沙质草地3 种植物群落土壤有机碳氮密度特征
武雅琳 , 1 , 2 , 4 , 赵学勇 1 , 2 , 3 , 张蕊 , 1 , 2 , 3 , 4
1.中国科学院西北生态环境资源研究院,干旱区生态安全与可持续发展全国重点实验室,甘肃 兰州 730000
2.中国科学院西北生态环境资源研究院,奈曼沙漠化研究站,甘肃 兰州 730000
3.中国科学院西北生态环境资源研究院,乌拉特荒漠草原研究站,甘肃 兰州 730000
4.中国科学院大学,北京 100049
Characteristics of soil organic carbon and nitrogen density in three plant communities of sandy grasslands
Wu Yalin , 1 , 2 , 4 , Zhao Xueyong 1 , 2 , 3 , Zhang Rui , 1 , 2 , 3 , 4
1.State Key Laboratory of Ecological Safety and Sustainable Development in Arid Lands /, Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
2.Naiman Desertification Research Station /, Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
3.Urat Desert-Grassland Research Station, Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
4.University of Chinese Academy of Sciences,Beijing 100049,China
通讯作者: 张蕊(E-mail: zhangrui@lzb.ac.cn )
收稿日期: 2025-05-11
修回日期: 2025-07-01
基金资助:
国家自然科学基金项目 . 42207538
Received: 2025-05-11
Revised: 2025-07-01
摘要
土壤有机碳和全氮是土壤系统功能实现的核心要素,探讨植物群落对土壤有机碳和全氮的影响有利于深入理解科尔沁沙地土壤碳氮密度的变化规律。本研究以科尔沁沙地黄蒿+禾本科(Artemisia scoparia +Gramineae,ASP)、小叶锦鸡儿+杂类草(Caragana microphylla +Forbs,CMF)、沙米+狗尾草(Agriophyllum squarrosum +Setaria viridis ,ASGB)为主的3种植物群落为研究对象,分析不同植物群落的土壤碳氮密度特征及其影响因素。结果表明:(1)0~100 cm土壤有机碳(SOC)、全氮(TN)、全磷(TP)依次为ASP群落>CMF群落>ASGB群落。SOC、TN随土壤深度的增加而减少,TP、TK在各土层深度均保持稳定。(2)ASP群落土壤C∶P、C∶K、N∶P、N∶K、P∶K值最大,ASGB群落20~40 cm土壤C∶N显著高于ASP群落。土壤C∶P、C∶K、N∶K、P∶K随土层深度的增加呈下降趋势。(3)0~10、10~20、20~40 cm土壤有机碳密度(SCD)和土壤氮密度(SND)依次为ASP群落>CMF群落>ASGB群落。3种植物群落SCD随土壤深度的增加先降低后升高,而ASP群落SND表现出相反趋势。(4)土壤C∶N和黏粒对SCD影响的贡献率分别为15.5%和11.7%;土壤N∶P、N∶K和黏粒对SND影响的贡献率分别为23.7%、22.3%和8.4%。
关键词:
科尔沁沙地 土壤养分 化学计量 土壤氮密度 有机碳密度
Abstract
Soil organic carbon (SOC) and nitrogen (N) are core elements for the functioning of soil systems. Investigating the effects of different plant communities and soil depths on SOC and total nitrogen (TN) is essential for understanding the variation patterns of soil carbon and nitrogen density in the Horqin Sandy Land. This study chose three plant communities in the Horqin Sandy Land: dominant species of Artemisia scoparia + Gramineae (ASP), Caragana microphylla + Forbs (CMF), and Agriophyllum squarrosum + Setaria viridis (ASGB) to investigate the characteristics of soil carbon and nitrogen density and their influencing factors. The results showed that: (1) In the 0-100 cm soil layer, SOC, TN, and total phosphorus (TP) followed the order: ASP community > CMF community > ASGB community. SOC and TN decreased with increasing soil depth, while TP and total potassium (TK) remained relatively stable. (2) The ASP community exhibited the highest soil stoichiometric ratios (C∶P, C∶K, N∶P, N∶K, P∶K), whereas the ASGB community showed significantly higher C∶N in the 20-40 cm layer compared to ASP. Soil C∶P, C∶K, N∶K, and P∶K ratios generally decreased with depth. (3) Soil organic carbon density (SCD) and soil nitrogen density (SND) in the 0-10, 10-20 cm and 20-40 cm layers followed the order: ASP > CMF > ASGB. SCD in all three communities initially decreased and then increased with depth, while the ASP community displayed an opposite trend for SND. (4) The contribution rates of soil C∶N and clay to SCD were 15.5% and 11.7%, respectively; the contribution rates of soil N∶P, N∶K and clay to SND were 23.7%,22.3% and 8.4%, respectively.
Keywords:
Horqin Sandy Land soil nutrients stoichiometry soil nitrogen density organic carbon density
本文引用格式
武雅琳, 赵学勇, 张蕊. 沙质草地3 种植物群落土壤有机碳氮密度特征 . 中国沙漠 [J], 2025, 45(4): 357-367 doi:10.7522/j.issn.1000-694X.2025.00079
Wu Yalin, Zhao Xueyong, Zhang Rui. Characteristics of soil organic carbon and nitrogen density in three plant communities of sandy grasslands . Journal of Desert Research
0 引言
近年来,全球气候变化加剧,气温升高、降水格局改变及极端气候事件频发,沙质草地生态系统面临着前所未有的压力和挑战。气候变化直接影响植物生长发育、群落组成和分布格局,进而间接改变土壤有机碳氮循环过程和存储格局。此外,人类活动如过度放牧、开垦、城市化扩张等,也在不断干扰和破坏沙质草地生态系统原有结构和功能,进一步加剧土壤有机碳氮的流失从而导致生态系统退化[1 ] 。土壤有机碳、全氮是陆地生态系统中至关重要的组成部分[2 ] ,不仅在土壤肥力、植物生长和生态系统生产力方面发挥关键作用,还参与全球碳氮循环过程,对气候变化反馈调节机制有重要作用。
沙质草地生态系统结构和功能具有独特性和脆弱性,对于维持区域生态平衡和生物多样性保护具有不可忽视的意义[3 ] 。沙质草地生态系统植被覆盖度相对较低,土壤质地疏松,保水保肥能力较差,使得其土壤有机碳氮含量与动态变化相较于其他生态系统更为敏感,受环境因素和人类活动影响更为显著[4 ] 。研究沙质草地不同植物群落有机碳氮密度特征,对植被与土壤恢复具有重要参考意义[5 ] 。
由于气候变化和过度放牧等影响,科尔沁沙地沙质草地植被退化,土地沙化严重。随着围栏封育等固沙措施的实施,该区出现了不同物种组成的群落[6 ] 。随着封育年限增加,群落植被盖度和物种丰富度明显增加[7 -8 ] ,土壤种子库也随之改变[9 ] 。科尔沁沙地沙质草地的研究主要集中在植物群落对不同环境的响应[10 -11 ] 、退化沙质草地恢复过程中生物多样性变化[12 -15 ] 、土壤化学计量研究[16 -17 ] 、土壤营养元素随草地沙化程度的变化[18 ] 、沙质草地恢复过程中土壤微生物群落变化[19 ] 等方面,但鲜有关于沙质草地不同植物群落土壤有机碳和全氮密度特征的研究。
深入研究沙质草地不同植物群落土壤有机碳和全氮密度特征及其与土壤化学计量之间的内在联系,对于全面揭示沙质草地不同植物群落的土壤碳氮固存能力、准确评估其碳汇潜力、有效预测沙地土壤系统对气候变化和人类活动的响应机制具有重要的理论意义。同时可为沙质草地生态系统的合理利用和可持续发展提供关键科学依据,以期维护沙质草地生态系统的稳定性和实现区域生态环境健康发展。
1 材料与方法
1.1 试验区概况
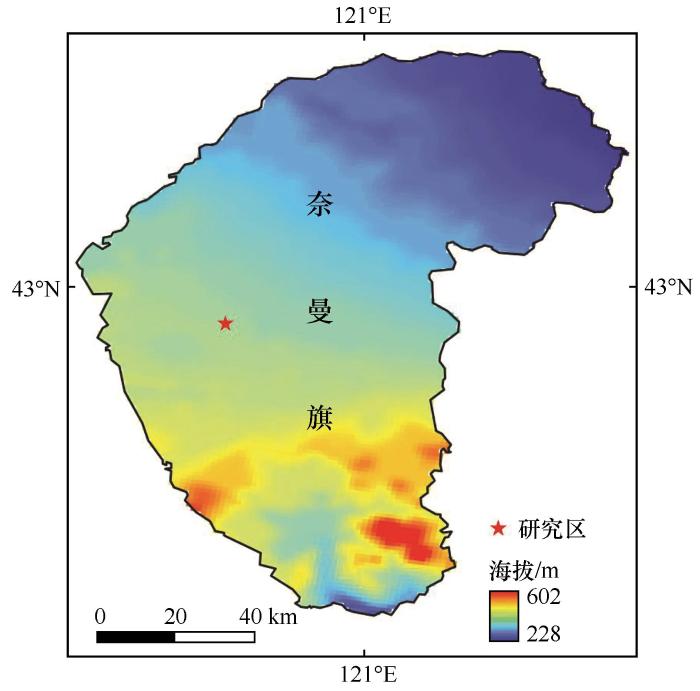
研究区域(42°41′N、120°55′E)位于中国北方半干旱区科尔沁沙地,该区属于温带大陆性半干旱季风气候,具有春季干燥多风、夏季炎热多雨、秋季凉爽短暂、冬季寒冷漫长的特点。研究区海拔358 m,多年平均气温约为6.4 ℃,多年平均降水量为360 mm,多年平均潜在蒸发量为2 000 mm[20 ] ,干旱程度较高。土壤主要为风沙土、沙质栗钙土和草甸土。植被组成以狗尾草(Setaria viridis )、黄蒿(Artemisia scoparia )、盐蒿(Suaeda salsa )、沙米(Agriophyllum squarrosum )、白草(Pennisetum flaccidum )、扁蓿豆(Medicago ruthenica )、尖头叶藜(Chenopodium acuminatum )、小叶锦鸡儿(Caragana microphylla )等为主。研究区位置如图1 所示。
图1
图1
研究区位置
注: 该图基于自然资源部标准地图服务网站标准地图(审图号GS(2024)0650号)制作,底图边界无修改
Fig.1
Location of the study area
1.2 试验设计
试验分别选择以黄蒿+禾本科(Artemisia scoparia +Gramineae,ASP)、小叶锦鸡儿+杂类草(Caragana microphylla +Forbs,CMF)和以沙米+狗尾草(Agriophyllum squarrosum +Setaria viridis ,ASGB)为主的3种植物群落为研究对象。于2020年8月中旬在3种群落设置3个1 m×1 m的样方,各样方之间距离20 m以上。每个样方中土壤样品分5层(0~10、10~20、20~40、40~60、60~100 cm)采集土壤样品。用直径2.5 cm的土钻在每个样方内按“S”形分层采集土样,同一土层深度样品混合为一个样品,剔除可见根系和砾石,共计45个土壤样品。采集的土样在避光、阴凉、通风处自然风干至恒重,研磨并过筛后测量土壤有机碳(SOC)、全氮(TN)、全磷(TP)和全钾(TK)含量。
1.3 测定指标与方法
土壤容重(BD)采用环刀法测定。土壤机械组成测定采用干筛法,共分为三级:砂粒(2~0.05 mm)、粉粒(0.05~0.002 mm)和黏粒(<0.002 mm)。土壤有机碳含量测定采用重铬酸钾氧化-外加热法;土壤全氮含量的测定采用凯氏定氮法;土壤磷含量测定采用HClO4 -H2 SO4 消煮-钼锑抗比色法。
1.4 数据分析
S C D = S O C × B D × H × 10 (1)
S N D = T N × B D × H × 10 (2)
式中:SCD 为土壤有机碳密度(g·m-2 );SND 为土壤氮密度(g·m-2 );BD 为土壤容重(g·cm-3 );H 为土层厚度(cm);SOC 为土壤有机碳含量(g·kg-1 );TN 为土壤全氮含量(g·kg-1 );10为换算系数[21 ] 。
文中数据使用Excel 2019软件整理,使用SPSS软件(27.0.1)、Origin2021和R软件(4.4.1)进行冗余分析以及广义线性混合模型[22 -23 ] 建立和绘图。不同植物群落和不同土层土壤容重、土壤机械组成、土壤碳氮磷钾含量以及土壤碳氮密度采用单因素方差分析法进行差异性比较。显著性检验采用邓肯法,显著水平为0.05。
2 结果与分析
2.1 土壤容重和机械组成
ASP群落BD在不同土壤深度垂直梯度变化为1.28~1.50 g·cm-3 ,变异系数为5.74%,最大值出现在10~20 cm土层;CMF群落BD垂直梯度变化为1.51~1.54 g·cm-3 ,变异系数为0.80%,最大值出现在60~100 cm土层;ASGB群落BD垂直梯度变化为1.51~1.61 g·cm-3 ,40~60 cm BD较0~10 cm增加了6.6%(表1 )。
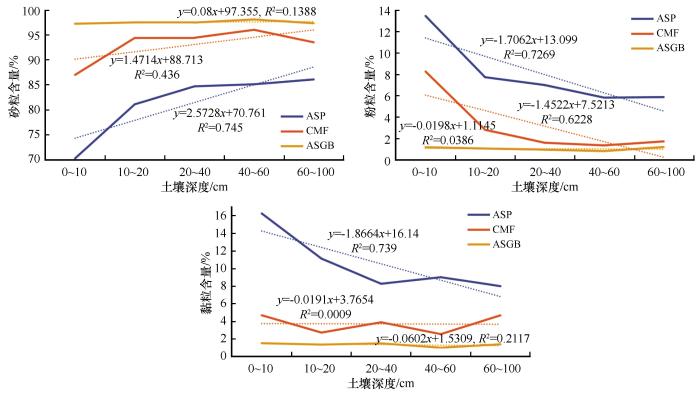
土壤砂粒含量依次为ASGB群落>CMF群落>ASP群落;土壤黏粉粒含量依次为ASP群落>CMF群落>ASGB群落(图2 )。ASP群落的砂粒含量随深度增加而增加,而CMF群落的最大值出现在40~60 cm土层。ASP群落的黏粉粒和CMF群落粉粒含量随深度增加而减少。不同植物群落土壤砂粒与土壤深度极显著正相关(r =0.808,P <0.001),粉粒、黏粒与土壤深度极显著负相关(r =-0.7,P <0.001;r =-0.841,P <0.001)。
图2
图2
3种植物群落土壤机械组成特征
注: ASP代表黄蒿+禾本科群落;CMF代表小叶锦鸡儿+杂类草群落;ASGB代表沙米+狗尾草群落
Fig.2
Mechanical composition characteristics of soil in three plant communities
2.2 土壤有机碳与全量养分
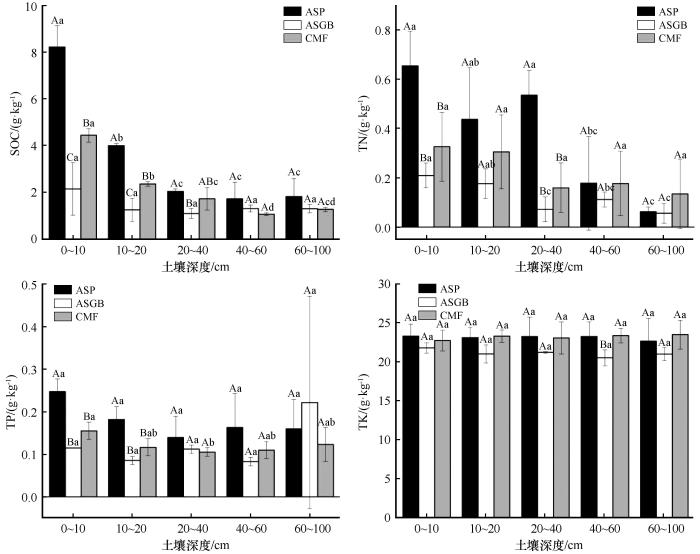
0~10、10~20 cm SOC、TN、TP和TK含量依次为ASP群落>CMF群落>ASGB群落(图3 )。随土壤深度增加,ASP群落SOC含量显著减少,从0~10 cm到60~100 cm降幅达78.5%,其中0~40 cm土层占总量的80.23%;TN含量呈现降—升—降模式,10~20 cm较0~10 cm降低33.2%,在40~60 cm和60~100 cm显著降低,0~40 cm累计占比87.2%;TP含量随土壤深度增加呈现先减小后增加再减小的趋势;TK含量无显著差异。CMF群落SOC含量从0~10 cm到60~100 cm降低71.8%,88.4%分布在0~60 cm;TN含量从0~20 cm到20~40 cm降低51.1%,0~60 cm占比87.8%;TP含量随土壤深度增加呈降低趋势。ASGB群落SOC含量从0~10 cm到60~100 cm降低39.8%,81.7%分布在0~60 cm;TN含量呈波动降低趋势,从0~10 cm到10~20 cm降低15.9%,20~40 cm显著减少后40~60 cm短暂回升,60~100 cm较0~10 cm降低73.9%,0~60 cm占比91.4%。
图3
图3
3种植物群落不同土层土壤养分差异
注: ASP代表黄蒿+禾本科群落;ASGB代表沙米+狗尾草群落;CMF代表小叶锦鸡儿+杂类草群落。大写字母表示同一土层不同植物群落间差异显著,小写字母表示同一植物群落不同土层间差异显著(P <0.05)
Fig.3
Nutrient differences of soil in different soil layers of three plant communities
2.3 土壤化学计量特征
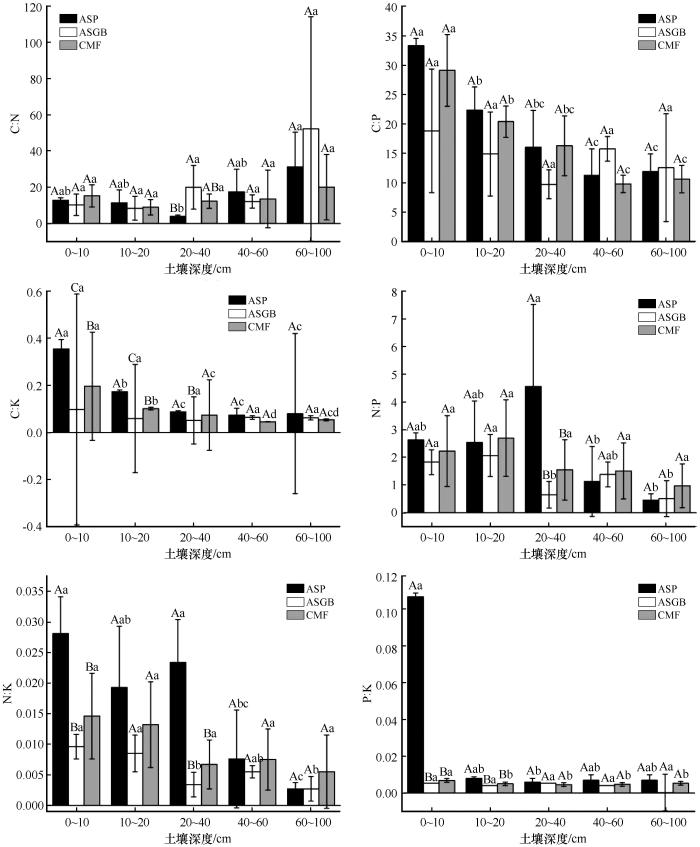
沙质草地不同植物群落土壤C∶N随着土壤深度增加先降低后升高,20~40 cm土层3种植物群落间差异显著,ASGB群落>CMF群落>ASP群落。N∶P随着土壤深度的增加先升高后降低,且在ASP群落20~40 cm有最大值。C∶P、C∶K、N∶K和P∶K随着土壤深度的增加而降低,在3种植物群落不同土层之间差异显著,依次为ASP群落>CMF群落>ASGB群落,且最大值均出现在ASP群落0~10 cm土层(图4 )。
图4
图4
3种植物群落土壤化学计量比特征
注: ASP代表黄蒿+禾本科群落;ASGB代表沙米+狗尾草群落;CMF代表小叶锦鸡儿+杂类草群落。大写字母表示同一土层不同植物群落间差异显著,小写字母表示同一植物群落不同土层间差异显著(P <0.05)
Fig.4
Chemical metrology characteristics of soil in three plant communities
2.4 土壤有机碳氮密度
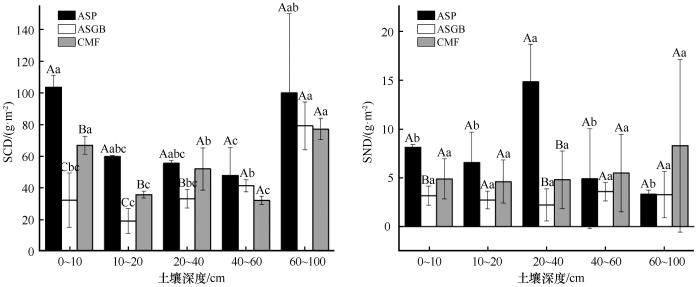
SCD最大值出现在ASP群落的0~10 cm(103.66 g·m-2 );最小值出现在ASGB群落的10~20 cm(19.24 g·m-2 )。SND最大值出现在ASP群落的20~40 cm(14.85 g·m-2 );最小值出现在ASGB群落的20~40 cm(2.23 g·m-2 ,图5 )。
图5
图5
3种植物群落土壤碳氮密度
注: ASP代表黄蒿+禾本科群落;ASGB代表沙米+狗尾草群落;CMF代表小叶锦鸡儿+杂类草群落。大写字母表示同一土层不同植物群落间差异显著,小写字母表示同一植物群落不同土层间差异显著(P <0.05)
Fig.5
Soil carbon and nitrogen density of three plant communities
ASP群落SCD为48~103.66 g·m-2 ,0~100 cm SCD的平均值为73.48 g·m-2 ,10~20、20~40、40~60、60~100 cm SCD较0~10 cm的降低42.2%、49.2%、53.7%,3.4%;SND变化范围为3.33~14.85 g·m-2 ,0~100 cm SND平均值为7.56 g·m-2 ,10~20、40~60、60~100 cm SND较0~10 cm的降低18.9%、39.5%、59.1%,而20~40 cm SND较0~10 cm的升高82.8%。
CMF群落SCD为32.3~77.29 g·m-2 ,0~100 cm SCD的平均值为52.92 g·m-2 ,10~20、20~40、40~60 cm SCD较0~10 cm的降低46.6%、22.3%、51.8%,而60~100 cm SCD较0~10 cm的升高15.3%;SND为4.62~8.3 g·m-2 ,0~100 cm SND平均值为5.63 g·m-2 ,10~20、20~40 cm SND较0~10 cm的降低5.7%、1.7%,而40~60、60~100 cm SND较0~10 cm的升高12.1%、69.5%。
ASGB群落SCD为19.24~79.32 g·m-2 ,0~100 cm SCD平均值为41.16 g·m-2 ,10~20 cm SCD较0~10 cm的降低14.1%,而20~40、40~60、60~100 cm SCD较0~10 cm的升高2.8%、28.2%、144.8%;SND变化范围为2.23~3.6 g·m-2 ,0~100 cm SND平均值为3 g·m-2 ,10~20、20~40 cm SND较0~10 cm的降低14.1%、29.9%,而40~60、60~100 cm SND较0~10 cm的升高13.1%、2.9%。
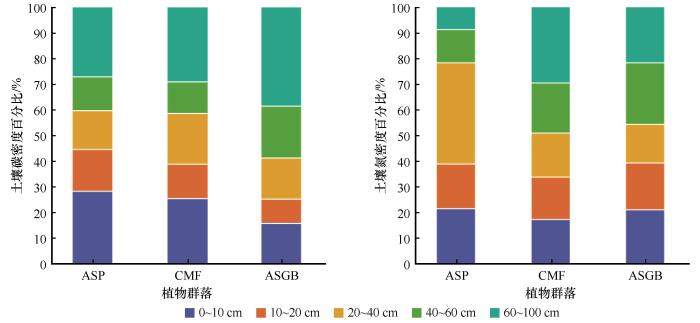
ASP群落0~10 cm SCD占比最高(28.2%),其次是60~100 cm(27.3%),10~20、20~40 cm和40~60 cm的占比较少。CMF群落60~100 cm SCD占比最高(29.2%),其次是0~10 cm(25.3%),20~40、10~20 cm和40~60 cm的占比较少。ASGB群落60~100 cm SCD占比最高(38.5%),40~60、20~40、0~10 cm和10~20 cm的占比逐渐减少(图6 )。
图6
图6
3种植物群落土壤碳氮密度垂直分布特征
注: ASP代表黄蒿+禾本科群落;CMF代表小叶锦鸡儿+杂类草群落;ASGB代表沙米+狗尾草群落
Fig.6
Vertical distribution characteristics of soil carbon and nitrogen density in three plant communities
ASP群落20~40 cm SND占比最高(39.3%),0~10、10~20、40~60 cm和60~100 cm的占比逐渐减少。CMF群落60~100 cm SND占比最高(29.5%),其他4个土层占比相当。ASGB群落40~60 cm SND占比最高(24%),60~100、0~10、10~20 cm和20~40 cm的占比逐渐减少(图6 )。
2.5 土壤要素的相关性
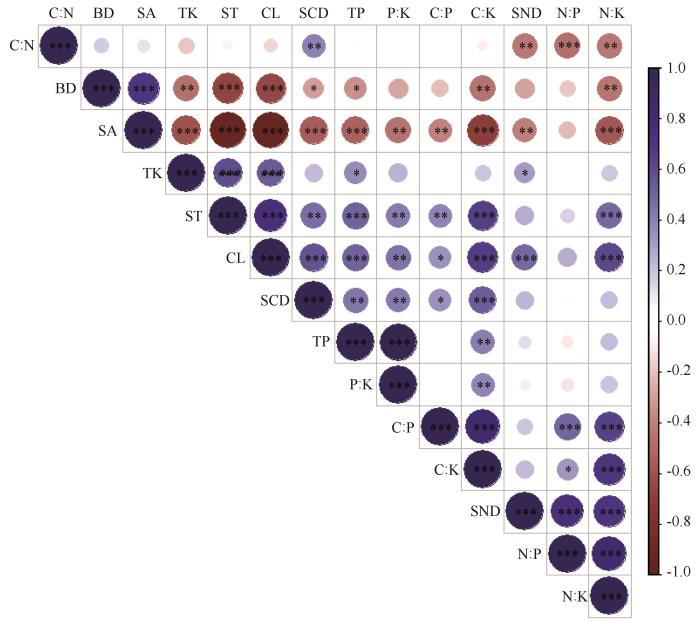
3种植物群落BD与砂粒含量极显著正相关(P <0.001),与粉粒和黏粒极显著负相关(P <0.001),砂粒含量与C∶K、N∶K极显著负相关(P <0.01),而粉粒和黏粒含量与C∶K、N∶K极显著正相关(P <0.001)。TP与P∶K极显著正相关(P <0.001),C∶P与C∶K和N∶K极显著正相关(P <0.001,图7 )。SCD和土壤砂粒极显著负相关(P <0.001),与粉粒和黏粒含量极显著正相关(P <0.01),表明SCD随土壤黏粉粒含量的增加而增加,随砂粒含量增加而减少。SCD与C∶N和C∶K极显著正相关(P <0.05)。SCD与BD显著负相关(P <0.05),表明随BD的升高,SCD降低。SND与砂粒极显著负相关(P <0.01),与黏粒极显著正相关(P <0.001)。SND与N∶P和N∶K极显著正相关(P <0.001)。
图7
图7
3种植物群落土壤有机碳和氮密度与环境因子的相关性
注: *:在0.05水平差异显著;**:在0.01水平差异显著;***:在0.001水平差异显著。BD、SA、ST、CL、TP、TK、C∶N、C∶P、C∶K、N∶P、N∶K、P∶K、SCD、SND分别代表土壤容重、土壤砂粒含量、土壤粉粒含量、土壤黏粒含量、全磷含量、全钾含量、碳氮比、碳磷比、碳钾比、氮磷比、氮钾比、磷钾比、土壤有机碳密度和土壤氮密度
Fig.7
Correlation between soil organic carbon and nitrogen density and environmental factors in three plant communities
2.6 土壤有机碳密度、氮密度的影响因素
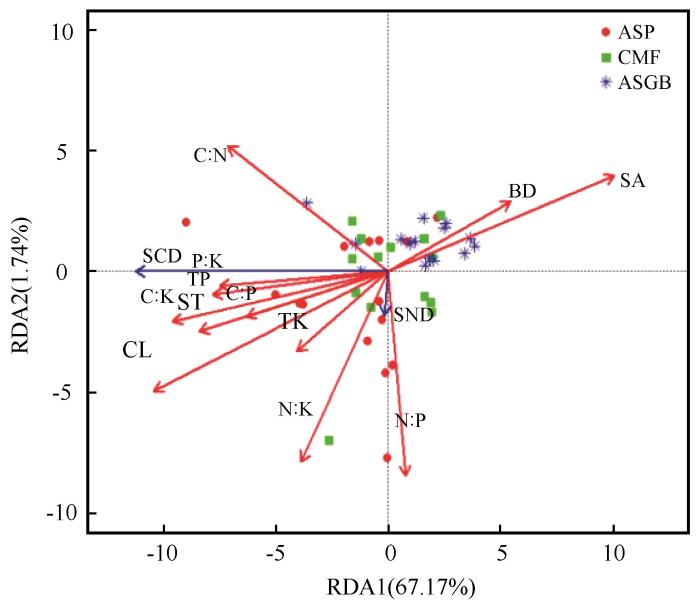
以SCD和SND为响应变量,BD、SA、ST、CL、TP、TK、C∶N、C∶P、C∶K、N∶P、N∶K、P∶K为解释变量进行冗余分析(图8 ),结果表明,第一轴解释了响应变量变异的67.17%,第二轴解释了响应变量变异的1.74%,累计解释率达到68.91%,表明第一轴在解释SCD和SND变异方面起到了主导作用,而第二轴的贡献相对较小。其中,C∶N与SCD和SND显著相关(P <0.05),对SCD和SND的解释率达15.71%;黏粒与SCD和SND极显著相关(P <0.01),对SCD和SND的解释率达32.85%。ASP群落多分布在RDA1负向、RDA2中低区间,落点更靠近左侧,体现出与CMF、ASGB群落空间分布的差异,靠近C∶N、ST等因子,说明其碳氮密度与C∶N、粉粒含量等因子相关。CMF群落集中在RDA1近原点及正向,与ASGB群落有部分重叠,受多因子综合作用,可能是相对广适性或受复杂因子协同影响的群落。ASGB群落主要分布在RDA1正向、RDA2中部范围,落点偏向右侧,贴近SA、BD因子,说明土壤砂粒、BD是驱动其碳氮密度的关键因子。
图8
图8
不同植物群落土壤物理性质、碳、氮、磷、钾及其化学计量比的冗余分析
注: 图中蓝线箭头为响应变量(土壤有机碳密度、土壤氮密度),红线箭头为解释变量(土壤全磷、全钾含量、砂粒粉粒黏粒含量、土壤容重以及碳氮磷钾化学计量特征),ASP代表黄蒿+禾本科群落、CMF代表小叶锦鸡儿+杂类草群落、ASGB代表沙米+狗尾草群落。BD、SA、ST、CL、TP、TK、C∶N、C∶P、C∶K、N∶P、N∶K、P∶K、SCD、SND分别代表土壤容重、土壤砂粒含量、土壤粉粒含量、土壤黏粒含量、全磷含量、全钾含量、碳氮比、碳磷比、碳钾比、氮磷比、氮钾比、磷钾比、土壤有机碳密度和土壤氮密度
Fig.8
Redundancy analysis of soil physical properties, carbon, nitrogen, phosphorus, potassium and their stoichiometric ratios in different plant communities
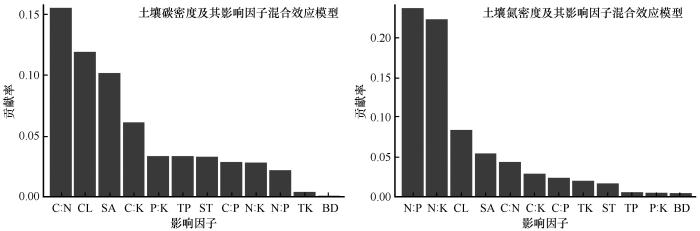
C∶N、CL和SA等12个预测变量解释了SCD变异的61.4%,C∶N、土壤黏粒含量和砂粒含量对SCD的贡献率最高,分别为15.5%、11.7%和10.1%(图9 )。N∶P、N∶K、CL等12个预测变量解释了SND变异的74.4%,N∶P、N∶K和土壤黏粒对SND的贡献率最高,分别为23.7%、22.3%和8.4%(图9 )。
图9
图9
土壤碳氮密度影响因子的重要性排序
注: BD、SA、ST、CL、TP、TK、C∶N、C∶P、C∶K、N∶P、N∶K、P∶K分别代表土壤容重、土壤砂粒含量、土壤粉粒含量、土壤黏粒含量、全磷含量、全钾含量、碳氮比、碳磷比、碳钾比、氮磷比、氮钾比、磷钾比
Fig.9
Importance ranking of factors affecting soil carbon and nitrogen density
3 讨论
3.1 土壤理化性质随土壤深度的变化特征
BD不仅反映了土壤颗粒的组成和通气状况,还是评估土壤结构和肥力的重要指标[24 ] 。3种植物群落的BD依次为ASGB群落>CMF群落>ASP群落,表明ASP群落的土壤通气状况较好,随土壤深度增加BD先增加后减小。3种植物群落的土壤砂粒含量随着土壤深度的增加而增加,粉粒和黏粒含量随着土壤深度的增加而减少,ASP群落砂粒含量最低,粉粒和黏粒含量最高,其次是CMF群落,ASGB群落砂粒含量最高,粉粒黏粒含量最少,这是因为ASGB植物群落资源处于极其匮乏的状态,现存群落拓殖能力有限,土壤种子库基础也相对薄弱,植物输入较少[25 ] 。不同土壤机械组成对有机物和化学元素的积累效果不同,黏粒具有较大的比表面积,可以较好地吸附土壤养分[26 -28 ] ,因此ASP群落的SOC、TN、TP以及TK均高于其他两个植物群落,CMF群落的含量高于ASGB群落。
科尔沁沙地3种植物群落的土壤养分存在明显差异。3种植物群落SOC、TN随着土壤深度的增加而减少,TP、TK无明显变化,这与以前的研究结果一致,这是因为SOC和TN主要来源于动植物残体,表层土壤植物根系比深层的丰富[16 ] 。
C∶N均沿土壤深度呈现先降低后升高的趋势,且最大值出现在60~100 cm,这可能是下层土壤有机质分解速率降低所致,在深层土壤氮的流失和被搬运速率高于碳[29 ] 。ASGB群落土壤C∶N在不同土层间变异系数为88.44%,这是因为表层土壤微生物活动旺盛,快速分解新鲜有机质,导致碳损失增加,从而降低碳氮比,而深层土壤缺氧环境显著抑制微生物活性,有机质以难分解的芳香化物质为主,这些物质碳含量高、氮含量低,导致碳氮比升高。N∶P随土壤深度的增加先升高后降低,C∶P、C∶K、N∶K、P∶K均随着土壤深度的增加而降低,且最大值均出现在ASP群落。TP主要受土壤母质的影响,而SOC和TN除受土壤母质的影响外,还受凋落物分解及植物根系输入的影响,这可能是造成草地沙化过程中土壤C∶P和N∶P下降的原因[30 ] 。
3.2 驱动土壤属性变化的主要因素
SCD和SND受土壤容重、土壤机械组成和化学计量等因素的影响,并随着沙地植被的演变而产生较大差异,SCD最大值均出现在ASP群落,而最小值均出现在ASGB群落。SCD在3种植被群落中随着土壤深度的增加均呈现先减小后增加的趋势,并且CMF群落和ASGB群落60~100 cm的SCD超过了0~10 cm,这可能是由于植被组成发生变化导致土壤碳在垂直梯度上发生了改变[31 ] 。SND在ASP群落随着土壤深度的增加呈现先减小再增加再减小的趋势,这可能与土壤中氮的迁移能力较强有关;CMF和ASGB群落SND随着土壤深度的增加先减小后增加,这可能与土壤中氮固定和释放过程有关[32 ] 。CMF群落60~100 cm土层SND高于0~10 cm土层,这可能与深层土壤中氮积累和循环过程有关。
ASP土壤碳氮密度最大,这是因为其群落生物量、物种多样性以及多年生植物优势度较高[33 ] 。随着土壤中C∶N的升高,SCD增大,SND减小;随着土壤中黏粒含量的增加,SCD和SND都增大。BD与SCD显著负相关,这是因为BD增加土壤孔隙度减小,破坏大团聚体结构,使包裹于内部的有机碳暴露,易被微生物分解,从而减少土壤碳密度[34 -35 ] 。C∶N、土壤黏粒含量和砂粒含量是影响SCD最主要的因素;N∶P、N∶K和土壤黏粒是影响SND最主要的因素。土壤碳氮密度特征也受到土壤微生物及其他环境因子的影响。随着土壤深度的增加土壤微环境发生改变,进而影响微生物群落及生物量,导致土壤碳氮密度差异,丁爽等[36 ] 在中国西南典型森林土壤的研究发现,微生物各类群生物量均在表层最高,随土壤深度的增加逐渐减少,进而影响土壤碳氮密度。此外,土壤含水量、土壤温度及pH等环境因子也影响土壤碳氮含量[37 ] ,这是因为不同植物群落不同土层的土壤含水量、温度和pH的不同,影响土壤微生物种类及其活性,进而影响土壤碳氮循环,最终导致土壤碳氮密度发生改变。
ASP群落为多年生草本优势群落,地上与地下生物量显著高于其他群落,其凋落物输入量更大且碳氮含量更高。其高凋落物输入和密集分布的须根系具有快速的周转速率,提升了表层土壤微生物的活性,促进了有机质的快速分解与转化,从而导致表层土壤碳氮积累。而CMF群落以灌木为主,其深根系通过周转分泌物、根瘤固氮直接向中深层土壤输入碳氮,其凋落物木质素含量高,分解速率较慢,碳氮存储更偏向于深层土壤。ASGB群落的根系可深入60~100 cm土层,通过深层根系周转促进该层土壤碳氮积累。未来需探究不同植物群落深层土壤碳氮库稳定性,为准确评估沙地生态系统碳氮储量提供数据支撑。
4 结论
SOC、TN、TP、TK及其化学计量和土壤碳氮密度在3种植物群落之间存在明显差异。ASP群落0~40 cm SOC、TN、TP含量最高。3种植物群落土壤C∶N均随着土壤深度的增加先降低后升高,N∶P随土壤深度的增加先升高后降低,土壤C∶P、C∶K、N∶K、P∶K在3种群落土壤中随着土壤深度的增加而降低。SCD与粉粒、黏粒、TP、C∶N、C∶K、P∶K显著正相关,与BD和砂粒负相关;SND与黏粒、N∶P、N∶K极显著正相关,与砂粒、C∶N极显著负相关。BD与SCD显著负相关,SCD主要受土壤C∶N、黏粒含量的影响,而SND主要受N∶P、N∶K和黏粒含量变化影响。
参考文献
View Option
[1]
左小安 ,赵学勇 ,赵哈林 ,等 科尔沁沙地草地退化过程中的物种组成及功能多样性变化特征
[J].水土保持学报 ,2006 ,20 (1 ):181 -185 .
[本文引用: 1]
[2]
张蕊 ,曹静娟 ,郭瑞英 ,等 祁连山北坡亚高山草地退耕还林草混合植被对土壤碳氮磷的影响
[J].生态环境学报 ,2014 ,23 (6 ):938 -944 .
[本文引用: 1]
[3]
刘玲莉 ,井新 ,任海燕 ,等 草地生物多样性与稳定性及对草地保护与修复的启示
[J].中国科学基金 ,2023 ,37 (4 ):560 -570 .
[本文引用: 1]
[4]
张铜会 ,赵哈林 ,大黑俊哉 ,等 连续放牧对沙质草地植被盖度、土壤性质及其空间分布的影响
[J].干旱区资源与环境 ,2003 ,17 (4 ):117 -121 .
[本文引用: 1]
[5]
赵啸龙 ,谢玉鸿 ,马旭君 ,等 科尔沁沙质草地不同恢复年限草本层群落结构及其与土壤理化性质的关系
[J].中国沙漠 ,2022 ,42 (2 ):134 -141 .
[本文引用: 1]
[6]
吕朋 ,左小安 ,岳喜元 ,等 科尔沁沙地封育过程中植被特征的动态变化
[J].生态学杂志 ,2018 ,37 (10 ):2880 -2888 .
[本文引用: 1]
[7]
李雪华 ,韩士杰 ,蒋德明 ,等 科尔沁沙地不同演替阶段植被特征及固沙作用
[J].辽宁工程技术大学学报 ,2006 ,25 (5 ):789 -791 .
[本文引用: 1]
[8]
左小安 ,赵学勇 ,赵哈林 ,等 沙地退化植被恢复过程中植被的空间异质性
[J].生态环境学报 ,2010 ,19 (7 ):1513 -1518 .
[本文引用: 1]
[9]
王明明 ,刘新平 ,何玉惠 ,等 科尔沁沙地封育恢复过程中植物群落特征变化及影响因素
[J].植物生态学报 ,2019 ,43 (8 ):672 -684 .
[本文引用: 1]
[10]
包天玲 ,刘继亮 ,苑峰 ,等 科尔沁沙质草地植物群落对增温的响应
[J].中国沙漠 ,2024 ,44 (1 ):151 -160 .
[本文引用: 1]
[11]
朱远忠 ,黄文达 ,于海伦 ,等 科尔沁沙质草地不同水热梯度植物群落叶功能性状特征
[J].中国沙漠 ,2024 ,44 (2 ):143 -150 .
[本文引用: 1]
[12]
周静 ,孙永峰 ,丁杰萍 ,等 退化沙质草地恢复过程中植被生物量变化及其与土壤碳的关系
[J].干旱区研究 ,2023 ,40 (9 ):1457 -1464 .
[本文引用: 1]
[13]
詹瑾 ,韩丹 ,杨红玲 ,等 科尔沁沙地植被恢复过程中群落组成及多样性演变特征
[J].中国沙漠 ,2022 ,42 (2 ):194 -206 .
[14]
赵丽娅 ,钟韩珊 ,赵美玉 ,等 围封和放牧对科尔沁沙地群落物种多样性与地上生物量的影响
[J].生态环境学报 ,2018 ,27 (10 ):1783 -1790 .
[15]
左小安 ,赵哈林 ,赵学勇 ,等 科尔沁沙地不同恢复年限退化植被的物种多样性
[J].草业学报 ,2009 ,18 (4 ):9 -16 .
[本文引用: 1]
[16]
曹雯婕 ,李玉强 ,陈银萍 ,等 科尔沁沙地不同土地利用类型土壤化学计量特征
[J].应用生态学报 ,2022 ,33 (12 ):3312 -3320 .
[本文引用: 2]
[17]
宁志英 ,李玉霖 ,杨红玲 ,等 科尔沁沙地主要植物细根和叶片碳、氮、磷化学计量特征
[J].植物生态学报 ,2017 ,41 (10 ):1069 -1080 .
[本文引用: 1]
[18]
王毅 ,刘碧颖 ,刘苗 ,等 若尔盖地区沙化草地土壤酶协同和抑制效应
[J].草业科学 ,2019 ,36 (4 ):939 -951 .
[本文引用: 1]
[19]
王亚妮 ,胡宜刚 ,王增如 ,等 沙化和人工植被重建对高寒草地土壤细菌群落特征的影响
[J].草业学报 ,2022 ,31 (5 ):26 -39 .
[本文引用: 1]
[20]
张晶 ,左小安 ,吕朋 生长季降水格局变化对科尔沁沙地典型生境植物群落结构、功能和地上生物量的影响
[J].中国沙漠 ,2024 ,44 (4 ):1 -13 .
[本文引用: 1]
[21]
张亦凡 ,王海燕 ,高子滢 ,等 华南地区森林土壤有机碳和全氮密度及储量估算
[J].中国土壤与肥料 ,2024 ,(12 ):17 -26 .
[本文引用: 1]
[22]
Lai J S Zhu W J Cui D F et al Extension of the glmm.hp package to zero-inflated generalized linear mixed models and multiple regression
[J].Journal of Plant Ecology ,2022 ,16 (6 ):rtad038 .
[本文引用: 1]
[23]
Lai J S Zou Y Zhang S et al glmm.hp:an R package for computing individual effect of predictors in generalized linear mixed models
[J].Journal of Plant Ecology ,2022 ,15 (6 ):1302 -1307 .
[本文引用: 1]
[24]
赵丽娅 ,李兆华 ,李锋瑞 ,等 科尔沁沙地植被恢复演替进程中群落土壤种子库研究
[J].生态学报 ,2005 ,25 (12 ):3204 -3211 .
[本文引用: 1]
[25]
Bronick C J Lal R Soil structure and management:a review
[J].Geoderma ,2005 ,124 :3 -22 .
[本文引用: 1]
[26]
Six J Conant R T Paul E A et al Stabilization mechanisms of soil organic matter:implications for C-saturation of soils
[J].Plant and Soil ,2002 ,241 (1/2 ):155 -176 .
[本文引用: 1]
[27]
John B Yamashita T Ludwig B et al Storage of organic carbon in aggregate and density fractions of silty soils under different types of land use
[J].Geoderma ,2005 ,128 :63 -79 .
[28]
徐海军 ,姚琴 ,孙宇峰 ,等 大庆地区不同土地利用下土壤空间异质性研究
[J].中国土壤与肥料 ,2020 (1 ):24 -30 .
[本文引用: 1]
[29]
冯德枫 ,包维楷 土壤碳氮磷化学计量比时空格局及影响因素研究进展
[J].应用与环境生物学报 ,2017 ,23 (2 ):400 -408 .
[本文引用: 1]
[30]
宁志英 ,李玉霖 ,杨红玲 ,等 沙化草地土壤碳氮磷化学计量特征及其对植被生产力和多样性的影响
[J].生态学报 ,2019 ,39 (10 ):3537 -3546 .
[本文引用: 1]
[31]
Sun R J Sun K Liu L et al Changes in soil carbon and nitrogen along a 3-m vertical profile and environmental regulation in alpine grassland on the Tibetan Plateau
[J].Journal of Geophysical Research:Biogeosciences ,2024 ,129 (7 ):e2023JG007579 .
[本文引用: 1]
[32]
Wu J H Pang H Zhou Y et al Effects of vegetation degradation on carbon and nitrogen distribution of roots-soil system in temperate typical steppe
[J].Acta Ecologica Sinica ,2018 ,38 (15 ):5340 -5350 .
[本文引用: 1]
[33]
胡亚伟 ,孙若修 ,申明爽 ,等 晋西黄土区土地利用方式对土壤C∶N∶P化学计量特征及土壤理化性质的影响
[J].干旱区研究 ,2021 ,38 (4 ):990 -999 .
[本文引用: 1]
[34]
阎欣 ,安慧 ,刘任涛 荒漠草原沙漠化对土壤物理和化学特性的影响
[J].土壤 ,2019 ,51 (5 ):1006 -1012 .
[本文引用: 1]
[35]
曹生奎 ,陈克龙 ,曹广超 ,等 草地退化对青海湖流域小蒿草草甸土壤碳密度的影响
[J].水土保持研究 ,2014 ,21 (1 ):71 -75 .
[本文引用: 1]
[36]
丁爽 ,魏圣钊 ,陈真亮 ,等 中国西南典型森林土壤微生物在不同土壤深度下的变化特征
[J].应用生态学报 ,2023 ,34 (3 ):614 -622 .
[本文引用: 1]
[37]
刘旻霞 ,蒋晓轩 ,李全弟 ,等 兰州北山不同植被土壤可培养微生物丰度变化特征及影响因素
[J].中国环境科学 ,2020 ,40 (6 ):2683 -2691 .
[本文引用: 1]
科尔沁沙地草地退化过程中的物种组成及功能多样性变化特征
1
2006
... 近年来,全球气候变化加剧,气温升高、降水格局改变及极端气候事件频发,沙质草地生态系统面临着前所未有的压力和挑战.气候变化直接影响植物生长发育、群落组成和分布格局,进而间接改变土壤有机碳氮循环过程和存储格局.此外,人类活动如过度放牧、开垦、城市化扩张等,也在不断干扰和破坏沙质草地生态系统原有结构和功能,进一步加剧土壤有机碳氮的流失从而导致生态系统退化[1 ] .土壤有机碳、全氮是陆地生态系统中至关重要的组成部分[2 ] ,不仅在土壤肥力、植物生长和生态系统生产力方面发挥关键作用,还参与全球碳氮循环过程,对气候变化反馈调节机制有重要作用. ...
祁连山北坡亚高山草地退耕还林草混合植被对土壤碳氮磷的影响
1
2014
... 近年来,全球气候变化加剧,气温升高、降水格局改变及极端气候事件频发,沙质草地生态系统面临着前所未有的压力和挑战.气候变化直接影响植物生长发育、群落组成和分布格局,进而间接改变土壤有机碳氮循环过程和存储格局.此外,人类活动如过度放牧、开垦、城市化扩张等,也在不断干扰和破坏沙质草地生态系统原有结构和功能,进一步加剧土壤有机碳氮的流失从而导致生态系统退化[1 ] .土壤有机碳、全氮是陆地生态系统中至关重要的组成部分[2 ] ,不仅在土壤肥力、植物生长和生态系统生产力方面发挥关键作用,还参与全球碳氮循环过程,对气候变化反馈调节机制有重要作用. ...
草地生物多样性与稳定性及对草地保护与修复的启示
1
2023
... 沙质草地生态系统结构和功能具有独特性和脆弱性,对于维持区域生态平衡和生物多样性保护具有不可忽视的意义[3 ] .沙质草地生态系统植被覆盖度相对较低,土壤质地疏松,保水保肥能力较差,使得其土壤有机碳氮含量与动态变化相较于其他生态系统更为敏感,受环境因素和人类活动影响更为显著[4 ] .研究沙质草地不同植物群落有机碳氮密度特征,对植被与土壤恢复具有重要参考意义[5 ] . ...
连续放牧对沙质草地植被盖度、土壤性质及其空间分布的影响
1
2003
... 沙质草地生态系统结构和功能具有独特性和脆弱性,对于维持区域生态平衡和生物多样性保护具有不可忽视的意义[3 ] .沙质草地生态系统植被覆盖度相对较低,土壤质地疏松,保水保肥能力较差,使得其土壤有机碳氮含量与动态变化相较于其他生态系统更为敏感,受环境因素和人类活动影响更为显著[4 ] .研究沙质草地不同植物群落有机碳氮密度特征,对植被与土壤恢复具有重要参考意义[5 ] . ...
科尔沁沙质草地不同恢复年限草本层群落结构及其与土壤理化性质的关系
1
2022
... 沙质草地生态系统结构和功能具有独特性和脆弱性,对于维持区域生态平衡和生物多样性保护具有不可忽视的意义[3 ] .沙质草地生态系统植被覆盖度相对较低,土壤质地疏松,保水保肥能力较差,使得其土壤有机碳氮含量与动态变化相较于其他生态系统更为敏感,受环境因素和人类活动影响更为显著[4 ] .研究沙质草地不同植物群落有机碳氮密度特征,对植被与土壤恢复具有重要参考意义[5 ] . ...
科尔沁沙地封育过程中植被特征的动态变化
1
2018
... 由于气候变化和过度放牧等影响,科尔沁沙地沙质草地植被退化,土地沙化严重.随着围栏封育等固沙措施的实施,该区出现了不同物种组成的群落[6 ] .随着封育年限增加,群落植被盖度和物种丰富度明显增加[7 -8 ] ,土壤种子库也随之改变[9 ] .科尔沁沙地沙质草地的研究主要集中在植物群落对不同环境的响应[10 -11 ] 、退化沙质草地恢复过程中生物多样性变化[12 -15 ] 、土壤化学计量研究[16 -17 ] 、土壤营养元素随草地沙化程度的变化[18 ] 、沙质草地恢复过程中土壤微生物群落变化[19 ] 等方面,但鲜有关于沙质草地不同植物群落土壤有机碳和全氮密度特征的研究. ...
科尔沁沙地不同演替阶段植被特征及固沙作用
1
2006
... 由于气候变化和过度放牧等影响,科尔沁沙地沙质草地植被退化,土地沙化严重.随着围栏封育等固沙措施的实施,该区出现了不同物种组成的群落[6 ] .随着封育年限增加,群落植被盖度和物种丰富度明显增加[7 -8 ] ,土壤种子库也随之改变[9 ] .科尔沁沙地沙质草地的研究主要集中在植物群落对不同环境的响应[10 -11 ] 、退化沙质草地恢复过程中生物多样性变化[12 -15 ] 、土壤化学计量研究[16 -17 ] 、土壤营养元素随草地沙化程度的变化[18 ] 、沙质草地恢复过程中土壤微生物群落变化[19 ] 等方面,但鲜有关于沙质草地不同植物群落土壤有机碳和全氮密度特征的研究. ...
沙地退化植被恢复过程中植被的空间异质性
1
2010
... 由于气候变化和过度放牧等影响,科尔沁沙地沙质草地植被退化,土地沙化严重.随着围栏封育等固沙措施的实施,该区出现了不同物种组成的群落[6 ] .随着封育年限增加,群落植被盖度和物种丰富度明显增加[7 -8 ] ,土壤种子库也随之改变[9 ] .科尔沁沙地沙质草地的研究主要集中在植物群落对不同环境的响应[10 -11 ] 、退化沙质草地恢复过程中生物多样性变化[12 -15 ] 、土壤化学计量研究[16 -17 ] 、土壤营养元素随草地沙化程度的变化[18 ] 、沙质草地恢复过程中土壤微生物群落变化[19 ] 等方面,但鲜有关于沙质草地不同植物群落土壤有机碳和全氮密度特征的研究. ...
科尔沁沙地封育恢复过程中植物群落特征变化及影响因素
1
2019
... 由于气候变化和过度放牧等影响,科尔沁沙地沙质草地植被退化,土地沙化严重.随着围栏封育等固沙措施的实施,该区出现了不同物种组成的群落[6 ] .随着封育年限增加,群落植被盖度和物种丰富度明显增加[7 -8 ] ,土壤种子库也随之改变[9 ] .科尔沁沙地沙质草地的研究主要集中在植物群落对不同环境的响应[10 -11 ] 、退化沙质草地恢复过程中生物多样性变化[12 -15 ] 、土壤化学计量研究[16 -17 ] 、土壤营养元素随草地沙化程度的变化[18 ] 、沙质草地恢复过程中土壤微生物群落变化[19 ] 等方面,但鲜有关于沙质草地不同植物群落土壤有机碳和全氮密度特征的研究. ...
科尔沁沙质草地植物群落对增温的响应
1
2024
... 由于气候变化和过度放牧等影响,科尔沁沙地沙质草地植被退化,土地沙化严重.随着围栏封育等固沙措施的实施,该区出现了不同物种组成的群落[6 ] .随着封育年限增加,群落植被盖度和物种丰富度明显增加[7 -8 ] ,土壤种子库也随之改变[9 ] .科尔沁沙地沙质草地的研究主要集中在植物群落对不同环境的响应[10 -11 ] 、退化沙质草地恢复过程中生物多样性变化[12 -15 ] 、土壤化学计量研究[16 -17 ] 、土壤营养元素随草地沙化程度的变化[18 ] 、沙质草地恢复过程中土壤微生物群落变化[19 ] 等方面,但鲜有关于沙质草地不同植物群落土壤有机碳和全氮密度特征的研究. ...
科尔沁沙质草地不同水热梯度植物群落叶功能性状特征
1
2024
... 由于气候变化和过度放牧等影响,科尔沁沙地沙质草地植被退化,土地沙化严重.随着围栏封育等固沙措施的实施,该区出现了不同物种组成的群落[6 ] .随着封育年限增加,群落植被盖度和物种丰富度明显增加[7 -8 ] ,土壤种子库也随之改变[9 ] .科尔沁沙地沙质草地的研究主要集中在植物群落对不同环境的响应[10 -11 ] 、退化沙质草地恢复过程中生物多样性变化[12 -15 ] 、土壤化学计量研究[16 -17 ] 、土壤营养元素随草地沙化程度的变化[18 ] 、沙质草地恢复过程中土壤微生物群落变化[19 ] 等方面,但鲜有关于沙质草地不同植物群落土壤有机碳和全氮密度特征的研究. ...
退化沙质草地恢复过程中植被生物量变化及其与土壤碳的关系
1
2023
... 由于气候变化和过度放牧等影响,科尔沁沙地沙质草地植被退化,土地沙化严重.随着围栏封育等固沙措施的实施,该区出现了不同物种组成的群落[6 ] .随着封育年限增加,群落植被盖度和物种丰富度明显增加[7 -8 ] ,土壤种子库也随之改变[9 ] .科尔沁沙地沙质草地的研究主要集中在植物群落对不同环境的响应[10 -11 ] 、退化沙质草地恢复过程中生物多样性变化[12 -15 ] 、土壤化学计量研究[16 -17 ] 、土壤营养元素随草地沙化程度的变化[18 ] 、沙质草地恢复过程中土壤微生物群落变化[19 ] 等方面,但鲜有关于沙质草地不同植物群落土壤有机碳和全氮密度特征的研究. ...
科尔沁沙地植被恢复过程中群落组成及多样性演变特征
0
2022
围封和放牧对科尔沁沙地群落物种多样性与地上生物量的影响
0
2018
科尔沁沙地不同恢复年限退化植被的物种多样性
1
2009
... 由于气候变化和过度放牧等影响,科尔沁沙地沙质草地植被退化,土地沙化严重.随着围栏封育等固沙措施的实施,该区出现了不同物种组成的群落[6 ] .随着封育年限增加,群落植被盖度和物种丰富度明显增加[7 -8 ] ,土壤种子库也随之改变[9 ] .科尔沁沙地沙质草地的研究主要集中在植物群落对不同环境的响应[10 -11 ] 、退化沙质草地恢复过程中生物多样性变化[12 -15 ] 、土壤化学计量研究[16 -17 ] 、土壤营养元素随草地沙化程度的变化[18 ] 、沙质草地恢复过程中土壤微生物群落变化[19 ] 等方面,但鲜有关于沙质草地不同植物群落土壤有机碳和全氮密度特征的研究. ...
科尔沁沙地不同土地利用类型土壤化学计量特征
2
2022
... 由于气候变化和过度放牧等影响,科尔沁沙地沙质草地植被退化,土地沙化严重.随着围栏封育等固沙措施的实施,该区出现了不同物种组成的群落[6 ] .随着封育年限增加,群落植被盖度和物种丰富度明显增加[7 -8 ] ,土壤种子库也随之改变[9 ] .科尔沁沙地沙质草地的研究主要集中在植物群落对不同环境的响应[10 -11 ] 、退化沙质草地恢复过程中生物多样性变化[12 -15 ] 、土壤化学计量研究[16 -17 ] 、土壤营养元素随草地沙化程度的变化[18 ] 、沙质草地恢复过程中土壤微生物群落变化[19 ] 等方面,但鲜有关于沙质草地不同植物群落土壤有机碳和全氮密度特征的研究. ...
... 科尔沁沙地3种植物群落的土壤养分存在明显差异.3种植物群落SOC、TN随着土壤深度的增加而减少,TP、TK无明显变化,这与以前的研究结果一致,这是因为SOC和TN主要来源于动植物残体,表层土壤植物根系比深层的丰富[16 ] . ...
科尔沁沙地主要植物细根和叶片碳、氮、磷化学计量特征
1
2017
... 由于气候变化和过度放牧等影响,科尔沁沙地沙质草地植被退化,土地沙化严重.随着围栏封育等固沙措施的实施,该区出现了不同物种组成的群落[6 ] .随着封育年限增加,群落植被盖度和物种丰富度明显增加[7 -8 ] ,土壤种子库也随之改变[9 ] .科尔沁沙地沙质草地的研究主要集中在植物群落对不同环境的响应[10 -11 ] 、退化沙质草地恢复过程中生物多样性变化[12 -15 ] 、土壤化学计量研究[16 -17 ] 、土壤营养元素随草地沙化程度的变化[18 ] 、沙质草地恢复过程中土壤微生物群落变化[19 ] 等方面,但鲜有关于沙质草地不同植物群落土壤有机碳和全氮密度特征的研究. ...
若尔盖地区沙化草地土壤酶协同和抑制效应
1
2019
... 由于气候变化和过度放牧等影响,科尔沁沙地沙质草地植被退化,土地沙化严重.随着围栏封育等固沙措施的实施,该区出现了不同物种组成的群落[6 ] .随着封育年限增加,群落植被盖度和物种丰富度明显增加[7 -8 ] ,土壤种子库也随之改变[9 ] .科尔沁沙地沙质草地的研究主要集中在植物群落对不同环境的响应[10 -11 ] 、退化沙质草地恢复过程中生物多样性变化[12 -15 ] 、土壤化学计量研究[16 -17 ] 、土壤营养元素随草地沙化程度的变化[18 ] 、沙质草地恢复过程中土壤微生物群落变化[19 ] 等方面,但鲜有关于沙质草地不同植物群落土壤有机碳和全氮密度特征的研究. ...
沙化和人工植被重建对高寒草地土壤细菌群落特征的影响
1
2022
... 由于气候变化和过度放牧等影响,科尔沁沙地沙质草地植被退化,土地沙化严重.随着围栏封育等固沙措施的实施,该区出现了不同物种组成的群落[6 ] .随着封育年限增加,群落植被盖度和物种丰富度明显增加[7 -8 ] ,土壤种子库也随之改变[9 ] .科尔沁沙地沙质草地的研究主要集中在植物群落对不同环境的响应[10 -11 ] 、退化沙质草地恢复过程中生物多样性变化[12 -15 ] 、土壤化学计量研究[16 -17 ] 、土壤营养元素随草地沙化程度的变化[18 ] 、沙质草地恢复过程中土壤微生物群落变化[19 ] 等方面,但鲜有关于沙质草地不同植物群落土壤有机碳和全氮密度特征的研究. ...
生长季降水格局变化对科尔沁沙地典型生境植物群落结构、功能和地上生物量的影响
1
2024
... 研究区域(42°41′N、120°55′E)位于中国北方半干旱区科尔沁沙地,该区属于温带大陆性半干旱季风气候,具有春季干燥多风、夏季炎热多雨、秋季凉爽短暂、冬季寒冷漫长的特点.研究区海拔358 m,多年平均气温约为6.4 ℃,多年平均降水量为360 mm,多年平均潜在蒸发量为2 000 mm[20 ] ,干旱程度较高.土壤主要为风沙土、沙质栗钙土和草甸土.植被组成以狗尾草(Setaria viridis )、黄蒿(Artemisia scoparia )、盐蒿(Suaeda salsa )、沙米(Agriophyllum squarrosum )、白草(Pennisetum flaccidum )、扁蓿豆(Medicago ruthenica )、尖头叶藜(Chenopodium acuminatum )、小叶锦鸡儿(Caragana microphylla )等为主.研究区位置如图1 所示. ...
华南地区森林土壤有机碳和全氮密度及储量估算
1
2024
... 式中:SCD 为土壤有机碳密度(g·m-2 );SND 为土壤氮密度(g·m-2 );BD 为土壤容重(g·cm-3 );H 为土层厚度(cm);SOC 为土壤有机碳含量(g·kg-1 );TN 为土壤全氮含量(g·kg-1 );10为换算系数[21 ] . ...
Extension of the glmm.hp package to zero-inflated generalized linear mixed models and multiple regression
1
2022
... 文中数据使用Excel 2019软件整理,使用SPSS软件(27.0.1)、Origin2021和R软件(4.4.1)进行冗余分析以及广义线性混合模型[22 -23 ] 建立和绘图.不同植物群落和不同土层土壤容重、土壤机械组成、土壤碳氮磷钾含量以及土壤碳氮密度采用单因素方差分析法进行差异性比较.显著性检验采用邓肯法,显著水平为0.05. ...
glmm.hp:an R package for computing individual effect of predictors in generalized linear mixed models
1
2022
... 文中数据使用Excel 2019软件整理,使用SPSS软件(27.0.1)、Origin2021和R软件(4.4.1)进行冗余分析以及广义线性混合模型[22 -23 ] 建立和绘图.不同植物群落和不同土层土壤容重、土壤机械组成、土壤碳氮磷钾含量以及土壤碳氮密度采用单因素方差分析法进行差异性比较.显著性检验采用邓肯法,显著水平为0.05. ...
科尔沁沙地植被恢复演替进程中群落土壤种子库研究
1
2005
... BD不仅反映了土壤颗粒的组成和通气状况,还是评估土壤结构和肥力的重要指标[24 ] .3种植物群落的BD依次为ASGB群落>CMF群落>ASP群落,表明ASP群落的土壤通气状况较好,随土壤深度增加BD先增加后减小.3种植物群落的土壤砂粒含量随着土壤深度的增加而增加,粉粒和黏粒含量随着土壤深度的增加而减少,ASP群落砂粒含量最低,粉粒和黏粒含量最高,其次是CMF群落,ASGB群落砂粒含量最高,粉粒黏粒含量最少,这是因为ASGB植物群落资源处于极其匮乏的状态,现存群落拓殖能力有限,土壤种子库基础也相对薄弱,植物输入较少[25 ] .不同土壤机械组成对有机物和化学元素的积累效果不同,黏粒具有较大的比表面积,可以较好地吸附土壤养分[26 -28 ] ,因此ASP群落的SOC、TN、TP以及TK均高于其他两个植物群落,CMF群落的含量高于ASGB群落. ...
Soil structure and management:a review
1
2005
... BD不仅反映了土壤颗粒的组成和通气状况,还是评估土壤结构和肥力的重要指标[24 ] .3种植物群落的BD依次为ASGB群落>CMF群落>ASP群落,表明ASP群落的土壤通气状况较好,随土壤深度增加BD先增加后减小.3种植物群落的土壤砂粒含量随着土壤深度的增加而增加,粉粒和黏粒含量随着土壤深度的增加而减少,ASP群落砂粒含量最低,粉粒和黏粒含量最高,其次是CMF群落,ASGB群落砂粒含量最高,粉粒黏粒含量最少,这是因为ASGB植物群落资源处于极其匮乏的状态,现存群落拓殖能力有限,土壤种子库基础也相对薄弱,植物输入较少[25 ] .不同土壤机械组成对有机物和化学元素的积累效果不同,黏粒具有较大的比表面积,可以较好地吸附土壤养分[26 -28 ] ,因此ASP群落的SOC、TN、TP以及TK均高于其他两个植物群落,CMF群落的含量高于ASGB群落. ...
Stabilization mechanisms of soil organic matter:implications for C-saturation of soils
1
2002
... BD不仅反映了土壤颗粒的组成和通气状况,还是评估土壤结构和肥力的重要指标[24 ] .3种植物群落的BD依次为ASGB群落>CMF群落>ASP群落,表明ASP群落的土壤通气状况较好,随土壤深度增加BD先增加后减小.3种植物群落的土壤砂粒含量随着土壤深度的增加而增加,粉粒和黏粒含量随着土壤深度的增加而减少,ASP群落砂粒含量最低,粉粒和黏粒含量最高,其次是CMF群落,ASGB群落砂粒含量最高,粉粒黏粒含量最少,这是因为ASGB植物群落资源处于极其匮乏的状态,现存群落拓殖能力有限,土壤种子库基础也相对薄弱,植物输入较少[25 ] .不同土壤机械组成对有机物和化学元素的积累效果不同,黏粒具有较大的比表面积,可以较好地吸附土壤养分[26 -28 ] ,因此ASP群落的SOC、TN、TP以及TK均高于其他两个植物群落,CMF群落的含量高于ASGB群落. ...
Storage of organic carbon in aggregate and density fractions of silty soils under different types of land use
0
2005
大庆地区不同土地利用下土壤空间异质性研究
1
2020
... BD不仅反映了土壤颗粒的组成和通气状况,还是评估土壤结构和肥力的重要指标[24 ] .3种植物群落的BD依次为ASGB群落>CMF群落>ASP群落,表明ASP群落的土壤通气状况较好,随土壤深度增加BD先增加后减小.3种植物群落的土壤砂粒含量随着土壤深度的增加而增加,粉粒和黏粒含量随着土壤深度的增加而减少,ASP群落砂粒含量最低,粉粒和黏粒含量最高,其次是CMF群落,ASGB群落砂粒含量最高,粉粒黏粒含量最少,这是因为ASGB植物群落资源处于极其匮乏的状态,现存群落拓殖能力有限,土壤种子库基础也相对薄弱,植物输入较少[25 ] .不同土壤机械组成对有机物和化学元素的积累效果不同,黏粒具有较大的比表面积,可以较好地吸附土壤养分[26 -28 ] ,因此ASP群落的SOC、TN、TP以及TK均高于其他两个植物群落,CMF群落的含量高于ASGB群落. ...
土壤碳氮磷化学计量比时空格局及影响因素研究进展
1
2017
... C∶N均沿土壤深度呈现先降低后升高的趋势,且最大值出现在60~100 cm,这可能是下层土壤有机质分解速率降低所致,在深层土壤氮的流失和被搬运速率高于碳[29 ] .ASGB群落土壤C∶N在不同土层间变异系数为88.44%,这是因为表层土壤微生物活动旺盛,快速分解新鲜有机质,导致碳损失增加,从而降低碳氮比,而深层土壤缺氧环境显著抑制微生物活性,有机质以难分解的芳香化物质为主,这些物质碳含量高、氮含量低,导致碳氮比升高.N∶P随土壤深度的增加先升高后降低,C∶P、C∶K、N∶K、P∶K均随着土壤深度的增加而降低,且最大值均出现在ASP群落.TP主要受土壤母质的影响,而SOC和TN除受土壤母质的影响外,还受凋落物分解及植物根系输入的影响,这可能是造成草地沙化过程中土壤C∶P和N∶P下降的原因[30 ] . ...
沙化草地土壤碳氮磷化学计量特征及其对植被生产力和多样性的影响
1
2019
... C∶N均沿土壤深度呈现先降低后升高的趋势,且最大值出现在60~100 cm,这可能是下层土壤有机质分解速率降低所致,在深层土壤氮的流失和被搬运速率高于碳[29 ] .ASGB群落土壤C∶N在不同土层间变异系数为88.44%,这是因为表层土壤微生物活动旺盛,快速分解新鲜有机质,导致碳损失增加,从而降低碳氮比,而深层土壤缺氧环境显著抑制微生物活性,有机质以难分解的芳香化物质为主,这些物质碳含量高、氮含量低,导致碳氮比升高.N∶P随土壤深度的增加先升高后降低,C∶P、C∶K、N∶K、P∶K均随着土壤深度的增加而降低,且最大值均出现在ASP群落.TP主要受土壤母质的影响,而SOC和TN除受土壤母质的影响外,还受凋落物分解及植物根系输入的影响,这可能是造成草地沙化过程中土壤C∶P和N∶P下降的原因[30 ] . ...
Changes in soil carbon and nitrogen along a 3-m vertical profile and environmental regulation in alpine grassland on the Tibetan Plateau
1
2024
... SCD和SND受土壤容重、土壤机械组成和化学计量等因素的影响,并随着沙地植被的演变而产生较大差异,SCD最大值均出现在ASP群落,而最小值均出现在ASGB群落.SCD在3种植被群落中随着土壤深度的增加均呈现先减小后增加的趋势,并且CMF群落和ASGB群落60~100 cm的SCD超过了0~10 cm,这可能是由于植被组成发生变化导致土壤碳在垂直梯度上发生了改变[31 ] .SND在ASP群落随着土壤深度的增加呈现先减小再增加再减小的趋势,这可能与土壤中氮的迁移能力较强有关;CMF和ASGB群落SND随着土壤深度的增加先减小后增加,这可能与土壤中氮固定和释放过程有关[32 ] .CMF群落60~100 cm土层SND高于0~10 cm土层,这可能与深层土壤中氮积累和循环过程有关. ...
Effects of vegetation degradation on carbon and nitrogen distribution of roots-soil system in temperate typical steppe
1
2018
... SCD和SND受土壤容重、土壤机械组成和化学计量等因素的影响,并随着沙地植被的演变而产生较大差异,SCD最大值均出现在ASP群落,而最小值均出现在ASGB群落.SCD在3种植被群落中随着土壤深度的增加均呈现先减小后增加的趋势,并且CMF群落和ASGB群落60~100 cm的SCD超过了0~10 cm,这可能是由于植被组成发生变化导致土壤碳在垂直梯度上发生了改变[31 ] .SND在ASP群落随着土壤深度的增加呈现先减小再增加再减小的趋势,这可能与土壤中氮的迁移能力较强有关;CMF和ASGB群落SND随着土壤深度的增加先减小后增加,这可能与土壤中氮固定和释放过程有关[32 ] .CMF群落60~100 cm土层SND高于0~10 cm土层,这可能与深层土壤中氮积累和循环过程有关. ...
晋西黄土区土地利用方式对土壤C∶N∶P化学计量特征及土壤理化性质的影响
1
2021
... ASP土壤碳氮密度最大,这是因为其群落生物量、物种多样性以及多年生植物优势度较高[33 ] .随着土壤中C∶N的升高,SCD增大,SND减小;随着土壤中黏粒含量的增加,SCD和SND都增大.BD与SCD显著负相关,这是因为BD增加土壤孔隙度减小,破坏大团聚体结构,使包裹于内部的有机碳暴露,易被微生物分解,从而减少土壤碳密度[34 -35 ] .C∶N、土壤黏粒含量和砂粒含量是影响SCD最主要的因素;N∶P、N∶K和土壤黏粒是影响SND最主要的因素.土壤碳氮密度特征也受到土壤微生物及其他环境因子的影响.随着土壤深度的增加土壤微环境发生改变,进而影响微生物群落及生物量,导致土壤碳氮密度差异,丁爽等[36 ] 在中国西南典型森林土壤的研究发现,微生物各类群生物量均在表层最高,随土壤深度的增加逐渐减少,进而影响土壤碳氮密度.此外,土壤含水量、土壤温度及pH等环境因子也影响土壤碳氮含量[37 ] ,这是因为不同植物群落不同土层的土壤含水量、温度和pH的不同,影响土壤微生物种类及其活性,进而影响土壤碳氮循环,最终导致土壤碳氮密度发生改变. ...
荒漠草原沙漠化对土壤物理和化学特性的影响
1
2019
... ASP土壤碳氮密度最大,这是因为其群落生物量、物种多样性以及多年生植物优势度较高[33 ] .随着土壤中C∶N的升高,SCD增大,SND减小;随着土壤中黏粒含量的增加,SCD和SND都增大.BD与SCD显著负相关,这是因为BD增加土壤孔隙度减小,破坏大团聚体结构,使包裹于内部的有机碳暴露,易被微生物分解,从而减少土壤碳密度[34 -35 ] .C∶N、土壤黏粒含量和砂粒含量是影响SCD最主要的因素;N∶P、N∶K和土壤黏粒是影响SND最主要的因素.土壤碳氮密度特征也受到土壤微生物及其他环境因子的影响.随着土壤深度的增加土壤微环境发生改变,进而影响微生物群落及生物量,导致土壤碳氮密度差异,丁爽等[36 ] 在中国西南典型森林土壤的研究发现,微生物各类群生物量均在表层最高,随土壤深度的增加逐渐减少,进而影响土壤碳氮密度.此外,土壤含水量、土壤温度及pH等环境因子也影响土壤碳氮含量[37 ] ,这是因为不同植物群落不同土层的土壤含水量、温度和pH的不同,影响土壤微生物种类及其活性,进而影响土壤碳氮循环,最终导致土壤碳氮密度发生改变. ...
草地退化对青海湖流域小蒿草草甸土壤碳密度的影响
1
2014
... ASP土壤碳氮密度最大,这是因为其群落生物量、物种多样性以及多年生植物优势度较高[33 ] .随着土壤中C∶N的升高,SCD增大,SND减小;随着土壤中黏粒含量的增加,SCD和SND都增大.BD与SCD显著负相关,这是因为BD增加土壤孔隙度减小,破坏大团聚体结构,使包裹于内部的有机碳暴露,易被微生物分解,从而减少土壤碳密度[34 -35 ] .C∶N、土壤黏粒含量和砂粒含量是影响SCD最主要的因素;N∶P、N∶K和土壤黏粒是影响SND最主要的因素.土壤碳氮密度特征也受到土壤微生物及其他环境因子的影响.随着土壤深度的增加土壤微环境发生改变,进而影响微生物群落及生物量,导致土壤碳氮密度差异,丁爽等[36 ] 在中国西南典型森林土壤的研究发现,微生物各类群生物量均在表层最高,随土壤深度的增加逐渐减少,进而影响土壤碳氮密度.此外,土壤含水量、土壤温度及pH等环境因子也影响土壤碳氮含量[37 ] ,这是因为不同植物群落不同土层的土壤含水量、温度和pH的不同,影响土壤微生物种类及其活性,进而影响土壤碳氮循环,最终导致土壤碳氮密度发生改变. ...
中国西南典型森林土壤微生物在不同土壤深度下的变化特征
1
2023
... ASP土壤碳氮密度最大,这是因为其群落生物量、物种多样性以及多年生植物优势度较高[33 ] .随着土壤中C∶N的升高,SCD增大,SND减小;随着土壤中黏粒含量的增加,SCD和SND都增大.BD与SCD显著负相关,这是因为BD增加土壤孔隙度减小,破坏大团聚体结构,使包裹于内部的有机碳暴露,易被微生物分解,从而减少土壤碳密度[34 -35 ] .C∶N、土壤黏粒含量和砂粒含量是影响SCD最主要的因素;N∶P、N∶K和土壤黏粒是影响SND最主要的因素.土壤碳氮密度特征也受到土壤微生物及其他环境因子的影响.随着土壤深度的增加土壤微环境发生改变,进而影响微生物群落及生物量,导致土壤碳氮密度差异,丁爽等[36 ] 在中国西南典型森林土壤的研究发现,微生物各类群生物量均在表层最高,随土壤深度的增加逐渐减少,进而影响土壤碳氮密度.此外,土壤含水量、土壤温度及pH等环境因子也影响土壤碳氮含量[37 ] ,这是因为不同植物群落不同土层的土壤含水量、温度和pH的不同,影响土壤微生物种类及其活性,进而影响土壤碳氮循环,最终导致土壤碳氮密度发生改变. ...
兰州北山不同植被土壤可培养微生物丰度变化特征及影响因素
1
2020
... ASP土壤碳氮密度最大,这是因为其群落生物量、物种多样性以及多年生植物优势度较高[33 ] .随着土壤中C∶N的升高,SCD增大,SND减小;随着土壤中黏粒含量的增加,SCD和SND都增大.BD与SCD显著负相关,这是因为BD增加土壤孔隙度减小,破坏大团聚体结构,使包裹于内部的有机碳暴露,易被微生物分解,从而减少土壤碳密度[34 -35 ] .C∶N、土壤黏粒含量和砂粒含量是影响SCD最主要的因素;N∶P、N∶K和土壤黏粒是影响SND最主要的因素.土壤碳氮密度特征也受到土壤微生物及其他环境因子的影响.随着土壤深度的增加土壤微环境发生改变,进而影响微生物群落及生物量,导致土壤碳氮密度差异,丁爽等[36 ] 在中国西南典型森林土壤的研究发现,微生物各类群生物量均在表层最高,随土壤深度的增加逐渐减少,进而影响土壤碳氮密度.此外,土壤含水量、土壤温度及pH等环境因子也影响土壤碳氮含量[37 ] ,这是因为不同植物群落不同土层的土壤含水量、温度和pH的不同,影响土壤微生物种类及其活性,进而影响土壤碳氮循环,最终导致土壤碳氮密度发生改变. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}