


0 引言
植被修复和重建是沙化土地治理的重要举措,不仅有阻断沙源、涵养水源的作用,还承担改善区域生态环境、提高生物多样性的任务。植被修复措施能够显著提升土壤质量、养分含量及微生物活性[1-3],降低土壤容重,增强团聚体稳定性,通过地表凋落物和地下根系分解以及根系分泌物等提高土壤入渗等[4]。土壤作为陆地生态系统的支撑结构,为植物根系生长提供了机械支撑和养分来源,然而,沙地修复过程中土壤诸多生态效应也深刻影响着植被的发展与演替[5]。浑善达克沙地位于中国北方防沙带-京津风沙源治理工程重点区域,自20世纪80年代末至2000年,该地区土地沙化加速,植被多样性和覆盖度降低、固定沙丘活化等相继发生[6];2000年后,因京津风沙源等生态修复工程的实施,土地沙化趋势得到缓解[7]。多年来,浑善达克沙地实施了一系列生态恢复和植被建设项目,包括人工造林、飞播灌草、退耕还林、扦插造林等生态工程;而樟子松(Pinus sylvestris)和杨树(Populus)固沙林、飞播杨柴(Hedysarum laeve)以及扦插黄柳(Salix gordejevii)活沙障也是该地区常见的、有效的植被恢复措施。
沙地植被重建能够改善人类生活环境和气候条件[8-9]、提高植被多样性[10-11]、改变土壤水热条件[12-13]、提高土壤肥力等[14-15]。目前,针对沙地植被重建改良土壤方面的研究主要集中于理化性质的改善,而增加对土壤微生物及其生态功能的分析能全面反映沙地土壤修复的真实情况。土壤微生物在植被凋落物分解驱动碳循环、生物固氮、种子萌发和植物促生等方面发挥重要作用[16-19]。土壤细菌在土壤微生物中数量最多,分布最广,其群落结构和多样性是影响土壤生态功能的重要指标[20-21]。并且,不同植被类型显著影响土壤养分和细菌群落组成[22]。典型农牧交错带樟子松和落叶松恢复策略较榆树和杨树恢复策略显著提高了土壤细菌群落丰富度和多样性[23];广西喀斯特地区桑葚和核桃造林显著提升了土壤微生物多样性,而封育林措施增加了土壤有机碳、氮和磷含量[24];荒漠绿洲过渡带梭梭、花棒以及梭梭-沙拐枣混交林土壤细菌群落结构差异显著[25]。然而,关于流动沙地不同植被恢复策略下土壤细菌生态功能的研究相对不足。基于此,本研究以浑善达克沙地造林20年的樟子松、杨树固沙林地、飞播杨柴固沙林地、扦插黄柳活沙障固沙林地以及附近自然恢复20年的流动沙地为研究对象,通过16S rRNA基因高通量测序技术、PICRUSt功能预测和土壤养分分析相结合的方法,阐释不同植被恢复策略对土壤养分、细菌群落结构及其潜在生态功能的影响,以期为沙化土地功能修复全面评价以及沙区植被重建提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于内蒙古自治区锡林郭勒盟多伦县浑善达克沙地生态系统定位观测研究站(42°08′ 42″N、116°29′47″E,平均海拔1 277 m,图1)。该区域属于温带大陆性气候,寒冷、干旱少雨、多风沙天气;多年平均气温1.6 ℃,最高气温35.4 ℃,最低气温-39.8 ℃;年平均日照3 400 h;无霜期100 d;多年平均降水量386 mm,多年平均蒸发量1 748 mm;年均风速3.6 m∙s-1,全年风速大于17 m∙s-1的日数67 d;土壤为风沙土。
图1
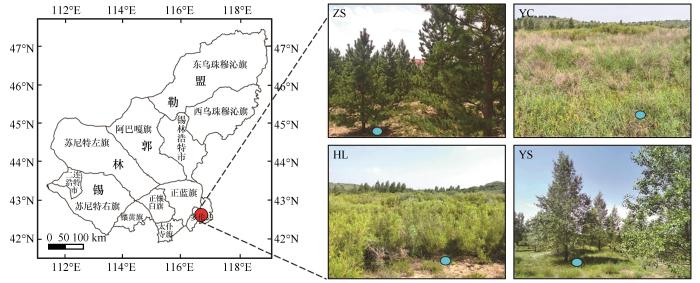
图1
研究区位置及采样点
Fig.1
Study location and sampling sites
研究样地2001年以前为流动沙地,2001—2002年分别营造樟子松(ZS)、杨柴(YC)、黄柳(HL)和杨树(YS)固沙林;其中樟子松林地采用6~7年生苗木栽植,株行距为2 m×4 m,林地面积约1.33 hm2;杨柴林为飞播固沙林地,飞播种子用量约为7.5 kg∙hm-2,林地面积约6.67 hm2;黄柳林为高度50 cm黄柳条扦插的4 m×4 m活沙障,林地面积约2.33 hm2;杨树林采用2年生苗木栽植,株行距为4 m×4 m,林地面积约1.33 hm2。造林后禁牧、自然恢复。恢复20年后,杨柴固沙林杨柴覆盖度约为95%;林下草本植被主要有褐沙蒿(Artemisia halodendron)、灰绿藜(Oxybasis glauca)、羊草(Leymus chinensis)、披碱草(Elymus dahuricus)、叉分蓼(Koenigia divaricata)、菊叶委陵菜(Potentilla tanacetifolia)等。对照样地原为流沙地,自然恢复20年后为固定沙地,植被类型以菊科、藜科和蓼科为主,植被覆盖度约为50%。
1.2 土壤样品采集
于2022年7月11日对樟子松、黄柳、杨柴、杨树固沙林地以及对照样地进行土壤样品采集。采用多点混合法,每个样地在100 m2范围内选择25个距离目标树种30 cm以内的采样点(图1),钻取0~20 cm深度的土壤,并将每5个相近取样点获得的土壤等量均匀混合成1份作为1个生物学重复。5个样地共采集125个采样点,混合成25份,去除植物根系、碎石、枯枝落叶等杂物,4 ℃保存,用于土壤养分检测和土壤细菌DNA的提取和测序分析。
1.3 土壤养分测定
采用碱解扩散法测定土壤速效氮含量,采用钼锑抗比色法测定土壤速效磷含量,采用火焰光度法测定土壤速效钾含量,采用重铬酸钾外加热法测定土壤有机质含量,以上指标测定均参考章家恩[26]的实验方法;参与土壤碳循环的蔗糖酶(Sucrase)和纤维素酶(Cellulase)活性、参与土壤氮循环的中性蛋白酶(Neutral protease)和脲酶(Urease)活性、参与土壤磷循环的中性磷酸酶(Neutral phosphatase)活性以及参与土壤氧化还原反应的过氧化氢酶(Catalase)和脱氢酶(Dehydrogenase)活性均使用北京索莱宝科技有限公司相应土壤酶活性检测试剂盒进行测定。
1.4 土壤细菌测序分析
均匀称取0.5 g土壤样品,采用土壤细菌DNA提取试剂盒(Soil DNA Kit,D5625-01)对不同样地土壤细菌DNA进行提取,3次重复。以引物338F:5′-ACT CCT ACG GGA GGC AGC AG-3′和806R:5′-GGA CTA CHV GGG TWT CTA AT-3′为模板[27],对细菌16S rRNA基因的V3~V4区进行扩增。PCR反应体系为20 μL:5×Fast Pfu Buffer 4 μL,dNTPs(2.5 mmol∙L-1)2 μL,正、反向引物(5 μmol∙L-1)各8 μL,Fast Pfu Polymerase 0.4 μL,DNA模板10 ng。PCR反应条件为:95 ℃ 5 min;95 ℃ 30 s,55 ℃ 30 s,72 ℃ 30 s,35个循环;72 ℃ 10 min。PCR产物以1%琼脂糖凝胶电泳检测,并将检测合格的产物送至上海美吉生物医药科技有限公司,采用Illumina MiSeq测序平台对扩增产物进行测序。序列上传至国际生物技术信息中心(National Center for Biotechnology Information,NCBI)序列读取存档(Sequence Read Archive,SRA)处,获得的登录号为:PRJNA1184990。
按照美吉生信云平台(
1.5 图形绘制和数据分析
采用SPSS(v19.0)软件进行统计学分析;采用R(v3.3.2)软件和Adobe Illustrator 2020进行图形绘制。
2 结果与分析
2.1 不同植被恢复策略下土壤养分差异
不同植被恢复策略土壤养分情况如表1,杨树固沙林土壤速效氮含量、有机质含量、脲酶活性、蔗糖酶活性、纤维素酶活性、过氧化氢酶活性和脱氢酶活性均显著高于对照样地、樟子松、杨柴、黄柳固沙林地,各指标比对照样地分别显著提高58.15%(速效氮)、78.49%(有机质)、52.25%(脲酶活性)、153.65%(蔗糖酶活性)、223.04%(纤维素酶活性)、12.74%(过氧化氢酶活性)和51.61%(脱氢酶活性)。杨柴和杨树固沙林地速效磷和速效钾含量显著高于对照样地;杨树固沙林地和对照样地的中性蛋白酶活性显著高于樟子松、杨柴、黄柳固沙林地,分别提高96.17%~120.22%、184.92%~291.84%和103.98%~128.98%;杨树、杨柴固沙林地和对照样地的中性磷酸酶活性显著高于樟子松和黄柳固沙林地,较樟子松固沙林地分别提高105.19%、59.78%和73.32%,较黄柳固沙林地分别提高109.02%、62.76%和76.56%。结果表明,杨树固沙林地能够更快积累有机质以及氮、磷、钾养分;杨柴固沙林地碳、氮消耗较快,土壤中富集了磷、钾养分;樟子松固沙林地有机质、氮、磷、钾养分消耗较快,黄柳固沙林地养分消耗相对较慢。
表1 不同植被恢复策略土壤养分含量
Table 1
| 土壤指标 | 植被类型 | ||||
|---|---|---|---|---|---|
| CK | ZS | YC | HL | YS | |
| 速效氮/(mg∙kg-1) | 15.46±0.83c | 14.19±0.92c | 17.73±1.17b | 16.83±0.97b | 24.45±1.10a |
| 速效磷/(mg∙kg-1) | 0.90±0.29b | 1.41±0.47ab | 1.74±0.45a | 1.45±0.53ab | 1.61±0.31a |
| 速效钾/(mg∙kg-1) | 10.56±0.45b | 9.60±2.34b | 14.00±1.41a | 11.68±2.19ab | 14.07±1.93a |
| 有机质/(g∙kg-1) | 2.51±0.53bc | 1.66±0.27c | 3.04±0.49b | 2.58±0.51b | 4.48±0.91a |
| 中性蛋白酶/(mg∙d-1∙g-1) | 4.03±0.88a | 1.83±0.44b | 1.26±0.51b | 1.76±0.47b | 3.59±0.70a |
| 脲酶/(μg∙g-1) | 353.17±34.60c | 382.27±17.40bc | 347.09±11.92c | 399.91±43.37b | 537.70±31.77a |
| 蔗糖酶/(mg∙d-1∙g-1) | 3.56±0.13bc | 2.76±0.36cd | 2.13±0.42d | 4.44±0.56b | 9.03±1.71a |
| 纤维素酶/(μg∙d-1∙g-1) | 8.94±2.13d | 13.40±3.75cd | 21.38±4.90b | 17.31±4.98bc | 28.88±3.25a |
| 中性磷酸酶/(mg∙g-1) | 17.02±4.19a | 9.82±1.89b | 15.69±3.29a | 9.64±2.71b | 20.15±4.07a |
| 过氧化氢酶/(μg∙h-1∙g-1) | 1 144.20±23.44b | 1 066.35±35.55b | 883.64±115.60c | 1 116.47±45.52b | 1 289.93±19.18a |
| 脱氢酶/(μg∙d-1∙g-1) | 0.93±0.21b | 0.59±0.13c | 0.67±0.17bc | 0.74±0.15bc | 1.41±0.32a |
2.2 不同植被恢复策略下土壤细菌多样性
不同植被恢复策略下土壤细菌α多样性存在显著差异(P<0.05,图2A),Shannon多样性指数在对照样地(5.04)较樟子松固沙林地(4.89)显著提高;Chao丰富度指数在571~625,在不同植被修复区无显著变化。这说明不同植被恢复策略对土壤细菌群落结构的影响主要表现在多样性上,并且对照样地和杨柴固沙林地的细菌多样性更高,其次是黄柳固沙林地和杨树固沙林地,樟子松固沙林地土壤细菌多样性最低。
图2
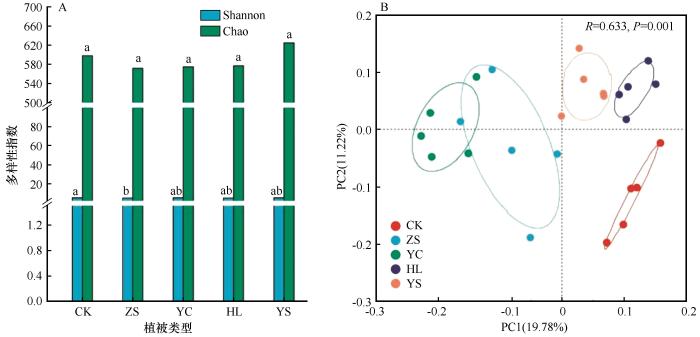
图2
不同植被恢复策略土壤细菌群落α多样性(A)和主坐标分析(B)
Fig.2
α-diversity (A) and Principal coordinate analysis (B) on soil bacterial communities of different vegetation restoration strategies
基于Bray-curtis距离算法对不同植被修复区土壤细菌群落进行主坐标分析(图2B),第一主坐标的解释度为19.78%,第二主坐标的解释度为11.22%,两者共解释31.00%;对照样地、樟子松固沙林、杨柴固沙林、黄柳固沙林和杨树固沙林细菌群落分布相对离散,也进一步证实不同植被恢复策略对土壤细菌群落差异的影响表现在多样性上。基于ANOSIM组间差异检验,证实不同植被恢复策略下土壤细菌群落之间存在显著差异(R=0.633,P=0.001)。
2.3 不同植被恢复策略下土壤细菌群落组成
对不同植被修复区土壤细菌群落进行分析(图3),共获得39门、124纲、292目、465科、845属、1 885种和7119 OTU。平均相对丰度≥1%的门水平物种包括放线菌门(Actinobacteriota)、变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)、拟杆菌门(Bacteroidota)、芽单胞菌门(Gemmatimonadota)、厚壁菌门(Firmicutes)、髌骨菌门(Patescibacteria)、黏球菌门(Myxococcota)和疣微菌门(Verrucomicrobiota),相对丰度分别为34.32%~40.75%、21.16%~29.87%、9.49%~15.62%、7.27%~11.28%、2.72%~4.16%、2.61%~3.34%、1.33%~2.98%、1.07%~3.40%、1.54%~2.20%和0.65%~1.47%。其中,放线菌门、变形菌门和酸杆菌门为优势门;变形菌门、酸杆菌门、绿弯菌门、拟杆菌门、芽单胞菌门和厚壁菌门的相对丰度在不同植被修复区存在显著差异(P<0.05)。杨柴固沙林地变形菌门和拟杆菌门的相对丰度显著高于黄柳固沙林地,分别提高了41.16%和52.94%;而黄柳固沙林地酸杆菌门和绿弯菌门相对丰度显著高于杨柴固沙林地,分别提高了64.59%和55.16%,说明杨柴固沙林地富营养型细菌较为丰富,黄柳固沙林地寡营养型细菌较丰富。对照样地和杨柴固沙林地芽单胞菌门的相对丰度显著高于樟子松固沙林地和杨树固沙林地,并且对照样地、黄柳固沙林地和杨树固沙林地的厚壁菌门相对丰度显著高于樟子松固沙林地,分别提高了116.33%、106.77%和124.06%,芽单胞菌门和厚壁菌门在对照样地明显增多,也说明对照样地出现了更多的抗逆性较强的细菌。
图3
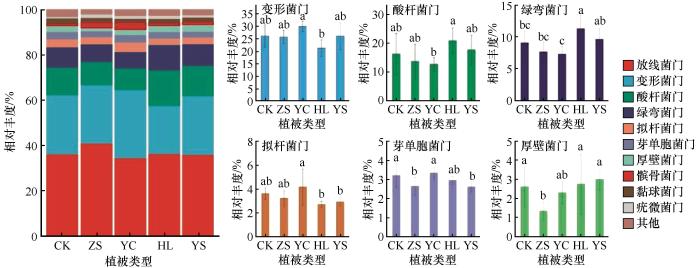
图3
不同植被恢复策略下门水平细菌群落组成
Fig.3
Bacterial community composition of different vegetation restoration strategies at phylum level
如图4所示,属水平相对丰度>0.1%已分类的物种共有35属,包括相对丰度>1%的节杆菌属(Arthrobacter)、慢生根瘤菌属(Bradyrhizobium)、链霉菌属(Streptomyces)、鞘氨醇单胞菌属(Sphingomonas)、类诺卡氏菌属(Nocardioides)、分枝杆菌属(Mycobacterium)、芽孢杆菌属(Bacillus)、假诺卡氏菌属(Pseudonocardia)、红球菌属(Rhodococcus)、Kribbella属,相对丰度>0.5%的土壤红杆菌属(Solirubrobacter)、Gaiella、根瘤菌属(Rhizobium)、中慢生根瘤菌属(Mesorhizobium)、芽单胞菌属(Gemmatimonas)、红色杆菌属(Rubrobacter)、苔藓杆菌属(Bryobacter)、中村氏菌属(Nakamurella)、红游动菌属(Rhodoplanes)、甲基杆菌属(Methylorubrum)、酸杆菌属(Acidibacter)、拟无枝酸菌属(Amycolatopsis)、海球菌属(Marmoricola)、微枝杆菌属(Microvirga),以及相对丰度0.1%~0.5%的德沃斯氏菌属(Devosia)、类芽孢杆菌属(Paenibacillus)等11个属。
图4
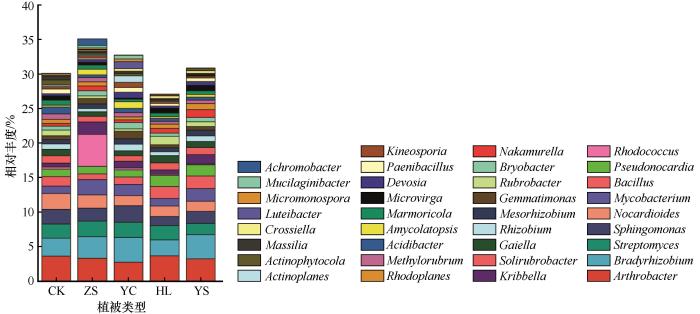
图4
不同植被恢复策略下属水平细菌群落组成
Fig.4
Bacterial community composition of different vegetation restoration strategies at genus level
樟子松固沙林地分枝杆菌属、红球菌属和Kribbella的丰度显著提高(P<0.05);杨柴固沙林地慢生根瘤菌属、鞘氨醇单胞菌属的丰度显著提高(P<0.05);黄柳固沙林地慢生根瘤菌属、鞘氨醇单胞菌属、分枝杆菌属的相对丰度显著降低(P<0.05),芽孢杆菌属和假诺卡氏菌属的相对丰度显著升高;杨树固沙林地慢生根瘤菌属、分枝杆菌属、芽孢杆菌属和假诺卡氏菌属的丰度显著提高(P<0.05)。此外,能够与植物共生固氮的慢生根瘤菌属、根瘤菌属和中慢生根瘤菌属的相对丰度在杨柴固沙林和杨树固沙林显著高于其他样地(P<0.05),慢生根瘤菌在杨柴固沙林和杨树固沙林中的丰度分别为3.59%和3.50%,其他样地为2.30%~3.17%;根瘤菌属在杨柴固沙林和杨树固沙林中的丰度分别为0.97%和0.82%,在其他样地为0.47%~0.78%;中慢生根瘤菌属在杨柴固沙林和杨树固沙林中的丰度分别为0.80%和0.83%,其他样地为0.55%~0.69%:说明杨柴和杨树固沙林地具有固氮功能的细菌类群较多。
2.4 不同植被恢复策略下土壤细菌群落差异
采用Lefse多级物种差异判别分析不同植被恢复策略下土壤细菌群落的差异性(图5),线性判别分析(LDA)得分值越大,说明此类细菌在对照、樟子松、杨柴、黄柳和杨树5个样地中差异越大。不同样地的细菌群落经Lefse分析得到的LDA值大于4.0的分别为樟子松、杨柴和黄柳固沙林地细菌。樟子松固沙林地差异细菌为γ-变形菌纲(Gammaproteobacteria);棒状菌目(Corynebacteriales)、伯克氏菌目(Burkholderiales)和丙酸杆菌目(Propionibacteriales);科诺卡氏菌科(Nocardiaceae)和伯克氏菌科(Burkholderiaceae);红球菌属(Rhodococcus)和Burkholderia-Caballeronia-Paraburkholderia。杨柴固沙林地差异细菌为变形菌门和髌骨菌门;α-变形菌纲(Alphaproteobacteria)和Saccharimonadia;Saccharimonadales目。黄柳固沙林地差异细菌为绿弯菌门;芽球菌纲(Blastocatellia)Vicinamibacterales和拟单胞菌目(Pyrinomonadales);拟单胞菌科(Pyrinomonadaceae);RB41属。结果表明,樟子松、杨柴和黄柳固沙林恢复20年后改变了沙地土壤细菌类群。
图5
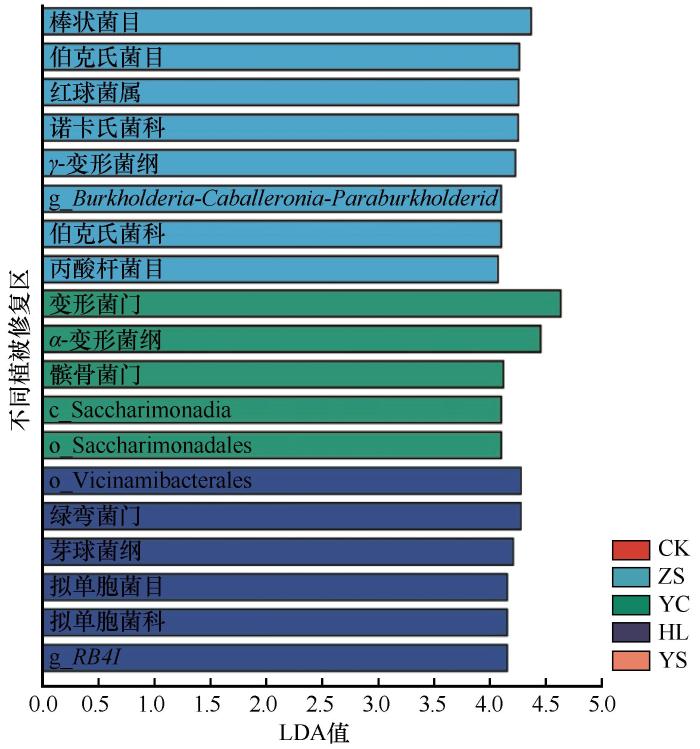
图5
不同植被恢复策略下细菌群落Lefse多级物种差异判别分析
Fig.5
LDA effect size analysis on bacterial community of different vegetation restoration strategies
变形菌门、髌骨菌门和绿弯菌门在门水平对不同植被恢复策略差异贡献度最大,LDA值分别为4.63、4.12和4.27。樟子松固沙林地LDA值大于4.0的差异细菌全部属于变形菌门(γ-变形菌纲、伯克氏菌目、伯克氏菌科、Burkholderia-Caballeronia-Paraburkholderia)和放线菌门(棒状杆菌目、丙酸杆菌目、诺卡氏菌科、红球菌属);杨柴固沙林地LDA值大于4.0的差异细菌全部属于髌骨菌门(髌骨菌门、Saccharimonadia纲和Saccharimonadale目)和变形菌门(变形菌门、α-变形菌纲);黄柳固沙林地LDA值大于4.0的差异细菌全部属于绿弯菌门(绿弯菌门)和酸杆菌门(芽球菌纲、拟单胞菌目、Vicinamibacterales、拟单胞菌科、RB41)。结合各样地细菌群落组成和差异分析,樟子松固沙林地标志细菌为γ-变形菌纲和红球菌属,杨柴固沙林地标志细菌为变形菌门和α-变形菌纲,黄柳固沙林地标志细菌为绿弯菌门和酸杆菌门。
2.5 不同植被恢复策略下土壤细菌群落结构与土壤养分的关系
利用方差膨胀因子分析(VIF默认值为10)筛选保留了共线性较小的环境因子,速效氮(AN)、速效磷(AP)、速效钾(AK)、有机质(OM)、中性蛋白酶(Pr)、脲酶(Ur)、纤维素酶(Ce)、中性磷酸酶(Ph)、过氧化氢酶(Ca)和脱氢酶(De)等10个土壤养分参数用于Mantel test热图分析(图6)。
图6
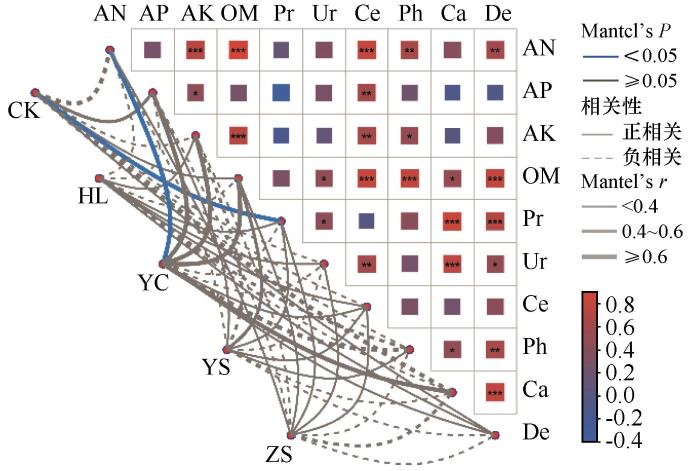
图6
土壤细菌群落与养分的Mantel test热图分析
Fig.6
Mantel test heat map analysis of soil bacterial communities and nutrients
土壤速效氮与多数土壤养分指标呈显著正相关,其次为有机质,说明不同植被恢复策略对土壤氮和有机质的改变是影响土壤肥力的重要因素。对照样地的土壤细菌群落与中性蛋白酶活性显著正相关,杨柴固沙林地细菌群落则与速效氮含量显著正相关,而中性蛋白酶能够水解蛋白质生成氨基酸类有机氮,说明氮是影响对照样地和杨柴固沙林地细菌群落的重要土壤养分;其他样地细菌群落与土壤养分含量相关性不显著。通过Mantel’s r值可以看出,杨柴固沙林地细菌群落与土壤速效氮、速效磷、速效钾、有机质和过氧化氢酶具有较强的相关性,杨树固沙林地细菌与土壤有机质具有较强的相关性,黄柳固沙林地和樟子松固沙林地细菌群落与土壤养分的相关性不显著。结合各样地土壤养分含量,杨柴固沙林地细菌群落对土壤养分的响应更明显,并且与土壤有机质、氮、磷、钾等重要营养元素紧密相关;杨树固沙林地细菌群落则对有机质的响应更明显;黄柳固沙林地和樟子松固沙林地养分含量较低,其细菌群落对养分的响应不明显。
2.6 不同植被恢复策略下土壤细菌群落潜在功能分析
通过PICRUSt2对5种沙地植被恢复策略下土壤细菌生态功能进行预测,结果表明不同植被修复区沙地土壤细菌群落一级功能均具显著性差异(P<0.05)。新陈代谢为主要功能,平均相对丰度为78.67%~79.38%;樟子松、黄柳固沙林地细菌新陈代谢基因相对丰度显著高于杨柴固沙林地,杨柴、杨树固沙林地的环境信息处理基因相对丰度显著高于黄柳固沙林地。考虑到沙地养分贫瘠、高温等恶劣环境,本研究关注新陈代谢和环境信息处理二级功能(图7)。从图7可以看出,杨柴固沙林地信号转导和膜运输作用显著增强,而脂代谢、能量代谢、氨基酸代谢和碳水化合物代谢相对较低;黄柳固沙林地信号转导和膜运输作用显著降低,而能量代谢和碳水化合物代谢作用相对较强;杨树固沙林地膜运输作用显著增强,脂代谢作用较弱;樟子松固沙林地信号转导作用显著降低,脂代谢和氨基酸代谢作用相对较强。综上,养分丰富的杨柴、杨树固沙林地细菌对于环境信息处理能力更强,养分相对贫瘠的樟子松、黄柳固沙林地细菌以碳、氮、脂以及能量代谢为主。
图7
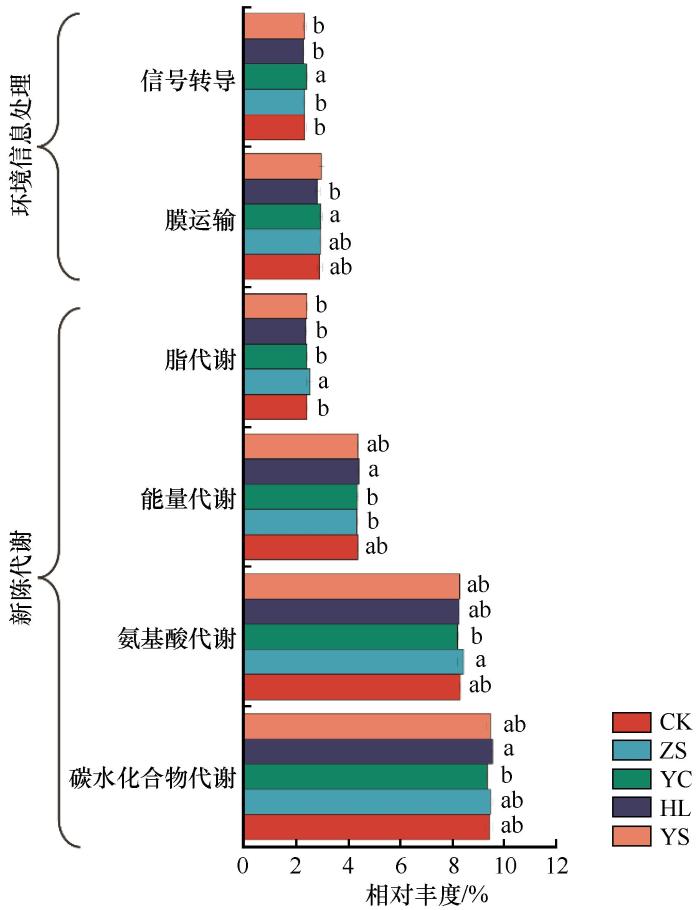
图7
不同植被恢复策略土壤细菌群落潜在功能变化
Fig.7
Potential functional changes of soil bacterial communities in different vegetation restoration strategies
3 讨论
3.1 沙地不同植被恢复策略下土壤细菌群落组成
沙地植被恢复措施不仅能提高土壤微生物多样性和速效氮含量,还能改变细菌群落组成,随着恢复时间的延长,土壤优势细菌由绿弯菌门逐渐转变为变形菌门和放线菌门[28]。本研究中,杨柴固沙林地细菌多样性更高,并且变形菌门和拟杆菌门细菌丰度显著高于黄柳固沙林地;而黄柳固沙林地的酸杆菌门和绿弯菌门相对丰度显著高于杨柴固沙林地。此外,变形菌门也是杨柴固沙林地的标志细菌,绿弯菌门和酸杆菌门也是黄柳固沙林地的标志细菌。酸杆菌门属于典型的寡营养型细菌,而变形菌门属于富营养型细菌,适生于养分含量较高的土壤环境[29];绿弯菌门成员能够通过光合作用固定空气中的CO2转化为有机物质,并供给自身生命活动[30]。结合表1土壤养分含量,可知黄柳固沙林地土壤有机质含量和纤维素酶活性相对杨柴固沙林地较低,使得自养型绿弯菌门细菌丰度在黄柳固沙林地升高,从而适应贫瘠环境。造成这一现象的原因可能是杨柴更新演替较黄柳更快,促进了土壤中的碳回归,提高了土壤有机碳含量;另外,杨柴固沙林地植被覆盖度高达95%,地面覆盖的枯枝落叶有效减少了沙地水分蒸发,间接促进了枯枝落叶的分解,为富营养型细菌的生长提供了有利条件。
通过Lefse多级物种差异判别分析发现,樟子松固沙林地标志细菌为γ-变形菌纲,而杨柴固沙林地标志细菌为α-变形菌纲。Pinhassi等[31]研究发现,γ-变形菌纲成员主要采取“机会主义生存策略”,当遇到易降解的土壤有机碳时具高生长速率,但在低营养条件下,会被α-变形菌纲选择性淘汰,说明α-变形菌纲较γ-变形菌纲成员更具宽泛的营养生长范围,同时也说明杨柴固沙林地的养分供给较樟子松固沙林更稳定。樟子松为针叶树种,其叶片为硬质叶片,不易分解;且叶片氮含量低,进一步抑制了叶片中纤维素和木质素的分解[32];此外,结合表1可知,樟子松固沙林土壤纤维素酶活性较低,经过20年的沙地恢复过程,其土壤养分归还较慢,导致樟子松固沙林地较杨柴固沙林贫瘠,因此出现了大量的机会主义生存者γ-变形菌纲细菌。
本研究中,能够与植物共生固氮的慢生根瘤菌属、根瘤菌属和中慢生根瘤菌属的丰度在杨柴、杨树固沙林地显著高于其他样地,说明杨柴、杨树固沙林地富含固氮菌。生物固氮作为一种无污染、高效提高土壤氮素的方式,在土壤肥力提升、植物生长以及养分循环中具有重要意义。杨柴为豆科植物,其根系与根瘤菌共生形成根瘤帮助杨柴获取更多的氮素,又因为杨柴对氮素的较高需求,导致杨柴固沙林地中富集了多种固氮菌。而杨树为阔叶乔木,其叶片易分解,促进了土壤碳、氮以及多种酶的积累;固氮菌多为富营养型细菌,因此在养分富足的杨树固沙林地富集了较多的固氮菌类群;此外,杨树叶片易分解导致土壤碳含量升高[33],固氮菌便通过增加种群数量达到维持沙地土壤碳氮比平衡的目的。
3.2 沙地不同植被恢复策略下土壤细菌潜在生态功能差异
微生物多样性提升了生态系统功能的稳定性,这种现象在微生物丰度较低的植物-土壤生态系统中尤为明显[34];土壤微生物群落不仅是土壤生态功能的基础,还能调节和指示土壤功能[35]。本研究基于PICRUSt功能预测发现,养分丰富的杨柴、杨树固沙林地细菌环境信息处理能力更强,其二级功能“膜运输”基因丰度显著高于其他样地。Wu等[36]认为膜运输功能增强有利于细菌各项生命活动,是维持细菌发挥正常生理功能的基础。杨树、杨柴固沙林地土壤细菌膜运输功能显著高于黄柳固沙林地,也说明杨树、杨柴固沙林地细菌更能适应环境变化,其活跃的代谢功能有利于维持细胞稳态、抵御逆境[37],细胞内积累的有害代谢产物还可通过细胞膜运输及时排出,利于其自身生长,从而加速细菌群落演替[38]。信号转导是指当外界环境发生变化时,细菌通过改变基因转录表达以减轻环境胁迫对自身造成损伤的生理应答[39]。本研究中杨柴固沙林地“信号转导”基因丰度显著高于樟子松、黄柳、杨树固沙林和对照样地,说明杨柴固沙林地细菌相较其他处理细菌群落更稳定、抵御环境和生物胁迫的能力更强。导致这一现象的原因可能是杨柴固沙林地细菌多样性较高、富营养型细菌占比较大,并且与土壤养分之间的相关性较强,引起了细菌-养分之间的强烈互作,进而增强了细菌膜运输和信号转导基因的表达。
本研究中,杨柴固沙林地细菌群落与土壤速效氮含量呈显著正相关。杜宇佳等[40]在呼伦贝尔沙地土壤细菌群落结构和功能的研究中发现,樟子松固沙林土壤细菌群落丰富度指数与土壤速效氮含量呈显著正相关。Niu等[41]在科尔沁半固定沙地以及沙化草地短期施氮,发现土壤中β-1,4-葡萄糖苷酶活性提高,可能是因为氮添加促进了植物凋落物在土壤中的富集和固定,从而导致产酶相关微生物对氮添加的响应。此外,还有研究表明,沙漠细菌主要参与大分子降解、光合作用、氮素的地球化学循环[42]。杨柴为豆科半灌木,全株富含蛋白质[43],需吸收大量土壤氮素供给其蛋白质合成。沙地土壤贫瘠,而本研究中杨柴固沙林地固氮细菌种类丰富,能够将空气中的氮气固定成铵态氮供杨柴吸收利用;因此,杨柴固沙林地细菌群落与土壤速效氮含量呈显著正相关,也与其自身氮素需求导致根际细菌群落发生改变,进而满足自身高氮需求有关。本研究鉴于PICRUSt功能预测的局限性,未来可结合宏基因、宏代谢、宏蛋白组学相关技术,深入分析沙地不同植被恢复策略土壤细菌生态功能特征,为沙地土壤功能修复以及流动沙地营林建设植物种选择提供科学指导。
4 结论
沙地不同植被恢复策略下土壤细菌群落组成差异显著:杨柴固沙林地富含富营养型细菌;黄柳固沙林地富含寡营养型细菌;樟子松固沙林地“机会主义”生存策略型细菌含量较高;杨树固沙林地和杨柴固沙林地均含有较高比例的固氮细菌。不同植被恢复策略下土壤细菌群落组成差异导致了其潜在生态功能显著变化,杨柴、杨树固沙林地环境信息处理基因显著升高,其细菌群落更稳定,具有较强的抵御环境和生物胁迫的能力;此外,相较樟子松、杨树和黄柳造林,豆科植物杨柴协同土壤固氮细菌对于氮固定和土壤氮积累发挥了重要作用。
参考文献
Impacts of different types of vegetation restoration on the physicochemical properties of sandy soil
[J].
Drivers of soil microbial metabolic limitation changes along a vegetation restoration gradient on the Loess Plateau,China
[J].
Intensive forest harvesting increases susceptibility of northern forest soils to carbon,nitrogen and phosphorus loss
[J].
Effects of different vegetation restoration types on soil hydro-physical properties in the hilly region of the Loess Plateau,China
[J].
NDVI-based vegetation dynamics and their responses to climate change and human activities from 1982 to 2020: a case study in the Mu Us Sandy Land,China
[J].
Impacts of climate change and human activities on vegetation NDVI in China's Mu Us Sandy Land during 2000-2019
[J].
Effects of vegetation restoration on soil properties and vegetation attributes in the arid and semi-arid regions of China
[J].
Environmental changes promoted vegetation growth and reduced water yield over the temperate semi-arid grassland of China during 1901-2016
[J].
Vegetation-cover control of between-site soil temperature evolution in a sandy desertland
[J].
Analyzing vegetation cover-induced organic matter mineralization dynamics in sandy soils from tropical dry coastal ecosystems
[J].
Plant litter crust role in nutrients cycling potentials by bacterial communities in a sandy land ecosystem
[J].
Synergistic effects of plant growth promoting rhizobacteria and mycorrhizal fungi on roselle performance in sandy soils
[J].
Plant growth promoting bacteria promote rice growth cultivated in two different sandy soils subjected distinct climates conditions
[J].
Enhancement of nitrogen cycling and functional microbial flora by artificial inoculation of biological soil crusts in sandy soils of highway slopes
[J].
Evaluating the role of bacterial diversity in supporting soil ecosystem functions under anthropogenic stress
[J].
Soil nutrients,enzyme activities,and bacterial communities in varied plant communities in karst rocky desertification regions in Wushan County,Southwest China
[J].
干旱区不同植被类型土壤细菌群落结构及代谢功能特征
[J/OL].
Dicyandiamide addition delay nitrous oxide emission and shift its production pathway from denitrification to incomplete nitrification in maturation phase of composting
[J].
Effects of different artificial vegetation restoration modes on soil microbial community structure in the soil erosion area of southern China
[J].
Microbial growth and enzyme kinetics in rhizosphere hotspots are modulated by soil organics and nutrient availability
[J].
Non-photosynthetic chemoautotrophic CO2 assimilation microorganisms carbon fixation efficiency and control factors in deep-sea hydrothermal vent
[J].
Differential growth response of colony-forming α-and γ-proteobacteria in dilution culture and nutrient addition experiments from Lake Kinneret (Israel),the eastern Mediterranean Sea,and the Gulf of Eilat
[J].
Decomposition and dynamics of poplar leaves on short rotation conditions
[C].
Diversity and asynchrony in soil microbial communities stabilizes ecosystem functioning
[J].
Assessment of soil microbial diversity measurements as indicators of soil functioning in organic and conventional horticulture systems
[J].
Comparison on the structure and function of the rhizosphere microbial community between healthy and root-rot Panax notoginseng
[J].
New insights into the impact of nZVI on soil microbial biodiversity and functionality
[J].
Interplay between regulatory RNAs and signal transduction systems during bacterial infection
[J].
Soil microbial community responses to short-term nitrogen addition in China's Horqin Sandy Land
[J].
Distribution patterns and functional characteristics of soil bacterial communities in desert ecosystems of northern China
[J].

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}