AR6 Synthesis Report:Climate Change 2023
1
2023
... 全球变化的加剧正在深刻改变降雨格局与氮素水平,显著影响干旱区草地生态系统的结构与功能[1-3],并进一步反馈于区域生态安全与可持续发展[4-5].极端干旱和极端降雨事件的频率与强度显著增加,不仅重塑了草地群落结构,也改变了生态系统功能[4,6-7].工农业活动的加剧提升了大气与地表氮负荷,虽在一定程度上提高了草地生产力,但也造成了生物多样性与系统稳定性的下降[8-9].草地生物量作为反映植被动态和生态系统功能变化的重要指标[10],是揭示生态过程的核心参数.因而,在全球变化背景下,降雨格局和氮添加的改变可能通过影响物种间关系、群落组成与结构及土壤特性,进而对生物量、生态系统的结构与功能产生深远影响. ...
Anthropogenic climate and land-use change drive short-and long-term biodiversity shifts across taxa
0
2024
Nitrogen and water additions with or without mowing altered soil microbial community characteristics in a semi-arid steppe
1
2025
... 全球变化的加剧正在深刻改变降雨格局与氮素水平,显著影响干旱区草地生态系统的结构与功能[1-3],并进一步反馈于区域生态安全与可持续发展[4-5].极端干旱和极端降雨事件的频率与强度显著增加,不仅重塑了草地群落结构,也改变了生态系统功能[4,6-7].工农业活动的加剧提升了大气与地表氮负荷,虽在一定程度上提高了草地生产力,但也造成了生物多样性与系统稳定性的下降[8-9].草地生物量作为反映植被动态和生态系统功能变化的重要指标[10],是揭示生态过程的核心参数.因而,在全球变化背景下,降雨格局和氮添加的改变可能通过影响物种间关系、群落组成与结构及土壤特性,进而对生物量、生态系统的结构与功能产生深远影响. ...
Changes of extreme precipitation and nonlinear influence of climate variables over monsoon region in China
2
2017
... 全球变化的加剧正在深刻改变降雨格局与氮素水平,显著影响干旱区草地生态系统的结构与功能[1-3],并进一步反馈于区域生态安全与可持续发展[4-5].极端干旱和极端降雨事件的频率与强度显著增加,不仅重塑了草地群落结构,也改变了生态系统功能[4,6-7].工农业活动的加剧提升了大气与地表氮负荷,虽在一定程度上提高了草地生产力,但也造成了生物多样性与系统稳定性的下降[8-9].草地生物量作为反映植被动态和生态系统功能变化的重要指标[10],是揭示生态过程的核心参数.因而,在全球变化背景下,降雨格局和氮添加的改变可能通过影响物种间关系、群落组成与结构及土壤特性,进而对生物量、生态系统的结构与功能产生深远影响. ...
... [4,6-7].工农业活动的加剧提升了大气与地表氮负荷,虽在一定程度上提高了草地生产力,但也造成了生物多样性与系统稳定性的下降[8-9].草地生物量作为反映植被动态和生态系统功能变化的重要指标[10],是揭示生态过程的核心参数.因而,在全球变化背景下,降雨格局和氮添加的改变可能通过影响物种间关系、群落组成与结构及土壤特性,进而对生物量、生态系统的结构与功能产生深远影响. ...
1
2016
... 全球变化的加剧正在深刻改变降雨格局与氮素水平,显著影响干旱区草地生态系统的结构与功能[1-3],并进一步反馈于区域生态安全与可持续发展[4-5].极端干旱和极端降雨事件的频率与强度显著增加,不仅重塑了草地群落结构,也改变了生态系统功能[4,6-7].工农业活动的加剧提升了大气与地表氮负荷,虽在一定程度上提高了草地生产力,但也造成了生物多样性与系统稳定性的下降[8-9].草地生物量作为反映植被动态和生态系统功能变化的重要指标[10],是揭示生态过程的核心参数.因而,在全球变化背景下,降雨格局和氮添加的改变可能通过影响物种间关系、群落组成与结构及土壤特性,进而对生物量、生态系统的结构与功能产生深远影响. ...
IPCC AR6报告解读:极端天气气候事件变化
1
2021
... 全球变化的加剧正在深刻改变降雨格局与氮素水平,显著影响干旱区草地生态系统的结构与功能[1-3],并进一步反馈于区域生态安全与可持续发展[4-5].极端干旱和极端降雨事件的频率与强度显著增加,不仅重塑了草地群落结构,也改变了生态系统功能[4,6-7].工农业活动的加剧提升了大气与地表氮负荷,虽在一定程度上提高了草地生产力,但也造成了生物多样性与系统稳定性的下降[8-9].草地生物量作为反映植被动态和生态系统功能变化的重要指标[10],是揭示生态过程的核心参数.因而,在全球变化背景下,降雨格局和氮添加的改变可能通过影响物种间关系、群落组成与结构及土壤特性,进而对生物量、生态系统的结构与功能产生深远影响. ...
植物多样性与全球变化因素对松嫩草地土壤多功能性的影响与机制
1
2022
... 全球变化的加剧正在深刻改变降雨格局与氮素水平,显著影响干旱区草地生态系统的结构与功能[1-3],并进一步反馈于区域生态安全与可持续发展[4-5].极端干旱和极端降雨事件的频率与强度显著增加,不仅重塑了草地群落结构,也改变了生态系统功能[4,6-7].工农业活动的加剧提升了大气与地表氮负荷,虽在一定程度上提高了草地生产力,但也造成了生物多样性与系统稳定性的下降[8-9].草地生物量作为反映植被动态和生态系统功能变化的重要指标[10],是揭示生态过程的核心参数.因而,在全球变化背景下,降雨格局和氮添加的改变可能通过影响物种间关系、群落组成与结构及土壤特性,进而对生物量、生态系统的结构与功能产生深远影响. ...
Enhanced nitrogen deposition over China
1
2013
... 全球变化的加剧正在深刻改变降雨格局与氮素水平,显著影响干旱区草地生态系统的结构与功能[1-3],并进一步反馈于区域生态安全与可持续发展[4-5].极端干旱和极端降雨事件的频率与强度显著增加,不仅重塑了草地群落结构,也改变了生态系统功能[4,6-7].工农业活动的加剧提升了大气与地表氮负荷,虽在一定程度上提高了草地生产力,但也造成了生物多样性与系统稳定性的下降[8-9].草地生物量作为反映植被动态和生态系统功能变化的重要指标[10],是揭示生态过程的核心参数.因而,在全球变化背景下,降雨格局和氮添加的改变可能通过影响物种间关系、群落组成与结构及土壤特性,进而对生物量、生态系统的结构与功能产生深远影响. ...
Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands
1
2008
... 全球变化的加剧正在深刻改变降雨格局与氮素水平,显著影响干旱区草地生态系统的结构与功能[1-3],并进一步反馈于区域生态安全与可持续发展[4-5].极端干旱和极端降雨事件的频率与强度显著增加,不仅重塑了草地群落结构,也改变了生态系统功能[4,6-7].工农业活动的加剧提升了大气与地表氮负荷,虽在一定程度上提高了草地生产力,但也造成了生物多样性与系统稳定性的下降[8-9].草地生物量作为反映植被动态和生态系统功能变化的重要指标[10],是揭示生态过程的核心参数.因而,在全球变化背景下,降雨格局和氮添加的改变可能通过影响物种间关系、群落组成与结构及土壤特性,进而对生物量、生态系统的结构与功能产生深远影响. ...
半干旱沙地生境变化对植物地上生物量及其碳、氮储量的影响
1
2014
... 全球变化的加剧正在深刻改变降雨格局与氮素水平,显著影响干旱区草地生态系统的结构与功能[1-3],并进一步反馈于区域生态安全与可持续发展[4-5].极端干旱和极端降雨事件的频率与强度显著增加,不仅重塑了草地群落结构,也改变了生态系统功能[4,6-7].工农业活动的加剧提升了大气与地表氮负荷,虽在一定程度上提高了草地生产力,但也造成了生物多样性与系统稳定性的下降[8-9].草地生物量作为反映植被动态和生态系统功能变化的重要指标[10],是揭示生态过程的核心参数.因而,在全球变化背景下,降雨格局和氮添加的改变可能通过影响物种间关系、群落组成与结构及土壤特性,进而对生物量、生态系统的结构与功能产生深远影响. ...
科尔沁固定沙地植被特征对降雨变化的响应
1
2014
... 降雨变化对草地生态系统的调控作用体现为多维生态响应,涉及植物群落结构、土壤理化过程及地上生物量动态的综合调节.在植被方面,降雨增加通常有助于提高植被盖度和高度.强降雨事件能够迅速提升土壤水分含量,促进休眠种子萌发,加速群落发育过程,推动群落结构复杂化,提升物种多样性与系统稳定性[11].在土壤过程中,降雨变化通过调节水分状况,改变土壤气液相比例与理化性质,进而驱动养分循环的动态变化[12].然而,不同区域对降雨变化的响应存在差异.在内蒙古典型草原和荒漠草原,降雨增加显著提升地上生物量;而在科尔沁沙地和黄土高原的草原-荒漠过渡带,过多的降雨反而可能抑制植物生长[13],原因可能在于超出生态适宜范围的水分过剩引发根系通气受阻、营养淋溶或土壤结构破坏[14]. ...
Effects of precipitation and restoration vegetation on soil erosion in a semi-arid environment in the Loess Plateau,China
1
2016
... 降雨变化对草地生态系统的调控作用体现为多维生态响应,涉及植物群落结构、土壤理化过程及地上生物量动态的综合调节.在植被方面,降雨增加通常有助于提高植被盖度和高度.强降雨事件能够迅速提升土壤水分含量,促进休眠种子萌发,加速群落发育过程,推动群落结构复杂化,提升物种多样性与系统稳定性[11].在土壤过程中,降雨变化通过调节水分状况,改变土壤气液相比例与理化性质,进而驱动养分循环的动态变化[12].然而,不同区域对降雨变化的响应存在差异.在内蒙古典型草原和荒漠草原,降雨增加显著提升地上生物量;而在科尔沁沙地和黄土高原的草原-荒漠过渡带,过多的降雨反而可能抑制植物生长[13],原因可能在于超出生态适宜范围的水分过剩引发根系通气受阻、营养淋溶或土壤结构破坏[14]. ...
黄土高原4种植被带草本群落特征及其对降水变化的响应
1
2019
... 降雨变化对草地生态系统的调控作用体现为多维生态响应,涉及植物群落结构、土壤理化过程及地上生物量动态的综合调节.在植被方面,降雨增加通常有助于提高植被盖度和高度.强降雨事件能够迅速提升土壤水分含量,促进休眠种子萌发,加速群落发育过程,推动群落结构复杂化,提升物种多样性与系统稳定性[11].在土壤过程中,降雨变化通过调节水分状况,改变土壤气液相比例与理化性质,进而驱动养分循环的动态变化[12].然而,不同区域对降雨变化的响应存在差异.在内蒙古典型草原和荒漠草原,降雨增加显著提升地上生物量;而在科尔沁沙地和黄土高原的草原-荒漠过渡带,过多的降雨反而可能抑制植物生长[13],原因可能在于超出生态适宜范围的水分过剩引发根系通气受阻、营养淋溶或土壤结构破坏[14]. ...
模拟短期降雨量变化对准噶尔荒漠植物群落的影响
1
2019
... 降雨变化对草地生态系统的调控作用体现为多维生态响应,涉及植物群落结构、土壤理化过程及地上生物量动态的综合调节.在植被方面,降雨增加通常有助于提高植被盖度和高度.强降雨事件能够迅速提升土壤水分含量,促进休眠种子萌发,加速群落发育过程,推动群落结构复杂化,提升物种多样性与系统稳定性[11].在土壤过程中,降雨变化通过调节水分状况,改变土壤气液相比例与理化性质,进而驱动养分循环的动态变化[12].然而,不同区域对降雨变化的响应存在差异.在内蒙古典型草原和荒漠草原,降雨增加显著提升地上生物量;而在科尔沁沙地和黄土高原的草原-荒漠过渡带,过多的降雨反而可能抑制植物生长[13],原因可能在于超出生态适宜范围的水分过剩引发根系通气受阻、营养淋溶或土壤结构破坏[14]. ...
Response of soil nutrients and stoichiometry to elevated nitrogen deposition in alpine grassland on the Qinghai-Tibetan Plateau
1
2019
... 氮添加通过改变土壤酸碱性和调控微生物呼吸速率,进而影响土壤酶活性[15-17],最终作用于生态系统功能.全球范围的研究普遍认为,氮添加会导致草地植物物种多样性下降.其主要机制包括:①生态位压缩:氮添加减少生态位维度,加剧物种竞争,最终导致物种丧失;②幼苗更新受阻:氮添加增加凋落物积累,抑制幼苗萌发和存活;③土壤化学环境变化:长期或高浓度氮添加可能引发土壤酸化或铵毒害,抑制植物生长[18].然而,氮添加对物种多样性的影响具有环境依赖性,主要受氮素水平、施加持续时间、植物氮素利用效率、土壤养分状况及水热条件等因素共同调控,在特定条件下可能出现与上述趋势相悖的结果[19].关于氮添加对植物生产力的影响,大多数研究表明,适量氮添加可促进植物生长,提高生态系统生产力,但当氮添加超过植物的需求阈值时,可能对生态系统产生负面效应[20].部分研究发现,过量氮添加可能降低植物的氮素利用效率,未能显著提升生产力[21-22].此外,氮添加对地下生物量的影响仍存在争议,不同研究得出了相互矛盾的结论.有研究表明,氮添加能促进叶片生长,使植物将更多资源分配至地上部分,导致地下生物量减少[23];而另一些研究则认为,氮添加有助于根系发育从而增加地下生物量[24].因此,氮添加是否以及如何通过调节草地生态系统中植被与土壤结构,进而影响其生产力仍不明确.半干旱区草地生态系统具有较强的养分限制性和较高的生态敏感性,是研究氮添加生态效应的重要区域.在该地区开展氮添加情景下的系统响应研究,有助于深入理解草地生态系统对氮添加的多维响应机制,并为干旱区生态系统的管理与可持续利用提供科学依据. ...
Soil abiotic properties and plant functional diversity co-regulate the impacts of nitrogen addition on ecosystem multifunctionality in an alpine meadow
0
2021
Effects of nitrogen enrichment on soil enzyme activities in grassland ecosystems in China:a multilevel meta-analysis
1
2025
... 氮添加通过改变土壤酸碱性和调控微生物呼吸速率,进而影响土壤酶活性[15-17],最终作用于生态系统功能.全球范围的研究普遍认为,氮添加会导致草地植物物种多样性下降.其主要机制包括:①生态位压缩:氮添加减少生态位维度,加剧物种竞争,最终导致物种丧失;②幼苗更新受阻:氮添加增加凋落物积累,抑制幼苗萌发和存活;③土壤化学环境变化:长期或高浓度氮添加可能引发土壤酸化或铵毒害,抑制植物生长[18].然而,氮添加对物种多样性的影响具有环境依赖性,主要受氮素水平、施加持续时间、植物氮素利用效率、土壤养分状况及水热条件等因素共同调控,在特定条件下可能出现与上述趋势相悖的结果[19].关于氮添加对植物生产力的影响,大多数研究表明,适量氮添加可促进植物生长,提高生态系统生产力,但当氮添加超过植物的需求阈值时,可能对生态系统产生负面效应[20].部分研究发现,过量氮添加可能降低植物的氮素利用效率,未能显著提升生产力[21-22].此外,氮添加对地下生物量的影响仍存在争议,不同研究得出了相互矛盾的结论.有研究表明,氮添加能促进叶片生长,使植物将更多资源分配至地上部分,导致地下生物量减少[23];而另一些研究则认为,氮添加有助于根系发育从而增加地下生物量[24].因此,氮添加是否以及如何通过调节草地生态系统中植被与土壤结构,进而影响其生产力仍不明确.半干旱区草地生态系统具有较强的养分限制性和较高的生态敏感性,是研究氮添加生态效应的重要区域.在该地区开展氮添加情景下的系统响应研究,有助于深入理解草地生态系统对氮添加的多维响应机制,并为干旱区生态系统的管理与可持续利用提供科学依据. ...
Addition of multiple limiting resources reduces grassland diversity
1
2016
... 氮添加通过改变土壤酸碱性和调控微生物呼吸速率,进而影响土壤酶活性[15-17],最终作用于生态系统功能.全球范围的研究普遍认为,氮添加会导致草地植物物种多样性下降.其主要机制包括:①生态位压缩:氮添加减少生态位维度,加剧物种竞争,最终导致物种丧失;②幼苗更新受阻:氮添加增加凋落物积累,抑制幼苗萌发和存活;③土壤化学环境变化:长期或高浓度氮添加可能引发土壤酸化或铵毒害,抑制植物生长[18].然而,氮添加对物种多样性的影响具有环境依赖性,主要受氮素水平、施加持续时间、植物氮素利用效率、土壤养分状况及水热条件等因素共同调控,在特定条件下可能出现与上述趋势相悖的结果[19].关于氮添加对植物生产力的影响,大多数研究表明,适量氮添加可促进植物生长,提高生态系统生产力,但当氮添加超过植物的需求阈值时,可能对生态系统产生负面效应[20].部分研究发现,过量氮添加可能降低植物的氮素利用效率,未能显著提升生产力[21-22].此外,氮添加对地下生物量的影响仍存在争议,不同研究得出了相互矛盾的结论.有研究表明,氮添加能促进叶片生长,使植物将更多资源分配至地上部分,导致地下生物量减少[23];而另一些研究则认为,氮添加有助于根系发育从而增加地下生物量[24].因此,氮添加是否以及如何通过调节草地生态系统中植被与土壤结构,进而影响其生产力仍不明确.半干旱区草地生态系统具有较强的养分限制性和较高的生态敏感性,是研究氮添加生态效应的重要区域.在该地区开展氮添加情景下的系统响应研究,有助于深入理解草地生态系统对氮添加的多维响应机制,并为干旱区生态系统的管理与可持续利用提供科学依据. ...
Changes and drivers of vegetation productivity in China's drylands under climate change
1
2024
... 氮添加通过改变土壤酸碱性和调控微生物呼吸速率,进而影响土壤酶活性[15-17],最终作用于生态系统功能.全球范围的研究普遍认为,氮添加会导致草地植物物种多样性下降.其主要机制包括:①生态位压缩:氮添加减少生态位维度,加剧物种竞争,最终导致物种丧失;②幼苗更新受阻:氮添加增加凋落物积累,抑制幼苗萌发和存活;③土壤化学环境变化:长期或高浓度氮添加可能引发土壤酸化或铵毒害,抑制植物生长[18].然而,氮添加对物种多样性的影响具有环境依赖性,主要受氮素水平、施加持续时间、植物氮素利用效率、土壤养分状况及水热条件等因素共同调控,在特定条件下可能出现与上述趋势相悖的结果[19].关于氮添加对植物生产力的影响,大多数研究表明,适量氮添加可促进植物生长,提高生态系统生产力,但当氮添加超过植物的需求阈值时,可能对生态系统产生负面效应[20].部分研究发现,过量氮添加可能降低植物的氮素利用效率,未能显著提升生产力[21-22].此外,氮添加对地下生物量的影响仍存在争议,不同研究得出了相互矛盾的结论.有研究表明,氮添加能促进叶片生长,使植物将更多资源分配至地上部分,导致地下生物量减少[23];而另一些研究则认为,氮添加有助于根系发育从而增加地下生物量[24].因此,氮添加是否以及如何通过调节草地生态系统中植被与土壤结构,进而影响其生产力仍不明确.半干旱区草地生态系统具有较强的养分限制性和较高的生态敏感性,是研究氮添加生态效应的重要区域.在该地区开展氮添加情景下的系统响应研究,有助于深入理解草地生态系统对氮添加的多维响应机制,并为干旱区生态系统的管理与可持续利用提供科学依据. ...
内蒙古荒漠草原植物群落结构和功能对增温和氮素添加的响应
2
2014
... 氮添加通过改变土壤酸碱性和调控微生物呼吸速率,进而影响土壤酶活性[15-17],最终作用于生态系统功能.全球范围的研究普遍认为,氮添加会导致草地植物物种多样性下降.其主要机制包括:①生态位压缩:氮添加减少生态位维度,加剧物种竞争,最终导致物种丧失;②幼苗更新受阻:氮添加增加凋落物积累,抑制幼苗萌发和存活;③土壤化学环境变化:长期或高浓度氮添加可能引发土壤酸化或铵毒害,抑制植物生长[18].然而,氮添加对物种多样性的影响具有环境依赖性,主要受氮素水平、施加持续时间、植物氮素利用效率、土壤养分状况及水热条件等因素共同调控,在特定条件下可能出现与上述趋势相悖的结果[19].关于氮添加对植物生产力的影响,大多数研究表明,适量氮添加可促进植物生长,提高生态系统生产力,但当氮添加超过植物的需求阈值时,可能对生态系统产生负面效应[20].部分研究发现,过量氮添加可能降低植物的氮素利用效率,未能显著提升生产力[21-22].此外,氮添加对地下生物量的影响仍存在争议,不同研究得出了相互矛盾的结论.有研究表明,氮添加能促进叶片生长,使植物将更多资源分配至地上部分,导致地下生物量减少[23];而另一些研究则认为,氮添加有助于根系发育从而增加地下生物量[24].因此,氮添加是否以及如何通过调节草地生态系统中植被与土壤结构,进而影响其生产力仍不明确.半干旱区草地生态系统具有较强的养分限制性和较高的生态敏感性,是研究氮添加生态效应的重要区域.在该地区开展氮添加情景下的系统响应研究,有助于深入理解草地生态系统对氮添加的多维响应机制,并为干旱区生态系统的管理与可持续利用提供科学依据. ...
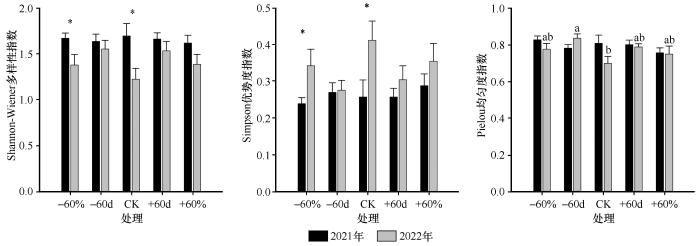
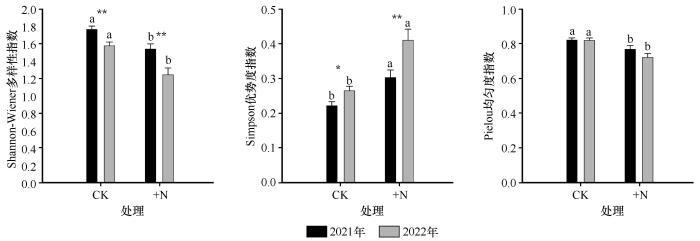
... 本研究发现,氮添加显著提高了植被盖度,这一结果与以往研究中氮添加促进植物群落盖度的结论一致[35].大量研究表明,氮素输入通常能够促进地上部分植被的生长,增加生物量[36].然而,氮添加也可能对群落结构和生态系统稳定性产生负面影响[20].例如,部分研究指出,氮添加会导致稀有物种逐渐消失,从而引发物种多样性下降和生态系统稳定性减弱[37];也有研究认为,养分输入的增加并不必然导致多样性的变化,具体结果受到区域水热条件、初始土壤养分状况和植物氮利用效率等因素的影响[38].本研究中,氮添加在显著提高群落总生物量的同时,也导致了物种多样性的下降.其主要机制在于氮素输入不仅促进了地上部分植物的生长,提高了整体生物量,还有助于根系的发育,从而增加了地下生物量[28].然而,氮添加也改变了植物间的竞争格局,增强了优势种的竞争能力,导致稀有物种逐渐被排挤出群落,最终引发群落多样性的下降[39-40].此外,氮添加降低了Shannon-Wiener多样性指数,提升了Simpson优势度指数,并降低了Pielou均匀度指数,反映出群落结构趋于单一化[41].其主要原因在于氮素输入改变了资源分配格局,强化了群落内的资源竞争.生长迅速、竞争能力强的优势种(如高大禾本科植物)对氮素具有更高的响应性,在氮添加条件下能够迅速扩张,占据更多的空间与资源,从而抑制其他物种的生存与生长,导致群落中物种数量减少、结构不均.随着优势种生物量的大幅增加,其高大的植株逐渐形成上层冠层,遮蔽了下层光照,限制了矮小或生长缓慢物种的光合作用和空间获取能力.这种垂直层级上的“光竞争”进一步抑制了弱势物种的生长,降低了群落物种在资源利用和空间分布上的均衡性,最终表现为Pielou均匀度指数的显著下降[42]. ...
Nitrogen deposition and its ecological impact in China:an overview
1
2011
... 氮添加通过改变土壤酸碱性和调控微生物呼吸速率,进而影响土壤酶活性[15-17],最终作用于生态系统功能.全球范围的研究普遍认为,氮添加会导致草地植物物种多样性下降.其主要机制包括:①生态位压缩:氮添加减少生态位维度,加剧物种竞争,最终导致物种丧失;②幼苗更新受阻:氮添加增加凋落物积累,抑制幼苗萌发和存活;③土壤化学环境变化:长期或高浓度氮添加可能引发土壤酸化或铵毒害,抑制植物生长[18].然而,氮添加对物种多样性的影响具有环境依赖性,主要受氮素水平、施加持续时间、植物氮素利用效率、土壤养分状况及水热条件等因素共同调控,在特定条件下可能出现与上述趋势相悖的结果[19].关于氮添加对植物生产力的影响,大多数研究表明,适量氮添加可促进植物生长,提高生态系统生产力,但当氮添加超过植物的需求阈值时,可能对生态系统产生负面效应[20].部分研究发现,过量氮添加可能降低植物的氮素利用效率,未能显著提升生产力[21-22].此外,氮添加对地下生物量的影响仍存在争议,不同研究得出了相互矛盾的结论.有研究表明,氮添加能促进叶片生长,使植物将更多资源分配至地上部分,导致地下生物量减少[23];而另一些研究则认为,氮添加有助于根系发育从而增加地下生物量[24].因此,氮添加是否以及如何通过调节草地生态系统中植被与土壤结构,进而影响其生产力仍不明确.半干旱区草地生态系统具有较强的养分限制性和较高的生态敏感性,是研究氮添加生态效应的重要区域.在该地区开展氮添加情景下的系统响应研究,有助于深入理解草地生态系统对氮添加的多维响应机制,并为干旱区生态系统的管理与可持续利用提供科学依据. ...
Linkages of stoichiometric imbalances to soil microbial respiration with increasing nitrogen addition:evidence from a long-term grassland experiment
1
2019
... 氮添加通过改变土壤酸碱性和调控微生物呼吸速率,进而影响土壤酶活性[15-17],最终作用于生态系统功能.全球范围的研究普遍认为,氮添加会导致草地植物物种多样性下降.其主要机制包括:①生态位压缩:氮添加减少生态位维度,加剧物种竞争,最终导致物种丧失;②幼苗更新受阻:氮添加增加凋落物积累,抑制幼苗萌发和存活;③土壤化学环境变化:长期或高浓度氮添加可能引发土壤酸化或铵毒害,抑制植物生长[18].然而,氮添加对物种多样性的影响具有环境依赖性,主要受氮素水平、施加持续时间、植物氮素利用效率、土壤养分状况及水热条件等因素共同调控,在特定条件下可能出现与上述趋势相悖的结果[19].关于氮添加对植物生产力的影响,大多数研究表明,适量氮添加可促进植物生长,提高生态系统生产力,但当氮添加超过植物的需求阈值时,可能对生态系统产生负面效应[20].部分研究发现,过量氮添加可能降低植物的氮素利用效率,未能显著提升生产力[21-22].此外,氮添加对地下生物量的影响仍存在争议,不同研究得出了相互矛盾的结论.有研究表明,氮添加能促进叶片生长,使植物将更多资源分配至地上部分,导致地下生物量减少[23];而另一些研究则认为,氮添加有助于根系发育从而增加地下生物量[24].因此,氮添加是否以及如何通过调节草地生态系统中植被与土壤结构,进而影响其生产力仍不明确.半干旱区草地生态系统具有较强的养分限制性和较高的生态敏感性,是研究氮添加生态效应的重要区域.在该地区开展氮添加情景下的系统响应研究,有助于深入理解草地生态系统对氮添加的多维响应机制,并为干旱区生态系统的管理与可持续利用提供科学依据. ...
养分添加对内蒙古草原生物量和多样性的影响
1
2014
... 氮添加通过改变土壤酸碱性和调控微生物呼吸速率,进而影响土壤酶活性[15-17],最终作用于生态系统功能.全球范围的研究普遍认为,氮添加会导致草地植物物种多样性下降.其主要机制包括:①生态位压缩:氮添加减少生态位维度,加剧物种竞争,最终导致物种丧失;②幼苗更新受阻:氮添加增加凋落物积累,抑制幼苗萌发和存活;③土壤化学环境变化:长期或高浓度氮添加可能引发土壤酸化或铵毒害,抑制植物生长[18].然而,氮添加对物种多样性的影响具有环境依赖性,主要受氮素水平、施加持续时间、植物氮素利用效率、土壤养分状况及水热条件等因素共同调控,在特定条件下可能出现与上述趋势相悖的结果[19].关于氮添加对植物生产力的影响,大多数研究表明,适量氮添加可促进植物生长,提高生态系统生产力,但当氮添加超过植物的需求阈值时,可能对生态系统产生负面效应[20].部分研究发现,过量氮添加可能降低植物的氮素利用效率,未能显著提升生产力[21-22].此外,氮添加对地下生物量的影响仍存在争议,不同研究得出了相互矛盾的结论.有研究表明,氮添加能促进叶片生长,使植物将更多资源分配至地上部分,导致地下生物量减少[23];而另一些研究则认为,氮添加有助于根系发育从而增加地下生物量[24].因此,氮添加是否以及如何通过调节草地生态系统中植被与土壤结构,进而影响其生产力仍不明确.半干旱区草地生态系统具有较强的养分限制性和较高的生态敏感性,是研究氮添加生态效应的重要区域.在该地区开展氮添加情景下的系统响应研究,有助于深入理解草地生态系统对氮添加的多维响应机制,并为干旱区生态系统的管理与可持续利用提供科学依据. ...
氮、磷添加对亚高山草甸地上/地下生物量分配及植物功能群组成的影响
1
2011
... 氮添加通过改变土壤酸碱性和调控微生物呼吸速率,进而影响土壤酶活性[15-17],最终作用于生态系统功能.全球范围的研究普遍认为,氮添加会导致草地植物物种多样性下降.其主要机制包括:①生态位压缩:氮添加减少生态位维度,加剧物种竞争,最终导致物种丧失;②幼苗更新受阻:氮添加增加凋落物积累,抑制幼苗萌发和存活;③土壤化学环境变化:长期或高浓度氮添加可能引发土壤酸化或铵毒害,抑制植物生长[18].然而,氮添加对物种多样性的影响具有环境依赖性,主要受氮素水平、施加持续时间、植物氮素利用效率、土壤养分状况及水热条件等因素共同调控,在特定条件下可能出现与上述趋势相悖的结果[19].关于氮添加对植物生产力的影响,大多数研究表明,适量氮添加可促进植物生长,提高生态系统生产力,但当氮添加超过植物的需求阈值时,可能对生态系统产生负面效应[20].部分研究发现,过量氮添加可能降低植物的氮素利用效率,未能显著提升生产力[21-22].此外,氮添加对地下生物量的影响仍存在争议,不同研究得出了相互矛盾的结论.有研究表明,氮添加能促进叶片生长,使植物将更多资源分配至地上部分,导致地下生物量减少[23];而另一些研究则认为,氮添加有助于根系发育从而增加地下生物量[24].因此,氮添加是否以及如何通过调节草地生态系统中植被与土壤结构,进而影响其生产力仍不明确.半干旱区草地生态系统具有较强的养分限制性和较高的生态敏感性,是研究氮添加生态效应的重要区域.在该地区开展氮添加情景下的系统响应研究,有助于深入理解草地生态系统对氮添加的多维响应机制,并为干旱区生态系统的管理与可持续利用提供科学依据. ...
科尔沁沙地植被恢复系列上群落演替与物种多样性的恢复动态
1
2004
... 科尔沁沙地位于中国北方半干旱地区的农牧交错带东南端,是中国北方地区重要的生态屏障.沙质草地是该区域分布最广的典型生态生境,具有代表性强、生态过程敏感等特征,是反映科尔沁沙地沙漠化进程的重要景观指标[25].近年来,剧烈的气候变化和人类活动深刻改变了该区域草地的地上/地下群落结构、物种组成、土壤特性及主要生态过程,显著削弱了区域生态系统的服务功能[26].在这一生态脆弱区,水分和氮素是影响草地发育的关键限制性资源,降雨变化和氮添加通过累加效应、协同效应或拮抗效应共同作用于草地生态系统[27].然而,目前针对该区域草地生态系统对水、氮多要素干扰的植被特征、土壤特性与生产力的相互作用、生产力维持机制的认识仍然十分有限.沙质草地是该区域分布最广的典型生态生境,是反映科尔沁沙地沙漠化进程的关键景观特征.因此,本研究以沙质草地为研究对象,结合野外样带调查与原位模拟实验,对降雨、氮添加两种因素进行控制研究,系统解析半干旱沙质草地在多重环境胁迫下植被、土壤对生产力的调控机理. ...
Plant composition change mediates climate drought,nitrogen addition,and grazing effects on soil net nitrogen mineralization in a semi-arid grassland in North China
1
2024
... 科尔沁沙地位于中国北方半干旱地区的农牧交错带东南端,是中国北方地区重要的生态屏障.沙质草地是该区域分布最广的典型生态生境,具有代表性强、生态过程敏感等特征,是反映科尔沁沙地沙漠化进程的重要景观指标[25].近年来,剧烈的气候变化和人类活动深刻改变了该区域草地的地上/地下群落结构、物种组成、土壤特性及主要生态过程,显著削弱了区域生态系统的服务功能[26].在这一生态脆弱区,水分和氮素是影响草地发育的关键限制性资源,降雨变化和氮添加通过累加效应、协同效应或拮抗效应共同作用于草地生态系统[27].然而,目前针对该区域草地生态系统对水、氮多要素干扰的植被特征、土壤特性与生产力的相互作用、生产力维持机制的认识仍然十分有限.沙质草地是该区域分布最广的典型生态生境,是反映科尔沁沙地沙漠化进程的关键景观特征.因此,本研究以沙质草地为研究对象,结合野外样带调查与原位模拟实验,对降雨、氮添加两种因素进行控制研究,系统解析半干旱沙质草地在多重环境胁迫下植被、土壤对生产力的调控机理. ...
沙质草地植物功能性状对放牧,增水,氮添加及其耦合效应的响应机制
1
2021
... 科尔沁沙地位于中国北方半干旱地区的农牧交错带东南端,是中国北方地区重要的生态屏障.沙质草地是该区域分布最广的典型生态生境,具有代表性强、生态过程敏感等特征,是反映科尔沁沙地沙漠化进程的重要景观指标[25].近年来,剧烈的气候变化和人类活动深刻改变了该区域草地的地上/地下群落结构、物种组成、土壤特性及主要生态过程,显著削弱了区域生态系统的服务功能[26].在这一生态脆弱区,水分和氮素是影响草地发育的关键限制性资源,降雨变化和氮添加通过累加效应、协同效应或拮抗效应共同作用于草地生态系统[27].然而,目前针对该区域草地生态系统对水、氮多要素干扰的植被特征、土壤特性与生产力的相互作用、生产力维持机制的认识仍然十分有限.沙质草地是该区域分布最广的典型生态生境,是反映科尔沁沙地沙漠化进程的关键景观特征.因此,本研究以沙质草地为研究对象,结合野外样带调查与原位模拟实验,对降雨、氮添加两种因素进行控制研究,系统解析半干旱沙质草地在多重环境胁迫下植被、土壤对生产力的调控机理. ...
土壤酸碱性对植物生长影响的研究进展
2
2020
... 本研究采用国际通行的NH₄NO₃(硝酸铵)溶液形式,于每年5月和7月分别施加氮10 g·m-2,年施氮总量为20 g·m-2.该施氮水平的设定综合考虑了试验区生态环境条件与草地恢复的实际需求:一方面,区域土壤为碱性沙质土壤,在碱性条件下,硝化作用受抑制,氮素转化效率降低,易加剧氨挥发损失,从而进一步限制植物对氮素的有效吸收[28];另一方面,土壤养分普遍匮乏,尤其氮素显著不足,严重制约了生态系统的生产力与恢复潜力.已有调查显示,科尔沁沙地表层(0~20 cm)土壤总氮含量仅为0.9 g·kg-1,远低于全国(1.9 g·kg-1)和全球(4.1 g·kg-1)平均水平[29],难以支撑退化草地的自然恢复进程.基于此,本研究选用20 g·m2·a-1的氮添加水平,既体现了区域氮限制的实际情况,也处于生态恢复推荐范围内,有助于提升土壤养分供给、改善植被生长环境,进而推动退化草地生态系统功能的重建与恢复. ...
... 本研究发现,氮添加显著提高了植被盖度,这一结果与以往研究中氮添加促进植物群落盖度的结论一致[35].大量研究表明,氮素输入通常能够促进地上部分植被的生长,增加生物量[36].然而,氮添加也可能对群落结构和生态系统稳定性产生负面影响[20].例如,部分研究指出,氮添加会导致稀有物种逐渐消失,从而引发物种多样性下降和生态系统稳定性减弱[37];也有研究认为,养分输入的增加并不必然导致多样性的变化,具体结果受到区域水热条件、初始土壤养分状况和植物氮利用效率等因素的影响[38].本研究中,氮添加在显著提高群落总生物量的同时,也导致了物种多样性的下降.其主要机制在于氮素输入不仅促进了地上部分植物的生长,提高了整体生物量,还有助于根系的发育,从而增加了地下生物量[28].然而,氮添加也改变了植物间的竞争格局,增强了优势种的竞争能力,导致稀有物种逐渐被排挤出群落,最终引发群落多样性的下降[39-40].此外,氮添加降低了Shannon-Wiener多样性指数,提升了Simpson优势度指数,并降低了Pielou均匀度指数,反映出群落结构趋于单一化[41].其主要原因在于氮素输入改变了资源分配格局,强化了群落内的资源竞争.生长迅速、竞争能力强的优势种(如高大禾本科植物)对氮素具有更高的响应性,在氮添加条件下能够迅速扩张,占据更多的空间与资源,从而抑制其他物种的生存与生长,导致群落中物种数量减少、结构不均.随着优势种生物量的大幅增加,其高大的植株逐渐形成上层冠层,遮蔽了下层光照,限制了矮小或生长缓慢物种的光合作用和空间获取能力.这种垂直层级上的“光竞争”进一步抑制了弱势物种的生长,降低了群落物种在资源利用和空间分布上的均衡性,最终表现为Pielou均匀度指数的显著下降[42]. ...
Spatial patterns of soil stoichiometry and their responses to land use in a desertified area:a case study of China's Horqin Sandy Land
1
2024
... 本研究采用国际通行的NH₄NO₃(硝酸铵)溶液形式,于每年5月和7月分别施加氮10 g·m-2,年施氮总量为20 g·m-2.该施氮水平的设定综合考虑了试验区生态环境条件与草地恢复的实际需求:一方面,区域土壤为碱性沙质土壤,在碱性条件下,硝化作用受抑制,氮素转化效率降低,易加剧氨挥发损失,从而进一步限制植物对氮素的有效吸收[28];另一方面,土壤养分普遍匮乏,尤其氮素显著不足,严重制约了生态系统的生产力与恢复潜力.已有调查显示,科尔沁沙地表层(0~20 cm)土壤总氮含量仅为0.9 g·kg-1,远低于全国(1.9 g·kg-1)和全球(4.1 g·kg-1)平均水平[29],难以支撑退化草地的自然恢复进程.基于此,本研究选用20 g·m2·a-1的氮添加水平,既体现了区域氮限制的实际情况,也处于生态恢复推荐范围内,有助于提升土壤养分供给、改善植被生长环境,进而推动退化草地生态系统功能的重建与恢复. ...
水、氮施用量对科尔沁沙地植被特征和叶性状的影响
1
2020
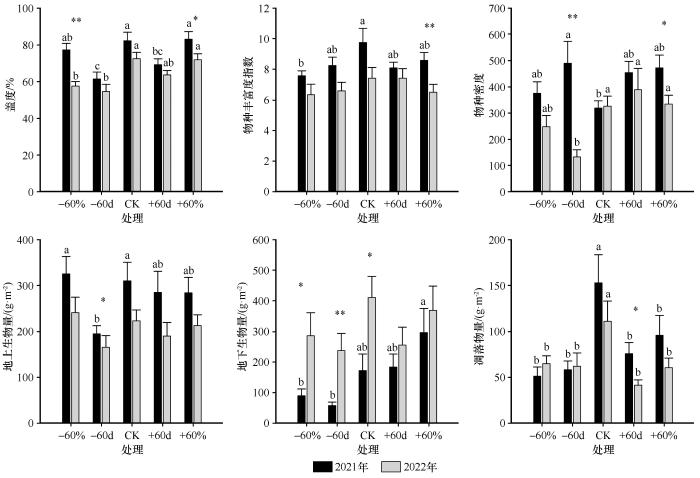
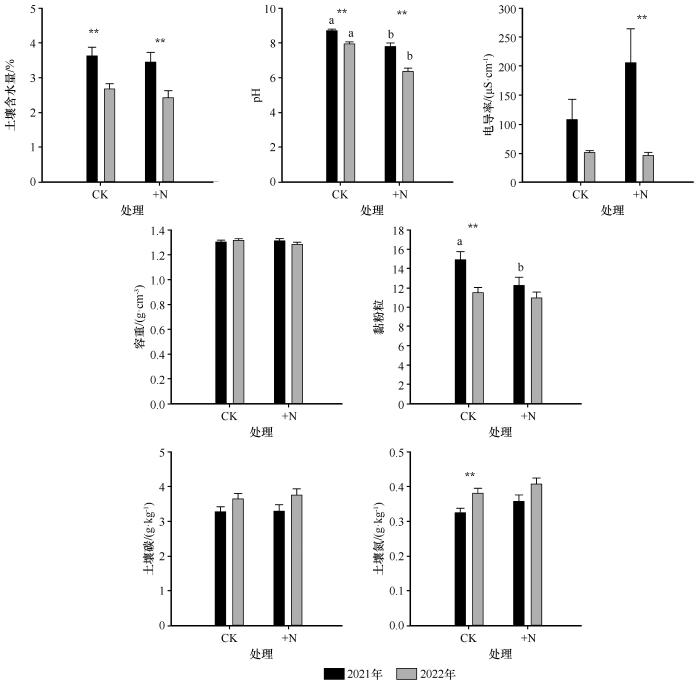
... 2021年和2022年,降雨变化对植被群落特征的影响表现出显著的年度差异.与已有研究中生长季降雨减少会降低植被盖度的结论一致[30],本研究发现,生长季前期(5—6月)持续干旱处理(-60d)在两年中均显著降低了植被盖度.然而,对于整个生长季减雨处理(-60%),其影响在两年间存在显著差异:2021年该处理对盖度无显著影响,而2022年则显著降低了盖度.该差异可能归因于两年降雨的时空分布不同.2021年7月降雨量高达159 mm,即便经历了60%的减雨处理,仍有充足水分维持植物生长,从而抵消了前期干旱的负效应.在物种密度方面,生长季前期的持续干旱(-60d)也表现出不同的响应趋势:2021年该处理下物种密度升高,而2022年则降低.主要原因在于2021年7月的大量降雨促进了一年生和二年生草本植物的快速萌发与生长,这类植物具有较高的资源利用效率,能够在短时间内完成生命周期[31],从而显著提升了植被密度;而2022年缺乏类似的降雨补偿,导致密度下降.在物种丰富度方面,2021年的-60%处理显著降低了物种丰富度,而2022年则未观察到显著变化.这一差异可能是由于两年生长季前期累积降雨量差异所致:2021年5—6月降雨总量为35.76 mm,而2022年为75.36 mm,前者更为干旱,对物种更新和建群过程影响更强.地上生物量的变化也呈现类似趋势:2021年生长季前期干旱显著抑制了地上生物量积累,而2022年降雨变化对其影响不显著.这与已有研究中持续干旱显著降低地上生物量的结论一致[32],表明降雨间隔及其时间分布对生产力的调控至关重要.相比之下,两年间降雨变化对地下生物量、Shannon-Wiener多样性指数、Pielou均匀度指数和Simpson优势度指数均未产生显著影响,表明在半干旱沙质草地生态系统中,植被盖度、丰富度、物种密度和地上生物量等指标对降雨变化更为敏感,而地下生物量及多样性指数相对稳定,响应滞后或调节能力更强. ...
科尔沁沙地中南部34种植物叶功能性状及其相互关系
1
2015
... 2021年和2022年,降雨变化对植被群落特征的影响表现出显著的年度差异.与已有研究中生长季降雨减少会降低植被盖度的结论一致[30],本研究发现,生长季前期(5—6月)持续干旱处理(-60d)在两年中均显著降低了植被盖度.然而,对于整个生长季减雨处理(-60%),其影响在两年间存在显著差异:2021年该处理对盖度无显著影响,而2022年则显著降低了盖度.该差异可能归因于两年降雨的时空分布不同.2021年7月降雨量高达159 mm,即便经历了60%的减雨处理,仍有充足水分维持植物生长,从而抵消了前期干旱的负效应.在物种密度方面,生长季前期的持续干旱(-60d)也表现出不同的响应趋势:2021年该处理下物种密度升高,而2022年则降低.主要原因在于2021年7月的大量降雨促进了一年生和二年生草本植物的快速萌发与生长,这类植物具有较高的资源利用效率,能够在短时间内完成生命周期[31],从而显著提升了植被密度;而2022年缺乏类似的降雨补偿,导致密度下降.在物种丰富度方面,2021年的-60%处理显著降低了物种丰富度,而2022年则未观察到显著变化.这一差异可能是由于两年生长季前期累积降雨量差异所致:2021年5—6月降雨总量为35.76 mm,而2022年为75.36 mm,前者更为干旱,对物种更新和建群过程影响更强.地上生物量的变化也呈现类似趋势:2021年生长季前期干旱显著抑制了地上生物量积累,而2022年降雨变化对其影响不显著.这与已有研究中持续干旱显著降低地上生物量的结论一致[32],表明降雨间隔及其时间分布对生产力的调控至关重要.相比之下,两年间降雨变化对地下生物量、Shannon-Wiener多样性指数、Pielou均匀度指数和Simpson优势度指数均未产生显著影响,表明在半干旱沙质草地生态系统中,植被盖度、丰富度、物种密度和地上生物量等指标对降雨变化更为敏感,而地下生物量及多样性指数相对稳定,响应滞后或调节能力更强. ...
科尔沁沙质草地生物量积累过程对降水变化的响应模拟
1
2020
... 2021年和2022年,降雨变化对植被群落特征的影响表现出显著的年度差异.与已有研究中生长季降雨减少会降低植被盖度的结论一致[30],本研究发现,生长季前期(5—6月)持续干旱处理(-60d)在两年中均显著降低了植被盖度.然而,对于整个生长季减雨处理(-60%),其影响在两年间存在显著差异:2021年该处理对盖度无显著影响,而2022年则显著降低了盖度.该差异可能归因于两年降雨的时空分布不同.2021年7月降雨量高达159 mm,即便经历了60%的减雨处理,仍有充足水分维持植物生长,从而抵消了前期干旱的负效应.在物种密度方面,生长季前期的持续干旱(-60d)也表现出不同的响应趋势:2021年该处理下物种密度升高,而2022年则降低.主要原因在于2021年7月的大量降雨促进了一年生和二年生草本植物的快速萌发与生长,这类植物具有较高的资源利用效率,能够在短时间内完成生命周期[31],从而显著提升了植被密度;而2022年缺乏类似的降雨补偿,导致密度下降.在物种丰富度方面,2021年的-60%处理显著降低了物种丰富度,而2022年则未观察到显著变化.这一差异可能是由于两年生长季前期累积降雨量差异所致:2021年5—6月降雨总量为35.76 mm,而2022年为75.36 mm,前者更为干旱,对物种更新和建群过程影响更强.地上生物量的变化也呈现类似趋势:2021年生长季前期干旱显著抑制了地上生物量积累,而2022年降雨变化对其影响不显著.这与已有研究中持续干旱显著降低地上生物量的结论一致[32],表明降雨间隔及其时间分布对生产力的调控至关重要.相比之下,两年间降雨变化对地下生物量、Shannon-Wiener多样性指数、Pielou均匀度指数和Simpson优势度指数均未产生显著影响,表明在半干旱沙质草地生态系统中,植被盖度、丰富度、物种密度和地上生物量等指标对降雨变化更为敏感,而地下生物量及多样性指数相对稳定,响应滞后或调节能力更强. ...
降水变化对典型草原土壤特征及植物群落结构的影响
1
2023
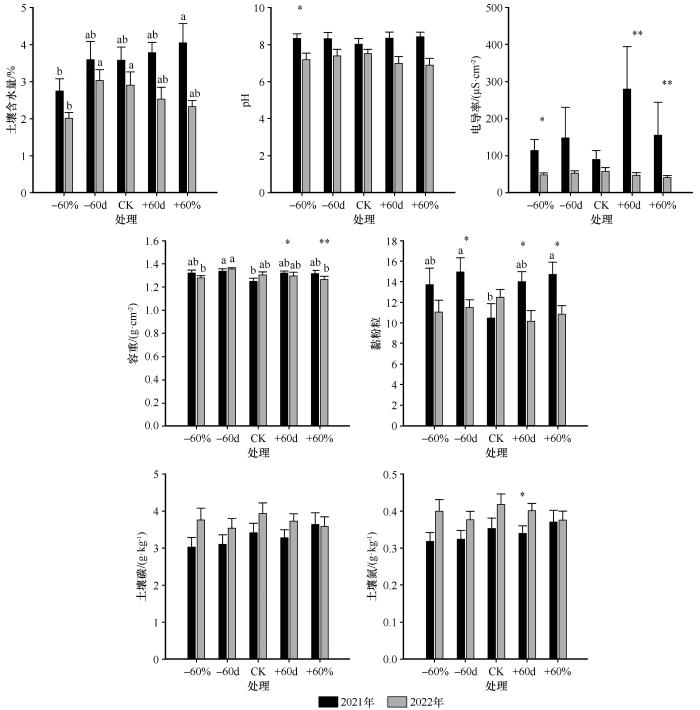
... 2021年和2022年,降雨变化对土壤理化性质的影响也呈现出显著的年度差异.2021年,降雨处理对土壤含水量无显著影响,而2022年,-60%处理显著降低了土壤含水量.这一差异的主要原因在于土壤采样均于8月初进行,而2021年7月降水量较大,有效补充了前期水分亏缺,从而缓解了减雨处理对土壤水分的影响.关于土壤容重的变化,已有典型草原研究表明,降雨变化对土壤容重影响不显著[33];而在荒漠草原中,降雨减少常伴随土壤容重的上升[34].本研究结果与后者一致.2021年生长季前期的极端干旱条件导致植被生长受限、地表覆盖度下降,随之减少了土壤中有机质和根系的物理支撑作用,进而引起土壤结构压实,导致容重升高.同时,干旱还促使土壤中水分迁移和风蚀风险加剧,可能导致部分粗颗粒流失,相对增加了细颗粒(如黏粒和粉粒)的比例,从而表现为黏粉粒含量上升. ...
降水量对荒漠草原土壤呼吸的影响机制研究
1
2022
... 2021年和2022年,降雨变化对土壤理化性质的影响也呈现出显著的年度差异.2021年,降雨处理对土壤含水量无显著影响,而2022年,-60%处理显著降低了土壤含水量.这一差异的主要原因在于土壤采样均于8月初进行,而2021年7月降水量较大,有效补充了前期水分亏缺,从而缓解了减雨处理对土壤水分的影响.关于土壤容重的变化,已有典型草原研究表明,降雨变化对土壤容重影响不显著[33];而在荒漠草原中,降雨减少常伴随土壤容重的上升[34].本研究结果与后者一致.2021年生长季前期的极端干旱条件导致植被生长受限、地表覆盖度下降,随之减少了土壤中有机质和根系的物理支撑作用,进而引起土壤结构压实,导致容重升高.同时,干旱还促使土壤中水分迁移和风蚀风险加剧,可能导致部分粗颗粒流失,相对增加了细颗粒(如黏粒和粉粒)的比例,从而表现为黏粉粒含量上升. ...
沙质草地植物群落特征和土壤质量对氮磷养分添加的响应
1
2021
... 本研究发现,氮添加显著提高了植被盖度,这一结果与以往研究中氮添加促进植物群落盖度的结论一致[35].大量研究表明,氮素输入通常能够促进地上部分植被的生长,增加生物量[36].然而,氮添加也可能对群落结构和生态系统稳定性产生负面影响[20].例如,部分研究指出,氮添加会导致稀有物种逐渐消失,从而引发物种多样性下降和生态系统稳定性减弱[37];也有研究认为,养分输入的增加并不必然导致多样性的变化,具体结果受到区域水热条件、初始土壤养分状况和植物氮利用效率等因素的影响[38].本研究中,氮添加在显著提高群落总生物量的同时,也导致了物种多样性的下降.其主要机制在于氮素输入不仅促进了地上部分植物的生长,提高了整体生物量,还有助于根系的发育,从而增加了地下生物量[28].然而,氮添加也改变了植物间的竞争格局,增强了优势种的竞争能力,导致稀有物种逐渐被排挤出群落,最终引发群落多样性的下降[39-40].此外,氮添加降低了Shannon-Wiener多样性指数,提升了Simpson优势度指数,并降低了Pielou均匀度指数,反映出群落结构趋于单一化[41].其主要原因在于氮素输入改变了资源分配格局,强化了群落内的资源竞争.生长迅速、竞争能力强的优势种(如高大禾本科植物)对氮素具有更高的响应性,在氮添加条件下能够迅速扩张,占据更多的空间与资源,从而抑制其他物种的生存与生长,导致群落中物种数量减少、结构不均.随着优势种生物量的大幅增加,其高大的植株逐渐形成上层冠层,遮蔽了下层光照,限制了矮小或生长缓慢物种的光合作用和空间获取能力.这种垂直层级上的“光竞争”进一步抑制了弱势物种的生长,降低了群落物种在资源利用和空间分布上的均衡性,最终表现为Pielou均匀度指数的显著下降[42]. ...
干旱半干旱区草地植物-土壤响应降水和管理措施的研究综述
2
2025
... 本研究发现,氮添加显著提高了植被盖度,这一结果与以往研究中氮添加促进植物群落盖度的结论一致[35].大量研究表明,氮素输入通常能够促进地上部分植被的生长,增加生物量[36].然而,氮添加也可能对群落结构和生态系统稳定性产生负面影响[20].例如,部分研究指出,氮添加会导致稀有物种逐渐消失,从而引发物种多样性下降和生态系统稳定性减弱[37];也有研究认为,养分输入的增加并不必然导致多样性的变化,具体结果受到区域水热条件、初始土壤养分状况和植物氮利用效率等因素的影响[38].本研究中,氮添加在显著提高群落总生物量的同时,也导致了物种多样性的下降.其主要机制在于氮素输入不仅促进了地上部分植物的生长,提高了整体生物量,还有助于根系的发育,从而增加了地下生物量[28].然而,氮添加也改变了植物间的竞争格局,增强了优势种的竞争能力,导致稀有物种逐渐被排挤出群落,最终引发群落多样性的下降[39-40].此外,氮添加降低了Shannon-Wiener多样性指数,提升了Simpson优势度指数,并降低了Pielou均匀度指数,反映出群落结构趋于单一化[41].其主要原因在于氮素输入改变了资源分配格局,强化了群落内的资源竞争.生长迅速、竞争能力强的优势种(如高大禾本科植物)对氮素具有更高的响应性,在氮添加条件下能够迅速扩张,占据更多的空间与资源,从而抑制其他物种的生存与生长,导致群落中物种数量减少、结构不均.随着优势种生物量的大幅增加,其高大的植株逐渐形成上层冠层,遮蔽了下层光照,限制了矮小或生长缓慢物种的光合作用和空间获取能力.这种垂直层级上的“光竞争”进一步抑制了弱势物种的生长,降低了群落物种在资源利用和空间分布上的均衡性,最终表现为Pielou均匀度指数的显著下降[42]. ...
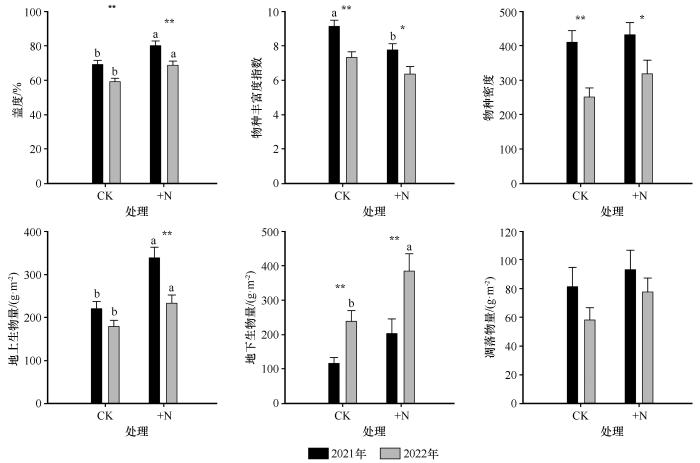
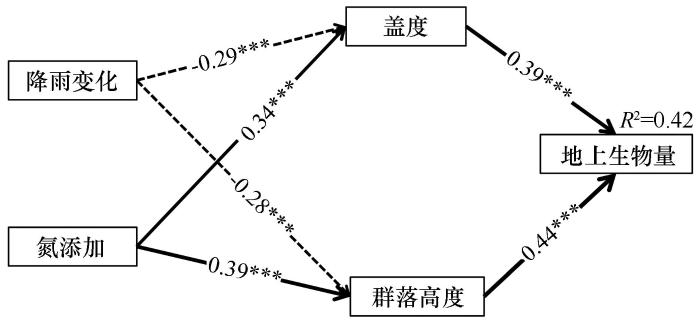
... 本研究发现,降雨处理在研究区内对植被盖度和群落高度均表现出显著的负效应,而氮添加则对上述两个群落结构指标均具有显著的正效应.同时,盖度和群落高度与地上生物量均呈显著正相关关系,表明群落结构的变化对生产力具有直接影响.然而,这一结果与以往部分研究的结论存在不一致.例如,已有研究普遍认为降雨增加有助于提升盖度和群落高度,从而促进地上生物量积累[46-47].本研究中的差异性结果提示,在特定生态背景下,降雨变化对群落结构和功能的影响可能更加复杂,受制于区域性物种组成与资源限制格局.进一步分析表明,降雨处理导致生物量降低的主要机制可能与群落内物种更替密切相关[48-49].干旱胁迫引发了群落结构的重组,促进了对水分敏感的劣势草种的消退,而耐旱性强、生物个体较大的黄蒿和蒺藜等植物则在资源竞争中占据上风.该类植物虽然个体生物量较大,但整体种类较少,群落异质性降低,生物多样性下降,从而削弱了群落对环境波动的响应能力.相比之下,本研究中氮添加显著提高了盖度和群落高度,并间接推动了生物量的增加.作为限制性营养元素,氮素增加能够改善植物养分状况,促进地上部分的生长,并可能缓解部分干旱带来的资源压力,尤其是对于在养分获取上具较强优势的草本植物更为明显[36,50].综合来看,研究结果提示,在半干旱区生态系统中,降雨变化所引发的植物功能群更替和竞争格局变化,是驱动群落结构和生产力变化的重要过程,需在未来的生态响应与恢复策略中予以充分考虑. ...
Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning:evidence from Inner Mongolia Grasslands
1
2010
... 本研究发现,氮添加显著提高了植被盖度,这一结果与以往研究中氮添加促进植物群落盖度的结论一致[35].大量研究表明,氮素输入通常能够促进地上部分植被的生长,增加生物量[36].然而,氮添加也可能对群落结构和生态系统稳定性产生负面影响[20].例如,部分研究指出,氮添加会导致稀有物种逐渐消失,从而引发物种多样性下降和生态系统稳定性减弱[37];也有研究认为,养分输入的增加并不必然导致多样性的变化,具体结果受到区域水热条件、初始土壤养分状况和植物氮利用效率等因素的影响[38].本研究中,氮添加在显著提高群落总生物量的同时,也导致了物种多样性的下降.其主要机制在于氮素输入不仅促进了地上部分植物的生长,提高了整体生物量,还有助于根系的发育,从而增加了地下生物量[28].然而,氮添加也改变了植物间的竞争格局,增强了优势种的竞争能力,导致稀有物种逐渐被排挤出群落,最终引发群落多样性的下降[39-40].此外,氮添加降低了Shannon-Wiener多样性指数,提升了Simpson优势度指数,并降低了Pielou均匀度指数,反映出群落结构趋于单一化[41].其主要原因在于氮素输入改变了资源分配格局,强化了群落内的资源竞争.生长迅速、竞争能力强的优势种(如高大禾本科植物)对氮素具有更高的响应性,在氮添加条件下能够迅速扩张,占据更多的空间与资源,从而抑制其他物种的生存与生长,导致群落中物种数量减少、结构不均.随着优势种生物量的大幅增加,其高大的植株逐渐形成上层冠层,遮蔽了下层光照,限制了矮小或生长缓慢物种的光合作用和空间获取能力.这种垂直层级上的“光竞争”进一步抑制了弱势物种的生长,降低了群落物种在资源利用和空间分布上的均衡性,最终表现为Pielou均匀度指数的显著下降[42]. ...
金沙江干旱河谷退化草地群落对氮磷施肥的反应
1
2004
... 本研究发现,氮添加显著提高了植被盖度,这一结果与以往研究中氮添加促进植物群落盖度的结论一致[35].大量研究表明,氮素输入通常能够促进地上部分植被的生长,增加生物量[36].然而,氮添加也可能对群落结构和生态系统稳定性产生负面影响[20].例如,部分研究指出,氮添加会导致稀有物种逐渐消失,从而引发物种多样性下降和生态系统稳定性减弱[37];也有研究认为,养分输入的增加并不必然导致多样性的变化,具体结果受到区域水热条件、初始土壤养分状况和植物氮利用效率等因素的影响[38].本研究中,氮添加在显著提高群落总生物量的同时,也导致了物种多样性的下降.其主要机制在于氮素输入不仅促进了地上部分植物的生长,提高了整体生物量,还有助于根系的发育,从而增加了地下生物量[28].然而,氮添加也改变了植物间的竞争格局,增强了优势种的竞争能力,导致稀有物种逐渐被排挤出群落,最终引发群落多样性的下降[39-40].此外,氮添加降低了Shannon-Wiener多样性指数,提升了Simpson优势度指数,并降低了Pielou均匀度指数,反映出群落结构趋于单一化[41].其主要原因在于氮素输入改变了资源分配格局,强化了群落内的资源竞争.生长迅速、竞争能力强的优势种(如高大禾本科植物)对氮素具有更高的响应性,在氮添加条件下能够迅速扩张,占据更多的空间与资源,从而抑制其他物种的生存与生长,导致群落中物种数量减少、结构不均.随着优势种生物量的大幅增加,其高大的植株逐渐形成上层冠层,遮蔽了下层光照,限制了矮小或生长缓慢物种的光合作用和空间获取能力.这种垂直层级上的“光竞争”进一步抑制了弱势物种的生长,降低了群落物种在资源利用和空间分布上的均衡性,最终表现为Pielou均匀度指数的显著下降[42]. ...
Increased productivity in wet years drives a decline in ecosystem stability with nitrogen additions in arid grasslands
1
2017
... 本研究发现,氮添加显著提高了植被盖度,这一结果与以往研究中氮添加促进植物群落盖度的结论一致[35].大量研究表明,氮素输入通常能够促进地上部分植被的生长,增加生物量[36].然而,氮添加也可能对群落结构和生态系统稳定性产生负面影响[20].例如,部分研究指出,氮添加会导致稀有物种逐渐消失,从而引发物种多样性下降和生态系统稳定性减弱[37];也有研究认为,养分输入的增加并不必然导致多样性的变化,具体结果受到区域水热条件、初始土壤养分状况和植物氮利用效率等因素的影响[38].本研究中,氮添加在显著提高群落总生物量的同时,也导致了物种多样性的下降.其主要机制在于氮素输入不仅促进了地上部分植物的生长,提高了整体生物量,还有助于根系的发育,从而增加了地下生物量[28].然而,氮添加也改变了植物间的竞争格局,增强了优势种的竞争能力,导致稀有物种逐渐被排挤出群落,最终引发群落多样性的下降[39-40].此外,氮添加降低了Shannon-Wiener多样性指数,提升了Simpson优势度指数,并降低了Pielou均匀度指数,反映出群落结构趋于单一化[41].其主要原因在于氮素输入改变了资源分配格局,强化了群落内的资源竞争.生长迅速、竞争能力强的优势种(如高大禾本科植物)对氮素具有更高的响应性,在氮添加条件下能够迅速扩张,占据更多的空间与资源,从而抑制其他物种的生存与生长,导致群落中物种数量减少、结构不均.随着优势种生物量的大幅增加,其高大的植株逐渐形成上层冠层,遮蔽了下层光照,限制了矮小或生长缓慢物种的光合作用和空间获取能力.这种垂直层级上的“光竞争”进一步抑制了弱势物种的生长,降低了群落物种在资源利用和空间分布上的均衡性,最终表现为Pielou均匀度指数的显著下降[42]. ...
氮添加对内蒙古退化草地植物群落多样性和生物量的影响
1
2018
... 本研究发现,氮添加显著提高了植被盖度,这一结果与以往研究中氮添加促进植物群落盖度的结论一致[35].大量研究表明,氮素输入通常能够促进地上部分植被的生长,增加生物量[36].然而,氮添加也可能对群落结构和生态系统稳定性产生负面影响[20].例如,部分研究指出,氮添加会导致稀有物种逐渐消失,从而引发物种多样性下降和生态系统稳定性减弱[37];也有研究认为,养分输入的增加并不必然导致多样性的变化,具体结果受到区域水热条件、初始土壤养分状况和植物氮利用效率等因素的影响[38].本研究中,氮添加在显著提高群落总生物量的同时,也导致了物种多样性的下降.其主要机制在于氮素输入不仅促进了地上部分植物的生长,提高了整体生物量,还有助于根系的发育,从而增加了地下生物量[28].然而,氮添加也改变了植物间的竞争格局,增强了优势种的竞争能力,导致稀有物种逐渐被排挤出群落,最终引发群落多样性的下降[39-40].此外,氮添加降低了Shannon-Wiener多样性指数,提升了Simpson优势度指数,并降低了Pielou均匀度指数,反映出群落结构趋于单一化[41].其主要原因在于氮素输入改变了资源分配格局,强化了群落内的资源竞争.生长迅速、竞争能力强的优势种(如高大禾本科植物)对氮素具有更高的响应性,在氮添加条件下能够迅速扩张,占据更多的空间与资源,从而抑制其他物种的生存与生长,导致群落中物种数量减少、结构不均.随着优势种生物量的大幅增加,其高大的植株逐渐形成上层冠层,遮蔽了下层光照,限制了矮小或生长缓慢物种的光合作用和空间获取能力.这种垂直层级上的“光竞争”进一步抑制了弱势物种的生长,降低了群落物种在资源利用和空间分布上的均衡性,最终表现为Pielou均匀度指数的显著下降[42]. ...
围封和水氮添加对重度退化草地植物多样性的影响
1
2020
... 本研究发现,氮添加显著提高了植被盖度,这一结果与以往研究中氮添加促进植物群落盖度的结论一致[35].大量研究表明,氮素输入通常能够促进地上部分植被的生长,增加生物量[36].然而,氮添加也可能对群落结构和生态系统稳定性产生负面影响[20].例如,部分研究指出,氮添加会导致稀有物种逐渐消失,从而引发物种多样性下降和生态系统稳定性减弱[37];也有研究认为,养分输入的增加并不必然导致多样性的变化,具体结果受到区域水热条件、初始土壤养分状况和植物氮利用效率等因素的影响[38].本研究中,氮添加在显著提高群落总生物量的同时,也导致了物种多样性的下降.其主要机制在于氮素输入不仅促进了地上部分植物的生长,提高了整体生物量,还有助于根系的发育,从而增加了地下生物量[28].然而,氮添加也改变了植物间的竞争格局,增强了优势种的竞争能力,导致稀有物种逐渐被排挤出群落,最终引发群落多样性的下降[39-40].此外,氮添加降低了Shannon-Wiener多样性指数,提升了Simpson优势度指数,并降低了Pielou均匀度指数,反映出群落结构趋于单一化[41].其主要原因在于氮素输入改变了资源分配格局,强化了群落内的资源竞争.生长迅速、竞争能力强的优势种(如高大禾本科植物)对氮素具有更高的响应性,在氮添加条件下能够迅速扩张,占据更多的空间与资源,从而抑制其他物种的生存与生长,导致群落中物种数量减少、结构不均.随着优势种生物量的大幅增加,其高大的植株逐渐形成上层冠层,遮蔽了下层光照,限制了矮小或生长缓慢物种的光合作用和空间获取能力.这种垂直层级上的“光竞争”进一步抑制了弱势物种的生长,降低了群落物种在资源利用和空间分布上的均衡性,最终表现为Pielou均匀度指数的显著下降[42]. ...
Effects of rainfall manipulation and nitrogen addition on plant biomass allocation in a semiarid sandy grassland
1
2020
... 本研究发现,氮添加显著提高了植被盖度,这一结果与以往研究中氮添加促进植物群落盖度的结论一致[35].大量研究表明,氮素输入通常能够促进地上部分植被的生长,增加生物量[36].然而,氮添加也可能对群落结构和生态系统稳定性产生负面影响[20].例如,部分研究指出,氮添加会导致稀有物种逐渐消失,从而引发物种多样性下降和生态系统稳定性减弱[37];也有研究认为,养分输入的增加并不必然导致多样性的变化,具体结果受到区域水热条件、初始土壤养分状况和植物氮利用效率等因素的影响[38].本研究中,氮添加在显著提高群落总生物量的同时,也导致了物种多样性的下降.其主要机制在于氮素输入不仅促进了地上部分植物的生长,提高了整体生物量,还有助于根系的发育,从而增加了地下生物量[28].然而,氮添加也改变了植物间的竞争格局,增强了优势种的竞争能力,导致稀有物种逐渐被排挤出群落,最终引发群落多样性的下降[39-40].此外,氮添加降低了Shannon-Wiener多样性指数,提升了Simpson优势度指数,并降低了Pielou均匀度指数,反映出群落结构趋于单一化[41].其主要原因在于氮素输入改变了资源分配格局,强化了群落内的资源竞争.生长迅速、竞争能力强的优势种(如高大禾本科植物)对氮素具有更高的响应性,在氮添加条件下能够迅速扩张,占据更多的空间与资源,从而抑制其他物种的生存与生长,导致群落中物种数量减少、结构不均.随着优势种生物量的大幅增加,其高大的植株逐渐形成上层冠层,遮蔽了下层光照,限制了矮小或生长缓慢物种的光合作用和空间获取能力.这种垂直层级上的“光竞争”进一步抑制了弱势物种的生长,降低了群落物种在资源利用和空间分布上的均衡性,最终表现为Pielou均匀度指数的显著下降[42]. ...
长期氮沉降和地上凋落物处理对半干旱区沙质草地表层土壤碳氮组分的影响
1
2023
... 本研究发现,氮添加显著降低了土壤pH值,这一结果与先前关于半干旱区沙质草地中氮添加引发土壤酸化的研究结论一致[43].此外,氮添加还导致了土壤黏粉粒比例的下降,可能的机制在于氮引起的土壤酸化改变了土壤有机质与金属阳离子之间的结合状态,削弱了微团聚体的结构稳定性,进而使部分细粒径颗粒(如黏粒和粉粒)在降水作用下发生径流流失或垂向淋溶,从而降低了其在土壤中的相对含量[44].本研究还发现,氮添加对土壤有机碳和全氮含量未表现出显著影响.可能的原因在于氮添加一方面通过促进植物生长增强了初级生产力,增加了有机碳输入,另一方面又可能加速了微生物分解作用,增强有机质矿化速率.这两种效应在一定程度上相互抵消,导致土壤碳氮含量总体保持动态平衡状态[45]. ...
气候和植被影响土壤理化性质及微生物对氮添加的响应全球控制实验Meta分析
1
2025
... 本研究发现,氮添加显著降低了土壤pH值,这一结果与先前关于半干旱区沙质草地中氮添加引发土壤酸化的研究结论一致[43].此外,氮添加还导致了土壤黏粉粒比例的下降,可能的机制在于氮引起的土壤酸化改变了土壤有机质与金属阳离子之间的结合状态,削弱了微团聚体的结构稳定性,进而使部分细粒径颗粒(如黏粒和粉粒)在降水作用下发生径流流失或垂向淋溶,从而降低了其在土壤中的相对含量[44].本研究还发现,氮添加对土壤有机碳和全氮含量未表现出显著影响.可能的原因在于氮添加一方面通过促进植物生长增强了初级生产力,增加了有机碳输入,另一方面又可能加速了微生物分解作用,增强有机质矿化速率.这两种效应在一定程度上相互抵消,导致土壤碳氮含量总体保持动态平衡状态[45]. ...
温带半干旱草地生态系统碳固定及土壤有机碳库对外源氮输入的响应
1
2010
... 本研究发现,氮添加显著降低了土壤pH值,这一结果与先前关于半干旱区沙质草地中氮添加引发土壤酸化的研究结论一致[43].此外,氮添加还导致了土壤黏粉粒比例的下降,可能的机制在于氮引起的土壤酸化改变了土壤有机质与金属阳离子之间的结合状态,削弱了微团聚体的结构稳定性,进而使部分细粒径颗粒(如黏粒和粉粒)在降水作用下发生径流流失或垂向淋溶,从而降低了其在土壤中的相对含量[44].本研究还发现,氮添加对土壤有机碳和全氮含量未表现出显著影响.可能的原因在于氮添加一方面通过促进植物生长增强了初级生产力,增加了有机碳输入,另一方面又可能加速了微生物分解作用,增强有机质矿化速率.这两种效应在一定程度上相互抵消,导致土壤碳氮含量总体保持动态平衡状态[45]. ...
Effect of precipita tion variability on net primary production and soil respiration in a Chihuahuan Desert grassland
1
2011
... 本研究发现,降雨处理在研究区内对植被盖度和群落高度均表现出显著的负效应,而氮添加则对上述两个群落结构指标均具有显著的正效应.同时,盖度和群落高度与地上生物量均呈显著正相关关系,表明群落结构的变化对生产力具有直接影响.然而,这一结果与以往部分研究的结论存在不一致.例如,已有研究普遍认为降雨增加有助于提升盖度和群落高度,从而促进地上生物量积累[46-47].本研究中的差异性结果提示,在特定生态背景下,降雨变化对群落结构和功能的影响可能更加复杂,受制于区域性物种组成与资源限制格局.进一步分析表明,降雨处理导致生物量降低的主要机制可能与群落内物种更替密切相关[48-49].干旱胁迫引发了群落结构的重组,促进了对水分敏感的劣势草种的消退,而耐旱性强、生物个体较大的黄蒿和蒺藜等植物则在资源竞争中占据上风.该类植物虽然个体生物量较大,但整体种类较少,群落异质性降低,生物多样性下降,从而削弱了群落对环境波动的响应能力.相比之下,本研究中氮添加显著提高了盖度和群落高度,并间接推动了生物量的增加.作为限制性营养元素,氮素增加能够改善植物养分状况,促进地上部分的生长,并可能缓解部分干旱带来的资源压力,尤其是对于在养分获取上具较强优势的草本植物更为明显[36,50].综合来看,研究结果提示,在半干旱区生态系统中,降雨变化所引发的植物功能群更替和竞争格局变化,是驱动群落结构和生产力变化的重要过程,需在未来的生态响应与恢复策略中予以充分考虑. ...
水氮添加对毛乌素沙地柠条固沙区草本层植被和土壤特征的影响
1
2022
... 本研究发现,降雨处理在研究区内对植被盖度和群落高度均表现出显著的负效应,而氮添加则对上述两个群落结构指标均具有显著的正效应.同时,盖度和群落高度与地上生物量均呈显著正相关关系,表明群落结构的变化对生产力具有直接影响.然而,这一结果与以往部分研究的结论存在不一致.例如,已有研究普遍认为降雨增加有助于提升盖度和群落高度,从而促进地上生物量积累[46-47].本研究中的差异性结果提示,在特定生态背景下,降雨变化对群落结构和功能的影响可能更加复杂,受制于区域性物种组成与资源限制格局.进一步分析表明,降雨处理导致生物量降低的主要机制可能与群落内物种更替密切相关[48-49].干旱胁迫引发了群落结构的重组,促进了对水分敏感的劣势草种的消退,而耐旱性强、生物个体较大的黄蒿和蒺藜等植物则在资源竞争中占据上风.该类植物虽然个体生物量较大,但整体种类较少,群落异质性降低,生物多样性下降,从而削弱了群落对环境波动的响应能力.相比之下,本研究中氮添加显著提高了盖度和群落高度,并间接推动了生物量的增加.作为限制性营养元素,氮素增加能够改善植物养分状况,促进地上部分的生长,并可能缓解部分干旱带来的资源压力,尤其是对于在养分获取上具较强优势的草本植物更为明显[36,50].综合来看,研究结果提示,在半干旱区生态系统中,降雨变化所引发的植物功能群更替和竞争格局变化,是驱动群落结构和生产力变化的重要过程,需在未来的生态响应与恢复策略中予以充分考虑. ...
Microbial diversity drives multifunctionality in terrestrial ecosystems
1
2016
... 本研究发现,降雨处理在研究区内对植被盖度和群落高度均表现出显著的负效应,而氮添加则对上述两个群落结构指标均具有显著的正效应.同时,盖度和群落高度与地上生物量均呈显著正相关关系,表明群落结构的变化对生产力具有直接影响.然而,这一结果与以往部分研究的结论存在不一致.例如,已有研究普遍认为降雨增加有助于提升盖度和群落高度,从而促进地上生物量积累[46-47].本研究中的差异性结果提示,在特定生态背景下,降雨变化对群落结构和功能的影响可能更加复杂,受制于区域性物种组成与资源限制格局.进一步分析表明,降雨处理导致生物量降低的主要机制可能与群落内物种更替密切相关[48-49].干旱胁迫引发了群落结构的重组,促进了对水分敏感的劣势草种的消退,而耐旱性强、生物个体较大的黄蒿和蒺藜等植物则在资源竞争中占据上风.该类植物虽然个体生物量较大,但整体种类较少,群落异质性降低,生物多样性下降,从而削弱了群落对环境波动的响应能力.相比之下,本研究中氮添加显著提高了盖度和群落高度,并间接推动了生物量的增加.作为限制性营养元素,氮素增加能够改善植物养分状况,促进地上部分的生长,并可能缓解部分干旱带来的资源压力,尤其是对于在养分获取上具较强优势的草本植物更为明显[36,50].综合来看,研究结果提示,在半干旱区生态系统中,降雨变化所引发的植物功能群更替和竞争格局变化,是驱动群落结构和生产力变化的重要过程,需在未来的生态响应与恢复策略中予以充分考虑. ...
Soil microbial network complexity predicts ecosystem function along elevation gradients on the Tibetan Plateau
1
2022
... 本研究发现,降雨处理在研究区内对植被盖度和群落高度均表现出显著的负效应,而氮添加则对上述两个群落结构指标均具有显著的正效应.同时,盖度和群落高度与地上生物量均呈显著正相关关系,表明群落结构的变化对生产力具有直接影响.然而,这一结果与以往部分研究的结论存在不一致.例如,已有研究普遍认为降雨增加有助于提升盖度和群落高度,从而促进地上生物量积累[46-47].本研究中的差异性结果提示,在特定生态背景下,降雨变化对群落结构和功能的影响可能更加复杂,受制于区域性物种组成与资源限制格局.进一步分析表明,降雨处理导致生物量降低的主要机制可能与群落内物种更替密切相关[48-49].干旱胁迫引发了群落结构的重组,促进了对水分敏感的劣势草种的消退,而耐旱性强、生物个体较大的黄蒿和蒺藜等植物则在资源竞争中占据上风.该类植物虽然个体生物量较大,但整体种类较少,群落异质性降低,生物多样性下降,从而削弱了群落对环境波动的响应能力.相比之下,本研究中氮添加显著提高了盖度和群落高度,并间接推动了生物量的增加.作为限制性营养元素,氮素增加能够改善植物养分状况,促进地上部分的生长,并可能缓解部分干旱带来的资源压力,尤其是对于在养分获取上具较强优势的草本植物更为明显[36,50].综合来看,研究结果提示,在半干旱区生态系统中,降雨变化所引发的植物功能群更替和竞争格局变化,是驱动群落结构和生产力变化的重要过程,需在未来的生态响应与恢复策略中予以充分考虑. ...
Growing-season drought and nitrogen addition interactively impair grassland ecosystem stability by reducing species diversity,asynchrony,and stability
1
2023
... 本研究发现,降雨处理在研究区内对植被盖度和群落高度均表现出显著的负效应,而氮添加则对上述两个群落结构指标均具有显著的正效应.同时,盖度和群落高度与地上生物量均呈显著正相关关系,表明群落结构的变化对生产力具有直接影响.然而,这一结果与以往部分研究的结论存在不一致.例如,已有研究普遍认为降雨增加有助于提升盖度和群落高度,从而促进地上生物量积累[46-47].本研究中的差异性结果提示,在特定生态背景下,降雨变化对群落结构和功能的影响可能更加复杂,受制于区域性物种组成与资源限制格局.进一步分析表明,降雨处理导致生物量降低的主要机制可能与群落内物种更替密切相关[48-49].干旱胁迫引发了群落结构的重组,促进了对水分敏感的劣势草种的消退,而耐旱性强、生物个体较大的黄蒿和蒺藜等植物则在资源竞争中占据上风.该类植物虽然个体生物量较大,但整体种类较少,群落异质性降低,生物多样性下降,从而削弱了群落对环境波动的响应能力.相比之下,本研究中氮添加显著提高了盖度和群落高度,并间接推动了生物量的增加.作为限制性营养元素,氮素增加能够改善植物养分状况,促进地上部分的生长,并可能缓解部分干旱带来的资源压力,尤其是对于在养分获取上具较强优势的草本植物更为明显[36,50].综合来看,研究结果提示,在半干旱区生态系统中,降雨变化所引发的植物功能群更替和竞争格局变化,是驱动群落结构和生产力变化的重要过程,需在未来的生态响应与恢复策略中予以充分考虑. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}