极端干旱对科尔沁沙地小叶锦鸡儿( Caragana microphylla
夏咪 , 1 , 2 , 罗亚勇 , 1 , 2 , 赵鑫宇 3 , 王鹤松 4 , 陈丙昊 5 , 石璨瑜 1 , 2
1.中国科学院西北生态环境资源研究院 奈曼沙漠化研究站,甘肃 兰州 730000
2.中国科学院大学 资源与环境学院,北京 101408
3.内蒙古农业大学 沙漠治理学院,内蒙古 呼和浩特 010018
4.内蒙古民族大学 草业学院,内蒙古 通辽 028043
5.内蒙古大学 生态与环境学院,内蒙古 呼和浩特 010020
The impact of extreme drought on soil respiration in Caragana microphylla habitats in the Horqin Sandy Land
Xia Mi , 1 , 2 , Luo Yayong , 1 , 2 , Zhao Xinyu 3 , Wang Hesong 4 , Chen Binghao 5 , Shi Canyu 1 , 2
1.Naiman Desertification Research Station,Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
2.College of Resources and Environment,University of Chinese Academy of Sciences,Beijing 101408,China
3.College of Desert Control Science and Engineering,Inner Mongolia Agricultural University,Hohhot 010018,China
4.College of Grassland Science,Inner Mongolia Minzu University,Tongliao 028043,Inner Mongolia,China
5.School of Ecology and Environment,Inner Mongolia University,Hohhot 010020,China
通讯作者: 罗亚勇(E-mail: luoyy@lzb.ac.cn )
收稿日期: 2025-04-02
修回日期: 2025-06-24
基金资助:
国家自然科学基金项目 . 42275132 内蒙古自治区重点研发和成果转化计划项目 . 2022YFDZ0028 中国科学院“西部之光”人才培养引进计划项目
Received: 2025-04-02
Revised: 2025-06-24
作者简介 About authors
夏咪(2001—),女,河南南阳人,硕士研究生,研究方向为土壤生态学E-mail:3209056738@qq.com
, E-mail:3209056738@qq.com
摘要
土壤呼吸是陆地生态系统碳循环的重要组成部分,研究极端干旱下科尔沁沙地土壤呼吸与环境因子的动态变化,以期揭示降水格局变化对小叶锦鸡儿灌丛土壤呼吸的影响。本研究基于2024年在科尔沁沙地腹地开展的野外控制试验,设置了自然降水、减雨50%、6月干旱、7月干旱和8月干旱5个水平。结果表明:生长季土壤呼吸速率均呈现先上升后下降的趋势;生长季长于1个月的持续干旱期间灌丛土壤呼吸显著降低,减雨50%对土壤呼吸速率有影响但不显著;土壤呼吸速率与土壤温度呈显著的指数关系,与土壤含水量线性关系不显著;土壤水热因子、植物特征和酶活性可解释土壤呼吸速率变异的48%,其中土壤水热因子对其有最高的独立解释力(37%)。
关键词:
极端干旱 土壤呼吸 科尔沁沙地 小叶锦鸡儿
Abstract
Soil respiration is a crucial component of the terrestrial ecosystem carbon cycle. This study investigated the dynamic changes in soil respiration and environmental factors under extreme drought conditions in the Horqin Sandy Land, aiming to reveal the impacts of precipitation pattern alterations on soil respiration in Caragana microphylla . Based on field control experiments conducted in 2024 in the hinterland of Horqin Sandy Land, five treatments were established: natural precipitation, 50% rainfall reduction, June drought, July drought, and August drought. The results showed that: The soil respiration rate during the growing season initially increased and then decreased; Continuous drought lasting over one month during the growing season significantly reduced soil respiration, while 50% rainfall reduction had non-significant effects; The soil respiration rate exhibited a significant exponential relationship with soil temperature, but showed no significant linear correlation with soil moisture content; Soil hydrothermal factors, plant characteristics, and enzyme activities collectively explained 48% of the variation in soil respiration rates, with soil hydrothermal factors demonstrating the highest independent explanatory power (37%). This study provides important scientific references for accurately estimating carbon emissions in Horqin Sandy Land under future precipitation change scenarios.
Keywords:
extreme drought soil respiration Horqin Sandy Land Caragana microphylla
本文引用格式
夏咪, 罗亚勇, 赵鑫宇, 王鹤松, 陈丙昊, 石璨瑜. 极端干旱对科尔沁沙地小叶锦鸡儿( Caragana microphylla . 中国沙漠 [J], 2025, 45(4): 334-342 doi:10.7522/j.issn.1000-694X.2025.00174
Xia Mi, Luo Yayong, Zhao Xinyu, Wang Hesong, Chen Binghao, Shi Canyu. The impact of extreme drought on soil respiration in Caragana microphylla habitats in the Horqin Sandy Land . Journal of Desert Research
0 引言
干旱半干旱地区约占全球陆地面积的41%,贡献了全球超过25%的碳储量,对陆地生态系统碳循环起重要作用[1 ] 。土壤呼吸是陆地生态系统向大气输出CO2 的主要途径,占整个生态系统呼吸总量的60%~90%,是陆地生态系统碳循环的重要组成部分[2 -3 ] 。2021年IPCC报告提出,与土壤呼吸有关的碳通量大约是与化石燃料燃烧相关的碳通量的10倍[4 ] ,这凸显了其在调控全球碳循环过程中的关键地位,土壤呼吸微弱的变化将会引起大气CO2 浓度的显著改变。因此,研究干旱半干旱地区土壤呼吸对环境变化的响应对准确估算陆地生态系统碳排放量至关重要。
土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用。在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] 。环境因子的这种变化必然会对土壤呼吸过程产生深刻影响。陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平。值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] 。Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] 。土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] 。尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏。科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区。1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] 。在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种。目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落。而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差。因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要。
基于此,本文以科尔沁沙地的小叶锦鸡儿灌丛为研究对象,通过设置不同极端干旱处理,量化不同极端干旱处理下科尔沁沙地灌丛生态系统土壤呼吸速率的时间动态变化特征,分析土壤水热因子、植物特征和酶活性对土壤呼吸动态的调控作用,并明确其主要影响因素,以期为准确预测未来降水格局变化驱动的干旱半干旱地区土壤碳排放变化提供数据支撑。
1 材料与方法
1.1 研究区概况
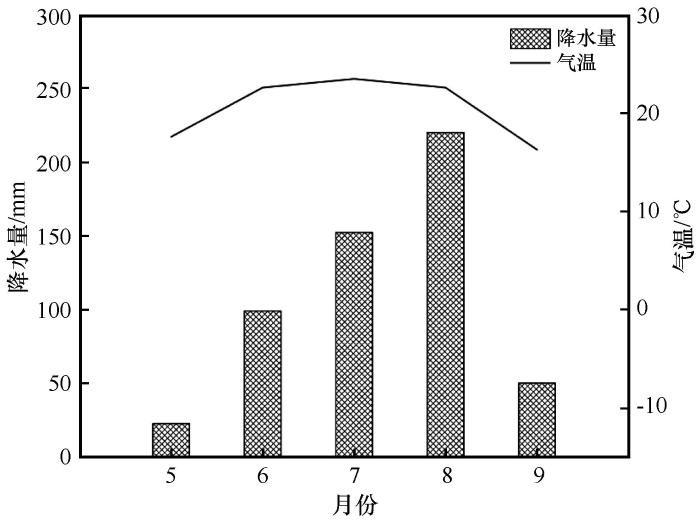
研究区位于中国科尔沁沙地腹地(42°56′N,120°45′E),海拔约360 m。该地区属于温带大陆性半干旱气候,春季干燥多风,夏季炎热多雨,秋季凉爽短暂,冬季寒冷漫长[20 ] 。年降水量360 mm,集中在6—8月且分布不均(图1 ),年潜在蒸发量1 500~2 500 mm。年均气温6.4 ℃,≥10 ℃积温3 000~3 400 ℃。主要土壤类型为风沙土,土壤全碳和全氮含量分别为0.019、 0.252 g·kg-1 ,饱和含水量0.234 g·g-1 ,土壤容重为1.518 g·cm-3 ,土壤pH为7.1,电导率是30.44 μS·cm-1 。植物群落结构组成简单,以灌丛和草本为主,优势植物种有小叶锦鸡儿、猪毛蒿(Artemisia Scoparia )、糙隐子草(Cleistogenes squarrosa )、狗尾草(Setaria viridis )、尖头叶藜(Cheno podium acuminatum )等。
图1
图1
2024年研究区5—9月平均气温和月降水量
Fig.1
Monthly precipitation and average temperature in the study area in 2024
1.2 试验设计
本试验选取统一种植年限的小叶锦鸡儿灌丛典型样地,建立围栏以开展长期控制试验。科尔沁沙地近10年降水分布呈现出显著的年际波动性和年内分配不均的特征[20 ,23 ] ,且科尔沁沙地生态系统的草本或灌木在生长季的不同月份处于不同的关键物候阶段(如返青、快速生长、开花、结实)和生理敏感期(如分蘖、花芽分化),该区域生长季(6—8月)降水是生态系统水分供应的绝对关键期,但其总量和月度分布存在高度不确定性。基于此,共设置了5种极端干旱处理,分别为自然对照组(W1)、减雨50%处理组(W2)、6月干旱处理组(W3)、7月干旱处理组(W4)和8月干旱处理组(W5)。试验采用完全随机设计,每种处理设置5个重复,共计25个试验小区。每个小区面积为7 m×7 m,小区间隔约10 m,以避免不同处理间土壤水分的相互干扰。减雨50%处理组(W2)通过在试验小区上方搭建间隔式遮雨棚(宽度50 cm、间隔50 cm)实现,遮雨棚采用透光率为90%的聚丙烯塑料板材构建,板材之间预留与板材宽度相同的空隙,以确保减少50%的自然降水量。干旱处理组(W3、W4、W5)同样在试验小区上方搭建全覆盖遮雨棚,并使用透光率为90%的塑料棚完全覆盖棚顶和两侧,两头保持通风状态。其中,6月干旱处理组仅在6月实施干旱处理,其余月份与自然对照组保持一致;7月干旱处理组和8月干旱处理组分别于7月和8月实施干旱处理,其余月份亦与自然对照组一致。
1.3 测定方法
1.3.1 土壤呼吸的测定
土壤呼吸速率采用Li-6400X便携式光合作用测定仪(LI-COR.,美国)测定,2024年4月上旬于每个处理小区内在小叶锦鸡儿灌丛下布置1个土壤呼吸测定所用的PVC材质的呼吸环(高度10 cm,直径20 cm),每个土壤呼吸环露出地表高度3 cm,共布置25个土壤呼吸环,安装时尽可能避免地面植被的破坏,整个试验期间呼吸环位置、高度不变,以实现土壤呼吸速率的长期定点监测。生长季6—9月每个月定期测定土壤呼吸速率2~3次,08:00—11:00土壤呼吸速率变化斜率最小,且此时测量值最接近日均值[24 ] ,故每次测量均选择晴朗无风的08:00—11:00测定。土壤呼吸是通过在90 s间隔内腔室顶部空间CO2 的增加来测量的,因此,在每次测量前1天,用剪刀齐地面剪去土壤呼吸环内的植物的地上部分,以消除地上植物部分对土壤呼吸速率的影响[24 -25 ] 。
1.3.2 土壤水热因子、植物特征及酶活性的测定
于每个样地在小叶锦鸡儿灌丛下10 cm处分别放置一个iButton纽扣式温度记录仪(美国Maxim公司),设定为每2 h记录1次土壤的温度,土壤含水量的测定采用烘干法。于生长季末期8月底测定土壤呼吸的同时,测定根生物量,采用土钻法(内径为10 cm)在试验样地采集0~10 cm的根芯,挑出植物根系用清水冲洗干净,将干净的根系于65 ℃烘箱中烘干至恒重后称重。
同时用土钻采集灌丛下土壤样品,在每个处理小区随机选取3钻0~10 cm土层土壤,混合均匀后过2 mm筛分两部分保存,放入干冰中保鲜用于测土壤脱氢酶、蔗糖酶、纤维素酶含量,其中脱氢酶、蔗糖酶、纤维素酶含量采用可见分光光度法测定。
最后于生长季末期在每个小区灌丛下选择有代表性的1 m×1 m的样方做植被调查,记录并测量植物的种类名称、物种个体数、物种高度,物种多样性以Shannon-Wiener多样性指数(H ' D )、Pielou均匀度指数(E )来衡量。
重要值( P i ) (1)
H ' = - ∑ i = 1 S P i l n P i (2)
D = 1 - ∑ i = 1 S P i 2 (3)
E = H ' / l n S (4)
式中:P i i 种植物占总种数的比例;S 为总种数。
1.4 数据分析
用Excel 2019对数据进行初步整理,使用SPSS 22.0软件进行重复测量方差分析(Repeated measure ANOVA)研究干旱处理、生长季及其交互作用对土壤呼吸速率的影响,采用单因素方差分析(One-way ANOVA)研究干旱处理对土壤呼吸速率和土壤水热因子、植物特征及酶活性的影响。利用Origin2022进行图表绘制和数据拟合,分别采用线性回归方程模型和指数回归模型拟合土壤呼吸与土壤含水量、土壤温度之间的关系。在R 2.0软件中进行相关性分析,评价土壤呼吸速率与土壤水热因子、植物特征及酶活性的相关性,剔除强相关的因子后采用“vegan”包进行数据的方差分解。
2 结果与分析
2.1 极端干旱对小叶锦鸡儿灌丛土壤呼吸的影响
生长季和干旱处理均对土壤呼吸速率有显著影响(P <0.01),二者的交互作用也对土壤呼吸速率有显著影响(表1 )。
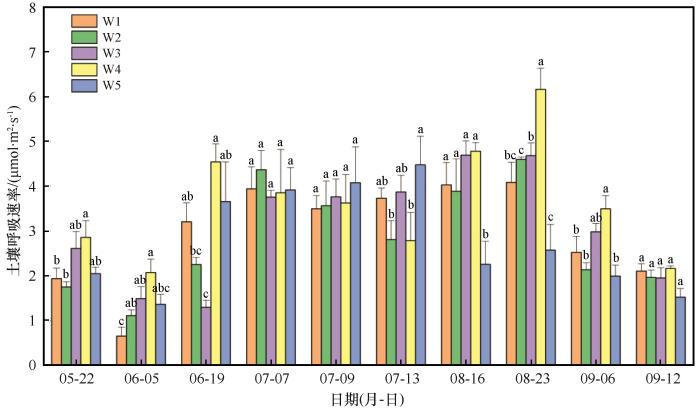
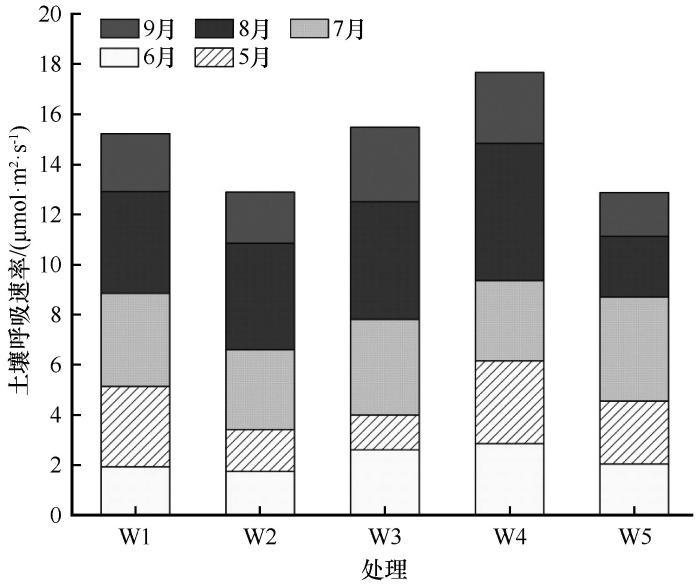
随着生长季的推移,不同干旱处理下小叶锦鸡儿灌丛土壤呼吸速率总体上均呈现先上升后下降的趋势(图2 ),最大值出现在7月下旬或8月(分别是4.08、4.59、4.69、6.16、4.47 μmol·m-2 ·s-1 ),最小值出现在6月(分别是0.65、1.11、1.29、2.07、1.36 μmol·m-2 ·s-1 ),与自然降水对照组相比,除了6月5日,减雨50%对各时期土壤呼吸速率无显著影响(P >0.05),而生长季长于1个月的持续干旱期间灌丛土壤呼吸显著降低,降幅为25.36%~61.59%。与自然降水对照组相比,6、7月干旱处理下的累积土壤呼吸分别提高了1.67%和15.99%,而8月干旱处理及减雨50%处理组的累积土壤呼吸值分别降低了15.43%和15.31%(图3 )。
图2
图2
干旱处理对小叶锦鸡儿灌丛土壤呼吸速率的影响(均值±标准误)
注: W1、W2、W3、W4、W5分别代表自然处理、减雨50%、6月干旱、7月干旱、8月干旱。不同小写字母表示同一测量时间不同干旱处理间具有显著差异(P <0.05)
Fig.2
The impact of drought treatments on soil respiration rate in Caragana microphylla
图3
图3
干旱处理对小叶锦鸡儿月累积土壤呼吸的影响
注: W1、W2、W3、W4、W5分别代表自然处理、减雨50%、6月干旱、7月干旱、8月干旱
Fig.3
The effect of drought treatment on cumulative soil respiration in Caragana microphylla
2.2 极端干旱对小叶锦鸡儿灌丛土壤水热因子、植物特征和酶活性的影响
与自然处理组(W1)相比,8月干旱处理组(W5)显著增高了土壤温度和根生物量,显著降低了蔗糖酶含量(P <0.05);减雨50%处理组(W2)显著增高了土壤温度(表2 )。
2.3 极端干旱下土壤呼吸与影响因子间的关系
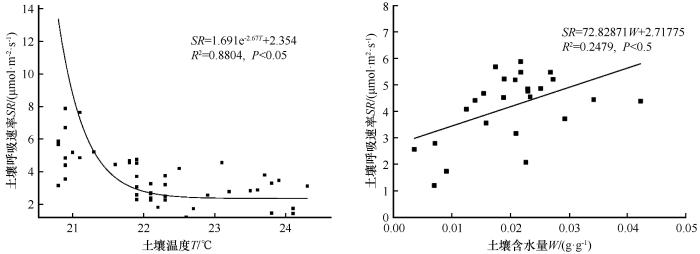
土壤呼吸速率(SR )与土壤温度(T )之间可用指数函数(SR =a +b e cT 图4 )。
图4
图4
土壤呼吸速率(SR )与土壤温度(T )和含水量(W )的拟合关系
注: W1、W2、W3、W4、W5分别代表自然处理、减雨50%、6月干旱、7月干旱、8月干旱
Fig4
The fitting relationship between soil respiration rate and soil temperature and moisture content
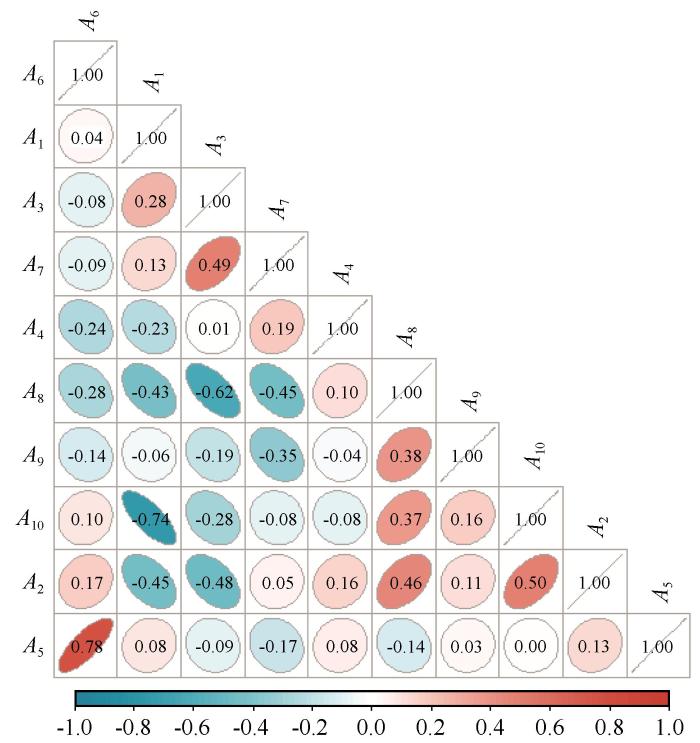
由土壤呼吸速率与土壤水热因子、植物特征和酶活性的相关性分析可知(图5 ),土壤呼吸速率与土壤温度呈显著负相关,与土壤含水量、蔗糖酶含量呈正相关,但不显著。此外,土壤温度与蔗糖酶含量、土壤含水量呈负相关,根生物量与蔗糖酶含量显著负相关,而土壤含水量与根生物量负相关。
图5
图5
土壤呼吸速率与土壤水热因子、植物特征、酶活性的相关性分析
注: A 1 ~A 10 分别代表土壤温度、土壤含水量、根生物量、Simpson优势度指数、Shannon-Wiener多样性指数、均匀度指数、脱氢酶(U·g-1 )、蔗糖酶(U·g-1 )、纤维素酶(U·g-1 )、土壤呼吸速率
Fig.5
Correlation analysis of soil respiration rate with Soil hydrothermal factors, plant characteristics, and enzyme activity
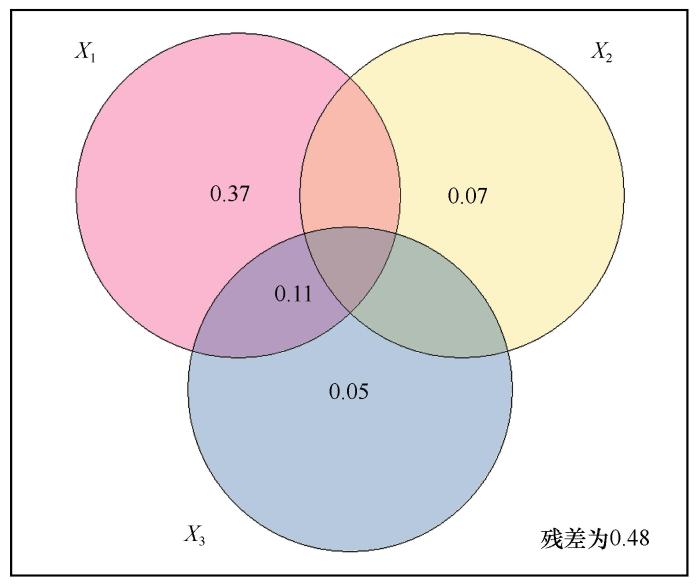
方差分解结果显示,土壤水热因子、植物特征和酶活性能够解释土壤呼吸速率变异的48%,其中土壤水热因子独立的解释量最大(37%),土壤酶特征独立的解释量最小(5%),植物特征独立的解释量为7%;其中3组环境因子中两两共同的解释量较小(图6 )。
图6
图6
土壤水热因子、植物特征和酶活性对土壤呼吸速率的方差分解
注: 单个圆圈内数字代表该环境因子组合能解释的方差。圆圈重合部分内数字代表几个环境因子组合共同解释的方差。X 1 组包括土壤温度、含水量。X 2 组包括根生物量、Shannon-Wiener多样性指数和Pielou均匀度指数。X 3 组包括土壤脱氢酶、纤维素酶和蔗糖酶含量
Fig.6
Variation partitioning of soil respiration rate by environmental factor groups
3 讨论
3.1 干旱处理下土壤呼吸的动态变化
生长季小叶锦鸡儿灌丛土壤呼吸速率在不同干旱处理下均呈现先上升后下降的趋势(图2 ),其最大值出现在7月下旬或8月,最小值出现在6月。这与以往研究一致[13 ,26 -27 ] ,这可能是因为随着生长季的推移,气温升高、降水增多,从而促进植物光合作用和生产力,加速植物生长,导致土壤中新碳比例的增加,与旧碳相关的土壤呼吸增强,且水分增多促进土壤中可溶性有机碳的有效性和流动性,促进地下部分植物生长和微生物的代谢活动[28 ] ,加快与凋落物、微生物有关碳的分解,导致土壤呼吸随生长季呈现先上升的趋势。之后气温下降,植物地上部分和微生物活性减弱,土壤呼吸速率下降[29 ] 。
先前的研究表明,土壤水分是限制干旱与半干旱地区生命活动的主要非生物因子[27 ] ,降水是土壤水分的主要来源,对地下生物化学过程起着重要的作用,降水的改变势必会引起土壤水分发生变化,进而影响土壤呼吸在内的生态系统碳循环发生变化。在本研究中,长时间持续干旱显著降低了小叶锦鸡儿的土壤呼吸速率,这与李冰等[13 ] 的研究相似,而减雨50%对各时期土壤呼吸速率无显著影响,这可能是由试验期间降水较多、试验方法以及植被类型等不同所导致,土壤呼吸对模拟降水变化的响应取决于处理的量级[30 ] 、生态系统类型[31 ] 和降水处理时间[15 ] ;另外科尔沁沙地位于干旱半干旱区,小叶锦鸡儿灌丛对于干旱具有一定的适应性。类似的,一项在荒漠草原中的研究也表明适度干旱对土壤呼吸速率影响较小[13 ] 。
干旱可抑制土壤微生物代谢活性并降低土壤呼吸作用,水分条件恢复后,其微生物生物量及活性呈现激增式响应,促使CO₂、N₂O和CH₄等含碳、氮气态产物被大量释放至大气中[32 ] 。本研究发现,生长季6、7月干旱会造成累积土壤呼吸增高,而8月干旱和减雨50%会造成累积土壤呼吸降低。值得注意的是,小叶锦鸡儿灌丛历经长于1个月的持续干旱,在次月干旱解除后会出现土壤呼吸的激发效应,且在不同时期的持续干旱间有明显差异,其中7月持续干旱的激发效应最大。进一步分析表明,这可能是由土壤呼吸的激发效应、土壤呼吸底物增加、生长季温度和降水变化规律等综合所导致。类似的,王旭等[33 ] 发现降水对土壤呼吸存在激发效应和抑制效应,且土壤呼吸季节变异性与降雨频率和平均降雨强度密切相关。
3.2 干旱处理下环境因子对土壤呼吸的影响
干旱处理下,土壤呼吸作用受生物因子和非生物因子的共同调节[34 ] ,在受水分限制的干旱半干旱区,干旱处理主要通过影响土壤含水量来改变土壤呼吸,从而影响生态系统碳排放[35 ] 。本研究发现,8月干旱处理组小叶锦鸡儿灌丛的土壤含水量显著低于自然处理组,同时其小叶锦鸡儿灌丛的土壤呼吸速率也显著低于自然处理组,但土壤呼吸速率与土壤含水量线性关系不显著。这与沈健等[36 ] 、段北星等[37 ] 的研究结果相似,是因为只要土壤水分处于正常阈值,其对土壤呼吸速率的贡献低于土壤温度。但当土壤水分一旦处于极低或极高的水平,就会替代温度成为影响土壤呼吸速率的主导因子[38 ] ,土壤水分过低时会抑制土壤根系呼吸和微生物活动,过高时会堵塞土壤孔隙,从而抑制土壤CO2 向大气中的排放[12 ] 。
先前的研究表明,变暖对土壤呼吸的影响既有正面也有负面。变暖会加速土壤有机碳的微生物分解速率,从而刺激土壤呼吸。然而,变暖引起的干旱也会限制土壤呼吸,从而导致土壤呼吸减少,并在土壤含水量低的情况下产生正的碳-气候反馈[28 ] 。在本研究中,日尺度上小叶锦鸡儿灌丛占主导地位的微生境的土壤呼吸速率在变暖条件下显著受到抑制。这一结果与陶冬雪等[10 ] 在呼伦贝尔草甸草原的研究结论一致,但与李冰等[13 ] 在荒漠草原的发现相反,推测这种差异可能源于研究时间尺度的不同。具体而言,在生长季尺度上,土壤呼吸速率随温度升高呈显著上升趋势;而在日尺度观测中,极端干旱处理过程中伴随的增温通过降低土壤含水量对植物和微生物形成水分胁迫,从而部分抵消了温度升高对土壤呼吸的正效应,最终表现为土壤呼吸速率的整体抑制[26 ] 。
本研究发现土壤水热因子、植物特征和酶活性可解释土壤呼吸变异的48%,其中土壤水热因子独立的解释量最大,这与他人研究结果相似[10 ,13 ] 。进一步分析表明,极端干旱处理可以通过影响土壤含水量和土壤温度来直接影响土壤呼吸速率,也可以通过影响根生物量、植物多样性、酶活性等间接影响土壤呼吸速率。本研究中,与自然处理组相比,8月干旱处理组显著提高了土壤温度和根生物量,降低了土壤含水量和蔗糖酶含量。这与上述分析相佐证。值得注意的是,在干旱半干旱区,水分减少会直接影响土壤酶活性,也会通过与养分循环密切相关的微生物群落组成与结构来间接影响土壤酶活性[39 ] ;另外植物倾向于在潮湿条件下向地上分配更多的碳水化合物,以利于光捕获,但在干燥条件下会优先将碳分配给根,以促进地下资源的获取[40 ] 。当植物受到干旱胁迫时,光合产物向根系分配的比例会增加,从而促进根系生长,加速对深层土壤水分的利用,提高自身在干旱环境的存活率[41 ] 。本研究未深入研究土壤呼吸组分(自养呼吸和异养呼吸)对极端干旱事件的响应,有待通过长期的定位观测对此进行深入分析。
4 结论
生长季不同水平干旱处理下小叶锦鸡儿灌丛的土壤呼吸速率均呈现先上升后下降的趋势;生长季长于1个月的持续干旱期间灌丛土壤呼吸显著降低,次月干旱解除后土壤呼吸的激发效应在不同时期的持续干旱间有明显差异,7月持续干旱的激发效应最大。土壤水热因子、植物特征和酶活性共同调控土壤呼吸(解释率为48%),其中土壤水热因子独立的解释量最大(解释率可达37%),土壤酶特征独立的解释量最小。
参考文献
View Option
[1]
李兵强 ,姚小萌 ,孟凡 ,等 黄土高原不同盖度生物结皮土壤呼吸特征及其影响因素
[J].生态学报 ,2025 ,45 (8 ):1 -11 .
[本文引用: 1]
[2]
Zhou X H Wan S Q Luo Y Q Source components and interannual variability of soil CO2 efflux under experimental warming and clipping in a grass land ecosystem
[J].Global Change Biology ,2007 ,13 (4 ): 761 -775 .
[本文引用: 1]
[3]
Phillips C L Bond-Lamberty B Desai A R et al The value of soil respiration measurements for interpreting and modeling terrestrial carbon cycling
[J].Plant and Soil ,2017 ,413 :1 -25 .
[本文引用: 1]
[4]
Solomon S Qin D H Manning M et al Climate Change 2007:The Physical Science Basis
[R].IPCC ,2007 :95 -123 .
[本文引用: 1]
[5]
王兴 ,钟泽坤 ,朱玉帆 ,等 增温和增雨对黄土丘陵区撂荒草地土壤呼吸的影响
[J].环境科学 ,2022 ,43 (3 ):1657 -1667 .
[本文引用: 1]
[6]
李丹 ,马晓明 ,雷佳 ,等 不同沟垄覆盖模式下旱作马铃薯田土壤呼吸及有机碳矿化特征
[J].环境科学 ,2024 ,45 (10 ):6038 -6049 .
[本文引用: 1]
[7]
闫宝龙 ,赵清格 ,张波 ,等 不同植被类型对土壤理化性质和土壤呼吸的影响
[J].生态环境学报 ,2017 ,26 (2 ):189 -195 .
[本文引用: 1]
[8]
黄飞 ,刘静 ,王博 ,等 灌枝覆盖对宁夏荒漠草地土壤呼吸的影响
[J].生态学报 ,2025 ,45 (7 ):1 -11 .
[本文引用: 1]
[9]
Yuan X Wang Y Wu P et al A global transition to flash droughts under climate change
[J].Science ,2023 ,380 :187 -191 .
[本文引用: 1]
[10]
陶冬雪 ,李文瑾 ,杨恬 ,等 降水变化和养分添加对呼伦贝尔草甸草原土壤呼吸的影响
[J].生态学杂志 ,2022 ,41 (3 ):465 -472 .
[本文引用: 3]
[11]
彭信浩 ,徐小芳 ,韩海荣 ,等 降雨减少对华北落叶松人工林土壤呼吸的影响
[J].生态环境学报 ,2017 ,26 (8 ):1310 -1316 .
[本文引用: 1]
[12]
蒿廉伊 ,张丽华 ,谢忠奎 ,等 降水变化对荒漠草原土壤呼吸的影响
[J].环境科学 ,2021 ,42 (9 ):4527 -4537 .
[本文引用: 2]
[13]
李冰 ,朱湾湾 ,韩翠 ,等 降水量变化下荒漠草原土壤呼吸及其影响因素
[J].植物生态学报 ,2023 ,47 (9 ):1310 -1321 .
[本文引用: 6]
[14]
赵智慧 ,陈俏艳 ,程思源 ,等 基于Meta分析的土壤呼吸对降水改变的非对称响应
[J].生态学报 ,2024 ,44 (7 ):2886 -2897 .
[本文引用: 1]
[15]
Morris K A Hornum S Crystal-Ornelas R et al Soil respiration response to simulated precipitation change depends on ecosystem type and study duration
[J].Journal of Geophysical Research:Biogeosciences ,2022 ,127 :e2022 JG006887.
[本文引用: 2]
[16]
Li Z L Qin P Dong Y S et al Response of soil respiration to water and nitrogen addition and its influencing factors:a four-year field experiment in a temperate steppe
[J].Plant and Soil ,2022 ,471 :427 -442 .
[本文引用: 1]
[17]
Escolar C Maestre F T Rey A Biocrusts modulate warming and rainfall exclusion effects on soil respiration in a semi-arid grassland
[J].Soil Biology and Biochemistry ,2015 ,80 :9 -17 .
[18]
Chen F Yan G Y Xing Y J et al Effects of N addition and precipitation reduction on soil respiration and its components in a temperate forest
[J].Agricultural and Forest Meteorology ,2019 ,271 :336 -345 .
[本文引用: 1]
[19]
李新鸽 ,朱连奇 ,朱文博 ,等 模拟降雨量变化对土壤呼吸的影响:进展与展望
[J].应用生态学报 ,2024 ,35 (9 ):2445 -2454 .
[本文引用: 1]
[20]
牛亚毅 ,刘蔚 ,董佳蕊 ,等 科尔沁沙地1961-2021年主要气象要素的变化特征:以奈曼旗为例
[J].中国沙漠 ,2023 ,43 (4 ):263 -273 .
[本文引用: 3]
[21]
李玉强 ,王旭洋 ,郑成卓 ,等 科尔沁沙地防沙治沙实践与生态可持续修复浅议
[J].中国沙漠 ,2024 ,44 (4 ):302 -314 .
[本文引用: 1]
[22]
包永志 ,段利民 ,刘廷玺 ,等 小叶锦鸡儿(Caragana microphylla )群落蒸散发模拟
[J].中国沙漠 ,2019 ,39 (4 ):177 -186 .
[本文引用: 1]
[23]
Su Y Zhao C Wang Y et al Spatiotemporal variations of precipitation in china using surface gauge observations from 1961 to 2016
[J].Atmosphere ,2020 ,11 (3 ):303 .
[本文引用: 1]
[24]
高雅玉 ,何文博 ,郝建斌 ,等 极端干旱区土壤呼吸动态特征及对水热因子的响应
[J].节水灌溉 ,2025 (1 ):1 -17 .
[本文引用: 2]
[25]
Zheng P F Wang D D Yu X X et al Effects of drought and rainfall events on soil autotrophic respiration and heterotrophic respiration
[J].Agriculture ,Ecosystems & Environment,2021 ,308 :107267 .
[本文引用: 1]
[26]
王忠武 ,宋晓辉 ,王悦骅 ,等 模拟降水对短花针茅荒漠草原土壤呼吸的影响
[J].中国草地学报 ,2020 ,42 (1 ):111 -116 .
[本文引用: 2]
[27]
Wang H H Huang W D He Y Z et al Effects of warming and precipitation reduction on soil respiration in Horqin sandy grassland,northern China
[J].CATENA ,2023 ,233 :107470 .
[本文引用: 2]
[28]
Lin X W Zhang Z H Wang S P et al Response of ecosystem respiration to warming and grazing during the growing seasons in the alpine meadow on the Tibetan Plateau
[J].Agricultural and Forest Meteorology ,2011 ,151 (7 ):792 -802 .
[本文引用: 2]
[29]
郭文章 ,井长青 ,王公鑫 ,等 天山北坡荒漠草原土壤呼吸和生态系统呼吸对降水的响应
[J].草地学报 ,2021 ,29 (9 ):2031 -2039 .
[本文引用: 1]
[30]
孟伟庆 ,莫训强 ,胡蓓蓓 ,等 模拟干湿交替对湿地土壤呼吸及有机碳含量的影响
[J].土壤通报 ,2015 ,46 (4 ):910 -915 .
[本文引用: 1]
[31]
Wang Z Mckenna T P Schellenberg M P et al Soil respiration response to alterations in precipitation and nitrogen addition in a desert steppe in northern China
[J].Science of The Total Environment ,2019 ,688 :231 -242 .
[本文引用: 1]
[32]
周祉蕴 ,王奕钧 ,杨艳丽 ,等 脉冲降水和凋落物对温性草原土壤碳矿化激发效应的影响
[J].草地学报 ,2024 ,32 (3 ):879 -888 .
[本文引用: 1]
[33]
王旭 ,闫玉春 ,闫瑞瑞 ,等 降雨对草地土壤呼吸季节变异性的影响
[J].生态学报 ,2013 ,33 (18 ):5631 -5635 .
[本文引用: 1]
[34]
杨青霄 ,田大栓 ,曾辉 ,等 降水格局改变背景下土壤呼吸变化的主要影响因素及其调控过程
[J].植物生态学报 ,2017 ,41 (12 ):1239 -1250 .
[本文引用: 1]
[35]
侯建峰 ,吕晓涛 ,王超 ,等 中国北方草地土壤呼吸的空间变异及成因
[J].应用生态学报 ,2014 ,25 (10 ):2840 -2846 .
[本文引用: 1]
[36]
沈健 ,何宗明 ,董强 ,等 轻度火烧对滨海沙地人工林土壤呼吸速率和非生物因子的影响
[J].植物生态学报 ,2023 ,47 (7 ):1032 -1042 .
[本文引用: 1]
[37]
段北星 ,满秀玲 ,宋浩 ,等 大兴安岭北部不同类型兴安落叶松林土壤呼吸及其组分特征
[J].北京林业大学学报 ,2018 ,40 (2 ):40 -50 .
[本文引用: 1]
[38]
Liu W Fox J E D Xu Z Litterfall and nutrient dynamics in a montane moist evergreen broad-leaved forest in Ailao Mountains,SW China
[J].Plant Ecology ,2003 ,164 (2 ):157 -170 .
[本文引用: 1]
[39]
霍皇煜 ,米琦 ,周天财 ,等 青藏高原高寒草地土壤酶活性海拔地带性特征
[J].生态学报 ,2024 ,44 (1 ):405 -415 .
[本文引用: 1]
[40]
Farrior C E Rodriguez-Iturbe I Dybzinski R et al Decreased water limitation under elevated CO2 amplifies potential for forest carbon sinks
[J].Proceedings of the National Academy of Sciences of the United States of America ,2015 ,112 (23 ):7213 -7218 .
[本文引用: 1]
[41]
Huang B Gao H Root physiological characteristics associated with drought resistance in tall fescue cultivars
[J].Crop Sciences ,2000 ,40 :196 -203 .
[本文引用: 1]
黄土高原不同盖度生物结皮土壤呼吸特征及其影响因素
1
2025
... 干旱半干旱地区约占全球陆地面积的41%,贡献了全球超过25%的碳储量,对陆地生态系统碳循环起重要作用[1 ] .土壤呼吸是陆地生态系统向大气输出CO2 的主要途径,占整个生态系统呼吸总量的60%~90%,是陆地生态系统碳循环的重要组成部分[2 -3 ] .2021年IPCC报告提出,与土壤呼吸有关的碳通量大约是与化石燃料燃烧相关的碳通量的10倍[4 ] ,这凸显了其在调控全球碳循环过程中的关键地位,土壤呼吸微弱的变化将会引起大气CO2 浓度的显著改变.因此,研究干旱半干旱地区土壤呼吸对环境变化的响应对准确估算陆地生态系统碳排放量至关重要. ...
Source components and interannual variability of soil CO2 efflux under experimental warming and clipping in a grass land ecosystem
1
2007
... 干旱半干旱地区约占全球陆地面积的41%,贡献了全球超过25%的碳储量,对陆地生态系统碳循环起重要作用[1 ] .土壤呼吸是陆地生态系统向大气输出CO2 的主要途径,占整个生态系统呼吸总量的60%~90%,是陆地生态系统碳循环的重要组成部分[2 -3 ] .2021年IPCC报告提出,与土壤呼吸有关的碳通量大约是与化石燃料燃烧相关的碳通量的10倍[4 ] ,这凸显了其在调控全球碳循环过程中的关键地位,土壤呼吸微弱的变化将会引起大气CO2 浓度的显著改变.因此,研究干旱半干旱地区土壤呼吸对环境变化的响应对准确估算陆地生态系统碳排放量至关重要. ...
The value of soil respiration measurements for interpreting and modeling terrestrial carbon cycling
1
2017
... 干旱半干旱地区约占全球陆地面积的41%,贡献了全球超过25%的碳储量,对陆地生态系统碳循环起重要作用[1 ] .土壤呼吸是陆地生态系统向大气输出CO2 的主要途径,占整个生态系统呼吸总量的60%~90%,是陆地生态系统碳循环的重要组成部分[2 -3 ] .2021年IPCC报告提出,与土壤呼吸有关的碳通量大约是与化石燃料燃烧相关的碳通量的10倍[4 ] ,这凸显了其在调控全球碳循环过程中的关键地位,土壤呼吸微弱的变化将会引起大气CO2 浓度的显著改变.因此,研究干旱半干旱地区土壤呼吸对环境变化的响应对准确估算陆地生态系统碳排放量至关重要. ...
Climate Change 2007:The Physical Science Basis
1
2007
... 干旱半干旱地区约占全球陆地面积的41%,贡献了全球超过25%的碳储量,对陆地生态系统碳循环起重要作用[1 ] .土壤呼吸是陆地生态系统向大气输出CO2 的主要途径,占整个生态系统呼吸总量的60%~90%,是陆地生态系统碳循环的重要组成部分[2 -3 ] .2021年IPCC报告提出,与土壤呼吸有关的碳通量大约是与化石燃料燃烧相关的碳通量的10倍[4 ] ,这凸显了其在调控全球碳循环过程中的关键地位,土壤呼吸微弱的变化将会引起大气CO2 浓度的显著改变.因此,研究干旱半干旱地区土壤呼吸对环境变化的响应对准确估算陆地生态系统碳排放量至关重要. ...
增温和增雨对黄土丘陵区撂荒草地土壤呼吸的影响
1
2022
... 土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用.在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] .环境因子的这种变化必然会对土壤呼吸过程产生深刻影响.陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平.值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] .Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] .土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] .尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏.科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区.1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] .在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种.目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落.而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差.因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要. ...
不同沟垄覆盖模式下旱作马铃薯田土壤呼吸及有机碳矿化特征
1
2024
... 土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用.在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] .环境因子的这种变化必然会对土壤呼吸过程产生深刻影响.陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平.值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] .Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] .土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] .尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏.科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区.1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] .在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种.目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落.而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差.因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要. ...
不同植被类型对土壤理化性质和土壤呼吸的影响
1
2017
... 土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用.在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] .环境因子的这种变化必然会对土壤呼吸过程产生深刻影响.陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平.值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] .Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] .土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] .尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏.科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区.1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] .在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种.目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落.而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差.因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要. ...
灌枝覆盖对宁夏荒漠草地土壤呼吸的影响
1
2025
... 土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用.在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] .环境因子的这种变化必然会对土壤呼吸过程产生深刻影响.陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平.值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] .Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] .土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] .尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏.科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区.1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] .在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种.目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落.而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差.因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要. ...
A global transition to flash droughts under climate change
1
2023
... 土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用.在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] .环境因子的这种变化必然会对土壤呼吸过程产生深刻影响.陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平.值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] .Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] .土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] .尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏.科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区.1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] .在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种.目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落.而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差.因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要. ...
降水变化和养分添加对呼伦贝尔草甸草原土壤呼吸的影响
3
2022
... 土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用.在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] .环境因子的这种变化必然会对土壤呼吸过程产生深刻影响.陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平.值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] .Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] .土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] .尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏.科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区.1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] .在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种.目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落.而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差.因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要. ...
... 先前的研究表明,变暖对土壤呼吸的影响既有正面也有负面.变暖会加速土壤有机碳的微生物分解速率,从而刺激土壤呼吸.然而,变暖引起的干旱也会限制土壤呼吸,从而导致土壤呼吸减少,并在土壤含水量低的情况下产生正的碳-气候反馈[28 ] .在本研究中,日尺度上小叶锦鸡儿灌丛占主导地位的微生境的土壤呼吸速率在变暖条件下显著受到抑制.这一结果与陶冬雪等[10 ] 在呼伦贝尔草甸草原的研究结论一致,但与李冰等[13 ] 在荒漠草原的发现相反,推测这种差异可能源于研究时间尺度的不同.具体而言,在生长季尺度上,土壤呼吸速率随温度升高呈显著上升趋势;而在日尺度观测中,极端干旱处理过程中伴随的增温通过降低土壤含水量对植物和微生物形成水分胁迫,从而部分抵消了温度升高对土壤呼吸的正效应,最终表现为土壤呼吸速率的整体抑制[26 ] . ...
... 本研究发现土壤水热因子、植物特征和酶活性可解释土壤呼吸变异的48%,其中土壤水热因子独立的解释量最大,这与他人研究结果相似[10 ,13 ] .进一步分析表明,极端干旱处理可以通过影响土壤含水量和土壤温度来直接影响土壤呼吸速率,也可以通过影响根生物量、植物多样性、酶活性等间接影响土壤呼吸速率.本研究中,与自然处理组相比,8月干旱处理组显著提高了土壤温度和根生物量,降低了土壤含水量和蔗糖酶含量.这与上述分析相佐证.值得注意的是,在干旱半干旱区,水分减少会直接影响土壤酶活性,也会通过与养分循环密切相关的微生物群落组成与结构来间接影响土壤酶活性[39 ] ;另外植物倾向于在潮湿条件下向地上分配更多的碳水化合物,以利于光捕获,但在干燥条件下会优先将碳分配给根,以促进地下资源的获取[40 ] .当植物受到干旱胁迫时,光合产物向根系分配的比例会增加,从而促进根系生长,加速对深层土壤水分的利用,提高自身在干旱环境的存活率[41 ] .本研究未深入研究土壤呼吸组分(自养呼吸和异养呼吸)对极端干旱事件的响应,有待通过长期的定位观测对此进行深入分析. ...
降雨减少对华北落叶松人工林土壤呼吸的影响
1
2017
... 土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用.在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] .环境因子的这种变化必然会对土壤呼吸过程产生深刻影响.陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平.值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] .Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] .土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] .尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏.科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区.1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] .在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种.目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落.而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差.因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要. ...
降水变化对荒漠草原土壤呼吸的影响
2
2021
... 土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用.在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] .环境因子的这种变化必然会对土壤呼吸过程产生深刻影响.陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平.值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] .Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] .土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] .尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏.科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区.1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] .在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种.目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落.而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差.因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要. ...
... 干旱处理下,土壤呼吸作用受生物因子和非生物因子的共同调节[34 ] ,在受水分限制的干旱半干旱区,干旱处理主要通过影响土壤含水量来改变土壤呼吸,从而影响生态系统碳排放[35 ] .本研究发现,8月干旱处理组小叶锦鸡儿灌丛的土壤含水量显著低于自然处理组,同时其小叶锦鸡儿灌丛的土壤呼吸速率也显著低于自然处理组,但土壤呼吸速率与土壤含水量线性关系不显著.这与沈健等[36 ] 、段北星等[37 ] 的研究结果相似,是因为只要土壤水分处于正常阈值,其对土壤呼吸速率的贡献低于土壤温度.但当土壤水分一旦处于极低或极高的水平,就会替代温度成为影响土壤呼吸速率的主导因子[38 ] ,土壤水分过低时会抑制土壤根系呼吸和微生物活动,过高时会堵塞土壤孔隙,从而抑制土壤CO2 向大气中的排放[12 ] . ...
降水量变化下荒漠草原土壤呼吸及其影响因素
6
2023
... 土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用.在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] .环境因子的这种变化必然会对土壤呼吸过程产生深刻影响.陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平.值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] .Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] .土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] .尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏.科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区.1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] .在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种.目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落.而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差.因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要. ...
... 生长季小叶锦鸡儿灌丛土壤呼吸速率在不同干旱处理下均呈现先上升后下降的趋势(图2 ),其最大值出现在7月下旬或8月,最小值出现在6月.这与以往研究一致[13 ,26 -27 ] ,这可能是因为随着生长季的推移,气温升高、降水增多,从而促进植物光合作用和生产力,加速植物生长,导致土壤中新碳比例的增加,与旧碳相关的土壤呼吸增强,且水分增多促进土壤中可溶性有机碳的有效性和流动性,促进地下部分植物生长和微生物的代谢活动[28 ] ,加快与凋落物、微生物有关碳的分解,导致土壤呼吸随生长季呈现先上升的趋势.之后气温下降,植物地上部分和微生物活性减弱,土壤呼吸速率下降[29 ] . ...
... 先前的研究表明,土壤水分是限制干旱与半干旱地区生命活动的主要非生物因子[27 ] ,降水是土壤水分的主要来源,对地下生物化学过程起着重要的作用,降水的改变势必会引起土壤水分发生变化,进而影响土壤呼吸在内的生态系统碳循环发生变化.在本研究中,长时间持续干旱显著降低了小叶锦鸡儿的土壤呼吸速率,这与李冰等[13 ] 的研究相似,而减雨50%对各时期土壤呼吸速率无显著影响,这可能是由试验期间降水较多、试验方法以及植被类型等不同所导致,土壤呼吸对模拟降水变化的响应取决于处理的量级[30 ] 、生态系统类型[31 ] 和降水处理时间[15 ] ;另外科尔沁沙地位于干旱半干旱区,小叶锦鸡儿灌丛对于干旱具有一定的适应性.类似的,一项在荒漠草原中的研究也表明适度干旱对土壤呼吸速率影响较小[13 ] . ...
... [13 ]. ...
... 先前的研究表明,变暖对土壤呼吸的影响既有正面也有负面.变暖会加速土壤有机碳的微生物分解速率,从而刺激土壤呼吸.然而,变暖引起的干旱也会限制土壤呼吸,从而导致土壤呼吸减少,并在土壤含水量低的情况下产生正的碳-气候反馈[28 ] .在本研究中,日尺度上小叶锦鸡儿灌丛占主导地位的微生境的土壤呼吸速率在变暖条件下显著受到抑制.这一结果与陶冬雪等[10 ] 在呼伦贝尔草甸草原的研究结论一致,但与李冰等[13 ] 在荒漠草原的发现相反,推测这种差异可能源于研究时间尺度的不同.具体而言,在生长季尺度上,土壤呼吸速率随温度升高呈显著上升趋势;而在日尺度观测中,极端干旱处理过程中伴随的增温通过降低土壤含水量对植物和微生物形成水分胁迫,从而部分抵消了温度升高对土壤呼吸的正效应,最终表现为土壤呼吸速率的整体抑制[26 ] . ...
... 本研究发现土壤水热因子、植物特征和酶活性可解释土壤呼吸变异的48%,其中土壤水热因子独立的解释量最大,这与他人研究结果相似[10 ,13 ] .进一步分析表明,极端干旱处理可以通过影响土壤含水量和土壤温度来直接影响土壤呼吸速率,也可以通过影响根生物量、植物多样性、酶活性等间接影响土壤呼吸速率.本研究中,与自然处理组相比,8月干旱处理组显著提高了土壤温度和根生物量,降低了土壤含水量和蔗糖酶含量.这与上述分析相佐证.值得注意的是,在干旱半干旱区,水分减少会直接影响土壤酶活性,也会通过与养分循环密切相关的微生物群落组成与结构来间接影响土壤酶活性[39 ] ;另外植物倾向于在潮湿条件下向地上分配更多的碳水化合物,以利于光捕获,但在干燥条件下会优先将碳分配给根,以促进地下资源的获取[40 ] .当植物受到干旱胁迫时,光合产物向根系分配的比例会增加,从而促进根系生长,加速对深层土壤水分的利用,提高自身在干旱环境的存活率[41 ] .本研究未深入研究土壤呼吸组分(自养呼吸和异养呼吸)对极端干旱事件的响应,有待通过长期的定位观测对此进行深入分析. ...
基于Meta分析的土壤呼吸对降水改变的非对称响应
1
2024
... 土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用.在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] .环境因子的这种变化必然会对土壤呼吸过程产生深刻影响.陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平.值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] .Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] .土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] .尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏.科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区.1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] .在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种.目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落.而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差.因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要. ...
Soil respiration response to simulated precipitation change depends on ecosystem type and study duration
2
2022
... 土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用.在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] .环境因子的这种变化必然会对土壤呼吸过程产生深刻影响.陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平.值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] .Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] .土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] .尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏.科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区.1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] .在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种.目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落.而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差.因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要. ...
... 先前的研究表明,土壤水分是限制干旱与半干旱地区生命活动的主要非生物因子[27 ] ,降水是土壤水分的主要来源,对地下生物化学过程起着重要的作用,降水的改变势必会引起土壤水分发生变化,进而影响土壤呼吸在内的生态系统碳循环发生变化.在本研究中,长时间持续干旱显著降低了小叶锦鸡儿的土壤呼吸速率,这与李冰等[13 ] 的研究相似,而减雨50%对各时期土壤呼吸速率无显著影响,这可能是由试验期间降水较多、试验方法以及植被类型等不同所导致,土壤呼吸对模拟降水变化的响应取决于处理的量级[30 ] 、生态系统类型[31 ] 和降水处理时间[15 ] ;另外科尔沁沙地位于干旱半干旱区,小叶锦鸡儿灌丛对于干旱具有一定的适应性.类似的,一项在荒漠草原中的研究也表明适度干旱对土壤呼吸速率影响较小[13 ] . ...
Response of soil respiration to water and nitrogen addition and its influencing factors:a four-year field experiment in a temperate steppe
1
2022
... 土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用.在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] .环境因子的这种变化必然会对土壤呼吸过程产生深刻影响.陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平.值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] .Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] .土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] .尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏.科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区.1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] .在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种.目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落.而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差.因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要. ...
Biocrusts modulate warming and rainfall exclusion effects on soil respiration in a semi-arid grassland
0
2015
Effects of N addition and precipitation reduction on soil respiration and its components in a temperate forest
1
2019
... 土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用.在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] .环境因子的这种变化必然会对土壤呼吸过程产生深刻影响.陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平.值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] .Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] .土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] .尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏.科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区.1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] .在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种.目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落.而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差.因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要. ...
模拟降雨量变化对土壤呼吸的影响:进展与展望
1
2024
... 土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用.在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] .环境因子的这种变化必然会对土壤呼吸过程产生深刻影响.陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平.值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] .Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] .土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] .尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏.科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区.1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] .在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种.目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落.而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差.因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要. ...
科尔沁沙地1961-2021年主要气象要素的变化特征:以奈曼旗为例
3
2023
... 土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用.在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] .环境因子的这种变化必然会对土壤呼吸过程产生深刻影响.陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平.值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] .Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] .土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] .尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏.科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区.1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] .在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种.目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落.而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差.因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要. ...
... 研究区位于中国科尔沁沙地腹地(42°56′N,120°45′E),海拔约360 m.该地区属于温带大陆性半干旱气候,春季干燥多风,夏季炎热多雨,秋季凉爽短暂,冬季寒冷漫长[20 ] .年降水量360 mm,集中在6—8月且分布不均(图1 ),年潜在蒸发量1 500~2 500 mm.年均气温6.4 ℃,≥10 ℃积温3 000~3 400 ℃.主要土壤类型为风沙土,土壤全碳和全氮含量分别为0.019、 0.252 g·kg-1 ,饱和含水量0.234 g·g-1 ,土壤容重为1.518 g·cm-3 ,土壤pH为7.1,电导率是30.44 μS·cm-1 .植物群落结构组成简单,以灌丛和草本为主,优势植物种有小叶锦鸡儿、猪毛蒿(Artemisia Scoparia )、糙隐子草(Cleistogenes squarrosa )、狗尾草(Setaria viridis )、尖头叶藜(Cheno podium acuminatum )等. ...
... 本试验选取统一种植年限的小叶锦鸡儿灌丛典型样地,建立围栏以开展长期控制试验.科尔沁沙地近10年降水分布呈现出显著的年际波动性和年内分配不均的特征[20 ,23 ] ,且科尔沁沙地生态系统的草本或灌木在生长季的不同月份处于不同的关键物候阶段(如返青、快速生长、开花、结实)和生理敏感期(如分蘖、花芽分化),该区域生长季(6—8月)降水是生态系统水分供应的绝对关键期,但其总量和月度分布存在高度不确定性.基于此,共设置了5种极端干旱处理,分别为自然对照组(W1)、减雨50%处理组(W2)、6月干旱处理组(W3)、7月干旱处理组(W4)和8月干旱处理组(W5).试验采用完全随机设计,每种处理设置5个重复,共计25个试验小区.每个小区面积为7 m×7 m,小区间隔约10 m,以避免不同处理间土壤水分的相互干扰.减雨50%处理组(W2)通过在试验小区上方搭建间隔式遮雨棚(宽度50 cm、间隔50 cm)实现,遮雨棚采用透光率为90%的聚丙烯塑料板材构建,板材之间预留与板材宽度相同的空隙,以确保减少50%的自然降水量.干旱处理组(W3、W4、W5)同样在试验小区上方搭建全覆盖遮雨棚,并使用透光率为90%的塑料棚完全覆盖棚顶和两侧,两头保持通风状态.其中,6月干旱处理组仅在6月实施干旱处理,其余月份与自然对照组保持一致;7月干旱处理组和8月干旱处理组分别于7月和8月实施干旱处理,其余月份亦与自然对照组一致. ...
科尔沁沙地防沙治沙实践与生态可持续修复浅议
1
2024
... 土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用.在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] .环境因子的这种变化必然会对土壤呼吸过程产生深刻影响.陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平.值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] .Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] .土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] .尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏.科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区.1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] .在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种.目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落.而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差.因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要. ...
小叶锦鸡儿(Caragana microphylla )群落蒸散发模拟
1
2019
... 土壤呼吸对环境变化敏感,受土壤水分[5 ] 、温度[6 ] 、植物特征[7 ] 、酶活性[8 ] 等多种因素的共同调控,且多因素间存在复杂的协同作用.在全球气候变暖的背景下,极端干旱事件发生频率显著增加,其中干旱事件的强度、频次和持续时间均呈现加剧趋势[9 ] .环境因子的这种变化必然会对土壤呼吸过程产生深刻影响.陶冬雪等[10 ] 在呼伦贝尔草甸草原的定位观测表明,干旱可显著降低草原土壤呼吸强度;而彭信浩等[11 ] 在华北落叶松人工林开展的降水控制试验结果显示,减少降水虽使土壤呼吸速率呈下降趋势,但未达到显著水平.值得注意的是,尽管极端干旱事件普遍导致不同生态系统土壤呼吸速率降低[12 -13 ] ,但比较研究表明,草地生态系统土壤呼吸对干旱的敏感性显著高于森林生态系统,极端干旱事件普遍会降低不同生态系统的土壤呼吸速率,但草地土壤呼吸对极端干旱的响应大于森林[14 ] .Meta分析表明,土壤呼吸对极端干旱的响应取决于生态系统类型和研究持续时间,降水充足的生态系统对土壤呼吸的响应随着时间的变化表现出适应性,而降水较少的生态系统随着时间的推移影响加剧[15 ] .土壤呼吸对极端干旱的响应与生态系统水分状况密切相关[16 -18 ] ,具体而言,极端干旱通过改变土壤水分有效性,直接影响土壤微生物活动和根系呼吸;同时,干旱还通过调控植物生长、土壤有机碳动态及微生物群落结构,间接影响土壤呼吸[19 ] .尽管在森林、草地已有多项关于土壤呼吸对极端干旱事件响应的研究,但在干旱半干旱地区,特别是科尔沁沙地,此类研究仍然匮乏.科尔沁沙地位于西辽河平原向蒙古高原过渡地带,地处温带季风气候边缘区,属于典型的干旱半干旱地区.1961—2021年的气候观测数据表明,该区域正经历显著的气温升高、降水减少及极端降水事件增加过程[20 ] ,这种变化进一步加剧了生态脆弱性,使其成为中国沙漠化最为严重的区域,亦是国家防沙治沙的重点区域[21 ] .在荒漠化治理实践中,以小叶锦鸡儿(Caragana microphylla )为代表的固沙灌木因其卓越的抗逆性(耐寒、耐旱、耐沙埋)被广泛引种.目前该物种分布面积已达510万hm²[22 ] ,成为区域优势灌木群落.而针对沙地生态系统特别是灌木主导系统的土壤呼吸对极端降水响应的研究不足,这种关键生态类型的缺失使区域碳通量评估存在系统性偏差.因此,研究科尔沁沙地小叶锦鸡儿灌丛土壤呼吸对极端干旱的响应对准确估算干旱半干旱地区陆地生态系统碳排放量至关重要. ...
Spatiotemporal variations of precipitation in china using surface gauge observations from 1961 to 2016
1
2020
... 本试验选取统一种植年限的小叶锦鸡儿灌丛典型样地,建立围栏以开展长期控制试验.科尔沁沙地近10年降水分布呈现出显著的年际波动性和年内分配不均的特征[20 ,23 ] ,且科尔沁沙地生态系统的草本或灌木在生长季的不同月份处于不同的关键物候阶段(如返青、快速生长、开花、结实)和生理敏感期(如分蘖、花芽分化),该区域生长季(6—8月)降水是生态系统水分供应的绝对关键期,但其总量和月度分布存在高度不确定性.基于此,共设置了5种极端干旱处理,分别为自然对照组(W1)、减雨50%处理组(W2)、6月干旱处理组(W3)、7月干旱处理组(W4)和8月干旱处理组(W5).试验采用完全随机设计,每种处理设置5个重复,共计25个试验小区.每个小区面积为7 m×7 m,小区间隔约10 m,以避免不同处理间土壤水分的相互干扰.减雨50%处理组(W2)通过在试验小区上方搭建间隔式遮雨棚(宽度50 cm、间隔50 cm)实现,遮雨棚采用透光率为90%的聚丙烯塑料板材构建,板材之间预留与板材宽度相同的空隙,以确保减少50%的自然降水量.干旱处理组(W3、W4、W5)同样在试验小区上方搭建全覆盖遮雨棚,并使用透光率为90%的塑料棚完全覆盖棚顶和两侧,两头保持通风状态.其中,6月干旱处理组仅在6月实施干旱处理,其余月份与自然对照组保持一致;7月干旱处理组和8月干旱处理组分别于7月和8月实施干旱处理,其余月份亦与自然对照组一致. ...
极端干旱区土壤呼吸动态特征及对水热因子的响应
2
2025
... 土壤呼吸速率采用Li-6400X便携式光合作用测定仪(LI-COR.,美国)测定,2024年4月上旬于每个处理小区内在小叶锦鸡儿灌丛下布置1个土壤呼吸测定所用的PVC材质的呼吸环(高度10 cm,直径20 cm),每个土壤呼吸环露出地表高度3 cm,共布置25个土壤呼吸环,安装时尽可能避免地面植被的破坏,整个试验期间呼吸环位置、高度不变,以实现土壤呼吸速率的长期定点监测.生长季6—9月每个月定期测定土壤呼吸速率2~3次,08:00—11:00土壤呼吸速率变化斜率最小,且此时测量值最接近日均值[24 ] ,故每次测量均选择晴朗无风的08:00—11:00测定.土壤呼吸是通过在90 s间隔内腔室顶部空间CO2 的增加来测量的,因此,在每次测量前1天,用剪刀齐地面剪去土壤呼吸环内的植物的地上部分,以消除地上植物部分对土壤呼吸速率的影响[24 -25 ] . ...
... [24 -25 ]. ...
Effects of drought and rainfall events on soil autotrophic respiration and heterotrophic respiration
1
2021
... 土壤呼吸速率采用Li-6400X便携式光合作用测定仪(LI-COR.,美国)测定,2024年4月上旬于每个处理小区内在小叶锦鸡儿灌丛下布置1个土壤呼吸测定所用的PVC材质的呼吸环(高度10 cm,直径20 cm),每个土壤呼吸环露出地表高度3 cm,共布置25个土壤呼吸环,安装时尽可能避免地面植被的破坏,整个试验期间呼吸环位置、高度不变,以实现土壤呼吸速率的长期定点监测.生长季6—9月每个月定期测定土壤呼吸速率2~3次,08:00—11:00土壤呼吸速率变化斜率最小,且此时测量值最接近日均值[24 ] ,故每次测量均选择晴朗无风的08:00—11:00测定.土壤呼吸是通过在90 s间隔内腔室顶部空间CO2 的增加来测量的,因此,在每次测量前1天,用剪刀齐地面剪去土壤呼吸环内的植物的地上部分,以消除地上植物部分对土壤呼吸速率的影响[24 -25 ] . ...
模拟降水对短花针茅荒漠草原土壤呼吸的影响
2
2020
... 生长季小叶锦鸡儿灌丛土壤呼吸速率在不同干旱处理下均呈现先上升后下降的趋势(图2 ),其最大值出现在7月下旬或8月,最小值出现在6月.这与以往研究一致[13 ,26 -27 ] ,这可能是因为随着生长季的推移,气温升高、降水增多,从而促进植物光合作用和生产力,加速植物生长,导致土壤中新碳比例的增加,与旧碳相关的土壤呼吸增强,且水分增多促进土壤中可溶性有机碳的有效性和流动性,促进地下部分植物生长和微生物的代谢活动[28 ] ,加快与凋落物、微生物有关碳的分解,导致土壤呼吸随生长季呈现先上升的趋势.之后气温下降,植物地上部分和微生物活性减弱,土壤呼吸速率下降[29 ] . ...
... 先前的研究表明,变暖对土壤呼吸的影响既有正面也有负面.变暖会加速土壤有机碳的微生物分解速率,从而刺激土壤呼吸.然而,变暖引起的干旱也会限制土壤呼吸,从而导致土壤呼吸减少,并在土壤含水量低的情况下产生正的碳-气候反馈[28 ] .在本研究中,日尺度上小叶锦鸡儿灌丛占主导地位的微生境的土壤呼吸速率在变暖条件下显著受到抑制.这一结果与陶冬雪等[10 ] 在呼伦贝尔草甸草原的研究结论一致,但与李冰等[13 ] 在荒漠草原的发现相反,推测这种差异可能源于研究时间尺度的不同.具体而言,在生长季尺度上,土壤呼吸速率随温度升高呈显著上升趋势;而在日尺度观测中,极端干旱处理过程中伴随的增温通过降低土壤含水量对植物和微生物形成水分胁迫,从而部分抵消了温度升高对土壤呼吸的正效应,最终表现为土壤呼吸速率的整体抑制[26 ] . ...
Effects of warming and precipitation reduction on soil respiration in Horqin sandy grassland,northern China
2
2023
... 生长季小叶锦鸡儿灌丛土壤呼吸速率在不同干旱处理下均呈现先上升后下降的趋势(图2 ),其最大值出现在7月下旬或8月,最小值出现在6月.这与以往研究一致[13 ,26 -27 ] ,这可能是因为随着生长季的推移,气温升高、降水增多,从而促进植物光合作用和生产力,加速植物生长,导致土壤中新碳比例的增加,与旧碳相关的土壤呼吸增强,且水分增多促进土壤中可溶性有机碳的有效性和流动性,促进地下部分植物生长和微生物的代谢活动[28 ] ,加快与凋落物、微生物有关碳的分解,导致土壤呼吸随生长季呈现先上升的趋势.之后气温下降,植物地上部分和微生物活性减弱,土壤呼吸速率下降[29 ] . ...
... 先前的研究表明,土壤水分是限制干旱与半干旱地区生命活动的主要非生物因子[27 ] ,降水是土壤水分的主要来源,对地下生物化学过程起着重要的作用,降水的改变势必会引起土壤水分发生变化,进而影响土壤呼吸在内的生态系统碳循环发生变化.在本研究中,长时间持续干旱显著降低了小叶锦鸡儿的土壤呼吸速率,这与李冰等[13 ] 的研究相似,而减雨50%对各时期土壤呼吸速率无显著影响,这可能是由试验期间降水较多、试验方法以及植被类型等不同所导致,土壤呼吸对模拟降水变化的响应取决于处理的量级[30 ] 、生态系统类型[31 ] 和降水处理时间[15 ] ;另外科尔沁沙地位于干旱半干旱区,小叶锦鸡儿灌丛对于干旱具有一定的适应性.类似的,一项在荒漠草原中的研究也表明适度干旱对土壤呼吸速率影响较小[13 ] . ...
Response of ecosystem respiration to warming and grazing during the growing seasons in the alpine meadow on the Tibetan Plateau
2
2011
... 生长季小叶锦鸡儿灌丛土壤呼吸速率在不同干旱处理下均呈现先上升后下降的趋势(图2 ),其最大值出现在7月下旬或8月,最小值出现在6月.这与以往研究一致[13 ,26 -27 ] ,这可能是因为随着生长季的推移,气温升高、降水增多,从而促进植物光合作用和生产力,加速植物生长,导致土壤中新碳比例的增加,与旧碳相关的土壤呼吸增强,且水分增多促进土壤中可溶性有机碳的有效性和流动性,促进地下部分植物生长和微生物的代谢活动[28 ] ,加快与凋落物、微生物有关碳的分解,导致土壤呼吸随生长季呈现先上升的趋势.之后气温下降,植物地上部分和微生物活性减弱,土壤呼吸速率下降[29 ] . ...
... 先前的研究表明,变暖对土壤呼吸的影响既有正面也有负面.变暖会加速土壤有机碳的微生物分解速率,从而刺激土壤呼吸.然而,变暖引起的干旱也会限制土壤呼吸,从而导致土壤呼吸减少,并在土壤含水量低的情况下产生正的碳-气候反馈[28 ] .在本研究中,日尺度上小叶锦鸡儿灌丛占主导地位的微生境的土壤呼吸速率在变暖条件下显著受到抑制.这一结果与陶冬雪等[10 ] 在呼伦贝尔草甸草原的研究结论一致,但与李冰等[13 ] 在荒漠草原的发现相反,推测这种差异可能源于研究时间尺度的不同.具体而言,在生长季尺度上,土壤呼吸速率随温度升高呈显著上升趋势;而在日尺度观测中,极端干旱处理过程中伴随的增温通过降低土壤含水量对植物和微生物形成水分胁迫,从而部分抵消了温度升高对土壤呼吸的正效应,最终表现为土壤呼吸速率的整体抑制[26 ] . ...
天山北坡荒漠草原土壤呼吸和生态系统呼吸对降水的响应
1
2021
... 生长季小叶锦鸡儿灌丛土壤呼吸速率在不同干旱处理下均呈现先上升后下降的趋势(图2 ),其最大值出现在7月下旬或8月,最小值出现在6月.这与以往研究一致[13 ,26 -27 ] ,这可能是因为随着生长季的推移,气温升高、降水增多,从而促进植物光合作用和生产力,加速植物生长,导致土壤中新碳比例的增加,与旧碳相关的土壤呼吸增强,且水分增多促进土壤中可溶性有机碳的有效性和流动性,促进地下部分植物生长和微生物的代谢活动[28 ] ,加快与凋落物、微生物有关碳的分解,导致土壤呼吸随生长季呈现先上升的趋势.之后气温下降,植物地上部分和微生物活性减弱,土壤呼吸速率下降[29 ] . ...
模拟干湿交替对湿地土壤呼吸及有机碳含量的影响
1
2015
... 先前的研究表明,土壤水分是限制干旱与半干旱地区生命活动的主要非生物因子[27 ] ,降水是土壤水分的主要来源,对地下生物化学过程起着重要的作用,降水的改变势必会引起土壤水分发生变化,进而影响土壤呼吸在内的生态系统碳循环发生变化.在本研究中,长时间持续干旱显著降低了小叶锦鸡儿的土壤呼吸速率,这与李冰等[13 ] 的研究相似,而减雨50%对各时期土壤呼吸速率无显著影响,这可能是由试验期间降水较多、试验方法以及植被类型等不同所导致,土壤呼吸对模拟降水变化的响应取决于处理的量级[30 ] 、生态系统类型[31 ] 和降水处理时间[15 ] ;另外科尔沁沙地位于干旱半干旱区,小叶锦鸡儿灌丛对于干旱具有一定的适应性.类似的,一项在荒漠草原中的研究也表明适度干旱对土壤呼吸速率影响较小[13 ] . ...
Soil respiration response to alterations in precipitation and nitrogen addition in a desert steppe in northern China
1
2019
... 先前的研究表明,土壤水分是限制干旱与半干旱地区生命活动的主要非生物因子[27 ] ,降水是土壤水分的主要来源,对地下生物化学过程起着重要的作用,降水的改变势必会引起土壤水分发生变化,进而影响土壤呼吸在内的生态系统碳循环发生变化.在本研究中,长时间持续干旱显著降低了小叶锦鸡儿的土壤呼吸速率,这与李冰等[13 ] 的研究相似,而减雨50%对各时期土壤呼吸速率无显著影响,这可能是由试验期间降水较多、试验方法以及植被类型等不同所导致,土壤呼吸对模拟降水变化的响应取决于处理的量级[30 ] 、生态系统类型[31 ] 和降水处理时间[15 ] ;另外科尔沁沙地位于干旱半干旱区,小叶锦鸡儿灌丛对于干旱具有一定的适应性.类似的,一项在荒漠草原中的研究也表明适度干旱对土壤呼吸速率影响较小[13 ] . ...
脉冲降水和凋落物对温性草原土壤碳矿化激发效应的影响
1
2024
... 干旱可抑制土壤微生物代谢活性并降低土壤呼吸作用,水分条件恢复后,其微生物生物量及活性呈现激增式响应,促使CO₂、N₂O和CH₄等含碳、氮气态产物被大量释放至大气中[32 ] .本研究发现,生长季6、7月干旱会造成累积土壤呼吸增高,而8月干旱和减雨50%会造成累积土壤呼吸降低.值得注意的是,小叶锦鸡儿灌丛历经长于1个月的持续干旱,在次月干旱解除后会出现土壤呼吸的激发效应,且在不同时期的持续干旱间有明显差异,其中7月持续干旱的激发效应最大.进一步分析表明,这可能是由土壤呼吸的激发效应、土壤呼吸底物增加、生长季温度和降水变化规律等综合所导致.类似的,王旭等[33 ] 发现降水对土壤呼吸存在激发效应和抑制效应,且土壤呼吸季节变异性与降雨频率和平均降雨强度密切相关. ...
降雨对草地土壤呼吸季节变异性的影响
1
2013
... 干旱可抑制土壤微生物代谢活性并降低土壤呼吸作用,水分条件恢复后,其微生物生物量及活性呈现激增式响应,促使CO₂、N₂O和CH₄等含碳、氮气态产物被大量释放至大气中[32 ] .本研究发现,生长季6、7月干旱会造成累积土壤呼吸增高,而8月干旱和减雨50%会造成累积土壤呼吸降低.值得注意的是,小叶锦鸡儿灌丛历经长于1个月的持续干旱,在次月干旱解除后会出现土壤呼吸的激发效应,且在不同时期的持续干旱间有明显差异,其中7月持续干旱的激发效应最大.进一步分析表明,这可能是由土壤呼吸的激发效应、土壤呼吸底物增加、生长季温度和降水变化规律等综合所导致.类似的,王旭等[33 ] 发现降水对土壤呼吸存在激发效应和抑制效应,且土壤呼吸季节变异性与降雨频率和平均降雨强度密切相关. ...
降水格局改变背景下土壤呼吸变化的主要影响因素及其调控过程
1
2017
... 干旱处理下,土壤呼吸作用受生物因子和非生物因子的共同调节[34 ] ,在受水分限制的干旱半干旱区,干旱处理主要通过影响土壤含水量来改变土壤呼吸,从而影响生态系统碳排放[35 ] .本研究发现,8月干旱处理组小叶锦鸡儿灌丛的土壤含水量显著低于自然处理组,同时其小叶锦鸡儿灌丛的土壤呼吸速率也显著低于自然处理组,但土壤呼吸速率与土壤含水量线性关系不显著.这与沈健等[36 ] 、段北星等[37 ] 的研究结果相似,是因为只要土壤水分处于正常阈值,其对土壤呼吸速率的贡献低于土壤温度.但当土壤水分一旦处于极低或极高的水平,就会替代温度成为影响土壤呼吸速率的主导因子[38 ] ,土壤水分过低时会抑制土壤根系呼吸和微生物活动,过高时会堵塞土壤孔隙,从而抑制土壤CO2 向大气中的排放[12 ] . ...
中国北方草地土壤呼吸的空间变异及成因
1
2014
... 干旱处理下,土壤呼吸作用受生物因子和非生物因子的共同调节[34 ] ,在受水分限制的干旱半干旱区,干旱处理主要通过影响土壤含水量来改变土壤呼吸,从而影响生态系统碳排放[35 ] .本研究发现,8月干旱处理组小叶锦鸡儿灌丛的土壤含水量显著低于自然处理组,同时其小叶锦鸡儿灌丛的土壤呼吸速率也显著低于自然处理组,但土壤呼吸速率与土壤含水量线性关系不显著.这与沈健等[36 ] 、段北星等[37 ] 的研究结果相似,是因为只要土壤水分处于正常阈值,其对土壤呼吸速率的贡献低于土壤温度.但当土壤水分一旦处于极低或极高的水平,就会替代温度成为影响土壤呼吸速率的主导因子[38 ] ,土壤水分过低时会抑制土壤根系呼吸和微生物活动,过高时会堵塞土壤孔隙,从而抑制土壤CO2 向大气中的排放[12 ] . ...
轻度火烧对滨海沙地人工林土壤呼吸速率和非生物因子的影响
1
2023
... 干旱处理下,土壤呼吸作用受生物因子和非生物因子的共同调节[34 ] ,在受水分限制的干旱半干旱区,干旱处理主要通过影响土壤含水量来改变土壤呼吸,从而影响生态系统碳排放[35 ] .本研究发现,8月干旱处理组小叶锦鸡儿灌丛的土壤含水量显著低于自然处理组,同时其小叶锦鸡儿灌丛的土壤呼吸速率也显著低于自然处理组,但土壤呼吸速率与土壤含水量线性关系不显著.这与沈健等[36 ] 、段北星等[37 ] 的研究结果相似,是因为只要土壤水分处于正常阈值,其对土壤呼吸速率的贡献低于土壤温度.但当土壤水分一旦处于极低或极高的水平,就会替代温度成为影响土壤呼吸速率的主导因子[38 ] ,土壤水分过低时会抑制土壤根系呼吸和微生物活动,过高时会堵塞土壤孔隙,从而抑制土壤CO2 向大气中的排放[12 ] . ...
大兴安岭北部不同类型兴安落叶松林土壤呼吸及其组分特征
1
2018
... 干旱处理下,土壤呼吸作用受生物因子和非生物因子的共同调节[34 ] ,在受水分限制的干旱半干旱区,干旱处理主要通过影响土壤含水量来改变土壤呼吸,从而影响生态系统碳排放[35 ] .本研究发现,8月干旱处理组小叶锦鸡儿灌丛的土壤含水量显著低于自然处理组,同时其小叶锦鸡儿灌丛的土壤呼吸速率也显著低于自然处理组,但土壤呼吸速率与土壤含水量线性关系不显著.这与沈健等[36 ] 、段北星等[37 ] 的研究结果相似,是因为只要土壤水分处于正常阈值,其对土壤呼吸速率的贡献低于土壤温度.但当土壤水分一旦处于极低或极高的水平,就会替代温度成为影响土壤呼吸速率的主导因子[38 ] ,土壤水分过低时会抑制土壤根系呼吸和微生物活动,过高时会堵塞土壤孔隙,从而抑制土壤CO2 向大气中的排放[12 ] . ...
Litterfall and nutrient dynamics in a montane moist evergreen broad-leaved forest in Ailao Mountains,SW China
1
2003
... 干旱处理下,土壤呼吸作用受生物因子和非生物因子的共同调节[34 ] ,在受水分限制的干旱半干旱区,干旱处理主要通过影响土壤含水量来改变土壤呼吸,从而影响生态系统碳排放[35 ] .本研究发现,8月干旱处理组小叶锦鸡儿灌丛的土壤含水量显著低于自然处理组,同时其小叶锦鸡儿灌丛的土壤呼吸速率也显著低于自然处理组,但土壤呼吸速率与土壤含水量线性关系不显著.这与沈健等[36 ] 、段北星等[37 ] 的研究结果相似,是因为只要土壤水分处于正常阈值,其对土壤呼吸速率的贡献低于土壤温度.但当土壤水分一旦处于极低或极高的水平,就会替代温度成为影响土壤呼吸速率的主导因子[38 ] ,土壤水分过低时会抑制土壤根系呼吸和微生物活动,过高时会堵塞土壤孔隙,从而抑制土壤CO2 向大气中的排放[12 ] . ...
青藏高原高寒草地土壤酶活性海拔地带性特征
1
2024
... 本研究发现土壤水热因子、植物特征和酶活性可解释土壤呼吸变异的48%,其中土壤水热因子独立的解释量最大,这与他人研究结果相似[10 ,13 ] .进一步分析表明,极端干旱处理可以通过影响土壤含水量和土壤温度来直接影响土壤呼吸速率,也可以通过影响根生物量、植物多样性、酶活性等间接影响土壤呼吸速率.本研究中,与自然处理组相比,8月干旱处理组显著提高了土壤温度和根生物量,降低了土壤含水量和蔗糖酶含量.这与上述分析相佐证.值得注意的是,在干旱半干旱区,水分减少会直接影响土壤酶活性,也会通过与养分循环密切相关的微生物群落组成与结构来间接影响土壤酶活性[39 ] ;另外植物倾向于在潮湿条件下向地上分配更多的碳水化合物,以利于光捕获,但在干燥条件下会优先将碳分配给根,以促进地下资源的获取[40 ] .当植物受到干旱胁迫时,光合产物向根系分配的比例会增加,从而促进根系生长,加速对深层土壤水分的利用,提高自身在干旱环境的存活率[41 ] .本研究未深入研究土壤呼吸组分(自养呼吸和异养呼吸)对极端干旱事件的响应,有待通过长期的定位观测对此进行深入分析. ...
Decreased water limitation under elevated CO2 amplifies potential for forest carbon sinks
1
2015
... 本研究发现土壤水热因子、植物特征和酶活性可解释土壤呼吸变异的48%,其中土壤水热因子独立的解释量最大,这与他人研究结果相似[10 ,13 ] .进一步分析表明,极端干旱处理可以通过影响土壤含水量和土壤温度来直接影响土壤呼吸速率,也可以通过影响根生物量、植物多样性、酶活性等间接影响土壤呼吸速率.本研究中,与自然处理组相比,8月干旱处理组显著提高了土壤温度和根生物量,降低了土壤含水量和蔗糖酶含量.这与上述分析相佐证.值得注意的是,在干旱半干旱区,水分减少会直接影响土壤酶活性,也会通过与养分循环密切相关的微生物群落组成与结构来间接影响土壤酶活性[39 ] ;另外植物倾向于在潮湿条件下向地上分配更多的碳水化合物,以利于光捕获,但在干燥条件下会优先将碳分配给根,以促进地下资源的获取[40 ] .当植物受到干旱胁迫时,光合产物向根系分配的比例会增加,从而促进根系生长,加速对深层土壤水分的利用,提高自身在干旱环境的存活率[41 ] .本研究未深入研究土壤呼吸组分(自养呼吸和异养呼吸)对极端干旱事件的响应,有待通过长期的定位观测对此进行深入分析. ...
Root physiological characteristics associated with drought resistance in tall fescue cultivars
1
2000
... 本研究发现土壤水热因子、植物特征和酶活性可解释土壤呼吸变异的48%,其中土壤水热因子独立的解释量最大,这与他人研究结果相似[10 ,13 ] .进一步分析表明,极端干旱处理可以通过影响土壤含水量和土壤温度来直接影响土壤呼吸速率,也可以通过影响根生物量、植物多样性、酶活性等间接影响土壤呼吸速率.本研究中,与自然处理组相比,8月干旱处理组显著提高了土壤温度和根生物量,降低了土壤含水量和蔗糖酶含量.这与上述分析相佐证.值得注意的是,在干旱半干旱区,水分减少会直接影响土壤酶活性,也会通过与养分循环密切相关的微生物群落组成与结构来间接影响土壤酶活性[39 ] ;另外植物倾向于在潮湿条件下向地上分配更多的碳水化合物,以利于光捕获,但在干燥条件下会优先将碳分配给根,以促进地下资源的获取[40 ] .当植物受到干旱胁迫时,光合产物向根系分配的比例会增加,从而促进根系生长,加速对深层土壤水分的利用,提高自身在干旱环境的存活率[41 ] .本研究未深入研究土壤呼吸组分(自养呼吸和异养呼吸)对极端干旱事件的响应,有待通过长期的定位观测对此进行深入分析. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}