Climate Change 2021.The Physical Science Basis.Summary for The Policy Makers,Technical Summary and Frequently Asked Questions
2
2021
... 在全球气候变暖的背景下,干旱、半干旱地区极端气候事件发生的频率、强度和持续时间将会显著增加[1-2],使得更多地区的人类遭遇严重的生存危机.联合国政府间气候变化专门委员会(IPCC)的评估报告指出,全球增温4 ℃水平下,人类50%的居住区域将面临农业生态干旱的威胁,尤其是降水量分布频次低于10%的极端干旱事件将对中国北方生态脆弱区造成尤为深远的影响[3].未来20年,中国大部分区域干旱出现的频率和强度将逐渐增多[4].因此,频繁出现的极端干旱将会对经济社会发展、粮食生产、生态环境及人民生命财产安全等造成严重威胁.极端气候事件时空变化及其驱动因素研究已成为国内外学者关注的热点问题[5-6]. ...
... 生物多样性和生态系统功能的关系研究是当前备受生态学关注的热点[7-10].近年来,生态学家利用植物、动物、微生物开展了大量的野外观察、人工调控及理论模型研究,进一步验证了生物多样性与生态系统功能关系的相关假说[11-13].目前,在全球变化大背景下,降水的季节分配及降水模式的改变使得区域水分循环的关键过程发生改变[14],进而对生态系统的生物多样性[12,15-16]、生产力[17-18]、碳氮循环[19]、土壤微生物群落组成[20]等生态系统功能产生深远影响.经验证据表明,频繁的极端干旱事件可以导致群落中关键物种或常见物种重新排序、诱导优势物种发生变化、驱动向新的生态系统类型的转变[21],未来愈加严重的水文干旱也将导致脆弱生态系统碳汇功能降低[1].极端干旱不仅会影响生物多样性和生态系统多功能性,还可调节生物多样性与生态系统多功能性之间的关系[2,22].因此,厘清生物多样性与生态系统多功能性的作用关系不仅具有重大的科学意义,也可以为生态系统管理和保护提供重要的理论指导. ...
Contrasting drought sensitivity of Eurasian and North American grasslands
2
2025
... 在全球气候变暖的背景下,干旱、半干旱地区极端气候事件发生的频率、强度和持续时间将会显著增加[1-2],使得更多地区的人类遭遇严重的生存危机.联合国政府间气候变化专门委员会(IPCC)的评估报告指出,全球增温4 ℃水平下,人类50%的居住区域将面临农业生态干旱的威胁,尤其是降水量分布频次低于10%的极端干旱事件将对中国北方生态脆弱区造成尤为深远的影响[3].未来20年,中国大部分区域干旱出现的频率和强度将逐渐增多[4].因此,频繁出现的极端干旱将会对经济社会发展、粮食生产、生态环境及人民生命财产安全等造成严重威胁.极端气候事件时空变化及其驱动因素研究已成为国内外学者关注的热点问题[5-6]. ...
... 生物多样性和生态系统功能的关系研究是当前备受生态学关注的热点[7-10].近年来,生态学家利用植物、动物、微生物开展了大量的野外观察、人工调控及理论模型研究,进一步验证了生物多样性与生态系统功能关系的相关假说[11-13].目前,在全球变化大背景下,降水的季节分配及降水模式的改变使得区域水分循环的关键过程发生改变[14],进而对生态系统的生物多样性[12,15-16]、生产力[17-18]、碳氮循环[19]、土壤微生物群落组成[20]等生态系统功能产生深远影响.经验证据表明,频繁的极端干旱事件可以导致群落中关键物种或常见物种重新排序、诱导优势物种发生变化、驱动向新的生态系统类型的转变[21],未来愈加严重的水文干旱也将导致脆弱生态系统碳汇功能降低[1].极端干旱不仅会影响生物多样性和生态系统多功能性,还可调节生物多样性与生态系统多功能性之间的关系[2,22].因此,厘清生物多样性与生态系统多功能性的作用关系不仅具有重大的科学意义,也可以为生态系统管理和保护提供重要的理论指导. ...
IPCC AR6报告解读:极端天气气候事件变化
1
2021
... 在全球气候变暖的背景下,干旱、半干旱地区极端气候事件发生的频率、强度和持续时间将会显著增加[1-2],使得更多地区的人类遭遇严重的生存危机.联合国政府间气候变化专门委员会(IPCC)的评估报告指出,全球增温4 ℃水平下,人类50%的居住区域将面临农业生态干旱的威胁,尤其是降水量分布频次低于10%的极端干旱事件将对中国北方生态脆弱区造成尤为深远的影响[3].未来20年,中国大部分区域干旱出现的频率和强度将逐渐增多[4].因此,频繁出现的极端干旱将会对经济社会发展、粮食生产、生态环境及人民生命财产安全等造成严重威胁.极端气候事件时空变化及其驱动因素研究已成为国内外学者关注的热点问题[5-6]. ...
A projection of future changes in summer precipitation and monsoon in East Asia
1
2010
... 在全球气候变暖的背景下,干旱、半干旱地区极端气候事件发生的频率、强度和持续时间将会显著增加[1-2],使得更多地区的人类遭遇严重的生存危机.联合国政府间气候变化专门委员会(IPCC)的评估报告指出,全球增温4 ℃水平下,人类50%的居住区域将面临农业生态干旱的威胁,尤其是降水量分布频次低于10%的极端干旱事件将对中国北方生态脆弱区造成尤为深远的影响[3].未来20年,中国大部分区域干旱出现的频率和强度将逐渐增多[4].因此,频繁出现的极端干旱将会对经济社会发展、粮食生产、生态环境及人民生命财产安全等造成严重威胁.极端气候事件时空变化及其驱动因素研究已成为国内外学者关注的热点问题[5-6]. ...
Flood inundation assessment in the low-lying river basin considering extreme rainfall impacts and topographic vulnerability
1
2021
... 在全球气候变暖的背景下,干旱、半干旱地区极端气候事件发生的频率、强度和持续时间将会显著增加[1-2],使得更多地区的人类遭遇严重的生存危机.联合国政府间气候变化专门委员会(IPCC)的评估报告指出,全球增温4 ℃水平下,人类50%的居住区域将面临农业生态干旱的威胁,尤其是降水量分布频次低于10%的极端干旱事件将对中国北方生态脆弱区造成尤为深远的影响[3].未来20年,中国大部分区域干旱出现的频率和强度将逐渐增多[4].因此,频繁出现的极端干旱将会对经济社会发展、粮食生产、生态环境及人民生命财产安全等造成严重威胁.极端气候事件时空变化及其驱动因素研究已成为国内外学者关注的热点问题[5-6]. ...
Trends and zonal variability of extreme rainfall events over East Africa during 1960-2017
1
2021
... 在全球气候变暖的背景下,干旱、半干旱地区极端气候事件发生的频率、强度和持续时间将会显著增加[1-2],使得更多地区的人类遭遇严重的生存危机.联合国政府间气候变化专门委员会(IPCC)的评估报告指出,全球增温4 ℃水平下,人类50%的居住区域将面临农业生态干旱的威胁,尤其是降水量分布频次低于10%的极端干旱事件将对中国北方生态脆弱区造成尤为深远的影响[3].未来20年,中国大部分区域干旱出现的频率和强度将逐渐增多[4].因此,频繁出现的极端干旱将会对经济社会发展、粮食生产、生态环境及人民生命财产安全等造成严重威胁.极端气候事件时空变化及其驱动因素研究已成为国内外学者关注的热点问题[5-6]. ...
生物多样性与生态系统功能的实验研究
1
2013
... 生物多样性和生态系统功能的关系研究是当前备受生态学关注的热点[7-10].近年来,生态学家利用植物、动物、微生物开展了大量的野外观察、人工调控及理论模型研究,进一步验证了生物多样性与生态系统功能关系的相关假说[11-13].目前,在全球变化大背景下,降水的季节分配及降水模式的改变使得区域水分循环的关键过程发生改变[14],进而对生态系统的生物多样性[12,15-16]、生产力[17-18]、碳氮循环[19]、土壤微生物群落组成[20]等生态系统功能产生深远影响.经验证据表明,频繁的极端干旱事件可以导致群落中关键物种或常见物种重新排序、诱导优势物种发生变化、驱动向新的生态系统类型的转变[21],未来愈加严重的水文干旱也将导致脆弱生态系统碳汇功能降低[1].极端干旱不仅会影响生物多样性和生态系统多功能性,还可调节生物多样性与生态系统多功能性之间的关系[2,22].因此,厘清生物多样性与生态系统多功能性的作用关系不仅具有重大的科学意义,也可以为生态系统管理和保护提供重要的理论指导. ...
Above-and below-ground biodiversity jointly regulate temperate forest multifunctionality along a local-scale environmental gradient
0
2020
Building a bridge between biodiversity and ecosystem multifunctionality
1
2023
... 降雨模式变化首先会改变土壤含水量[96]和土壤净氮矿化速率[97],影响枯落物和细根分解[98],进而调整微生物群落结构、有机物分解和养分循环过程[99],最终制约土壤微生物多样性[64].区域尺度上的土壤微生物群落组成以及细菌、真菌、古菌和线虫等不同维度的多样性主要受降水模式和植被类型(荒漠、草甸和草原等)的驱动[100-101].例如降雨模式变化对林地和草地土壤微生物群落影响次序为生长季干旱>极端降雨>全年增雨>全年减雨[65].极端干旱条件更有利于耐旱性微生物如真菌、厚壁菌、放线菌和革兰氏阴性菌等的生长[102-103].长期干旱降低了草地生态系统土壤真菌的丰富度,延滞细菌群落的共现网络[104-105];改变了草地土壤真菌和细菌比例[9,106];降低了中国北方草原干旱和半干旱区土壤线虫丰富度[107].生长季干旱提高了林地和草地的土壤细菌α多样性和放线菌门相对丰度,改变了丰富和稀有细菌群落组成[65]. ...
Biodiversity loss reduces global terrestrial carbon storage
1
2024
... 生物多样性和生态系统功能的关系研究是当前备受生态学关注的热点[7-10].近年来,生态学家利用植物、动物、微生物开展了大量的野外观察、人工调控及理论模型研究,进一步验证了生物多样性与生态系统功能关系的相关假说[11-13].目前,在全球变化大背景下,降水的季节分配及降水模式的改变使得区域水分循环的关键过程发生改变[14],进而对生态系统的生物多样性[12,15-16]、生产力[17-18]、碳氮循环[19]、土壤微生物群落组成[20]等生态系统功能产生深远影响.经验证据表明,频繁的极端干旱事件可以导致群落中关键物种或常见物种重新排序、诱导优势物种发生变化、驱动向新的生态系统类型的转变[21],未来愈加严重的水文干旱也将导致脆弱生态系统碳汇功能降低[1].极端干旱不仅会影响生物多样性和生态系统多功能性,还可调节生物多样性与生态系统多功能性之间的关系[2,22].因此,厘清生物多样性与生态系统多功能性的作用关系不仅具有重大的科学意义,也可以为生态系统管理和保护提供重要的理论指导. ...
Lack of functional redundancy in the relationship between microbial diversity and ecosystem functioning
3
2016
... 生物多样性和生态系统功能的关系研究是当前备受生态学关注的热点[7-10].近年来,生态学家利用植物、动物、微生物开展了大量的野外观察、人工调控及理论模型研究,进一步验证了生物多样性与生态系统功能关系的相关假说[11-13].目前,在全球变化大背景下,降水的季节分配及降水模式的改变使得区域水分循环的关键过程发生改变[14],进而对生态系统的生物多样性[12,15-16]、生产力[17-18]、碳氮循环[19]、土壤微生物群落组成[20]等生态系统功能产生深远影响.经验证据表明,频繁的极端干旱事件可以导致群落中关键物种或常见物种重新排序、诱导优势物种发生变化、驱动向新的生态系统类型的转变[21],未来愈加严重的水文干旱也将导致脆弱生态系统碳汇功能降低[1].极端干旱不仅会影响生物多样性和生态系统多功能性,还可调节生物多样性与生态系统多功能性之间的关系[2,22].因此,厘清生物多样性与生态系统多功能性的作用关系不仅具有重大的科学意义,也可以为生态系统管理和保护提供重要的理论指导. ...
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
... [11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
Soil multifunctionality and drought resistance are determined by plant structural traits in restoring grassland
4
2018
... 生物多样性和生态系统功能的关系研究是当前备受生态学关注的热点[7-10].近年来,生态学家利用植物、动物、微生物开展了大量的野外观察、人工调控及理论模型研究,进一步验证了生物多样性与生态系统功能关系的相关假说[11-13].目前,在全球变化大背景下,降水的季节分配及降水模式的改变使得区域水分循环的关键过程发生改变[14],进而对生态系统的生物多样性[12,15-16]、生产力[17-18]、碳氮循环[19]、土壤微生物群落组成[20]等生态系统功能产生深远影响.经验证据表明,频繁的极端干旱事件可以导致群落中关键物种或常见物种重新排序、诱导优势物种发生变化、驱动向新的生态系统类型的转变[21],未来愈加严重的水文干旱也将导致脆弱生态系统碳汇功能降低[1].极端干旱不仅会影响生物多样性和生态系统多功能性,还可调节生物多样性与生态系统多功能性之间的关系[2,22].因此,厘清生物多样性与生态系统多功能性的作用关系不仅具有重大的科学意义,也可以为生态系统管理和保护提供重要的理论指导. ...
... 面对全球物种灭绝加快,针对生物多样性与生态系统多功能性关系的相关研究已经广泛开展,并总结出了许多假说和量化方法,例如“质量比假说”“阈值法”“平均值法”等.气候变化尤其是降水模式变化会改变植物和土壤微生物群落多样性与生态功能的耦合关系[16,23],降水可通过改变土壤理化性质等对生态系统多功能性产生直接的负效应[19],也可通过改变生物多样性,尤其是植物-土壤微生物多样性来间接调控生态系统多功能性[12,24].探究全球变化背景下生物多样性与生态系统多功能性作用关系及机理对生物多样性保护和生态系统功能评估有重要意义. ...
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
... 降水可以通过改变生物多样性影响生态系统功能[12],也可以通过改变土壤性质、植物光合速率、根系分泌物等影响生物量,从而改变生态系统功能[19].众所周知生物量是研究生态系统碳收支和碳循环的重要环节.生物量分配反映了地上资源(光和CO2)与地下资源(水和养分)之间的平衡,被广泛用作指示陆地生态系统碳循环对全球变化响应的关键指标.大量野外实地研究和文献综述表明,群落生物量和生产力与极端降水事件具有很强的相关性,例如极端干旱事件使得全球典型草地生产力降低了40%[108]、中国荒漠草原沙生针茅(S. caucasica)群落地上生物量降低了50%以上[73];改变了荒漠生态系统典型植物骆驼刺(Alhagi sparsifolia)的叶磷分配模式[38];降低了全世界范围内83个草地C3禾草、C4杂草的生物量[109]和温带草原生态系统[110]、半干旱沙质草地[111]的地上、地下生产力.季节性极端干旱对不同生态系统群落生物量也产生了一定影响,例如生长季初期的春季极端干旱和生长季中期的夏季极端干旱显著降低了温带草原生态系统和草甸草原的地上生物量[112-113]. ...
生物多样性与生态系统多功能性和多服务性的关系:回顾与展望
1
2021
... 生物多样性和生态系统功能的关系研究是当前备受生态学关注的热点[7-10].近年来,生态学家利用植物、动物、微生物开展了大量的野外观察、人工调控及理论模型研究,进一步验证了生物多样性与生态系统功能关系的相关假说[11-13].目前,在全球变化大背景下,降水的季节分配及降水模式的改变使得区域水分循环的关键过程发生改变[14],进而对生态系统的生物多样性[12,15-16]、生产力[17-18]、碳氮循环[19]、土壤微生物群落组成[20]等生态系统功能产生深远影响.经验证据表明,频繁的极端干旱事件可以导致群落中关键物种或常见物种重新排序、诱导优势物种发生变化、驱动向新的生态系统类型的转变[21],未来愈加严重的水文干旱也将导致脆弱生态系统碳汇功能降低[1].极端干旱不仅会影响生物多样性和生态系统多功能性,还可调节生物多样性与生态系统多功能性之间的关系[2,22].因此,厘清生物多样性与生态系统多功能性的作用关系不仅具有重大的科学意义,也可以为生态系统管理和保护提供重要的理论指导. ...
极端降水和极端干旱事件对草原生态系统的影响
1
2014
... 生物多样性和生态系统功能的关系研究是当前备受生态学关注的热点[7-10].近年来,生态学家利用植物、动物、微生物开展了大量的野外观察、人工调控及理论模型研究,进一步验证了生物多样性与生态系统功能关系的相关假说[11-13].目前,在全球变化大背景下,降水的季节分配及降水模式的改变使得区域水分循环的关键过程发生改变[14],进而对生态系统的生物多样性[12,15-16]、生产力[17-18]、碳氮循环[19]、土壤微生物群落组成[20]等生态系统功能产生深远影响.经验证据表明,频繁的极端干旱事件可以导致群落中关键物种或常见物种重新排序、诱导优势物种发生变化、驱动向新的生态系统类型的转变[21],未来愈加严重的水文干旱也将导致脆弱生态系统碳汇功能降低[1].极端干旱不仅会影响生物多样性和生态系统多功能性,还可调节生物多样性与生态系统多功能性之间的关系[2,22].因此,厘清生物多样性与生态系统多功能性的作用关系不仅具有重大的科学意义,也可以为生态系统管理和保护提供重要的理论指导. ...
气候变化对陆地生物多样性影响研究的若干进展
1
2008
... 生物多样性和生态系统功能的关系研究是当前备受生态学关注的热点[7-10].近年来,生态学家利用植物、动物、微生物开展了大量的野外观察、人工调控及理论模型研究,进一步验证了生物多样性与生态系统功能关系的相关假说[11-13].目前,在全球变化大背景下,降水的季节分配及降水模式的改变使得区域水分循环的关键过程发生改变[14],进而对生态系统的生物多样性[12,15-16]、生产力[17-18]、碳氮循环[19]、土壤微生物群落组成[20]等生态系统功能产生深远影响.经验证据表明,频繁的极端干旱事件可以导致群落中关键物种或常见物种重新排序、诱导优势物种发生变化、驱动向新的生态系统类型的转变[21],未来愈加严重的水文干旱也将导致脆弱生态系统碳汇功能降低[1].极端干旱不仅会影响生物多样性和生态系统多功能性,还可调节生物多样性与生态系统多功能性之间的关系[2,22].因此,厘清生物多样性与生态系统多功能性的作用关系不仅具有重大的科学意义,也可以为生态系统管理和保护提供重要的理论指导. ...
Precipitation preferences alter the relative importance of herbaceous plant diversity for multifunctionality in the drylands of China
5
2023
... 生物多样性和生态系统功能的关系研究是当前备受生态学关注的热点[7-10].近年来,生态学家利用植物、动物、微生物开展了大量的野外观察、人工调控及理论模型研究,进一步验证了生物多样性与生态系统功能关系的相关假说[11-13].目前,在全球变化大背景下,降水的季节分配及降水模式的改变使得区域水分循环的关键过程发生改变[14],进而对生态系统的生物多样性[12,15-16]、生产力[17-18]、碳氮循环[19]、土壤微生物群落组成[20]等生态系统功能产生深远影响.经验证据表明,频繁的极端干旱事件可以导致群落中关键物种或常见物种重新排序、诱导优势物种发生变化、驱动向新的生态系统类型的转变[21],未来愈加严重的水文干旱也将导致脆弱生态系统碳汇功能降低[1].极端干旱不仅会影响生物多样性和生态系统多功能性,还可调节生物多样性与生态系统多功能性之间的关系[2,22].因此,厘清生物多样性与生态系统多功能性的作用关系不仅具有重大的科学意义,也可以为生态系统管理和保护提供重要的理论指导. ...
... 面对全球物种灭绝加快,针对生物多样性与生态系统多功能性关系的相关研究已经广泛开展,并总结出了许多假说和量化方法,例如“质量比假说”“阈值法”“平均值法”等.气候变化尤其是降水模式变化会改变植物和土壤微生物群落多样性与生态功能的耦合关系[16,23],降水可通过改变土壤理化性质等对生态系统多功能性产生直接的负效应[19],也可通过改变生物多样性,尤其是植物-土壤微生物多样性来间接调控生态系统多功能性[12,24].探究全球变化背景下生物多样性与生态系统多功能性作用关系及机理对生物多样性保护和生态系统功能评估有重要意义. ...
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
... [16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
... [16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
Interpreting the effects of plant species diversity and genotypic diversity within a dominant species on above-and belowground overyielding
2
2021
... 生物多样性和生态系统功能的关系研究是当前备受生态学关注的热点[7-10].近年来,生态学家利用植物、动物、微生物开展了大量的野外观察、人工调控及理论模型研究,进一步验证了生物多样性与生态系统功能关系的相关假说[11-13].目前,在全球变化大背景下,降水的季节分配及降水模式的改变使得区域水分循环的关键过程发生改变[14],进而对生态系统的生物多样性[12,15-16]、生产力[17-18]、碳氮循环[19]、土壤微生物群落组成[20]等生态系统功能产生深远影响.经验证据表明,频繁的极端干旱事件可以导致群落中关键物种或常见物种重新排序、诱导优势物种发生变化、驱动向新的生态系统类型的转变[21],未来愈加严重的水文干旱也将导致脆弱生态系统碳汇功能降低[1].极端干旱不仅会影响生物多样性和生态系统多功能性,还可调节生物多样性与生态系统多功能性之间的关系[2,22].因此,厘清生物多样性与生态系统多功能性的作用关系不仅具有重大的科学意义,也可以为生态系统管理和保护提供重要的理论指导. ...
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
Plant species richness mediates the responses of microbial necromass carbon accumulation to climate aridity in alpine meadows
1
2025
... 生物多样性和生态系统功能的关系研究是当前备受生态学关注的热点[7-10].近年来,生态学家利用植物、动物、微生物开展了大量的野外观察、人工调控及理论模型研究,进一步验证了生物多样性与生态系统功能关系的相关假说[11-13].目前,在全球变化大背景下,降水的季节分配及降水模式的改变使得区域水分循环的关键过程发生改变[14],进而对生态系统的生物多样性[12,15-16]、生产力[17-18]、碳氮循环[19]、土壤微生物群落组成[20]等生态系统功能产生深远影响.经验证据表明,频繁的极端干旱事件可以导致群落中关键物种或常见物种重新排序、诱导优势物种发生变化、驱动向新的生态系统类型的转变[21],未来愈加严重的水文干旱也将导致脆弱生态系统碳汇功能降低[1].极端干旱不仅会影响生物多样性和生态系统多功能性,还可调节生物多样性与生态系统多功能性之间的关系[2,22].因此,厘清生物多样性与生态系统多功能性的作用关系不仅具有重大的科学意义,也可以为生态系统管理和保护提供重要的理论指导. ...
Plant functional β diversity is an important mediator of effects of aridity on soil multifunctionality
5
2020
... 生物多样性和生态系统功能的关系研究是当前备受生态学关注的热点[7-10].近年来,生态学家利用植物、动物、微生物开展了大量的野外观察、人工调控及理论模型研究,进一步验证了生物多样性与生态系统功能关系的相关假说[11-13].目前,在全球变化大背景下,降水的季节分配及降水模式的改变使得区域水分循环的关键过程发生改变[14],进而对生态系统的生物多样性[12,15-16]、生产力[17-18]、碳氮循环[19]、土壤微生物群落组成[20]等生态系统功能产生深远影响.经验证据表明,频繁的极端干旱事件可以导致群落中关键物种或常见物种重新排序、诱导优势物种发生变化、驱动向新的生态系统类型的转变[21],未来愈加严重的水文干旱也将导致脆弱生态系统碳汇功能降低[1].极端干旱不仅会影响生物多样性和生态系统多功能性,还可调节生物多样性与生态系统多功能性之间的关系[2,22].因此,厘清生物多样性与生态系统多功能性的作用关系不仅具有重大的科学意义,也可以为生态系统管理和保护提供重要的理论指导. ...
... 面对全球物种灭绝加快,针对生物多样性与生态系统多功能性关系的相关研究已经广泛开展,并总结出了许多假说和量化方法,例如“质量比假说”“阈值法”“平均值法”等.气候变化尤其是降水模式变化会改变植物和土壤微生物群落多样性与生态功能的耦合关系[16,23],降水可通过改变土壤理化性质等对生态系统多功能性产生直接的负效应[19],也可通过改变生物多样性,尤其是植物-土壤微生物多样性来间接调控生态系统多功能性[12,24].探究全球变化背景下生物多样性与生态系统多功能性作用关系及机理对生物多样性保护和生态系统功能评估有重要意义. ...
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
... [19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
... 降水可以通过改变生物多样性影响生态系统功能[12],也可以通过改变土壤性质、植物光合速率、根系分泌物等影响生物量,从而改变生态系统功能[19].众所周知生物量是研究生态系统碳收支和碳循环的重要环节.生物量分配反映了地上资源(光和CO2)与地下资源(水和养分)之间的平衡,被广泛用作指示陆地生态系统碳循环对全球变化响应的关键指标.大量野外实地研究和文献综述表明,群落生物量和生产力与极端降水事件具有很强的相关性,例如极端干旱事件使得全球典型草地生产力降低了40%[108]、中国荒漠草原沙生针茅(S. caucasica)群落地上生物量降低了50%以上[73];改变了荒漠生态系统典型植物骆驼刺(Alhagi sparsifolia)的叶磷分配模式[38];降低了全世界范围内83个草地C3禾草、C4杂草的生物量[109]和温带草原生态系统[110]、半干旱沙质草地[111]的地上、地下生产力.季节性极端干旱对不同生态系统群落生物量也产生了一定影响,例如生长季初期的春季极端干旱和生长季中期的夏季极端干旱显著降低了温带草原生态系统和草甸草原的地上生物量[112-113]. ...
Linking microbial communities to ecosystem functions:what we can learn from genotype-phenotype mapping in organisms
2
2020
... 生物多样性和生态系统功能的关系研究是当前备受生态学关注的热点[7-10].近年来,生态学家利用植物、动物、微生物开展了大量的野外观察、人工调控及理论模型研究,进一步验证了生物多样性与生态系统功能关系的相关假说[11-13].目前,在全球变化大背景下,降水的季节分配及降水模式的改变使得区域水分循环的关键过程发生改变[14],进而对生态系统的生物多样性[12,15-16]、生产力[17-18]、碳氮循环[19]、土壤微生物群落组成[20]等生态系统功能产生深远影响.经验证据表明,频繁的极端干旱事件可以导致群落中关键物种或常见物种重新排序、诱导优势物种发生变化、驱动向新的生态系统类型的转变[21],未来愈加严重的水文干旱也将导致脆弱生态系统碳汇功能降低[1].极端干旱不仅会影响生物多样性和生态系统多功能性,还可调节生物多样性与生态系统多功能性之间的关系[2,22].因此,厘清生物多样性与生态系统多功能性的作用关系不仅具有重大的科学意义,也可以为生态系统管理和保护提供重要的理论指导. ...
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
Transitions from grassland to savanna under drought through passive facilitation by grasses
1
2014
... 生物多样性和生态系统功能的关系研究是当前备受生态学关注的热点[7-10].近年来,生态学家利用植物、动物、微生物开展了大量的野外观察、人工调控及理论模型研究,进一步验证了生物多样性与生态系统功能关系的相关假说[11-13].目前,在全球变化大背景下,降水的季节分配及降水模式的改变使得区域水分循环的关键过程发生改变[14],进而对生态系统的生物多样性[12,15-16]、生产力[17-18]、碳氮循环[19]、土壤微生物群落组成[20]等生态系统功能产生深远影响.经验证据表明,频繁的极端干旱事件可以导致群落中关键物种或常见物种重新排序、诱导优势物种发生变化、驱动向新的生态系统类型的转变[21],未来愈加严重的水文干旱也将导致脆弱生态系统碳汇功能降低[1].极端干旱不仅会影响生物多样性和生态系统多功能性,还可调节生物多样性与生态系统多功能性之间的关系[2,22].因此,厘清生物多样性与生态系统多功能性的作用关系不仅具有重大的科学意义,也可以为生态系统管理和保护提供重要的理论指导. ...
The links between ecosystem multifunctionality and above-and belowground biodiversity are mediated by climate
2
2015
... 生物多样性和生态系统功能的关系研究是当前备受生态学关注的热点[7-10].近年来,生态学家利用植物、动物、微生物开展了大量的野外观察、人工调控及理论模型研究,进一步验证了生物多样性与生态系统功能关系的相关假说[11-13].目前,在全球变化大背景下,降水的季节分配及降水模式的改变使得区域水分循环的关键过程发生改变[14],进而对生态系统的生物多样性[12,15-16]、生产力[17-18]、碳氮循环[19]、土壤微生物群落组成[20]等生态系统功能产生深远影响.经验证据表明,频繁的极端干旱事件可以导致群落中关键物种或常见物种重新排序、诱导优势物种发生变化、驱动向新的生态系统类型的转变[21],未来愈加严重的水文干旱也将导致脆弱生态系统碳汇功能降低[1].极端干旱不仅会影响生物多样性和生态系统多功能性,还可调节生物多样性与生态系统多功能性之间的关系[2,22].因此,厘清生物多样性与生态系统多功能性的作用关系不仅具有重大的科学意义,也可以为生态系统管理和保护提供重要的理论指导. ...
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
Nutrient enrichment,biodiversity loss,and consequent declines in ecosystem productivity
1
2013
... 面对全球物种灭绝加快,针对生物多样性与生态系统多功能性关系的相关研究已经广泛开展,并总结出了许多假说和量化方法,例如“质量比假说”“阈值法”“平均值法”等.气候变化尤其是降水模式变化会改变植物和土壤微生物群落多样性与生态功能的耦合关系[16,23],降水可通过改变土壤理化性质等对生态系统多功能性产生直接的负效应[19],也可通过改变生物多样性,尤其是植物-土壤微生物多样性来间接调控生态系统多功能性[12,24].探究全球变化背景下生物多样性与生态系统多功能性作用关系及机理对生物多样性保护和生态系统功能评估有重要意义. ...
Sustainable land management enhances ecological and economic multifunctionality under ambient and future climate
2
2024
... 面对全球物种灭绝加快,针对生物多样性与生态系统多功能性关系的相关研究已经广泛开展,并总结出了许多假说和量化方法,例如“质量比假说”“阈值法”“平均值法”等.气候变化尤其是降水模式变化会改变植物和土壤微生物群落多样性与生态功能的耦合关系[16,23],降水可通过改变土壤理化性质等对生态系统多功能性产生直接的负效应[19],也可通过改变生物多样性,尤其是植物-土壤微生物多样性来间接调控生态系统多功能性[12,24].探究全球变化背景下生物多样性与生态系统多功能性作用关系及机理对生物多样性保护和生态系统功能评估有重要意义. ...
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
Plant traits and soil fertility mediate productivity losses under extreme drought in C3 grasslands
2
2021
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
... 干旱除了对群落生物量产生影响外,也会对生态系统碳循环、氮循环等产生不同程度的影响,进而影响生态系统的养分循环和水循环,对生态系统多功能性起关键作用[114-115].生长季极端干旱会减弱土壤微生物活动[103]、降低氮矿化速率[111]和凋落物生物量的碳氮密度[116].极端干旱会显著影响氮素的转化过程[117]、土壤碳稳定性以及土壤团聚体的碳封存功能[56]、土壤总碳和总氮含量[118],增加贝加尔针茅(S. baicalensis)草原植物叶片碳含量[25,119].极端干旱也能导致土壤动物功能群重组与效率失衡[120]:杂食性和腐食性类群减少[121],微生物互作网络受损,氮利用率低,养分循环阻滞,结构稳定性下降[122].土壤动物群落冗余度降低,抗干扰能力下降,植物病原菌繁殖加速,植物生产力显著降低[123].蚂蚁、蚯蚓、千足虫等清道夫数量减少,土壤沉积物、有机质垂直运输的能力变弱[124-125]. ...
Phylogenetic,functional,and taxonomic richness have both positive and negative effects on ecosystem multifunctionality
1
2019
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
Plant functional trait diversity and structural diversity co-underpin ecosystem multifunctionality in subtropical forests
0
2023
Nitrogen availability and plant functional composition modify biodiversity-multifunctionality relationships
1
2024
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
Aridity-driven shift in biodiversity-soil multifunctionality relationships
1
2021
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
Plant genetic diversity affects multiple trophic levels and trophic interactions
1
2022
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
Biodiversity drives ecosystem multifunctionality in sandy grassland?
4
2024
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
... [31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
... [31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
Effects of environmental factors on genetic diversity of Canagana microphylla in Horqin Sandy Land,Northeast China
1
2016
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
阿拉善高原灌木种的丰富度和多度格局对环境因子变化的响应:极端干旱荒漠地区灌木多样性保育的前提
1
2009
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
Higher biodiversity is required to sustain multiple ecosystem processes across temperature regimes
1
2015
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
不同放牧模式对贵南县高寒草甸优势种羊茅叶属性的影响
1
2021
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
Functional diversity enhances the resistance of ecosystem multifunctionality to aridity in Mediterranean drylands
1
2015
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
A graphical null model for scaling biodiversity-ecosystem functioning relationships
1
2021
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
Allocation of foliar-P fractions of Alhagi sparsifolia and its relationship with soil-P fractions and soil properties in a hyperarid desert ecosystem
2
2022
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
... 降水可以通过改变生物多样性影响生态系统功能[12],也可以通过改变土壤性质、植物光合速率、根系分泌物等影响生物量,从而改变生态系统功能[19].众所周知生物量是研究生态系统碳收支和碳循环的重要环节.生物量分配反映了地上资源(光和CO2)与地下资源(水和养分)之间的平衡,被广泛用作指示陆地生态系统碳循环对全球变化响应的关键指标.大量野外实地研究和文献综述表明,群落生物量和生产力与极端降水事件具有很强的相关性,例如极端干旱事件使得全球典型草地生产力降低了40%[108]、中国荒漠草原沙生针茅(S. caucasica)群落地上生物量降低了50%以上[73];改变了荒漠生态系统典型植物骆驼刺(Alhagi sparsifolia)的叶磷分配模式[38];降低了全世界范围内83个草地C3禾草、C4杂草的生物量[109]和温带草原生态系统[110]、半干旱沙质草地[111]的地上、地下生产力.季节性极端干旱对不同生态系统群落生物量也产生了一定影响,例如生长季初期的春季极端干旱和生长季中期的夏季极端干旱显著降低了温带草原生态系统和草甸草原的地上生物量[112-113]. ...
植物多样性对土壤有机碳及其稳定性影响的研究进展
1
2024
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
Several scales of biodiversity affect ecosystem multifunctionality
1
2013
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
Elevated ozone decreases the multifunctionality of belowground
1
2023
... 不同维度植物多样性(物种多样性、功能多样性、遗传多样性及其相互作用)与生态系统多功能性都呈现显著的作用关系[16,25].例如功能多样性对单个生态系统功能和生态系统多功能性变异具有很高的解释度和关联性[16],能有效驱动陆地生态系统多功能性[26-28];系统发育多样性与旱地生态系统多功能性以及单个碳、氮循环功能正相关[29];遗传多样性增加会改变群落生产力[30-31];沙质草地生态系统小叶锦鸡儿(Caragana microphylla)遗传多样性与土壤有效磷含量显著正相关[32];阿拉善高原灌木种多度与土壤有机质、全氮含量显著正相关[33].物种丰富度和植物功能多样性是调控草地和旱地生态系统多功能性的重要指标[16,34].个体大小(植株高度或侧向扩展)和叶片属性(叶片干物质含量和比叶面积)等植物性状是驱动高寒草甸生态系统[35]、内蒙古草原生态系统[19]和地中海地区旱地生态系统[36]多功能性响应干旱的主要因素.物种丰富度、均匀度等植物多样性指数与稀树草原[37]、典型草原生态系统多功能性之间的关系均具有尺度依赖性[31].内蒙古草原植物物种多样性、功能多样性和系统发育多样性在α和β尺度上均与生态系统多功能性呈显著正相关[19].内蒙古中东部典型草原物种多样性和系统发育多样性与土壤多功能性呈负相关关系[17].植物丰富度(α多样性)对沙质草地生态系统[31]、荒漠草原生态系统[38]、草原生态系统[39]单个生态系统功能和生态系统多功能性均有显著的正效应,而群落多样性(β多样性)和大尺度生物多样性(γ多样性)仅对生态系统多功能性具有显著的正作用[40].除地上部分外,植物根系的功能性状,如比根长、分枝比、真菌侵染率等,也会通过调控植物根系的生长及养分的吸收,影响生态系统的物质循环和能量流动,对草地生态系统多功能性维持具有重要的作用[12,41]. ...
Animating the carbon cycle
1
2014
... 土壤动物通过功能互补、网络缓冲和动态反馈三重机制,深度参与物质循环、能量流动、生境调控等核心生态过程,而其多样性特征直接决定草地生态系统多功能性的表达效率.在生物地球化学层面,土壤动物多样化的功能群生态位与土壤元素周转效率显著正相关.微生物捕食性线虫通过口器分化调节细菌与真菌群落多样性,进而与土壤碳氮分解、矿化等形成正向作用关系[42-43],食腐动物类(如蚯蚓)构建生物孔隙网络能显著提升土壤通气性与导水能力,进而与土壤氮素转化显著正相关[44-45],中小型土壤动物(如螨类和跳虫)会加快木质素和纤维素的降解速率,且当物种丰富度超过阈值时,其多样性与碳固存呈非线性相关关系[46],而且耐旱弹尾虫在干旱期可补偿螨类的分解功能,使得刈割后草地土壤动物多样性与植物生产力显著正相关[47].在生态网络维度上,由植物-土壤动物-微生物构成的互作关系网络[48-49]对生态系统多功能性的协同效应远超单一营养级,尤其在干旱区草地中,土壤动物功能多样性与生态系统多功能性的正向作用关系能增强生态系统的抵抗力和恢复力[50-52].与此同时,生态系统多功能性既是土壤动物功能的体现,也是维持其多样性的基础.一旦生态系统多功能性受损,环境因子变化会通过生物网络级联效应威胁整体多样性,比如调节功能(如气候调节、土壤稳定性维持)的破坏会降低微生物网络复杂性,引发线虫-螨类营养耦合效率锐减,使得动物多样性与生态系统多功能性呈负相关关系[49];供应功能(如食物与栖息地供给)缺失将导致土壤动物多样性降低[53];循环功能(如营养周转)效率的降低造成干旱草原等敏感区域中小型动物物种丰度下降,使得动物多样性与生态功能呈直接负相关.这些过程凸显了土壤动物多样性通过多维度功能服务维系生态系统多功能性的核心作用. ...
Synergistic effects of microbial and animal decomposers on plant and herbivore performance
1
2010
... 土壤动物通过功能互补、网络缓冲和动态反馈三重机制,深度参与物质循环、能量流动、生境调控等核心生态过程,而其多样性特征直接决定草地生态系统多功能性的表达效率.在生物地球化学层面,土壤动物多样化的功能群生态位与土壤元素周转效率显著正相关.微生物捕食性线虫通过口器分化调节细菌与真菌群落多样性,进而与土壤碳氮分解、矿化等形成正向作用关系[42-43],食腐动物类(如蚯蚓)构建生物孔隙网络能显著提升土壤通气性与导水能力,进而与土壤氮素转化显著正相关[44-45],中小型土壤动物(如螨类和跳虫)会加快木质素和纤维素的降解速率,且当物种丰富度超过阈值时,其多样性与碳固存呈非线性相关关系[46],而且耐旱弹尾虫在干旱期可补偿螨类的分解功能,使得刈割后草地土壤动物多样性与植物生产力显著正相关[47].在生态网络维度上,由植物-土壤动物-微生物构成的互作关系网络[48-49]对生态系统多功能性的协同效应远超单一营养级,尤其在干旱区草地中,土壤动物功能多样性与生态系统多功能性的正向作用关系能增强生态系统的抵抗力和恢复力[50-52].与此同时,生态系统多功能性既是土壤动物功能的体现,也是维持其多样性的基础.一旦生态系统多功能性受损,环境因子变化会通过生物网络级联效应威胁整体多样性,比如调节功能(如气候调节、土壤稳定性维持)的破坏会降低微生物网络复杂性,引发线虫-螨类营养耦合效率锐减,使得动物多样性与生态系统多功能性呈负相关关系[49];供应功能(如食物与栖息地供给)缺失将导致土壤动物多样性降低[53];循环功能(如营养周转)效率的降低造成干旱草原等敏感区域中小型动物物种丰度下降,使得动物多样性与生态功能呈直接负相关.这些过程凸显了土壤动物多样性通过多维度功能服务维系生态系统多功能性的核心作用. ...
How do earthworms affect decomposition of residues with different quality apart from fragmentation and incorporation?
1
2018
... 土壤动物通过功能互补、网络缓冲和动态反馈三重机制,深度参与物质循环、能量流动、生境调控等核心生态过程,而其多样性特征直接决定草地生态系统多功能性的表达效率.在生物地球化学层面,土壤动物多样化的功能群生态位与土壤元素周转效率显著正相关.微生物捕食性线虫通过口器分化调节细菌与真菌群落多样性,进而与土壤碳氮分解、矿化等形成正向作用关系[42-43],食腐动物类(如蚯蚓)构建生物孔隙网络能显著提升土壤通气性与导水能力,进而与土壤氮素转化显著正相关[44-45],中小型土壤动物(如螨类和跳虫)会加快木质素和纤维素的降解速率,且当物种丰富度超过阈值时,其多样性与碳固存呈非线性相关关系[46],而且耐旱弹尾虫在干旱期可补偿螨类的分解功能,使得刈割后草地土壤动物多样性与植物生产力显著正相关[47].在生态网络维度上,由植物-土壤动物-微生物构成的互作关系网络[48-49]对生态系统多功能性的协同效应远超单一营养级,尤其在干旱区草地中,土壤动物功能多样性与生态系统多功能性的正向作用关系能增强生态系统的抵抗力和恢复力[50-52].与此同时,生态系统多功能性既是土壤动物功能的体现,也是维持其多样性的基础.一旦生态系统多功能性受损,环境因子变化会通过生物网络级联效应威胁整体多样性,比如调节功能(如气候调节、土壤稳定性维持)的破坏会降低微生物网络复杂性,引发线虫-螨类营养耦合效率锐减,使得动物多样性与生态系统多功能性呈负相关关系[49];供应功能(如食物与栖息地供给)缺失将导致土壤动物多样性降低[53];循环功能(如营养周转)效率的降低造成干旱草原等敏感区域中小型动物物种丰度下降,使得动物多样性与生态功能呈直接负相关.这些过程凸显了土壤动物多样性通过多维度功能服务维系生态系统多功能性的核心作用. ...
Depth-differentiated,multivariate control of biopore number under different land-use practices
1
2022
... 土壤动物通过功能互补、网络缓冲和动态反馈三重机制,深度参与物质循环、能量流动、生境调控等核心生态过程,而其多样性特征直接决定草地生态系统多功能性的表达效率.在生物地球化学层面,土壤动物多样化的功能群生态位与土壤元素周转效率显著正相关.微生物捕食性线虫通过口器分化调节细菌与真菌群落多样性,进而与土壤碳氮分解、矿化等形成正向作用关系[42-43],食腐动物类(如蚯蚓)构建生物孔隙网络能显著提升土壤通气性与导水能力,进而与土壤氮素转化显著正相关[44-45],中小型土壤动物(如螨类和跳虫)会加快木质素和纤维素的降解速率,且当物种丰富度超过阈值时,其多样性与碳固存呈非线性相关关系[46],而且耐旱弹尾虫在干旱期可补偿螨类的分解功能,使得刈割后草地土壤动物多样性与植物生产力显著正相关[47].在生态网络维度上,由植物-土壤动物-微生物构成的互作关系网络[48-49]对生态系统多功能性的协同效应远超单一营养级,尤其在干旱区草地中,土壤动物功能多样性与生态系统多功能性的正向作用关系能增强生态系统的抵抗力和恢复力[50-52].与此同时,生态系统多功能性既是土壤动物功能的体现,也是维持其多样性的基础.一旦生态系统多功能性受损,环境因子变化会通过生物网络级联效应威胁整体多样性,比如调节功能(如气候调节、土壤稳定性维持)的破坏会降低微生物网络复杂性,引发线虫-螨类营养耦合效率锐减,使得动物多样性与生态系统多功能性呈负相关关系[49];供应功能(如食物与栖息地供给)缺失将导致土壤动物多样性降低[53];循环功能(如营养周转)效率的降低造成干旱草原等敏感区域中小型动物物种丰度下降,使得动物多样性与生态功能呈直接负相关.这些过程凸显了土壤动物多样性通过多维度功能服务维系生态系统多功能性的核心作用. ...
土壤动物多样性及其生态功能
1
2015
... 土壤动物通过功能互补、网络缓冲和动态反馈三重机制,深度参与物质循环、能量流动、生境调控等核心生态过程,而其多样性特征直接决定草地生态系统多功能性的表达效率.在生物地球化学层面,土壤动物多样化的功能群生态位与土壤元素周转效率显著正相关.微生物捕食性线虫通过口器分化调节细菌与真菌群落多样性,进而与土壤碳氮分解、矿化等形成正向作用关系[42-43],食腐动物类(如蚯蚓)构建生物孔隙网络能显著提升土壤通气性与导水能力,进而与土壤氮素转化显著正相关[44-45],中小型土壤动物(如螨类和跳虫)会加快木质素和纤维素的降解速率,且当物种丰富度超过阈值时,其多样性与碳固存呈非线性相关关系[46],而且耐旱弹尾虫在干旱期可补偿螨类的分解功能,使得刈割后草地土壤动物多样性与植物生产力显著正相关[47].在生态网络维度上,由植物-土壤动物-微生物构成的互作关系网络[48-49]对生态系统多功能性的协同效应远超单一营养级,尤其在干旱区草地中,土壤动物功能多样性与生态系统多功能性的正向作用关系能增强生态系统的抵抗力和恢复力[50-52].与此同时,生态系统多功能性既是土壤动物功能的体现,也是维持其多样性的基础.一旦生态系统多功能性受损,环境因子变化会通过生物网络级联效应威胁整体多样性,比如调节功能(如气候调节、土壤稳定性维持)的破坏会降低微生物网络复杂性,引发线虫-螨类营养耦合效率锐减,使得动物多样性与生态系统多功能性呈负相关关系[49];供应功能(如食物与栖息地供给)缺失将导致土壤动物多样性降低[53];循环功能(如营养周转)效率的降低造成干旱草原等敏感区域中小型动物物种丰度下降,使得动物多样性与生态功能呈直接负相关.这些过程凸显了土壤动物多样性通过多维度功能服务维系生态系统多功能性的核心作用. ...
Resource modification by ecosystem engineers generates hotspots of stream community assembly and ecosystem function
1
2023
... 土壤动物通过功能互补、网络缓冲和动态反馈三重机制,深度参与物质循环、能量流动、生境调控等核心生态过程,而其多样性特征直接决定草地生态系统多功能性的表达效率.在生物地球化学层面,土壤动物多样化的功能群生态位与土壤元素周转效率显著正相关.微生物捕食性线虫通过口器分化调节细菌与真菌群落多样性,进而与土壤碳氮分解、矿化等形成正向作用关系[42-43],食腐动物类(如蚯蚓)构建生物孔隙网络能显著提升土壤通气性与导水能力,进而与土壤氮素转化显著正相关[44-45],中小型土壤动物(如螨类和跳虫)会加快木质素和纤维素的降解速率,且当物种丰富度超过阈值时,其多样性与碳固存呈非线性相关关系[46],而且耐旱弹尾虫在干旱期可补偿螨类的分解功能,使得刈割后草地土壤动物多样性与植物生产力显著正相关[47].在生态网络维度上,由植物-土壤动物-微生物构成的互作关系网络[48-49]对生态系统多功能性的协同效应远超单一营养级,尤其在干旱区草地中,土壤动物功能多样性与生态系统多功能性的正向作用关系能增强生态系统的抵抗力和恢复力[50-52].与此同时,生态系统多功能性既是土壤动物功能的体现,也是维持其多样性的基础.一旦生态系统多功能性受损,环境因子变化会通过生物网络级联效应威胁整体多样性,比如调节功能(如气候调节、土壤稳定性维持)的破坏会降低微生物网络复杂性,引发线虫-螨类营养耦合效率锐减,使得动物多样性与生态系统多功能性呈负相关关系[49];供应功能(如食物与栖息地供给)缺失将导致土壤动物多样性降低[53];循环功能(如营养周转)效率的降低造成干旱草原等敏感区域中小型动物物种丰度下降,使得动物多样性与生态功能呈直接负相关.这些过程凸显了土壤动物多样性通过多维度功能服务维系生态系统多功能性的核心作用. ...
不同土壤线虫功能团对水稻生长及地上部植食者的影响
1
2019
... 土壤动物通过功能互补、网络缓冲和动态反馈三重机制,深度参与物质循环、能量流动、生境调控等核心生态过程,而其多样性特征直接决定草地生态系统多功能性的表达效率.在生物地球化学层面,土壤动物多样化的功能群生态位与土壤元素周转效率显著正相关.微生物捕食性线虫通过口器分化调节细菌与真菌群落多样性,进而与土壤碳氮分解、矿化等形成正向作用关系[42-43],食腐动物类(如蚯蚓)构建生物孔隙网络能显著提升土壤通气性与导水能力,进而与土壤氮素转化显著正相关[44-45],中小型土壤动物(如螨类和跳虫)会加快木质素和纤维素的降解速率,且当物种丰富度超过阈值时,其多样性与碳固存呈非线性相关关系[46],而且耐旱弹尾虫在干旱期可补偿螨类的分解功能,使得刈割后草地土壤动物多样性与植物生产力显著正相关[47].在生态网络维度上,由植物-土壤动物-微生物构成的互作关系网络[48-49]对生态系统多功能性的协同效应远超单一营养级,尤其在干旱区草地中,土壤动物功能多样性与生态系统多功能性的正向作用关系能增强生态系统的抵抗力和恢复力[50-52].与此同时,生态系统多功能性既是土壤动物功能的体现,也是维持其多样性的基础.一旦生态系统多功能性受损,环境因子变化会通过生物网络级联效应威胁整体多样性,比如调节功能(如气候调节、土壤稳定性维持)的破坏会降低微生物网络复杂性,引发线虫-螨类营养耦合效率锐减,使得动物多样性与生态系统多功能性呈负相关关系[49];供应功能(如食物与栖息地供给)缺失将导致土壤动物多样性降低[53];循环功能(如营养周转)效率的降低造成干旱草原等敏感区域中小型动物物种丰度下降,使得动物多样性与生态功能呈直接负相关.这些过程凸显了土壤动物多样性通过多维度功能服务维系生态系统多功能性的核心作用. ...
Upland grassland habitats and agri-environment schemes change soil microarthropod abundance
2
2021
... 土壤动物通过功能互补、网络缓冲和动态反馈三重机制,深度参与物质循环、能量流动、生境调控等核心生态过程,而其多样性特征直接决定草地生态系统多功能性的表达效率.在生物地球化学层面,土壤动物多样化的功能群生态位与土壤元素周转效率显著正相关.微生物捕食性线虫通过口器分化调节细菌与真菌群落多样性,进而与土壤碳氮分解、矿化等形成正向作用关系[42-43],食腐动物类(如蚯蚓)构建生物孔隙网络能显著提升土壤通气性与导水能力,进而与土壤氮素转化显著正相关[44-45],中小型土壤动物(如螨类和跳虫)会加快木质素和纤维素的降解速率,且当物种丰富度超过阈值时,其多样性与碳固存呈非线性相关关系[46],而且耐旱弹尾虫在干旱期可补偿螨类的分解功能,使得刈割后草地土壤动物多样性与植物生产力显著正相关[47].在生态网络维度上,由植物-土壤动物-微生物构成的互作关系网络[48-49]对生态系统多功能性的协同效应远超单一营养级,尤其在干旱区草地中,土壤动物功能多样性与生态系统多功能性的正向作用关系能增强生态系统的抵抗力和恢复力[50-52].与此同时,生态系统多功能性既是土壤动物功能的体现,也是维持其多样性的基础.一旦生态系统多功能性受损,环境因子变化会通过生物网络级联效应威胁整体多样性,比如调节功能(如气候调节、土壤稳定性维持)的破坏会降低微生物网络复杂性,引发线虫-螨类营养耦合效率锐减,使得动物多样性与生态系统多功能性呈负相关关系[49];供应功能(如食物与栖息地供给)缺失将导致土壤动物多样性降低[53];循环功能(如营养周转)效率的降低造成干旱草原等敏感区域中小型动物物种丰度下降,使得动物多样性与生态功能呈直接负相关.这些过程凸显了土壤动物多样性通过多维度功能服务维系生态系统多功能性的核心作用. ...
... [49];供应功能(如食物与栖息地供给)缺失将导致土壤动物多样性降低[53];循环功能(如营养周转)效率的降低造成干旱草原等敏感区域中小型动物物种丰度下降,使得动物多样性与生态功能呈直接负相关.这些过程凸显了土壤动物多样性通过多维度功能服务维系生态系统多功能性的核心作用. ...
Soil food web properties explain ecosystem services across European land use systems
1
2013
... 土壤动物通过功能互补、网络缓冲和动态反馈三重机制,深度参与物质循环、能量流动、生境调控等核心生态过程,而其多样性特征直接决定草地生态系统多功能性的表达效率.在生物地球化学层面,土壤动物多样化的功能群生态位与土壤元素周转效率显著正相关.微生物捕食性线虫通过口器分化调节细菌与真菌群落多样性,进而与土壤碳氮分解、矿化等形成正向作用关系[42-43],食腐动物类(如蚯蚓)构建生物孔隙网络能显著提升土壤通气性与导水能力,进而与土壤氮素转化显著正相关[44-45],中小型土壤动物(如螨类和跳虫)会加快木质素和纤维素的降解速率,且当物种丰富度超过阈值时,其多样性与碳固存呈非线性相关关系[46],而且耐旱弹尾虫在干旱期可补偿螨类的分解功能,使得刈割后草地土壤动物多样性与植物生产力显著正相关[47].在生态网络维度上,由植物-土壤动物-微生物构成的互作关系网络[48-49]对生态系统多功能性的协同效应远超单一营养级,尤其在干旱区草地中,土壤动物功能多样性与生态系统多功能性的正向作用关系能增强生态系统的抵抗力和恢复力[50-52].与此同时,生态系统多功能性既是土壤动物功能的体现,也是维持其多样性的基础.一旦生态系统多功能性受损,环境因子变化会通过生物网络级联效应威胁整体多样性,比如调节功能(如气候调节、土壤稳定性维持)的破坏会降低微生物网络复杂性,引发线虫-螨类营养耦合效率锐减,使得动物多样性与生态系统多功能性呈负相关关系[49];供应功能(如食物与栖息地供给)缺失将导致土壤动物多样性降低[53];循环功能(如营养周转)效率的降低造成干旱草原等敏感区域中小型动物物种丰度下降,使得动物多样性与生态功能呈直接负相关.这些过程凸显了土壤动物多样性通过多维度功能服务维系生态系统多功能性的核心作用. ...
生态系统稳定性及其与生物多样性的关系
0
2021
土壤动物多样性的地理分布及其生态功能研究进展
1
2022
... 土壤动物通过功能互补、网络缓冲和动态反馈三重机制,深度参与物质循环、能量流动、生境调控等核心生态过程,而其多样性特征直接决定草地生态系统多功能性的表达效率.在生物地球化学层面,土壤动物多样化的功能群生态位与土壤元素周转效率显著正相关.微生物捕食性线虫通过口器分化调节细菌与真菌群落多样性,进而与土壤碳氮分解、矿化等形成正向作用关系[42-43],食腐动物类(如蚯蚓)构建生物孔隙网络能显著提升土壤通气性与导水能力,进而与土壤氮素转化显著正相关[44-45],中小型土壤动物(如螨类和跳虫)会加快木质素和纤维素的降解速率,且当物种丰富度超过阈值时,其多样性与碳固存呈非线性相关关系[46],而且耐旱弹尾虫在干旱期可补偿螨类的分解功能,使得刈割后草地土壤动物多样性与植物生产力显著正相关[47].在生态网络维度上,由植物-土壤动物-微生物构成的互作关系网络[48-49]对生态系统多功能性的协同效应远超单一营养级,尤其在干旱区草地中,土壤动物功能多样性与生态系统多功能性的正向作用关系能增强生态系统的抵抗力和恢复力[50-52].与此同时,生态系统多功能性既是土壤动物功能的体现,也是维持其多样性的基础.一旦生态系统多功能性受损,环境因子变化会通过生物网络级联效应威胁整体多样性,比如调节功能(如气候调节、土壤稳定性维持)的破坏会降低微生物网络复杂性,引发线虫-螨类营养耦合效率锐减,使得动物多样性与生态系统多功能性呈负相关关系[49];供应功能(如食物与栖息地供给)缺失将导致土壤动物多样性降低[53];循环功能(如营养周转)效率的降低造成干旱草原等敏感区域中小型动物物种丰度下降,使得动物多样性与生态功能呈直接负相关.这些过程凸显了土壤动物多样性通过多维度功能服务维系生态系统多功能性的核心作用. ...
A closer look at the functions behind ecosystem multifunctionality:a review
1
2021
... 土壤动物通过功能互补、网络缓冲和动态反馈三重机制,深度参与物质循环、能量流动、生境调控等核心生态过程,而其多样性特征直接决定草地生态系统多功能性的表达效率.在生物地球化学层面,土壤动物多样化的功能群生态位与土壤元素周转效率显著正相关.微生物捕食性线虫通过口器分化调节细菌与真菌群落多样性,进而与土壤碳氮分解、矿化等形成正向作用关系[42-43],食腐动物类(如蚯蚓)构建生物孔隙网络能显著提升土壤通气性与导水能力,进而与土壤氮素转化显著正相关[44-45],中小型土壤动物(如螨类和跳虫)会加快木质素和纤维素的降解速率,且当物种丰富度超过阈值时,其多样性与碳固存呈非线性相关关系[46],而且耐旱弹尾虫在干旱期可补偿螨类的分解功能,使得刈割后草地土壤动物多样性与植物生产力显著正相关[47].在生态网络维度上,由植物-土壤动物-微生物构成的互作关系网络[48-49]对生态系统多功能性的协同效应远超单一营养级,尤其在干旱区草地中,土壤动物功能多样性与生态系统多功能性的正向作用关系能增强生态系统的抵抗力和恢复力[50-52].与此同时,生态系统多功能性既是土壤动物功能的体现,也是维持其多样性的基础.一旦生态系统多功能性受损,环境因子变化会通过生物网络级联效应威胁整体多样性,比如调节功能(如气候调节、土壤稳定性维持)的破坏会降低微生物网络复杂性,引发线虫-螨类营养耦合效率锐减,使得动物多样性与生态系统多功能性呈负相关关系[49];供应功能(如食物与栖息地供给)缺失将导致土壤动物多样性降低[53];循环功能(如营养周转)效率的降低造成干旱草原等敏感区域中小型动物物种丰度下降,使得动物多样性与生态功能呈直接负相关.这些过程凸显了土壤动物多样性通过多维度功能服务维系生态系统多功能性的核心作用. ...
Fungal-bacterial diversity and microbiome complexity predict ecosystem functioning
1
2019
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
Soil microbial network complexity predicts ecosystem function along elevation gradients on the Tibetan Plateau
3
2022
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
... [55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
... ,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
Geographical,climatic,and soil factors control the altitudinal pattern of rhizosphere microbial diversity and its driving effect on root zone soil multifunctionality in mountain ecosystems
4
2023
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
... [56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
... [56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
... 干旱除了对群落生物量产生影响外,也会对生态系统碳循环、氮循环等产生不同程度的影响,进而影响生态系统的养分循环和水循环,对生态系统多功能性起关键作用[114-115].生长季极端干旱会减弱土壤微生物活动[103]、降低氮矿化速率[111]和凋落物生物量的碳氮密度[116].极端干旱会显著影响氮素的转化过程[117]、土壤碳稳定性以及土壤团聚体的碳封存功能[56]、土壤总碳和总氮含量[118],增加贝加尔针茅(S. baicalensis)草原植物叶片碳含量[25,119].极端干旱也能导致土壤动物功能群重组与效率失衡[120]:杂食性和腐食性类群减少[121],微生物互作网络受损,氮利用率低,养分循环阻滞,结构稳定性下降[122].土壤动物群落冗余度降低,抗干扰能力下降,植物病原菌繁殖加速,植物生产力显著降低[123].蚂蚁、蚯蚓、千足虫等清道夫数量减少,土壤沉积物、有机质垂直运输的能力变弱[124-125]. ...
Fungal community determines soil multifunctionality during vegetation restoration in metallic tailing reservoir
2
2024
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
... [57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
The persistence of bacterial diversity and ecosystem multifunctionality along a disturbance intensity gradient in karst soil
1
2020
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
The loss of above-and belowground biodiversity in degraded grasslands drives the decline of ecosystem multifunctionality
1
2022
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
Nutrient-induced acidification modulates soil biodiversity-function relationships
1
2024
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
退化高寒草甸土壤微生物及酶活性特征
1
2017
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
青藏高原高寒草甸土壤生物多样性对土壤生态系统功能的影响
1
2016
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
Soil microbial responses to drought and exotic plants shift carbon metabolism
1
2019
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
Soil microbial diversity and network complexity drive the ecosystem multifunctionality of temperate grasslands under changing precipitation
3
2024
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
... [64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
... 降雨模式变化首先会改变土壤含水量[96]和土壤净氮矿化速率[97],影响枯落物和细根分解[98],进而调整微生物群落结构、有机物分解和养分循环过程[99],最终制约土壤微生物多样性[64].区域尺度上的土壤微生物群落组成以及细菌、真菌、古菌和线虫等不同维度的多样性主要受降水模式和植被类型(荒漠、草甸和草原等)的驱动[100-101].例如降雨模式变化对林地和草地土壤微生物群落影响次序为生长季干旱>极端降雨>全年增雨>全年减雨[65].极端干旱条件更有利于耐旱性微生物如真菌、厚壁菌、放线菌和革兰氏阴性菌等的生长[102-103].长期干旱降低了草地生态系统土壤真菌的丰富度,延滞细菌群落的共现网络[104-105];改变了草地土壤真菌和细菌比例[9,106];降低了中国北方草原干旱和半干旱区土壤线虫丰富度[107].生长季干旱提高了林地和草地的土壤细菌α多样性和放线菌门相对丰度,改变了丰富和稀有细菌群落组成[65]. ...
陕西关中地区降雨模式对土壤微生物群落和生态多功能性的影响机制
3
2021
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
... 降雨模式变化首先会改变土壤含水量[96]和土壤净氮矿化速率[97],影响枯落物和细根分解[98],进而调整微生物群落结构、有机物分解和养分循环过程[99],最终制约土壤微生物多样性[64].区域尺度上的土壤微生物群落组成以及细菌、真菌、古菌和线虫等不同维度的多样性主要受降水模式和植被类型(荒漠、草甸和草原等)的驱动[100-101].例如降雨模式变化对林地和草地土壤微生物群落影响次序为生长季干旱>极端降雨>全年增雨>全年减雨[65].极端干旱条件更有利于耐旱性微生物如真菌、厚壁菌、放线菌和革兰氏阴性菌等的生长[102-103].长期干旱降低了草地生态系统土壤真菌的丰富度,延滞细菌群落的共现网络[104-105];改变了草地土壤真菌和细菌比例[9,106];降低了中国北方草原干旱和半干旱区土壤线虫丰富度[107].生长季干旱提高了林地和草地的土壤细菌α多样性和放线菌门相对丰度,改变了丰富和稀有细菌群落组成[65]. ...
... [65]. ...
Towards a multidimensional view of biodiversity and ecosystem functioning in a changing world
1
2020
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
Vegetation biomass and soil moisture coregulate bacterial community succession under altered precipitation regimes in a desert steppe in northwestern China
1
2019
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
樟树和马尾松人工林土壤酶活性和微生物群落对干季和湿季的响应
1
2022
... 土壤微生物多样性在维持土壤健康、提供多种功能和服务方面发挥着重要作用[24],比如土壤微生物能够驱动陆地生态系统初级生产力、有机质分解和养分循环等生态系统多功能性[11,20,54].不同生态系统土壤微生物多样性与生态系统多功能性之间相关关系存在差异[11,55-56],土壤细菌、真菌和古菌群落在驱动生态多功能性方面发挥着关键作用[22,57].例如山地生态系统、高寒草地、喀斯特生态修复区土壤细菌群落α多样性与土壤生态多功能性显著正相关[56,58-59].有报道指出,土壤微生物多样性改变了退化植被修复过程中生态系统多功能性,比如土壤真菌群落多样性显著提高了矿山植被修复过程中土壤多功能性[57];细菌群落多样性提升了红树林恢复过程中土壤多功能性[60];不同退化程度高寒草地土壤Shannon指数和Simpson指数制约了土壤脲酶活性、降低了土壤微生物量氮含量[61]、增加了植物生物量,使得土壤微生物多样性与植物地上-地下生物量正相关[62]、并间接驱动生态系统多功能性[55].不同草地类型土壤细菌多样性与生态系统多功能性关系存在差异,比如在典型草原二者显著负相关,而在高寒草地显著正相关[56].全球变化背景下,土壤微生物群落对降水的反应主要通过产生更小、更脆弱的微生物量碳库,从而增强了土壤异养呼吸活性[63],进而影响生态系统多功能性[64].例如干旱会降低典型草原细菌和真菌多样性,进而改变生态系统功能指标[64];生长季干旱时林地和草地土壤细菌群落组成对生态多功能性具有直接的显著正向影响,极端干旱时则表现为土壤真菌群落占主导地位[65].土壤理化性质与细菌、真菌群落组成和多样性也有显著相关性[31,55-66],例如土壤理化性质是降雨模式变化过程中土壤微生物群落改变的资源影响因素[67];土壤全磷是调控人工林地土壤微生物群落结构的主要因素[68]. ...
Will extreme climatic events facilitate biological invasions?
1
2012
... 极端降水事件通过创造“入侵窗口”,增加外来物种入侵群落的可能性,导致群落组成和结构发生明显变化[69].但由于降水模式、群落类型、生态系统类型等不同,植物物种多样性、功能多样性和遗传多样性均呈现不同的响应方式.如长期极端干旱降低了原生草地、灌木丛和灌-草丛的物种丰富度[70];显著降低了非洲大草原和沙质草地物种优势度和多样性[71-72];促使草原优势种发生更替、群落结构单一化[71].生长季极端干旱和生长中期干旱降低了荒漠草原群落的物种多样性指数和丰富度指数[73].生长季降水量和降水格局显著影响了中国新疆和吉尔吉斯斯坦鸭茅(Dactylis glomerata)种群的遗传多样性[74];与维管植物期望杂合度、观测杂合度相关性最高[75];对内蒙古短花针茅(Stipa breviflora)[76]、金发草(Pogonatherum paniceum)[77]和差不嘎蒿(Artemisia halodendron)[78]遗传多样性没有显著影响.极端降水变化对不同生活型植物的叶功能性状影响趋势不同[79];显著提高了荒漠草原的优势物种比叶面积、叶氮、叶磷和叶干物质含量[80].极端干旱发生时,植物通过调整比根长和叶片、根系养分含量来提高水分和养分的吸收能力,保障根系生长的物质来源[81]. ...
Regional vegetation die-off in response to global-change-type drought
1
2005
... 极端降水事件通过创造“入侵窗口”,增加外来物种入侵群落的可能性,导致群落组成和结构发生明显变化[69].但由于降水模式、群落类型、生态系统类型等不同,植物物种多样性、功能多样性和遗传多样性均呈现不同的响应方式.如长期极端干旱降低了原生草地、灌木丛和灌-草丛的物种丰富度[70];显著降低了非洲大草原和沙质草地物种优势度和多样性[71-72];促使草原优势种发生更替、群落结构单一化[71].生长季极端干旱和生长中期干旱降低了荒漠草原群落的物种多样性指数和丰富度指数[73].生长季降水量和降水格局显著影响了中国新疆和吉尔吉斯斯坦鸭茅(Dactylis glomerata)种群的遗传多样性[74];与维管植物期望杂合度、观测杂合度相关性最高[75];对内蒙古短花针茅(Stipa breviflora)[76]、金发草(Pogonatherum paniceum)[77]和差不嘎蒿(Artemisia halodendron)[78]遗传多样性没有显著影响.极端降水变化对不同生活型植物的叶功能性状影响趋势不同[79];显著提高了荒漠草原的优势物种比叶面积、叶氮、叶磷和叶干物质含量[80].极端干旱发生时,植物通过调整比根长和叶片、根系养分含量来提高水分和养分的吸收能力,保障根系生长的物质来源[81]. ...
Resistance and resilience of a grassland ecosystem to climate extremes
2
2014
... 极端降水事件通过创造“入侵窗口”,增加外来物种入侵群落的可能性,导致群落组成和结构发生明显变化[69].但由于降水模式、群落类型、生态系统类型等不同,植物物种多样性、功能多样性和遗传多样性均呈现不同的响应方式.如长期极端干旱降低了原生草地、灌木丛和灌-草丛的物种丰富度[70];显著降低了非洲大草原和沙质草地物种优势度和多样性[71-72];促使草原优势种发生更替、群落结构单一化[71].生长季极端干旱和生长中期干旱降低了荒漠草原群落的物种多样性指数和丰富度指数[73].生长季降水量和降水格局显著影响了中国新疆和吉尔吉斯斯坦鸭茅(Dactylis glomerata)种群的遗传多样性[74];与维管植物期望杂合度、观测杂合度相关性最高[75];对内蒙古短花针茅(Stipa breviflora)[76]、金发草(Pogonatherum paniceum)[77]和差不嘎蒿(Artemisia halodendron)[78]遗传多样性没有显著影响.极端降水变化对不同生活型植物的叶功能性状影响趋势不同[79];显著提高了荒漠草原的优势物种比叶面积、叶氮、叶磷和叶干物质含量[80].极端干旱发生时,植物通过调整比根长和叶片、根系养分含量来提高水分和养分的吸收能力,保障根系生长的物质来源[81]. ...
... [71].生长季极端干旱和生长中期干旱降低了荒漠草原群落的物种多样性指数和丰富度指数[73].生长季降水量和降水格局显著影响了中国新疆和吉尔吉斯斯坦鸭茅(Dactylis glomerata)种群的遗传多样性[74];与维管植物期望杂合度、观测杂合度相关性最高[75];对内蒙古短花针茅(Stipa breviflora)[76]、金发草(Pogonatherum paniceum)[77]和差不嘎蒿(Artemisia halodendron)[78]遗传多样性没有显著影响.极端降水变化对不同生活型植物的叶功能性状影响趋势不同[79];显著提高了荒漠草原的优势物种比叶面积、叶氮、叶磷和叶干物质含量[80].极端干旱发生时,植物通过调整比根长和叶片、根系养分含量来提高水分和养分的吸收能力,保障根系生长的物质来源[81]. ...
半干旱沙地草本植物群落特征对短期降水变化的响应
1
2020
... 极端降水事件通过创造“入侵窗口”,增加外来物种入侵群落的可能性,导致群落组成和结构发生明显变化[69].但由于降水模式、群落类型、生态系统类型等不同,植物物种多样性、功能多样性和遗传多样性均呈现不同的响应方式.如长期极端干旱降低了原生草地、灌木丛和灌-草丛的物种丰富度[70];显著降低了非洲大草原和沙质草地物种优势度和多样性[71-72];促使草原优势种发生更替、群落结构单一化[71].生长季极端干旱和生长中期干旱降低了荒漠草原群落的物种多样性指数和丰富度指数[73].生长季降水量和降水格局显著影响了中国新疆和吉尔吉斯斯坦鸭茅(Dactylis glomerata)种群的遗传多样性[74];与维管植物期望杂合度、观测杂合度相关性最高[75];对内蒙古短花针茅(Stipa breviflora)[76]、金发草(Pogonatherum paniceum)[77]和差不嘎蒿(Artemisia halodendron)[78]遗传多样性没有显著影响.极端降水变化对不同生活型植物的叶功能性状影响趋势不同[79];显著提高了荒漠草原的优势物种比叶面积、叶氮、叶磷和叶干物质含量[80].极端干旱发生时,植物通过调整比根长和叶片、根系养分含量来提高水分和养分的吸收能力,保障根系生长的物质来源[81]. ...
极端干旱对荒漠草原群落物种多样性和地上生物量碳氮的影响
2
2019
... 极端降水事件通过创造“入侵窗口”,增加外来物种入侵群落的可能性,导致群落组成和结构发生明显变化[69].但由于降水模式、群落类型、生态系统类型等不同,植物物种多样性、功能多样性和遗传多样性均呈现不同的响应方式.如长期极端干旱降低了原生草地、灌木丛和灌-草丛的物种丰富度[70];显著降低了非洲大草原和沙质草地物种优势度和多样性[71-72];促使草原优势种发生更替、群落结构单一化[71].生长季极端干旱和生长中期干旱降低了荒漠草原群落的物种多样性指数和丰富度指数[73].生长季降水量和降水格局显著影响了中国新疆和吉尔吉斯斯坦鸭茅(Dactylis glomerata)种群的遗传多样性[74];与维管植物期望杂合度、观测杂合度相关性最高[75];对内蒙古短花针茅(Stipa breviflora)[76]、金发草(Pogonatherum paniceum)[77]和差不嘎蒿(Artemisia halodendron)[78]遗传多样性没有显著影响.极端降水变化对不同生活型植物的叶功能性状影响趋势不同[79];显著提高了荒漠草原的优势物种比叶面积、叶氮、叶磷和叶干物质含量[80].极端干旱发生时,植物通过调整比根长和叶片、根系养分含量来提高水分和养分的吸收能力,保障根系生长的物质来源[81]. ...
... 降水可以通过改变生物多样性影响生态系统功能[12],也可以通过改变土壤性质、植物光合速率、根系分泌物等影响生物量,从而改变生态系统功能[19].众所周知生物量是研究生态系统碳收支和碳循环的重要环节.生物量分配反映了地上资源(光和CO2)与地下资源(水和养分)之间的平衡,被广泛用作指示陆地生态系统碳循环对全球变化响应的关键指标.大量野外实地研究和文献综述表明,群落生物量和生产力与极端降水事件具有很强的相关性,例如极端干旱事件使得全球典型草地生产力降低了40%[108]、中国荒漠草原沙生针茅(S. caucasica)群落地上生物量降低了50%以上[73];改变了荒漠生态系统典型植物骆驼刺(Alhagi sparsifolia)的叶磷分配模式[38];降低了全世界范围内83个草地C3禾草、C4杂草的生物量[109]和温带草原生态系统[110]、半干旱沙质草地[111]的地上、地下生产力.季节性极端干旱对不同生态系统群落生物量也产生了一定影响,例如生长季初期的春季极端干旱和生长季中期的夏季极端干旱显著降低了温带草原生态系统和草甸草原的地上生物量[112-113]. ...
鸭茅野生种质遗传多样性的AFLP分析
1
2017
... 极端降水事件通过创造“入侵窗口”,增加外来物种入侵群落的可能性,导致群落组成和结构发生明显变化[69].但由于降水模式、群落类型、生态系统类型等不同,植物物种多样性、功能多样性和遗传多样性均呈现不同的响应方式.如长期极端干旱降低了原生草地、灌木丛和灌-草丛的物种丰富度[70];显著降低了非洲大草原和沙质草地物种优势度和多样性[71-72];促使草原优势种发生更替、群落结构单一化[71].生长季极端干旱和生长中期干旱降低了荒漠草原群落的物种多样性指数和丰富度指数[73].生长季降水量和降水格局显著影响了中国新疆和吉尔吉斯斯坦鸭茅(Dactylis glomerata)种群的遗传多样性[74];与维管植物期望杂合度、观测杂合度相关性最高[75];对内蒙古短花针茅(Stipa breviflora)[76]、金发草(Pogonatherum paniceum)[77]和差不嘎蒿(Artemisia halodendron)[78]遗传多样性没有显著影响.极端降水变化对不同生活型植物的叶功能性状影响趋势不同[79];显著提高了荒漠草原的优势物种比叶面积、叶氮、叶磷和叶干物质含量[80].极端干旱发生时,植物通过调整比根长和叶片、根系养分含量来提高水分和养分的吸收能力,保障根系生长的物质来源[81]. ...
Relationships between genetic diversity of vascular plant species and climate factors
1
2018
... 极端降水事件通过创造“入侵窗口”,增加外来物种入侵群落的可能性,导致群落组成和结构发生明显变化[69].但由于降水模式、群落类型、生态系统类型等不同,植物物种多样性、功能多样性和遗传多样性均呈现不同的响应方式.如长期极端干旱降低了原生草地、灌木丛和灌-草丛的物种丰富度[70];显著降低了非洲大草原和沙质草地物种优势度和多样性[71-72];促使草原优势种发生更替、群落结构单一化[71].生长季极端干旱和生长中期干旱降低了荒漠草原群落的物种多样性指数和丰富度指数[73].生长季降水量和降水格局显著影响了中国新疆和吉尔吉斯斯坦鸭茅(Dactylis glomerata)种群的遗传多样性[74];与维管植物期望杂合度、观测杂合度相关性最高[75];对内蒙古短花针茅(Stipa breviflora)[76]、金发草(Pogonatherum paniceum)[77]和差不嘎蒿(Artemisia halodendron)[78]遗传多样性没有显著影响.极端降水变化对不同生活型植物的叶功能性状影响趋势不同[79];显著提高了荒漠草原的优势物种比叶面积、叶氮、叶磷和叶干物质含量[80].极端干旱发生时,植物通过调整比根长和叶片、根系养分含量来提高水分和养分的吸收能力,保障根系生长的物质来源[81]. ...
内蒙古地区短花针茅(Stipa breviflora)种群遗传多样性
1
2008
... 极端降水事件通过创造“入侵窗口”,增加外来物种入侵群落的可能性,导致群落组成和结构发生明显变化[69].但由于降水模式、群落类型、生态系统类型等不同,植物物种多样性、功能多样性和遗传多样性均呈现不同的响应方式.如长期极端干旱降低了原生草地、灌木丛和灌-草丛的物种丰富度[70];显著降低了非洲大草原和沙质草地物种优势度和多样性[71-72];促使草原优势种发生更替、群落结构单一化[71].生长季极端干旱和生长中期干旱降低了荒漠草原群落的物种多样性指数和丰富度指数[73].生长季降水量和降水格局显著影响了中国新疆和吉尔吉斯斯坦鸭茅(Dactylis glomerata)种群的遗传多样性[74];与维管植物期望杂合度、观测杂合度相关性最高[75];对内蒙古短花针茅(Stipa breviflora)[76]、金发草(Pogonatherum paniceum)[77]和差不嘎蒿(Artemisia halodendron)[78]遗传多样性没有显著影响.极端降水变化对不同生活型植物的叶功能性状影响趋势不同[79];显著提高了荒漠草原的优势物种比叶面积、叶氮、叶磷和叶干物质含量[80].极端干旱发生时,植物通过调整比根长和叶片、根系养分含量来提高水分和养分的吸收能力,保障根系生长的物质来源[81]. ...
环境因子对岩生植物金发草遗传多样性的影响
1
2006
... 极端降水事件通过创造“入侵窗口”,增加外来物种入侵群落的可能性,导致群落组成和结构发生明显变化[69].但由于降水模式、群落类型、生态系统类型等不同,植物物种多样性、功能多样性和遗传多样性均呈现不同的响应方式.如长期极端干旱降低了原生草地、灌木丛和灌-草丛的物种丰富度[70];显著降低了非洲大草原和沙质草地物种优势度和多样性[71-72];促使草原优势种发生更替、群落结构单一化[71].生长季极端干旱和生长中期干旱降低了荒漠草原群落的物种多样性指数和丰富度指数[73].生长季降水量和降水格局显著影响了中国新疆和吉尔吉斯斯坦鸭茅(Dactylis glomerata)种群的遗传多样性[74];与维管植物期望杂合度、观测杂合度相关性最高[75];对内蒙古短花针茅(Stipa breviflora)[76]、金发草(Pogonatherum paniceum)[77]和差不嘎蒿(Artemisia halodendron)[78]遗传多样性没有显著影响.极端降水变化对不同生活型植物的叶功能性状影响趋势不同[79];显著提高了荒漠草原的优势物种比叶面积、叶氮、叶磷和叶干物质含量[80].极端干旱发生时,植物通过调整比根长和叶片、根系养分含量来提高水分和养分的吸收能力,保障根系生长的物质来源[81]. ...
Relationship between the genetic diversity of Artemisia halodendron and climatic factors
1
2014
... 极端降水事件通过创造“入侵窗口”,增加外来物种入侵群落的可能性,导致群落组成和结构发生明显变化[69].但由于降水模式、群落类型、生态系统类型等不同,植物物种多样性、功能多样性和遗传多样性均呈现不同的响应方式.如长期极端干旱降低了原生草地、灌木丛和灌-草丛的物种丰富度[70];显著降低了非洲大草原和沙质草地物种优势度和多样性[71-72];促使草原优势种发生更替、群落结构单一化[71].生长季极端干旱和生长中期干旱降低了荒漠草原群落的物种多样性指数和丰富度指数[73].生长季降水量和降水格局显著影响了中国新疆和吉尔吉斯斯坦鸭茅(Dactylis glomerata)种群的遗传多样性[74];与维管植物期望杂合度、观测杂合度相关性最高[75];对内蒙古短花针茅(Stipa breviflora)[76]、金发草(Pogonatherum paniceum)[77]和差不嘎蒿(Artemisia halodendron)[78]遗传多样性没有显著影响.极端降水变化对不同生活型植物的叶功能性状影响趋势不同[79];显著提高了荒漠草原的优势物种比叶面积、叶氮、叶磷和叶干物质含量[80].极端干旱发生时,植物通过调整比根长和叶片、根系养分含量来提高水分和养分的吸收能力,保障根系生长的物质来源[81]. ...
降水梯度对油松天然林内主要植物叶功能性状的影响
1
2016
... 极端降水事件通过创造“入侵窗口”,增加外来物种入侵群落的可能性,导致群落组成和结构发生明显变化[69].但由于降水模式、群落类型、生态系统类型等不同,植物物种多样性、功能多样性和遗传多样性均呈现不同的响应方式.如长期极端干旱降低了原生草地、灌木丛和灌-草丛的物种丰富度[70];显著降低了非洲大草原和沙质草地物种优势度和多样性[71-72];促使草原优势种发生更替、群落结构单一化[71].生长季极端干旱和生长中期干旱降低了荒漠草原群落的物种多样性指数和丰富度指数[73].生长季降水量和降水格局显著影响了中国新疆和吉尔吉斯斯坦鸭茅(Dactylis glomerata)种群的遗传多样性[74];与维管植物期望杂合度、观测杂合度相关性最高[75];对内蒙古短花针茅(Stipa breviflora)[76]、金发草(Pogonatherum paniceum)[77]和差不嘎蒿(Artemisia halodendron)[78]遗传多样性没有显著影响.极端降水变化对不同生活型植物的叶功能性状影响趋势不同[79];显著提高了荒漠草原的优势物种比叶面积、叶氮、叶磷和叶干物质含量[80].极端干旱发生时,植物通过调整比根长和叶片、根系养分含量来提高水分和养分的吸收能力,保障根系生长的物质来源[81]. ...
降水量对荒漠草原骆驼蓬(Peganum harmala)地上生物量的影响
1
2022
... 极端降水事件通过创造“入侵窗口”,增加外来物种入侵群落的可能性,导致群落组成和结构发生明显变化[69].但由于降水模式、群落类型、生态系统类型等不同,植物物种多样性、功能多样性和遗传多样性均呈现不同的响应方式.如长期极端干旱降低了原生草地、灌木丛和灌-草丛的物种丰富度[70];显著降低了非洲大草原和沙质草地物种优势度和多样性[71-72];促使草原优势种发生更替、群落结构单一化[71].生长季极端干旱和生长中期干旱降低了荒漠草原群落的物种多样性指数和丰富度指数[73].生长季降水量和降水格局显著影响了中国新疆和吉尔吉斯斯坦鸭茅(Dactylis glomerata)种群的遗传多样性[74];与维管植物期望杂合度、观测杂合度相关性最高[75];对内蒙古短花针茅(Stipa breviflora)[76]、金发草(Pogonatherum paniceum)[77]和差不嘎蒿(Artemisia halodendron)[78]遗传多样性没有显著影响.极端降水变化对不同生活型植物的叶功能性状影响趋势不同[79];显著提高了荒漠草原的优势物种比叶面积、叶氮、叶磷和叶干物质含量[80].极端干旱发生时,植物通过调整比根长和叶片、根系养分含量来提高水分和养分的吸收能力,保障根系生长的物质来源[81]. ...
羌塘高原降水梯度植物叶片、根系性状变异和生态适应对策
1
2020
... 极端降水事件通过创造“入侵窗口”,增加外来物种入侵群落的可能性,导致群落组成和结构发生明显变化[69].但由于降水模式、群落类型、生态系统类型等不同,植物物种多样性、功能多样性和遗传多样性均呈现不同的响应方式.如长期极端干旱降低了原生草地、灌木丛和灌-草丛的物种丰富度[70];显著降低了非洲大草原和沙质草地物种优势度和多样性[71-72];促使草原优势种发生更替、群落结构单一化[71].生长季极端干旱和生长中期干旱降低了荒漠草原群落的物种多样性指数和丰富度指数[73].生长季降水量和降水格局显著影响了中国新疆和吉尔吉斯斯坦鸭茅(Dactylis glomerata)种群的遗传多样性[74];与维管植物期望杂合度、观测杂合度相关性最高[75];对内蒙古短花针茅(Stipa breviflora)[76]、金发草(Pogonatherum paniceum)[77]和差不嘎蒿(Artemisia halodendron)[78]遗传多样性没有显著影响.极端降水变化对不同生活型植物的叶功能性状影响趋势不同[79];显著提高了荒漠草原的优势物种比叶面积、叶氮、叶磷和叶干物质含量[80].极端干旱发生时,植物通过调整比根长和叶片、根系养分含量来提高水分和养分的吸收能力,保障根系生长的物质来源[81]. ...
Nematode communities indicate diverse soil functioning across a fog gradient in the Namib Desert gravel plains
1
2022
... 极端干旱通过改变土壤物理化学环境、植被结构和微生物群落,直接或间接影响土壤动物群落的组成、丰度及分布.在类群多样性方面,土壤线虫作为依赖于水进行生命活动的动物[82]受极端干旱影响极大,虽然线虫会进入无水生物状态停止活动[83]以适应干旱环境,但干旱带来的土壤盐浓度上升、有害离子增多会促进线虫群落的死亡[84];螨类随着干旱程度增加,多样性指数与丰富度指数显著下降,群落结构存在明显差异[85],类群密度也呈下降趋势,并且干旱情况下降水后类群丰富度的增长困难[86].蚂蚁和甲虫有较强的干旱区生存能力,但极端干旱带来的植被盖度、多样性和丰富度的变化会影响它们的活动[87],主要表现在随着高等植物分布减少而迁移,这些动物栖息地在极端环境中分布不均,集中在植物附近的“资源岛”[88].随着干旱加剧,喜湿类群(如蚯蚓、线虫)数量锐减,而耐旱节肢动物(如拟步甲科甲虫)相对丰度上升.在功能多样性方面,腐食性动物由于凋落物分解率下降,碳周转减缓[89],捕食性动物由于初级生产量小,限制了营养金字塔的基础[90],丰度都显著降低.对于植食性昆虫群落结构会随着植物应对极端干旱的策略改变而改变,干旱下植物汁液会积累更多可溶性糖以及含氮营养化合物,有利于吸汁昆虫,但更严重的干旱导致植物化学防御的加强,会限制吸汁昆虫的增长[91].叶片在干旱下会提高细胞外壁厚度,加强角质层[92],降低食叶昆虫的适口性[93],以及化感作用增强,对于这些昆虫来说营养物质增多不足以弥补压力带来的损失,水分胁迫对食叶昆虫造成负面影响,不过干旱后湿润降水促进的新嫩枝叶长出也会有利于食叶昆虫繁荣[94].食根昆虫中不适应土壤干旱的属将退出竞争,而水分需求较低的属将能够利用植物向根部分配更多养分、温度升高和捕食者更少的优势,成为优势物种[89,95]. ...
The use of anhydrobiosis by soil nematodes in the Antarctic Dry Valleys
1
2000
... 极端干旱通过改变土壤物理化学环境、植被结构和微生物群落,直接或间接影响土壤动物群落的组成、丰度及分布.在类群多样性方面,土壤线虫作为依赖于水进行生命活动的动物[82]受极端干旱影响极大,虽然线虫会进入无水生物状态停止活动[83]以适应干旱环境,但干旱带来的土壤盐浓度上升、有害离子增多会促进线虫群落的死亡[84];螨类随着干旱程度增加,多样性指数与丰富度指数显著下降,群落结构存在明显差异[85],类群密度也呈下降趋势,并且干旱情况下降水后类群丰富度的增长困难[86].蚂蚁和甲虫有较强的干旱区生存能力,但极端干旱带来的植被盖度、多样性和丰富度的变化会影响它们的活动[87],主要表现在随着高等植物分布减少而迁移,这些动物栖息地在极端环境中分布不均,集中在植物附近的“资源岛”[88].随着干旱加剧,喜湿类群(如蚯蚓、线虫)数量锐减,而耐旱节肢动物(如拟步甲科甲虫)相对丰度上升.在功能多样性方面,腐食性动物由于凋落物分解率下降,碳周转减缓[89],捕食性动物由于初级生产量小,限制了营养金字塔的基础[90],丰度都显著降低.对于植食性昆虫群落结构会随着植物应对极端干旱的策略改变而改变,干旱下植物汁液会积累更多可溶性糖以及含氮营养化合物,有利于吸汁昆虫,但更严重的干旱导致植物化学防御的加强,会限制吸汁昆虫的增长[91].叶片在干旱下会提高细胞外壁厚度,加强角质层[92],降低食叶昆虫的适口性[93],以及化感作用增强,对于这些昆虫来说营养物质增多不足以弥补压力带来的损失,水分胁迫对食叶昆虫造成负面影响,不过干旱后湿润降水促进的新嫩枝叶长出也会有利于食叶昆虫繁荣[94].食根昆虫中不适应土壤干旱的属将退出竞争,而水分需求较低的属将能够利用植物向根部分配更多养分、温度升高和捕食者更少的优势,成为优势物种[89,95]. ...
土地荒漠化及其恢复过程土壤动物生态学研究进展
1
2025
... 极端干旱通过改变土壤物理化学环境、植被结构和微生物群落,直接或间接影响土壤动物群落的组成、丰度及分布.在类群多样性方面,土壤线虫作为依赖于水进行生命活动的动物[82]受极端干旱影响极大,虽然线虫会进入无水生物状态停止活动[83]以适应干旱环境,但干旱带来的土壤盐浓度上升、有害离子增多会促进线虫群落的死亡[84];螨类随着干旱程度增加,多样性指数与丰富度指数显著下降,群落结构存在明显差异[85],类群密度也呈下降趋势,并且干旱情况下降水后类群丰富度的增长困难[86].蚂蚁和甲虫有较强的干旱区生存能力,但极端干旱带来的植被盖度、多样性和丰富度的变化会影响它们的活动[87],主要表现在随着高等植物分布减少而迁移,这些动物栖息地在极端环境中分布不均,集中在植物附近的“资源岛”[88].随着干旱加剧,喜湿类群(如蚯蚓、线虫)数量锐减,而耐旱节肢动物(如拟步甲科甲虫)相对丰度上升.在功能多样性方面,腐食性动物由于凋落物分解率下降,碳周转减缓[89],捕食性动物由于初级生产量小,限制了营养金字塔的基础[90],丰度都显著降低.对于植食性昆虫群落结构会随着植物应对极端干旱的策略改变而改变,干旱下植物汁液会积累更多可溶性糖以及含氮营养化合物,有利于吸汁昆虫,但更严重的干旱导致植物化学防御的加强,会限制吸汁昆虫的增长[91].叶片在干旱下会提高细胞外壁厚度,加强角质层[92],降低食叶昆虫的适口性[93],以及化感作用增强,对于这些昆虫来说营养物质增多不足以弥补压力带来的损失,水分胁迫对食叶昆虫造成负面影响,不过干旱后湿润降水促进的新嫩枝叶长出也会有利于食叶昆虫繁荣[94].食根昆虫中不适应土壤干旱的属将退出竞争,而水分需求较低的属将能够利用植物向根部分配更多养分、温度升高和捕食者更少的优势,成为优势物种[89,95]. ...
Community structure of epilithic moss mites and their response to environmental factors in different grades of rocky desertification habitats
1
2022
... 极端干旱通过改变土壤物理化学环境、植被结构和微生物群落,直接或间接影响土壤动物群落的组成、丰度及分布.在类群多样性方面,土壤线虫作为依赖于水进行生命活动的动物[82]受极端干旱影响极大,虽然线虫会进入无水生物状态停止活动[83]以适应干旱环境,但干旱带来的土壤盐浓度上升、有害离子增多会促进线虫群落的死亡[84];螨类随着干旱程度增加,多样性指数与丰富度指数显著下降,群落结构存在明显差异[85],类群密度也呈下降趋势,并且干旱情况下降水后类群丰富度的增长困难[86].蚂蚁和甲虫有较强的干旱区生存能力,但极端干旱带来的植被盖度、多样性和丰富度的变化会影响它们的活动[87],主要表现在随着高等植物分布减少而迁移,这些动物栖息地在极端环境中分布不均,集中在植物附近的“资源岛”[88].随着干旱加剧,喜湿类群(如蚯蚓、线虫)数量锐减,而耐旱节肢动物(如拟步甲科甲虫)相对丰度上升.在功能多样性方面,腐食性动物由于凋落物分解率下降,碳周转减缓[89],捕食性动物由于初级生产量小,限制了营养金字塔的基础[90],丰度都显著降低.对于植食性昆虫群落结构会随着植物应对极端干旱的策略改变而改变,干旱下植物汁液会积累更多可溶性糖以及含氮营养化合物,有利于吸汁昆虫,但更严重的干旱导致植物化学防御的加强,会限制吸汁昆虫的增长[91].叶片在干旱下会提高细胞外壁厚度,加强角质层[92],降低食叶昆虫的适口性[93],以及化感作用增强,对于这些昆虫来说营养物质增多不足以弥补压力带来的损失,水分胁迫对食叶昆虫造成负面影响,不过干旱后湿润降水促进的新嫩枝叶长出也会有利于食叶昆虫繁荣[94].食根昆虫中不适应土壤干旱的属将退出竞争,而水分需求较低的属将能够利用植物向根部分配更多养分、温度升高和捕食者更少的优势,成为优势物种[89,95]. ...
干旱荒漠螨类和跳虫对降雨的响应
1
2017
... 极端干旱通过改变土壤物理化学环境、植被结构和微生物群落,直接或间接影响土壤动物群落的组成、丰度及分布.在类群多样性方面,土壤线虫作为依赖于水进行生命活动的动物[82]受极端干旱影响极大,虽然线虫会进入无水生物状态停止活动[83]以适应干旱环境,但干旱带来的土壤盐浓度上升、有害离子增多会促进线虫群落的死亡[84];螨类随着干旱程度增加,多样性指数与丰富度指数显著下降,群落结构存在明显差异[85],类群密度也呈下降趋势,并且干旱情况下降水后类群丰富度的增长困难[86].蚂蚁和甲虫有较强的干旱区生存能力,但极端干旱带来的植被盖度、多样性和丰富度的变化会影响它们的活动[87],主要表现在随着高等植物分布减少而迁移,这些动物栖息地在极端环境中分布不均,集中在植物附近的“资源岛”[88].随着干旱加剧,喜湿类群(如蚯蚓、线虫)数量锐减,而耐旱节肢动物(如拟步甲科甲虫)相对丰度上升.在功能多样性方面,腐食性动物由于凋落物分解率下降,碳周转减缓[89],捕食性动物由于初级生产量小,限制了营养金字塔的基础[90],丰度都显著降低.对于植食性昆虫群落结构会随着植物应对极端干旱的策略改变而改变,干旱下植物汁液会积累更多可溶性糖以及含氮营养化合物,有利于吸汁昆虫,但更严重的干旱导致植物化学防御的加强,会限制吸汁昆虫的增长[91].叶片在干旱下会提高细胞外壁厚度,加强角质层[92],降低食叶昆虫的适口性[93],以及化感作用增强,对于这些昆虫来说营养物质增多不足以弥补压力带来的损失,水分胁迫对食叶昆虫造成负面影响,不过干旱后湿润降水促进的新嫩枝叶长出也会有利于食叶昆虫繁荣[94].食根昆虫中不适应土壤干旱的属将退出竞争,而水分需求较低的属将能够利用植物向根部分配更多养分、温度升高和捕食者更少的优势,成为优势物种[89,95]. ...
Social life in arid environments:the case study of Cataglyphis ants
1
2017
... 极端干旱通过改变土壤物理化学环境、植被结构和微生物群落,直接或间接影响土壤动物群落的组成、丰度及分布.在类群多样性方面,土壤线虫作为依赖于水进行生命活动的动物[82]受极端干旱影响极大,虽然线虫会进入无水生物状态停止活动[83]以适应干旱环境,但干旱带来的土壤盐浓度上升、有害离子增多会促进线虫群落的死亡[84];螨类随着干旱程度增加,多样性指数与丰富度指数显著下降,群落结构存在明显差异[85],类群密度也呈下降趋势,并且干旱情况下降水后类群丰富度的增长困难[86].蚂蚁和甲虫有较强的干旱区生存能力,但极端干旱带来的植被盖度、多样性和丰富度的变化会影响它们的活动[87],主要表现在随着高等植物分布减少而迁移,这些动物栖息地在极端环境中分布不均,集中在植物附近的“资源岛”[88].随着干旱加剧,喜湿类群(如蚯蚓、线虫)数量锐减,而耐旱节肢动物(如拟步甲科甲虫)相对丰度上升.在功能多样性方面,腐食性动物由于凋落物分解率下降,碳周转减缓[89],捕食性动物由于初级生产量小,限制了营养金字塔的基础[90],丰度都显著降低.对于植食性昆虫群落结构会随着植物应对极端干旱的策略改变而改变,干旱下植物汁液会积累更多可溶性糖以及含氮营养化合物,有利于吸汁昆虫,但更严重的干旱导致植物化学防御的加强,会限制吸汁昆虫的增长[91].叶片在干旱下会提高细胞外壁厚度,加强角质层[92],降低食叶昆虫的适口性[93],以及化感作用增强,对于这些昆虫来说营养物质增多不足以弥补压力带来的损失,水分胁迫对食叶昆虫造成负面影响,不过干旱后湿润降水促进的新嫩枝叶长出也会有利于食叶昆虫繁荣[94].食根昆虫中不适应土壤干旱的属将退出竞争,而水分需求较低的属将能够利用植物向根部分配更多养分、温度升高和捕食者更少的优势,成为优势物种[89,95]. ...
Controls on soil biodiversity:insights from extreme environments
1
1999
... 极端干旱通过改变土壤物理化学环境、植被结构和微生物群落,直接或间接影响土壤动物群落的组成、丰度及分布.在类群多样性方面,土壤线虫作为依赖于水进行生命活动的动物[82]受极端干旱影响极大,虽然线虫会进入无水生物状态停止活动[83]以适应干旱环境,但干旱带来的土壤盐浓度上升、有害离子增多会促进线虫群落的死亡[84];螨类随着干旱程度增加,多样性指数与丰富度指数显著下降,群落结构存在明显差异[85],类群密度也呈下降趋势,并且干旱情况下降水后类群丰富度的增长困难[86].蚂蚁和甲虫有较强的干旱区生存能力,但极端干旱带来的植被盖度、多样性和丰富度的变化会影响它们的活动[87],主要表现在随着高等植物分布减少而迁移,这些动物栖息地在极端环境中分布不均,集中在植物附近的“资源岛”[88].随着干旱加剧,喜湿类群(如蚯蚓、线虫)数量锐减,而耐旱节肢动物(如拟步甲科甲虫)相对丰度上升.在功能多样性方面,腐食性动物由于凋落物分解率下降,碳周转减缓[89],捕食性动物由于初级生产量小,限制了营养金字塔的基础[90],丰度都显著降低.对于植食性昆虫群落结构会随着植物应对极端干旱的策略改变而改变,干旱下植物汁液会积累更多可溶性糖以及含氮营养化合物,有利于吸汁昆虫,但更严重的干旱导致植物化学防御的加强,会限制吸汁昆虫的增长[91].叶片在干旱下会提高细胞外壁厚度,加强角质层[92],降低食叶昆虫的适口性[93],以及化感作用增强,对于这些昆虫来说营养物质增多不足以弥补压力带来的损失,水分胁迫对食叶昆虫造成负面影响,不过干旱后湿润降水促进的新嫩枝叶长出也会有利于食叶昆虫繁荣[94].食根昆虫中不适应土壤干旱的属将退出竞争,而水分需求较低的属将能够利用植物向根部分配更多养分、温度升高和捕食者更少的优势,成为优势物种[89,95]. ...
Suppression of soil decomposers and promotion of long-lived,root herbivorous nematodes by climate change
2
2012
... 极端干旱通过改变土壤物理化学环境、植被结构和微生物群落,直接或间接影响土壤动物群落的组成、丰度及分布.在类群多样性方面,土壤线虫作为依赖于水进行生命活动的动物[82]受极端干旱影响极大,虽然线虫会进入无水生物状态停止活动[83]以适应干旱环境,但干旱带来的土壤盐浓度上升、有害离子增多会促进线虫群落的死亡[84];螨类随着干旱程度增加,多样性指数与丰富度指数显著下降,群落结构存在明显差异[85],类群密度也呈下降趋势,并且干旱情况下降水后类群丰富度的增长困难[86].蚂蚁和甲虫有较强的干旱区生存能力,但极端干旱带来的植被盖度、多样性和丰富度的变化会影响它们的活动[87],主要表现在随着高等植物分布减少而迁移,这些动物栖息地在极端环境中分布不均,集中在植物附近的“资源岛”[88].随着干旱加剧,喜湿类群(如蚯蚓、线虫)数量锐减,而耐旱节肢动物(如拟步甲科甲虫)相对丰度上升.在功能多样性方面,腐食性动物由于凋落物分解率下降,碳周转减缓[89],捕食性动物由于初级生产量小,限制了营养金字塔的基础[90],丰度都显著降低.对于植食性昆虫群落结构会随着植物应对极端干旱的策略改变而改变,干旱下植物汁液会积累更多可溶性糖以及含氮营养化合物,有利于吸汁昆虫,但更严重的干旱导致植物化学防御的加强,会限制吸汁昆虫的增长[91].叶片在干旱下会提高细胞外壁厚度,加强角质层[92],降低食叶昆虫的适口性[93],以及化感作用增强,对于这些昆虫来说营养物质增多不足以弥补压力带来的损失,水分胁迫对食叶昆虫造成负面影响,不过干旱后湿润降水促进的新嫩枝叶长出也会有利于食叶昆虫繁荣[94].食根昆虫中不适应土壤干旱的属将退出竞争,而水分需求较低的属将能够利用植物向根部分配更多养分、温度升高和捕食者更少的优势,成为优势物种[89,95]. ...
... [89,95]. ...
Responses of a desert nematode community to changes in water availability
1
2015
... 极端干旱通过改变土壤物理化学环境、植被结构和微生物群落,直接或间接影响土壤动物群落的组成、丰度及分布.在类群多样性方面,土壤线虫作为依赖于水进行生命活动的动物[82]受极端干旱影响极大,虽然线虫会进入无水生物状态停止活动[83]以适应干旱环境,但干旱带来的土壤盐浓度上升、有害离子增多会促进线虫群落的死亡[84];螨类随着干旱程度增加,多样性指数与丰富度指数显著下降,群落结构存在明显差异[85],类群密度也呈下降趋势,并且干旱情况下降水后类群丰富度的增长困难[86].蚂蚁和甲虫有较强的干旱区生存能力,但极端干旱带来的植被盖度、多样性和丰富度的变化会影响它们的活动[87],主要表现在随着高等植物分布减少而迁移,这些动物栖息地在极端环境中分布不均,集中在植物附近的“资源岛”[88].随着干旱加剧,喜湿类群(如蚯蚓、线虫)数量锐减,而耐旱节肢动物(如拟步甲科甲虫)相对丰度上升.在功能多样性方面,腐食性动物由于凋落物分解率下降,碳周转减缓[89],捕食性动物由于初级生产量小,限制了营养金字塔的基础[90],丰度都显著降低.对于植食性昆虫群落结构会随着植物应对极端干旱的策略改变而改变,干旱下植物汁液会积累更多可溶性糖以及含氮营养化合物,有利于吸汁昆虫,但更严重的干旱导致植物化学防御的加强,会限制吸汁昆虫的增长[91].叶片在干旱下会提高细胞外壁厚度,加强角质层[92],降低食叶昆虫的适口性[93],以及化感作用增强,对于这些昆虫来说营养物质增多不足以弥补压力带来的损失,水分胁迫对食叶昆虫造成负面影响,不过干旱后湿润降水促进的新嫩枝叶长出也会有利于食叶昆虫繁荣[94].食根昆虫中不适应土壤干旱的属将退出竞争,而水分需求较低的属将能够利用植物向根部分配更多养分、温度升高和捕食者更少的优势,成为优势物种[89,95]. ...
Plant water stress and its consequences for herbivorous insects:a new synthesis
1
2004
... 极端干旱通过改变土壤物理化学环境、植被结构和微生物群落,直接或间接影响土壤动物群落的组成、丰度及分布.在类群多样性方面,土壤线虫作为依赖于水进行生命活动的动物[82]受极端干旱影响极大,虽然线虫会进入无水生物状态停止活动[83]以适应干旱环境,但干旱带来的土壤盐浓度上升、有害离子增多会促进线虫群落的死亡[84];螨类随着干旱程度增加,多样性指数与丰富度指数显著下降,群落结构存在明显差异[85],类群密度也呈下降趋势,并且干旱情况下降水后类群丰富度的增长困难[86].蚂蚁和甲虫有较强的干旱区生存能力,但极端干旱带来的植被盖度、多样性和丰富度的变化会影响它们的活动[87],主要表现在随着高等植物分布减少而迁移,这些动物栖息地在极端环境中分布不均,集中在植物附近的“资源岛”[88].随着干旱加剧,喜湿类群(如蚯蚓、线虫)数量锐减,而耐旱节肢动物(如拟步甲科甲虫)相对丰度上升.在功能多样性方面,腐食性动物由于凋落物分解率下降,碳周转减缓[89],捕食性动物由于初级生产量小,限制了营养金字塔的基础[90],丰度都显著降低.对于植食性昆虫群落结构会随着植物应对极端干旱的策略改变而改变,干旱下植物汁液会积累更多可溶性糖以及含氮营养化合物,有利于吸汁昆虫,但更严重的干旱导致植物化学防御的加强,会限制吸汁昆虫的增长[91].叶片在干旱下会提高细胞外壁厚度,加强角质层[92],降低食叶昆虫的适口性[93],以及化感作用增强,对于这些昆虫来说营养物质增多不足以弥补压力带来的损失,水分胁迫对食叶昆虫造成负面影响,不过干旱后湿润降水促进的新嫩枝叶长出也会有利于食叶昆虫繁荣[94].食根昆虫中不适应土壤干旱的属将退出竞争,而水分需求较低的属将能够利用植物向根部分配更多养分、温度升高和捕食者更少的优势,成为优势物种[89,95]. ...
我国旱生植物的形态解剖学研究
1
2002
... 极端干旱通过改变土壤物理化学环境、植被结构和微生物群落,直接或间接影响土壤动物群落的组成、丰度及分布.在类群多样性方面,土壤线虫作为依赖于水进行生命活动的动物[82]受极端干旱影响极大,虽然线虫会进入无水生物状态停止活动[83]以适应干旱环境,但干旱带来的土壤盐浓度上升、有害离子增多会促进线虫群落的死亡[84];螨类随着干旱程度增加,多样性指数与丰富度指数显著下降,群落结构存在明显差异[85],类群密度也呈下降趋势,并且干旱情况下降水后类群丰富度的增长困难[86].蚂蚁和甲虫有较强的干旱区生存能力,但极端干旱带来的植被盖度、多样性和丰富度的变化会影响它们的活动[87],主要表现在随着高等植物分布减少而迁移,这些动物栖息地在极端环境中分布不均,集中在植物附近的“资源岛”[88].随着干旱加剧,喜湿类群(如蚯蚓、线虫)数量锐减,而耐旱节肢动物(如拟步甲科甲虫)相对丰度上升.在功能多样性方面,腐食性动物由于凋落物分解率下降,碳周转减缓[89],捕食性动物由于初级生产量小,限制了营养金字塔的基础[90],丰度都显著降低.对于植食性昆虫群落结构会随着植物应对极端干旱的策略改变而改变,干旱下植物汁液会积累更多可溶性糖以及含氮营养化合物,有利于吸汁昆虫,但更严重的干旱导致植物化学防御的加强,会限制吸汁昆虫的增长[91].叶片在干旱下会提高细胞外壁厚度,加强角质层[92],降低食叶昆虫的适口性[93],以及化感作用增强,对于这些昆虫来说营养物质增多不足以弥补压力带来的损失,水分胁迫对食叶昆虫造成负面影响,不过干旱后湿润降水促进的新嫩枝叶长出也会有利于食叶昆虫繁荣[94].食根昆虫中不适应土壤干旱的属将退出竞争,而水分需求较低的属将能够利用植物向根部分配更多养分、温度升高和捕食者更少的优势,成为优势物种[89,95]. ...
Plant trait-digestibility relationships across management and climate gradients in permanent grasslands
1
2014
... 极端干旱通过改变土壤物理化学环境、植被结构和微生物群落,直接或间接影响土壤动物群落的组成、丰度及分布.在类群多样性方面,土壤线虫作为依赖于水进行生命活动的动物[82]受极端干旱影响极大,虽然线虫会进入无水生物状态停止活动[83]以适应干旱环境,但干旱带来的土壤盐浓度上升、有害离子增多会促进线虫群落的死亡[84];螨类随着干旱程度增加,多样性指数与丰富度指数显著下降,群落结构存在明显差异[85],类群密度也呈下降趋势,并且干旱情况下降水后类群丰富度的增长困难[86].蚂蚁和甲虫有较强的干旱区生存能力,但极端干旱带来的植被盖度、多样性和丰富度的变化会影响它们的活动[87],主要表现在随着高等植物分布减少而迁移,这些动物栖息地在极端环境中分布不均,集中在植物附近的“资源岛”[88].随着干旱加剧,喜湿类群(如蚯蚓、线虫)数量锐减,而耐旱节肢动物(如拟步甲科甲虫)相对丰度上升.在功能多样性方面,腐食性动物由于凋落物分解率下降,碳周转减缓[89],捕食性动物由于初级生产量小,限制了营养金字塔的基础[90],丰度都显著降低.对于植食性昆虫群落结构会随着植物应对极端干旱的策略改变而改变,干旱下植物汁液会积累更多可溶性糖以及含氮营养化合物,有利于吸汁昆虫,但更严重的干旱导致植物化学防御的加强,会限制吸汁昆虫的增长[91].叶片在干旱下会提高细胞外壁厚度,加强角质层[92],降低食叶昆虫的适口性[93],以及化感作用增强,对于这些昆虫来说营养物质增多不足以弥补压力带来的损失,水分胁迫对食叶昆虫造成负面影响,不过干旱后湿润降水促进的新嫩枝叶长出也会有利于食叶昆虫繁荣[94].食根昆虫中不适应土壤干旱的属将退出竞争,而水分需求较低的属将能够利用植物向根部分配更多养分、温度升高和捕食者更少的优势,成为优势物种[89,95]. ...
How do herbivorous insects respond to drought stress in trees?
1
2020
... 极端干旱通过改变土壤物理化学环境、植被结构和微生物群落,直接或间接影响土壤动物群落的组成、丰度及分布.在类群多样性方面,土壤线虫作为依赖于水进行生命活动的动物[82]受极端干旱影响极大,虽然线虫会进入无水生物状态停止活动[83]以适应干旱环境,但干旱带来的土壤盐浓度上升、有害离子增多会促进线虫群落的死亡[84];螨类随着干旱程度增加,多样性指数与丰富度指数显著下降,群落结构存在明显差异[85],类群密度也呈下降趋势,并且干旱情况下降水后类群丰富度的增长困难[86].蚂蚁和甲虫有较强的干旱区生存能力,但极端干旱带来的植被盖度、多样性和丰富度的变化会影响它们的活动[87],主要表现在随着高等植物分布减少而迁移,这些动物栖息地在极端环境中分布不均,集中在植物附近的“资源岛”[88].随着干旱加剧,喜湿类群(如蚯蚓、线虫)数量锐减,而耐旱节肢动物(如拟步甲科甲虫)相对丰度上升.在功能多样性方面,腐食性动物由于凋落物分解率下降,碳周转减缓[89],捕食性动物由于初级生产量小,限制了营养金字塔的基础[90],丰度都显著降低.对于植食性昆虫群落结构会随着植物应对极端干旱的策略改变而改变,干旱下植物汁液会积累更多可溶性糖以及含氮营养化合物,有利于吸汁昆虫,但更严重的干旱导致植物化学防御的加强,会限制吸汁昆虫的增长[91].叶片在干旱下会提高细胞外壁厚度,加强角质层[92],降低食叶昆虫的适口性[93],以及化感作用增强,对于这些昆虫来说营养物质增多不足以弥补压力带来的损失,水分胁迫对食叶昆虫造成负面影响,不过干旱后湿润降水促进的新嫩枝叶长出也会有利于食叶昆虫繁荣[94].食根昆虫中不适应土壤干旱的属将退出竞争,而水分需求较低的属将能够利用植物向根部分配更多养分、温度升高和捕食者更少的优势,成为优势物种[89,95]. ...
Soil factors determined nematode community composition in a two year pot experiment
1
2003
... 极端干旱通过改变土壤物理化学环境、植被结构和微生物群落,直接或间接影响土壤动物群落的组成、丰度及分布.在类群多样性方面,土壤线虫作为依赖于水进行生命活动的动物[82]受极端干旱影响极大,虽然线虫会进入无水生物状态停止活动[83]以适应干旱环境,但干旱带来的土壤盐浓度上升、有害离子增多会促进线虫群落的死亡[84];螨类随着干旱程度增加,多样性指数与丰富度指数显著下降,群落结构存在明显差异[85],类群密度也呈下降趋势,并且干旱情况下降水后类群丰富度的增长困难[86].蚂蚁和甲虫有较强的干旱区生存能力,但极端干旱带来的植被盖度、多样性和丰富度的变化会影响它们的活动[87],主要表现在随着高等植物分布减少而迁移,这些动物栖息地在极端环境中分布不均,集中在植物附近的“资源岛”[88].随着干旱加剧,喜湿类群(如蚯蚓、线虫)数量锐减,而耐旱节肢动物(如拟步甲科甲虫)相对丰度上升.在功能多样性方面,腐食性动物由于凋落物分解率下降,碳周转减缓[89],捕食性动物由于初级生产量小,限制了营养金字塔的基础[90],丰度都显著降低.对于植食性昆虫群落结构会随着植物应对极端干旱的策略改变而改变,干旱下植物汁液会积累更多可溶性糖以及含氮营养化合物,有利于吸汁昆虫,但更严重的干旱导致植物化学防御的加强,会限制吸汁昆虫的增长[91].叶片在干旱下会提高细胞外壁厚度,加强角质层[92],降低食叶昆虫的适口性[93],以及化感作用增强,对于这些昆虫来说营养物质增多不足以弥补压力带来的损失,水分胁迫对食叶昆虫造成负面影响,不过干旱后湿润降水促进的新嫩枝叶长出也会有利于食叶昆虫繁荣[94].食根昆虫中不适应土壤干旱的属将退出竞争,而水分需求较低的属将能够利用植物向根部分配更多养分、温度升高和捕食者更少的优势,成为优势物种[89,95]. ...
生长季不同时期降雨变化对土壤微生物群落结构和异养呼吸的影响
1
2022
... 降雨模式变化首先会改变土壤含水量[96]和土壤净氮矿化速率[97],影响枯落物和细根分解[98],进而调整微生物群落结构、有机物分解和养分循环过程[99],最终制约土壤微生物多样性[64].区域尺度上的土壤微生物群落组成以及细菌、真菌、古菌和线虫等不同维度的多样性主要受降水模式和植被类型(荒漠、草甸和草原等)的驱动[100-101].例如降雨模式变化对林地和草地土壤微生物群落影响次序为生长季干旱>极端降雨>全年增雨>全年减雨[65].极端干旱条件更有利于耐旱性微生物如真菌、厚壁菌、放线菌和革兰氏阴性菌等的生长[102-103].长期干旱降低了草地生态系统土壤真菌的丰富度,延滞细菌群落的共现网络[104-105];改变了草地土壤真菌和细菌比例[9,106];降低了中国北方草原干旱和半干旱区土壤线虫丰富度[107].生长季干旱提高了林地和草地的土壤细菌α多样性和放线菌门相对丰度,改变了丰富和稀有细菌群落组成[65]. ...
青海湖高寒草地土壤理化性质及微生物群落特征对模拟降水的响应
1
2021
... 降雨模式变化首先会改变土壤含水量[96]和土壤净氮矿化速率[97],影响枯落物和细根分解[98],进而调整微生物群落结构、有机物分解和养分循环过程[99],最终制约土壤微生物多样性[64].区域尺度上的土壤微生物群落组成以及细菌、真菌、古菌和线虫等不同维度的多样性主要受降水模式和植被类型(荒漠、草甸和草原等)的驱动[100-101].例如降雨模式变化对林地和草地土壤微生物群落影响次序为生长季干旱>极端降雨>全年增雨>全年减雨[65].极端干旱条件更有利于耐旱性微生物如真菌、厚壁菌、放线菌和革兰氏阴性菌等的生长[102-103].长期干旱降低了草地生态系统土壤真菌的丰富度,延滞细菌群落的共现网络[104-105];改变了草地土壤真菌和细菌比例[9,106];降低了中国北方草原干旱和半干旱区土壤线虫丰富度[107].生长季干旱提高了林地和草地的土壤细菌α多样性和放线菌门相对丰度,改变了丰富和稀有细菌群落组成[65]. ...
Differential responses of litter decomposition to warming,elevated CO2,and changed precipitation regime
1
2020
... 降雨模式变化首先会改变土壤含水量[96]和土壤净氮矿化速率[97],影响枯落物和细根分解[98],进而调整微生物群落结构、有机物分解和养分循环过程[99],最终制约土壤微生物多样性[64].区域尺度上的土壤微生物群落组成以及细菌、真菌、古菌和线虫等不同维度的多样性主要受降水模式和植被类型(荒漠、草甸和草原等)的驱动[100-101].例如降雨模式变化对林地和草地土壤微生物群落影响次序为生长季干旱>极端降雨>全年增雨>全年减雨[65].极端干旱条件更有利于耐旱性微生物如真菌、厚壁菌、放线菌和革兰氏阴性菌等的生长[102-103].长期干旱降低了草地生态系统土壤真菌的丰富度,延滞细菌群落的共现网络[104-105];改变了草地土壤真菌和细菌比例[9,106];降低了中国北方草原干旱和半干旱区土壤线虫丰富度[107].生长季干旱提高了林地和草地的土壤细菌α多样性和放线菌门相对丰度,改变了丰富和稀有细菌群落组成[65]. ...
Soil microbial activity in a Mediterranean garrigue responds more to changing shrub community than to reduced rainfall
1
2020
... 降雨模式变化首先会改变土壤含水量[96]和土壤净氮矿化速率[97],影响枯落物和细根分解[98],进而调整微生物群落结构、有机物分解和养分循环过程[99],最终制约土壤微生物多样性[64].区域尺度上的土壤微生物群落组成以及细菌、真菌、古菌和线虫等不同维度的多样性主要受降水模式和植被类型(荒漠、草甸和草原等)的驱动[100-101].例如降雨模式变化对林地和草地土壤微生物群落影响次序为生长季干旱>极端降雨>全年增雨>全年减雨[65].极端干旱条件更有利于耐旱性微生物如真菌、厚壁菌、放线菌和革兰氏阴性菌等的生长[102-103].长期干旱降低了草地生态系统土壤真菌的丰富度,延滞细菌群落的共现网络[104-105];改变了草地土壤真菌和细菌比例[9,106];降低了中国北方草原干旱和半干旱区土壤线虫丰富度[107].生长季干旱提高了林地和草地的土壤细菌α多样性和放线菌门相对丰度,改变了丰富和稀有细菌群落组成[65]. ...
Soil microbial biomass and functional diversity in shrub-encroached grasslands along a precipitation gradient
1
2017
... 降雨模式变化首先会改变土壤含水量[96]和土壤净氮矿化速率[97],影响枯落物和细根分解[98],进而调整微生物群落结构、有机物分解和养分循环过程[99],最终制约土壤微生物多样性[64].区域尺度上的土壤微生物群落组成以及细菌、真菌、古菌和线虫等不同维度的多样性主要受降水模式和植被类型(荒漠、草甸和草原等)的驱动[100-101].例如降雨模式变化对林地和草地土壤微生物群落影响次序为生长季干旱>极端降雨>全年增雨>全年减雨[65].极端干旱条件更有利于耐旱性微生物如真菌、厚壁菌、放线菌和革兰氏阴性菌等的生长[102-103].长期干旱降低了草地生态系统土壤真菌的丰富度,延滞细菌群落的共现网络[104-105];改变了草地土壤真菌和细菌比例[9,106];降低了中国北方草原干旱和半干旱区土壤线虫丰富度[107].生长季干旱提高了林地和草地的土壤细菌α多样性和放线菌门相对丰度,改变了丰富和稀有细菌群落组成[65]. ...
青藏高原高寒草地土壤微生物群落及影响因子
1
2019
... 降雨模式变化首先会改变土壤含水量[96]和土壤净氮矿化速率[97],影响枯落物和细根分解[98],进而调整微生物群落结构、有机物分解和养分循环过程[99],最终制约土壤微生物多样性[64].区域尺度上的土壤微生物群落组成以及细菌、真菌、古菌和线虫等不同维度的多样性主要受降水模式和植被类型(荒漠、草甸和草原等)的驱动[100-101].例如降雨模式变化对林地和草地土壤微生物群落影响次序为生长季干旱>极端降雨>全年增雨>全年减雨[65].极端干旱条件更有利于耐旱性微生物如真菌、厚壁菌、放线菌和革兰氏阴性菌等的生长[102-103].长期干旱降低了草地生态系统土壤真菌的丰富度,延滞细菌群落的共现网络[104-105];改变了草地土壤真菌和细菌比例[9,106];降低了中国北方草原干旱和半干旱区土壤线虫丰富度[107].生长季干旱提高了林地和草地的土壤细菌α多样性和放线菌门相对丰度,改变了丰富和稀有细菌群落组成[65]. ...
Multiple climate change factors interact to alter soil microbial community structure in an old-field ecosystem
1
2011
... 降雨模式变化首先会改变土壤含水量[96]和土壤净氮矿化速率[97],影响枯落物和细根分解[98],进而调整微生物群落结构、有机物分解和养分循环过程[99],最终制约土壤微生物多样性[64].区域尺度上的土壤微生物群落组成以及细菌、真菌、古菌和线虫等不同维度的多样性主要受降水模式和植被类型(荒漠、草甸和草原等)的驱动[100-101].例如降雨模式变化对林地和草地土壤微生物群落影响次序为生长季干旱>极端降雨>全年增雨>全年减雨[65].极端干旱条件更有利于耐旱性微生物如真菌、厚壁菌、放线菌和革兰氏阴性菌等的生长[102-103].长期干旱降低了草地生态系统土壤真菌的丰富度,延滞细菌群落的共现网络[104-105];改变了草地土壤真菌和细菌比例[9,106];降低了中国北方草原干旱和半干旱区土壤线虫丰富度[107].生长季干旱提高了林地和草地的土壤细菌α多样性和放线菌门相对丰度,改变了丰富和稀有细菌群落组成[65]. ...
Biogeographic patterns of soil microbe communities in the deserts of the Hexi Corridor,northern China
2
2022
... 降雨模式变化首先会改变土壤含水量[96]和土壤净氮矿化速率[97],影响枯落物和细根分解[98],进而调整微生物群落结构、有机物分解和养分循环过程[99],最终制约土壤微生物多样性[64].区域尺度上的土壤微生物群落组成以及细菌、真菌、古菌和线虫等不同维度的多样性主要受降水模式和植被类型(荒漠、草甸和草原等)的驱动[100-101].例如降雨模式变化对林地和草地土壤微生物群落影响次序为生长季干旱>极端降雨>全年增雨>全年减雨[65].极端干旱条件更有利于耐旱性微生物如真菌、厚壁菌、放线菌和革兰氏阴性菌等的生长[102-103].长期干旱降低了草地生态系统土壤真菌的丰富度,延滞细菌群落的共现网络[104-105];改变了草地土壤真菌和细菌比例[9,106];降低了中国北方草原干旱和半干旱区土壤线虫丰富度[107].生长季干旱提高了林地和草地的土壤细菌α多样性和放线菌门相对丰度,改变了丰富和稀有细菌群落组成[65]. ...
... 干旱除了对群落生物量产生影响外,也会对生态系统碳循环、氮循环等产生不同程度的影响,进而影响生态系统的养分循环和水循环,对生态系统多功能性起关键作用[114-115].生长季极端干旱会减弱土壤微生物活动[103]、降低氮矿化速率[111]和凋落物生物量的碳氮密度[116].极端干旱会显著影响氮素的转化过程[117]、土壤碳稳定性以及土壤团聚体的碳封存功能[56]、土壤总碳和总氮含量[118],增加贝加尔针茅(S. baicalensis)草原植物叶片碳含量[25,119].极端干旱也能导致土壤动物功能群重组与效率失衡[120]:杂食性和腐食性类群减少[121],微生物互作网络受损,氮利用率低,养分循环阻滞,结构稳定性下降[122].土壤动物群落冗余度降低,抗干扰能力下降,植物病原菌繁殖加速,植物生产力显著降低[123].蚂蚁、蚯蚓、千足虫等清道夫数量减少,土壤沉积物、有机质垂直运输的能力变弱[124-125]. ...
Links between soil microbial communities and plant traits in a species-rich grassland under long-term climate change
1
2017
... 降雨模式变化首先会改变土壤含水量[96]和土壤净氮矿化速率[97],影响枯落物和细根分解[98],进而调整微生物群落结构、有机物分解和养分循环过程[99],最终制约土壤微生物多样性[64].区域尺度上的土壤微生物群落组成以及细菌、真菌、古菌和线虫等不同维度的多样性主要受降水模式和植被类型(荒漠、草甸和草原等)的驱动[100-101].例如降雨模式变化对林地和草地土壤微生物群落影响次序为生长季干旱>极端降雨>全年增雨>全年减雨[65].极端干旱条件更有利于耐旱性微生物如真菌、厚壁菌、放线菌和革兰氏阴性菌等的生长[102-103].长期干旱降低了草地生态系统土壤真菌的丰富度,延滞细菌群落的共现网络[104-105];改变了草地土壤真菌和细菌比例[9,106];降低了中国北方草原干旱和半干旱区土壤线虫丰富度[107].生长季干旱提高了林地和草地的土壤细菌α多样性和放线菌门相对丰度,改变了丰富和稀有细菌群落组成[65]. ...
Soil bacterial networks are less stable under drought than fungal networks
1
2018
... 降雨模式变化首先会改变土壤含水量[96]和土壤净氮矿化速率[97],影响枯落物和细根分解[98],进而调整微生物群落结构、有机物分解和养分循环过程[99],最终制约土壤微生物多样性[64].区域尺度上的土壤微生物群落组成以及细菌、真菌、古菌和线虫等不同维度的多样性主要受降水模式和植被类型(荒漠、草甸和草原等)的驱动[100-101].例如降雨模式变化对林地和草地土壤微生物群落影响次序为生长季干旱>极端降雨>全年增雨>全年减雨[65].极端干旱条件更有利于耐旱性微生物如真菌、厚壁菌、放线菌和革兰氏阴性菌等的生长[102-103].长期干旱降低了草地生态系统土壤真菌的丰富度,延滞细菌群落的共现网络[104-105];改变了草地土壤真菌和细菌比例[9,106];降低了中国北方草原干旱和半干旱区土壤线虫丰富度[107].生长季干旱提高了林地和草地的土壤细菌α多样性和放线菌门相对丰度,改变了丰富和稀有细菌群落组成[65]. ...
Drought stress induced increase of fungi:bacteria ratio in a poplar plantation
1
2020
... 降雨模式变化首先会改变土壤含水量[96]和土壤净氮矿化速率[97],影响枯落物和细根分解[98],进而调整微生物群落结构、有机物分解和养分循环过程[99],最终制约土壤微生物多样性[64].区域尺度上的土壤微生物群落组成以及细菌、真菌、古菌和线虫等不同维度的多样性主要受降水模式和植被类型(荒漠、草甸和草原等)的驱动[100-101].例如降雨模式变化对林地和草地土壤微生物群落影响次序为生长季干旱>极端降雨>全年增雨>全年减雨[65].极端干旱条件更有利于耐旱性微生物如真菌、厚壁菌、放线菌和革兰氏阴性菌等的生长[102-103].长期干旱降低了草地生态系统土壤真菌的丰富度,延滞细菌群落的共现网络[104-105];改变了草地土壤真菌和细菌比例[9,106];降低了中国北方草原干旱和半干旱区土壤线虫丰富度[107].生长季干旱提高了林地和草地的土壤细菌α多样性和放线菌门相对丰度,改变了丰富和稀有细菌群落组成[65]. ...
Nonlinear responses of soil nematode community composition to increasing aridity
1
2020
... 降雨模式变化首先会改变土壤含水量[96]和土壤净氮矿化速率[97],影响枯落物和细根分解[98],进而调整微生物群落结构、有机物分解和养分循环过程[99],最终制约土壤微生物多样性[64].区域尺度上的土壤微生物群落组成以及细菌、真菌、古菌和线虫等不同维度的多样性主要受降水模式和植被类型(荒漠、草甸和草原等)的驱动[100-101].例如降雨模式变化对林地和草地土壤微生物群落影响次序为生长季干旱>极端降雨>全年增雨>全年减雨[65].极端干旱条件更有利于耐旱性微生物如真菌、厚壁菌、放线菌和革兰氏阴性菌等的生长[102-103].长期干旱降低了草地生态系统土壤真菌的丰富度,延滞细菌群落的共现网络[104-105];改变了草地土壤真菌和细菌比例[9,106];降低了中国北方草原干旱和半干旱区土壤线虫丰富度[107].生长季干旱提高了林地和草地的土壤细菌α多样性和放线菌门相对丰度,改变了丰富和稀有细菌群落组成[65]. ...
Aridity-dependent resistance but strong resilience of grassland ANPP to naturally occurring precipitation extremes
1
2024
... 降水可以通过改变生物多样性影响生态系统功能[12],也可以通过改变土壤性质、植物光合速率、根系分泌物等影响生物量,从而改变生态系统功能[19].众所周知生物量是研究生态系统碳收支和碳循环的重要环节.生物量分配反映了地上资源(光和CO2)与地下资源(水和养分)之间的平衡,被广泛用作指示陆地生态系统碳循环对全球变化响应的关键指标.大量野外实地研究和文献综述表明,群落生物量和生产力与极端降水事件具有很强的相关性,例如极端干旱事件使得全球典型草地生产力降低了40%[108]、中国荒漠草原沙生针茅(S. caucasica)群落地上生物量降低了50%以上[73];改变了荒漠生态系统典型植物骆驼刺(Alhagi sparsifolia)的叶磷分配模式[38];降低了全世界范围内83个草地C3禾草、C4杂草的生物量[109]和温带草原生态系统[110]、半干旱沙质草地[111]的地上、地下生产力.季节性极端干旱对不同生态系统群落生物量也产生了一定影响,例如生长季初期的春季极端干旱和生长季中期的夏季极端干旱显著降低了温带草原生态系统和草甸草原的地上生物量[112-113]. ...
Asymmetric responses of primary productivity to precipitation extremes:a synthesis of grassland precipitation manipulation experiments
1
2017
... 降水可以通过改变生物多样性影响生态系统功能[12],也可以通过改变土壤性质、植物光合速率、根系分泌物等影响生物量,从而改变生态系统功能[19].众所周知生物量是研究生态系统碳收支和碳循环的重要环节.生物量分配反映了地上资源(光和CO2)与地下资源(水和养分)之间的平衡,被广泛用作指示陆地生态系统碳循环对全球变化响应的关键指标.大量野外实地研究和文献综述表明,群落生物量和生产力与极端降水事件具有很强的相关性,例如极端干旱事件使得全球典型草地生产力降低了40%[108]、中国荒漠草原沙生针茅(S. caucasica)群落地上生物量降低了50%以上[73];改变了荒漠生态系统典型植物骆驼刺(Alhagi sparsifolia)的叶磷分配模式[38];降低了全世界范围内83个草地C3禾草、C4杂草的生物量[109]和温带草原生态系统[110]、半干旱沙质草地[111]的地上、地下生产力.季节性极端干旱对不同生态系统群落生物量也产生了一定影响,例如生长季初期的春季极端干旱和生长季中期的夏季极端干旱显著降低了温带草原生态系统和草甸草原的地上生物量[112-113]. ...
Species richness effects on grassland recovery from drought depend on community productivity in a multisite experiment
1
2017
... 降水可以通过改变生物多样性影响生态系统功能[12],也可以通过改变土壤性质、植物光合速率、根系分泌物等影响生物量,从而改变生态系统功能[19].众所周知生物量是研究生态系统碳收支和碳循环的重要环节.生物量分配反映了地上资源(光和CO2)与地下资源(水和养分)之间的平衡,被广泛用作指示陆地生态系统碳循环对全球变化响应的关键指标.大量野外实地研究和文献综述表明,群落生物量和生产力与极端降水事件具有很强的相关性,例如极端干旱事件使得全球典型草地生产力降低了40%[108]、中国荒漠草原沙生针茅(S. caucasica)群落地上生物量降低了50%以上[73];改变了荒漠生态系统典型植物骆驼刺(Alhagi sparsifolia)的叶磷分配模式[38];降低了全世界范围内83个草地C3禾草、C4杂草的生物量[109]和温带草原生态系统[110]、半干旱沙质草地[111]的地上、地下生产力.季节性极端干旱对不同生态系统群落生物量也产生了一定影响,例如生长季初期的春季极端干旱和生长季中期的夏季极端干旱显著降低了温带草原生态系统和草甸草原的地上生物量[112-113]. ...
Effects of rainfall manipulation and nitrogen addition on plant biomass allocation in a semiarid sandy grassland
2
2020
... 降水可以通过改变生物多样性影响生态系统功能[12],也可以通过改变土壤性质、植物光合速率、根系分泌物等影响生物量,从而改变生态系统功能[19].众所周知生物量是研究生态系统碳收支和碳循环的重要环节.生物量分配反映了地上资源(光和CO2)与地下资源(水和养分)之间的平衡,被广泛用作指示陆地生态系统碳循环对全球变化响应的关键指标.大量野外实地研究和文献综述表明,群落生物量和生产力与极端降水事件具有很强的相关性,例如极端干旱事件使得全球典型草地生产力降低了40%[108]、中国荒漠草原沙生针茅(S. caucasica)群落地上生物量降低了50%以上[73];改变了荒漠生态系统典型植物骆驼刺(Alhagi sparsifolia)的叶磷分配模式[38];降低了全世界范围内83个草地C3禾草、C4杂草的生物量[109]和温带草原生态系统[110]、半干旱沙质草地[111]的地上、地下生产力.季节性极端干旱对不同生态系统群落生物量也产生了一定影响,例如生长季初期的春季极端干旱和生长季中期的夏季极端干旱显著降低了温带草原生态系统和草甸草原的地上生物量[112-113]. ...
... 干旱除了对群落生物量产生影响外,也会对生态系统碳循环、氮循环等产生不同程度的影响,进而影响生态系统的养分循环和水循环,对生态系统多功能性起关键作用[114-115].生长季极端干旱会减弱土壤微生物活动[103]、降低氮矿化速率[111]和凋落物生物量的碳氮密度[116].极端干旱会显著影响氮素的转化过程[117]、土壤碳稳定性以及土壤团聚体的碳封存功能[56]、土壤总碳和总氮含量[118],增加贝加尔针茅(S. baicalensis)草原植物叶片碳含量[25,119].极端干旱也能导致土壤动物功能群重组与效率失衡[120]:杂食性和腐食性类群减少[121],微生物互作网络受损,氮利用率低,养分循环阻滞,结构稳定性下降[122].土壤动物群落冗余度降低,抗干扰能力下降,植物病原菌繁殖加速,植物生产力显著降低[123].蚂蚁、蚯蚓、千足虫等清道夫数量减少,土壤沉积物、有机质垂直运输的能力变弱[124-125]. ...
Drought sensitivity of aboveground productivity in Leymus chinensis meadow steppe depends on drought timing
1
2019
... 降水可以通过改变生物多样性影响生态系统功能[12],也可以通过改变土壤性质、植物光合速率、根系分泌物等影响生物量,从而改变生态系统功能[19].众所周知生物量是研究生态系统碳收支和碳循环的重要环节.生物量分配反映了地上资源(光和CO2)与地下资源(水和养分)之间的平衡,被广泛用作指示陆地生态系统碳循环对全球变化响应的关键指标.大量野外实地研究和文献综述表明,群落生物量和生产力与极端降水事件具有很强的相关性,例如极端干旱事件使得全球典型草地生产力降低了40%[108]、中国荒漠草原沙生针茅(S. caucasica)群落地上生物量降低了50%以上[73];改变了荒漠生态系统典型植物骆驼刺(Alhagi sparsifolia)的叶磷分配模式[38];降低了全世界范围内83个草地C3禾草、C4杂草的生物量[109]和温带草原生态系统[110]、半干旱沙质草地[111]的地上、地下生产力.季节性极端干旱对不同生态系统群落生物量也产生了一定影响,例如生长季初期的春季极端干旱和生长季中期的夏季极端干旱显著降低了温带草原生态系统和草甸草原的地上生物量[112-113]. ...
Effects of extreme rainfall events are independent of plant species richness in an experimental grassland community
1
2019
... 降水可以通过改变生物多样性影响生态系统功能[12],也可以通过改变土壤性质、植物光合速率、根系分泌物等影响生物量,从而改变生态系统功能[19].众所周知生物量是研究生态系统碳收支和碳循环的重要环节.生物量分配反映了地上资源(光和CO2)与地下资源(水和养分)之间的平衡,被广泛用作指示陆地生态系统碳循环对全球变化响应的关键指标.大量野外实地研究和文献综述表明,群落生物量和生产力与极端降水事件具有很强的相关性,例如极端干旱事件使得全球典型草地生产力降低了40%[108]、中国荒漠草原沙生针茅(S. caucasica)群落地上生物量降低了50%以上[73];改变了荒漠生态系统典型植物骆驼刺(Alhagi sparsifolia)的叶磷分配模式[38];降低了全世界范围内83个草地C3禾草、C4杂草的生物量[109]和温带草原生态系统[110]、半干旱沙质草地[111]的地上、地下生产力.季节性极端干旱对不同生态系统群落生物量也产生了一定影响,例如生长季初期的春季极端干旱和生长季中期的夏季极端干旱显著降低了温带草原生态系统和草甸草原的地上生物量[112-113]. ...
极端气候事件对陆地生态系统碳循环的影响
1
2019
... 干旱除了对群落生物量产生影响外,也会对生态系统碳循环、氮循环等产生不同程度的影响,进而影响生态系统的养分循环和水循环,对生态系统多功能性起关键作用[114-115].生长季极端干旱会减弱土壤微生物活动[103]、降低氮矿化速率[111]和凋落物生物量的碳氮密度[116].极端干旱会显著影响氮素的转化过程[117]、土壤碳稳定性以及土壤团聚体的碳封存功能[56]、土壤总碳和总氮含量[118],增加贝加尔针茅(S. baicalensis)草原植物叶片碳含量[25,119].极端干旱也能导致土壤动物功能群重组与效率失衡[120]:杂食性和腐食性类群减少[121],微生物互作网络受损,氮利用率低,养分循环阻滞,结构稳定性下降[122].土壤动物群落冗余度降低,抗干扰能力下降,植物病原菌繁殖加速,植物生产力显著降低[123].蚂蚁、蚯蚓、千足虫等清道夫数量减少,土壤沉积物、有机质垂直运输的能力变弱[124-125]. ...
Net loss of biomass predicted for tropical biomes in a changing climate
1
2023
... 干旱除了对群落生物量产生影响外,也会对生态系统碳循环、氮循环等产生不同程度的影响,进而影响生态系统的养分循环和水循环,对生态系统多功能性起关键作用[114-115].生长季极端干旱会减弱土壤微生物活动[103]、降低氮矿化速率[111]和凋落物生物量的碳氮密度[116].极端干旱会显著影响氮素的转化过程[117]、土壤碳稳定性以及土壤团聚体的碳封存功能[56]、土壤总碳和总氮含量[118],增加贝加尔针茅(S. baicalensis)草原植物叶片碳含量[25,119].极端干旱也能导致土壤动物功能群重组与效率失衡[120]:杂食性和腐食性类群减少[121],微生物互作网络受损,氮利用率低,养分循环阻滞,结构稳定性下降[122].土壤动物群落冗余度降低,抗干扰能力下降,植物病原菌繁殖加速,植物生产力显著降低[123].蚂蚁、蚯蚓、千足虫等清道夫数量减少,土壤沉积物、有机质垂直运输的能力变弱[124-125]. ...
Effects of short-term drought,nitrogen application and their interactions on the composition and functional genes of soil microbial communities in alfalfa grassland on the Loess Plateau
1
2023
... 干旱除了对群落生物量产生影响外,也会对生态系统碳循环、氮循环等产生不同程度的影响,进而影响生态系统的养分循环和水循环,对生态系统多功能性起关键作用[114-115].生长季极端干旱会减弱土壤微生物活动[103]、降低氮矿化速率[111]和凋落物生物量的碳氮密度[116].极端干旱会显著影响氮素的转化过程[117]、土壤碳稳定性以及土壤团聚体的碳封存功能[56]、土壤总碳和总氮含量[118],增加贝加尔针茅(S. baicalensis)草原植物叶片碳含量[25,119].极端干旱也能导致土壤动物功能群重组与效率失衡[120]:杂食性和腐食性类群减少[121],微生物互作网络受损,氮利用率低,养分循环阻滞,结构稳定性下降[122].土壤动物群落冗余度降低,抗干扰能力下降,植物病原菌繁殖加速,植物生产力显著降低[123].蚂蚁、蚯蚓、千足虫等清道夫数量减少,土壤沉积物、有机质垂直运输的能力变弱[124-125]. ...
Effects of altered precipitation patterns on soil nitrogen transformation in different landscape types during the growing season in northern China
1
2023
... 干旱除了对群落生物量产生影响外,也会对生态系统碳循环、氮循环等产生不同程度的影响,进而影响生态系统的养分循环和水循环,对生态系统多功能性起关键作用[114-115].生长季极端干旱会减弱土壤微生物活动[103]、降低氮矿化速率[111]和凋落物生物量的碳氮密度[116].极端干旱会显著影响氮素的转化过程[117]、土壤碳稳定性以及土壤团聚体的碳封存功能[56]、土壤总碳和总氮含量[118],增加贝加尔针茅(S. baicalensis)草原植物叶片碳含量[25,119].极端干旱也能导致土壤动物功能群重组与效率失衡[120]:杂食性和腐食性类群减少[121],微生物互作网络受损,氮利用率低,养分循环阻滞,结构稳定性下降[122].土壤动物群落冗余度降低,抗干扰能力下降,植物病原菌繁殖加速,植物生产力显著降低[123].蚂蚁、蚯蚓、千足虫等清道夫数量减少,土壤沉积物、有机质垂直运输的能力变弱[124-125]. ...
Soil microbial community resistance to drought and links to C stabilization in an Australian grassland
1
2016
... 干旱除了对群落生物量产生影响外,也会对生态系统碳循环、氮循环等产生不同程度的影响,进而影响生态系统的养分循环和水循环,对生态系统多功能性起关键作用[114-115].生长季极端干旱会减弱土壤微生物活动[103]、降低氮矿化速率[111]和凋落物生物量的碳氮密度[116].极端干旱会显著影响氮素的转化过程[117]、土壤碳稳定性以及土壤团聚体的碳封存功能[56]、土壤总碳和总氮含量[118],增加贝加尔针茅(S. baicalensis)草原植物叶片碳含量[25,119].极端干旱也能导致土壤动物功能群重组与效率失衡[120]:杂食性和腐食性类群减少[121],微生物互作网络受损,氮利用率低,养分循环阻滞,结构稳定性下降[122].土壤动物群落冗余度降低,抗干扰能力下降,植物病原菌繁殖加速,植物生产力显著降低[123].蚂蚁、蚯蚓、千足虫等清道夫数量减少,土壤沉积物、有机质垂直运输的能力变弱[124-125]. ...
氮添加和干旱对呼伦贝尔草原5种植物性状的影响
1
2020
... 干旱除了对群落生物量产生影响外,也会对生态系统碳循环、氮循环等产生不同程度的影响,进而影响生态系统的养分循环和水循环,对生态系统多功能性起关键作用[114-115].生长季极端干旱会减弱土壤微生物活动[103]、降低氮矿化速率[111]和凋落物生物量的碳氮密度[116].极端干旱会显著影响氮素的转化过程[117]、土壤碳稳定性以及土壤团聚体的碳封存功能[56]、土壤总碳和总氮含量[118],增加贝加尔针茅(S. baicalensis)草原植物叶片碳含量[25,119].极端干旱也能导致土壤动物功能群重组与效率失衡[120]:杂食性和腐食性类群减少[121],微生物互作网络受损,氮利用率低,养分循环阻滞,结构稳定性下降[122].土壤动物群落冗余度降低,抗干扰能力下降,植物病原菌繁殖加速,植物生产力显著降低[123].蚂蚁、蚯蚓、千足虫等清道夫数量减少,土壤沉积物、有机质垂直运输的能力变弱[124-125]. ...
Conserving dryland biodiversity:a future vision of sustainable dryland development
1
2014
... 干旱除了对群落生物量产生影响外,也会对生态系统碳循环、氮循环等产生不同程度的影响,进而影响生态系统的养分循环和水循环,对生态系统多功能性起关键作用[114-115].生长季极端干旱会减弱土壤微生物活动[103]、降低氮矿化速率[111]和凋落物生物量的碳氮密度[116].极端干旱会显著影响氮素的转化过程[117]、土壤碳稳定性以及土壤团聚体的碳封存功能[56]、土壤总碳和总氮含量[118],增加贝加尔针茅(S. baicalensis)草原植物叶片碳含量[25,119].极端干旱也能导致土壤动物功能群重组与效率失衡[120]:杂食性和腐食性类群减少[121],微生物互作网络受损,氮利用率低,养分循环阻滞,结构稳定性下降[122].土壤动物群落冗余度降低,抗干扰能力下降,植物病原菌繁殖加速,植物生产力显著降低[123].蚂蚁、蚯蚓、千足虫等清道夫数量减少,土壤沉积物、有机质垂直运输的能力变弱[124-125]. ...
Soil microarthropods alter the outcome of plant-soil feedback experiments
1
2018
... 干旱除了对群落生物量产生影响外,也会对生态系统碳循环、氮循环等产生不同程度的影响,进而影响生态系统的养分循环和水循环,对生态系统多功能性起关键作用[114-115].生长季极端干旱会减弱土壤微生物活动[103]、降低氮矿化速率[111]和凋落物生物量的碳氮密度[116].极端干旱会显著影响氮素的转化过程[117]、土壤碳稳定性以及土壤团聚体的碳封存功能[56]、土壤总碳和总氮含量[118],增加贝加尔针茅(S. baicalensis)草原植物叶片碳含量[25,119].极端干旱也能导致土壤动物功能群重组与效率失衡[120]:杂食性和腐食性类群减少[121],微生物互作网络受损,氮利用率低,养分循环阻滞,结构稳定性下降[122].土壤动物群落冗余度降低,抗干扰能力下降,植物病原菌繁殖加速,植物生产力显著降低[123].蚂蚁、蚯蚓、千足虫等清道夫数量减少,土壤沉积物、有机质垂直运输的能力变弱[124-125]. ...
Soil fauna and its potential use in the ecological restoration of dryland ecosystems
1
2022
... 干旱除了对群落生物量产生影响外,也会对生态系统碳循环、氮循环等产生不同程度的影响,进而影响生态系统的养分循环和水循环,对生态系统多功能性起关键作用[114-115].生长季极端干旱会减弱土壤微生物活动[103]、降低氮矿化速率[111]和凋落物生物量的碳氮密度[116].极端干旱会显著影响氮素的转化过程[117]、土壤碳稳定性以及土壤团聚体的碳封存功能[56]、土壤总碳和总氮含量[118],增加贝加尔针茅(S. baicalensis)草原植物叶片碳含量[25,119].极端干旱也能导致土壤动物功能群重组与效率失衡[120]:杂食性和腐食性类群减少[121],微生物互作网络受损,氮利用率低,养分循环阻滞,结构稳定性下降[122].土壤动物群落冗余度降低,抗干扰能力下降,植物病原菌繁殖加速,植物生产力显著降低[123].蚂蚁、蚯蚓、千足虫等清道夫数量减少,土壤沉积物、有机质垂直运输的能力变弱[124-125]. ...
Biodiversity of soil animals and its function
1
2001
... 干旱除了对群落生物量产生影响外,也会对生态系统碳循环、氮循环等产生不同程度的影响,进而影响生态系统的养分循环和水循环,对生态系统多功能性起关键作用[114-115].生长季极端干旱会减弱土壤微生物活动[103]、降低氮矿化速率[111]和凋落物生物量的碳氮密度[116].极端干旱会显著影响氮素的转化过程[117]、土壤碳稳定性以及土壤团聚体的碳封存功能[56]、土壤总碳和总氮含量[118],增加贝加尔针茅(S. baicalensis)草原植物叶片碳含量[25,119].极端干旱也能导致土壤动物功能群重组与效率失衡[120]:杂食性和腐食性类群减少[121],微生物互作网络受损,氮利用率低,养分循环阻滞,结构稳定性下降[122].土壤动物群落冗余度降低,抗干扰能力下降,植物病原菌繁殖加速,植物生产力显著降低[123].蚂蚁、蚯蚓、千足虫等清道夫数量减少,土壤沉积物、有机质垂直运输的能力变弱[124-125]. ...
Increased soil aggregate stability is strongly correlated with root and soil properties along a gradient of secondary succession on the Loess Plateau
1
2020
... 干旱除了对群落生物量产生影响外,也会对生态系统碳循环、氮循环等产生不同程度的影响,进而影响生态系统的养分循环和水循环,对生态系统多功能性起关键作用[114-115].生长季极端干旱会减弱土壤微生物活动[103]、降低氮矿化速率[111]和凋落物生物量的碳氮密度[116].极端干旱会显著影响氮素的转化过程[117]、土壤碳稳定性以及土壤团聚体的碳封存功能[56]、土壤总碳和总氮含量[118],增加贝加尔针茅(S. baicalensis)草原植物叶片碳含量[25,119].极端干旱也能导致土壤动物功能群重组与效率失衡[120]:杂食性和腐食性类群减少[121],微生物互作网络受损,氮利用率低,养分循环阻滞,结构稳定性下降[122].土壤动物群落冗余度降低,抗干扰能力下降,植物病原菌繁殖加速,植物生产力显著降低[123].蚂蚁、蚯蚓、千足虫等清道夫数量减少,土壤沉积物、有机质垂直运输的能力变弱[124-125]. ...
Soil aggregation,ecosystem engineers and the C cycle
1
2020
... 干旱除了对群落生物量产生影响外,也会对生态系统碳循环、氮循环等产生不同程度的影响,进而影响生态系统的养分循环和水循环,对生态系统多功能性起关键作用[114-115].生长季极端干旱会减弱土壤微生物活动[103]、降低氮矿化速率[111]和凋落物生物量的碳氮密度[116].极端干旱会显著影响氮素的转化过程[117]、土壤碳稳定性以及土壤团聚体的碳封存功能[56]、土壤总碳和总氮含量[118],增加贝加尔针茅(S. baicalensis)草原植物叶片碳含量[25,119].极端干旱也能导致土壤动物功能群重组与效率失衡[120]:杂食性和腐食性类群减少[121],微生物互作网络受损,氮利用率低,养分循环阻滞,结构稳定性下降[122].土壤动物群落冗余度降低,抗干扰能力下降,植物病原菌繁殖加速,植物生产力显著降低[123].蚂蚁、蚯蚓、千足虫等清道夫数量减少,土壤沉积物、有机质垂直运输的能力变弱[124-125]. ...



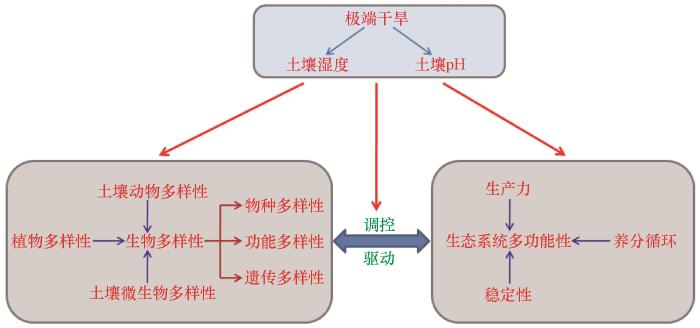

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}