而逐渐提升物种丰富度[4 ] .类似地,高寒沙区的研究也显示,在人工治理初期以柠条和油蒿(Artemisia ordosica )为主的人工固沙植被区,植物群落的物种丰富度、Shannon多样性指数和Pielou均匀度指数均不断增加并逐渐达到峰值,随后赖草(Leymus racemosus )占优势地位[5 ] .植被恢复过程中,土壤环境改善、种间竞争与生态位分化等因素共同作用,推动群落结构复杂化与功能完善化,进一步证实植被重建能够有效促进土壤环境改良和植被群落恢复.在干旱沙区,柠条作为建群种,能够降低地表温度、减少水分蒸发,有效固定沙土,减少风蚀和水蚀,从而提高土壤质量[6 ] .对樟子松(Pinus sylvestris )人工林林下植物多样性和生物量动态变化的研究显示,随着林龄增加,林下草本植物科、属、种数呈先减少后增加的规律,林下草本层Shannon多样性指数、Simpson优势度指数均随着林龄增加呈先减少后增加的规律,Pielou均匀度指数随林龄增加而逐渐增加[7 ] .这说明植物群落随植被的动态演替逐渐趋于稳定.那么在本研究区,随着柠条林建植年限的增加,其对草本植物群落结构、多样性指数以及土壤理化性质如何影响,在干旱区是否具有明显的演替特征等有待深入研究. ...
不同牧压强度对草原土壤水分、养分及其地上生物量的影响
1
2000
... 除了植被恢复时间外,放牧对植被演替过程的影响同样不可忽视.放牧改变土壤水分和养分状况,导致植被高度、盖度及地上生物量的降低[8 ] .盐池自2002年开始采取水土保持生态修复以来,植被覆盖度从修复前的30%提升到目前的65%[9 ] .尽管过度放牧会导致植被退化和土壤侵蚀,适度放牧却可能促进草本植物的生长[10 ] .在本研究区域内,部分样地仍然存在放牧现象,需要了解放牧如何影响该地区草本群落的动态变化,以及植被恢复过程中群落结构和多样性.因此,本研究采用时空替代法,重点关注人工种植柠条灌木林,系统探究不同年限下草本群落结构、Shannon多样性指数、Simpson多样性指数、Pielou均匀度指数以及土壤理化性质的变化特征.旨在明晰人工固沙植物群落随时间的变化规律,揭示驱动群落变化的关键因子与影响因素,为沙化地区的生态修复实践提供科学依据与理论支持. ...
盐池县抗旱治沙生态修复的探索与实践
1
2011
... 除了植被恢复时间外,放牧对植被演替过程的影响同样不可忽视.放牧改变土壤水分和养分状况,导致植被高度、盖度及地上生物量的降低[8 ] .盐池自2002年开始采取水土保持生态修复以来,植被覆盖度从修复前的30%提升到目前的65%[9 ] .尽管过度放牧会导致植被退化和土壤侵蚀,适度放牧却可能促进草本植物的生长[10 ] .在本研究区域内,部分样地仍然存在放牧现象,需要了解放牧如何影响该地区草本群落的动态变化,以及植被恢复过程中群落结构和多样性.因此,本研究采用时空替代法,重点关注人工种植柠条灌木林,系统探究不同年限下草本群落结构、Shannon多样性指数、Simpson多样性指数、Pielou均匀度指数以及土壤理化性质的变化特征.旨在明晰人工固沙植物群落随时间的变化规律,揭示驱动群落变化的关键因子与影响因素,为沙化地区的生态修复实践提供科学依据与理论支持. ...
放牧对草地土壤种子库的影响
1
2014
... 除了植被恢复时间外,放牧对植被演替过程的影响同样不可忽视.放牧改变土壤水分和养分状况,导致植被高度、盖度及地上生物量的降低[8 ] .盐池自2002年开始采取水土保持生态修复以来,植被覆盖度从修复前的30%提升到目前的65%[9 ] .尽管过度放牧会导致植被退化和土壤侵蚀,适度放牧却可能促进草本植物的生长[10 ] .在本研究区域内,部分样地仍然存在放牧现象,需要了解放牧如何影响该地区草本群落的动态变化,以及植被恢复过程中群落结构和多样性.因此,本研究采用时空替代法,重点关注人工种植柠条灌木林,系统探究不同年限下草本群落结构、Shannon多样性指数、Simpson多样性指数、Pielou均匀度指数以及土壤理化性质的变化特征.旨在明晰人工固沙植物群落随时间的变化规律,揭示驱动群落变化的关键因子与影响因素,为沙化地区的生态修复实践提供科学依据与理论支持. ...
沙区生物土壤结皮演替对表层土壤养分和微生物群落组成的影响
1
2024
... 土壤pH采用玻璃电极法测定,有机碳采用重铬酸钾容量法-外加热法测定,全氮采用凯氏定氮法测定,全磷采用酸消解-钼锑抗分光光度法测定[11 ] ;微生物量碳氮采用氯仿熏蒸法提取-碳氮分析仪测定[12 ] ;含水率采用烘干法测定[13 ] . ...
An extraction method for measuring soil microbial biomass
1
1987
... 土壤pH采用玻璃电极法测定,有机碳采用重铬酸钾容量法-外加热法测定,全氮采用凯氏定氮法测定,全磷采用酸消解-钼锑抗分光光度法测定[11 ] ;微生物量碳氮采用氯仿熏蒸法提取-碳氮分析仪测定[12 ] ;含水率采用烘干法测定[13 ] . ...
西北干旱区藓类结皮覆盖下土壤多功能性特征及影响因子
1
2024
... 土壤pH采用玻璃电极法测定,有机碳采用重铬酸钾容量法-外加热法测定,全氮采用凯氏定氮法测定,全磷采用酸消解-钼锑抗分光光度法测定[11 ] ;微生物量碳氮采用氯仿熏蒸法提取-碳氮分析仪测定[12 ] ;含水率采用烘干法测定[13 ] . ...
生物群落多样性的测度方法Ⅰ:α多样性的测度方法(下)
1
1994
... 根据样方调查的草本及木本物种的种类、盖度、多度计算出物种丰富度(R )、 Shannon多样性指数(H )、Simpson指数(D ),公式如下[14 -18 ] : ...
生物群落多样性的测度方法Ⅰ:α多样性的测度方法(上)
1994
锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究
2000
Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from Inner Mongolia grasslands
2010
喜马拉雅山哺乳动物物种多样性垂直分布格局
1
2018
... 根据样方调查的草本及木本物种的种类、盖度、多度计算出物种丰富度(R )、 Shannon多样性指数(H )、Simpson指数(D ),公式如下[14 -18 ] : ...
黑河桥湿地植物群落组成与多样性研究
1
2016
... 基于Shannon多样性指数计算Pielou均匀度指数,公式如下[19 ] : ...
昆明西山森林公园东坡蚂蚁物种多样性研究
1
2006
... 群落相似性采用Jaccard相似性系数计算,公式如下[20 ] : ...
鄂尔多斯高原砒砂岩地区草本物种组成及多样性
1
2023
... 通过对4个年份的人工柠条灌木林的调查发现,研究区共记录草本植物43种,隶属于19科、39属,总体上物种组成较为简单.这主要是由于研究区位于鄂尔多斯台地向黄土丘陵区的过渡地带,该地区常年干旱少雨,水分匮乏,影响了沙化土地的恢复以及人工固沙植被的稳定性,限制了植物种类的丰富度[21 -22 ] .随着植被恢复年限的增加,草本植被的盖度总体呈现上升趋势.其中,刺蓬、虫实、雾冰藜等一年生植物的比例逐渐减少,而诸如苦豆子和茵陈蒿这类多年生草本植物增多.多年生植物凭借发达的根系,能够高效吸收深层土壤水分和养分,其较长的生命周期有益于生物量的持续积累,从而在草本群落中逐渐占据主导地位[23 ] .如茵陈蒿、小画眉、刺蓬、针茅、地锦草等在沙区植被演替过程中逐渐成为优势种(表2 ),这与其强大的生命力和耐旱特性有关.例如,小画眉草发育出较多的浅层须根系,能够在降雨季节快速吸收水分,这种资源利用策略使其在沙区植被演替中具有显著竞争优势[24 ] . ...
宁夏盐池沙地3种植物群落的数量特征
1
2013
... 通过对4个年份的人工柠条灌木林的调查发现,研究区共记录草本植物43种,隶属于19科、39属,总体上物种组成较为简单.这主要是由于研究区位于鄂尔多斯台地向黄土丘陵区的过渡地带,该地区常年干旱少雨,水分匮乏,影响了沙化土地的恢复以及人工固沙植被的稳定性,限制了植物种类的丰富度[21 -22 ] .随着植被恢复年限的增加,草本植被的盖度总体呈现上升趋势.其中,刺蓬、虫实、雾冰藜等一年生植物的比例逐渐减少,而诸如苦豆子和茵陈蒿这类多年生草本植物增多.多年生植物凭借发达的根系,能够高效吸收深层土壤水分和养分,其较长的生命周期有益于生物量的持续积累,从而在草本群落中逐渐占据主导地位[23 ] .如茵陈蒿、小画眉、刺蓬、针茅、地锦草等在沙区植被演替过程中逐渐成为优势种(表2 ),这与其强大的生命力和耐旱特性有关.例如,小画眉草发育出较多的浅层须根系,能够在降雨季节快速吸收水分,这种资源利用策略使其在沙区植被演替中具有显著竞争优势[24 ] . ...
科尔沁沙质草地不同恢复年限草本层群落结构及其与土壤理化性质的关系
1
2022
... 通过对4个年份的人工柠条灌木林的调查发现,研究区共记录草本植物43种,隶属于19科、39属,总体上物种组成较为简单.这主要是由于研究区位于鄂尔多斯台地向黄土丘陵区的过渡地带,该地区常年干旱少雨,水分匮乏,影响了沙化土地的恢复以及人工固沙植被的稳定性,限制了植物种类的丰富度[21 -22 ] .随着植被恢复年限的增加,草本植被的盖度总体呈现上升趋势.其中,刺蓬、虫实、雾冰藜等一年生植物的比例逐渐减少,而诸如苦豆子和茵陈蒿这类多年生草本植物增多.多年生植物凭借发达的根系,能够高效吸收深层土壤水分和养分,其较长的生命周期有益于生物量的持续积累,从而在草本群落中逐渐占据主导地位[23 ] .如茵陈蒿、小画眉、刺蓬、针茅、地锦草等在沙区植被演替过程中逐渐成为优势种(表2 ),这与其强大的生命力和耐旱特性有关.例如,小画眉草发育出较多的浅层须根系,能够在降雨季节快速吸收水分,这种资源利用策略使其在沙区植被演替中具有显著竞争优势[24 ] . ...
人工固沙区一年生植物小画眉草种群异速生长动态研究
1
2002
... 通过对4个年份的人工柠条灌木林的调查发现,研究区共记录草本植物43种,隶属于19科、39属,总体上物种组成较为简单.这主要是由于研究区位于鄂尔多斯台地向黄土丘陵区的过渡地带,该地区常年干旱少雨,水分匮乏,影响了沙化土地的恢复以及人工固沙植被的稳定性,限制了植物种类的丰富度[21 -22 ] .随着植被恢复年限的增加,草本植被的盖度总体呈现上升趋势.其中,刺蓬、虫实、雾冰藜等一年生植物的比例逐渐减少,而诸如苦豆子和茵陈蒿这类多年生草本植物增多.多年生植物凭借发达的根系,能够高效吸收深层土壤水分和养分,其较长的生命周期有益于生物量的持续积累,从而在草本群落中逐渐占据主导地位[23 ] .如茵陈蒿、小画眉、刺蓬、针茅、地锦草等在沙区植被演替过程中逐渐成为优势种(表2 ),这与其强大的生命力和耐旱特性有关.例如,小画眉草发育出较多的浅层须根系,能够在降雨季节快速吸收水分,这种资源利用策略使其在沙区植被演替中具有显著竞争优势[24 ] . ...
不同干扰对黄土区典型草原物种多样性和生物量的影响
1
2013
... 物种多样性对维持生态平衡和人类生存至关重要,而多样性指数用于衡量生态系统中物种的多样性和异质性[25 ] .本研究显示,草本物种丰富度随植被恢复年限的增加具有波动性,且1984年和2015年样地的物种丰富度、Shannon多样性指数和Simpson多样性指数均高于其余两个样地(图1 ),与王树力等[26 ] 关于植被物种丰富度随着植被恢复年限的增加而增大的结果不同.在柠条灌木林重建后,沙地中自然侵入的植物种数逐渐增多,尤其是一些先锋草本植物的拓殖,对沙丘表面的固定起到了关键作用,有效改善了土壤水分和养分状况,为植被的定居与繁殖提供了理想的环境,从而提升了物种的丰富度.但在本研究中,2015年样地的丰富度及多样性指数最高,1984年次之.1984年样地中生物土壤结皮盖度超过90%,且存在较多苔藓结皮,其致密结构阻碍降水入渗和种子着床,改变了土壤中降水的再分配过程,使水分浅层化[27 ] ,进而引起草本植物退化.此外,致密的结皮组织还会影响草本种子的定殖.然而,2015年样地中较高土壤水分和较少外界干扰,促进了草本植物的快速生长和扩散,表现为更高的物种丰富度、Shannon多样性指数和Simpson多样性指数.这表明,在植被恢复的过程中,虽然植被恢复年限增加有利于草本群落的发展,但生物土壤结皮在这个过程中扮演着不可忽视的角色,对植被群落的动态有一定的调节作用. ...
浑善达克沙地植被恢复特征研究
1
2014
... 物种多样性对维持生态平衡和人类生存至关重要,而多样性指数用于衡量生态系统中物种的多样性和异质性[25 ] .本研究显示,草本物种丰富度随植被恢复年限的增加具有波动性,且1984年和2015年样地的物种丰富度、Shannon多样性指数和Simpson多样性指数均高于其余两个样地(图1 ),与王树力等[26 ] 关于植被物种丰富度随着植被恢复年限的增加而增大的结果不同.在柠条灌木林重建后,沙地中自然侵入的植物种数逐渐增多,尤其是一些先锋草本植物的拓殖,对沙丘表面的固定起到了关键作用,有效改善了土壤水分和养分状况,为植被的定居与繁殖提供了理想的环境,从而提升了物种的丰富度.但在本研究中,2015年样地的丰富度及多样性指数最高,1984年次之.1984年样地中生物土壤结皮盖度超过90%,且存在较多苔藓结皮,其致密结构阻碍降水入渗和种子着床,改变了土壤中降水的再分配过程,使水分浅层化[27 ] ,进而引起草本植物退化.此外,致密的结皮组织还会影响草本种子的定殖.然而,2015年样地中较高土壤水分和较少外界干扰,促进了草本植物的快速生长和扩散,表现为更高的物种丰富度、Shannon多样性指数和Simpson多样性指数.这表明,在植被恢复的过程中,虽然植被恢复年限增加有利于草本群落的发展,但生物土壤结皮在这个过程中扮演着不可忽视的角色,对植被群落的动态有一定的调节作用. ...
干旱·半干旱地区生物结皮对植被演替的影响
1
2013
... 物种多样性对维持生态平衡和人类生存至关重要,而多样性指数用于衡量生态系统中物种的多样性和异质性[25 ] .本研究显示,草本物种丰富度随植被恢复年限的增加具有波动性,且1984年和2015年样地的物种丰富度、Shannon多样性指数和Simpson多样性指数均高于其余两个样地(图1 ),与王树力等[26 ] 关于植被物种丰富度随着植被恢复年限的增加而增大的结果不同.在柠条灌木林重建后,沙地中自然侵入的植物种数逐渐增多,尤其是一些先锋草本植物的拓殖,对沙丘表面的固定起到了关键作用,有效改善了土壤水分和养分状况,为植被的定居与繁殖提供了理想的环境,从而提升了物种的丰富度.但在本研究中,2015年样地的丰富度及多样性指数最高,1984年次之.1984年样地中生物土壤结皮盖度超过90%,且存在较多苔藓结皮,其致密结构阻碍降水入渗和种子着床,改变了土壤中降水的再分配过程,使水分浅层化[27 ] ,进而引起草本植物退化.此外,致密的结皮组织还会影响草本种子的定殖.然而,2015年样地中较高土壤水分和较少外界干扰,促进了草本植物的快速生长和扩散,表现为更高的物种丰富度、Shannon多样性指数和Simpson多样性指数.这表明,在植被恢复的过程中,虽然植被恢复年限增加有利于草本群落的发展,但生物土壤结皮在这个过程中扮演着不可忽视的角色,对植被群落的动态有一定的调节作用. ...
Experimental impacts of grazing on grassland biodiversity and function are explained by aridity
1
2023
... 此外,值得注意的是,1996年和2005年样地的草本物种丰富度、多样性指数、地上生物量及凋落物均低于1984年和2015年的样地(表2 、图1 ).调查显示,1996年和2005年样地存在放牧现象.放牧显著影响了草本植被的物种丰富度、多样性及凋落物(表4 ).许多研究表明,放牧会破坏土壤结构,导致土壤保水保肥能力下降,并加剧土壤养分流失.这种变化使得植物无法获得生长所需的养分和水分,导致其生长和繁殖受到限制,从而降低了植被的丰富度和多样性[28 ] .这与孙宗玖等[29 ] 的结论一致,他们发现放牧后草本群落盖度及地上生物量明显降低,并显著影响了Shannon 指数.尽管有研究指出,适度放牧可增加植被多样性,促进荒漠草原的可持续利用[30 ] .但在本研究区域,由于环境恶劣和水分极度缺乏,放牧干扰使得原本脆弱的生态系统变得更加紧张,因此抑制作用更加突出.然而,本研究中放牧对草本植被的 Simpson多样性指数及Pielou均匀度指数的影响并不显著,这可能与放牧的时间、强度以及种源等因素有关[31 -32 ] . ...
短期放牧强度对昭苏草甸草原植被特征及多样性影响
1
2014
... 此外,值得注意的是,1996年和2005年样地的草本物种丰富度、多样性指数、地上生物量及凋落物均低于1984年和2015年的样地(表2 、图1 ).调查显示,1996年和2005年样地存在放牧现象.放牧显著影响了草本植被的物种丰富度、多样性及凋落物(表4 ).许多研究表明,放牧会破坏土壤结构,导致土壤保水保肥能力下降,并加剧土壤养分流失.这种变化使得植物无法获得生长所需的养分和水分,导致其生长和繁殖受到限制,从而降低了植被的丰富度和多样性[28 ] .这与孙宗玖等[29 ] 的结论一致,他们发现放牧后草本群落盖度及地上生物量明显降低,并显著影响了Shannon 指数.尽管有研究指出,适度放牧可增加植被多样性,促进荒漠草原的可持续利用[30 ] .但在本研究区域,由于环境恶劣和水分极度缺乏,放牧干扰使得原本脆弱的生态系统变得更加紧张,因此抑制作用更加突出.然而,本研究中放牧对草本植被的 Simpson多样性指数及Pielou均匀度指数的影响并不显著,这可能与放牧的时间、强度以及种源等因素有关[31 -32 ] . ...
放牧强度对荒漠草原土壤含水量及群落特征的影响
1
2022
... 此外,值得注意的是,1996年和2005年样地的草本物种丰富度、多样性指数、地上生物量及凋落物均低于1984年和2015年的样地(表2 、图1 ).调查显示,1996年和2005年样地存在放牧现象.放牧显著影响了草本植被的物种丰富度、多样性及凋落物(表4 ).许多研究表明,放牧会破坏土壤结构,导致土壤保水保肥能力下降,并加剧土壤养分流失.这种变化使得植物无法获得生长所需的养分和水分,导致其生长和繁殖受到限制,从而降低了植被的丰富度和多样性[28 ] .这与孙宗玖等[29 ] 的结论一致,他们发现放牧后草本群落盖度及地上生物量明显降低,并显著影响了Shannon 指数.尽管有研究指出,适度放牧可增加植被多样性,促进荒漠草原的可持续利用[30 ] .但在本研究区域,由于环境恶劣和水分极度缺乏,放牧干扰使得原本脆弱的生态系统变得更加紧张,因此抑制作用更加突出.然而,本研究中放牧对草本植被的 Simpson多样性指数及Pielou均匀度指数的影响并不显著,这可能与放牧的时间、强度以及种源等因素有关[31 -32 ] . ...
放牧强度对科尔沁沙地沙质草地植被的影响
1
2016
... 此外,值得注意的是,1996年和2005年样地的草本物种丰富度、多样性指数、地上生物量及凋落物均低于1984年和2015年的样地(表2 、图1 ).调查显示,1996年和2005年样地存在放牧现象.放牧显著影响了草本植被的物种丰富度、多样性及凋落物(表4 ).许多研究表明,放牧会破坏土壤结构,导致土壤保水保肥能力下降,并加剧土壤养分流失.这种变化使得植物无法获得生长所需的养分和水分,导致其生长和繁殖受到限制,从而降低了植被的丰富度和多样性[28 ] .这与孙宗玖等[29 ] 的结论一致,他们发现放牧后草本群落盖度及地上生物量明显降低,并显著影响了Shannon 指数.尽管有研究指出,适度放牧可增加植被多样性,促进荒漠草原的可持续利用[30 ] .但在本研究区域,由于环境恶劣和水分极度缺乏,放牧干扰使得原本脆弱的生态系统变得更加紧张,因此抑制作用更加突出.然而,本研究中放牧对草本植被的 Simpson多样性指数及Pielou均匀度指数的影响并不显著,这可能与放牧的时间、强度以及种源等因素有关[31 -32 ] . ...
草甸草原区不同放牧方式对植被群落特征的影响
1
2013
... 此外,值得注意的是,1996年和2005年样地的草本物种丰富度、多样性指数、地上生物量及凋落物均低于1984年和2015年的样地(表2 、图1 ).调查显示,1996年和2005年样地存在放牧现象.放牧显著影响了草本植被的物种丰富度、多样性及凋落物(表4 ).许多研究表明,放牧会破坏土壤结构,导致土壤保水保肥能力下降,并加剧土壤养分流失.这种变化使得植物无法获得生长所需的养分和水分,导致其生长和繁殖受到限制,从而降低了植被的丰富度和多样性[28 ] .这与孙宗玖等[29 ] 的结论一致,他们发现放牧后草本群落盖度及地上生物量明显降低,并显著影响了Shannon 指数.尽管有研究指出,适度放牧可增加植被多样性,促进荒漠草原的可持续利用[30 ] .但在本研究区域,由于环境恶劣和水分极度缺乏,放牧干扰使得原本脆弱的生态系统变得更加紧张,因此抑制作用更加突出.然而,本研究中放牧对草本植被的 Simpson多样性指数及Pielou均匀度指数的影响并不显著,这可能与放牧的时间、强度以及种源等因素有关[31 -32 ] . ...
不同生境类型金莲花群落物种多样性比较
1
2011
... 植物群落相似性指数用于量化植物群落之间的异质性,反映植物群落生境条件的异质性[33 ] .本研究结果表明,草本层不同恢复年限的群落样地间相似系数为0.139~0.412(表3 ),样地间整体相似性较低,草本群落结构存在较大差异.此外,由于封育措施,2015年样地的草本群落特征与恢复时间较长的1984年样地表现出较高相似性. ...
Changes in soil organic carbon, nitrogen,pH and bulk density with the development of larch (Larix gmelinii ) plantations in China
1
2011
... 土壤作为植物赖以生存的物质基础,其性质的变化能够反映植被的变化.本研究发现,除1996年样地外,土壤有机碳、全氮、微生物量碳氮的含量在植被恢复过程中不断富集,且随着土壤深度的增加而降低(表5 ).这一现象印证了植被重建对土壤的理化性质的改良.柠条灌木林通过凋落物输入和根系分泌物释放,促进微生物分解代谢,加速腐殖质形成,驱动碳氮养分的富集,从而更加有利于草本植物的生长[34 -35 ] .土壤pH则呈相反的变化趋势,可能是由于表层土壤中的植物呼吸作用释放出的二氧化碳会降低土壤pH[36 ] .而1996年样地因受放牧的影响,土壤pH升高而土壤养分含量均低于其余年限样地.这是因为放牧破坏土壤积盐与脱盐的平衡,使可溶性盐类迁移,并累积于土表,导致土壤pH值升高,从而引起土壤肥力下降[37 ] .同时,牲畜的踩踏对表层土壤造成严重破坏,也加剧了土壤养分流失,被破坏后的土壤难以维持植被的生长,进而导致草本植被丰富度和多样性降低. ...
腾格里沙漠东南缘生物土壤结皮对土壤理化性质的影响
1
2018
... 土壤作为植物赖以生存的物质基础,其性质的变化能够反映植被的变化.本研究发现,除1996年样地外,土壤有机碳、全氮、微生物量碳氮的含量在植被恢复过程中不断富集,且随着土壤深度的增加而降低(表5 ).这一现象印证了植被重建对土壤的理化性质的改良.柠条灌木林通过凋落物输入和根系分泌物释放,促进微生物分解代谢,加速腐殖质形成,驱动碳氮养分的富集,从而更加有利于草本植物的生长[34 -35 ] .土壤pH则呈相反的变化趋势,可能是由于表层土壤中的植物呼吸作用释放出的二氧化碳会降低土壤pH[36 ] .而1996年样地因受放牧的影响,土壤pH升高而土壤养分含量均低于其余年限样地.这是因为放牧破坏土壤积盐与脱盐的平衡,使可溶性盐类迁移,并累积于土表,导致土壤pH值升高,从而引起土壤肥力下降[37 ] .同时,牲畜的踩踏对表层土壤造成严重破坏,也加剧了土壤养分流失,被破坏后的土壤难以维持植被的生长,进而导致草本植被丰富度和多样性降低. ...
Laboratory analysis of the effects of elevated atmospheric carbon dioxide on respiration in biological soil crusts
1
2013
... 土壤作为植物赖以生存的物质基础,其性质的变化能够反映植被的变化.本研究发现,除1996年样地外,土壤有机碳、全氮、微生物量碳氮的含量在植被恢复过程中不断富集,且随着土壤深度的增加而降低(表5 ).这一现象印证了植被重建对土壤的理化性质的改良.柠条灌木林通过凋落物输入和根系分泌物释放,促进微生物分解代谢,加速腐殖质形成,驱动碳氮养分的富集,从而更加有利于草本植物的生长[34 -35 ] .土壤pH则呈相反的变化趋势,可能是由于表层土壤中的植物呼吸作用释放出的二氧化碳会降低土壤pH[36 ] .而1996年样地因受放牧的影响,土壤pH升高而土壤养分含量均低于其余年限样地.这是因为放牧破坏土壤积盐与脱盐的平衡,使可溶性盐类迁移,并累积于土表,导致土壤pH值升高,从而引起土壤肥力下降[37 ] .同时,牲畜的踩踏对表层土壤造成严重破坏,也加剧了土壤养分流失,被破坏后的土壤难以维持植被的生长,进而导致草本植被丰富度和多样性降低. ...
放牧对荒漠草原区植被、土壤及根系的影响研究综述
1
2025
... 土壤作为植物赖以生存的物质基础,其性质的变化能够反映植被的变化.本研究发现,除1996年样地外,土壤有机碳、全氮、微生物量碳氮的含量在植被恢复过程中不断富集,且随着土壤深度的增加而降低(表5 ).这一现象印证了植被重建对土壤的理化性质的改良.柠条灌木林通过凋落物输入和根系分泌物释放,促进微生物分解代谢,加速腐殖质形成,驱动碳氮养分的富集,从而更加有利于草本植物的生长[34 -35 ] .土壤pH则呈相反的变化趋势,可能是由于表层土壤中的植物呼吸作用释放出的二氧化碳会降低土壤pH[36 ] .而1996年样地因受放牧的影响,土壤pH升高而土壤养分含量均低于其余年限样地.这是因为放牧破坏土壤积盐与脱盐的平衡,使可溶性盐类迁移,并累积于土表,导致土壤pH值升高,从而引起土壤肥力下降[37 ] .同时,牲畜的踩踏对表层土壤造成严重破坏,也加剧了土壤养分流失,被破坏后的土壤难以维持植被的生长,进而导致草本植被丰富度和多样性降低. ...
中亚热带4种林分类型土壤微生物生物量碳氮特征及季节变化
1
2019
... 本研究通过冗余分析(RDA)研究不同植被恢复年限样地的植被-土壤关系,草本植被的盖度、地上生物量、丰富度、Shannon多样性指数和Simpson多样性指数与土壤有机碳、全氮、微生物量碳氮之间表现出较强的正相关性(图2 ).这表明土壤的有机碳、微生物量碳氮是驱动草本植被群落结构变化的关键环境因子.有研究指出,土壤微生物量碳氮在土壤养分的供应和转化过程中发挥着决定作用[38 ] ,他们不仅为植物提供必要的营养,还增强了植物的抗逆性,这对于植物的生长至关重要.同时,高含量的土壤全氮和有机碳能够促进植物生长,从而提高植物群落的多样性[39 ] .在高寒草甸退化和沙化草地中,植物群落的地上生物量、盖度、物种丰富度、Shannon多样性指数与土壤有机碳和全氮含量呈极显著正相关[40 ] .这与本研究中土壤全氮、有机碳含量与草本多度、盖度及生物量的正相关一致.在干旱半干旱地区,土壤含水量是植物生长和群落稳定性的关键限制因子[41 ] .充足的土壤水分不仅满足植物的水分需求,提高植物的成活率和生长速度,还影响土壤中养分的有效性,从而间接影响植物的生长.本研究发现,草本植被多度、盖度、地上生物量和丰富度与表层土壤(0~20 cm)水分有较强的相关性,此外,草本植被的Shannon多样性指数和Simpson多样性指数也与浅层土壤(20~60 cm)水分呈较强的相关性.这可能是因为草本植物更容易获取表层土壤(0~20 cm)水分,有利于种子的萌发、幼苗的生长以及植株的发育,进而提高了草本植被的多度、盖度和地上生物量.而浅层土壤(20~60 cm)水分为具有不同根系深度的草本植物提供了多样化的生存空间,一些根系较深的多年生草本植物利用浅层土壤水分,与主要依赖表层土壤水分的植物形成生态位分化,减少种间竞争,使更多种类的草本植物能够共存,从而提高了草本植被的多样性.这与腾格里沙漠土壤水分含量与植被的研究结果相似[42 ] .在民勤青土湖区退耕40年的研究中也发现草本植被的Shannon多样性指数与0~20 cm土层的土壤含水量呈极显著正相关,强调了表层土壤水分在植被自然演替中的关键性作用[43 ] . ...
Changes in soil organic carbon, total nitrogen, and abundance of arbuscular mycorrhizal fungi along a large-scale aridity gradient
1
2011
... 本研究通过冗余分析(RDA)研究不同植被恢复年限样地的植被-土壤关系,草本植被的盖度、地上生物量、丰富度、Shannon多样性指数和Simpson多样性指数与土壤有机碳、全氮、微生物量碳氮之间表现出较强的正相关性(图2 ).这表明土壤的有机碳、微生物量碳氮是驱动草本植被群落结构变化的关键环境因子.有研究指出,土壤微生物量碳氮在土壤养分的供应和转化过程中发挥着决定作用[38 ] ,他们不仅为植物提供必要的营养,还增强了植物的抗逆性,这对于植物的生长至关重要.同时,高含量的土壤全氮和有机碳能够促进植物生长,从而提高植物群落的多样性[39 ] .在高寒草甸退化和沙化草地中,植物群落的地上生物量、盖度、物种丰富度、Shannon多样性指数与土壤有机碳和全氮含量呈极显著正相关[40 ] .这与本研究中土壤全氮、有机碳含量与草本多度、盖度及生物量的正相关一致.在干旱半干旱地区,土壤含水量是植物生长和群落稳定性的关键限制因子[41 ] .充足的土壤水分不仅满足植物的水分需求,提高植物的成活率和生长速度,还影响土壤中养分的有效性,从而间接影响植物的生长.本研究发现,草本植被多度、盖度、地上生物量和丰富度与表层土壤(0~20 cm)水分有较强的相关性,此外,草本植被的Shannon多样性指数和Simpson多样性指数也与浅层土壤(20~60 cm)水分呈较强的相关性.这可能是因为草本植物更容易获取表层土壤(0~20 cm)水分,有利于种子的萌发、幼苗的生长以及植株的发育,进而提高了草本植被的多度、盖度和地上生物量.而浅层土壤(20~60 cm)水分为具有不同根系深度的草本植物提供了多样化的生存空间,一些根系较深的多年生草本植物利用浅层土壤水分,与主要依赖表层土壤水分的植物形成生态位分化,减少种间竞争,使更多种类的草本植物能够共存,从而提高了草本植被的多样性.这与腾格里沙漠土壤水分含量与植被的研究结果相似[42 ] .在民勤青土湖区退耕40年的研究中也发现草本植被的Shannon多样性指数与0~20 cm土层的土壤含水量呈极显著正相关,强调了表层土壤水分在植被自然演替中的关键性作用[43 ] . ...
围封对高寒草甸轻度退化和沙化草地植物群落及土壤理化性质的影响
1
1
... 本研究通过冗余分析(RDA)研究不同植被恢复年限样地的植被-土壤关系,草本植被的盖度、地上生物量、丰富度、Shannon多样性指数和Simpson多样性指数与土壤有机碳、全氮、微生物量碳氮之间表现出较强的正相关性(图2 ).这表明土壤的有机碳、微生物量碳氮是驱动草本植被群落结构变化的关键环境因子.有研究指出,土壤微生物量碳氮在土壤养分的供应和转化过程中发挥着决定作用[38 ] ,他们不仅为植物提供必要的营养,还增强了植物的抗逆性,这对于植物的生长至关重要.同时,高含量的土壤全氮和有机碳能够促进植物生长,从而提高植物群落的多样性[39 ] .在高寒草甸退化和沙化草地中,植物群落的地上生物量、盖度、物种丰富度、Shannon多样性指数与土壤有机碳和全氮含量呈极显著正相关[40 ] .这与本研究中土壤全氮、有机碳含量与草本多度、盖度及生物量的正相关一致.在干旱半干旱地区,土壤含水量是植物生长和群落稳定性的关键限制因子[41 ] .充足的土壤水分不仅满足植物的水分需求,提高植物的成活率和生长速度,还影响土壤中养分的有效性,从而间接影响植物的生长.本研究发现,草本植被多度、盖度、地上生物量和丰富度与表层土壤(0~20 cm)水分有较强的相关性,此外,草本植被的Shannon多样性指数和Simpson多样性指数也与浅层土壤(20~60 cm)水分呈较强的相关性.这可能是因为草本植物更容易获取表层土壤(0~20 cm)水分,有利于种子的萌发、幼苗的生长以及植株的发育,进而提高了草本植被的多度、盖度和地上生物量.而浅层土壤(20~60 cm)水分为具有不同根系深度的草本植物提供了多样化的生存空间,一些根系较深的多年生草本植物利用浅层土壤水分,与主要依赖表层土壤水分的植物形成生态位分化,减少种间竞争,使更多种类的草本植物能够共存,从而提高了草本植被的多样性.这与腾格里沙漠土壤水分含量与植被的研究结果相似[42 ] .在民勤青土湖区退耕40年的研究中也发现草本植被的Shannon多样性指数与0~20 cm土层的土壤含水量呈极显著正相关,强调了表层土壤水分在植被自然演替中的关键性作用[43 ] . ...
干旱区生态水文过程研究若干问题评述
1
2001
... 本研究通过冗余分析(RDA)研究不同植被恢复年限样地的植被-土壤关系,草本植被的盖度、地上生物量、丰富度、Shannon多样性指数和Simpson多样性指数与土壤有机碳、全氮、微生物量碳氮之间表现出较强的正相关性(图2 ).这表明土壤的有机碳、微生物量碳氮是驱动草本植被群落结构变化的关键环境因子.有研究指出,土壤微生物量碳氮在土壤养分的供应和转化过程中发挥着决定作用[38 ] ,他们不仅为植物提供必要的营养,还增强了植物的抗逆性,这对于植物的生长至关重要.同时,高含量的土壤全氮和有机碳能够促进植物生长,从而提高植物群落的多样性[39 ] .在高寒草甸退化和沙化草地中,植物群落的地上生物量、盖度、物种丰富度、Shannon多样性指数与土壤有机碳和全氮含量呈极显著正相关[40 ] .这与本研究中土壤全氮、有机碳含量与草本多度、盖度及生物量的正相关一致.在干旱半干旱地区,土壤含水量是植物生长和群落稳定性的关键限制因子[41 ] .充足的土壤水分不仅满足植物的水分需求,提高植物的成活率和生长速度,还影响土壤中养分的有效性,从而间接影响植物的生长.本研究发现,草本植被多度、盖度、地上生物量和丰富度与表层土壤(0~20 cm)水分有较强的相关性,此外,草本植被的Shannon多样性指数和Simpson多样性指数也与浅层土壤(20~60 cm)水分呈较强的相关性.这可能是因为草本植物更容易获取表层土壤(0~20 cm)水分,有利于种子的萌发、幼苗的生长以及植株的发育,进而提高了草本植被的多度、盖度和地上生物量.而浅层土壤(20~60 cm)水分为具有不同根系深度的草本植物提供了多样化的生存空间,一些根系较深的多年生草本植物利用浅层土壤水分,与主要依赖表层土壤水分的植物形成生态位分化,减少种间竞争,使更多种类的草本植物能够共存,从而提高了草本植被的多样性.这与腾格里沙漠土壤水分含量与植被的研究结果相似[42 ] .在民勤青土湖区退耕40年的研究中也发现草本植被的Shannon多样性指数与0~20 cm土层的土壤含水量呈极显著正相关,强调了表层土壤水分在植被自然演替中的关键性作用[43 ] . ...
腾格里沙漠土壤水分含量与地形、植被的关系
1
2024
... 本研究通过冗余分析(RDA)研究不同植被恢复年限样地的植被-土壤关系,草本植被的盖度、地上生物量、丰富度、Shannon多样性指数和Simpson多样性指数与土壤有机碳、全氮、微生物量碳氮之间表现出较强的正相关性(图2 ).这表明土壤的有机碳、微生物量碳氮是驱动草本植被群落结构变化的关键环境因子.有研究指出,土壤微生物量碳氮在土壤养分的供应和转化过程中发挥着决定作用[38 ] ,他们不仅为植物提供必要的营养,还增强了植物的抗逆性,这对于植物的生长至关重要.同时,高含量的土壤全氮和有机碳能够促进植物生长,从而提高植物群落的多样性[39 ] .在高寒草甸退化和沙化草地中,植物群落的地上生物量、盖度、物种丰富度、Shannon多样性指数与土壤有机碳和全氮含量呈极显著正相关[40 ] .这与本研究中土壤全氮、有机碳含量与草本多度、盖度及生物量的正相关一致.在干旱半干旱地区,土壤含水量是植物生长和群落稳定性的关键限制因子[41 ] .充足的土壤水分不仅满足植物的水分需求,提高植物的成活率和生长速度,还影响土壤中养分的有效性,从而间接影响植物的生长.本研究发现,草本植被多度、盖度、地上生物量和丰富度与表层土壤(0~20 cm)水分有较强的相关性,此外,草本植被的Shannon多样性指数和Simpson多样性指数也与浅层土壤(20~60 cm)水分呈较强的相关性.这可能是因为草本植物更容易获取表层土壤(0~20 cm)水分,有利于种子的萌发、幼苗的生长以及植株的发育,进而提高了草本植被的多度、盖度和地上生物量.而浅层土壤(20~60 cm)水分为具有不同根系深度的草本植物提供了多样化的生存空间,一些根系较深的多年生草本植物利用浅层土壤水分,与主要依赖表层土壤水分的植物形成生态位分化,减少种间竞争,使更多种类的草本植物能够共存,从而提高了草本植被的多样性.这与腾格里沙漠土壤水分含量与植被的研究结果相似[42 ] .在民勤青土湖区退耕40年的研究中也发现草本植被的Shannon多样性指数与0~20 cm土层的土壤含水量呈极显著正相关,强调了表层土壤水分在植被自然演替中的关键性作用[43 ] . ...
民勤青土湖区不同年限退耕地对土壤水分与物种多样性的影响
1
2022
... 本研究通过冗余分析(RDA)研究不同植被恢复年限样地的植被-土壤关系,草本植被的盖度、地上生物量、丰富度、Shannon多样性指数和Simpson多样性指数与土壤有机碳、全氮、微生物量碳氮之间表现出较强的正相关性(图2 ).这表明土壤的有机碳、微生物量碳氮是驱动草本植被群落结构变化的关键环境因子.有研究指出,土壤微生物量碳氮在土壤养分的供应和转化过程中发挥着决定作用[38 ] ,他们不仅为植物提供必要的营养,还增强了植物的抗逆性,这对于植物的生长至关重要.同时,高含量的土壤全氮和有机碳能够促进植物生长,从而提高植物群落的多样性[39 ] .在高寒草甸退化和沙化草地中,植物群落的地上生物量、盖度、物种丰富度、Shannon多样性指数与土壤有机碳和全氮含量呈极显著正相关[40 ] .这与本研究中土壤全氮、有机碳含量与草本多度、盖度及生物量的正相关一致.在干旱半干旱地区,土壤含水量是植物生长和群落稳定性的关键限制因子[41 ] .充足的土壤水分不仅满足植物的水分需求,提高植物的成活率和生长速度,还影响土壤中养分的有效性,从而间接影响植物的生长.本研究发现,草本植被多度、盖度、地上生物量和丰富度与表层土壤(0~20 cm)水分有较强的相关性,此外,草本植被的Shannon多样性指数和Simpson多样性指数也与浅层土壤(20~60 cm)水分呈较强的相关性.这可能是因为草本植物更容易获取表层土壤(0~20 cm)水分,有利于种子的萌发、幼苗的生长以及植株的发育,进而提高了草本植被的多度、盖度和地上生物量.而浅层土壤(20~60 cm)水分为具有不同根系深度的草本植物提供了多样化的生存空间,一些根系较深的多年生草本植物利用浅层土壤水分,与主要依赖表层土壤水分的植物形成生态位分化,减少种间竞争,使更多种类的草本植物能够共存,从而提高了草本植被的多样性.这与腾格里沙漠土壤水分含量与植被的研究结果相似[42 ] .在民勤青土湖区退耕40年的研究中也发现草本植被的Shannon多样性指数与0~20 cm土层的土壤含水量呈极显著正相关,强调了表层土壤水分在植被自然演替中的关键性作用[43 ] . ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}