差巴嘎蒿( Artemisia halodendron Caragana microphylla
白浩江 , 1 , 4 , 罗永清 , 1 , 4 , 程莉 1 , 4 , 汪正蛟一 2 , 3 , 4
1.中国科学院西北生态环境资源研究院,奈曼沙漠化研究站,甘肃 兰州 730000
2.中国科学院西北生态环境资源研究院,乌拉特荒漠草原研究站,甘肃 兰州 730000
3.中国科学院西北生态环境资源研究院,甘肃省寒区旱区逆境生理与生态重点实验室,甘肃 兰州 730000
4.中国科学院大学,北京 100049
The phenological responses of the dominant shrubs Artemisia halodendron and Caragana microphylla in the Horqin Sandy Land to climate change
Bai Haojiang , 1 , 4 , Luo Yongqing , 1 , 4 , Cheng Li 1 , 4 , Wang Zhengjiaoyi 2 , 3 , 4
1.Naiman Desertification Research Station /, Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
2.Urat Desert-Grassland Research Station /, Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
3.Gansu Province Key Laboratory of Stress Physiology and Ecology in Cold and Arid Region, Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
4.University of Chinese Academy of Sciences,Beijing 100049,China
Received: 2024-09-18
Revised: 2025-02-25
摘要
植物物候可敏感、精确地反映气候变化,反映生态系统对环境变化的响应和适应。为探究半干旱沙质草地优势灌木物候对气候变化的响应,通过定位观测得到了科尔沁沙地2种优势灌木差巴嘎蒿(Artemisia halodendron )和小叶锦鸡儿(Caragana microphylla ) 的物候时序变化,同时收集同期气候数据,采用偏最小二乘回归(PLS)方法,分析了该区域优势灌木物候对气候变化的响应。结果表明:2005—2019年,科尔沁沙地多数月份呈现出气温、降水、太阳辐射增加的趋势。随着时间的推移,2种优势灌木的展叶期显著提前(P <0.05),秋季叶变色、落叶期显著推迟(P <0.05),生长季延长(P <0.05)。温度的变化使差巴嘎蒿和小叶锦鸡儿展叶期提前,秋季叶变色、落叶期推迟,生长季延长。降水变化使其秋季落叶推迟,生长季延长。太阳辐射对优势灌木差巴嘎蒿和小叶锦鸡儿关键物候期的影响与温度和降水作用方向相反,即展叶期推迟,秋季叶变色、落叶期提前,生长季缩短。温度和降水是半干旱沙质草地优势灌木差巴嘎蒿和小叶锦鸡儿展叶期提前,秋季叶变色、落叶期推迟,生长季延长的关键气候因子。
关键词:
物候 科尔沁沙地 气候变化 偏最小二乘回归 优势灌木
Abstract
Plant phenology is one of the most sensitive and accurate indicators of climate change, and it can reflect the response and adaptation of ecosystems to environmental changes. In order to explore the meteorological driving factors of dominant shrub phenological change in semi-arid sandy grassland. We collected the phenological observation data and corresponding meteorological data from 2005 to 2019 recorded by Naiman Desertification Research Station. We used the partial least squares regression (PLS) method to analyze the effects of meteorological variables, such as temperature, precipitation and solar radiation on shrub phenology. The results showed that: temperature, precipitation, and solar radiation increased in most months in Horqin Sandy Land during 2005-2019. The leaf unfolding of two dominant shrubs was significantly advanced (P <0.05), leaf discoloration and senescence was significantly delayed (P <0.05), the length of growing season was significantly prolonged (P <0.05). Over the past 15 years, the change of temperature made Artemisia halodendron and Caragana microphylla show a trend of earlier leaf unfolding, later leaf discoloration, later senescence and longer growing season. The change of precipitation made the later senescence and longer growing season of A. halodendron and C. microphylla . With the increase of solar radiation, A. halodendron and C. microphylla showed a trend of later leaf unfolding, earlier leaf discoloration,earlier senescence and shorter growing season. In conclusion, the effects of solar radiation on A. halodendron and C. microphylla were in the opposite direction to the effects of temperature and precipitation. Temperature and precipitation were the key meteorological factor to advance the leaf unfolding, delay the leaf discoloration and senescence, as well as prolong the length of growing season of A. halodendron and C. microphylla in the semi-arid sandy grassland.
Keywords:
phenology Horqin Sandy Land climate change partial least squares regression dominant shrubs
本文引用格式
白浩江, 罗永清, 程莉, 汪正蛟一. 差巴嘎蒿( Artemisia halodendron Caragana microphylla . 中国沙漠 [J], 2025, 45(4): 305-313 doi:10.7522/j.issn.1000-694X.2025.00034
Bai Haojiang, Luo Yongqing, Cheng Li, Wang Zhengjiaoyi. The phenological responses of the dominant shrubs Artemisia halodendron and Caragana microphylla in the Horqin Sandy Land to climate change . Journal of Desert Research
0 引言
植物物候指植物受环境影响而出现的以年为周期的自然现象[1 ] 。植物物候包括休眠解除过程(如出芽)、活动过程(如展叶、开花、结果)、休眠过程(如秋季叶变色和落叶[2 ] ),是植物长期适应气候变化与环境条件而形成的生长发育节律[3 -4 ] ,关系到植物体自身的生存繁衍和植物群落的稳定[5 -7 ] 。植物物候与气候变化之间具有双向指示的特点,一方面植物物候可作为气候变化的敏感和精确指标[8 -9 ] ,另一方面气候变化直接影响植物物候。探究植物物候与气象因子的关系,有助于深入了解植物物候对气候变化的响应特征,进一步建立起更精确的草地植物物候动态模型,对监测草原的生态环境以及采取措施来应对气候变化具有重要的科学意义和实用价值。
近60年来,世界范围内的植物呈现出春季物候提前、秋季物候推迟、生长季延长的趋势[3 ] 。遥感监测和地面定点人工观测作为常用的物候观测手段[8 ] ,均监测到了这一物候变化趋势。具体而言,在基于地面观测的物候变化的信息中,通过气候变化对中国植被影响的观测证据集成分析发现,87.4%的记录表明木本植物春、夏季物候呈提前趋势;71.5%的记录表明其秋季物候呈推迟趋势;有57.7%的记录表明其生长季长度呈延长趋势[10 ] 。对中国温带植被1982—1999年物候变化的遥感数据分析显示,该区域植被春季物候提前,秋季物候推迟,生长季长度延长[11 ] 。植物物候变化受气候(温度、降水、太阳辐射等)、植物生活史、植物的适应与进化策略等因素的影响[12 ] 。其中,温度、降水和光照等气候因子是植物物候变化的主要驱动力[13 ] 。不同地区植物物候变化的主要驱动因素不同[1 ] 。在热带地区,温度是控制植物物候变化的主要因素[14 ] ;在温带和寒带地区,植物物候变化依赖于温度、低温持续和光周期的综合作用[1 ] ;在草原生态系统,温度和湿度是植物物候的主要驱动因子[15 ] ;在干旱和半干旱生态系统中,水分是植物物候的关键驱动因子[16 ] 。植物物候与气候因子的关系是全球变化的一个研究热点,反映陆地生态系统对气候变化的动态响应[11 ] 。
科尔沁沙地位于中国北方农牧交错带,生态环境脆弱,属于典型的半干旱沙地生态系统,是全球变化的敏感区域[17 ] 。近年来,在全球气候变暖背景下,科尔沁沙地植被特别是优势灌木生育期也发生了显著变化。然而,已有研究多集中于单个物候初始期的变化对气候变化的响应,很少研究不同生活史植物物候序列对气候变化的响应[18 -19 ] ,生长季不同时期气象因子对优势灌木物候期的影响仍不充分。本研究的目的是分析2005—2019年科尔沁沙地沙质草地优势灌木物候的时序变化,进一步用PLS分析探讨该区域优势灌木物候与气候因子(温度、降水、太阳辐射)之间的关系,得到驱动优势灌木物候变化的关键气候因子,进一步认知气候变化对沙地植被的影响,以期为半干旱沙地植被保护和恢复提供理论依据。
1 材料和方法
1.1 研究区概况
本试验在中国科学院奈曼沙漠化研究站的沙地综合观测场(42.92°N,120.70°E)进行,观测场地地形平坦,无明显起伏。该区域属于温带大陆性季风气候区[20 ] ,年均气温5.8~6.4 ℃,最热月(7月)平均气温23.5℃,最冷月(1月)平均气温-16.8 ℃[21 ] ,≥ 10 ℃年积温3 000~4 000 ℃,无霜期151 d[22 ] ,多年平均降水量280~360 mm,年均蒸发量1 600~2 400 mm,年均日照2 951.2 h,年均风速3.5~4.5 m·s-1[23 ] 。该地区主要物种有灌木小叶锦鸡儿(Caragana microphylla )、灌木差巴嘎蒿(Artemisia halodendron )和多年生或一年生草本如扁蓿豆(Melissitus ruthenicus )、大果虫实(Corispermum macrocarpum )、狗尾草(Setaria viridis )。相对于草本植物,灌木具有更加耐旱和抗风蚀、沙埋的特点,是沙质草地植被的主要生活型[24 ] 。其中差巴嘎蒿是一种典型的沙生中灌木,花期7—8月,果期8—9月,是良好的固沙先锋植物;小叶锦鸡儿一般4月萌发,5月下旬至6月上旬开花,6月中旬至7月中旬种子成熟,9月中旬以后逐渐落叶干枯,是沙质草地较早完成生活史周期的植物种。
1.2 气象与物候数据
本文使用的气象数据来自中国科学院奈曼沙漠化研究站标准气象站观测的气象数据(http://nmd.cern.ac.cn/meta/metaData ),选取2005—2019年月平均气温、月平均最高气温、月平均最低气温、月降水量和月平均太阳辐射5个气候变量。
在中国科学院奈曼沙漠化研究站沙地综合观测场,在目标物种分布较为均匀的区域选取了30株差巴嘎蒿和30株小叶锦鸡儿。对每一株选定个体进行编号,并记录其具体位置,采用定株观测的方式。在植物生长相对平稳、物候变化较缓慢时期,每10 d观测1次;而在临近或正处于出芽、展叶、开花、落叶等关键物候事件发生时期,则将观测频次提高至每3~5 d 1次。
灌木物候期观测以中国气象局农业气象观测规范为依据,当一半以上的所选植株达到某一发育期时,就记录该物种到了这一物候期。当明显看到植株长出绿色叶芽就是出芽期;当所观测植株上的芽从芽苞中发出卷曲着的或按叶脉褶叠着的小叶,出现第一批有一二片的叶片平展时就是展叶期;当观测到一半以上的植株有一朵或同时有几朵花的花瓣开始完全开放即为始花期;在观测的植株上有一半以上的花蕾都展开花瓣,或一半以上的花序散出花粉为盛花期;当观测的植株上有一半的果实或种子变为成熟时的颜色为结果期;所观测植株的叶子在秋天开始变色为秋季叶变色;当所观测植株秋季开始落叶记录为落叶期[25 ] 。
1.3 数据分析
本研究采用一元线性方程定量描述气象和物候要素的多年变化特征,其斜率可以反映各要素的变化速率。采用偏最小二乘回归(PLS)分析最高气温、最低气温、平均气温、降水量和太阳辐射等气候要素对植物物候的潜在影响[8 ,26 -29 ] 。由于研究区灌木的生长季末期平均出现在10月,因此选取2种优势灌木上年生长季结束至次年物候期发生前的温度、降水量和太阳辐射,应用PLS分析其与物候参数的关系。研究中气象要素和物候要素的一元线性回归在Origin 2024中完成,PLS分析在R-4.0.3中进行,插图用Origin 2024绘制。
2 结果与分析
2.1 气候因子变化特征
2005—2019年研究区平均气温、最高气温、最低气温、降水量和太阳辐射均呈现出一定的年际波动。总体上,科尔沁沙地气候呈现出暖湿变化特征。15年来,整个冬(12月至次年2月)、春(3—5月)季平均气温、最高气温、最低气温均呈现出升高趋势(2月除外),其中平均气温和最高气温变化最大是4月,其年增加速率分别为0.193、0.320 ℃;除此之外,7月平均气温、最高气温显著升高。就降水量情况而言,该地区不同月份降水量普遍呈增加趋势,降水量显著增加主要在8月,而4、7月降水量分别呈现出1.107、0.512 mm的年减少趋势。就太阳辐射而言,不同月份太阳辐射均呈增加趋势(表1 )。
2.2 灌木物候年际变化
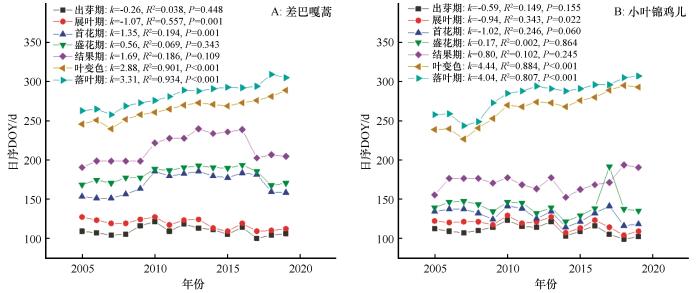
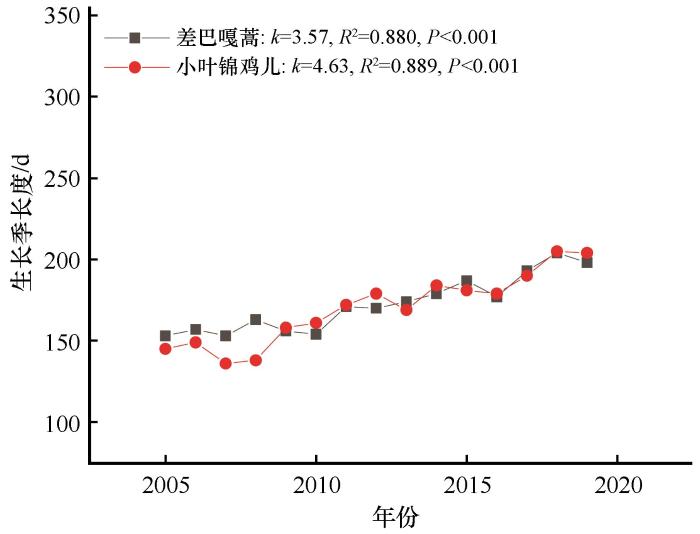
科尔沁沙地优势灌木差巴嘎蒿和小叶锦鸡儿2005—2019年物候变化如图1 所示。回归分析表明,15年间2种优势灌木秋季叶变色、落叶期显著推迟(P <0.05);生长季显著延长(P <0.05);出芽期、首花期、盛花期、结果期变化趋势不明显(P >0.05),但存在一定的年际波动。展叶期均表现为-1 d·a-1 的变化速率,并且差巴嘎蒿展叶期显著提前(P <0.01);对于秋季叶变色和落叶期而言,差巴嘎蒿的年变化率约为3 d,小叶锦鸡儿的年变化率为4 d(图1 )。受出芽期和落叶期变化趋势影响,15年来半干旱沙质草地优势灌木差巴嘎蒿、小叶锦鸡儿生长季分别延长54 d和69 d(图2 )。
图1
图1
科尔沁沙地半干旱沙质草地优势灌木不同物候期年际变化
Fig.1
Inter-annual changes of phenology of dominant shrub in the semi-arid sandy grassland of Horqin Sandy Land
图2
图2
科尔沁沙地半干旱沙质草地优势灌木生长季长度年际变化
Fig. 2
Inter-annual changes of the length of growing season of dominant shrub in the semi-arid sandy grassland of Horqin Sandy
2.3 灌木物候与气候因子关系
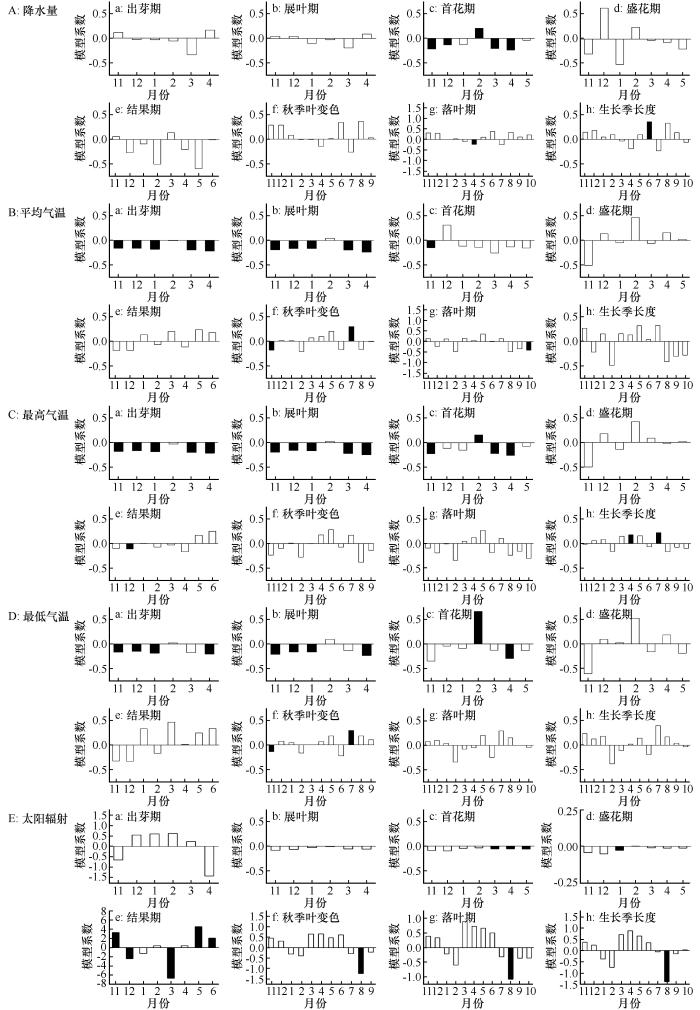
结合差巴嘎蒿和小叶锦鸡儿物候要素的年际变化特征和物候期与气象因子的PLS分析可知,气象因子的变化对差巴嘎蒿和小叶锦鸡儿的出芽期、首花期、盛花期、结果期均有影响,会使其产生一定的年际波动,但变化趋势并不显著(P >0.05),而对展叶期、秋季叶变色、落叶期和生长季长度存在显著影响(P <0.05,图1 、3 ~4 )。
图3
图3
科尔沁沙地差巴嘎蒿物候期与气象因子PLS回归分析(VIP值大于0.8用黑色填充)
Fig.3
Results of PLS regression correlating phenology of Artemisia halodendron . Coefficients filled by black indicate VIP>0.8
图4
图4
科尔沁沙地小叶锦鸡儿物候期与气象因子PLS回归分析(VIP值大于0.8用黑色填充)
Fig.4
Results of PLS regression correlating phenology of Caragana microphylla . Coefficients filled by orange indicate VIP>0.8
差巴嘎蒿的展叶期和出芽期与降水量不存在显著的相关性;秋季叶变色、生长季长度与8月降雨显著正相关,平均降水量每增加1 mm,差巴嘎蒿秋季叶变色推迟0.429 d,生长季长度增加0.528 d;落叶期与6月降雨显著正相关,说明6月降雨增加对落叶期有推迟作用。差巴嘎蒿展叶期与12、1、3、4月平均气温、最高气温以及最低气温负相关,生长季长度与3、4、7月平均气温、最高气温呈正相关关系,其中,3月平均气温升高1 ℃,差巴嘎蒿展叶期提前0.208 d,生长季长度增加0.347 d;秋季叶变色与7月平均气温、最高气温、最低气温显著正相关;落叶期与2、10月平均气温显著负相关,与4、7月最高气温显著正相关。这说明气温升高使差巴嘎蒿展叶期提前,秋季叶变色、落叶期推迟,生长季长度延长。就太阳辐射而言,差巴嘎蒿秋季叶变色、落叶期、生长季长度主要受8月份太阳辐射的影响,8月太阳辐射量每增加1 MJ·m-2 ,秋季叶变色提前1.300 d,落叶期提前1.240 d,生长季长度缩短1.626 d(图3 )。
图4 表明,降水量与小叶锦鸡儿的展叶期、出芽期不存在显著的相关性,说明降水量不是影响小叶锦鸡儿物候始期发生显著变化的关键气象因子。小叶锦鸡儿落叶期与4月降雨显著负相关,4月降水量每增加1 mm,小叶锦鸡儿落叶期提前0.228 d;生长季长度与6月份降雨显著正相关,6月降水量每增加1 mm,生长季长度延长0.358 d。就气温而言,小叶锦鸡儿展叶期与11、12、1、3、4月平均气温、最高气温、最低气温负相关;秋季叶变色与7月平均气温、最高气温、最低气温正相关;落叶期与10月平均气温显著负相关;生长季长度仅与4、7月最高气温显著正相关,4、7月温度每升高1 ℃,小叶锦鸡儿生长季长度分别延长0.176 d与0.223 d。这说明气温升高会使小叶锦鸡儿展叶期提前,秋季叶变色、落叶期推迟,生长季延长。就太阳辐射而言,秋季叶变色、落叶期、生长季长度与8月份太阳辐射呈显著负相关,8月份太阳辐射每增加1 MJ·m-2 ,小叶锦鸡儿秋季叶变色提前1.242 d,落叶期提前1.079 d,生长季长度缩短1.384 d(图4 )。
3 讨论
温度是影响植物物候期的主导因素[30 ] ,无论是温度高低、有效积温的多少,还是低温刺激,都能显著影响植物物候的演变。早期植物物候(如出芽、展叶、开花)的提前或推迟主要受春季气温的影响;中期植物物候(如结果)主要与生长季积温相关;晚期植物物候(如秋季叶变色、落叶)主要取决于阈值温度的短期作用,一旦气温下降到某个临界值就会引发这些变化[13 ] 。已有研究表明,气温升高会导致荒漠区植被提前返青。与本研究结果相吻合的是,我们观察到差巴嘎蒿和小叶锦鸡儿在生长季初期的物候指标(展叶期)均有所提前,且这一变化与12、1、3、4月的气温升高密切相关(图3 ~4 )。究其原因,可能包括以下两个方面:一方面,温度升高会使植被发育所需的有效积温更早达到,从而促进植被返青[31 ] ;另一方面,植物的生理活动涉及一系列复杂的生化反应,温度升高能够增强酶的活性,从而加速植物的发育进程[32 ] 。而生长季末期物候指标(秋季叶变色)主要与7月气温升高有关,原因可能是温度升高会使植株光合作用增强,降低了植物体内叶绿素分解速率[33 ] 、提高了酶活性[34 ] 、并推迟了霜冻的出现[35 ] ,从而使植物秋季叶变色推后[36 ] 。此外,生长季延长主要与春夏季节气温升高有关,春季(3、4月)气温上升主要影响生长季初期的物候变化,夏季(7月)气温升高则影响生长季末期,从而共同调节生长季长度,这与以往的研究结果一致[37 -42 ] 。总体而言,温度升高会在春季解除休眠并推迟秋季休眠,从而延长植物的生活史周期[1 ] 。
水分同样会影响植物物候[1 ] ,植物从生存到光合作用以及营养的传输等过程都离不开水分[37 ] 。本研究结果显示(图3 ~4 ),两种灌木的发芽期和展叶期与4月降水量之间并无显著相关。然而,已有诸多研究指出,降水增加会导致早期植物物候提前[43 -45 ] 。这一差异或许与研究区域和植物种类的差异有关[46 ] 。在本研究中,差巴嘎蒿与小叶锦鸡儿的返青时间显著提前(图1 ),很可能是由于4月气温升高及降雪减少等因素所致,而与水分变化并无显著关联。6月的降雨增加使差巴嘎蒿秋季落叶期推迟,小叶锦鸡儿生长季变长;8月降雨增加导致差巴嘎蒿秋季叶变色推迟,并延长其生长季,这与此前的研究结果相符[43 ,47 ] ,即夏季降水增加会导致晚期植物物候推迟。这可能是因为在满足植物生长所需热量条件下,水分的增加能为植物提供更多可利用的水资源,从而使秋季叶变色和落叶期推后,生长季得以延长。
光作为植物重要的能量来源,可通过影响植物的光敏色素,进而通过辐射强度、辐射长度以及光谱成分来影响植物生活史的很多阶段[1 ,38 ] 。本研究发现8月太阳辐射增加使秋季叶变色、落叶期提前,生长季长度缩短(图3 ~4 )。推测其主要原因是因为太阳辐射升高,植被蒸腾作用增强,植物可利用水资源减少而导致[31 ] 。
综上所述,物候变化是温度、降水量、太阳辐射等因素综合作用的结果,其中温度和降水是差巴嘎蒿和小叶锦鸡儿展叶期提前,秋季叶变色、落叶期推迟,生长季长度延长的关键气象因子,太阳辐射则是相对次要的因子。
4 结论
2005—2019年,科尔沁沙地气候呈现暖湿化发展趋势,导致该地区优势灌木差巴嘎蒿和小叶锦鸡儿的物候期发生明显改变。差巴嘎蒿的展叶期提前15 d,落叶期推迟43 d,生长季延长54 d;小叶锦鸡儿的展叶期提前13 d,落叶期推迟49 d,生长季延长69 d。
气温和降水量是影响差巴嘎蒿物候期的关键因素。春季气温升高会使差巴嘎蒿出芽期和展叶期提前,而夏季降水量的增加会显著推迟其秋季叶变色与落叶期,从而延长生长季。小叶锦鸡儿主要受气温影响:春季平均温度和最高温度上升均可使其出芽期和展叶期提前,夏季最高温度升高则会显著延长其生长季;降水量也会在一定程度上延长小叶锦鸡儿生长季,但除对花期有所影响外,对其他物候期并无显著作用。
展叶期提前与落叶期推迟表明植物通过延长光合作用时间来增强有机物累积和提高资源利用效率。然而,生长季的延长会在科尔沁沙地引发更高的水分蒸散压力,特别是在干旱年份,可能对当地环境稳定性产生不利影响。
参考文献
View Option
[1]
翟佳 ,袁凤辉 ,吴家兵 植物物候变化研究进展
[J].生态学杂志 ,2015 ,34 (11 ):257 -263 .
[本文引用: 6]
[2]
杨扬 ,罗贤 ,李荣平 ,等 气象要素对植物物候影响及其驱动机制研究进展
[J].气象与环境学报 ,2016 ,32 (5 ):154 -159 .
[本文引用: 1]
[3]
王连喜 ,陈怀亮 ,李琪 ,等 植物物候与气候研究进展
[J].生态学报 ,2010 ,20 (2 ):447 -454 .
[本文引用: 2]
[4]
Gonsamo A Chen J M Price D T et al Land surface phenology from optical satellite measurement and CO2 eddy covariance technique
[J].Journal of Geophysical Research Biogeosciences ,2015 ,117 (G3 ):1 -12 .
[本文引用: 1]
[5]
张德露 ,刘芳 ,武倩 ,等 气候变化下草原植物物候响应规律研究进展
[J].内蒙古林业调查设计 ,2018 ,41 (6 ):102 -104 .
[本文引用: 1]
[6]
Fenner M The phenology of growth and reproduction in plants
[J].Perspectives in Plant Ecology,Evolution and Systematics ,1998 ,1 (1 ):78 -91 .
[7]
刘志民 ,蒋德明 ,高红瑛 ,等 植物生活史繁殖对策与干扰关系的研究
[J].应用生态学报 ,2003 ,14 (3 ):418 -422 .
[本文引用: 1]
[8]
俎佳星 ,杨健 东北地区植物物候时序变化
[J].生态学报 ,2016 ,36 (7 ):2015 -2023 .
[本文引用: 3]
[9]
Beaubien E G Freeland H J Spring phenology trends in Alberta,Canada:links to ocean temperature
[J].International Journal of Biometeorology ,2000 ,44 (2 ):53 -59 .
[本文引用: 1]
[10]
徐佳 气候变化对我国植被影响的观测证据集成分析
[D].浙江金华 :浙江师范大学 ,2019 .
[本文引用: 1]
[11]
Piao S L Fang J Y Zhou L M et al Variations in satellite-derived phenology in China's temperate vegetation
[J].Global Change Biology ,2006 ,12 (4 ):672 -685 .
[本文引用: 2]
[12]
包晓影 ,崔树娟 ,王奇 ,等 草地植物物候研究进展及其存在的问题
[J].生态学杂志 ,2017 ,36 (8 ):2321 -2326 .
[本文引用: 1]
[13]
丁抗抗 ,高庆先 ,李辑 我国植物物候变化及对气候变化的响应综述
[J].安徽农业科学 ,2010 ,38 (14 ):7414 -7417 .
[本文引用: 2]
[14]
莫非 ,赵鸿 ,王建永 ,等 全球变化下植物物候研究的关键问题
[J].生态学报 ,2011 ,31 (9 ):2593 -2601 .
[本文引用: 1]
[15]
Wolkovich E M Cook B I Davies T J Progress towards an interdisciplinary science of plant phenology:building predictions across space,time and species diversity
[J].New Phytologists ,2014 ,201 (4 ):1156 -1162 .
[本文引用: 1]
[16]
代武君 ,金慧颖 ,张玉红 ,等 植物物候学研究进展
[J].生态学报 ,2020 ,40 (19 ):6705 -6719 .
[本文引用: 1]
[17]
穆少杰 ,李建龙 ,陈奕兆 ,等 2001-2010年内蒙古植被覆盖度时空变化特征
[J].地理学报 ,2012 ,67 (9 ):1255 -1268 .
[本文引用: 1]
[18]
Wang S Wang C Duan J et al Timing and duration of phenological sequences of alpine plants along an elevation gradient on the Tibetan Plateau
[J].Agricultural and Forest Meteorology ,2014 ,189 :220 -228 .
[本文引用: 1]
[19]
Jiang L L Wang S P Meng F D et al Relatively stable response of fruiting stage to warming and cooling relative to other phenological events
[J].Ecology ,2016 ,97 (8 ):1961 .
[本文引用: 1]
[20]
李玉霖 ,崔建垣 ,苏永中 不同沙丘生境主要植物比叶面积和叶干物质含量的比较
[J].生态学报 ,2005 ,25 (2 ):304 -311 .
[本文引用: 1]
[21]
Zuo X Yue X Lv P et al Contrasting effects of plant inter‐ and intraspecific variation on community trait responses to restoration of a sandy grassland ecosystem
[J].Ecology & Evolution ,2017 ,7 (4 ):1125 -1134 .
[本文引用: 1]
[22]
宁志英 ,李玉霖 ,杨红玲 ,等 科尔沁沙地主要植物细根和叶片碳,氮,磷化学计量特征
[J].植物生态学报 ,2017 ,41 (10 ):1069 -1081 .
[本文引用: 1]
[23]
马赟花 ,张铜会 ,刘新平 半干旱区沙地芦苇对浅水位变化的生理生态响应
[J].生态学报 ,2013 ,33 (21 ):6984 -6991 .
[本文引用: 1]
[24]
王青宁 ,衣学慧 ,王晗生 ,等 水分胁迫对几种固沙灌木幼苗生物量的影响
[J].甘肃农业大学学报 ,2014 ,49 (3 ):93 -100 .
[本文引用: 1]
[25]
宛敏渭 ,刘秀珍 中国物候观测方法 [M].北京 :科学出版社 ,1979 .
[本文引用: 1]
[26]
Luedeling E Gassner A Partial least squares regression for analyzing walnut phenology in California
[J].Agricultural & Forest Meteorology ,2012 ,158 :43 -52 .
[本文引用: 1]
[27]
Guo L Dai J Ranjitkar S et al Response of chestnut phenology in China to climate variation and change
[J].Agricultural & Forest Meteorology ,2013 ,180 :164 -172 .
[28]
张文奇 ,李丹 ,李浩 ,等 克里雅河流域植被物候时空变化及影响因素
[J].水土保持通报 ,2020 ,40 (5 ):291 -298 .
[29]
孙丕苓 ,许月卿 ,王数 环京津贫困带土地利用变化的地形梯度效应分析
[J].农业工程学报 ,2014 ,30 (14 ):277 -288 .
[本文引用: 1]
[30]
程建刚 ,解明恩 近50年云南区域气候变化特征分析
[J].地理科学进展 ,2008 ,27 (5 ):19 -26 .
[本文引用: 1]
[31]
Zhang Q Kong D Shi P et al Vegetation phenology on the Qinghai-Tibetan Plateau and its response to climate change (1982-2013)
[J].Agricultural And Forest Meteorology ,2018 ,248 :408 -417 .
[本文引用: 2]
[32]
李荣平 ,周广胜 1980-2005年中国东北木本植物物候特征及其对气温的响应
[J].生态学杂志 ,2010 ,29 (12 ):2317 -2326 .
[本文引用: 1]
[33]
Fracheboud Y Björkén L Sjödin A et al The control of autumn senescence in European aspens
[J].Plant Physiology ,2009 ,149 (4 ):1982 -1991 .
[本文引用: 1]
[34]
Shi C Sun G Zhang H et al Effects of warming on chlorophyll degradation and carbohydrate accumulation of alpine herbaceous species during plant senescence on the Tibetan Plateau
[J].Plos One ,2014 ,9 (9 ):e107874 .
[本文引用: 1]
[35]
Schwartz M Phenology:An Integrative Environmental Science [M].Netherlands :Springer ,2013 .
[本文引用: 1]
[36]
Liu Q Fu Y H Zeng Z et al Temperature,precipitation,and insolation effects on autumn vegetation phenology in temperate China
[J].Global Change Biology ,2016 ,22 (2 ):644 -655 .
[本文引用: 1]
[37]
符瑜 ,潘学标 草本植物物候及其物候模拟模型的研究进展
[J].中国农业气象 ,2011 ,32 (3 ):319 -325 .
[本文引用: 2]
[38]
李荣平 ,周广胜 ,张慧玲 植物物候研究进展
[J].应用生态学报 ,2006 ,17 (3 ):541 -544 .
[本文引用: 1]
[39]
Menzel A Sparks T H Estrella N et al European phenological response to climate change matches the warming pattern
[J].Global Change Biology ,2006 ,12 (10 ):1969 -1976 .
[40]
Schwartz M D Ahas R Aasa A Onset of spring starting earlier across the Northern Hemisphere
[J].Global Change Biology ,2006 ,12 (2 ):343 -351 .
[41]
Fu Y H Piao S Opdebeeck M et al Recent spring phenology shifts in western Central Europe based on multiscale observations
[J].Global Ecology and Biogeography ,2014 ,23 (11 ):1255 -1263 .
[42]
刘秀梅 ,郑红玉 ,李小锋 天山北坡荒漠草原主要植物物候特征对气候变化的响应
[J].草学 ,2022 ,39 (3 ):51 -57 .
[本文引用: 1]
[43]
张仁平 ,郭靖 ,冯琦胜 ,等 新疆地区草地植被物候时空变化
[J].草业学报 ,2018 ,27 (10 ):66 -75 .
[本文引用: 2]
[44]
范德芹 ,赵学胜 ,郑周涛 内蒙古羊草草原物候及其对气候变化的响应
[J].地理与地理信息科学 ,2016 ,32 (6 ):81 -86 .
[45]
张玉静 ,杨秀春 ,郭剑 ,等 呼伦贝尔草原物候变化及其与气象因子的关系
[J].干旱区地理 ,2019 ,42 (1 ):144 -153 .
[本文引用: 1]
[46]
秦格霞 ,吴静 ,李纯斌 ,等 中国北方草地植被物候变化及其对气候变化的响应
[J].应用生态学报 ,2019 ,30 (12 ):4099 -4107 .
[本文引用: 1]
[47]
李杭燕 ,颉耀文 ,马明国 时序NDVI数据集重建方法评价与实例研究
[J].遥感技术与应用 ,2009 ,24 (5 ):596 -602 .
[本文引用: 1]
植物物候变化研究进展
6
2015
... 植物物候指植物受环境影响而出现的以年为周期的自然现象[1 ] .植物物候包括休眠解除过程(如出芽)、活动过程(如展叶、开花、结果)、休眠过程(如秋季叶变色和落叶[2 ] ),是植物长期适应气候变化与环境条件而形成的生长发育节律[3 -4 ] ,关系到植物体自身的生存繁衍和植物群落的稳定[5 -7 ] .植物物候与气候变化之间具有双向指示的特点,一方面植物物候可作为气候变化的敏感和精确指标[8 -9 ] ,另一方面气候变化直接影响植物物候.探究植物物候与气象因子的关系,有助于深入了解植物物候对气候变化的响应特征,进一步建立起更精确的草地植物物候动态模型,对监测草原的生态环境以及采取措施来应对气候变化具有重要的科学意义和实用价值. ...
... 近60年来,世界范围内的植物呈现出春季物候提前、秋季物候推迟、生长季延长的趋势[3 ] .遥感监测和地面定点人工观测作为常用的物候观测手段[8 ] ,均监测到了这一物候变化趋势.具体而言,在基于地面观测的物候变化的信息中,通过气候变化对中国植被影响的观测证据集成分析发现,87.4%的记录表明木本植物春、夏季物候呈提前趋势;71.5%的记录表明其秋季物候呈推迟趋势;有57.7%的记录表明其生长季长度呈延长趋势[10 ] .对中国温带植被1982—1999年物候变化的遥感数据分析显示,该区域植被春季物候提前,秋季物候推迟,生长季长度延长[11 ] .植物物候变化受气候(温度、降水、太阳辐射等)、植物生活史、植物的适应与进化策略等因素的影响[12 ] .其中,温度、降水和光照等气候因子是植物物候变化的主要驱动力[13 ] .不同地区植物物候变化的主要驱动因素不同[1 ] .在热带地区,温度是控制植物物候变化的主要因素[14 ] ;在温带和寒带地区,植物物候变化依赖于温度、低温持续和光周期的综合作用[1 ] ;在草原生态系统,温度和湿度是植物物候的主要驱动因子[15 ] ;在干旱和半干旱生态系统中,水分是植物物候的关键驱动因子[16 ] .植物物候与气候因子的关系是全球变化的一个研究热点,反映陆地生态系统对气候变化的动态响应[11 ] . ...
... [1 ];在草原生态系统,温度和湿度是植物物候的主要驱动因子[15 ] ;在干旱和半干旱生态系统中,水分是植物物候的关键驱动因子[16 ] .植物物候与气候因子的关系是全球变化的一个研究热点,反映陆地生态系统对气候变化的动态响应[11 ] . ...
... 温度是影响植物物候期的主导因素[30 ] ,无论是温度高低、有效积温的多少,还是低温刺激,都能显著影响植物物候的演变.早期植物物候(如出芽、展叶、开花)的提前或推迟主要受春季气温的影响;中期植物物候(如结果)主要与生长季积温相关;晚期植物物候(如秋季叶变色、落叶)主要取决于阈值温度的短期作用,一旦气温下降到某个临界值就会引发这些变化[13 ] .已有研究表明,气温升高会导致荒漠区植被提前返青.与本研究结果相吻合的是,我们观察到差巴嘎蒿和小叶锦鸡儿在生长季初期的物候指标(展叶期)均有所提前,且这一变化与12、1、3、4月的气温升高密切相关(图3 ~4 ).究其原因,可能包括以下两个方面:一方面,温度升高会使植被发育所需的有效积温更早达到,从而促进植被返青[31 ] ;另一方面,植物的生理活动涉及一系列复杂的生化反应,温度升高能够增强酶的活性,从而加速植物的发育进程[32 ] .而生长季末期物候指标(秋季叶变色)主要与7月气温升高有关,原因可能是温度升高会使植株光合作用增强,降低了植物体内叶绿素分解速率[33 ] 、提高了酶活性[34 ] 、并推迟了霜冻的出现[35 ] ,从而使植物秋季叶变色推后[36 ] .此外,生长季延长主要与春夏季节气温升高有关,春季(3、4月)气温上升主要影响生长季初期的物候变化,夏季(7月)气温升高则影响生长季末期,从而共同调节生长季长度,这与以往的研究结果一致[37 -42 ] .总体而言,温度升高会在春季解除休眠并推迟秋季休眠,从而延长植物的生活史周期[1 ] . ...
... 水分同样会影响植物物候[1 ] ,植物从生存到光合作用以及营养的传输等过程都离不开水分[37 ] .本研究结果显示(图3 ~4 ),两种灌木的发芽期和展叶期与4月降水量之间并无显著相关.然而,已有诸多研究指出,降水增加会导致早期植物物候提前[43 -45 ] .这一差异或许与研究区域和植物种类的差异有关[46 ] .在本研究中,差巴嘎蒿与小叶锦鸡儿的返青时间显著提前(图1 ),很可能是由于4月气温升高及降雪减少等因素所致,而与水分变化并无显著关联.6月的降雨增加使差巴嘎蒿秋季落叶期推迟,小叶锦鸡儿生长季变长;8月降雨增加导致差巴嘎蒿秋季叶变色推迟,并延长其生长季,这与此前的研究结果相符[43 ,47 ] ,即夏季降水增加会导致晚期植物物候推迟.这可能是因为在满足植物生长所需热量条件下,水分的增加能为植物提供更多可利用的水资源,从而使秋季叶变色和落叶期推后,生长季得以延长. ...
... 光作为植物重要的能量来源,可通过影响植物的光敏色素,进而通过辐射强度、辐射长度以及光谱成分来影响植物生活史的很多阶段[1 ,38 ] .本研究发现8月太阳辐射增加使秋季叶变色、落叶期提前,生长季长度缩短(图3 ~4 ).推测其主要原因是因为太阳辐射升高,植被蒸腾作用增强,植物可利用水资源减少而导致[31 ] . ...
气象要素对植物物候影响及其驱动机制研究进展
1
2016
... 植物物候指植物受环境影响而出现的以年为周期的自然现象[1 ] .植物物候包括休眠解除过程(如出芽)、活动过程(如展叶、开花、结果)、休眠过程(如秋季叶变色和落叶[2 ] ),是植物长期适应气候变化与环境条件而形成的生长发育节律[3 -4 ] ,关系到植物体自身的生存繁衍和植物群落的稳定[5 -7 ] .植物物候与气候变化之间具有双向指示的特点,一方面植物物候可作为气候变化的敏感和精确指标[8 -9 ] ,另一方面气候变化直接影响植物物候.探究植物物候与气象因子的关系,有助于深入了解植物物候对气候变化的响应特征,进一步建立起更精确的草地植物物候动态模型,对监测草原的生态环境以及采取措施来应对气候变化具有重要的科学意义和实用价值. ...
植物物候与气候研究进展
2
2010
... 植物物候指植物受环境影响而出现的以年为周期的自然现象[1 ] .植物物候包括休眠解除过程(如出芽)、活动过程(如展叶、开花、结果)、休眠过程(如秋季叶变色和落叶[2 ] ),是植物长期适应气候变化与环境条件而形成的生长发育节律[3 -4 ] ,关系到植物体自身的生存繁衍和植物群落的稳定[5 -7 ] .植物物候与气候变化之间具有双向指示的特点,一方面植物物候可作为气候变化的敏感和精确指标[8 -9 ] ,另一方面气候变化直接影响植物物候.探究植物物候与气象因子的关系,有助于深入了解植物物候对气候变化的响应特征,进一步建立起更精确的草地植物物候动态模型,对监测草原的生态环境以及采取措施来应对气候变化具有重要的科学意义和实用价值. ...
... 近60年来,世界范围内的植物呈现出春季物候提前、秋季物候推迟、生长季延长的趋势[3 ] .遥感监测和地面定点人工观测作为常用的物候观测手段[8 ] ,均监测到了这一物候变化趋势.具体而言,在基于地面观测的物候变化的信息中,通过气候变化对中国植被影响的观测证据集成分析发现,87.4%的记录表明木本植物春、夏季物候呈提前趋势;71.5%的记录表明其秋季物候呈推迟趋势;有57.7%的记录表明其生长季长度呈延长趋势[10 ] .对中国温带植被1982—1999年物候变化的遥感数据分析显示,该区域植被春季物候提前,秋季物候推迟,生长季长度延长[11 ] .植物物候变化受气候(温度、降水、太阳辐射等)、植物生活史、植物的适应与进化策略等因素的影响[12 ] .其中,温度、降水和光照等气候因子是植物物候变化的主要驱动力[13 ] .不同地区植物物候变化的主要驱动因素不同[1 ] .在热带地区,温度是控制植物物候变化的主要因素[14 ] ;在温带和寒带地区,植物物候变化依赖于温度、低温持续和光周期的综合作用[1 ] ;在草原生态系统,温度和湿度是植物物候的主要驱动因子[15 ] ;在干旱和半干旱生态系统中,水分是植物物候的关键驱动因子[16 ] .植物物候与气候因子的关系是全球变化的一个研究热点,反映陆地生态系统对气候变化的动态响应[11 ] . ...
Land surface phenology from optical satellite measurement and CO2 eddy covariance technique
1
2015
... 植物物候指植物受环境影响而出现的以年为周期的自然现象[1 ] .植物物候包括休眠解除过程(如出芽)、活动过程(如展叶、开花、结果)、休眠过程(如秋季叶变色和落叶[2 ] ),是植物长期适应气候变化与环境条件而形成的生长发育节律[3 -4 ] ,关系到植物体自身的生存繁衍和植物群落的稳定[5 -7 ] .植物物候与气候变化之间具有双向指示的特点,一方面植物物候可作为气候变化的敏感和精确指标[8 -9 ] ,另一方面气候变化直接影响植物物候.探究植物物候与气象因子的关系,有助于深入了解植物物候对气候变化的响应特征,进一步建立起更精确的草地植物物候动态模型,对监测草原的生态环境以及采取措施来应对气候变化具有重要的科学意义和实用价值. ...
气候变化下草原植物物候响应规律研究进展
1
2018
... 植物物候指植物受环境影响而出现的以年为周期的自然现象[1 ] .植物物候包括休眠解除过程(如出芽)、活动过程(如展叶、开花、结果)、休眠过程(如秋季叶变色和落叶[2 ] ),是植物长期适应气候变化与环境条件而形成的生长发育节律[3 -4 ] ,关系到植物体自身的生存繁衍和植物群落的稳定[5 -7 ] .植物物候与气候变化之间具有双向指示的特点,一方面植物物候可作为气候变化的敏感和精确指标[8 -9 ] ,另一方面气候变化直接影响植物物候.探究植物物候与气象因子的关系,有助于深入了解植物物候对气候变化的响应特征,进一步建立起更精确的草地植物物候动态模型,对监测草原的生态环境以及采取措施来应对气候变化具有重要的科学意义和实用价值. ...
The phenology of growth and reproduction in plants
0
1998
植物生活史繁殖对策与干扰关系的研究
1
2003
... 植物物候指植物受环境影响而出现的以年为周期的自然现象[1 ] .植物物候包括休眠解除过程(如出芽)、活动过程(如展叶、开花、结果)、休眠过程(如秋季叶变色和落叶[2 ] ),是植物长期适应气候变化与环境条件而形成的生长发育节律[3 -4 ] ,关系到植物体自身的生存繁衍和植物群落的稳定[5 -7 ] .植物物候与气候变化之间具有双向指示的特点,一方面植物物候可作为气候变化的敏感和精确指标[8 -9 ] ,另一方面气候变化直接影响植物物候.探究植物物候与气象因子的关系,有助于深入了解植物物候对气候变化的响应特征,进一步建立起更精确的草地植物物候动态模型,对监测草原的生态环境以及采取措施来应对气候变化具有重要的科学意义和实用价值. ...
东北地区植物物候时序变化
3
2016
... 植物物候指植物受环境影响而出现的以年为周期的自然现象[1 ] .植物物候包括休眠解除过程(如出芽)、活动过程(如展叶、开花、结果)、休眠过程(如秋季叶变色和落叶[2 ] ),是植物长期适应气候变化与环境条件而形成的生长发育节律[3 -4 ] ,关系到植物体自身的生存繁衍和植物群落的稳定[5 -7 ] .植物物候与气候变化之间具有双向指示的特点,一方面植物物候可作为气候变化的敏感和精确指标[8 -9 ] ,另一方面气候变化直接影响植物物候.探究植物物候与气象因子的关系,有助于深入了解植物物候对气候变化的响应特征,进一步建立起更精确的草地植物物候动态模型,对监测草原的生态环境以及采取措施来应对气候变化具有重要的科学意义和实用价值. ...
... 近60年来,世界范围内的植物呈现出春季物候提前、秋季物候推迟、生长季延长的趋势[3 ] .遥感监测和地面定点人工观测作为常用的物候观测手段[8 ] ,均监测到了这一物候变化趋势.具体而言,在基于地面观测的物候变化的信息中,通过气候变化对中国植被影响的观测证据集成分析发现,87.4%的记录表明木本植物春、夏季物候呈提前趋势;71.5%的记录表明其秋季物候呈推迟趋势;有57.7%的记录表明其生长季长度呈延长趋势[10 ] .对中国温带植被1982—1999年物候变化的遥感数据分析显示,该区域植被春季物候提前,秋季物候推迟,生长季长度延长[11 ] .植物物候变化受气候(温度、降水、太阳辐射等)、植物生活史、植物的适应与进化策略等因素的影响[12 ] .其中,温度、降水和光照等气候因子是植物物候变化的主要驱动力[13 ] .不同地区植物物候变化的主要驱动因素不同[1 ] .在热带地区,温度是控制植物物候变化的主要因素[14 ] ;在温带和寒带地区,植物物候变化依赖于温度、低温持续和光周期的综合作用[1 ] ;在草原生态系统,温度和湿度是植物物候的主要驱动因子[15 ] ;在干旱和半干旱生态系统中,水分是植物物候的关键驱动因子[16 ] .植物物候与气候因子的关系是全球变化的一个研究热点,反映陆地生态系统对气候变化的动态响应[11 ] . ...
... 本研究采用一元线性方程定量描述气象和物候要素的多年变化特征,其斜率可以反映各要素的变化速率.采用偏最小二乘回归(PLS)分析最高气温、最低气温、平均气温、降水量和太阳辐射等气候要素对植物物候的潜在影响[8 ,26 -29 ] .由于研究区灌木的生长季末期平均出现在10月,因此选取2种优势灌木上年生长季结束至次年物候期发生前的温度、降水量和太阳辐射,应用PLS分析其与物候参数的关系.研究中气象要素和物候要素的一元线性回归在Origin 2024中完成,PLS分析在R-4.0.3中进行,插图用Origin 2024绘制. ...
Spring phenology trends in Alberta,Canada:links to ocean temperature
1
2000
... 植物物候指植物受环境影响而出现的以年为周期的自然现象[1 ] .植物物候包括休眠解除过程(如出芽)、活动过程(如展叶、开花、结果)、休眠过程(如秋季叶变色和落叶[2 ] ),是植物长期适应气候变化与环境条件而形成的生长发育节律[3 -4 ] ,关系到植物体自身的生存繁衍和植物群落的稳定[5 -7 ] .植物物候与气候变化之间具有双向指示的特点,一方面植物物候可作为气候变化的敏感和精确指标[8 -9 ] ,另一方面气候变化直接影响植物物候.探究植物物候与气象因子的关系,有助于深入了解植物物候对气候变化的响应特征,进一步建立起更精确的草地植物物候动态模型,对监测草原的生态环境以及采取措施来应对气候变化具有重要的科学意义和实用价值. ...
气候变化对我国植被影响的观测证据集成分析
1
2019
... 近60年来,世界范围内的植物呈现出春季物候提前、秋季物候推迟、生长季延长的趋势[3 ] .遥感监测和地面定点人工观测作为常用的物候观测手段[8 ] ,均监测到了这一物候变化趋势.具体而言,在基于地面观测的物候变化的信息中,通过气候变化对中国植被影响的观测证据集成分析发现,87.4%的记录表明木本植物春、夏季物候呈提前趋势;71.5%的记录表明其秋季物候呈推迟趋势;有57.7%的记录表明其生长季长度呈延长趋势[10 ] .对中国温带植被1982—1999年物候变化的遥感数据分析显示,该区域植被春季物候提前,秋季物候推迟,生长季长度延长[11 ] .植物物候变化受气候(温度、降水、太阳辐射等)、植物生活史、植物的适应与进化策略等因素的影响[12 ] .其中,温度、降水和光照等气候因子是植物物候变化的主要驱动力[13 ] .不同地区植物物候变化的主要驱动因素不同[1 ] .在热带地区,温度是控制植物物候变化的主要因素[14 ] ;在温带和寒带地区,植物物候变化依赖于温度、低温持续和光周期的综合作用[1 ] ;在草原生态系统,温度和湿度是植物物候的主要驱动因子[15 ] ;在干旱和半干旱生态系统中,水分是植物物候的关键驱动因子[16 ] .植物物候与气候因子的关系是全球变化的一个研究热点,反映陆地生态系统对气候变化的动态响应[11 ] . ...
Variations in satellite-derived phenology in China's temperate vegetation
2
2006
... 近60年来,世界范围内的植物呈现出春季物候提前、秋季物候推迟、生长季延长的趋势[3 ] .遥感监测和地面定点人工观测作为常用的物候观测手段[8 ] ,均监测到了这一物候变化趋势.具体而言,在基于地面观测的物候变化的信息中,通过气候变化对中国植被影响的观测证据集成分析发现,87.4%的记录表明木本植物春、夏季物候呈提前趋势;71.5%的记录表明其秋季物候呈推迟趋势;有57.7%的记录表明其生长季长度呈延长趋势[10 ] .对中国温带植被1982—1999年物候变化的遥感数据分析显示,该区域植被春季物候提前,秋季物候推迟,生长季长度延长[11 ] .植物物候变化受气候(温度、降水、太阳辐射等)、植物生活史、植物的适应与进化策略等因素的影响[12 ] .其中,温度、降水和光照等气候因子是植物物候变化的主要驱动力[13 ] .不同地区植物物候变化的主要驱动因素不同[1 ] .在热带地区,温度是控制植物物候变化的主要因素[14 ] ;在温带和寒带地区,植物物候变化依赖于温度、低温持续和光周期的综合作用[1 ] ;在草原生态系统,温度和湿度是植物物候的主要驱动因子[15 ] ;在干旱和半干旱生态系统中,水分是植物物候的关键驱动因子[16 ] .植物物候与气候因子的关系是全球变化的一个研究热点,反映陆地生态系统对气候变化的动态响应[11 ] . ...
... [11 ]. ...
草地植物物候研究进展及其存在的问题
1
2017
... 近60年来,世界范围内的植物呈现出春季物候提前、秋季物候推迟、生长季延长的趋势[3 ] .遥感监测和地面定点人工观测作为常用的物候观测手段[8 ] ,均监测到了这一物候变化趋势.具体而言,在基于地面观测的物候变化的信息中,通过气候变化对中国植被影响的观测证据集成分析发现,87.4%的记录表明木本植物春、夏季物候呈提前趋势;71.5%的记录表明其秋季物候呈推迟趋势;有57.7%的记录表明其生长季长度呈延长趋势[10 ] .对中国温带植被1982—1999年物候变化的遥感数据分析显示,该区域植被春季物候提前,秋季物候推迟,生长季长度延长[11 ] .植物物候变化受气候(温度、降水、太阳辐射等)、植物生活史、植物的适应与进化策略等因素的影响[12 ] .其中,温度、降水和光照等气候因子是植物物候变化的主要驱动力[13 ] .不同地区植物物候变化的主要驱动因素不同[1 ] .在热带地区,温度是控制植物物候变化的主要因素[14 ] ;在温带和寒带地区,植物物候变化依赖于温度、低温持续和光周期的综合作用[1 ] ;在草原生态系统,温度和湿度是植物物候的主要驱动因子[15 ] ;在干旱和半干旱生态系统中,水分是植物物候的关键驱动因子[16 ] .植物物候与气候因子的关系是全球变化的一个研究热点,反映陆地生态系统对气候变化的动态响应[11 ] . ...
我国植物物候变化及对气候变化的响应综述
2
2010
... 近60年来,世界范围内的植物呈现出春季物候提前、秋季物候推迟、生长季延长的趋势[3 ] .遥感监测和地面定点人工观测作为常用的物候观测手段[8 ] ,均监测到了这一物候变化趋势.具体而言,在基于地面观测的物候变化的信息中,通过气候变化对中国植被影响的观测证据集成分析发现,87.4%的记录表明木本植物春、夏季物候呈提前趋势;71.5%的记录表明其秋季物候呈推迟趋势;有57.7%的记录表明其生长季长度呈延长趋势[10 ] .对中国温带植被1982—1999年物候变化的遥感数据分析显示,该区域植被春季物候提前,秋季物候推迟,生长季长度延长[11 ] .植物物候变化受气候(温度、降水、太阳辐射等)、植物生活史、植物的适应与进化策略等因素的影响[12 ] .其中,温度、降水和光照等气候因子是植物物候变化的主要驱动力[13 ] .不同地区植物物候变化的主要驱动因素不同[1 ] .在热带地区,温度是控制植物物候变化的主要因素[14 ] ;在温带和寒带地区,植物物候变化依赖于温度、低温持续和光周期的综合作用[1 ] ;在草原生态系统,温度和湿度是植物物候的主要驱动因子[15 ] ;在干旱和半干旱生态系统中,水分是植物物候的关键驱动因子[16 ] .植物物候与气候因子的关系是全球变化的一个研究热点,反映陆地生态系统对气候变化的动态响应[11 ] . ...
... 温度是影响植物物候期的主导因素[30 ] ,无论是温度高低、有效积温的多少,还是低温刺激,都能显著影响植物物候的演变.早期植物物候(如出芽、展叶、开花)的提前或推迟主要受春季气温的影响;中期植物物候(如结果)主要与生长季积温相关;晚期植物物候(如秋季叶变色、落叶)主要取决于阈值温度的短期作用,一旦气温下降到某个临界值就会引发这些变化[13 ] .已有研究表明,气温升高会导致荒漠区植被提前返青.与本研究结果相吻合的是,我们观察到差巴嘎蒿和小叶锦鸡儿在生长季初期的物候指标(展叶期)均有所提前,且这一变化与12、1、3、4月的气温升高密切相关(图3 ~4 ).究其原因,可能包括以下两个方面:一方面,温度升高会使植被发育所需的有效积温更早达到,从而促进植被返青[31 ] ;另一方面,植物的生理活动涉及一系列复杂的生化反应,温度升高能够增强酶的活性,从而加速植物的发育进程[32 ] .而生长季末期物候指标(秋季叶变色)主要与7月气温升高有关,原因可能是温度升高会使植株光合作用增强,降低了植物体内叶绿素分解速率[33 ] 、提高了酶活性[34 ] 、并推迟了霜冻的出现[35 ] ,从而使植物秋季叶变色推后[36 ] .此外,生长季延长主要与春夏季节气温升高有关,春季(3、4月)气温上升主要影响生长季初期的物候变化,夏季(7月)气温升高则影响生长季末期,从而共同调节生长季长度,这与以往的研究结果一致[37 -42 ] .总体而言,温度升高会在春季解除休眠并推迟秋季休眠,从而延长植物的生活史周期[1 ] . ...
全球变化下植物物候研究的关键问题
1
2011
... 近60年来,世界范围内的植物呈现出春季物候提前、秋季物候推迟、生长季延长的趋势[3 ] .遥感监测和地面定点人工观测作为常用的物候观测手段[8 ] ,均监测到了这一物候变化趋势.具体而言,在基于地面观测的物候变化的信息中,通过气候变化对中国植被影响的观测证据集成分析发现,87.4%的记录表明木本植物春、夏季物候呈提前趋势;71.5%的记录表明其秋季物候呈推迟趋势;有57.7%的记录表明其生长季长度呈延长趋势[10 ] .对中国温带植被1982—1999年物候变化的遥感数据分析显示,该区域植被春季物候提前,秋季物候推迟,生长季长度延长[11 ] .植物物候变化受气候(温度、降水、太阳辐射等)、植物生活史、植物的适应与进化策略等因素的影响[12 ] .其中,温度、降水和光照等气候因子是植物物候变化的主要驱动力[13 ] .不同地区植物物候变化的主要驱动因素不同[1 ] .在热带地区,温度是控制植物物候变化的主要因素[14 ] ;在温带和寒带地区,植物物候变化依赖于温度、低温持续和光周期的综合作用[1 ] ;在草原生态系统,温度和湿度是植物物候的主要驱动因子[15 ] ;在干旱和半干旱生态系统中,水分是植物物候的关键驱动因子[16 ] .植物物候与气候因子的关系是全球变化的一个研究热点,反映陆地生态系统对气候变化的动态响应[11 ] . ...
Progress towards an interdisciplinary science of plant phenology:building predictions across space,time and species diversity
1
2014
... 近60年来,世界范围内的植物呈现出春季物候提前、秋季物候推迟、生长季延长的趋势[3 ] .遥感监测和地面定点人工观测作为常用的物候观测手段[8 ] ,均监测到了这一物候变化趋势.具体而言,在基于地面观测的物候变化的信息中,通过气候变化对中国植被影响的观测证据集成分析发现,87.4%的记录表明木本植物春、夏季物候呈提前趋势;71.5%的记录表明其秋季物候呈推迟趋势;有57.7%的记录表明其生长季长度呈延长趋势[10 ] .对中国温带植被1982—1999年物候变化的遥感数据分析显示,该区域植被春季物候提前,秋季物候推迟,生长季长度延长[11 ] .植物物候变化受气候(温度、降水、太阳辐射等)、植物生活史、植物的适应与进化策略等因素的影响[12 ] .其中,温度、降水和光照等气候因子是植物物候变化的主要驱动力[13 ] .不同地区植物物候变化的主要驱动因素不同[1 ] .在热带地区,温度是控制植物物候变化的主要因素[14 ] ;在温带和寒带地区,植物物候变化依赖于温度、低温持续和光周期的综合作用[1 ] ;在草原生态系统,温度和湿度是植物物候的主要驱动因子[15 ] ;在干旱和半干旱生态系统中,水分是植物物候的关键驱动因子[16 ] .植物物候与气候因子的关系是全球变化的一个研究热点,反映陆地生态系统对气候变化的动态响应[11 ] . ...
植物物候学研究进展
1
2020
... 近60年来,世界范围内的植物呈现出春季物候提前、秋季物候推迟、生长季延长的趋势[3 ] .遥感监测和地面定点人工观测作为常用的物候观测手段[8 ] ,均监测到了这一物候变化趋势.具体而言,在基于地面观测的物候变化的信息中,通过气候变化对中国植被影响的观测证据集成分析发现,87.4%的记录表明木本植物春、夏季物候呈提前趋势;71.5%的记录表明其秋季物候呈推迟趋势;有57.7%的记录表明其生长季长度呈延长趋势[10 ] .对中国温带植被1982—1999年物候变化的遥感数据分析显示,该区域植被春季物候提前,秋季物候推迟,生长季长度延长[11 ] .植物物候变化受气候(温度、降水、太阳辐射等)、植物生活史、植物的适应与进化策略等因素的影响[12 ] .其中,温度、降水和光照等气候因子是植物物候变化的主要驱动力[13 ] .不同地区植物物候变化的主要驱动因素不同[1 ] .在热带地区,温度是控制植物物候变化的主要因素[14 ] ;在温带和寒带地区,植物物候变化依赖于温度、低温持续和光周期的综合作用[1 ] ;在草原生态系统,温度和湿度是植物物候的主要驱动因子[15 ] ;在干旱和半干旱生态系统中,水分是植物物候的关键驱动因子[16 ] .植物物候与气候因子的关系是全球变化的一个研究热点,反映陆地生态系统对气候变化的动态响应[11 ] . ...
2001-2010年内蒙古植被覆盖度时空变化特征
1
2012
... 科尔沁沙地位于中国北方农牧交错带,生态环境脆弱,属于典型的半干旱沙地生态系统,是全球变化的敏感区域[17 ] .近年来,在全球气候变暖背景下,科尔沁沙地植被特别是优势灌木生育期也发生了显著变化.然而,已有研究多集中于单个物候初始期的变化对气候变化的响应,很少研究不同生活史植物物候序列对气候变化的响应[18 -19 ] ,生长季不同时期气象因子对优势灌木物候期的影响仍不充分.本研究的目的是分析2005—2019年科尔沁沙地沙质草地优势灌木物候的时序变化,进一步用PLS分析探讨该区域优势灌木物候与气候因子(温度、降水、太阳辐射)之间的关系,得到驱动优势灌木物候变化的关键气候因子,进一步认知气候变化对沙地植被的影响,以期为半干旱沙地植被保护和恢复提供理论依据. ...
Timing and duration of phenological sequences of alpine plants along an elevation gradient on the Tibetan Plateau
1
2014
... 科尔沁沙地位于中国北方农牧交错带,生态环境脆弱,属于典型的半干旱沙地生态系统,是全球变化的敏感区域[17 ] .近年来,在全球气候变暖背景下,科尔沁沙地植被特别是优势灌木生育期也发生了显著变化.然而,已有研究多集中于单个物候初始期的变化对气候变化的响应,很少研究不同生活史植物物候序列对气候变化的响应[18 -19 ] ,生长季不同时期气象因子对优势灌木物候期的影响仍不充分.本研究的目的是分析2005—2019年科尔沁沙地沙质草地优势灌木物候的时序变化,进一步用PLS分析探讨该区域优势灌木物候与气候因子(温度、降水、太阳辐射)之间的关系,得到驱动优势灌木物候变化的关键气候因子,进一步认知气候变化对沙地植被的影响,以期为半干旱沙地植被保护和恢复提供理论依据. ...
Relatively stable response of fruiting stage to warming and cooling relative to other phenological events
1
2016
... 科尔沁沙地位于中国北方农牧交错带,生态环境脆弱,属于典型的半干旱沙地生态系统,是全球变化的敏感区域[17 ] .近年来,在全球气候变暖背景下,科尔沁沙地植被特别是优势灌木生育期也发生了显著变化.然而,已有研究多集中于单个物候初始期的变化对气候变化的响应,很少研究不同生活史植物物候序列对气候变化的响应[18 -19 ] ,生长季不同时期气象因子对优势灌木物候期的影响仍不充分.本研究的目的是分析2005—2019年科尔沁沙地沙质草地优势灌木物候的时序变化,进一步用PLS分析探讨该区域优势灌木物候与气候因子(温度、降水、太阳辐射)之间的关系,得到驱动优势灌木物候变化的关键气候因子,进一步认知气候变化对沙地植被的影响,以期为半干旱沙地植被保护和恢复提供理论依据. ...
不同沙丘生境主要植物比叶面积和叶干物质含量的比较
1
2005
... 本试验在中国科学院奈曼沙漠化研究站的沙地综合观测场(42.92°N,120.70°E)进行,观测场地地形平坦,无明显起伏.该区域属于温带大陆性季风气候区[20 ] ,年均气温5.8~6.4 ℃,最热月(7月)平均气温23.5℃,最冷月(1月)平均气温-16.8 ℃[21 ] ,≥ 10 ℃年积温3 000~4 000 ℃,无霜期151 d[22 ] ,多年平均降水量280~360 mm,年均蒸发量1 600~2 400 mm,年均日照2 951.2 h,年均风速3.5~4.5 m·s-1[23 ] .该地区主要物种有灌木小叶锦鸡儿(Caragana microphylla )、灌木差巴嘎蒿(Artemisia halodendron )和多年生或一年生草本如扁蓿豆(Melissitus ruthenicus )、大果虫实(Corispermum macrocarpum )、狗尾草(Setaria viridis ).相对于草本植物,灌木具有更加耐旱和抗风蚀、沙埋的特点,是沙质草地植被的主要生活型[24 ] .其中差巴嘎蒿是一种典型的沙生中灌木,花期7—8月,果期8—9月,是良好的固沙先锋植物;小叶锦鸡儿一般4月萌发,5月下旬至6月上旬开花,6月中旬至7月中旬种子成熟,9月中旬以后逐渐落叶干枯,是沙质草地较早完成生活史周期的植物种. ...
Contrasting effects of plant inter‐ and intraspecific variation on community trait responses to restoration of a sandy grassland ecosystem
1
2017
... 本试验在中国科学院奈曼沙漠化研究站的沙地综合观测场(42.92°N,120.70°E)进行,观测场地地形平坦,无明显起伏.该区域属于温带大陆性季风气候区[20 ] ,年均气温5.8~6.4 ℃,最热月(7月)平均气温23.5℃,最冷月(1月)平均气温-16.8 ℃[21 ] ,≥ 10 ℃年积温3 000~4 000 ℃,无霜期151 d[22 ] ,多年平均降水量280~360 mm,年均蒸发量1 600~2 400 mm,年均日照2 951.2 h,年均风速3.5~4.5 m·s-1[23 ] .该地区主要物种有灌木小叶锦鸡儿(Caragana microphylla )、灌木差巴嘎蒿(Artemisia halodendron )和多年生或一年生草本如扁蓿豆(Melissitus ruthenicus )、大果虫实(Corispermum macrocarpum )、狗尾草(Setaria viridis ).相对于草本植物,灌木具有更加耐旱和抗风蚀、沙埋的特点,是沙质草地植被的主要生活型[24 ] .其中差巴嘎蒿是一种典型的沙生中灌木,花期7—8月,果期8—9月,是良好的固沙先锋植物;小叶锦鸡儿一般4月萌发,5月下旬至6月上旬开花,6月中旬至7月中旬种子成熟,9月中旬以后逐渐落叶干枯,是沙质草地较早完成生活史周期的植物种. ...
科尔沁沙地主要植物细根和叶片碳,氮,磷化学计量特征
1
2017
... 本试验在中国科学院奈曼沙漠化研究站的沙地综合观测场(42.92°N,120.70°E)进行,观测场地地形平坦,无明显起伏.该区域属于温带大陆性季风气候区[20 ] ,年均气温5.8~6.4 ℃,最热月(7月)平均气温23.5℃,最冷月(1月)平均气温-16.8 ℃[21 ] ,≥ 10 ℃年积温3 000~4 000 ℃,无霜期151 d[22 ] ,多年平均降水量280~360 mm,年均蒸发量1 600~2 400 mm,年均日照2 951.2 h,年均风速3.5~4.5 m·s-1[23 ] .该地区主要物种有灌木小叶锦鸡儿(Caragana microphylla )、灌木差巴嘎蒿(Artemisia halodendron )和多年生或一年生草本如扁蓿豆(Melissitus ruthenicus )、大果虫实(Corispermum macrocarpum )、狗尾草(Setaria viridis ).相对于草本植物,灌木具有更加耐旱和抗风蚀、沙埋的特点,是沙质草地植被的主要生活型[24 ] .其中差巴嘎蒿是一种典型的沙生中灌木,花期7—8月,果期8—9月,是良好的固沙先锋植物;小叶锦鸡儿一般4月萌发,5月下旬至6月上旬开花,6月中旬至7月中旬种子成熟,9月中旬以后逐渐落叶干枯,是沙质草地较早完成生活史周期的植物种. ...
半干旱区沙地芦苇对浅水位变化的生理生态响应
1
2013
... 本试验在中国科学院奈曼沙漠化研究站的沙地综合观测场(42.92°N,120.70°E)进行,观测场地地形平坦,无明显起伏.该区域属于温带大陆性季风气候区[20 ] ,年均气温5.8~6.4 ℃,最热月(7月)平均气温23.5℃,最冷月(1月)平均气温-16.8 ℃[21 ] ,≥ 10 ℃年积温3 000~4 000 ℃,无霜期151 d[22 ] ,多年平均降水量280~360 mm,年均蒸发量1 600~2 400 mm,年均日照2 951.2 h,年均风速3.5~4.5 m·s-1[23 ] .该地区主要物种有灌木小叶锦鸡儿(Caragana microphylla )、灌木差巴嘎蒿(Artemisia halodendron )和多年生或一年生草本如扁蓿豆(Melissitus ruthenicus )、大果虫实(Corispermum macrocarpum )、狗尾草(Setaria viridis ).相对于草本植物,灌木具有更加耐旱和抗风蚀、沙埋的特点,是沙质草地植被的主要生活型[24 ] .其中差巴嘎蒿是一种典型的沙生中灌木,花期7—8月,果期8—9月,是良好的固沙先锋植物;小叶锦鸡儿一般4月萌发,5月下旬至6月上旬开花,6月中旬至7月中旬种子成熟,9月中旬以后逐渐落叶干枯,是沙质草地较早完成生活史周期的植物种. ...
水分胁迫对几种固沙灌木幼苗生物量的影响
1
2014
... 本试验在中国科学院奈曼沙漠化研究站的沙地综合观测场(42.92°N,120.70°E)进行,观测场地地形平坦,无明显起伏.该区域属于温带大陆性季风气候区[20 ] ,年均气温5.8~6.4 ℃,最热月(7月)平均气温23.5℃,最冷月(1月)平均气温-16.8 ℃[21 ] ,≥ 10 ℃年积温3 000~4 000 ℃,无霜期151 d[22 ] ,多年平均降水量280~360 mm,年均蒸发量1 600~2 400 mm,年均日照2 951.2 h,年均风速3.5~4.5 m·s-1[23 ] .该地区主要物种有灌木小叶锦鸡儿(Caragana microphylla )、灌木差巴嘎蒿(Artemisia halodendron )和多年生或一年生草本如扁蓿豆(Melissitus ruthenicus )、大果虫实(Corispermum macrocarpum )、狗尾草(Setaria viridis ).相对于草本植物,灌木具有更加耐旱和抗风蚀、沙埋的特点,是沙质草地植被的主要生活型[24 ] .其中差巴嘎蒿是一种典型的沙生中灌木,花期7—8月,果期8—9月,是良好的固沙先锋植物;小叶锦鸡儿一般4月萌发,5月下旬至6月上旬开花,6月中旬至7月中旬种子成熟,9月中旬以后逐渐落叶干枯,是沙质草地较早完成生活史周期的植物种. ...
1
1979
... 灌木物候期观测以中国气象局农业气象观测规范为依据,当一半以上的所选植株达到某一发育期时,就记录该物种到了这一物候期.当明显看到植株长出绿色叶芽就是出芽期;当所观测植株上的芽从芽苞中发出卷曲着的或按叶脉褶叠着的小叶,出现第一批有一二片的叶片平展时就是展叶期;当观测到一半以上的植株有一朵或同时有几朵花的花瓣开始完全开放即为始花期;在观测的植株上有一半以上的花蕾都展开花瓣,或一半以上的花序散出花粉为盛花期;当观测的植株上有一半的果实或种子变为成熟时的颜色为结果期;所观测植株的叶子在秋天开始变色为秋季叶变色;当所观测植株秋季开始落叶记录为落叶期[25 ] . ...
Partial least squares regression for analyzing walnut phenology in California
1
2012
... 本研究采用一元线性方程定量描述气象和物候要素的多年变化特征,其斜率可以反映各要素的变化速率.采用偏最小二乘回归(PLS)分析最高气温、最低气温、平均气温、降水量和太阳辐射等气候要素对植物物候的潜在影响[8 ,26 -29 ] .由于研究区灌木的生长季末期平均出现在10月,因此选取2种优势灌木上年生长季结束至次年物候期发生前的温度、降水量和太阳辐射,应用PLS分析其与物候参数的关系.研究中气象要素和物候要素的一元线性回归在Origin 2024中完成,PLS分析在R-4.0.3中进行,插图用Origin 2024绘制. ...
Response of chestnut phenology in China to climate variation and change
0
2013
克里雅河流域植被物候时空变化及影响因素
0
2020
环京津贫困带土地利用变化的地形梯度效应分析
1
2014
... 本研究采用一元线性方程定量描述气象和物候要素的多年变化特征,其斜率可以反映各要素的变化速率.采用偏最小二乘回归(PLS)分析最高气温、最低气温、平均气温、降水量和太阳辐射等气候要素对植物物候的潜在影响[8 ,26 -29 ] .由于研究区灌木的生长季末期平均出现在10月,因此选取2种优势灌木上年生长季结束至次年物候期发生前的温度、降水量和太阳辐射,应用PLS分析其与物候参数的关系.研究中气象要素和物候要素的一元线性回归在Origin 2024中完成,PLS分析在R-4.0.3中进行,插图用Origin 2024绘制. ...
近50年云南区域气候变化特征分析
1
2008
... 温度是影响植物物候期的主导因素[30 ] ,无论是温度高低、有效积温的多少,还是低温刺激,都能显著影响植物物候的演变.早期植物物候(如出芽、展叶、开花)的提前或推迟主要受春季气温的影响;中期植物物候(如结果)主要与生长季积温相关;晚期植物物候(如秋季叶变色、落叶)主要取决于阈值温度的短期作用,一旦气温下降到某个临界值就会引发这些变化[13 ] .已有研究表明,气温升高会导致荒漠区植被提前返青.与本研究结果相吻合的是,我们观察到差巴嘎蒿和小叶锦鸡儿在生长季初期的物候指标(展叶期)均有所提前,且这一变化与12、1、3、4月的气温升高密切相关(图3 ~4 ).究其原因,可能包括以下两个方面:一方面,温度升高会使植被发育所需的有效积温更早达到,从而促进植被返青[31 ] ;另一方面,植物的生理活动涉及一系列复杂的生化反应,温度升高能够增强酶的活性,从而加速植物的发育进程[32 ] .而生长季末期物候指标(秋季叶变色)主要与7月气温升高有关,原因可能是温度升高会使植株光合作用增强,降低了植物体内叶绿素分解速率[33 ] 、提高了酶活性[34 ] 、并推迟了霜冻的出现[35 ] ,从而使植物秋季叶变色推后[36 ] .此外,生长季延长主要与春夏季节气温升高有关,春季(3、4月)气温上升主要影响生长季初期的物候变化,夏季(7月)气温升高则影响生长季末期,从而共同调节生长季长度,这与以往的研究结果一致[37 -42 ] .总体而言,温度升高会在春季解除休眠并推迟秋季休眠,从而延长植物的生活史周期[1 ] . ...
Vegetation phenology on the Qinghai-Tibetan Plateau and its response to climate change (1982-2013)
2
2018
... 温度是影响植物物候期的主导因素[30 ] ,无论是温度高低、有效积温的多少,还是低温刺激,都能显著影响植物物候的演变.早期植物物候(如出芽、展叶、开花)的提前或推迟主要受春季气温的影响;中期植物物候(如结果)主要与生长季积温相关;晚期植物物候(如秋季叶变色、落叶)主要取决于阈值温度的短期作用,一旦气温下降到某个临界值就会引发这些变化[13 ] .已有研究表明,气温升高会导致荒漠区植被提前返青.与本研究结果相吻合的是,我们观察到差巴嘎蒿和小叶锦鸡儿在生长季初期的物候指标(展叶期)均有所提前,且这一变化与12、1、3、4月的气温升高密切相关(图3 ~4 ).究其原因,可能包括以下两个方面:一方面,温度升高会使植被发育所需的有效积温更早达到,从而促进植被返青[31 ] ;另一方面,植物的生理活动涉及一系列复杂的生化反应,温度升高能够增强酶的活性,从而加速植物的发育进程[32 ] .而生长季末期物候指标(秋季叶变色)主要与7月气温升高有关,原因可能是温度升高会使植株光合作用增强,降低了植物体内叶绿素分解速率[33 ] 、提高了酶活性[34 ] 、并推迟了霜冻的出现[35 ] ,从而使植物秋季叶变色推后[36 ] .此外,生长季延长主要与春夏季节气温升高有关,春季(3、4月)气温上升主要影响生长季初期的物候变化,夏季(7月)气温升高则影响生长季末期,从而共同调节生长季长度,这与以往的研究结果一致[37 -42 ] .总体而言,温度升高会在春季解除休眠并推迟秋季休眠,从而延长植物的生活史周期[1 ] . ...
... 光作为植物重要的能量来源,可通过影响植物的光敏色素,进而通过辐射强度、辐射长度以及光谱成分来影响植物生活史的很多阶段[1 ,38 ] .本研究发现8月太阳辐射增加使秋季叶变色、落叶期提前,生长季长度缩短(图3 ~4 ).推测其主要原因是因为太阳辐射升高,植被蒸腾作用增强,植物可利用水资源减少而导致[31 ] . ...
1980-2005年中国东北木本植物物候特征及其对气温的响应
1
2010
... 温度是影响植物物候期的主导因素[30 ] ,无论是温度高低、有效积温的多少,还是低温刺激,都能显著影响植物物候的演变.早期植物物候(如出芽、展叶、开花)的提前或推迟主要受春季气温的影响;中期植物物候(如结果)主要与生长季积温相关;晚期植物物候(如秋季叶变色、落叶)主要取决于阈值温度的短期作用,一旦气温下降到某个临界值就会引发这些变化[13 ] .已有研究表明,气温升高会导致荒漠区植被提前返青.与本研究结果相吻合的是,我们观察到差巴嘎蒿和小叶锦鸡儿在生长季初期的物候指标(展叶期)均有所提前,且这一变化与12、1、3、4月的气温升高密切相关(图3 ~4 ).究其原因,可能包括以下两个方面:一方面,温度升高会使植被发育所需的有效积温更早达到,从而促进植被返青[31 ] ;另一方面,植物的生理活动涉及一系列复杂的生化反应,温度升高能够增强酶的活性,从而加速植物的发育进程[32 ] .而生长季末期物候指标(秋季叶变色)主要与7月气温升高有关,原因可能是温度升高会使植株光合作用增强,降低了植物体内叶绿素分解速率[33 ] 、提高了酶活性[34 ] 、并推迟了霜冻的出现[35 ] ,从而使植物秋季叶变色推后[36 ] .此外,生长季延长主要与春夏季节气温升高有关,春季(3、4月)气温上升主要影响生长季初期的物候变化,夏季(7月)气温升高则影响生长季末期,从而共同调节生长季长度,这与以往的研究结果一致[37 -42 ] .总体而言,温度升高会在春季解除休眠并推迟秋季休眠,从而延长植物的生活史周期[1 ] . ...
The control of autumn senescence in European aspens
1
2009
... 温度是影响植物物候期的主导因素[30 ] ,无论是温度高低、有效积温的多少,还是低温刺激,都能显著影响植物物候的演变.早期植物物候(如出芽、展叶、开花)的提前或推迟主要受春季气温的影响;中期植物物候(如结果)主要与生长季积温相关;晚期植物物候(如秋季叶变色、落叶)主要取决于阈值温度的短期作用,一旦气温下降到某个临界值就会引发这些变化[13 ] .已有研究表明,气温升高会导致荒漠区植被提前返青.与本研究结果相吻合的是,我们观察到差巴嘎蒿和小叶锦鸡儿在生长季初期的物候指标(展叶期)均有所提前,且这一变化与12、1、3、4月的气温升高密切相关(图3 ~4 ).究其原因,可能包括以下两个方面:一方面,温度升高会使植被发育所需的有效积温更早达到,从而促进植被返青[31 ] ;另一方面,植物的生理活动涉及一系列复杂的生化反应,温度升高能够增强酶的活性,从而加速植物的发育进程[32 ] .而生长季末期物候指标(秋季叶变色)主要与7月气温升高有关,原因可能是温度升高会使植株光合作用增强,降低了植物体内叶绿素分解速率[33 ] 、提高了酶活性[34 ] 、并推迟了霜冻的出现[35 ] ,从而使植物秋季叶变色推后[36 ] .此外,生长季延长主要与春夏季节气温升高有关,春季(3、4月)气温上升主要影响生长季初期的物候变化,夏季(7月)气温升高则影响生长季末期,从而共同调节生长季长度,这与以往的研究结果一致[37 -42 ] .总体而言,温度升高会在春季解除休眠并推迟秋季休眠,从而延长植物的生活史周期[1 ] . ...
Effects of warming on chlorophyll degradation and carbohydrate accumulation of alpine herbaceous species during plant senescence on the Tibetan Plateau
1
2014
... 温度是影响植物物候期的主导因素[30 ] ,无论是温度高低、有效积温的多少,还是低温刺激,都能显著影响植物物候的演变.早期植物物候(如出芽、展叶、开花)的提前或推迟主要受春季气温的影响;中期植物物候(如结果)主要与生长季积温相关;晚期植物物候(如秋季叶变色、落叶)主要取决于阈值温度的短期作用,一旦气温下降到某个临界值就会引发这些变化[13 ] .已有研究表明,气温升高会导致荒漠区植被提前返青.与本研究结果相吻合的是,我们观察到差巴嘎蒿和小叶锦鸡儿在生长季初期的物候指标(展叶期)均有所提前,且这一变化与12、1、3、4月的气温升高密切相关(图3 ~4 ).究其原因,可能包括以下两个方面:一方面,温度升高会使植被发育所需的有效积温更早达到,从而促进植被返青[31 ] ;另一方面,植物的生理活动涉及一系列复杂的生化反应,温度升高能够增强酶的活性,从而加速植物的发育进程[32 ] .而生长季末期物候指标(秋季叶变色)主要与7月气温升高有关,原因可能是温度升高会使植株光合作用增强,降低了植物体内叶绿素分解速率[33 ] 、提高了酶活性[34 ] 、并推迟了霜冻的出现[35 ] ,从而使植物秋季叶变色推后[36 ] .此外,生长季延长主要与春夏季节气温升高有关,春季(3、4月)气温上升主要影响生长季初期的物候变化,夏季(7月)气温升高则影响生长季末期,从而共同调节生长季长度,这与以往的研究结果一致[37 -42 ] .总体而言,温度升高会在春季解除休眠并推迟秋季休眠,从而延长植物的生活史周期[1 ] . ...
1
2013
... 温度是影响植物物候期的主导因素[30 ] ,无论是温度高低、有效积温的多少,还是低温刺激,都能显著影响植物物候的演变.早期植物物候(如出芽、展叶、开花)的提前或推迟主要受春季气温的影响;中期植物物候(如结果)主要与生长季积温相关;晚期植物物候(如秋季叶变色、落叶)主要取决于阈值温度的短期作用,一旦气温下降到某个临界值就会引发这些变化[13 ] .已有研究表明,气温升高会导致荒漠区植被提前返青.与本研究结果相吻合的是,我们观察到差巴嘎蒿和小叶锦鸡儿在生长季初期的物候指标(展叶期)均有所提前,且这一变化与12、1、3、4月的气温升高密切相关(图3 ~4 ).究其原因,可能包括以下两个方面:一方面,温度升高会使植被发育所需的有效积温更早达到,从而促进植被返青[31 ] ;另一方面,植物的生理活动涉及一系列复杂的生化反应,温度升高能够增强酶的活性,从而加速植物的发育进程[32 ] .而生长季末期物候指标(秋季叶变色)主要与7月气温升高有关,原因可能是温度升高会使植株光合作用增强,降低了植物体内叶绿素分解速率[33 ] 、提高了酶活性[34 ] 、并推迟了霜冻的出现[35 ] ,从而使植物秋季叶变色推后[36 ] .此外,生长季延长主要与春夏季节气温升高有关,春季(3、4月)气温上升主要影响生长季初期的物候变化,夏季(7月)气温升高则影响生长季末期,从而共同调节生长季长度,这与以往的研究结果一致[37 -42 ] .总体而言,温度升高会在春季解除休眠并推迟秋季休眠,从而延长植物的生活史周期[1 ] . ...
Temperature,precipitation,and insolation effects on autumn vegetation phenology in temperate China
1
2016
... 温度是影响植物物候期的主导因素[30 ] ,无论是温度高低、有效积温的多少,还是低温刺激,都能显著影响植物物候的演变.早期植物物候(如出芽、展叶、开花)的提前或推迟主要受春季气温的影响;中期植物物候(如结果)主要与生长季积温相关;晚期植物物候(如秋季叶变色、落叶)主要取决于阈值温度的短期作用,一旦气温下降到某个临界值就会引发这些变化[13 ] .已有研究表明,气温升高会导致荒漠区植被提前返青.与本研究结果相吻合的是,我们观察到差巴嘎蒿和小叶锦鸡儿在生长季初期的物候指标(展叶期)均有所提前,且这一变化与12、1、3、4月的气温升高密切相关(图3 ~4 ).究其原因,可能包括以下两个方面:一方面,温度升高会使植被发育所需的有效积温更早达到,从而促进植被返青[31 ] ;另一方面,植物的生理活动涉及一系列复杂的生化反应,温度升高能够增强酶的活性,从而加速植物的发育进程[32 ] .而生长季末期物候指标(秋季叶变色)主要与7月气温升高有关,原因可能是温度升高会使植株光合作用增强,降低了植物体内叶绿素分解速率[33 ] 、提高了酶活性[34 ] 、并推迟了霜冻的出现[35 ] ,从而使植物秋季叶变色推后[36 ] .此外,生长季延长主要与春夏季节气温升高有关,春季(3、4月)气温上升主要影响生长季初期的物候变化,夏季(7月)气温升高则影响生长季末期,从而共同调节生长季长度,这与以往的研究结果一致[37 -42 ] .总体而言,温度升高会在春季解除休眠并推迟秋季休眠,从而延长植物的生活史周期[1 ] . ...
草本植物物候及其物候模拟模型的研究进展
2
2011
... 温度是影响植物物候期的主导因素[30 ] ,无论是温度高低、有效积温的多少,还是低温刺激,都能显著影响植物物候的演变.早期植物物候(如出芽、展叶、开花)的提前或推迟主要受春季气温的影响;中期植物物候(如结果)主要与生长季积温相关;晚期植物物候(如秋季叶变色、落叶)主要取决于阈值温度的短期作用,一旦气温下降到某个临界值就会引发这些变化[13 ] .已有研究表明,气温升高会导致荒漠区植被提前返青.与本研究结果相吻合的是,我们观察到差巴嘎蒿和小叶锦鸡儿在生长季初期的物候指标(展叶期)均有所提前,且这一变化与12、1、3、4月的气温升高密切相关(图3 ~4 ).究其原因,可能包括以下两个方面:一方面,温度升高会使植被发育所需的有效积温更早达到,从而促进植被返青[31 ] ;另一方面,植物的生理活动涉及一系列复杂的生化反应,温度升高能够增强酶的活性,从而加速植物的发育进程[32 ] .而生长季末期物候指标(秋季叶变色)主要与7月气温升高有关,原因可能是温度升高会使植株光合作用增强,降低了植物体内叶绿素分解速率[33 ] 、提高了酶活性[34 ] 、并推迟了霜冻的出现[35 ] ,从而使植物秋季叶变色推后[36 ] .此外,生长季延长主要与春夏季节气温升高有关,春季(3、4月)气温上升主要影响生长季初期的物候变化,夏季(7月)气温升高则影响生长季末期,从而共同调节生长季长度,这与以往的研究结果一致[37 -42 ] .总体而言,温度升高会在春季解除休眠并推迟秋季休眠,从而延长植物的生活史周期[1 ] . ...
... 水分同样会影响植物物候[1 ] ,植物从生存到光合作用以及营养的传输等过程都离不开水分[37 ] .本研究结果显示(图3 ~4 ),两种灌木的发芽期和展叶期与4月降水量之间并无显著相关.然而,已有诸多研究指出,降水增加会导致早期植物物候提前[43 -45 ] .这一差异或许与研究区域和植物种类的差异有关[46 ] .在本研究中,差巴嘎蒿与小叶锦鸡儿的返青时间显著提前(图1 ),很可能是由于4月气温升高及降雪减少等因素所致,而与水分变化并无显著关联.6月的降雨增加使差巴嘎蒿秋季落叶期推迟,小叶锦鸡儿生长季变长;8月降雨增加导致差巴嘎蒿秋季叶变色推迟,并延长其生长季,这与此前的研究结果相符[43 ,47 ] ,即夏季降水增加会导致晚期植物物候推迟.这可能是因为在满足植物生长所需热量条件下,水分的增加能为植物提供更多可利用的水资源,从而使秋季叶变色和落叶期推后,生长季得以延长. ...
植物物候研究进展
1
2006
... 光作为植物重要的能量来源,可通过影响植物的光敏色素,进而通过辐射强度、辐射长度以及光谱成分来影响植物生活史的很多阶段[1 ,38 ] .本研究发现8月太阳辐射增加使秋季叶变色、落叶期提前,生长季长度缩短(图3 ~4 ).推测其主要原因是因为太阳辐射升高,植被蒸腾作用增强,植物可利用水资源减少而导致[31 ] . ...
European phenological response to climate change matches the warming pattern
0
2006
Onset of spring starting earlier across the Northern Hemisphere
0
2006
Recent spring phenology shifts in western Central Europe based on multiscale observations
0
2014
天山北坡荒漠草原主要植物物候特征对气候变化的响应
1
2022
... 温度是影响植物物候期的主导因素[30 ] ,无论是温度高低、有效积温的多少,还是低温刺激,都能显著影响植物物候的演变.早期植物物候(如出芽、展叶、开花)的提前或推迟主要受春季气温的影响;中期植物物候(如结果)主要与生长季积温相关;晚期植物物候(如秋季叶变色、落叶)主要取决于阈值温度的短期作用,一旦气温下降到某个临界值就会引发这些变化[13 ] .已有研究表明,气温升高会导致荒漠区植被提前返青.与本研究结果相吻合的是,我们观察到差巴嘎蒿和小叶锦鸡儿在生长季初期的物候指标(展叶期)均有所提前,且这一变化与12、1、3、4月的气温升高密切相关(图3 ~4 ).究其原因,可能包括以下两个方面:一方面,温度升高会使植被发育所需的有效积温更早达到,从而促进植被返青[31 ] ;另一方面,植物的生理活动涉及一系列复杂的生化反应,温度升高能够增强酶的活性,从而加速植物的发育进程[32 ] .而生长季末期物候指标(秋季叶变色)主要与7月气温升高有关,原因可能是温度升高会使植株光合作用增强,降低了植物体内叶绿素分解速率[33 ] 、提高了酶活性[34 ] 、并推迟了霜冻的出现[35 ] ,从而使植物秋季叶变色推后[36 ] .此外,生长季延长主要与春夏季节气温升高有关,春季(3、4月)气温上升主要影响生长季初期的物候变化,夏季(7月)气温升高则影响生长季末期,从而共同调节生长季长度,这与以往的研究结果一致[37 -42 ] .总体而言,温度升高会在春季解除休眠并推迟秋季休眠,从而延长植物的生活史周期[1 ] . ...
新疆地区草地植被物候时空变化
2
2018
... 水分同样会影响植物物候[1 ] ,植物从生存到光合作用以及营养的传输等过程都离不开水分[37 ] .本研究结果显示(图3 ~4 ),两种灌木的发芽期和展叶期与4月降水量之间并无显著相关.然而,已有诸多研究指出,降水增加会导致早期植物物候提前[43 -45 ] .这一差异或许与研究区域和植物种类的差异有关[46 ] .在本研究中,差巴嘎蒿与小叶锦鸡儿的返青时间显著提前(图1 ),很可能是由于4月气温升高及降雪减少等因素所致,而与水分变化并无显著关联.6月的降雨增加使差巴嘎蒿秋季落叶期推迟,小叶锦鸡儿生长季变长;8月降雨增加导致差巴嘎蒿秋季叶变色推迟,并延长其生长季,这与此前的研究结果相符[43 ,47 ] ,即夏季降水增加会导致晚期植物物候推迟.这可能是因为在满足植物生长所需热量条件下,水分的增加能为植物提供更多可利用的水资源,从而使秋季叶变色和落叶期推后,生长季得以延长. ...
... [43 ,47 ],即夏季降水增加会导致晚期植物物候推迟.这可能是因为在满足植物生长所需热量条件下,水分的增加能为植物提供更多可利用的水资源,从而使秋季叶变色和落叶期推后,生长季得以延长. ...
内蒙古羊草草原物候及其对气候变化的响应
0
2016
呼伦贝尔草原物候变化及其与气象因子的关系
1
2019
... 水分同样会影响植物物候[1 ] ,植物从生存到光合作用以及营养的传输等过程都离不开水分[37 ] .本研究结果显示(图3 ~4 ),两种灌木的发芽期和展叶期与4月降水量之间并无显著相关.然而,已有诸多研究指出,降水增加会导致早期植物物候提前[43 -45 ] .这一差异或许与研究区域和植物种类的差异有关[46 ] .在本研究中,差巴嘎蒿与小叶锦鸡儿的返青时间显著提前(图1 ),很可能是由于4月气温升高及降雪减少等因素所致,而与水分变化并无显著关联.6月的降雨增加使差巴嘎蒿秋季落叶期推迟,小叶锦鸡儿生长季变长;8月降雨增加导致差巴嘎蒿秋季叶变色推迟,并延长其生长季,这与此前的研究结果相符[43 ,47 ] ,即夏季降水增加会导致晚期植物物候推迟.这可能是因为在满足植物生长所需热量条件下,水分的增加能为植物提供更多可利用的水资源,从而使秋季叶变色和落叶期推后,生长季得以延长. ...
中国北方草地植被物候变化及其对气候变化的响应
1
2019
... 水分同样会影响植物物候[1 ] ,植物从生存到光合作用以及营养的传输等过程都离不开水分[37 ] .本研究结果显示(图3 ~4 ),两种灌木的发芽期和展叶期与4月降水量之间并无显著相关.然而,已有诸多研究指出,降水增加会导致早期植物物候提前[43 -45 ] .这一差异或许与研究区域和植物种类的差异有关[46 ] .在本研究中,差巴嘎蒿与小叶锦鸡儿的返青时间显著提前(图1 ),很可能是由于4月气温升高及降雪减少等因素所致,而与水分变化并无显著关联.6月的降雨增加使差巴嘎蒿秋季落叶期推迟,小叶锦鸡儿生长季变长;8月降雨增加导致差巴嘎蒿秋季叶变色推迟,并延长其生长季,这与此前的研究结果相符[43 ,47 ] ,即夏季降水增加会导致晚期植物物候推迟.这可能是因为在满足植物生长所需热量条件下,水分的增加能为植物提供更多可利用的水资源,从而使秋季叶变色和落叶期推后,生长季得以延长. ...
时序NDVI数据集重建方法评价与实例研究
1
2009
... 水分同样会影响植物物候[1 ] ,植物从生存到光合作用以及营养的传输等过程都离不开水分[37 ] .本研究结果显示(图3 ~4 ),两种灌木的发芽期和展叶期与4月降水量之间并无显著相关.然而,已有诸多研究指出,降水增加会导致早期植物物候提前[43 -45 ] .这一差异或许与研究区域和植物种类的差异有关[46 ] .在本研究中,差巴嘎蒿与小叶锦鸡儿的返青时间显著提前(图1 ),很可能是由于4月气温升高及降雪减少等因素所致,而与水分变化并无显著关联.6月的降雨增加使差巴嘎蒿秋季落叶期推迟,小叶锦鸡儿生长季变长;8月降雨增加导致差巴嘎蒿秋季叶变色推迟,并延长其生长季,这与此前的研究结果相符[43 ,47 ] ,即夏季降水增加会导致晚期植物物候推迟.这可能是因为在满足植物生长所需热量条件下,水分的增加能为植物提供更多可利用的水资源,从而使秋季叶变色和落叶期推后,生长季得以延长. ...





 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}