3
1994
... 荒漠绿洲过渡带是荒漠与绿洲之间的生态过渡区,是荒漠和绿洲生态系统的重要界面及生态缓冲带,是维护绿洲生态安全的重要屏障[1-3].人工固沙植被带是荒漠绿洲过渡带的重要组成部分,不仅是绿洲生态系统的天然保护屏障,也是沙漠化防治的前沿阵地,可有效阻止沙漠向绿洲侵蚀[1,4-7].大量研究表明,人工固沙植被能显著增加土壤表层有机质及氮、磷等养分含量,还会提高动植物多样性,促进土壤恢复,提高人工固沙植物群落的稳定性[8-11].此外,人工植被建植有效固定了流动沙丘,为物种繁殖和生物多样性恢复提供必要的环境条件,同时促进土壤形成及发育,进而影响植被格局演变[9-10].然而,人工固沙植被建植通常加剧土壤水分消耗,导致不同土层水分逐渐匮乏,群落结构趋于疏化,进而逐步进入衰退阶段[12-13].此外,梭梭(Haloxylon ammodendron)、柽柳(Tamarix chinensis)等部分泌盐植物的建植可能导致表层土壤盐分积聚,形成盐结皮,进而抑制降水入渗及草本植物等的种子萌发,阻碍林地自然更新过程,加速土壤功能退化[14-15].因此,开展人工固沙植被建植过程中土壤环境变化及影响要素研究,可为人工固沙植被建植过程中植被结构优化管理、土壤健康恢复及服务功能提升研究提供科学依据. ...
... [1,4-7].大量研究表明,人工固沙植被能显著增加土壤表层有机质及氮、磷等养分含量,还会提高动植物多样性,促进土壤恢复,提高人工固沙植物群落的稳定性[8-11].此外,人工植被建植有效固定了流动沙丘,为物种繁殖和生物多样性恢复提供必要的环境条件,同时促进土壤形成及发育,进而影响植被格局演变[9-10].然而,人工固沙植被建植通常加剧土壤水分消耗,导致不同土层水分逐渐匮乏,群落结构趋于疏化,进而逐步进入衰退阶段[12-13].此外,梭梭(Haloxylon ammodendron)、柽柳(Tamarix chinensis)等部分泌盐植物的建植可能导致表层土壤盐分积聚,形成盐结皮,进而抑制降水入渗及草本植物等的种子萌发,阻碍林地自然更新过程,加速土壤功能退化[14-15].因此,开展人工固沙植被建植过程中土壤环境变化及影响要素研究,可为人工固沙植被建植过程中植被结构优化管理、土壤健康恢复及服务功能提升研究提供科学依据. ...
... 为应对风沙侵蚀,张掖绿洲北部自20世纪70年代逐步开展人工固沙植被建植,形成从荒漠向绿洲过渡的人工梭梭林分布格局[1].基于该区域人工梭梭林随栽植年限形成的重建-演变梯度,选取林龄为5(Y5)、10(Y10)、20(Y20)、30(Y30)年林分作为取样区,并在人工梭梭林斑块边缘选择流动沙丘(MSD)和固定沙丘(FSD)作为对照取样区[35],以探讨不同建植阶段土壤养分特征及其影响因素.在每个取样区内分别选择4个地点,每个地点之间相距500~800 m.在每个地点内再分别选取3个相对独立的梭梭林斑块或沙丘作为植被和土壤样品的采样区,每个采样区间隔至少100 m.2018年秋季即植物多样性最高的季节调查灌木和草本群落组成,采集土壤样品测定土壤理化性质.在每个采样区内设置1个10 m×10 m的灌木调查样方,调查样方内灌木物种组成、密度及盖度,对每株灌木进行株高及冠幅测定;并按对角线设置5个1 m×1 m的草本样方,调查样方内草本盖度和物种丰富度.在每个草本样方内利用环刀法测定表层土壤容重,并利用土钻采集5个0~20 cm的土壤样品混合成1个样品,最终将每个采样区内的5个土壤样品再混合成1个样品.样品带回实验室后风干测定土壤理化性质,其中土壤机械组成[粗砂(>0.25 mm)、细砂(0.05~0.25 mm)和黏粉粒(<0.05 mm)]采用干筛法测定,土壤pH值采用电位法测定,土壤全盐含量采用干烧法测定,全氮采用硫酸-催化剂消解法测定,全磷采用硫酸-高氯酸消解法测定,有机质采用重铬酸钾-浓硫酸外加热法测定. ...
Ecological effects of desertification control and desertified land reclamation in an oasis-desert ecotone in an arid region:a case study in Hexi Corridor,Northwest China
0
2007
The effect of hydrologic process changes on NDVI in the desert-oasis ecotone of the Hexi Corridor
1
2014
... 荒漠绿洲过渡带是荒漠与绿洲之间的生态过渡区,是荒漠和绿洲生态系统的重要界面及生态缓冲带,是维护绿洲生态安全的重要屏障[1-3].人工固沙植被带是荒漠绿洲过渡带的重要组成部分,不仅是绿洲生态系统的天然保护屏障,也是沙漠化防治的前沿阵地,可有效阻止沙漠向绿洲侵蚀[1,4-7].大量研究表明,人工固沙植被能显著增加土壤表层有机质及氮、磷等养分含量,还会提高动植物多样性,促进土壤恢复,提高人工固沙植物群落的稳定性[8-11].此外,人工植被建植有效固定了流动沙丘,为物种繁殖和生物多样性恢复提供必要的环境条件,同时促进土壤形成及发育,进而影响植被格局演变[9-10].然而,人工固沙植被建植通常加剧土壤水分消耗,导致不同土层水分逐渐匮乏,群落结构趋于疏化,进而逐步进入衰退阶段[12-13].此外,梭梭(Haloxylon ammodendron)、柽柳(Tamarix chinensis)等部分泌盐植物的建植可能导致表层土壤盐分积聚,形成盐结皮,进而抑制降水入渗及草本植物等的种子萌发,阻碍林地自然更新过程,加速土壤功能退化[14-15].因此,开展人工固沙植被建植过程中土壤环境变化及影响要素研究,可为人工固沙植被建植过程中植被结构优化管理、土壤健康恢复及服务功能提升研究提供科学依据. ...
Shielding effect of oasis-protection systems composed of various forms of wind break on sand fixation in an arid region: a case study in the Hexi Corridor,Northwest China
1
2008
... 荒漠绿洲过渡带是荒漠与绿洲之间的生态过渡区,是荒漠和绿洲生态系统的重要界面及生态缓冲带,是维护绿洲生态安全的重要屏障[1-3].人工固沙植被带是荒漠绿洲过渡带的重要组成部分,不仅是绿洲生态系统的天然保护屏障,也是沙漠化防治的前沿阵地,可有效阻止沙漠向绿洲侵蚀[1,4-7].大量研究表明,人工固沙植被能显著增加土壤表层有机质及氮、磷等养分含量,还会提高动植物多样性,促进土壤恢复,提高人工固沙植物群落的稳定性[8-11].此外,人工植被建植有效固定了流动沙丘,为物种繁殖和生物多样性恢复提供必要的环境条件,同时促进土壤形成及发育,进而影响植被格局演变[9-10].然而,人工固沙植被建植通常加剧土壤水分消耗,导致不同土层水分逐渐匮乏,群落结构趋于疏化,进而逐步进入衰退阶段[12-13].此外,梭梭(Haloxylon ammodendron)、柽柳(Tamarix chinensis)等部分泌盐植物的建植可能导致表层土壤盐分积聚,形成盐结皮,进而抑制降水入渗及草本植物等的种子萌发,阻碍林地自然更新过程,加速土壤功能退化[14-15].因此,开展人工固沙植被建植过程中土壤环境变化及影响要素研究,可为人工固沙植被建植过程中植被结构优化管理、土壤健康恢复及服务功能提升研究提供科学依据. ...
我国西北干旱区的荒漠化过程及其空间分异规律
0
2011
Combating aeolian desertification in Northern China
0
2012
我国北方风沙危害区生态重建与恢复:腾格里沙漠土壤水分与植被承载力的探讨
1
2014
... 荒漠绿洲过渡带是荒漠与绿洲之间的生态过渡区,是荒漠和绿洲生态系统的重要界面及生态缓冲带,是维护绿洲生态安全的重要屏障[1-3].人工固沙植被带是荒漠绿洲过渡带的重要组成部分,不仅是绿洲生态系统的天然保护屏障,也是沙漠化防治的前沿阵地,可有效阻止沙漠向绿洲侵蚀[1,4-7].大量研究表明,人工固沙植被能显著增加土壤表层有机质及氮、磷等养分含量,还会提高动植物多样性,促进土壤恢复,提高人工固沙植物群落的稳定性[8-11].此外,人工植被建植有效固定了流动沙丘,为物种繁殖和生物多样性恢复提供必要的环境条件,同时促进土壤形成及发育,进而影响植被格局演变[9-10].然而,人工固沙植被建植通常加剧土壤水分消耗,导致不同土层水分逐渐匮乏,群落结构趋于疏化,进而逐步进入衰退阶段[12-13].此外,梭梭(Haloxylon ammodendron)、柽柳(Tamarix chinensis)等部分泌盐植物的建植可能导致表层土壤盐分积聚,形成盐结皮,进而抑制降水入渗及草本植物等的种子萌发,阻碍林地自然更新过程,加速土壤功能退化[14-15].因此,开展人工固沙植被建植过程中土壤环境变化及影响要素研究,可为人工固沙植被建植过程中植被结构优化管理、土壤健康恢复及服务功能提升研究提供科学依据. ...
et al.Long‐term ecosystem effects of sand‐binding vegetation in the Tengger Desert,northern China
1
2004
... 荒漠绿洲过渡带是荒漠与绿洲之间的生态过渡区,是荒漠和绿洲生态系统的重要界面及生态缓冲带,是维护绿洲生态安全的重要屏障[1-3].人工固沙植被带是荒漠绿洲过渡带的重要组成部分,不仅是绿洲生态系统的天然保护屏障,也是沙漠化防治的前沿阵地,可有效阻止沙漠向绿洲侵蚀[1,4-7].大量研究表明,人工固沙植被能显著增加土壤表层有机质及氮、磷等养分含量,还会提高动植物多样性,促进土壤恢复,提高人工固沙植物群落的稳定性[8-11].此外,人工植被建植有效固定了流动沙丘,为物种繁殖和生物多样性恢复提供必要的环境条件,同时促进土壤形成及发育,进而影响植被格局演变[9-10].然而,人工固沙植被建植通常加剧土壤水分消耗,导致不同土层水分逐渐匮乏,群落结构趋于疏化,进而逐步进入衰退阶段[12-13].此外,梭梭(Haloxylon ammodendron)、柽柳(Tamarix chinensis)等部分泌盐植物的建植可能导致表层土壤盐分积聚,形成盐结皮,进而抑制降水入渗及草本植物等的种子萌发,阻碍林地自然更新过程,加速土壤功能退化[14-15].因此,开展人工固沙植被建植过程中土壤环境变化及影响要素研究,可为人工固沙植被建植过程中植被结构优化管理、土壤健康恢复及服务功能提升研究提供科学依据. ...
Long-term effects of xerophytic shrub Haloxylon ammodendron plantations on soil properties and vegetation dynamics in Northwest China
1
2016
... 荒漠绿洲过渡带是荒漠与绿洲之间的生态过渡区,是荒漠和绿洲生态系统的重要界面及生态缓冲带,是维护绿洲生态安全的重要屏障[1-3].人工固沙植被带是荒漠绿洲过渡带的重要组成部分,不仅是绿洲生态系统的天然保护屏障,也是沙漠化防治的前沿阵地,可有效阻止沙漠向绿洲侵蚀[1,4-7].大量研究表明,人工固沙植被能显著增加土壤表层有机质及氮、磷等养分含量,还会提高动植物多样性,促进土壤恢复,提高人工固沙植物群落的稳定性[8-11].此外,人工植被建植有效固定了流动沙丘,为物种繁殖和生物多样性恢复提供必要的环境条件,同时促进土壤形成及发育,进而影响植被格局演变[9-10].然而,人工固沙植被建植通常加剧土壤水分消耗,导致不同土层水分逐渐匮乏,群落结构趋于疏化,进而逐步进入衰退阶段[12-13].此外,梭梭(Haloxylon ammodendron)、柽柳(Tamarix chinensis)等部分泌盐植物的建植可能导致表层土壤盐分积聚,形成盐结皮,进而抑制降水入渗及草本植物等的种子萌发,阻碍林地自然更新过程,加速土壤功能退化[14-15].因此,开展人工固沙植被建植过程中土壤环境变化及影响要素研究,可为人工固沙植被建植过程中植被结构优化管理、土壤健康恢复及服务功能提升研究提供科学依据. ...
Soil properties and herbaceous characteristics in an age sequence of Haloxylon ammodendron plantations in an oasis‐desert ecotone of Northwestern China
2
2016
... 荒漠绿洲过渡带是荒漠与绿洲之间的生态过渡区,是荒漠和绿洲生态系统的重要界面及生态缓冲带,是维护绿洲生态安全的重要屏障[1-3].人工固沙植被带是荒漠绿洲过渡带的重要组成部分,不仅是绿洲生态系统的天然保护屏障,也是沙漠化防治的前沿阵地,可有效阻止沙漠向绿洲侵蚀[1,4-7].大量研究表明,人工固沙植被能显著增加土壤表层有机质及氮、磷等养分含量,还会提高动植物多样性,促进土壤恢复,提高人工固沙植物群落的稳定性[8-11].此外,人工植被建植有效固定了流动沙丘,为物种繁殖和生物多样性恢复提供必要的环境条件,同时促进土壤形成及发育,进而影响植被格局演变[9-10].然而,人工固沙植被建植通常加剧土壤水分消耗,导致不同土层水分逐渐匮乏,群落结构趋于疏化,进而逐步进入衰退阶段[12-13].此外,梭梭(Haloxylon ammodendron)、柽柳(Tamarix chinensis)等部分泌盐植物的建植可能导致表层土壤盐分积聚,形成盐结皮,进而抑制降水入渗及草本植物等的种子萌发,阻碍林地自然更新过程,加速土壤功能退化[14-15].因此,开展人工固沙植被建植过程中土壤环境变化及影响要素研究,可为人工固沙植被建植过程中植被结构优化管理、土壤健康恢复及服务功能提升研究提供科学依据. ...
... 天然荒漠转变为人工梭梭林不仅改变了区域动植物多样性,还对土壤理化性质产生影响,这些改变会随着梭梭林建植年限的增加而变化[10,21,35,42].本研究表明,天然流动沙丘与固定沙丘之间土壤碳、氮、磷化学计量特征存在明显差异;随着人工梭梭林的建植,表层土壤化学计量特征亦发生改变.相较于流动沙丘,固定沙丘中灌木和草本植物的盖度及多样性明显提高,进而引发土壤C∶N下降和N∶P升高.上述结果表明,植被盖度提升加速表层土壤中氮素积累,其速率超过有机碳和全磷积累速率.然而,An等[43]在沙化草地研究发现,随着草地土壤沙化程度加剧,土壤C∶P和N∶P均下降,而C∶N变化不显著,这与本文的研究结果略有不同.泡泡刺和沙拐枣等灌木在荒漠生态系统中具有典型的“生态系统工程师”作用.这类灌木不仅富集枯落物并改善土壤微环境,也为其他生物提供了关键生态位即聚集了大型及中小型节肢动物,从而促进凋落物分解及氮素释放[21,44-45].在荒漠生态系统中,拟步甲科和蚁科等杂食性节肢动物是主要的食物网组成部分,其丰富度和多样性在灌木存在的环境中显著增加.这些节肢动物不仅直接参与凋落物的分解,还通过其死亡残体输入,提升了固定沙丘表层土壤中碳、氮、磷等养分含量[35].此外,研究还发现,研究区流动沙丘和固定沙丘土壤N∶P均低于1,显著低于全国土壤平均水平,进一步表明该区荒漠土壤存在明显的氮素限制[29,46].这种氮素限制特征可能与该区域干旱少雨、土壤有机质积累缓慢及微生物活动受限有关,同时也反映出人工固沙植被建植过程中,氮素补给和循环仍是限制生态系统功能提升的重要因素.梭梭是中国西北干旱区广泛分布的乔木型灌木种,不同梭梭种类及其分布区域会影响土壤碳、氮、磷化学计量比值.在流动和固定沙丘向人工梭梭林转变的过程中,表层土壤C∶N和N∶P发生了显著变化,而C∶P变化相对较小.这一结果与宁虎森等[47]在新疆甘家湖地区梭梭林的研究结果相似,但与魏亚娟等[32]在乌兰布和沙漠西南缘荒漠区梭梭林中的研究结果相反.宁虎森等[47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
Long-term impacts of shrub plantations in a desert-oasis ecotone: accumulation of soil nutrients,salinity,and development of herbaceour layer
1
2018
... 荒漠绿洲过渡带是荒漠与绿洲之间的生态过渡区,是荒漠和绿洲生态系统的重要界面及生态缓冲带,是维护绿洲生态安全的重要屏障[1-3].人工固沙植被带是荒漠绿洲过渡带的重要组成部分,不仅是绿洲生态系统的天然保护屏障,也是沙漠化防治的前沿阵地,可有效阻止沙漠向绿洲侵蚀[1,4-7].大量研究表明,人工固沙植被能显著增加土壤表层有机质及氮、磷等养分含量,还会提高动植物多样性,促进土壤恢复,提高人工固沙植物群落的稳定性[8-11].此外,人工植被建植有效固定了流动沙丘,为物种繁殖和生物多样性恢复提供必要的环境条件,同时促进土壤形成及发育,进而影响植被格局演变[9-10].然而,人工固沙植被建植通常加剧土壤水分消耗,导致不同土层水分逐渐匮乏,群落结构趋于疏化,进而逐步进入衰退阶段[12-13].此外,梭梭(Haloxylon ammodendron)、柽柳(Tamarix chinensis)等部分泌盐植物的建植可能导致表层土壤盐分积聚,形成盐结皮,进而抑制降水入渗及草本植物等的种子萌发,阻碍林地自然更新过程,加速土壤功能退化[14-15].因此,开展人工固沙植被建植过程中土壤环境变化及影响要素研究,可为人工固沙植被建植过程中植被结构优化管理、土壤健康恢复及服务功能提升研究提供科学依据. ...
我国沙区人工植被系统生态-水文过程和互馈机理研究评述
1
2013
... 荒漠绿洲过渡带是荒漠与绿洲之间的生态过渡区,是荒漠和绿洲生态系统的重要界面及生态缓冲带,是维护绿洲生态安全的重要屏障[1-3].人工固沙植被带是荒漠绿洲过渡带的重要组成部分,不仅是绿洲生态系统的天然保护屏障,也是沙漠化防治的前沿阵地,可有效阻止沙漠向绿洲侵蚀[1,4-7].大量研究表明,人工固沙植被能显著增加土壤表层有机质及氮、磷等养分含量,还会提高动植物多样性,促进土壤恢复,提高人工固沙植物群落的稳定性[8-11].此外,人工植被建植有效固定了流动沙丘,为物种繁殖和生物多样性恢复提供必要的环境条件,同时促进土壤形成及发育,进而影响植被格局演变[9-10].然而,人工固沙植被建植通常加剧土壤水分消耗,导致不同土层水分逐渐匮乏,群落结构趋于疏化,进而逐步进入衰退阶段[12-13].此外,梭梭(Haloxylon ammodendron)、柽柳(Tamarix chinensis)等部分泌盐植物的建植可能导致表层土壤盐分积聚,形成盐结皮,进而抑制降水入渗及草本植物等的种子萌发,阻碍林地自然更新过程,加速土壤功能退化[14-15].因此,开展人工固沙植被建植过程中土壤环境变化及影响要素研究,可为人工固沙植被建植过程中植被结构优化管理、土壤健康恢复及服务功能提升研究提供科学依据. ...
Effects of stand structure on ecological benefits and tree growth of Haloxylon ammodendron plantations: a meta-analysis
6
2024
... 荒漠绿洲过渡带是荒漠与绿洲之间的生态过渡区,是荒漠和绿洲生态系统的重要界面及生态缓冲带,是维护绿洲生态安全的重要屏障[1-3].人工固沙植被带是荒漠绿洲过渡带的重要组成部分,不仅是绿洲生态系统的天然保护屏障,也是沙漠化防治的前沿阵地,可有效阻止沙漠向绿洲侵蚀[1,4-7].大量研究表明,人工固沙植被能显著增加土壤表层有机质及氮、磷等养分含量,还会提高动植物多样性,促进土壤恢复,提高人工固沙植物群落的稳定性[8-11].此外,人工植被建植有效固定了流动沙丘,为物种繁殖和生物多样性恢复提供必要的环境条件,同时促进土壤形成及发育,进而影响植被格局演变[9-10].然而,人工固沙植被建植通常加剧土壤水分消耗,导致不同土层水分逐渐匮乏,群落结构趋于疏化,进而逐步进入衰退阶段[12-13].此外,梭梭(Haloxylon ammodendron)、柽柳(Tamarix chinensis)等部分泌盐植物的建植可能导致表层土壤盐分积聚,形成盐结皮,进而抑制降水入渗及草本植物等的种子萌发,阻碍林地自然更新过程,加速土壤功能退化[14-15].因此,开展人工固沙植被建植过程中土壤环境变化及影响要素研究,可为人工固沙植被建植过程中植被结构优化管理、土壤健康恢复及服务功能提升研究提供科学依据. ...
... 梭梭作为适应干旱环境的典型植物,因其耐旱、耐盐碱等特性,在荒漠化治理和生态恢复中具有重要的应用价值[16-19].人工梭梭建植通过修复退化生境、降低风蚀程度、提高动物多样性以及促进土壤碳氮等养分固存,进而显著提升多种生态系统服务功能[13,20-21].人工梭梭林在建植初期通常密度较高,随着建植时间延长,林分密度有所下降.生长至20年左右梭梭林开始出现不同程度衰退,其生态系统服务功能也随之减弱[13].然而,也有研究发现,在绿洲边缘地区20~30年梭梭林往往能够形成相对稳定的斑块状结构,并在30年左右出现幼苗密度回升,表现出一定的自然更新潜力[22-24].降水和地下水位变化以及不同立地条件影响人工梭梭林的重建、演变过程,同时也对土壤恢复过程产生重要影响[22,25-27].在人工梭梭林建植过程中,土壤恢复进程相对缓慢,而土壤质量提升对人工林地生态系统长期健康与稳定具有重要意义[28-29].土壤碳、氮、磷是土壤养分的重要组成部分,其比值(C∶N∶P)不仅反映土壤养分相对平衡状态,还能揭示生态系统养分限制特征与功能特性[30].此外,表层土壤碳、氮和磷储量也是衡量土壤功能变化的重要指标.近年来,关于人工梭梭林建植过程中土壤碳、氮、磷含量、比值及储量变化的研究逐渐增多,但不同的降水条件、地下水位及土壤环境背景均会显著影响其响应模式和变化规律[31-33].在人工梭梭林建植过程中,植被、土壤动物等生物要素与土壤结构、盐分等非生物要素之间相互作用及其对表层土壤碳、氮、磷含量和储量变化的耦合影响认识较为有限.鉴于此,本文以张掖绿洲北部荒漠绿洲过渡带的流动沙丘、固定沙丘以及5、10、20、30年梭梭林为研究对象,分析人工梭梭林重建、演变过程中表层土壤有机碳、全氮和全磷比值及土壤有机碳、全氮、无机氮和全磷储量的变化规律,确定生物和非生物因素对表层土壤碳、氮、磷比值及储量变化的影响和相对贡献率,为绿洲边缘人工梭梭林土壤质量演变及生态系统服务功能变化研究提供科学依据. ...
... [13].然而,也有研究发现,在绿洲边缘地区20~30年梭梭林往往能够形成相对稳定的斑块状结构,并在30年左右出现幼苗密度回升,表现出一定的自然更新潜力[22-24].降水和地下水位变化以及不同立地条件影响人工梭梭林的重建、演变过程,同时也对土壤恢复过程产生重要影响[22,25-27].在人工梭梭林建植过程中,土壤恢复进程相对缓慢,而土壤质量提升对人工林地生态系统长期健康与稳定具有重要意义[28-29].土壤碳、氮、磷是土壤养分的重要组成部分,其比值(C∶N∶P)不仅反映土壤养分相对平衡状态,还能揭示生态系统养分限制特征与功能特性[30].此外,表层土壤碳、氮和磷储量也是衡量土壤功能变化的重要指标.近年来,关于人工梭梭林建植过程中土壤碳、氮、磷含量、比值及储量变化的研究逐渐增多,但不同的降水条件、地下水位及土壤环境背景均会显著影响其响应模式和变化规律[31-33].在人工梭梭林建植过程中,植被、土壤动物等生物要素与土壤结构、盐分等非生物要素之间相互作用及其对表层土壤碳、氮、磷含量和储量变化的耦合影响认识较为有限.鉴于此,本文以张掖绿洲北部荒漠绿洲过渡带的流动沙丘、固定沙丘以及5、10、20、30年梭梭林为研究对象,分析人工梭梭林重建、演变过程中表层土壤有机碳、全氮和全磷比值及土壤有机碳、全氮、无机氮和全磷储量的变化规律,确定生物和非生物因素对表层土壤碳、氮、磷比值及储量变化的影响和相对贡献率,为绿洲边缘人工梭梭林土壤质量演变及生态系统服务功能变化研究提供科学依据. ...
... 天然荒漠转变为人工梭梭林不仅改变了区域动植物多样性,还对土壤理化性质产生影响,这些改变会随着梭梭林建植年限的增加而变化[10,21,35,42].本研究表明,天然流动沙丘与固定沙丘之间土壤碳、氮、磷化学计量特征存在明显差异;随着人工梭梭林的建植,表层土壤化学计量特征亦发生改变.相较于流动沙丘,固定沙丘中灌木和草本植物的盖度及多样性明显提高,进而引发土壤C∶N下降和N∶P升高.上述结果表明,植被盖度提升加速表层土壤中氮素积累,其速率超过有机碳和全磷积累速率.然而,An等[43]在沙化草地研究发现,随着草地土壤沙化程度加剧,土壤C∶P和N∶P均下降,而C∶N变化不显著,这与本文的研究结果略有不同.泡泡刺和沙拐枣等灌木在荒漠生态系统中具有典型的“生态系统工程师”作用.这类灌木不仅富集枯落物并改善土壤微环境,也为其他生物提供了关键生态位即聚集了大型及中小型节肢动物,从而促进凋落物分解及氮素释放[21,44-45].在荒漠生态系统中,拟步甲科和蚁科等杂食性节肢动物是主要的食物网组成部分,其丰富度和多样性在灌木存在的环境中显著增加.这些节肢动物不仅直接参与凋落物的分解,还通过其死亡残体输入,提升了固定沙丘表层土壤中碳、氮、磷等养分含量[35].此外,研究还发现,研究区流动沙丘和固定沙丘土壤N∶P均低于1,显著低于全国土壤平均水平,进一步表明该区荒漠土壤存在明显的氮素限制[29,46].这种氮素限制特征可能与该区域干旱少雨、土壤有机质积累缓慢及微生物活动受限有关,同时也反映出人工固沙植被建植过程中,氮素补给和循环仍是限制生态系统功能提升的重要因素.梭梭是中国西北干旱区广泛分布的乔木型灌木种,不同梭梭种类及其分布区域会影响土壤碳、氮、磷化学计量比值.在流动和固定沙丘向人工梭梭林转变的过程中,表层土壤C∶N和N∶P发生了显著变化,而C∶P变化相对较小.这一结果与宁虎森等[47]在新疆甘家湖地区梭梭林的研究结果相似,但与魏亚娟等[32]在乌兰布和沙漠西南缘荒漠区梭梭林中的研究结果相反.宁虎森等[47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
... 天然和人工梭梭林表层土壤碳、氮、磷含量均高于流动沙地.木本植被的存在及盖度增加,有助于提升荒漠土壤的养分含量与储量,而表层土壤碳、氮、磷储量对植被建植较为敏感[51,53-54].本研究发现,固定沙丘土壤有机碳、全氮和全磷储量均高于流动沙丘,这一趋势与腾格里沙漠和乌兰布和沙漠等区域的研究结果相近[32,54-55].然而,流动和固定沙丘表层土壤的无机氮储量变化相对较小,这可能与该区域土壤水分有效性较低,从而抑制土壤生物的活动有关[56].在人工梭梭林恢复的前期阶段(<20年),表层土壤有机碳、全磷和无机氮储量变化不明显,唯有土壤全氮储量显著增加,这与天然荒漠转变为梭梭林后,节肢动物、线虫及微生物多样性提高有关[35,50,57].魏亚娟等[32]在乌兰布和沙漠的研究也发现,在3~11年人工梭梭林中,土壤有机碳储量变化不大,而在16年时则显著提升,这与本研究结果相近.值得注意的是,在人工梭梭林恢复早期,表层土壤全氮和全磷储量随建植年限的增长而增加,但这一趋势与魏亚娟等[32]的研究稍有不同.Ma等[58]在石羊河流域的研究显示,人工梭梭林土壤有机碳储量随梭梭林建植年限增加而提升,但仍显著低于流动沙丘.本研究进一步表明,在人工梭梭林恢复的后期阶段(>20年),表层土壤的有机碳、无机氮和全磷储量随着建植年限延长而显著增加,但全氮储量在30年林龄时出现显著下降.这一变化趋势与Huang等[13]的研究相一致,即人工梭梭林的生态恢复效应在建植约20年时达到峰值,超过20年后则需要人工干预以增强其生态服务功能.在绿洲边缘区域,人工梭梭林表层土壤的有机碳、无机氮和全磷储量随梭梭林龄的延长而增长,而表层土壤全氮储量则先升后降.这可能与恢复后期表层土壤盐分的积累有关,表层土壤盐分积累抑制了土壤生物活动,进而降低了土壤全氮含量[15]. ...
... 在人工梭梭林建植过程中,非生物因子及其与生物因子的交互作用对表层土壤碳、氮、磷储量的影响大于生物因子的作用,且这些因子对土壤C∶N∶P比值的影响机制存在明显差异.本研究发现,土壤粒径组成和水盐条件等非生物因子在调控养分积累方面发挥着关键作用,这一结论与张掖绿洲边缘及其他干旱区的相关研究结果一致[52,54,59].梭梭林重建与演变过程能够有效削弱风沙活动,减少表层土壤流失,从而增强有机碳和氮积累,尤其在0~5 cm表层土壤中表现显著[54].此外,土壤黏粉粒比例增加可能是提升碳氮固存效率的重要因素[52].进一步分析表明,土壤黏粉粒含量、pH和全盐含量是非生物因子中影响人工梭梭林表层土壤有机碳、全氮和全磷储量变化的主要驱动因子.较高的黏粉粒含量有助于土壤结构的稳定性及有机质的高效积累,而pH值和盐分的变化则通过影响微生物活性及养分转化过程,进而调控土壤养分积累.特别是全盐含量的增加,促进了人工梭梭林下土壤盐结皮的形成,这一过程不仅抑制土壤水分入渗,还会限制草本植物及梭梭幼苗的萌发与生长,同时对土壤生物的活动产生负面影响,最终导致土壤全氮储量及其积累速率下降[15,51].安芳娇等[57]研究指出,在人工梭梭林的建植过程中,土壤pH值、有机碳含量和电导率是驱动线虫群落变化的主要环境因子,其中土壤电导率是影响线虫群落变化的关键因素.此外,灌木盖度、草本盖度及大型节肢动物数量是影响人工梭梭林建植过程中土壤碳、氮、磷储量变化的主要生物因子.灌木盖度与土壤黏粉粒之间的相互作用能够抑制风蚀,促进土壤细颗粒的聚集,进而提高表层土壤碳、氮、磷储量.同时,土壤中蚂蚁、蜘蛛及其他中小型土壤动物种类和数量的增加,加速了凋落物的分解过程[35].然而,土壤盐分增加及土壤水分有效性降低可能抑制了螨类和跳虫等主要分解类群的数量和多样性,从而导致人工梭梭林建植后期土壤全氮储量的下降[35,60-61].在人工梭梭林建植约20年时,通过降低灌木密度和盖度,增加挖掘兽类和节肢动物的多样性,以及提升关键捕食性动物种群的数量,可以显著增强人工梭梭林的生态系统服务功能,并提高其对气候变化和人类干扰的适应能力[13,21,48,62].总体而言,植被盖度、土壤质地与生物群落之间的耦合变化是驱动表层土壤养分储量变化的核心机制.因此,为了提升人工梭梭林的生态系统功能并实现可持续管理,有必要加强对建植过程中土壤环境和生物群落协同演变的长期监测,为相关理论研究和实践应用提供更为坚实的数据支持. ...
树干径流对梭梭“肥岛”和“盐岛”效应的作用机制
1
2012
... 荒漠绿洲过渡带是荒漠与绿洲之间的生态过渡区,是荒漠和绿洲生态系统的重要界面及生态缓冲带,是维护绿洲生态安全的重要屏障[1-3].人工固沙植被带是荒漠绿洲过渡带的重要组成部分,不仅是绿洲生态系统的天然保护屏障,也是沙漠化防治的前沿阵地,可有效阻止沙漠向绿洲侵蚀[1,4-7].大量研究表明,人工固沙植被能显著增加土壤表层有机质及氮、磷等养分含量,还会提高动植物多样性,促进土壤恢复,提高人工固沙植物群落的稳定性[8-11].此外,人工植被建植有效固定了流动沙丘,为物种繁殖和生物多样性恢复提供必要的环境条件,同时促进土壤形成及发育,进而影响植被格局演变[9-10].然而,人工固沙植被建植通常加剧土壤水分消耗,导致不同土层水分逐渐匮乏,群落结构趋于疏化,进而逐步进入衰退阶段[12-13].此外,梭梭(Haloxylon ammodendron)、柽柳(Tamarix chinensis)等部分泌盐植物的建植可能导致表层土壤盐分积聚,形成盐结皮,进而抑制降水入渗及草本植物等的种子萌发,阻碍林地自然更新过程,加速土壤功能退化[14-15].因此,开展人工固沙植被建植过程中土壤环境变化及影响要素研究,可为人工固沙植被建植过程中植被结构优化管理、土壤健康恢复及服务功能提升研究提供科学依据. ...
The establishment and development of Haloxylon ammodendron promotes salt accumulation in surface soil of arid sandy land
3
2019
... 荒漠绿洲过渡带是荒漠与绿洲之间的生态过渡区,是荒漠和绿洲生态系统的重要界面及生态缓冲带,是维护绿洲生态安全的重要屏障[1-3].人工固沙植被带是荒漠绿洲过渡带的重要组成部分,不仅是绿洲生态系统的天然保护屏障,也是沙漠化防治的前沿阵地,可有效阻止沙漠向绿洲侵蚀[1,4-7].大量研究表明,人工固沙植被能显著增加土壤表层有机质及氮、磷等养分含量,还会提高动植物多样性,促进土壤恢复,提高人工固沙植物群落的稳定性[8-11].此外,人工植被建植有效固定了流动沙丘,为物种繁殖和生物多样性恢复提供必要的环境条件,同时促进土壤形成及发育,进而影响植被格局演变[9-10].然而,人工固沙植被建植通常加剧土壤水分消耗,导致不同土层水分逐渐匮乏,群落结构趋于疏化,进而逐步进入衰退阶段[12-13].此外,梭梭(Haloxylon ammodendron)、柽柳(Tamarix chinensis)等部分泌盐植物的建植可能导致表层土壤盐分积聚,形成盐结皮,进而抑制降水入渗及草本植物等的种子萌发,阻碍林地自然更新过程,加速土壤功能退化[14-15].因此,开展人工固沙植被建植过程中土壤环境变化及影响要素研究,可为人工固沙植被建植过程中植被结构优化管理、土壤健康恢复及服务功能提升研究提供科学依据. ...
... 天然和人工梭梭林表层土壤碳、氮、磷含量均高于流动沙地.木本植被的存在及盖度增加,有助于提升荒漠土壤的养分含量与储量,而表层土壤碳、氮、磷储量对植被建植较为敏感[51,53-54].本研究发现,固定沙丘土壤有机碳、全氮和全磷储量均高于流动沙丘,这一趋势与腾格里沙漠和乌兰布和沙漠等区域的研究结果相近[32,54-55].然而,流动和固定沙丘表层土壤的无机氮储量变化相对较小,这可能与该区域土壤水分有效性较低,从而抑制土壤生物的活动有关[56].在人工梭梭林恢复的前期阶段(<20年),表层土壤有机碳、全磷和无机氮储量变化不明显,唯有土壤全氮储量显著增加,这与天然荒漠转变为梭梭林后,节肢动物、线虫及微生物多样性提高有关[35,50,57].魏亚娟等[32]在乌兰布和沙漠的研究也发现,在3~11年人工梭梭林中,土壤有机碳储量变化不大,而在16年时则显著提升,这与本研究结果相近.值得注意的是,在人工梭梭林恢复早期,表层土壤全氮和全磷储量随建植年限的增长而增加,但这一趋势与魏亚娟等[32]的研究稍有不同.Ma等[58]在石羊河流域的研究显示,人工梭梭林土壤有机碳储量随梭梭林建植年限增加而提升,但仍显著低于流动沙丘.本研究进一步表明,在人工梭梭林恢复的后期阶段(>20年),表层土壤的有机碳、无机氮和全磷储量随着建植年限延长而显著增加,但全氮储量在30年林龄时出现显著下降.这一变化趋势与Huang等[13]的研究相一致,即人工梭梭林的生态恢复效应在建植约20年时达到峰值,超过20年后则需要人工干预以增强其生态服务功能.在绿洲边缘区域,人工梭梭林表层土壤的有机碳、无机氮和全磷储量随梭梭林龄的延长而增长,而表层土壤全氮储量则先升后降.这可能与恢复后期表层土壤盐分的积累有关,表层土壤盐分积累抑制了土壤生物活动,进而降低了土壤全氮含量[15]. ...
... 在人工梭梭林建植过程中,非生物因子及其与生物因子的交互作用对表层土壤碳、氮、磷储量的影响大于生物因子的作用,且这些因子对土壤C∶N∶P比值的影响机制存在明显差异.本研究发现,土壤粒径组成和水盐条件等非生物因子在调控养分积累方面发挥着关键作用,这一结论与张掖绿洲边缘及其他干旱区的相关研究结果一致[52,54,59].梭梭林重建与演变过程能够有效削弱风沙活动,减少表层土壤流失,从而增强有机碳和氮积累,尤其在0~5 cm表层土壤中表现显著[54].此外,土壤黏粉粒比例增加可能是提升碳氮固存效率的重要因素[52].进一步分析表明,土壤黏粉粒含量、pH和全盐含量是非生物因子中影响人工梭梭林表层土壤有机碳、全氮和全磷储量变化的主要驱动因子.较高的黏粉粒含量有助于土壤结构的稳定性及有机质的高效积累,而pH值和盐分的变化则通过影响微生物活性及养分转化过程,进而调控土壤养分积累.特别是全盐含量的增加,促进了人工梭梭林下土壤盐结皮的形成,这一过程不仅抑制土壤水分入渗,还会限制草本植物及梭梭幼苗的萌发与生长,同时对土壤生物的活动产生负面影响,最终导致土壤全氮储量及其积累速率下降[15,51].安芳娇等[57]研究指出,在人工梭梭林的建植过程中,土壤pH值、有机碳含量和电导率是驱动线虫群落变化的主要环境因子,其中土壤电导率是影响线虫群落变化的关键因素.此外,灌木盖度、草本盖度及大型节肢动物数量是影响人工梭梭林建植过程中土壤碳、氮、磷储量变化的主要生物因子.灌木盖度与土壤黏粉粒之间的相互作用能够抑制风蚀,促进土壤细颗粒的聚集,进而提高表层土壤碳、氮、磷储量.同时,土壤中蚂蚁、蜘蛛及其他中小型土壤动物种类和数量的增加,加速了凋落物的分解过程[35].然而,土壤盐分增加及土壤水分有效性降低可能抑制了螨类和跳虫等主要分解类群的数量和多样性,从而导致人工梭梭林建植后期土壤全氮储量的下降[35,60-61].在人工梭梭林建植约20年时,通过降低灌木密度和盖度,增加挖掘兽类和节肢动物的多样性,以及提升关键捕食性动物种群的数量,可以显著增强人工梭梭林的生态系统服务功能,并提高其对气候变化和人类干扰的适应能力[13,21,48,62].总体而言,植被盖度、土壤质地与生物群落之间的耦合变化是驱动表层土壤养分储量变化的核心机制.因此,为了提升人工梭梭林的生态系统功能并实现可持续管理,有必要加强对建植过程中土壤环境和生物群落协同演变的长期监测,为相关理论研究和实践应用提供更为坚实的数据支持. ...
The role of Haloxylon species for combating desertification in central Asia
1
2002
... 梭梭作为适应干旱环境的典型植物,因其耐旱、耐盐碱等特性,在荒漠化治理和生态恢复中具有重要的应用价值[16-19].人工梭梭建植通过修复退化生境、降低风蚀程度、提高动物多样性以及促进土壤碳氮等养分固存,进而显著提升多种生态系统服务功能[13,20-21].人工梭梭林在建植初期通常密度较高,随着建植时间延长,林分密度有所下降.生长至20年左右梭梭林开始出现不同程度衰退,其生态系统服务功能也随之减弱[13].然而,也有研究发现,在绿洲边缘地区20~30年梭梭林往往能够形成相对稳定的斑块状结构,并在30年左右出现幼苗密度回升,表现出一定的自然更新潜力[22-24].降水和地下水位变化以及不同立地条件影响人工梭梭林的重建、演变过程,同时也对土壤恢复过程产生重要影响[22,25-27].在人工梭梭林建植过程中,土壤恢复进程相对缓慢,而土壤质量提升对人工林地生态系统长期健康与稳定具有重要意义[28-29].土壤碳、氮、磷是土壤养分的重要组成部分,其比值(C∶N∶P)不仅反映土壤养分相对平衡状态,还能揭示生态系统养分限制特征与功能特性[30].此外,表层土壤碳、氮和磷储量也是衡量土壤功能变化的重要指标.近年来,关于人工梭梭林建植过程中土壤碳、氮、磷含量、比值及储量变化的研究逐渐增多,但不同的降水条件、地下水位及土壤环境背景均会显著影响其响应模式和变化规律[31-33].在人工梭梭林建植过程中,植被、土壤动物等生物要素与土壤结构、盐分等非生物要素之间相互作用及其对表层土壤碳、氮、磷含量和储量变化的耦合影响认识较为有限.鉴于此,本文以张掖绿洲北部荒漠绿洲过渡带的流动沙丘、固定沙丘以及5、10、20、30年梭梭林为研究对象,分析人工梭梭林重建、演变过程中表层土壤有机碳、全氮和全磷比值及土壤有机碳、全氮、无机氮和全磷储量的变化规律,确定生物和非生物因素对表层土壤碳、氮、磷比值及储量变化的影响和相对贡献率,为绿洲边缘人工梭梭林土壤质量演变及生态系统服务功能变化研究提供科学依据. ...
我国现存梭梭荒漠植被地理分布及其斑块特征
0
2005
The potential geographical distribution of Haloxylon across central Asia under climate change in the 21st century
0
2019
中国梭梭属植物历史分布格局及其驱动机制
1
2022
... 梭梭作为适应干旱环境的典型植物,因其耐旱、耐盐碱等特性,在荒漠化治理和生态恢复中具有重要的应用价值[16-19].人工梭梭建植通过修复退化生境、降低风蚀程度、提高动物多样性以及促进土壤碳氮等养分固存,进而显著提升多种生态系统服务功能[13,20-21].人工梭梭林在建植初期通常密度较高,随着建植时间延长,林分密度有所下降.生长至20年左右梭梭林开始出现不同程度衰退,其生态系统服务功能也随之减弱[13].然而,也有研究发现,在绿洲边缘地区20~30年梭梭林往往能够形成相对稳定的斑块状结构,并在30年左右出现幼苗密度回升,表现出一定的自然更新潜力[22-24].降水和地下水位变化以及不同立地条件影响人工梭梭林的重建、演变过程,同时也对土壤恢复过程产生重要影响[22,25-27].在人工梭梭林建植过程中,土壤恢复进程相对缓慢,而土壤质量提升对人工林地生态系统长期健康与稳定具有重要意义[28-29].土壤碳、氮、磷是土壤养分的重要组成部分,其比值(C∶N∶P)不仅反映土壤养分相对平衡状态,还能揭示生态系统养分限制特征与功能特性[30].此外,表层土壤碳、氮和磷储量也是衡量土壤功能变化的重要指标.近年来,关于人工梭梭林建植过程中土壤碳、氮、磷含量、比值及储量变化的研究逐渐增多,但不同的降水条件、地下水位及土壤环境背景均会显著影响其响应模式和变化规律[31-33].在人工梭梭林建植过程中,植被、土壤动物等生物要素与土壤结构、盐分等非生物要素之间相互作用及其对表层土壤碳、氮、磷含量和储量变化的耦合影响认识较为有限.鉴于此,本文以张掖绿洲北部荒漠绿洲过渡带的流动沙丘、固定沙丘以及5、10、20、30年梭梭林为研究对象,分析人工梭梭林重建、演变过程中表层土壤有机碳、全氮和全磷比值及土壤有机碳、全氮、无机氮和全磷储量的变化规律,确定生物和非生物因素对表层土壤碳、氮、磷比值及储量变化的影响和相对贡献率,为绿洲边缘人工梭梭林土壤质量演变及生态系统服务功能变化研究提供科学依据. ...
荒漠绿洲过渡带人工梭梭(Haloxylon ammodendron)林下典型一年生草本植物空间格局
1
2025
... 梭梭作为适应干旱环境的典型植物,因其耐旱、耐盐碱等特性,在荒漠化治理和生态恢复中具有重要的应用价值[16-19].人工梭梭建植通过修复退化生境、降低风蚀程度、提高动物多样性以及促进土壤碳氮等养分固存,进而显著提升多种生态系统服务功能[13,20-21].人工梭梭林在建植初期通常密度较高,随着建植时间延长,林分密度有所下降.生长至20年左右梭梭林开始出现不同程度衰退,其生态系统服务功能也随之减弱[13].然而,也有研究发现,在绿洲边缘地区20~30年梭梭林往往能够形成相对稳定的斑块状结构,并在30年左右出现幼苗密度回升,表现出一定的自然更新潜力[22-24].降水和地下水位变化以及不同立地条件影响人工梭梭林的重建、演变过程,同时也对土壤恢复过程产生重要影响[22,25-27].在人工梭梭林建植过程中,土壤恢复进程相对缓慢,而土壤质量提升对人工林地生态系统长期健康与稳定具有重要意义[28-29].土壤碳、氮、磷是土壤养分的重要组成部分,其比值(C∶N∶P)不仅反映土壤养分相对平衡状态,还能揭示生态系统养分限制特征与功能特性[30].此外,表层土壤碳、氮和磷储量也是衡量土壤功能变化的重要指标.近年来,关于人工梭梭林建植过程中土壤碳、氮、磷含量、比值及储量变化的研究逐渐增多,但不同的降水条件、地下水位及土壤环境背景均会显著影响其响应模式和变化规律[31-33].在人工梭梭林建植过程中,植被、土壤动物等生物要素与土壤结构、盐分等非生物要素之间相互作用及其对表层土壤碳、氮、磷含量和储量变化的耦合影响认识较为有限.鉴于此,本文以张掖绿洲北部荒漠绿洲过渡带的流动沙丘、固定沙丘以及5、10、20、30年梭梭林为研究对象,分析人工梭梭林重建、演变过程中表层土壤有机碳、全氮和全磷比值及土壤有机碳、全氮、无机氮和全磷储量的变化规律,确定生物和非生物因素对表层土壤碳、氮、磷比值及储量变化的影响和相对贡献率,为绿洲边缘人工梭梭林土壤质量演变及生态系统服务功能变化研究提供科学依据. ...
黑河中游人工固沙植被恢复对爬行类和兽类动物多样性的影响
5
2024
... 梭梭作为适应干旱环境的典型植物,因其耐旱、耐盐碱等特性,在荒漠化治理和生态恢复中具有重要的应用价值[16-19].人工梭梭建植通过修复退化生境、降低风蚀程度、提高动物多样性以及促进土壤碳氮等养分固存,进而显著提升多种生态系统服务功能[13,20-21].人工梭梭林在建植初期通常密度较高,随着建植时间延长,林分密度有所下降.生长至20年左右梭梭林开始出现不同程度衰退,其生态系统服务功能也随之减弱[13].然而,也有研究发现,在绿洲边缘地区20~30年梭梭林往往能够形成相对稳定的斑块状结构,并在30年左右出现幼苗密度回升,表现出一定的自然更新潜力[22-24].降水和地下水位变化以及不同立地条件影响人工梭梭林的重建、演变过程,同时也对土壤恢复过程产生重要影响[22,25-27].在人工梭梭林建植过程中,土壤恢复进程相对缓慢,而土壤质量提升对人工林地生态系统长期健康与稳定具有重要意义[28-29].土壤碳、氮、磷是土壤养分的重要组成部分,其比值(C∶N∶P)不仅反映土壤养分相对平衡状态,还能揭示生态系统养分限制特征与功能特性[30].此外,表层土壤碳、氮和磷储量也是衡量土壤功能变化的重要指标.近年来,关于人工梭梭林建植过程中土壤碳、氮、磷含量、比值及储量变化的研究逐渐增多,但不同的降水条件、地下水位及土壤环境背景均会显著影响其响应模式和变化规律[31-33].在人工梭梭林建植过程中,植被、土壤动物等生物要素与土壤结构、盐分等非生物要素之间相互作用及其对表层土壤碳、氮、磷含量和储量变化的耦合影响认识较为有限.鉴于此,本文以张掖绿洲北部荒漠绿洲过渡带的流动沙丘、固定沙丘以及5、10、20、30年梭梭林为研究对象,分析人工梭梭林重建、演变过程中表层土壤有机碳、全氮和全磷比值及土壤有机碳、全氮、无机氮和全磷储量的变化规律,确定生物和非生物因素对表层土壤碳、氮、磷比值及储量变化的影响和相对贡献率,为绿洲边缘人工梭梭林土壤质量演变及生态系统服务功能变化研究提供科学依据. ...
... 研究区位于河西走廊中段,张掖绿洲北部的荒漠绿洲过渡带(39°21′53″—39°22′01″N,100°09′12″—100°09′14″E),该风沙带是巴丹吉林沙漠和张掖-临泽绿洲的交会处,海拔1 370 m.该区属温带大陆性荒漠气候,年平均气温为7.6 ℃,最低气温-10.7 ℃(1月),最高气温23.0 ℃(7月).多年平均降水量为117.0 mm,其中6、7、8、9月的降水量分别占全年降水量的13.6%、28.4%、16.7%、20.0%.年平均潜在蒸散量为2 390 mm,年平均日照时数达3 045 h[34].土壤类型为风沙土,地下水位4~5 m.天然木本植物有沙拐枣(Calligonum mongolicum)、泡泡刺(Nitraria sphaerocarpa)和小果白刺(Nitraria sibirica),草本植物主要有沙蓬(Agriophyllum pungens)、沙蒿(Artemisia desertorum)、白茎盐生草(Halogeton arachnoideus)、雾冰藜(Bassia dasyphylla)、沙鞭(Psammochloa villosa)和芦苇(Phragmites australis)等.人工栽植的灌木种有柽柳、柠条(Caragana korshinskii)、花棒(Corethrodendron scoparium)和梭梭等,梭梭是广泛使用的固沙灌木种.荒漠脊椎动物主要由爬行类、鸟类和兽类组成,荒漠沙蜥(Phrynocephalus przewalskii)、沙䳭(Oenanthe isabellina)、漠䳭(Oenanthe deserti)、楔尾伯劳(Lanius sphenocercus)、喜鹊(Pica pica)、鹅喉羚(Gazella subgutturosa)、虎鼬(Vormela peregusna)、赤狐(Vulpes vulpes)、亚洲狗獾(Meles leucurus)、兔狲(Otocolobus manul)和亚洲野猫(Felis silvestris)等是常见脊椎动物种,这些动物通过取食梭梭、草本和节肢动物以及挖掘土壤等直接和间接影响天然和人工固沙植被的健康与稳定[21]. ...
... 天然荒漠转变为人工梭梭林不仅改变了区域动植物多样性,还对土壤理化性质产生影响,这些改变会随着梭梭林建植年限的增加而变化[10,21,35,42].本研究表明,天然流动沙丘与固定沙丘之间土壤碳、氮、磷化学计量特征存在明显差异;随着人工梭梭林的建植,表层土壤化学计量特征亦发生改变.相较于流动沙丘,固定沙丘中灌木和草本植物的盖度及多样性明显提高,进而引发土壤C∶N下降和N∶P升高.上述结果表明,植被盖度提升加速表层土壤中氮素积累,其速率超过有机碳和全磷积累速率.然而,An等[43]在沙化草地研究发现,随着草地土壤沙化程度加剧,土壤C∶P和N∶P均下降,而C∶N变化不显著,这与本文的研究结果略有不同.泡泡刺和沙拐枣等灌木在荒漠生态系统中具有典型的“生态系统工程师”作用.这类灌木不仅富集枯落物并改善土壤微环境,也为其他生物提供了关键生态位即聚集了大型及中小型节肢动物,从而促进凋落物分解及氮素释放[21,44-45].在荒漠生态系统中,拟步甲科和蚁科等杂食性节肢动物是主要的食物网组成部分,其丰富度和多样性在灌木存在的环境中显著增加.这些节肢动物不仅直接参与凋落物的分解,还通过其死亡残体输入,提升了固定沙丘表层土壤中碳、氮、磷等养分含量[35].此外,研究还发现,研究区流动沙丘和固定沙丘土壤N∶P均低于1,显著低于全国土壤平均水平,进一步表明该区荒漠土壤存在明显的氮素限制[29,46].这种氮素限制特征可能与该区域干旱少雨、土壤有机质积累缓慢及微生物活动受限有关,同时也反映出人工固沙植被建植过程中,氮素补给和循环仍是限制生态系统功能提升的重要因素.梭梭是中国西北干旱区广泛分布的乔木型灌木种,不同梭梭种类及其分布区域会影响土壤碳、氮、磷化学计量比值.在流动和固定沙丘向人工梭梭林转变的过程中,表层土壤C∶N和N∶P发生了显著变化,而C∶P变化相对较小.这一结果与宁虎森等[47]在新疆甘家湖地区梭梭林的研究结果相似,但与魏亚娟等[32]在乌兰布和沙漠西南缘荒漠区梭梭林中的研究结果相反.宁虎森等[47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
... [21,44-45].在荒漠生态系统中,拟步甲科和蚁科等杂食性节肢动物是主要的食物网组成部分,其丰富度和多样性在灌木存在的环境中显著增加.这些节肢动物不仅直接参与凋落物的分解,还通过其死亡残体输入,提升了固定沙丘表层土壤中碳、氮、磷等养分含量[35].此外,研究还发现,研究区流动沙丘和固定沙丘土壤N∶P均低于1,显著低于全国土壤平均水平,进一步表明该区荒漠土壤存在明显的氮素限制[29,46].这种氮素限制特征可能与该区域干旱少雨、土壤有机质积累缓慢及微生物活动受限有关,同时也反映出人工固沙植被建植过程中,氮素补给和循环仍是限制生态系统功能提升的重要因素.梭梭是中国西北干旱区广泛分布的乔木型灌木种,不同梭梭种类及其分布区域会影响土壤碳、氮、磷化学计量比值.在流动和固定沙丘向人工梭梭林转变的过程中,表层土壤C∶N和N∶P发生了显著变化,而C∶P变化相对较小.这一结果与宁虎森等[47]在新疆甘家湖地区梭梭林的研究结果相似,但与魏亚娟等[32]在乌兰布和沙漠西南缘荒漠区梭梭林中的研究结果相反.宁虎森等[47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
... 在人工梭梭林建植过程中,非生物因子及其与生物因子的交互作用对表层土壤碳、氮、磷储量的影响大于生物因子的作用,且这些因子对土壤C∶N∶P比值的影响机制存在明显差异.本研究发现,土壤粒径组成和水盐条件等非生物因子在调控养分积累方面发挥着关键作用,这一结论与张掖绿洲边缘及其他干旱区的相关研究结果一致[52,54,59].梭梭林重建与演变过程能够有效削弱风沙活动,减少表层土壤流失,从而增强有机碳和氮积累,尤其在0~5 cm表层土壤中表现显著[54].此外,土壤黏粉粒比例增加可能是提升碳氮固存效率的重要因素[52].进一步分析表明,土壤黏粉粒含量、pH和全盐含量是非生物因子中影响人工梭梭林表层土壤有机碳、全氮和全磷储量变化的主要驱动因子.较高的黏粉粒含量有助于土壤结构的稳定性及有机质的高效积累,而pH值和盐分的变化则通过影响微生物活性及养分转化过程,进而调控土壤养分积累.特别是全盐含量的增加,促进了人工梭梭林下土壤盐结皮的形成,这一过程不仅抑制土壤水分入渗,还会限制草本植物及梭梭幼苗的萌发与生长,同时对土壤生物的活动产生负面影响,最终导致土壤全氮储量及其积累速率下降[15,51].安芳娇等[57]研究指出,在人工梭梭林的建植过程中,土壤pH值、有机碳含量和电导率是驱动线虫群落变化的主要环境因子,其中土壤电导率是影响线虫群落变化的关键因素.此外,灌木盖度、草本盖度及大型节肢动物数量是影响人工梭梭林建植过程中土壤碳、氮、磷储量变化的主要生物因子.灌木盖度与土壤黏粉粒之间的相互作用能够抑制风蚀,促进土壤细颗粒的聚集,进而提高表层土壤碳、氮、磷储量.同时,土壤中蚂蚁、蜘蛛及其他中小型土壤动物种类和数量的增加,加速了凋落物的分解过程[35].然而,土壤盐分增加及土壤水分有效性降低可能抑制了螨类和跳虫等主要分解类群的数量和多样性,从而导致人工梭梭林建植后期土壤全氮储量的下降[35,60-61].在人工梭梭林建植约20年时,通过降低灌木密度和盖度,增加挖掘兽类和节肢动物的多样性,以及提升关键捕食性动物种群的数量,可以显著增强人工梭梭林的生态系统服务功能,并提高其对气候变化和人类干扰的适应能力[13,21,48,62].总体而言,植被盖度、土壤质地与生物群落之间的耦合变化是驱动表层土壤养分储量变化的核心机制.因此,为了提升人工梭梭林的生态系统功能并实现可持续管理,有必要加强对建植过程中土壤环境和生物群落协同演变的长期监测,为相关理论研究和实践应用提供更为坚实的数据支持. ...
Spatial analysis of a Haloxylon ammodendron plantation in an oasis-desert ecotone in the Hexi Corridor,Northwestern China
3
2017
... 梭梭作为适应干旱环境的典型植物,因其耐旱、耐盐碱等特性,在荒漠化治理和生态恢复中具有重要的应用价值[16-19].人工梭梭建植通过修复退化生境、降低风蚀程度、提高动物多样性以及促进土壤碳氮等养分固存,进而显著提升多种生态系统服务功能[13,20-21].人工梭梭林在建植初期通常密度较高,随着建植时间延长,林分密度有所下降.生长至20年左右梭梭林开始出现不同程度衰退,其生态系统服务功能也随之减弱[13].然而,也有研究发现,在绿洲边缘地区20~30年梭梭林往往能够形成相对稳定的斑块状结构,并在30年左右出现幼苗密度回升,表现出一定的自然更新潜力[22-24].降水和地下水位变化以及不同立地条件影响人工梭梭林的重建、演变过程,同时也对土壤恢复过程产生重要影响[22,25-27].在人工梭梭林建植过程中,土壤恢复进程相对缓慢,而土壤质量提升对人工林地生态系统长期健康与稳定具有重要意义[28-29].土壤碳、氮、磷是土壤养分的重要组成部分,其比值(C∶N∶P)不仅反映土壤养分相对平衡状态,还能揭示生态系统养分限制特征与功能特性[30].此外,表层土壤碳、氮和磷储量也是衡量土壤功能变化的重要指标.近年来,关于人工梭梭林建植过程中土壤碳、氮、磷含量、比值及储量变化的研究逐渐增多,但不同的降水条件、地下水位及土壤环境背景均会显著影响其响应模式和变化规律[31-33].在人工梭梭林建植过程中,植被、土壤动物等生物要素与土壤结构、盐分等非生物要素之间相互作用及其对表层土壤碳、氮、磷含量和储量变化的耦合影响认识较为有限.鉴于此,本文以张掖绿洲北部荒漠绿洲过渡带的流动沙丘、固定沙丘以及5、10、20、30年梭梭林为研究对象,分析人工梭梭林重建、演变过程中表层土壤有机碳、全氮和全磷比值及土壤有机碳、全氮、无机氮和全磷储量的变化规律,确定生物和非生物因素对表层土壤碳、氮、磷比值及储量变化的影响和相对贡献率,为绿洲边缘人工梭梭林土壤质量演变及生态系统服务功能变化研究提供科学依据. ...
... [22,25-27].在人工梭梭林建植过程中,土壤恢复进程相对缓慢,而土壤质量提升对人工林地生态系统长期健康与稳定具有重要意义[28-29].土壤碳、氮、磷是土壤养分的重要组成部分,其比值(C∶N∶P)不仅反映土壤养分相对平衡状态,还能揭示生态系统养分限制特征与功能特性[30].此外,表层土壤碳、氮和磷储量也是衡量土壤功能变化的重要指标.近年来,关于人工梭梭林建植过程中土壤碳、氮、磷含量、比值及储量变化的研究逐渐增多,但不同的降水条件、地下水位及土壤环境背景均会显著影响其响应模式和变化规律[31-33].在人工梭梭林建植过程中,植被、土壤动物等生物要素与土壤结构、盐分等非生物要素之间相互作用及其对表层土壤碳、氮、磷含量和储量变化的耦合影响认识较为有限.鉴于此,本文以张掖绿洲北部荒漠绿洲过渡带的流动沙丘、固定沙丘以及5、10、20、30年梭梭林为研究对象,分析人工梭梭林重建、演变过程中表层土壤有机碳、全氮和全磷比值及土壤有机碳、全氮、无机氮和全磷储量的变化规律,确定生物和非生物因素对表层土壤碳、氮、磷比值及储量变化的影响和相对贡献率,为绿洲边缘人工梭梭林土壤质量演变及生态系统服务功能变化研究提供科学依据. ...
... 天然荒漠转变为人工梭梭林不仅改变了区域动植物多样性,还对土壤理化性质产生影响,这些改变会随着梭梭林建植年限的增加而变化[10,21,35,42].本研究表明,天然流动沙丘与固定沙丘之间土壤碳、氮、磷化学计量特征存在明显差异;随着人工梭梭林的建植,表层土壤化学计量特征亦发生改变.相较于流动沙丘,固定沙丘中灌木和草本植物的盖度及多样性明显提高,进而引发土壤C∶N下降和N∶P升高.上述结果表明,植被盖度提升加速表层土壤中氮素积累,其速率超过有机碳和全磷积累速率.然而,An等[43]在沙化草地研究发现,随着草地土壤沙化程度加剧,土壤C∶P和N∶P均下降,而C∶N变化不显著,这与本文的研究结果略有不同.泡泡刺和沙拐枣等灌木在荒漠生态系统中具有典型的“生态系统工程师”作用.这类灌木不仅富集枯落物并改善土壤微环境,也为其他生物提供了关键生态位即聚集了大型及中小型节肢动物,从而促进凋落物分解及氮素释放[21,44-45].在荒漠生态系统中,拟步甲科和蚁科等杂食性节肢动物是主要的食物网组成部分,其丰富度和多样性在灌木存在的环境中显著增加.这些节肢动物不仅直接参与凋落物的分解,还通过其死亡残体输入,提升了固定沙丘表层土壤中碳、氮、磷等养分含量[35].此外,研究还发现,研究区流动沙丘和固定沙丘土壤N∶P均低于1,显著低于全国土壤平均水平,进一步表明该区荒漠土壤存在明显的氮素限制[29,46].这种氮素限制特征可能与该区域干旱少雨、土壤有机质积累缓慢及微生物活动受限有关,同时也反映出人工固沙植被建植过程中,氮素补给和循环仍是限制生态系统功能提升的重要因素.梭梭是中国西北干旱区广泛分布的乔木型灌木种,不同梭梭种类及其分布区域会影响土壤碳、氮、磷化学计量比值.在流动和固定沙丘向人工梭梭林转变的过程中,表层土壤C∶N和N∶P发生了显著变化,而C∶P变化相对较小.这一结果与宁虎森等[47]在新疆甘家湖地区梭梭林的研究结果相似,但与魏亚娟等[32]在乌兰布和沙漠西南缘荒漠区梭梭林中的研究结果相反.宁虎森等[47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
人工梭梭林建植对荒漠土壤特征的长期影响
2
2025
... 梭梭作为适应干旱环境的典型植物,因其耐旱、耐盐碱等特性,在荒漠化治理和生态恢复中具有重要的应用价值[16-19].人工梭梭建植通过修复退化生境、降低风蚀程度、提高动物多样性以及促进土壤碳氮等养分固存,进而显著提升多种生态系统服务功能[13,20-21].人工梭梭林在建植初期通常密度较高,随着建植时间延长,林分密度有所下降.生长至20年左右梭梭林开始出现不同程度衰退,其生态系统服务功能也随之减弱[13].然而,也有研究发现,在绿洲边缘地区20~30年梭梭林往往能够形成相对稳定的斑块状结构,并在30年左右出现幼苗密度回升,表现出一定的自然更新潜力[22-24].降水和地下水位变化以及不同立地条件影响人工梭梭林的重建、演变过程,同时也对土壤恢复过程产生重要影响[22,25-27].在人工梭梭林建植过程中,土壤恢复进程相对缓慢,而土壤质量提升对人工林地生态系统长期健康与稳定具有重要意义[28-29].土壤碳、氮、磷是土壤养分的重要组成部分,其比值(C∶N∶P)不仅反映土壤养分相对平衡状态,还能揭示生态系统养分限制特征与功能特性[30].此外,表层土壤碳、氮和磷储量也是衡量土壤功能变化的重要指标.近年来,关于人工梭梭林建植过程中土壤碳、氮、磷含量、比值及储量变化的研究逐渐增多,但不同的降水条件、地下水位及土壤环境背景均会显著影响其响应模式和变化规律[31-33].在人工梭梭林建植过程中,植被、土壤动物等生物要素与土壤结构、盐分等非生物要素之间相互作用及其对表层土壤碳、氮、磷含量和储量变化的耦合影响认识较为有限.鉴于此,本文以张掖绿洲北部荒漠绿洲过渡带的流动沙丘、固定沙丘以及5、10、20、30年梭梭林为研究对象,分析人工梭梭林重建、演变过程中表层土壤有机碳、全氮和全磷比值及土壤有机碳、全氮、无机氮和全磷储量的变化规律,确定生物和非生物因素对表层土壤碳、氮、磷比值及储量变化的影响和相对贡献率,为绿洲边缘人工梭梭林土壤质量演变及生态系统服务功能变化研究提供科学依据. ...
... 天然荒漠转变为人工梭梭林不仅改变了区域动植物多样性,还对土壤理化性质产生影响,这些改变会随着梭梭林建植年限的增加而变化[10,21,35,42].本研究表明,天然流动沙丘与固定沙丘之间土壤碳、氮、磷化学计量特征存在明显差异;随着人工梭梭林的建植,表层土壤化学计量特征亦发生改变.相较于流动沙丘,固定沙丘中灌木和草本植物的盖度及多样性明显提高,进而引发土壤C∶N下降和N∶P升高.上述结果表明,植被盖度提升加速表层土壤中氮素积累,其速率超过有机碳和全磷积累速率.然而,An等[43]在沙化草地研究发现,随着草地土壤沙化程度加剧,土壤C∶P和N∶P均下降,而C∶N变化不显著,这与本文的研究结果略有不同.泡泡刺和沙拐枣等灌木在荒漠生态系统中具有典型的“生态系统工程师”作用.这类灌木不仅富集枯落物并改善土壤微环境,也为其他生物提供了关键生态位即聚集了大型及中小型节肢动物,从而促进凋落物分解及氮素释放[21,44-45].在荒漠生态系统中,拟步甲科和蚁科等杂食性节肢动物是主要的食物网组成部分,其丰富度和多样性在灌木存在的环境中显著增加.这些节肢动物不仅直接参与凋落物的分解,还通过其死亡残体输入,提升了固定沙丘表层土壤中碳、氮、磷等养分含量[35].此外,研究还发现,研究区流动沙丘和固定沙丘土壤N∶P均低于1,显著低于全国土壤平均水平,进一步表明该区荒漠土壤存在明显的氮素限制[29,46].这种氮素限制特征可能与该区域干旱少雨、土壤有机质积累缓慢及微生物活动受限有关,同时也反映出人工固沙植被建植过程中,氮素补给和循环仍是限制生态系统功能提升的重要因素.梭梭是中国西北干旱区广泛分布的乔木型灌木种,不同梭梭种类及其分布区域会影响土壤碳、氮、磷化学计量比值.在流动和固定沙丘向人工梭梭林转变的过程中,表层土壤C∶N和N∶P发生了显著变化,而C∶P变化相对较小.这一结果与宁虎森等[47]在新疆甘家湖地区梭梭林的研究结果相似,但与魏亚娟等[32]在乌兰布和沙漠西南缘荒漠区梭梭林中的研究结果相反.宁虎森等[47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
库姆塔格沙漠梭梭群落特征研究
1
2007
... 梭梭作为适应干旱环境的典型植物,因其耐旱、耐盐碱等特性,在荒漠化治理和生态恢复中具有重要的应用价值[16-19].人工梭梭建植通过修复退化生境、降低风蚀程度、提高动物多样性以及促进土壤碳氮等养分固存,进而显著提升多种生态系统服务功能[13,20-21].人工梭梭林在建植初期通常密度较高,随着建植时间延长,林分密度有所下降.生长至20年左右梭梭林开始出现不同程度衰退,其生态系统服务功能也随之减弱[13].然而,也有研究发现,在绿洲边缘地区20~30年梭梭林往往能够形成相对稳定的斑块状结构,并在30年左右出现幼苗密度回升,表现出一定的自然更新潜力[22-24].降水和地下水位变化以及不同立地条件影响人工梭梭林的重建、演变过程,同时也对土壤恢复过程产生重要影响[22,25-27].在人工梭梭林建植过程中,土壤恢复进程相对缓慢,而土壤质量提升对人工林地生态系统长期健康与稳定具有重要意义[28-29].土壤碳、氮、磷是土壤养分的重要组成部分,其比值(C∶N∶P)不仅反映土壤养分相对平衡状态,还能揭示生态系统养分限制特征与功能特性[30].此外,表层土壤碳、氮和磷储量也是衡量土壤功能变化的重要指标.近年来,关于人工梭梭林建植过程中土壤碳、氮、磷含量、比值及储量变化的研究逐渐增多,但不同的降水条件、地下水位及土壤环境背景均会显著影响其响应模式和变化规律[31-33].在人工梭梭林建植过程中,植被、土壤动物等生物要素与土壤结构、盐分等非生物要素之间相互作用及其对表层土壤碳、氮、磷含量和储量变化的耦合影响认识较为有限.鉴于此,本文以张掖绿洲北部荒漠绿洲过渡带的流动沙丘、固定沙丘以及5、10、20、30年梭梭林为研究对象,分析人工梭梭林重建、演变过程中表层土壤有机碳、全氮和全磷比值及土壤有机碳、全氮、无机氮和全磷储量的变化规律,确定生物和非生物因素对表层土壤碳、氮、磷比值及储量变化的影响和相对贡献率,为绿洲边缘人工梭梭林土壤质量演变及生态系统服务功能变化研究提供科学依据. ...
古尔班通古特沙漠土壤因素对退化梭梭更新局限的影响
0
2010
准噶尔盆地荒漠绿洲过渡带地下水埋深对人工梭梭林年龄结构及动态特征的影响
1
2024
... 梭梭作为适应干旱环境的典型植物,因其耐旱、耐盐碱等特性,在荒漠化治理和生态恢复中具有重要的应用价值[16-19].人工梭梭建植通过修复退化生境、降低风蚀程度、提高动物多样性以及促进土壤碳氮等养分固存,进而显著提升多种生态系统服务功能[13,20-21].人工梭梭林在建植初期通常密度较高,随着建植时间延长,林分密度有所下降.生长至20年左右梭梭林开始出现不同程度衰退,其生态系统服务功能也随之减弱[13].然而,也有研究发现,在绿洲边缘地区20~30年梭梭林往往能够形成相对稳定的斑块状结构,并在30年左右出现幼苗密度回升,表现出一定的自然更新潜力[22-24].降水和地下水位变化以及不同立地条件影响人工梭梭林的重建、演变过程,同时也对土壤恢复过程产生重要影响[22,25-27].在人工梭梭林建植过程中,土壤恢复进程相对缓慢,而土壤质量提升对人工林地生态系统长期健康与稳定具有重要意义[28-29].土壤碳、氮、磷是土壤养分的重要组成部分,其比值(C∶N∶P)不仅反映土壤养分相对平衡状态,还能揭示生态系统养分限制特征与功能特性[30].此外,表层土壤碳、氮和磷储量也是衡量土壤功能变化的重要指标.近年来,关于人工梭梭林建植过程中土壤碳、氮、磷含量、比值及储量变化的研究逐渐增多,但不同的降水条件、地下水位及土壤环境背景均会显著影响其响应模式和变化规律[31-33].在人工梭梭林建植过程中,植被、土壤动物等生物要素与土壤结构、盐分等非生物要素之间相互作用及其对表层土壤碳、氮、磷含量和储量变化的耦合影响认识较为有限.鉴于此,本文以张掖绿洲北部荒漠绿洲过渡带的流动沙丘、固定沙丘以及5、10、20、30年梭梭林为研究对象,分析人工梭梭林重建、演变过程中表层土壤有机碳、全氮和全磷比值及土壤有机碳、全氮、无机氮和全磷储量的变化规律,确定生物和非生物因素对表层土壤碳、氮、磷比值及储量变化的影响和相对贡献率,为绿洲边缘人工梭梭林土壤质量演变及生态系统服务功能变化研究提供科学依据. ...
中国荒漠植物-土壤系统生态化学计量学研究进展
1
2022
... 梭梭作为适应干旱环境的典型植物,因其耐旱、耐盐碱等特性,在荒漠化治理和生态恢复中具有重要的应用价值[16-19].人工梭梭建植通过修复退化生境、降低风蚀程度、提高动物多样性以及促进土壤碳氮等养分固存,进而显著提升多种生态系统服务功能[13,20-21].人工梭梭林在建植初期通常密度较高,随着建植时间延长,林分密度有所下降.生长至20年左右梭梭林开始出现不同程度衰退,其生态系统服务功能也随之减弱[13].然而,也有研究发现,在绿洲边缘地区20~30年梭梭林往往能够形成相对稳定的斑块状结构,并在30年左右出现幼苗密度回升,表现出一定的自然更新潜力[22-24].降水和地下水位变化以及不同立地条件影响人工梭梭林的重建、演变过程,同时也对土壤恢复过程产生重要影响[22,25-27].在人工梭梭林建植过程中,土壤恢复进程相对缓慢,而土壤质量提升对人工林地生态系统长期健康与稳定具有重要意义[28-29].土壤碳、氮、磷是土壤养分的重要组成部分,其比值(C∶N∶P)不仅反映土壤养分相对平衡状态,还能揭示生态系统养分限制特征与功能特性[30].此外,表层土壤碳、氮和磷储量也是衡量土壤功能变化的重要指标.近年来,关于人工梭梭林建植过程中土壤碳、氮、磷含量、比值及储量变化的研究逐渐增多,但不同的降水条件、地下水位及土壤环境背景均会显著影响其响应模式和变化规律[31-33].在人工梭梭林建植过程中,植被、土壤动物等生物要素与土壤结构、盐分等非生物要素之间相互作用及其对表层土壤碳、氮、磷含量和储量变化的耦合影响认识较为有限.鉴于此,本文以张掖绿洲北部荒漠绿洲过渡带的流动沙丘、固定沙丘以及5、10、20、30年梭梭林为研究对象,分析人工梭梭林重建、演变过程中表层土壤有机碳、全氮和全磷比值及土壤有机碳、全氮、无机氮和全磷储量的变化规律,确定生物和非生物因素对表层土壤碳、氮、磷比值及储量变化的影响和相对贡献率,为绿洲边缘人工梭梭林土壤质量演变及生态系统服务功能变化研究提供科学依据. ...
中国土壤碳氮磷及生态化学计量研究综述
2
2024
... 梭梭作为适应干旱环境的典型植物,因其耐旱、耐盐碱等特性,在荒漠化治理和生态恢复中具有重要的应用价值[16-19].人工梭梭建植通过修复退化生境、降低风蚀程度、提高动物多样性以及促进土壤碳氮等养分固存,进而显著提升多种生态系统服务功能[13,20-21].人工梭梭林在建植初期通常密度较高,随着建植时间延长,林分密度有所下降.生长至20年左右梭梭林开始出现不同程度衰退,其生态系统服务功能也随之减弱[13].然而,也有研究发现,在绿洲边缘地区20~30年梭梭林往往能够形成相对稳定的斑块状结构,并在30年左右出现幼苗密度回升,表现出一定的自然更新潜力[22-24].降水和地下水位变化以及不同立地条件影响人工梭梭林的重建、演变过程,同时也对土壤恢复过程产生重要影响[22,25-27].在人工梭梭林建植过程中,土壤恢复进程相对缓慢,而土壤质量提升对人工林地生态系统长期健康与稳定具有重要意义[28-29].土壤碳、氮、磷是土壤养分的重要组成部分,其比值(C∶N∶P)不仅反映土壤养分相对平衡状态,还能揭示生态系统养分限制特征与功能特性[30].此外,表层土壤碳、氮和磷储量也是衡量土壤功能变化的重要指标.近年来,关于人工梭梭林建植过程中土壤碳、氮、磷含量、比值及储量变化的研究逐渐增多,但不同的降水条件、地下水位及土壤环境背景均会显著影响其响应模式和变化规律[31-33].在人工梭梭林建植过程中,植被、土壤动物等生物要素与土壤结构、盐分等非生物要素之间相互作用及其对表层土壤碳、氮、磷含量和储量变化的耦合影响认识较为有限.鉴于此,本文以张掖绿洲北部荒漠绿洲过渡带的流动沙丘、固定沙丘以及5、10、20、30年梭梭林为研究对象,分析人工梭梭林重建、演变过程中表层土壤有机碳、全氮和全磷比值及土壤有机碳、全氮、无机氮和全磷储量的变化规律,确定生物和非生物因素对表层土壤碳、氮、磷比值及储量变化的影响和相对贡献率,为绿洲边缘人工梭梭林土壤质量演变及生态系统服务功能变化研究提供科学依据. ...
... 天然荒漠转变为人工梭梭林不仅改变了区域动植物多样性,还对土壤理化性质产生影响,这些改变会随着梭梭林建植年限的增加而变化[10,21,35,42].本研究表明,天然流动沙丘与固定沙丘之间土壤碳、氮、磷化学计量特征存在明显差异;随着人工梭梭林的建植,表层土壤化学计量特征亦发生改变.相较于流动沙丘,固定沙丘中灌木和草本植物的盖度及多样性明显提高,进而引发土壤C∶N下降和N∶P升高.上述结果表明,植被盖度提升加速表层土壤中氮素积累,其速率超过有机碳和全磷积累速率.然而,An等[43]在沙化草地研究发现,随着草地土壤沙化程度加剧,土壤C∶P和N∶P均下降,而C∶N变化不显著,这与本文的研究结果略有不同.泡泡刺和沙拐枣等灌木在荒漠生态系统中具有典型的“生态系统工程师”作用.这类灌木不仅富集枯落物并改善土壤微环境,也为其他生物提供了关键生态位即聚集了大型及中小型节肢动物,从而促进凋落物分解及氮素释放[21,44-45].在荒漠生态系统中,拟步甲科和蚁科等杂食性节肢动物是主要的食物网组成部分,其丰富度和多样性在灌木存在的环境中显著增加.这些节肢动物不仅直接参与凋落物的分解,还通过其死亡残体输入,提升了固定沙丘表层土壤中碳、氮、磷等养分含量[35].此外,研究还发现,研究区流动沙丘和固定沙丘土壤N∶P均低于1,显著低于全国土壤平均水平,进一步表明该区荒漠土壤存在明显的氮素限制[29,46].这种氮素限制特征可能与该区域干旱少雨、土壤有机质积累缓慢及微生物活动受限有关,同时也反映出人工固沙植被建植过程中,氮素补给和循环仍是限制生态系统功能提升的重要因素.梭梭是中国西北干旱区广泛分布的乔木型灌木种,不同梭梭种类及其分布区域会影响土壤碳、氮、磷化学计量比值.在流动和固定沙丘向人工梭梭林转变的过程中,表层土壤C∶N和N∶P发生了显著变化,而C∶P变化相对较小.这一结果与宁虎森等[47]在新疆甘家湖地区梭梭林的研究结果相似,但与魏亚娟等[32]在乌兰布和沙漠西南缘荒漠区梭梭林中的研究结果相反.宁虎森等[47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
不同生境下三种荒漠植物叶片及土壤C、N、P的化学计量特征
1
2014
... 梭梭作为适应干旱环境的典型植物,因其耐旱、耐盐碱等特性,在荒漠化治理和生态恢复中具有重要的应用价值[16-19].人工梭梭建植通过修复退化生境、降低风蚀程度、提高动物多样性以及促进土壤碳氮等养分固存,进而显著提升多种生态系统服务功能[13,20-21].人工梭梭林在建植初期通常密度较高,随着建植时间延长,林分密度有所下降.生长至20年左右梭梭林开始出现不同程度衰退,其生态系统服务功能也随之减弱[13].然而,也有研究发现,在绿洲边缘地区20~30年梭梭林往往能够形成相对稳定的斑块状结构,并在30年左右出现幼苗密度回升,表现出一定的自然更新潜力[22-24].降水和地下水位变化以及不同立地条件影响人工梭梭林的重建、演变过程,同时也对土壤恢复过程产生重要影响[22,25-27].在人工梭梭林建植过程中,土壤恢复进程相对缓慢,而土壤质量提升对人工林地生态系统长期健康与稳定具有重要意义[28-29].土壤碳、氮、磷是土壤养分的重要组成部分,其比值(C∶N∶P)不仅反映土壤养分相对平衡状态,还能揭示生态系统养分限制特征与功能特性[30].此外,表层土壤碳、氮和磷储量也是衡量土壤功能变化的重要指标.近年来,关于人工梭梭林建植过程中土壤碳、氮、磷含量、比值及储量变化的研究逐渐增多,但不同的降水条件、地下水位及土壤环境背景均会显著影响其响应模式和变化规律[31-33].在人工梭梭林建植过程中,植被、土壤动物等生物要素与土壤结构、盐分等非生物要素之间相互作用及其对表层土壤碳、氮、磷含量和储量变化的耦合影响认识较为有限.鉴于此,本文以张掖绿洲北部荒漠绿洲过渡带的流动沙丘、固定沙丘以及5、10、20、30年梭梭林为研究对象,分析人工梭梭林重建、演变过程中表层土壤有机碳、全氮和全磷比值及土壤有机碳、全氮、无机氮和全磷储量的变化规律,确定生物和非生物因素对表层土壤碳、氮、磷比值及储量变化的影响和相对贡献率,为绿洲边缘人工梭梭林土壤质量演变及生态系统服务功能变化研究提供科学依据. ...
荒漠绿洲区不同种植年限人工梭梭林土壤化学计量特征
3
2016
... 梭梭作为适应干旱环境的典型植物,因其耐旱、耐盐碱等特性,在荒漠化治理和生态恢复中具有重要的应用价值[16-19].人工梭梭建植通过修复退化生境、降低风蚀程度、提高动物多样性以及促进土壤碳氮等养分固存,进而显著提升多种生态系统服务功能[13,20-21].人工梭梭林在建植初期通常密度较高,随着建植时间延长,林分密度有所下降.生长至20年左右梭梭林开始出现不同程度衰退,其生态系统服务功能也随之减弱[13].然而,也有研究发现,在绿洲边缘地区20~30年梭梭林往往能够形成相对稳定的斑块状结构,并在30年左右出现幼苗密度回升,表现出一定的自然更新潜力[22-24].降水和地下水位变化以及不同立地条件影响人工梭梭林的重建、演变过程,同时也对土壤恢复过程产生重要影响[22,25-27].在人工梭梭林建植过程中,土壤恢复进程相对缓慢,而土壤质量提升对人工林地生态系统长期健康与稳定具有重要意义[28-29].土壤碳、氮、磷是土壤养分的重要组成部分,其比值(C∶N∶P)不仅反映土壤养分相对平衡状态,还能揭示生态系统养分限制特征与功能特性[30].此外,表层土壤碳、氮和磷储量也是衡量土壤功能变化的重要指标.近年来,关于人工梭梭林建植过程中土壤碳、氮、磷含量、比值及储量变化的研究逐渐增多,但不同的降水条件、地下水位及土壤环境背景均会显著影响其响应模式和变化规律[31-33].在人工梭梭林建植过程中,植被、土壤动物等生物要素与土壤结构、盐分等非生物要素之间相互作用及其对表层土壤碳、氮、磷含量和储量变化的耦合影响认识较为有限.鉴于此,本文以张掖绿洲北部荒漠绿洲过渡带的流动沙丘、固定沙丘以及5、10、20、30年梭梭林为研究对象,分析人工梭梭林重建、演变过程中表层土壤有机碳、全氮和全磷比值及土壤有机碳、全氮、无机氮和全磷储量的变化规律,确定生物和非生物因素对表层土壤碳、氮、磷比值及储量变化的影响和相对贡献率,为绿洲边缘人工梭梭林土壤质量演变及生态系统服务功能变化研究提供科学依据. ...
... 天然荒漠转变为人工梭梭林不仅改变了区域动植物多样性,还对土壤理化性质产生影响,这些改变会随着梭梭林建植年限的增加而变化[10,21,35,42].本研究表明,天然流动沙丘与固定沙丘之间土壤碳、氮、磷化学计量特征存在明显差异;随着人工梭梭林的建植,表层土壤化学计量特征亦发生改变.相较于流动沙丘,固定沙丘中灌木和草本植物的盖度及多样性明显提高,进而引发土壤C∶N下降和N∶P升高.上述结果表明,植被盖度提升加速表层土壤中氮素积累,其速率超过有机碳和全磷积累速率.然而,An等[43]在沙化草地研究发现,随着草地土壤沙化程度加剧,土壤C∶P和N∶P均下降,而C∶N变化不显著,这与本文的研究结果略有不同.泡泡刺和沙拐枣等灌木在荒漠生态系统中具有典型的“生态系统工程师”作用.这类灌木不仅富集枯落物并改善土壤微环境,也为其他生物提供了关键生态位即聚集了大型及中小型节肢动物,从而促进凋落物分解及氮素释放[21,44-45].在荒漠生态系统中,拟步甲科和蚁科等杂食性节肢动物是主要的食物网组成部分,其丰富度和多样性在灌木存在的环境中显著增加.这些节肢动物不仅直接参与凋落物的分解,还通过其死亡残体输入,提升了固定沙丘表层土壤中碳、氮、磷等养分含量[35].此外,研究还发现,研究区流动沙丘和固定沙丘土壤N∶P均低于1,显著低于全国土壤平均水平,进一步表明该区荒漠土壤存在明显的氮素限制[29,46].这种氮素限制特征可能与该区域干旱少雨、土壤有机质积累缓慢及微生物活动受限有关,同时也反映出人工固沙植被建植过程中,氮素补给和循环仍是限制生态系统功能提升的重要因素.梭梭是中国西北干旱区广泛分布的乔木型灌木种,不同梭梭种类及其分布区域会影响土壤碳、氮、磷化学计量比值.在流动和固定沙丘向人工梭梭林转变的过程中,表层土壤C∶N和N∶P发生了显著变化,而C∶P变化相对较小.这一结果与宁虎森等[47]在新疆甘家湖地区梭梭林的研究结果相似,但与魏亚娟等[32]在乌兰布和沙漠西南缘荒漠区梭梭林中的研究结果相反.宁虎森等[47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
... [31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
干旱荒漠区人工梭梭林土壤碳氮磷密度与生态化学计量特征
5
2022
... 天然荒漠转变为人工梭梭林不仅改变了区域动植物多样性,还对土壤理化性质产生影响,这些改变会随着梭梭林建植年限的增加而变化[10,21,35,42].本研究表明,天然流动沙丘与固定沙丘之间土壤碳、氮、磷化学计量特征存在明显差异;随着人工梭梭林的建植,表层土壤化学计量特征亦发生改变.相较于流动沙丘,固定沙丘中灌木和草本植物的盖度及多样性明显提高,进而引发土壤C∶N下降和N∶P升高.上述结果表明,植被盖度提升加速表层土壤中氮素积累,其速率超过有机碳和全磷积累速率.然而,An等[43]在沙化草地研究发现,随着草地土壤沙化程度加剧,土壤C∶P和N∶P均下降,而C∶N变化不显著,这与本文的研究结果略有不同.泡泡刺和沙拐枣等灌木在荒漠生态系统中具有典型的“生态系统工程师”作用.这类灌木不仅富集枯落物并改善土壤微环境,也为其他生物提供了关键生态位即聚集了大型及中小型节肢动物,从而促进凋落物分解及氮素释放[21,44-45].在荒漠生态系统中,拟步甲科和蚁科等杂食性节肢动物是主要的食物网组成部分,其丰富度和多样性在灌木存在的环境中显著增加.这些节肢动物不仅直接参与凋落物的分解,还通过其死亡残体输入,提升了固定沙丘表层土壤中碳、氮、磷等养分含量[35].此外,研究还发现,研究区流动沙丘和固定沙丘土壤N∶P均低于1,显著低于全国土壤平均水平,进一步表明该区荒漠土壤存在明显的氮素限制[29,46].这种氮素限制特征可能与该区域干旱少雨、土壤有机质积累缓慢及微生物活动受限有关,同时也反映出人工固沙植被建植过程中,氮素补给和循环仍是限制生态系统功能提升的重要因素.梭梭是中国西北干旱区广泛分布的乔木型灌木种,不同梭梭种类及其分布区域会影响土壤碳、氮、磷化学计量比值.在流动和固定沙丘向人工梭梭林转变的过程中,表层土壤C∶N和N∶P发生了显著变化,而C∶P变化相对较小.这一结果与宁虎森等[47]在新疆甘家湖地区梭梭林的研究结果相似,但与魏亚娟等[32]在乌兰布和沙漠西南缘荒漠区梭梭林中的研究结果相反.宁虎森等[47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
... [32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
... 天然和人工梭梭林表层土壤碳、氮、磷含量均高于流动沙地.木本植被的存在及盖度增加,有助于提升荒漠土壤的养分含量与储量,而表层土壤碳、氮、磷储量对植被建植较为敏感[51,53-54].本研究发现,固定沙丘土壤有机碳、全氮和全磷储量均高于流动沙丘,这一趋势与腾格里沙漠和乌兰布和沙漠等区域的研究结果相近[32,54-55].然而,流动和固定沙丘表层土壤的无机氮储量变化相对较小,这可能与该区域土壤水分有效性较低,从而抑制土壤生物的活动有关[56].在人工梭梭林恢复的前期阶段(<20年),表层土壤有机碳、全磷和无机氮储量变化不明显,唯有土壤全氮储量显著增加,这与天然荒漠转变为梭梭林后,节肢动物、线虫及微生物多样性提高有关[35,50,57].魏亚娟等[32]在乌兰布和沙漠的研究也发现,在3~11年人工梭梭林中,土壤有机碳储量变化不大,而在16年时则显著提升,这与本研究结果相近.值得注意的是,在人工梭梭林恢复早期,表层土壤全氮和全磷储量随建植年限的增长而增加,但这一趋势与魏亚娟等[32]的研究稍有不同.Ma等[58]在石羊河流域的研究显示,人工梭梭林土壤有机碳储量随梭梭林建植年限增加而提升,但仍显著低于流动沙丘.本研究进一步表明,在人工梭梭林恢复的后期阶段(>20年),表层土壤的有机碳、无机氮和全磷储量随着建植年限延长而显著增加,但全氮储量在30年林龄时出现显著下降.这一变化趋势与Huang等[13]的研究相一致,即人工梭梭林的生态恢复效应在建植约20年时达到峰值,超过20年后则需要人工干预以增强其生态服务功能.在绿洲边缘区域,人工梭梭林表层土壤的有机碳、无机氮和全磷储量随梭梭林龄的延长而增长,而表层土壤全氮储量则先升后降.这可能与恢复后期表层土壤盐分的积累有关,表层土壤盐分积累抑制了土壤生物活动,进而降低了土壤全氮含量[15]. ...
... [32]在乌兰布和沙漠的研究也发现,在3~11年人工梭梭林中,土壤有机碳储量变化不大,而在16年时则显著提升,这与本研究结果相近.值得注意的是,在人工梭梭林恢复早期,表层土壤全氮和全磷储量随建植年限的增长而增加,但这一趋势与魏亚娟等[32]的研究稍有不同.Ma等[58]在石羊河流域的研究显示,人工梭梭林土壤有机碳储量随梭梭林建植年限增加而提升,但仍显著低于流动沙丘.本研究进一步表明,在人工梭梭林恢复的后期阶段(>20年),表层土壤的有机碳、无机氮和全磷储量随着建植年限延长而显著增加,但全氮储量在30年林龄时出现显著下降.这一变化趋势与Huang等[13]的研究相一致,即人工梭梭林的生态恢复效应在建植约20年时达到峰值,超过20年后则需要人工干预以增强其生态服务功能.在绿洲边缘区域,人工梭梭林表层土壤的有机碳、无机氮和全磷储量随梭梭林龄的延长而增长,而表层土壤全氮储量则先升后降.这可能与恢复后期表层土壤盐分的积累有关,表层土壤盐分积累抑制了土壤生物活动,进而降低了土壤全氮含量[15]. ...
... [32]的研究稍有不同.Ma等[58]在石羊河流域的研究显示,人工梭梭林土壤有机碳储量随梭梭林建植年限增加而提升,但仍显著低于流动沙丘.本研究进一步表明,在人工梭梭林恢复的后期阶段(>20年),表层土壤的有机碳、无机氮和全磷储量随着建植年限延长而显著增加,但全氮储量在30年林龄时出现显著下降.这一变化趋势与Huang等[13]的研究相一致,即人工梭梭林的生态恢复效应在建植约20年时达到峰值,超过20年后则需要人工干预以增强其生态服务功能.在绿洲边缘区域,人工梭梭林表层土壤的有机碳、无机氮和全磷储量随梭梭林龄的延长而增长,而表层土壤全氮储量则先升后降.这可能与恢复后期表层土壤盐分的积累有关,表层土壤盐分积累抑制了土壤生物活动,进而降低了土壤全氮含量[15]. ...
荒漠生态系统C、N、P生态化学计量研究进展
1
2022
... 梭梭作为适应干旱环境的典型植物,因其耐旱、耐盐碱等特性,在荒漠化治理和生态恢复中具有重要的应用价值[16-19].人工梭梭建植通过修复退化生境、降低风蚀程度、提高动物多样性以及促进土壤碳氮等养分固存,进而显著提升多种生态系统服务功能[13,20-21].人工梭梭林在建植初期通常密度较高,随着建植时间延长,林分密度有所下降.生长至20年左右梭梭林开始出现不同程度衰退,其生态系统服务功能也随之减弱[13].然而,也有研究发现,在绿洲边缘地区20~30年梭梭林往往能够形成相对稳定的斑块状结构,并在30年左右出现幼苗密度回升,表现出一定的自然更新潜力[22-24].降水和地下水位变化以及不同立地条件影响人工梭梭林的重建、演变过程,同时也对土壤恢复过程产生重要影响[22,25-27].在人工梭梭林建植过程中,土壤恢复进程相对缓慢,而土壤质量提升对人工林地生态系统长期健康与稳定具有重要意义[28-29].土壤碳、氮、磷是土壤养分的重要组成部分,其比值(C∶N∶P)不仅反映土壤养分相对平衡状态,还能揭示生态系统养分限制特征与功能特性[30].此外,表层土壤碳、氮和磷储量也是衡量土壤功能变化的重要指标.近年来,关于人工梭梭林建植过程中土壤碳、氮、磷含量、比值及储量变化的研究逐渐增多,但不同的降水条件、地下水位及土壤环境背景均会显著影响其响应模式和变化规律[31-33].在人工梭梭林建植过程中,植被、土壤动物等生物要素与土壤结构、盐分等非生物要素之间相互作用及其对表层土壤碳、氮、磷含量和储量变化的耦合影响认识较为有限.鉴于此,本文以张掖绿洲北部荒漠绿洲过渡带的流动沙丘、固定沙丘以及5、10、20、30年梭梭林为研究对象,分析人工梭梭林重建、演变过程中表层土壤有机碳、全氮和全磷比值及土壤有机碳、全氮、无机氮和全磷储量的变化规律,确定生物和非生物因素对表层土壤碳、氮、磷比值及储量变化的影响和相对贡献率,为绿洲边缘人工梭梭林土壤质量演变及生态系统服务功能变化研究提供科学依据. ...
Interactive effects of vegetation and soil determine the composition and diversity of carabid and tenebrionid functional groups in an arid ecosystem
1
2016
... 研究区位于河西走廊中段,张掖绿洲北部的荒漠绿洲过渡带(39°21′53″—39°22′01″N,100°09′12″—100°09′14″E),该风沙带是巴丹吉林沙漠和张掖-临泽绿洲的交会处,海拔1 370 m.该区属温带大陆性荒漠气候,年平均气温为7.6 ℃,最低气温-10.7 ℃(1月),最高气温23.0 ℃(7月).多年平均降水量为117.0 mm,其中6、7、8、9月的降水量分别占全年降水量的13.6%、28.4%、16.7%、20.0%.年平均潜在蒸散量为2 390 mm,年平均日照时数达3 045 h[34].土壤类型为风沙土,地下水位4~5 m.天然木本植物有沙拐枣(Calligonum mongolicum)、泡泡刺(Nitraria sphaerocarpa)和小果白刺(Nitraria sibirica),草本植物主要有沙蓬(Agriophyllum pungens)、沙蒿(Artemisia desertorum)、白茎盐生草(Halogeton arachnoideus)、雾冰藜(Bassia dasyphylla)、沙鞭(Psammochloa villosa)和芦苇(Phragmites australis)等.人工栽植的灌木种有柽柳、柠条(Caragana korshinskii)、花棒(Corethrodendron scoparium)和梭梭等,梭梭是广泛使用的固沙灌木种.荒漠脊椎动物主要由爬行类、鸟类和兽类组成,荒漠沙蜥(Phrynocephalus przewalskii)、沙䳭(Oenanthe isabellina)、漠䳭(Oenanthe deserti)、楔尾伯劳(Lanius sphenocercus)、喜鹊(Pica pica)、鹅喉羚(Gazella subgutturosa)、虎鼬(Vormela peregusna)、赤狐(Vulpes vulpes)、亚洲狗獾(Meles leucurus)、兔狲(Otocolobus manul)和亚洲野猫(Felis silvestris)等是常见脊椎动物种,这些动物通过取食梭梭、草本和节肢动物以及挖掘土壤等直接和间接影响天然和人工固沙植被的健康与稳定[21]. ...
Changes in tenebrionid beetle and ant assembly influenced by different-aged Haloxylon ammodendron plantations in Northwest China
8
2025
... 为应对风沙侵蚀,张掖绿洲北部自20世纪70年代逐步开展人工固沙植被建植,形成从荒漠向绿洲过渡的人工梭梭林分布格局[1].基于该区域人工梭梭林随栽植年限形成的重建-演变梯度,选取林龄为5(Y5)、10(Y10)、20(Y20)、30(Y30)年林分作为取样区,并在人工梭梭林斑块边缘选择流动沙丘(MSD)和固定沙丘(FSD)作为对照取样区[35],以探讨不同建植阶段土壤养分特征及其影响因素.在每个取样区内分别选择4个地点,每个地点之间相距500~800 m.在每个地点内再分别选取3个相对独立的梭梭林斑块或沙丘作为植被和土壤样品的采样区,每个采样区间隔至少100 m.2018年秋季即植物多样性最高的季节调查灌木和草本群落组成,采集土壤样品测定土壤理化性质.在每个采样区内设置1个10 m×10 m的灌木调查样方,调查样方内灌木物种组成、密度及盖度,对每株灌木进行株高及冠幅测定;并按对角线设置5个1 m×1 m的草本样方,调查样方内草本盖度和物种丰富度.在每个草本样方内利用环刀法测定表层土壤容重,并利用土钻采集5个0~20 cm的土壤样品混合成1个样品,最终将每个采样区内的5个土壤样品再混合成1个样品.样品带回实验室后风干测定土壤理化性质,其中土壤机械组成[粗砂(>0.25 mm)、细砂(0.05~0.25 mm)和黏粉粒(<0.05 mm)]采用干筛法测定,土壤pH值采用电位法测定,土壤全盐含量采用干烧法测定,全氮采用硫酸-催化剂消解法测定,全磷采用硫酸-高氯酸消解法测定,有机质采用重铬酸钾-浓硫酸外加热法测定. ...
... 天然荒漠转变为人工梭梭林不仅改变了区域动植物多样性,还对土壤理化性质产生影响,这些改变会随着梭梭林建植年限的增加而变化[10,21,35,42].本研究表明,天然流动沙丘与固定沙丘之间土壤碳、氮、磷化学计量特征存在明显差异;随着人工梭梭林的建植,表层土壤化学计量特征亦发生改变.相较于流动沙丘,固定沙丘中灌木和草本植物的盖度及多样性明显提高,进而引发土壤C∶N下降和N∶P升高.上述结果表明,植被盖度提升加速表层土壤中氮素积累,其速率超过有机碳和全磷积累速率.然而,An等[43]在沙化草地研究发现,随着草地土壤沙化程度加剧,土壤C∶P和N∶P均下降,而C∶N变化不显著,这与本文的研究结果略有不同.泡泡刺和沙拐枣等灌木在荒漠生态系统中具有典型的“生态系统工程师”作用.这类灌木不仅富集枯落物并改善土壤微环境,也为其他生物提供了关键生态位即聚集了大型及中小型节肢动物,从而促进凋落物分解及氮素释放[21,44-45].在荒漠生态系统中,拟步甲科和蚁科等杂食性节肢动物是主要的食物网组成部分,其丰富度和多样性在灌木存在的环境中显著增加.这些节肢动物不仅直接参与凋落物的分解,还通过其死亡残体输入,提升了固定沙丘表层土壤中碳、氮、磷等养分含量[35].此外,研究还发现,研究区流动沙丘和固定沙丘土壤N∶P均低于1,显著低于全国土壤平均水平,进一步表明该区荒漠土壤存在明显的氮素限制[29,46].这种氮素限制特征可能与该区域干旱少雨、土壤有机质积累缓慢及微生物活动受限有关,同时也反映出人工固沙植被建植过程中,氮素补给和循环仍是限制生态系统功能提升的重要因素.梭梭是中国西北干旱区广泛分布的乔木型灌木种,不同梭梭种类及其分布区域会影响土壤碳、氮、磷化学计量比值.在流动和固定沙丘向人工梭梭林转变的过程中,表层土壤C∶N和N∶P发生了显著变化,而C∶P变化相对较小.这一结果与宁虎森等[47]在新疆甘家湖地区梭梭林的研究结果相似,但与魏亚娟等[32]在乌兰布和沙漠西南缘荒漠区梭梭林中的研究结果相反.宁虎森等[47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
... [35].此外,研究还发现,研究区流动沙丘和固定沙丘土壤N∶P均低于1,显著低于全国土壤平均水平,进一步表明该区荒漠土壤存在明显的氮素限制[29,46].这种氮素限制特征可能与该区域干旱少雨、土壤有机质积累缓慢及微生物活动受限有关,同时也反映出人工固沙植被建植过程中,氮素补给和循环仍是限制生态系统功能提升的重要因素.梭梭是中国西北干旱区广泛分布的乔木型灌木种,不同梭梭种类及其分布区域会影响土壤碳、氮、磷化学计量比值.在流动和固定沙丘向人工梭梭林转变的过程中,表层土壤C∶N和N∶P发生了显著变化,而C∶P变化相对较小.这一结果与宁虎森等[47]在新疆甘家湖地区梭梭林的研究结果相似,但与魏亚娟等[32]在乌兰布和沙漠西南缘荒漠区梭梭林中的研究结果相反.宁虎森等[47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
... 在人工梭梭林建植过程中,生物因子及其与非生物因子的交互作用显著影响表层土壤碳、氮、磷化学计量特征,相比之下,非生物因子的单独影响则相对较弱.随着人工梭梭林的重建与演变,灌木和草本植被盖度增加不仅丰富了生态系统中食物资源的数量和质量,还改变了脊椎动物和节肢动物的栖息环境,进而影响了土壤食物网的结构和功能[35].Wang等 [35]的研究表明,随着人工梭梭林建植年限的延长,拟步甲科等天然荒漠节肢动物的数量和多样性逐渐下降,而具有土壤扰动功能的蚁科节肢动物数量和多样性则逐步增加.荒漠收获蚁等不仅有助于植物种子扩散,还通过挖掘土壤提高了土壤养分含量、降低了土壤电导率,从而有利于提升土壤生物多样性及其生态功能[45,48].本文研究还发现,在人工梭梭林建植过程中,土壤大型节肢动物对表层土壤碳、氮、磷比值的影响大于中型节肢动物,这与土壤水分及盐分的限制密切相关.尽管已有研究表明,在人工梭梭林重建和演变的过程中,细菌和线虫多样性及其功能会发生变化,但土壤水分有效性较低限制了这些土壤生物功能的发挥[49-50].此外,随着灌木盖度的增加及植物群落结构的改变,表层土壤黏粉粒含量增加、粗砂含量减少,进而影响土壤无机碳和物理结构.这些因素共同改变了表层土壤碳、氮、磷比值的变化规律及土壤发育过程[51-52]. ...
... [35]的研究表明,随着人工梭梭林建植年限的延长,拟步甲科等天然荒漠节肢动物的数量和多样性逐渐下降,而具有土壤扰动功能的蚁科节肢动物数量和多样性则逐步增加.荒漠收获蚁等不仅有助于植物种子扩散,还通过挖掘土壤提高了土壤养分含量、降低了土壤电导率,从而有利于提升土壤生物多样性及其生态功能[45,48].本文研究还发现,在人工梭梭林建植过程中,土壤大型节肢动物对表层土壤碳、氮、磷比值的影响大于中型节肢动物,这与土壤水分及盐分的限制密切相关.尽管已有研究表明,在人工梭梭林重建和演变的过程中,细菌和线虫多样性及其功能会发生变化,但土壤水分有效性较低限制了这些土壤生物功能的发挥[49-50].此外,随着灌木盖度的增加及植物群落结构的改变,表层土壤黏粉粒含量增加、粗砂含量减少,进而影响土壤无机碳和物理结构.这些因素共同改变了表层土壤碳、氮、磷比值的变化规律及土壤发育过程[51-52]. ...
... 天然和人工梭梭林表层土壤碳、氮、磷含量均高于流动沙地.木本植被的存在及盖度增加,有助于提升荒漠土壤的养分含量与储量,而表层土壤碳、氮、磷储量对植被建植较为敏感[51,53-54].本研究发现,固定沙丘土壤有机碳、全氮和全磷储量均高于流动沙丘,这一趋势与腾格里沙漠和乌兰布和沙漠等区域的研究结果相近[32,54-55].然而,流动和固定沙丘表层土壤的无机氮储量变化相对较小,这可能与该区域土壤水分有效性较低,从而抑制土壤生物的活动有关[56].在人工梭梭林恢复的前期阶段(<20年),表层土壤有机碳、全磷和无机氮储量变化不明显,唯有土壤全氮储量显著增加,这与天然荒漠转变为梭梭林后,节肢动物、线虫及微生物多样性提高有关[35,50,57].魏亚娟等[32]在乌兰布和沙漠的研究也发现,在3~11年人工梭梭林中,土壤有机碳储量变化不大,而在16年时则显著提升,这与本研究结果相近.值得注意的是,在人工梭梭林恢复早期,表层土壤全氮和全磷储量随建植年限的增长而增加,但这一趋势与魏亚娟等[32]的研究稍有不同.Ma等[58]在石羊河流域的研究显示,人工梭梭林土壤有机碳储量随梭梭林建植年限增加而提升,但仍显著低于流动沙丘.本研究进一步表明,在人工梭梭林恢复的后期阶段(>20年),表层土壤的有机碳、无机氮和全磷储量随着建植年限延长而显著增加,但全氮储量在30年林龄时出现显著下降.这一变化趋势与Huang等[13]的研究相一致,即人工梭梭林的生态恢复效应在建植约20年时达到峰值,超过20年后则需要人工干预以增强其生态服务功能.在绿洲边缘区域,人工梭梭林表层土壤的有机碳、无机氮和全磷储量随梭梭林龄的延长而增长,而表层土壤全氮储量则先升后降.这可能与恢复后期表层土壤盐分的积累有关,表层土壤盐分积累抑制了土壤生物活动,进而降低了土壤全氮含量[15]. ...
... 在人工梭梭林建植过程中,非生物因子及其与生物因子的交互作用对表层土壤碳、氮、磷储量的影响大于生物因子的作用,且这些因子对土壤C∶N∶P比值的影响机制存在明显差异.本研究发现,土壤粒径组成和水盐条件等非生物因子在调控养分积累方面发挥着关键作用,这一结论与张掖绿洲边缘及其他干旱区的相关研究结果一致[52,54,59].梭梭林重建与演变过程能够有效削弱风沙活动,减少表层土壤流失,从而增强有机碳和氮积累,尤其在0~5 cm表层土壤中表现显著[54].此外,土壤黏粉粒比例增加可能是提升碳氮固存效率的重要因素[52].进一步分析表明,土壤黏粉粒含量、pH和全盐含量是非生物因子中影响人工梭梭林表层土壤有机碳、全氮和全磷储量变化的主要驱动因子.较高的黏粉粒含量有助于土壤结构的稳定性及有机质的高效积累,而pH值和盐分的变化则通过影响微生物活性及养分转化过程,进而调控土壤养分积累.特别是全盐含量的增加,促进了人工梭梭林下土壤盐结皮的形成,这一过程不仅抑制土壤水分入渗,还会限制草本植物及梭梭幼苗的萌发与生长,同时对土壤生物的活动产生负面影响,最终导致土壤全氮储量及其积累速率下降[15,51].安芳娇等[57]研究指出,在人工梭梭林的建植过程中,土壤pH值、有机碳含量和电导率是驱动线虫群落变化的主要环境因子,其中土壤电导率是影响线虫群落变化的关键因素.此外,灌木盖度、草本盖度及大型节肢动物数量是影响人工梭梭林建植过程中土壤碳、氮、磷储量变化的主要生物因子.灌木盖度与土壤黏粉粒之间的相互作用能够抑制风蚀,促进土壤细颗粒的聚集,进而提高表层土壤碳、氮、磷储量.同时,土壤中蚂蚁、蜘蛛及其他中小型土壤动物种类和数量的增加,加速了凋落物的分解过程[35].然而,土壤盐分增加及土壤水分有效性降低可能抑制了螨类和跳虫等主要分解类群的数量和多样性,从而导致人工梭梭林建植后期土壤全氮储量的下降[35,60-61].在人工梭梭林建植约20年时,通过降低灌木密度和盖度,增加挖掘兽类和节肢动物的多样性,以及提升关键捕食性动物种群的数量,可以显著增强人工梭梭林的生态系统服务功能,并提高其对气候变化和人类干扰的适应能力[13,21,48,62].总体而言,植被盖度、土壤质地与生物群落之间的耦合变化是驱动表层土壤养分储量变化的核心机制.因此,为了提升人工梭梭林的生态系统功能并实现可持续管理,有必要加强对建植过程中土壤环境和生物群落协同演变的长期监测,为相关理论研究和实践应用提供更为坚实的数据支持. ...
... [35,60-61].在人工梭梭林建植约20年时,通过降低灌木密度和盖度,增加挖掘兽类和节肢动物的多样性,以及提升关键捕食性动物种群的数量,可以显著增强人工梭梭林的生态系统服务功能,并提高其对气候变化和人类干扰的适应能力[13,21,48,62].总体而言,植被盖度、土壤质地与生物群落之间的耦合变化是驱动表层土壤养分储量变化的核心机制.因此,为了提升人工梭梭林的生态系统功能并实现可持续管理,有必要加强对建植过程中土壤环境和生物群落协同演变的长期监测,为相关理论研究和实践应用提供更为坚实的数据支持. ...
基于陷阱法采集的河西走廊戈壁荒漠甲虫数据集
1
2024
... 2018年5月下旬和8月中旬利用陷阱法收集大型和中型节肢动物样品,每次样品收集时间为3天.每个植被样方内布设5个陷阱收集器,其中3个位于灌木植株下方,2个位于灌木植株之间的空地上.陷阱收集器使用塑料杯和PVC管制作(直径7 cm,高度10 cm),使用75%的酒精溶液和少量甘油作为保存液,逐日收集样品[36].此外,在MSD的迎风坡、坡顶和背风坡分别放置诱捕器,每个微生境放置5个陷阱收集器.收集的节肢动物样品在体式显微镜下参照相关分类书籍确定亚目、科或属信息[37-40].在目或科水平统计大型和中型节肢动物数量及类群丰富度,大型节肢动物主要包括蛛形纲(蜘蛛)、鞘翅目(甲虫)和蚁科(蚂蚁),中型节肢动物主要包括蜱螨目(螨类)和弹尾目(跳虫). ...
1
1998
... 2018年5月下旬和8月中旬利用陷阱法收集大型和中型节肢动物样品,每次样品收集时间为3天.每个植被样方内布设5个陷阱收集器,其中3个位于灌木植株下方,2个位于灌木植株之间的空地上.陷阱收集器使用塑料杯和PVC管制作(直径7 cm,高度10 cm),使用75%的酒精溶液和少量甘油作为保存液,逐日收集样品[36].此外,在MSD的迎风坡、坡顶和背风坡分别放置诱捕器,每个微生境放置5个陷阱收集器.收集的节肢动物样品在体式显微镜下参照相关分类书籍确定亚目、科或属信息[37-40].在目或科水平统计大型和中型节肢动物数量及类群丰富度,大型节肢动物主要包括蛛形纲(蜘蛛)、鞘翅目(甲虫)和蚁科(蚂蚁),中型节肢动物主要包括蜱螨目(螨类)和弹尾目(跳虫). ...
1
1999
... 2018年5月下旬和8月中旬利用陷阱法收集大型和中型节肢动物样品,每次样品收集时间为3天.每个植被样方内布设5个陷阱收集器,其中3个位于灌木植株下方,2个位于灌木植株之间的空地上.陷阱收集器使用塑料杯和PVC管制作(直径7 cm,高度10 cm),使用75%的酒精溶液和少量甘油作为保存液,逐日收集样品[36].此外,在MSD的迎风坡、坡顶和背风坡分别放置诱捕器,每个微生境放置5个陷阱收集器.收集的节肢动物样品在体式显微镜下参照相关分类书籍确定亚目、科或属信息[37-40].在目或科水平统计大型和中型节肢动物数量及类群丰富度,大型节肢动物主要包括蛛形纲(蜘蛛)、鞘翅目(甲虫)和蚁科(蚂蚁),中型节肢动物主要包括蜱螨目(螨类)和弹尾目(跳虫). ...
Canoco Reference Manual and User's Guide: Software for Ordination
1
2012
... 选取灌木盖度、灌木密度、草本盖度、草本物种丰富度、大型和中型节肢动物数量与类群丰富度作为生物要素,土壤容重、土壤粗砂、细砂和黏粉粒含量、pH和全盐含量作为非生物要素.利用相关分析和主成分分析等确定灌木盖度、草本盖度、大型节肢动物数量、中型节肢动物数量、土壤黏粉粒含量、pH和全盐含量7个因子作为表层土壤碳氮磷化学计量特征及其储量变化的主要影响因子.使用冗余分析(RDA)排序分析7个生物和非生物因子对表层土壤碳氮磷化学计量特征及其储量变化的影响,利用蒙特卡洛检验(Monte-Carlo permutation test 999)第1排序轴和所有排序轴的显著性,检验结果显著表明排序结果较好反映环境要素和表层土壤碳氮磷化学计量特征及其储量变化关系.RDA分析过程中,利用变差分解(Variation Partitioning Analysis)确定生物因子和非生物因子及二者交互对表层土壤碳氮磷化学计量特征及其储量变化的影响及贡献率;pRDA偏相关分析中逐步向前选择(Interactive-forward-selection)确定7个生物、非生物因子对土壤有机碳、全氮和全磷储量变化的影响[41].排序分析过程中所有数据进行对数转化,统计分析使用CANOCO 5.0软件. ...
不同地下水位处梭梭(Haloxylon ammodendron)水分来源特征
1
2021
... 天然荒漠转变为人工梭梭林不仅改变了区域动植物多样性,还对土壤理化性质产生影响,这些改变会随着梭梭林建植年限的增加而变化[10,21,35,42].本研究表明,天然流动沙丘与固定沙丘之间土壤碳、氮、磷化学计量特征存在明显差异;随着人工梭梭林的建植,表层土壤化学计量特征亦发生改变.相较于流动沙丘,固定沙丘中灌木和草本植物的盖度及多样性明显提高,进而引发土壤C∶N下降和N∶P升高.上述结果表明,植被盖度提升加速表层土壤中氮素积累,其速率超过有机碳和全磷积累速率.然而,An等[43]在沙化草地研究发现,随着草地土壤沙化程度加剧,土壤C∶P和N∶P均下降,而C∶N变化不显著,这与本文的研究结果略有不同.泡泡刺和沙拐枣等灌木在荒漠生态系统中具有典型的“生态系统工程师”作用.这类灌木不仅富集枯落物并改善土壤微环境,也为其他生物提供了关键生态位即聚集了大型及中小型节肢动物,从而促进凋落物分解及氮素释放[21,44-45].在荒漠生态系统中,拟步甲科和蚁科等杂食性节肢动物是主要的食物网组成部分,其丰富度和多样性在灌木存在的环境中显著增加.这些节肢动物不仅直接参与凋落物的分解,还通过其死亡残体输入,提升了固定沙丘表层土壤中碳、氮、磷等养分含量[35].此外,研究还发现,研究区流动沙丘和固定沙丘土壤N∶P均低于1,显著低于全国土壤平均水平,进一步表明该区荒漠土壤存在明显的氮素限制[29,46].这种氮素限制特征可能与该区域干旱少雨、土壤有机质积累缓慢及微生物活动受限有关,同时也反映出人工固沙植被建植过程中,氮素补给和循环仍是限制生态系统功能提升的重要因素.梭梭是中国西北干旱区广泛分布的乔木型灌木种,不同梭梭种类及其分布区域会影响土壤碳、氮、磷化学计量比值.在流动和固定沙丘向人工梭梭林转变的过程中,表层土壤C∶N和N∶P发生了显著变化,而C∶P变化相对较小.这一结果与宁虎森等[47]在新疆甘家湖地区梭梭林的研究结果相似,但与魏亚娟等[32]在乌兰布和沙漠西南缘荒漠区梭梭林中的研究结果相反.宁虎森等[47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
Impact of desertification on soil and plant nutrient stoichiometry in a desert grassland
1
2019
... 天然荒漠转变为人工梭梭林不仅改变了区域动植物多样性,还对土壤理化性质产生影响,这些改变会随着梭梭林建植年限的增加而变化[10,21,35,42].本研究表明,天然流动沙丘与固定沙丘之间土壤碳、氮、磷化学计量特征存在明显差异;随着人工梭梭林的建植,表层土壤化学计量特征亦发生改变.相较于流动沙丘,固定沙丘中灌木和草本植物的盖度及多样性明显提高,进而引发土壤C∶N下降和N∶P升高.上述结果表明,植被盖度提升加速表层土壤中氮素积累,其速率超过有机碳和全磷积累速率.然而,An等[43]在沙化草地研究发现,随着草地土壤沙化程度加剧,土壤C∶P和N∶P均下降,而C∶N变化不显著,这与本文的研究结果略有不同.泡泡刺和沙拐枣等灌木在荒漠生态系统中具有典型的“生态系统工程师”作用.这类灌木不仅富集枯落物并改善土壤微环境,也为其他生物提供了关键生态位即聚集了大型及中小型节肢动物,从而促进凋落物分解及氮素释放[21,44-45].在荒漠生态系统中,拟步甲科和蚁科等杂食性节肢动物是主要的食物网组成部分,其丰富度和多样性在灌木存在的环境中显著增加.这些节肢动物不仅直接参与凋落物的分解,还通过其死亡残体输入,提升了固定沙丘表层土壤中碳、氮、磷等养分含量[35].此外,研究还发现,研究区流动沙丘和固定沙丘土壤N∶P均低于1,显著低于全国土壤平均水平,进一步表明该区荒漠土壤存在明显的氮素限制[29,46].这种氮素限制特征可能与该区域干旱少雨、土壤有机质积累缓慢及微生物活动受限有关,同时也反映出人工固沙植被建植过程中,氮素补给和循环仍是限制生态系统功能提升的重要因素.梭梭是中国西北干旱区广泛分布的乔木型灌木种,不同梭梭种类及其分布区域会影响土壤碳、氮、磷化学计量比值.在流动和固定沙丘向人工梭梭林转变的过程中,表层土壤C∶N和N∶P发生了显著变化,而C∶P变化相对较小.这一结果与宁虎森等[47]在新疆甘家湖地区梭梭林的研究结果相似,但与魏亚娟等[32]在乌兰布和沙漠西南缘荒漠区梭梭林中的研究结果相反.宁虎森等[47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
Effects of shrub presence and shrub species on ground beetle assemblages (Carabidae,Curculionidae and Tenebrionidae) in a sandy desert,Northwestern China
1
2015
... 天然荒漠转变为人工梭梭林不仅改变了区域动植物多样性,还对土壤理化性质产生影响,这些改变会随着梭梭林建植年限的增加而变化[10,21,35,42].本研究表明,天然流动沙丘与固定沙丘之间土壤碳、氮、磷化学计量特征存在明显差异;随着人工梭梭林的建植,表层土壤化学计量特征亦发生改变.相较于流动沙丘,固定沙丘中灌木和草本植物的盖度及多样性明显提高,进而引发土壤C∶N下降和N∶P升高.上述结果表明,植被盖度提升加速表层土壤中氮素积累,其速率超过有机碳和全磷积累速率.然而,An等[43]在沙化草地研究发现,随着草地土壤沙化程度加剧,土壤C∶P和N∶P均下降,而C∶N变化不显著,这与本文的研究结果略有不同.泡泡刺和沙拐枣等灌木在荒漠生态系统中具有典型的“生态系统工程师”作用.这类灌木不仅富集枯落物并改善土壤微环境,也为其他生物提供了关键生态位即聚集了大型及中小型节肢动物,从而促进凋落物分解及氮素释放[21,44-45].在荒漠生态系统中,拟步甲科和蚁科等杂食性节肢动物是主要的食物网组成部分,其丰富度和多样性在灌木存在的环境中显著增加.这些节肢动物不仅直接参与凋落物的分解,还通过其死亡残体输入,提升了固定沙丘表层土壤中碳、氮、磷等养分含量[35].此外,研究还发现,研究区流动沙丘和固定沙丘土壤N∶P均低于1,显著低于全国土壤平均水平,进一步表明该区荒漠土壤存在明显的氮素限制[29,46].这种氮素限制特征可能与该区域干旱少雨、土壤有机质积累缓慢及微生物活动受限有关,同时也反映出人工固沙植被建植过程中,氮素补给和循环仍是限制生态系统功能提升的重要因素.梭梭是中国西北干旱区广泛分布的乔木型灌木种,不同梭梭种类及其分布区域会影响土壤碳、氮、磷化学计量比值.在流动和固定沙丘向人工梭梭林转变的过程中,表层土壤C∶N和N∶P发生了显著变化,而C∶P变化相对较小.这一结果与宁虎森等[47]在新疆甘家湖地区梭梭林的研究结果相似,但与魏亚娟等[32]在乌兰布和沙漠西南缘荒漠区梭梭林中的研究结果相反.宁虎森等[47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
Ground arthropod assemblages shaped by ant nests and shrub microhabitats in Gobi Desert ecosystems
2
2025
... 天然荒漠转变为人工梭梭林不仅改变了区域动植物多样性,还对土壤理化性质产生影响,这些改变会随着梭梭林建植年限的增加而变化[10,21,35,42].本研究表明,天然流动沙丘与固定沙丘之间土壤碳、氮、磷化学计量特征存在明显差异;随着人工梭梭林的建植,表层土壤化学计量特征亦发生改变.相较于流动沙丘,固定沙丘中灌木和草本植物的盖度及多样性明显提高,进而引发土壤C∶N下降和N∶P升高.上述结果表明,植被盖度提升加速表层土壤中氮素积累,其速率超过有机碳和全磷积累速率.然而,An等[43]在沙化草地研究发现,随着草地土壤沙化程度加剧,土壤C∶P和N∶P均下降,而C∶N变化不显著,这与本文的研究结果略有不同.泡泡刺和沙拐枣等灌木在荒漠生态系统中具有典型的“生态系统工程师”作用.这类灌木不仅富集枯落物并改善土壤微环境,也为其他生物提供了关键生态位即聚集了大型及中小型节肢动物,从而促进凋落物分解及氮素释放[21,44-45].在荒漠生态系统中,拟步甲科和蚁科等杂食性节肢动物是主要的食物网组成部分,其丰富度和多样性在灌木存在的环境中显著增加.这些节肢动物不仅直接参与凋落物的分解,还通过其死亡残体输入,提升了固定沙丘表层土壤中碳、氮、磷等养分含量[35].此外,研究还发现,研究区流动沙丘和固定沙丘土壤N∶P均低于1,显著低于全国土壤平均水平,进一步表明该区荒漠土壤存在明显的氮素限制[29,46].这种氮素限制特征可能与该区域干旱少雨、土壤有机质积累缓慢及微生物活动受限有关,同时也反映出人工固沙植被建植过程中,氮素补给和循环仍是限制生态系统功能提升的重要因素.梭梭是中国西北干旱区广泛分布的乔木型灌木种,不同梭梭种类及其分布区域会影响土壤碳、氮、磷化学计量比值.在流动和固定沙丘向人工梭梭林转变的过程中,表层土壤C∶N和N∶P发生了显著变化,而C∶P变化相对较小.这一结果与宁虎森等[47]在新疆甘家湖地区梭梭林的研究结果相似,但与魏亚娟等[32]在乌兰布和沙漠西南缘荒漠区梭梭林中的研究结果相反.宁虎森等[47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
... 在人工梭梭林建植过程中,生物因子及其与非生物因子的交互作用显著影响表层土壤碳、氮、磷化学计量特征,相比之下,非生物因子的单独影响则相对较弱.随着人工梭梭林的重建与演变,灌木和草本植被盖度增加不仅丰富了生态系统中食物资源的数量和质量,还改变了脊椎动物和节肢动物的栖息环境,进而影响了土壤食物网的结构和功能[35].Wang等 [35]的研究表明,随着人工梭梭林建植年限的延长,拟步甲科等天然荒漠节肢动物的数量和多样性逐渐下降,而具有土壤扰动功能的蚁科节肢动物数量和多样性则逐步增加.荒漠收获蚁等不仅有助于植物种子扩散,还通过挖掘土壤提高了土壤养分含量、降低了土壤电导率,从而有利于提升土壤生物多样性及其生态功能[45,48].本文研究还发现,在人工梭梭林建植过程中,土壤大型节肢动物对表层土壤碳、氮、磷比值的影响大于中型节肢动物,这与土壤水分及盐分的限制密切相关.尽管已有研究表明,在人工梭梭林重建和演变的过程中,细菌和线虫多样性及其功能会发生变化,但土壤水分有效性较低限制了这些土壤生物功能的发挥[49-50].此外,随着灌木盖度的增加及植物群落结构的改变,表层土壤黏粉粒含量增加、粗砂含量减少,进而影响土壤无机碳和物理结构.这些因素共同改变了表层土壤碳、氮、磷比值的变化规律及土壤发育过程[51-52]. ...
C∶N∶P stoichiometry in Australian soils with respect to vegetation and environmental factors
1
2013
... 天然荒漠转变为人工梭梭林不仅改变了区域动植物多样性,还对土壤理化性质产生影响,这些改变会随着梭梭林建植年限的增加而变化[10,21,35,42].本研究表明,天然流动沙丘与固定沙丘之间土壤碳、氮、磷化学计量特征存在明显差异;随着人工梭梭林的建植,表层土壤化学计量特征亦发生改变.相较于流动沙丘,固定沙丘中灌木和草本植物的盖度及多样性明显提高,进而引发土壤C∶N下降和N∶P升高.上述结果表明,植被盖度提升加速表层土壤中氮素积累,其速率超过有机碳和全磷积累速率.然而,An等[43]在沙化草地研究发现,随着草地土壤沙化程度加剧,土壤C∶P和N∶P均下降,而C∶N变化不显著,这与本文的研究结果略有不同.泡泡刺和沙拐枣等灌木在荒漠生态系统中具有典型的“生态系统工程师”作用.这类灌木不仅富集枯落物并改善土壤微环境,也为其他生物提供了关键生态位即聚集了大型及中小型节肢动物,从而促进凋落物分解及氮素释放[21,44-45].在荒漠生态系统中,拟步甲科和蚁科等杂食性节肢动物是主要的食物网组成部分,其丰富度和多样性在灌木存在的环境中显著增加.这些节肢动物不仅直接参与凋落物的分解,还通过其死亡残体输入,提升了固定沙丘表层土壤中碳、氮、磷等养分含量[35].此外,研究还发现,研究区流动沙丘和固定沙丘土壤N∶P均低于1,显著低于全国土壤平均水平,进一步表明该区荒漠土壤存在明显的氮素限制[29,46].这种氮素限制特征可能与该区域干旱少雨、土壤有机质积累缓慢及微生物活动受限有关,同时也反映出人工固沙植被建植过程中,氮素补给和循环仍是限制生态系统功能提升的重要因素.梭梭是中国西北干旱区广泛分布的乔木型灌木种,不同梭梭种类及其分布区域会影响土壤碳、氮、磷化学计量比值.在流动和固定沙丘向人工梭梭林转变的过程中,表层土壤C∶N和N∶P发生了显著变化,而C∶P变化相对较小.这一结果与宁虎森等[47]在新疆甘家湖地区梭梭林的研究结果相似,但与魏亚娟等[32]在乌兰布和沙漠西南缘荒漠区梭梭林中的研究结果相反.宁虎森等[47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
新疆甘家湖梭梭林碳、氮、磷、钾生态化学计量特征
2
2017
... 天然荒漠转变为人工梭梭林不仅改变了区域动植物多样性,还对土壤理化性质产生影响,这些改变会随着梭梭林建植年限的增加而变化[10,21,35,42].本研究表明,天然流动沙丘与固定沙丘之间土壤碳、氮、磷化学计量特征存在明显差异;随着人工梭梭林的建植,表层土壤化学计量特征亦发生改变.相较于流动沙丘,固定沙丘中灌木和草本植物的盖度及多样性明显提高,进而引发土壤C∶N下降和N∶P升高.上述结果表明,植被盖度提升加速表层土壤中氮素积累,其速率超过有机碳和全磷积累速率.然而,An等[43]在沙化草地研究发现,随着草地土壤沙化程度加剧,土壤C∶P和N∶P均下降,而C∶N变化不显著,这与本文的研究结果略有不同.泡泡刺和沙拐枣等灌木在荒漠生态系统中具有典型的“生态系统工程师”作用.这类灌木不仅富集枯落物并改善土壤微环境,也为其他生物提供了关键生态位即聚集了大型及中小型节肢动物,从而促进凋落物分解及氮素释放[21,44-45].在荒漠生态系统中,拟步甲科和蚁科等杂食性节肢动物是主要的食物网组成部分,其丰富度和多样性在灌木存在的环境中显著增加.这些节肢动物不仅直接参与凋落物的分解,还通过其死亡残体输入,提升了固定沙丘表层土壤中碳、氮、磷等养分含量[35].此外,研究还发现,研究区流动沙丘和固定沙丘土壤N∶P均低于1,显著低于全国土壤平均水平,进一步表明该区荒漠土壤存在明显的氮素限制[29,46].这种氮素限制特征可能与该区域干旱少雨、土壤有机质积累缓慢及微生物活动受限有关,同时也反映出人工固沙植被建植过程中,氮素补给和循环仍是限制生态系统功能提升的重要因素.梭梭是中国西北干旱区广泛分布的乔木型灌木种,不同梭梭种类及其分布区域会影响土壤碳、氮、磷化学计量比值.在流动和固定沙丘向人工梭梭林转变的过程中,表层土壤C∶N和N∶P发生了显著变化,而C∶P变化相对较小.这一结果与宁虎森等[47]在新疆甘家湖地区梭梭林的研究结果相似,但与魏亚娟等[32]在乌兰布和沙漠西南缘荒漠区梭梭林中的研究结果相反.宁虎森等[47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
... [47]还发现,梭梭林盖度增加会显著提高表层土壤的N∶P,降低C∶P,而对C∶N影响不明显.本研究还发现,随着固定沙丘中灌木盖度的增加,表层土壤C∶N显著下降,而N∶P上升;人工梭梭林建植使得盖度增加,进一步加剧了这一变化趋势.然而,在人工梭梭林恢复的后期阶段,灌木盖度出现下降趋势,与之相应,表层土壤C∶N略有回升,而N∶P则呈下降态势.魏亚娟等[32]在对乌兰布和沙漠西南缘荒漠区不同建植年限(3年、6年、11年和16年)梭梭林研究发现,随着梭梭栽植年限的增加,土壤C∶N和C∶P逐步上升,而土壤N∶P变化较小,这与本文的研究结果存在差异.本文研究区位于绿洲边缘,受绿洲灌溉的影响,区域地下水位维持在较浅的深度(约4~5 m),为人工梭梭林的生长提供了相对稳定的水分条件,从而减缓了植被退化进程.研究表明,该区域人工梭梭林在建植约20年后可形成相对稳定的自组织结构,在建植30年以上林地中,梭梭幼苗数量明显增加,表明该区域的梭梭林具备一定的自然更新能力[22,24].张珂等[31]在张掖绿洲边缘的研究中发现,梭梭林栽植9年后土壤C∶P和N∶P表现出明显的成层化分布,且随着梭梭林龄增加,不同土层C∶P和N∶P均有所增大,这与本文研究结果相近.然而,土壤C∶N随着梭梭林龄增加保持相对稳定[31],这与本文的发现有所不同.此外,本文研究还发现,10年梭梭林的土壤C∶P最低,20年梭梭林的土壤C∶N最低、N∶P最高,而30年梭梭林的C∶N上升,则抑制了土壤生物的分解活动.人工梭梭林的生态效应在栽植约20年后达到最大值,之后开始下降,表明此时需要通过优化管理措施来提升梭梭林的生态系统服务功能[13]. ...
河西走廊中部荒漠收获蚁(Messor desertus)蚁穴对秋季地表节肢动物群落结构的影响
2
2023
... 在人工梭梭林建植过程中,生物因子及其与非生物因子的交互作用显著影响表层土壤碳、氮、磷化学计量特征,相比之下,非生物因子的单独影响则相对较弱.随着人工梭梭林的重建与演变,灌木和草本植被盖度增加不仅丰富了生态系统中食物资源的数量和质量,还改变了脊椎动物和节肢动物的栖息环境,进而影响了土壤食物网的结构和功能[35].Wang等 [35]的研究表明,随着人工梭梭林建植年限的延长,拟步甲科等天然荒漠节肢动物的数量和多样性逐渐下降,而具有土壤扰动功能的蚁科节肢动物数量和多样性则逐步增加.荒漠收获蚁等不仅有助于植物种子扩散,还通过挖掘土壤提高了土壤养分含量、降低了土壤电导率,从而有利于提升土壤生物多样性及其生态功能[45,48].本文研究还发现,在人工梭梭林建植过程中,土壤大型节肢动物对表层土壤碳、氮、磷比值的影响大于中型节肢动物,这与土壤水分及盐分的限制密切相关.尽管已有研究表明,在人工梭梭林重建和演变的过程中,细菌和线虫多样性及其功能会发生变化,但土壤水分有效性较低限制了这些土壤生物功能的发挥[49-50].此外,随着灌木盖度的增加及植物群落结构的改变,表层土壤黏粉粒含量增加、粗砂含量减少,进而影响土壤无机碳和物理结构.这些因素共同改变了表层土壤碳、氮、磷比值的变化规律及土壤发育过程[51-52]. ...
... 在人工梭梭林建植过程中,非生物因子及其与生物因子的交互作用对表层土壤碳、氮、磷储量的影响大于生物因子的作用,且这些因子对土壤C∶N∶P比值的影响机制存在明显差异.本研究发现,土壤粒径组成和水盐条件等非生物因子在调控养分积累方面发挥着关键作用,这一结论与张掖绿洲边缘及其他干旱区的相关研究结果一致[52,54,59].梭梭林重建与演变过程能够有效削弱风沙活动,减少表层土壤流失,从而增强有机碳和氮积累,尤其在0~5 cm表层土壤中表现显著[54].此外,土壤黏粉粒比例增加可能是提升碳氮固存效率的重要因素[52].进一步分析表明,土壤黏粉粒含量、pH和全盐含量是非生物因子中影响人工梭梭林表层土壤有机碳、全氮和全磷储量变化的主要驱动因子.较高的黏粉粒含量有助于土壤结构的稳定性及有机质的高效积累,而pH值和盐分的变化则通过影响微生物活性及养分转化过程,进而调控土壤养分积累.特别是全盐含量的增加,促进了人工梭梭林下土壤盐结皮的形成,这一过程不仅抑制土壤水分入渗,还会限制草本植物及梭梭幼苗的萌发与生长,同时对土壤生物的活动产生负面影响,最终导致土壤全氮储量及其积累速率下降[15,51].安芳娇等[57]研究指出,在人工梭梭林的建植过程中,土壤pH值、有机碳含量和电导率是驱动线虫群落变化的主要环境因子,其中土壤电导率是影响线虫群落变化的关键因素.此外,灌木盖度、草本盖度及大型节肢动物数量是影响人工梭梭林建植过程中土壤碳、氮、磷储量变化的主要生物因子.灌木盖度与土壤黏粉粒之间的相互作用能够抑制风蚀,促进土壤细颗粒的聚集,进而提高表层土壤碳、氮、磷储量.同时,土壤中蚂蚁、蜘蛛及其他中小型土壤动物种类和数量的增加,加速了凋落物的分解过程[35].然而,土壤盐分增加及土壤水分有效性降低可能抑制了螨类和跳虫等主要分解类群的数量和多样性,从而导致人工梭梭林建植后期土壤全氮储量的下降[35,60-61].在人工梭梭林建植约20年时,通过降低灌木密度和盖度,增加挖掘兽类和节肢动物的多样性,以及提升关键捕食性动物种群的数量,可以显著增强人工梭梭林的生态系统服务功能,并提高其对气候变化和人类干扰的适应能力[13,21,48,62].总体而言,植被盖度、土壤质地与生物群落之间的耦合变化是驱动表层土壤养分储量变化的核心机制.因此,为了提升人工梭梭林的生态系统功能并实现可持续管理,有必要加强对建植过程中土壤环境和生物群落协同演变的长期监测,为相关理论研究和实践应用提供更为坚实的数据支持. ...
准噶尔荒漠区域尺度浅层土壤化学计量特征及其空间分布格局
1
2016
... 在人工梭梭林建植过程中,生物因子及其与非生物因子的交互作用显著影响表层土壤碳、氮、磷化学计量特征,相比之下,非生物因子的单独影响则相对较弱.随着人工梭梭林的重建与演变,灌木和草本植被盖度增加不仅丰富了生态系统中食物资源的数量和质量,还改变了脊椎动物和节肢动物的栖息环境,进而影响了土壤食物网的结构和功能[35].Wang等 [35]的研究表明,随着人工梭梭林建植年限的延长,拟步甲科等天然荒漠节肢动物的数量和多样性逐渐下降,而具有土壤扰动功能的蚁科节肢动物数量和多样性则逐步增加.荒漠收获蚁等不仅有助于植物种子扩散,还通过挖掘土壤提高了土壤养分含量、降低了土壤电导率,从而有利于提升土壤生物多样性及其生态功能[45,48].本文研究还发现,在人工梭梭林建植过程中,土壤大型节肢动物对表层土壤碳、氮、磷比值的影响大于中型节肢动物,这与土壤水分及盐分的限制密切相关.尽管已有研究表明,在人工梭梭林重建和演变的过程中,细菌和线虫多样性及其功能会发生变化,但土壤水分有效性较低限制了这些土壤生物功能的发挥[49-50].此外,随着灌木盖度的增加及植物群落结构的改变,表层土壤黏粉粒含量增加、粗砂含量减少,进而影响土壤无机碳和物理结构.这些因素共同改变了表层土壤碳、氮、磷比值的变化规律及土壤发育过程[51-52]. ...
Succession of soil bacterial community along a 46-year choronsequence artificial revegetation in an arid oasis-desert ecotone
2
2022
... 在人工梭梭林建植过程中,生物因子及其与非生物因子的交互作用显著影响表层土壤碳、氮、磷化学计量特征,相比之下,非生物因子的单独影响则相对较弱.随着人工梭梭林的重建与演变,灌木和草本植被盖度增加不仅丰富了生态系统中食物资源的数量和质量,还改变了脊椎动物和节肢动物的栖息环境,进而影响了土壤食物网的结构和功能[35].Wang等 [35]的研究表明,随着人工梭梭林建植年限的延长,拟步甲科等天然荒漠节肢动物的数量和多样性逐渐下降,而具有土壤扰动功能的蚁科节肢动物数量和多样性则逐步增加.荒漠收获蚁等不仅有助于植物种子扩散,还通过挖掘土壤提高了土壤养分含量、降低了土壤电导率,从而有利于提升土壤生物多样性及其生态功能[45,48].本文研究还发现,在人工梭梭林建植过程中,土壤大型节肢动物对表层土壤碳、氮、磷比值的影响大于中型节肢动物,这与土壤水分及盐分的限制密切相关.尽管已有研究表明,在人工梭梭林重建和演变的过程中,细菌和线虫多样性及其功能会发生变化,但土壤水分有效性较低限制了这些土壤生物功能的发挥[49-50].此外,随着灌木盖度的增加及植物群落结构的改变,表层土壤黏粉粒含量增加、粗砂含量减少,进而影响土壤无机碳和物理结构.这些因素共同改变了表层土壤碳、氮、磷比值的变化规律及土壤发育过程[51-52]. ...
... 天然和人工梭梭林表层土壤碳、氮、磷含量均高于流动沙地.木本植被的存在及盖度增加,有助于提升荒漠土壤的养分含量与储量,而表层土壤碳、氮、磷储量对植被建植较为敏感[51,53-54].本研究发现,固定沙丘土壤有机碳、全氮和全磷储量均高于流动沙丘,这一趋势与腾格里沙漠和乌兰布和沙漠等区域的研究结果相近[32,54-55].然而,流动和固定沙丘表层土壤的无机氮储量变化相对较小,这可能与该区域土壤水分有效性较低,从而抑制土壤生物的活动有关[56].在人工梭梭林恢复的前期阶段(<20年),表层土壤有机碳、全磷和无机氮储量变化不明显,唯有土壤全氮储量显著增加,这与天然荒漠转变为梭梭林后,节肢动物、线虫及微生物多样性提高有关[35,50,57].魏亚娟等[32]在乌兰布和沙漠的研究也发现,在3~11年人工梭梭林中,土壤有机碳储量变化不大,而在16年时则显著提升,这与本研究结果相近.值得注意的是,在人工梭梭林恢复早期,表层土壤全氮和全磷储量随建植年限的增长而增加,但这一趋势与魏亚娟等[32]的研究稍有不同.Ma等[58]在石羊河流域的研究显示,人工梭梭林土壤有机碳储量随梭梭林建植年限增加而提升,但仍显著低于流动沙丘.本研究进一步表明,在人工梭梭林恢复的后期阶段(>20年),表层土壤的有机碳、无机氮和全磷储量随着建植年限延长而显著增加,但全氮储量在30年林龄时出现显著下降.这一变化趋势与Huang等[13]的研究相一致,即人工梭梭林的生态恢复效应在建植约20年时达到峰值,超过20年后则需要人工干预以增强其生态服务功能.在绿洲边缘区域,人工梭梭林表层土壤的有机碳、无机氮和全磷储量随梭梭林龄的延长而增长,而表层土壤全氮储量则先升后降.这可能与恢复后期表层土壤盐分的积累有关,表层土壤盐分积累抑制了土壤生物活动,进而降低了土壤全氮含量[15]. ...
流动沙地建植人工固沙梭梭林的土壤演变过程
3
2020
... 在人工梭梭林建植过程中,生物因子及其与非生物因子的交互作用显著影响表层土壤碳、氮、磷化学计量特征,相比之下,非生物因子的单独影响则相对较弱.随着人工梭梭林的重建与演变,灌木和草本植被盖度增加不仅丰富了生态系统中食物资源的数量和质量,还改变了脊椎动物和节肢动物的栖息环境,进而影响了土壤食物网的结构和功能[35].Wang等 [35]的研究表明,随着人工梭梭林建植年限的延长,拟步甲科等天然荒漠节肢动物的数量和多样性逐渐下降,而具有土壤扰动功能的蚁科节肢动物数量和多样性则逐步增加.荒漠收获蚁等不仅有助于植物种子扩散,还通过挖掘土壤提高了土壤养分含量、降低了土壤电导率,从而有利于提升土壤生物多样性及其生态功能[45,48].本文研究还发现,在人工梭梭林建植过程中,土壤大型节肢动物对表层土壤碳、氮、磷比值的影响大于中型节肢动物,这与土壤水分及盐分的限制密切相关.尽管已有研究表明,在人工梭梭林重建和演变的过程中,细菌和线虫多样性及其功能会发生变化,但土壤水分有效性较低限制了这些土壤生物功能的发挥[49-50].此外,随着灌木盖度的增加及植物群落结构的改变,表层土壤黏粉粒含量增加、粗砂含量减少,进而影响土壤无机碳和物理结构.这些因素共同改变了表层土壤碳、氮、磷比值的变化规律及土壤发育过程[51-52]. ...
... 天然和人工梭梭林表层土壤碳、氮、磷含量均高于流动沙地.木本植被的存在及盖度增加,有助于提升荒漠土壤的养分含量与储量,而表层土壤碳、氮、磷储量对植被建植较为敏感[51,53-54].本研究发现,固定沙丘土壤有机碳、全氮和全磷储量均高于流动沙丘,这一趋势与腾格里沙漠和乌兰布和沙漠等区域的研究结果相近[32,54-55].然而,流动和固定沙丘表层土壤的无机氮储量变化相对较小,这可能与该区域土壤水分有效性较低,从而抑制土壤生物的活动有关[56].在人工梭梭林恢复的前期阶段(<20年),表层土壤有机碳、全磷和无机氮储量变化不明显,唯有土壤全氮储量显著增加,这与天然荒漠转变为梭梭林后,节肢动物、线虫及微生物多样性提高有关[35,50,57].魏亚娟等[32]在乌兰布和沙漠的研究也发现,在3~11年人工梭梭林中,土壤有机碳储量变化不大,而在16年时则显著提升,这与本研究结果相近.值得注意的是,在人工梭梭林恢复早期,表层土壤全氮和全磷储量随建植年限的增长而增加,但这一趋势与魏亚娟等[32]的研究稍有不同.Ma等[58]在石羊河流域的研究显示,人工梭梭林土壤有机碳储量随梭梭林建植年限增加而提升,但仍显著低于流动沙丘.本研究进一步表明,在人工梭梭林恢复的后期阶段(>20年),表层土壤的有机碳、无机氮和全磷储量随着建植年限延长而显著增加,但全氮储量在30年林龄时出现显著下降.这一变化趋势与Huang等[13]的研究相一致,即人工梭梭林的生态恢复效应在建植约20年时达到峰值,超过20年后则需要人工干预以增强其生态服务功能.在绿洲边缘区域,人工梭梭林表层土壤的有机碳、无机氮和全磷储量随梭梭林龄的延长而增长,而表层土壤全氮储量则先升后降.这可能与恢复后期表层土壤盐分的积累有关,表层土壤盐分积累抑制了土壤生物活动,进而降低了土壤全氮含量[15]. ...
... 在人工梭梭林建植过程中,非生物因子及其与生物因子的交互作用对表层土壤碳、氮、磷储量的影响大于生物因子的作用,且这些因子对土壤C∶N∶P比值的影响机制存在明显差异.本研究发现,土壤粒径组成和水盐条件等非生物因子在调控养分积累方面发挥着关键作用,这一结论与张掖绿洲边缘及其他干旱区的相关研究结果一致[52,54,59].梭梭林重建与演变过程能够有效削弱风沙活动,减少表层土壤流失,从而增强有机碳和氮积累,尤其在0~5 cm表层土壤中表现显著[54].此外,土壤黏粉粒比例增加可能是提升碳氮固存效率的重要因素[52].进一步分析表明,土壤黏粉粒含量、pH和全盐含量是非生物因子中影响人工梭梭林表层土壤有机碳、全氮和全磷储量变化的主要驱动因子.较高的黏粉粒含量有助于土壤结构的稳定性及有机质的高效积累,而pH值和盐分的变化则通过影响微生物活性及养分转化过程,进而调控土壤养分积累.特别是全盐含量的增加,促进了人工梭梭林下土壤盐结皮的形成,这一过程不仅抑制土壤水分入渗,还会限制草本植物及梭梭幼苗的萌发与生长,同时对土壤生物的活动产生负面影响,最终导致土壤全氮储量及其积累速率下降[15,51].安芳娇等[57]研究指出,在人工梭梭林的建植过程中,土壤pH值、有机碳含量和电导率是驱动线虫群落变化的主要环境因子,其中土壤电导率是影响线虫群落变化的关键因素.此外,灌木盖度、草本盖度及大型节肢动物数量是影响人工梭梭林建植过程中土壤碳、氮、磷储量变化的主要生物因子.灌木盖度与土壤黏粉粒之间的相互作用能够抑制风蚀,促进土壤细颗粒的聚集,进而提高表层土壤碳、氮、磷储量.同时,土壤中蚂蚁、蜘蛛及其他中小型土壤动物种类和数量的增加,加速了凋落物的分解过程[35].然而,土壤盐分增加及土壤水分有效性降低可能抑制了螨类和跳虫等主要分解类群的数量和多样性,从而导致人工梭梭林建植后期土壤全氮储量的下降[35,60-61].在人工梭梭林建植约20年时,通过降低灌木密度和盖度,增加挖掘兽类和节肢动物的多样性,以及提升关键捕食性动物种群的数量,可以显著增强人工梭梭林的生态系统服务功能,并提高其对气候变化和人类干扰的适应能力[13,21,48,62].总体而言,植被盖度、土壤质地与生物群落之间的耦合变化是驱动表层土壤养分储量变化的核心机制.因此,为了提升人工梭梭林的生态系统功能并实现可持续管理,有必要加强对建植过程中土壤环境和生物群落协同演变的长期监测,为相关理论研究和实践应用提供更为坚实的数据支持. ...
干旱区流动沙地建植梭梭(Haloxylon ammodendron)林后细粒物质输入对土壤碳氮积累的影响
3
2021
... 在人工梭梭林建植过程中,生物因子及其与非生物因子的交互作用显著影响表层土壤碳、氮、磷化学计量特征,相比之下,非生物因子的单独影响则相对较弱.随着人工梭梭林的重建与演变,灌木和草本植被盖度增加不仅丰富了生态系统中食物资源的数量和质量,还改变了脊椎动物和节肢动物的栖息环境,进而影响了土壤食物网的结构和功能[35].Wang等 [35]的研究表明,随着人工梭梭林建植年限的延长,拟步甲科等天然荒漠节肢动物的数量和多样性逐渐下降,而具有土壤扰动功能的蚁科节肢动物数量和多样性则逐步增加.荒漠收获蚁等不仅有助于植物种子扩散,还通过挖掘土壤提高了土壤养分含量、降低了土壤电导率,从而有利于提升土壤生物多样性及其生态功能[45,48].本文研究还发现,在人工梭梭林建植过程中,土壤大型节肢动物对表层土壤碳、氮、磷比值的影响大于中型节肢动物,这与土壤水分及盐分的限制密切相关.尽管已有研究表明,在人工梭梭林重建和演变的过程中,细菌和线虫多样性及其功能会发生变化,但土壤水分有效性较低限制了这些土壤生物功能的发挥[49-50].此外,随着灌木盖度的增加及植物群落结构的改变,表层土壤黏粉粒含量增加、粗砂含量减少,进而影响土壤无机碳和物理结构.这些因素共同改变了表层土壤碳、氮、磷比值的变化规律及土壤发育过程[51-52]. ...
... 在人工梭梭林建植过程中,非生物因子及其与生物因子的交互作用对表层土壤碳、氮、磷储量的影响大于生物因子的作用,且这些因子对土壤C∶N∶P比值的影响机制存在明显差异.本研究发现,土壤粒径组成和水盐条件等非生物因子在调控养分积累方面发挥着关键作用,这一结论与张掖绿洲边缘及其他干旱区的相关研究结果一致[52,54,59].梭梭林重建与演变过程能够有效削弱风沙活动,减少表层土壤流失,从而增强有机碳和氮积累,尤其在0~5 cm表层土壤中表现显著[54].此外,土壤黏粉粒比例增加可能是提升碳氮固存效率的重要因素[52].进一步分析表明,土壤黏粉粒含量、pH和全盐含量是非生物因子中影响人工梭梭林表层土壤有机碳、全氮和全磷储量变化的主要驱动因子.较高的黏粉粒含量有助于土壤结构的稳定性及有机质的高效积累,而pH值和盐分的变化则通过影响微生物活性及养分转化过程,进而调控土壤养分积累.特别是全盐含量的增加,促进了人工梭梭林下土壤盐结皮的形成,这一过程不仅抑制土壤水分入渗,还会限制草本植物及梭梭幼苗的萌发与生长,同时对土壤生物的活动产生负面影响,最终导致土壤全氮储量及其积累速率下降[15,51].安芳娇等[57]研究指出,在人工梭梭林的建植过程中,土壤pH值、有机碳含量和电导率是驱动线虫群落变化的主要环境因子,其中土壤电导率是影响线虫群落变化的关键因素.此外,灌木盖度、草本盖度及大型节肢动物数量是影响人工梭梭林建植过程中土壤碳、氮、磷储量变化的主要生物因子.灌木盖度与土壤黏粉粒之间的相互作用能够抑制风蚀,促进土壤细颗粒的聚集,进而提高表层土壤碳、氮、磷储量.同时,土壤中蚂蚁、蜘蛛及其他中小型土壤动物种类和数量的增加,加速了凋落物的分解过程[35].然而,土壤盐分增加及土壤水分有效性降低可能抑制了螨类和跳虫等主要分解类群的数量和多样性,从而导致人工梭梭林建植后期土壤全氮储量的下降[35,60-61].在人工梭梭林建植约20年时,通过降低灌木密度和盖度,增加挖掘兽类和节肢动物的多样性,以及提升关键捕食性动物种群的数量,可以显著增强人工梭梭林的生态系统服务功能,并提高其对气候变化和人类干扰的适应能力[13,21,48,62].总体而言,植被盖度、土壤质地与生物群落之间的耦合变化是驱动表层土壤养分储量变化的核心机制.因此,为了提升人工梭梭林的生态系统功能并实现可持续管理,有必要加强对建植过程中土壤环境和生物群落协同演变的长期监测,为相关理论研究和实践应用提供更为坚实的数据支持. ...
... [52].进一步分析表明,土壤黏粉粒含量、pH和全盐含量是非生物因子中影响人工梭梭林表层土壤有机碳、全氮和全磷储量变化的主要驱动因子.较高的黏粉粒含量有助于土壤结构的稳定性及有机质的高效积累,而pH值和盐分的变化则通过影响微生物活性及养分转化过程,进而调控土壤养分积累.特别是全盐含量的增加,促进了人工梭梭林下土壤盐结皮的形成,这一过程不仅抑制土壤水分入渗,还会限制草本植物及梭梭幼苗的萌发与生长,同时对土壤生物的活动产生负面影响,最终导致土壤全氮储量及其积累速率下降[15,51].安芳娇等[57]研究指出,在人工梭梭林的建植过程中,土壤pH值、有机碳含量和电导率是驱动线虫群落变化的主要环境因子,其中土壤电导率是影响线虫群落变化的关键因素.此外,灌木盖度、草本盖度及大型节肢动物数量是影响人工梭梭林建植过程中土壤碳、氮、磷储量变化的主要生物因子.灌木盖度与土壤黏粉粒之间的相互作用能够抑制风蚀,促进土壤细颗粒的聚集,进而提高表层土壤碳、氮、磷储量.同时,土壤中蚂蚁、蜘蛛及其他中小型土壤动物种类和数量的增加,加速了凋落物的分解过程[35].然而,土壤盐分增加及土壤水分有效性降低可能抑制了螨类和跳虫等主要分解类群的数量和多样性,从而导致人工梭梭林建植后期土壤全氮储量的下降[35,60-61].在人工梭梭林建植约20年时,通过降低灌木密度和盖度,增加挖掘兽类和节肢动物的多样性,以及提升关键捕食性动物种群的数量,可以显著增强人工梭梭林的生态系统服务功能,并提高其对气候变化和人类干扰的适应能力[13,21,48,62].总体而言,植被盖度、土壤质地与生物群落之间的耦合变化是驱动表层土壤养分储量变化的核心机制.因此,为了提升人工梭梭林的生态系统功能并实现可持续管理,有必要加强对建植过程中土壤环境和生物群落协同演变的长期监测,为相关理论研究和实践应用提供更为坚实的数据支持. ...
准噶尔盆地西缘天然梭梭林土壤有机碳及养分的分布特征
1
2014
... 天然和人工梭梭林表层土壤碳、氮、磷含量均高于流动沙地.木本植被的存在及盖度增加,有助于提升荒漠土壤的养分含量与储量,而表层土壤碳、氮、磷储量对植被建植较为敏感[51,53-54].本研究发现,固定沙丘土壤有机碳、全氮和全磷储量均高于流动沙丘,这一趋势与腾格里沙漠和乌兰布和沙漠等区域的研究结果相近[32,54-55].然而,流动和固定沙丘表层土壤的无机氮储量变化相对较小,这可能与该区域土壤水分有效性较低,从而抑制土壤生物的活动有关[56].在人工梭梭林恢复的前期阶段(<20年),表层土壤有机碳、全磷和无机氮储量变化不明显,唯有土壤全氮储量显著增加,这与天然荒漠转变为梭梭林后,节肢动物、线虫及微生物多样性提高有关[35,50,57].魏亚娟等[32]在乌兰布和沙漠的研究也发现,在3~11年人工梭梭林中,土壤有机碳储量变化不大,而在16年时则显著提升,这与本研究结果相近.值得注意的是,在人工梭梭林恢复早期,表层土壤全氮和全磷储量随建植年限的增长而增加,但这一趋势与魏亚娟等[32]的研究稍有不同.Ma等[58]在石羊河流域的研究显示,人工梭梭林土壤有机碳储量随梭梭林建植年限增加而提升,但仍显著低于流动沙丘.本研究进一步表明,在人工梭梭林恢复的后期阶段(>20年),表层土壤的有机碳、无机氮和全磷储量随着建植年限延长而显著增加,但全氮储量在30年林龄时出现显著下降.这一变化趋势与Huang等[13]的研究相一致,即人工梭梭林的生态恢复效应在建植约20年时达到峰值,超过20年后则需要人工干预以增强其生态服务功能.在绿洲边缘区域,人工梭梭林表层土壤的有机碳、无机氮和全磷储量随梭梭林龄的延长而增长,而表层土壤全氮储量则先升后降.这可能与恢复后期表层土壤盐分的积累有关,表层土壤盐分积累抑制了土壤生物活动,进而降低了土壤全氮含量[15]. ...
风沙活动对人工梭梭(Haloxylon ammodendron)林土壤碳氮含量的影响
4
2022
... 天然和人工梭梭林表层土壤碳、氮、磷含量均高于流动沙地.木本植被的存在及盖度增加,有助于提升荒漠土壤的养分含量与储量,而表层土壤碳、氮、磷储量对植被建植较为敏感[51,53-54].本研究发现,固定沙丘土壤有机碳、全氮和全磷储量均高于流动沙丘,这一趋势与腾格里沙漠和乌兰布和沙漠等区域的研究结果相近[32,54-55].然而,流动和固定沙丘表层土壤的无机氮储量变化相对较小,这可能与该区域土壤水分有效性较低,从而抑制土壤生物的活动有关[56].在人工梭梭林恢复的前期阶段(<20年),表层土壤有机碳、全磷和无机氮储量变化不明显,唯有土壤全氮储量显著增加,这与天然荒漠转变为梭梭林后,节肢动物、线虫及微生物多样性提高有关[35,50,57].魏亚娟等[32]在乌兰布和沙漠的研究也发现,在3~11年人工梭梭林中,土壤有机碳储量变化不大,而在16年时则显著提升,这与本研究结果相近.值得注意的是,在人工梭梭林恢复早期,表层土壤全氮和全磷储量随建植年限的增长而增加,但这一趋势与魏亚娟等[32]的研究稍有不同.Ma等[58]在石羊河流域的研究显示,人工梭梭林土壤有机碳储量随梭梭林建植年限增加而提升,但仍显著低于流动沙丘.本研究进一步表明,在人工梭梭林恢复的后期阶段(>20年),表层土壤的有机碳、无机氮和全磷储量随着建植年限延长而显著增加,但全氮储量在30年林龄时出现显著下降.这一变化趋势与Huang等[13]的研究相一致,即人工梭梭林的生态恢复效应在建植约20年时达到峰值,超过20年后则需要人工干预以增强其生态服务功能.在绿洲边缘区域,人工梭梭林表层土壤的有机碳、无机氮和全磷储量随梭梭林龄的延长而增长,而表层土壤全氮储量则先升后降.这可能与恢复后期表层土壤盐分的积累有关,表层土壤盐分积累抑制了土壤生物活动,进而降低了土壤全氮含量[15]. ...
... ,54-55].然而,流动和固定沙丘表层土壤的无机氮储量变化相对较小,这可能与该区域土壤水分有效性较低,从而抑制土壤生物的活动有关[56].在人工梭梭林恢复的前期阶段(<20年),表层土壤有机碳、全磷和无机氮储量变化不明显,唯有土壤全氮储量显著增加,这与天然荒漠转变为梭梭林后,节肢动物、线虫及微生物多样性提高有关[35,50,57].魏亚娟等[32]在乌兰布和沙漠的研究也发现,在3~11年人工梭梭林中,土壤有机碳储量变化不大,而在16年时则显著提升,这与本研究结果相近.值得注意的是,在人工梭梭林恢复早期,表层土壤全氮和全磷储量随建植年限的增长而增加,但这一趋势与魏亚娟等[32]的研究稍有不同.Ma等[58]在石羊河流域的研究显示,人工梭梭林土壤有机碳储量随梭梭林建植年限增加而提升,但仍显著低于流动沙丘.本研究进一步表明,在人工梭梭林恢复的后期阶段(>20年),表层土壤的有机碳、无机氮和全磷储量随着建植年限延长而显著增加,但全氮储量在30年林龄时出现显著下降.这一变化趋势与Huang等[13]的研究相一致,即人工梭梭林的生态恢复效应在建植约20年时达到峰值,超过20年后则需要人工干预以增强其生态服务功能.在绿洲边缘区域,人工梭梭林表层土壤的有机碳、无机氮和全磷储量随梭梭林龄的延长而增长,而表层土壤全氮储量则先升后降.这可能与恢复后期表层土壤盐分的积累有关,表层土壤盐分积累抑制了土壤生物活动,进而降低了土壤全氮含量[15]. ...
... 在人工梭梭林建植过程中,非生物因子及其与生物因子的交互作用对表层土壤碳、氮、磷储量的影响大于生物因子的作用,且这些因子对土壤C∶N∶P比值的影响机制存在明显差异.本研究发现,土壤粒径组成和水盐条件等非生物因子在调控养分积累方面发挥着关键作用,这一结论与张掖绿洲边缘及其他干旱区的相关研究结果一致[52,54,59].梭梭林重建与演变过程能够有效削弱风沙活动,减少表层土壤流失,从而增强有机碳和氮积累,尤其在0~5 cm表层土壤中表现显著[54].此外,土壤黏粉粒比例增加可能是提升碳氮固存效率的重要因素[52].进一步分析表明,土壤黏粉粒含量、pH和全盐含量是非生物因子中影响人工梭梭林表层土壤有机碳、全氮和全磷储量变化的主要驱动因子.较高的黏粉粒含量有助于土壤结构的稳定性及有机质的高效积累,而pH值和盐分的变化则通过影响微生物活性及养分转化过程,进而调控土壤养分积累.特别是全盐含量的增加,促进了人工梭梭林下土壤盐结皮的形成,这一过程不仅抑制土壤水分入渗,还会限制草本植物及梭梭幼苗的萌发与生长,同时对土壤生物的活动产生负面影响,最终导致土壤全氮储量及其积累速率下降[15,51].安芳娇等[57]研究指出,在人工梭梭林的建植过程中,土壤pH值、有机碳含量和电导率是驱动线虫群落变化的主要环境因子,其中土壤电导率是影响线虫群落变化的关键因素.此外,灌木盖度、草本盖度及大型节肢动物数量是影响人工梭梭林建植过程中土壤碳、氮、磷储量变化的主要生物因子.灌木盖度与土壤黏粉粒之间的相互作用能够抑制风蚀,促进土壤细颗粒的聚集,进而提高表层土壤碳、氮、磷储量.同时,土壤中蚂蚁、蜘蛛及其他中小型土壤动物种类和数量的增加,加速了凋落物的分解过程[35].然而,土壤盐分增加及土壤水分有效性降低可能抑制了螨类和跳虫等主要分解类群的数量和多样性,从而导致人工梭梭林建植后期土壤全氮储量的下降[35,60-61].在人工梭梭林建植约20年时,通过降低灌木密度和盖度,增加挖掘兽类和节肢动物的多样性,以及提升关键捕食性动物种群的数量,可以显著增强人工梭梭林的生态系统服务功能,并提高其对气候变化和人类干扰的适应能力[13,21,48,62].总体而言,植被盖度、土壤质地与生物群落之间的耦合变化是驱动表层土壤养分储量变化的核心机制.因此,为了提升人工梭梭林的生态系统功能并实现可持续管理,有必要加强对建植过程中土壤环境和生物群落协同演变的长期监测,为相关理论研究和实践应用提供更为坚实的数据支持. ...
... [54].此外,土壤黏粉粒比例增加可能是提升碳氮固存效率的重要因素[52].进一步分析表明,土壤黏粉粒含量、pH和全盐含量是非生物因子中影响人工梭梭林表层土壤有机碳、全氮和全磷储量变化的主要驱动因子.较高的黏粉粒含量有助于土壤结构的稳定性及有机质的高效积累,而pH值和盐分的变化则通过影响微生物活性及养分转化过程,进而调控土壤养分积累.特别是全盐含量的增加,促进了人工梭梭林下土壤盐结皮的形成,这一过程不仅抑制土壤水分入渗,还会限制草本植物及梭梭幼苗的萌发与生长,同时对土壤生物的活动产生负面影响,最终导致土壤全氮储量及其积累速率下降[15,51].安芳娇等[57]研究指出,在人工梭梭林的建植过程中,土壤pH值、有机碳含量和电导率是驱动线虫群落变化的主要环境因子,其中土壤电导率是影响线虫群落变化的关键因素.此外,灌木盖度、草本盖度及大型节肢动物数量是影响人工梭梭林建植过程中土壤碳、氮、磷储量变化的主要生物因子.灌木盖度与土壤黏粉粒之间的相互作用能够抑制风蚀,促进土壤细颗粒的聚集,进而提高表层土壤碳、氮、磷储量.同时,土壤中蚂蚁、蜘蛛及其他中小型土壤动物种类和数量的增加,加速了凋落物的分解过程[35].然而,土壤盐分增加及土壤水分有效性降低可能抑制了螨类和跳虫等主要分解类群的数量和多样性,从而导致人工梭梭林建植后期土壤全氮储量的下降[35,60-61].在人工梭梭林建植约20年时,通过降低灌木密度和盖度,增加挖掘兽类和节肢动物的多样性,以及提升关键捕食性动物种群的数量,可以显著增强人工梭梭林的生态系统服务功能,并提高其对气候变化和人类干扰的适应能力[13,21,48,62].总体而言,植被盖度、土壤质地与生物群落之间的耦合变化是驱动表层土壤养分储量变化的核心机制.因此,为了提升人工梭梭林的生态系统功能并实现可持续管理,有必要加强对建植过程中土壤环境和生物群落协同演变的长期监测,为相关理论研究和实践应用提供更为坚实的数据支持. ...
腾格里沙漠南缘植被恢复对土壤有机碳组成及稳定性的影响
1
2024
... 天然和人工梭梭林表层土壤碳、氮、磷含量均高于流动沙地.木本植被的存在及盖度增加,有助于提升荒漠土壤的养分含量与储量,而表层土壤碳、氮、磷储量对植被建植较为敏感[51,53-54].本研究发现,固定沙丘土壤有机碳、全氮和全磷储量均高于流动沙丘,这一趋势与腾格里沙漠和乌兰布和沙漠等区域的研究结果相近[32,54-55].然而,流动和固定沙丘表层土壤的无机氮储量变化相对较小,这可能与该区域土壤水分有效性较低,从而抑制土壤生物的活动有关[56].在人工梭梭林恢复的前期阶段(<20年),表层土壤有机碳、全磷和无机氮储量变化不明显,唯有土壤全氮储量显著增加,这与天然荒漠转变为梭梭林后,节肢动物、线虫及微生物多样性提高有关[35,50,57].魏亚娟等[32]在乌兰布和沙漠的研究也发现,在3~11年人工梭梭林中,土壤有机碳储量变化不大,而在16年时则显著提升,这与本研究结果相近.值得注意的是,在人工梭梭林恢复早期,表层土壤全氮和全磷储量随建植年限的增长而增加,但这一趋势与魏亚娟等[32]的研究稍有不同.Ma等[58]在石羊河流域的研究显示,人工梭梭林土壤有机碳储量随梭梭林建植年限增加而提升,但仍显著低于流动沙丘.本研究进一步表明,在人工梭梭林恢复的后期阶段(>20年),表层土壤的有机碳、无机氮和全磷储量随着建植年限延长而显著增加,但全氮储量在30年林龄时出现显著下降.这一变化趋势与Huang等[13]的研究相一致,即人工梭梭林的生态恢复效应在建植约20年时达到峰值,超过20年后则需要人工干预以增强其生态服务功能.在绿洲边缘区域,人工梭梭林表层土壤的有机碳、无机氮和全磷储量随梭梭林龄的延长而增长,而表层土壤全氮储量则先升后降.这可能与恢复后期表层土壤盐分的积累有关,表层土壤盐分积累抑制了土壤生物活动,进而降低了土壤全氮含量[15]. ...
Impacts of altered precipitation regimes on soil communities and biogeochemistry in arid and semi‐arid ecosystems
1
2015
... 天然和人工梭梭林表层土壤碳、氮、磷含量均高于流动沙地.木本植被的存在及盖度增加,有助于提升荒漠土壤的养分含量与储量,而表层土壤碳、氮、磷储量对植被建植较为敏感[51,53-54].本研究发现,固定沙丘土壤有机碳、全氮和全磷储量均高于流动沙丘,这一趋势与腾格里沙漠和乌兰布和沙漠等区域的研究结果相近[32,54-55].然而,流动和固定沙丘表层土壤的无机氮储量变化相对较小,这可能与该区域土壤水分有效性较低,从而抑制土壤生物的活动有关[56].在人工梭梭林恢复的前期阶段(<20年),表层土壤有机碳、全磷和无机氮储量变化不明显,唯有土壤全氮储量显著增加,这与天然荒漠转变为梭梭林后,节肢动物、线虫及微生物多样性提高有关[35,50,57].魏亚娟等[32]在乌兰布和沙漠的研究也发现,在3~11年人工梭梭林中,土壤有机碳储量变化不大,而在16年时则显著提升,这与本研究结果相近.值得注意的是,在人工梭梭林恢复早期,表层土壤全氮和全磷储量随建植年限的增长而增加,但这一趋势与魏亚娟等[32]的研究稍有不同.Ma等[58]在石羊河流域的研究显示,人工梭梭林土壤有机碳储量随梭梭林建植年限增加而提升,但仍显著低于流动沙丘.本研究进一步表明,在人工梭梭林恢复的后期阶段(>20年),表层土壤的有机碳、无机氮和全磷储量随着建植年限延长而显著增加,但全氮储量在30年林龄时出现显著下降.这一变化趋势与Huang等[13]的研究相一致,即人工梭梭林的生态恢复效应在建植约20年时达到峰值,超过20年后则需要人工干预以增强其生态服务功能.在绿洲边缘区域,人工梭梭林表层土壤的有机碳、无机氮和全磷储量随梭梭林龄的延长而增长,而表层土壤全氮储量则先升后降.这可能与恢复后期表层土壤盐分的积累有关,表层土壤盐分积累抑制了土壤生物活动,进而降低了土壤全氮含量[15]. ...
干旱区荒漠绿洲过渡带建植梭梭(Haloxylon ammodendron)林后土壤线虫群落演变
2
2024
... 天然和人工梭梭林表层土壤碳、氮、磷含量均高于流动沙地.木本植被的存在及盖度增加,有助于提升荒漠土壤的养分含量与储量,而表层土壤碳、氮、磷储量对植被建植较为敏感[51,53-54].本研究发现,固定沙丘土壤有机碳、全氮和全磷储量均高于流动沙丘,这一趋势与腾格里沙漠和乌兰布和沙漠等区域的研究结果相近[32,54-55].然而,流动和固定沙丘表层土壤的无机氮储量变化相对较小,这可能与该区域土壤水分有效性较低,从而抑制土壤生物的活动有关[56].在人工梭梭林恢复的前期阶段(<20年),表层土壤有机碳、全磷和无机氮储量变化不明显,唯有土壤全氮储量显著增加,这与天然荒漠转变为梭梭林后,节肢动物、线虫及微生物多样性提高有关[35,50,57].魏亚娟等[32]在乌兰布和沙漠的研究也发现,在3~11年人工梭梭林中,土壤有机碳储量变化不大,而在16年时则显著提升,这与本研究结果相近.值得注意的是,在人工梭梭林恢复早期,表层土壤全氮和全磷储量随建植年限的增长而增加,但这一趋势与魏亚娟等[32]的研究稍有不同.Ma等[58]在石羊河流域的研究显示,人工梭梭林土壤有机碳储量随梭梭林建植年限增加而提升,但仍显著低于流动沙丘.本研究进一步表明,在人工梭梭林恢复的后期阶段(>20年),表层土壤的有机碳、无机氮和全磷储量随着建植年限延长而显著增加,但全氮储量在30年林龄时出现显著下降.这一变化趋势与Huang等[13]的研究相一致,即人工梭梭林的生态恢复效应在建植约20年时达到峰值,超过20年后则需要人工干预以增强其生态服务功能.在绿洲边缘区域,人工梭梭林表层土壤的有机碳、无机氮和全磷储量随梭梭林龄的延长而增长,而表层土壤全氮储量则先升后降.这可能与恢复后期表层土壤盐分的积累有关,表层土壤盐分积累抑制了土壤生物活动,进而降低了土壤全氮含量[15]. ...
... 在人工梭梭林建植过程中,非生物因子及其与生物因子的交互作用对表层土壤碳、氮、磷储量的影响大于生物因子的作用,且这些因子对土壤C∶N∶P比值的影响机制存在明显差异.本研究发现,土壤粒径组成和水盐条件等非生物因子在调控养分积累方面发挥着关键作用,这一结论与张掖绿洲边缘及其他干旱区的相关研究结果一致[52,54,59].梭梭林重建与演变过程能够有效削弱风沙活动,减少表层土壤流失,从而增强有机碳和氮积累,尤其在0~5 cm表层土壤中表现显著[54].此外,土壤黏粉粒比例增加可能是提升碳氮固存效率的重要因素[52].进一步分析表明,土壤黏粉粒含量、pH和全盐含量是非生物因子中影响人工梭梭林表层土壤有机碳、全氮和全磷储量变化的主要驱动因子.较高的黏粉粒含量有助于土壤结构的稳定性及有机质的高效积累,而pH值和盐分的变化则通过影响微生物活性及养分转化过程,进而调控土壤养分积累.特别是全盐含量的增加,促进了人工梭梭林下土壤盐结皮的形成,这一过程不仅抑制土壤水分入渗,还会限制草本植物及梭梭幼苗的萌发与生长,同时对土壤生物的活动产生负面影响,最终导致土壤全氮储量及其积累速率下降[15,51].安芳娇等[57]研究指出,在人工梭梭林的建植过程中,土壤pH值、有机碳含量和电导率是驱动线虫群落变化的主要环境因子,其中土壤电导率是影响线虫群落变化的关键因素.此外,灌木盖度、草本盖度及大型节肢动物数量是影响人工梭梭林建植过程中土壤碳、氮、磷储量变化的主要生物因子.灌木盖度与土壤黏粉粒之间的相互作用能够抑制风蚀,促进土壤细颗粒的聚集,进而提高表层土壤碳、氮、磷储量.同时,土壤中蚂蚁、蜘蛛及其他中小型土壤动物种类和数量的增加,加速了凋落物的分解过程[35].然而,土壤盐分增加及土壤水分有效性降低可能抑制了螨类和跳虫等主要分解类群的数量和多样性,从而导致人工梭梭林建植后期土壤全氮储量的下降[35,60-61].在人工梭梭林建植约20年时,通过降低灌木密度和盖度,增加挖掘兽类和节肢动物的多样性,以及提升关键捕食性动物种群的数量,可以显著增强人工梭梭林的生态系统服务功能,并提高其对气候变化和人类干扰的适应能力[13,21,48,62].总体而言,植被盖度、土壤质地与生物群落之间的耦合变化是驱动表层土壤养分储量变化的核心机制.因此,为了提升人工梭梭林的生态系统功能并实现可持续管理,有必要加强对建植过程中土壤环境和生物群落协同演变的长期监测,为相关理论研究和实践应用提供更为坚实的数据支持. ...
Carbon sequestration of sand-fixing plantation of Haloxylon ammodendron in Shiyang River Basin: storage,rate and potential
1
2021
... 天然和人工梭梭林表层土壤碳、氮、磷含量均高于流动沙地.木本植被的存在及盖度增加,有助于提升荒漠土壤的养分含量与储量,而表层土壤碳、氮、磷储量对植被建植较为敏感[51,53-54].本研究发现,固定沙丘土壤有机碳、全氮和全磷储量均高于流动沙丘,这一趋势与腾格里沙漠和乌兰布和沙漠等区域的研究结果相近[32,54-55].然而,流动和固定沙丘表层土壤的无机氮储量变化相对较小,这可能与该区域土壤水分有效性较低,从而抑制土壤生物的活动有关[56].在人工梭梭林恢复的前期阶段(<20年),表层土壤有机碳、全磷和无机氮储量变化不明显,唯有土壤全氮储量显著增加,这与天然荒漠转变为梭梭林后,节肢动物、线虫及微生物多样性提高有关[35,50,57].魏亚娟等[32]在乌兰布和沙漠的研究也发现,在3~11年人工梭梭林中,土壤有机碳储量变化不大,而在16年时则显著提升,这与本研究结果相近.值得注意的是,在人工梭梭林恢复早期,表层土壤全氮和全磷储量随建植年限的增长而增加,但这一趋势与魏亚娟等[32]的研究稍有不同.Ma等[58]在石羊河流域的研究显示,人工梭梭林土壤有机碳储量随梭梭林建植年限增加而提升,但仍显著低于流动沙丘.本研究进一步表明,在人工梭梭林恢复的后期阶段(>20年),表层土壤的有机碳、无机氮和全磷储量随着建植年限延长而显著增加,但全氮储量在30年林龄时出现显著下降.这一变化趋势与Huang等[13]的研究相一致,即人工梭梭林的生态恢复效应在建植约20年时达到峰值,超过20年后则需要人工干预以增强其生态服务功能.在绿洲边缘区域,人工梭梭林表层土壤的有机碳、无机氮和全磷储量随梭梭林龄的延长而增长,而表层土壤全氮储量则先升后降.这可能与恢复后期表层土壤盐分的积累有关,表层土壤盐分积累抑制了土壤生物活动,进而降低了土壤全氮含量[15]. ...
Ecological restoration enhances dryland carbon stock by reducing surface soil carbon loss due to wind erosion
1
2024
... 在人工梭梭林建植过程中,非生物因子及其与生物因子的交互作用对表层土壤碳、氮、磷储量的影响大于生物因子的作用,且这些因子对土壤C∶N∶P比值的影响机制存在明显差异.本研究发现,土壤粒径组成和水盐条件等非生物因子在调控养分积累方面发挥着关键作用,这一结论与张掖绿洲边缘及其他干旱区的相关研究结果一致[52,54,59].梭梭林重建与演变过程能够有效削弱风沙活动,减少表层土壤流失,从而增强有机碳和氮积累,尤其在0~5 cm表层土壤中表现显著[54].此外,土壤黏粉粒比例增加可能是提升碳氮固存效率的重要因素[52].进一步分析表明,土壤黏粉粒含量、pH和全盐含量是非生物因子中影响人工梭梭林表层土壤有机碳、全氮和全磷储量变化的主要驱动因子.较高的黏粉粒含量有助于土壤结构的稳定性及有机质的高效积累,而pH值和盐分的变化则通过影响微生物活性及养分转化过程,进而调控土壤养分积累.特别是全盐含量的增加,促进了人工梭梭林下土壤盐结皮的形成,这一过程不仅抑制土壤水分入渗,还会限制草本植物及梭梭幼苗的萌发与生长,同时对土壤生物的活动产生负面影响,最终导致土壤全氮储量及其积累速率下降[15,51].安芳娇等[57]研究指出,在人工梭梭林的建植过程中,土壤pH值、有机碳含量和电导率是驱动线虫群落变化的主要环境因子,其中土壤电导率是影响线虫群落变化的关键因素.此外,灌木盖度、草本盖度及大型节肢动物数量是影响人工梭梭林建植过程中土壤碳、氮、磷储量变化的主要生物因子.灌木盖度与土壤黏粉粒之间的相互作用能够抑制风蚀,促进土壤细颗粒的聚集,进而提高表层土壤碳、氮、磷储量.同时,土壤中蚂蚁、蜘蛛及其他中小型土壤动物种类和数量的增加,加速了凋落物的分解过程[35].然而,土壤盐分增加及土壤水分有效性降低可能抑制了螨类和跳虫等主要分解类群的数量和多样性,从而导致人工梭梭林建植后期土壤全氮储量的下降[35,60-61].在人工梭梭林建植约20年时,通过降低灌木密度和盖度,增加挖掘兽类和节肢动物的多样性,以及提升关键捕食性动物种群的数量,可以显著增强人工梭梭林的生态系统服务功能,并提高其对气候变化和人类干扰的适应能力[13,21,48,62].总体而言,植被盖度、土壤质地与生物群落之间的耦合变化是驱动表层土壤养分储量变化的核心机制.因此,为了提升人工梭梭林的生态系统功能并实现可持续管理,有必要加强对建植过程中土壤环境和生物群落协同演变的长期监测,为相关理论研究和实践应用提供更为坚实的数据支持. ...
黑河中游不同土地利用方式地面节肢动物对土壤盐渍化的响应
1
2011
... 在人工梭梭林建植过程中,非生物因子及其与生物因子的交互作用对表层土壤碳、氮、磷储量的影响大于生物因子的作用,且这些因子对土壤C∶N∶P比值的影响机制存在明显差异.本研究发现,土壤粒径组成和水盐条件等非生物因子在调控养分积累方面发挥着关键作用,这一结论与张掖绿洲边缘及其他干旱区的相关研究结果一致[52,54,59].梭梭林重建与演变过程能够有效削弱风沙活动,减少表层土壤流失,从而增强有机碳和氮积累,尤其在0~5 cm表层土壤中表现显著[54].此外,土壤黏粉粒比例增加可能是提升碳氮固存效率的重要因素[52].进一步分析表明,土壤黏粉粒含量、pH和全盐含量是非生物因子中影响人工梭梭林表层土壤有机碳、全氮和全磷储量变化的主要驱动因子.较高的黏粉粒含量有助于土壤结构的稳定性及有机质的高效积累,而pH值和盐分的变化则通过影响微生物活性及养分转化过程,进而调控土壤养分积累.特别是全盐含量的增加,促进了人工梭梭林下土壤盐结皮的形成,这一过程不仅抑制土壤水分入渗,还会限制草本植物及梭梭幼苗的萌发与生长,同时对土壤生物的活动产生负面影响,最终导致土壤全氮储量及其积累速率下降[15,51].安芳娇等[57]研究指出,在人工梭梭林的建植过程中,土壤pH值、有机碳含量和电导率是驱动线虫群落变化的主要环境因子,其中土壤电导率是影响线虫群落变化的关键因素.此外,灌木盖度、草本盖度及大型节肢动物数量是影响人工梭梭林建植过程中土壤碳、氮、磷储量变化的主要生物因子.灌木盖度与土壤黏粉粒之间的相互作用能够抑制风蚀,促进土壤细颗粒的聚集,进而提高表层土壤碳、氮、磷储量.同时,土壤中蚂蚁、蜘蛛及其他中小型土壤动物种类和数量的增加,加速了凋落物的分解过程[35].然而,土壤盐分增加及土壤水分有效性降低可能抑制了螨类和跳虫等主要分解类群的数量和多样性,从而导致人工梭梭林建植后期土壤全氮储量的下降[35,60-61].在人工梭梭林建植约20年时,通过降低灌木密度和盖度,增加挖掘兽类和节肢动物的多样性,以及提升关键捕食性动物种群的数量,可以显著增强人工梭梭林的生态系统服务功能,并提高其对气候变化和人类干扰的适应能力[13,21,48,62].总体而言,植被盖度、土壤质地与生物群落之间的耦合变化是驱动表层土壤养分储量变化的核心机制.因此,为了提升人工梭梭林的生态系统功能并实现可持续管理,有必要加强对建植过程中土壤环境和生物群落协同演变的长期监测,为相关理论研究和实践应用提供更为坚实的数据支持. ...
Responses of different collembola and mite taxa to experimental rain pulses in an arid ecosystem
1
2017
... 在人工梭梭林建植过程中,非生物因子及其与生物因子的交互作用对表层土壤碳、氮、磷储量的影响大于生物因子的作用,且这些因子对土壤C∶N∶P比值的影响机制存在明显差异.本研究发现,土壤粒径组成和水盐条件等非生物因子在调控养分积累方面发挥着关键作用,这一结论与张掖绿洲边缘及其他干旱区的相关研究结果一致[52,54,59].梭梭林重建与演变过程能够有效削弱风沙活动,减少表层土壤流失,从而增强有机碳和氮积累,尤其在0~5 cm表层土壤中表现显著[54].此外,土壤黏粉粒比例增加可能是提升碳氮固存效率的重要因素[52].进一步分析表明,土壤黏粉粒含量、pH和全盐含量是非生物因子中影响人工梭梭林表层土壤有机碳、全氮和全磷储量变化的主要驱动因子.较高的黏粉粒含量有助于土壤结构的稳定性及有机质的高效积累,而pH值和盐分的变化则通过影响微生物活性及养分转化过程,进而调控土壤养分积累.特别是全盐含量的增加,促进了人工梭梭林下土壤盐结皮的形成,这一过程不仅抑制土壤水分入渗,还会限制草本植物及梭梭幼苗的萌发与生长,同时对土壤生物的活动产生负面影响,最终导致土壤全氮储量及其积累速率下降[15,51].安芳娇等[57]研究指出,在人工梭梭林的建植过程中,土壤pH值、有机碳含量和电导率是驱动线虫群落变化的主要环境因子,其中土壤电导率是影响线虫群落变化的关键因素.此外,灌木盖度、草本盖度及大型节肢动物数量是影响人工梭梭林建植过程中土壤碳、氮、磷储量变化的主要生物因子.灌木盖度与土壤黏粉粒之间的相互作用能够抑制风蚀,促进土壤细颗粒的聚集,进而提高表层土壤碳、氮、磷储量.同时,土壤中蚂蚁、蜘蛛及其他中小型土壤动物种类和数量的增加,加速了凋落物的分解过程[35].然而,土壤盐分增加及土壤水分有效性降低可能抑制了螨类和跳虫等主要分解类群的数量和多样性,从而导致人工梭梭林建植后期土壤全氮储量的下降[35,60-61].在人工梭梭林建植约20年时,通过降低灌木密度和盖度,增加挖掘兽类和节肢动物的多样性,以及提升关键捕食性动物种群的数量,可以显著增强人工梭梭林的生态系统服务功能,并提高其对气候变化和人类干扰的适应能力[13,21,48,62].总体而言,植被盖度、土壤质地与生物群落之间的耦合变化是驱动表层土壤养分储量变化的核心机制.因此,为了提升人工梭梭林的生态系统功能并实现可持续管理,有必要加强对建植过程中土壤环境和生物群落协同演变的长期监测,为相关理论研究和实践应用提供更为坚实的数据支持. ...
Global meta‐analysis of soil‐disturbing vertebrates reveals strong effects on ecosystem patterns and processes
1
2019
... 在人工梭梭林建植过程中,非生物因子及其与生物因子的交互作用对表层土壤碳、氮、磷储量的影响大于生物因子的作用,且这些因子对土壤C∶N∶P比值的影响机制存在明显差异.本研究发现,土壤粒径组成和水盐条件等非生物因子在调控养分积累方面发挥着关键作用,这一结论与张掖绿洲边缘及其他干旱区的相关研究结果一致[52,54,59].梭梭林重建与演变过程能够有效削弱风沙活动,减少表层土壤流失,从而增强有机碳和氮积累,尤其在0~5 cm表层土壤中表现显著[54].此外,土壤黏粉粒比例增加可能是提升碳氮固存效率的重要因素[52].进一步分析表明,土壤黏粉粒含量、pH和全盐含量是非生物因子中影响人工梭梭林表层土壤有机碳、全氮和全磷储量变化的主要驱动因子.较高的黏粉粒含量有助于土壤结构的稳定性及有机质的高效积累,而pH值和盐分的变化则通过影响微生物活性及养分转化过程,进而调控土壤养分积累.特别是全盐含量的增加,促进了人工梭梭林下土壤盐结皮的形成,这一过程不仅抑制土壤水分入渗,还会限制草本植物及梭梭幼苗的萌发与生长,同时对土壤生物的活动产生负面影响,最终导致土壤全氮储量及其积累速率下降[15,51].安芳娇等[57]研究指出,在人工梭梭林的建植过程中,土壤pH值、有机碳含量和电导率是驱动线虫群落变化的主要环境因子,其中土壤电导率是影响线虫群落变化的关键因素.此外,灌木盖度、草本盖度及大型节肢动物数量是影响人工梭梭林建植过程中土壤碳、氮、磷储量变化的主要生物因子.灌木盖度与土壤黏粉粒之间的相互作用能够抑制风蚀,促进土壤细颗粒的聚集,进而提高表层土壤碳、氮、磷储量.同时,土壤中蚂蚁、蜘蛛及其他中小型土壤动物种类和数量的增加,加速了凋落物的分解过程[35].然而,土壤盐分增加及土壤水分有效性降低可能抑制了螨类和跳虫等主要分解类群的数量和多样性,从而导致人工梭梭林建植后期土壤全氮储量的下降[35,60-61].在人工梭梭林建植约20年时,通过降低灌木密度和盖度,增加挖掘兽类和节肢动物的多样性,以及提升关键捕食性动物种群的数量,可以显著增强人工梭梭林的生态系统服务功能,并提高其对气候变化和人类干扰的适应能力[13,21,48,62].总体而言,植被盖度、土壤质地与生物群落之间的耦合变化是驱动表层土壤养分储量变化的核心机制.因此,为了提升人工梭梭林的生态系统功能并实现可持续管理,有必要加强对建植过程中土壤环境和生物群落协同演变的长期监测,为相关理论研究和实践应用提供更为坚实的数据支持. ...






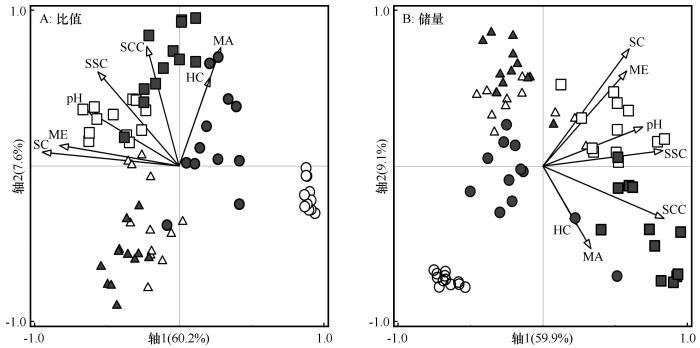

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}