Cyanobacteria:model microorganisms and beyond
2
2022
... 蓝藻或蓝绿藻也称蓝细菌(Cyanobacteria),在形态上主要可分为单细胞、群落和丝状蓝藻,适应性极强,可在荒漠、热泉、极地、冰川、高盐海域等区域中生存[1],是地球上最古老的原核放氧光合生物,约35亿年前就已出现在地球上[2].蓝藻具有重要的生态功能,对地球环境演化意义重大,通过光合作用,蓝藻主导了约20亿年前发生的地球大氧化事件,并改变了地球的大气成分比例,使大气成分从还原态转变成富氧化态,为需氧型生物的进化及水生生物登陆提供了前提条件[3],同时它也是真核光合生物叶绿体的祖先[4]. ...
... EPS在蓝藻的滑动过程中具有多种功能.蓝藻滑动时,分泌的EPS通常可覆盖整个藻丝表面,但不随藻丝进行位移,以塌陷的、有时扭曲的残留物形式被遗弃在前移藻丝末端后方[14].蓝藻细胞在滑动时倾向于使用其他蓝藻细胞滑动遗留的轨迹进行运动[18],原因可能是残留EPS的存在减少或免去了跟随细胞需分泌的EPS量,使得该细胞在被跟随细胞轨迹上的滑动过程更为经济.EPS还可提供有关近期存在于附近位置的细胞浓度的信息,类似于化学群体感应,因此,随着蓝藻群体的发展,EPS 踪迹能够提供一种持久的远程间接通信机制,可以引导滞后细胞与前端群体合并[21].此外,EPS还能通过改变细胞表面和周围光的折射率来影响细胞滑动[1].当前已知影响蓝藻EPS合成及分泌的主要因素包括NaCl含量、气体流通速率、温度、光强、pH值、重金属,以及氮、磷、硫、镁、钙、钾等元素的含量等[20-24]. ...
Undirected motility of filamentous cyanobacteria produces reticulate mats
1
2010
... 蓝藻或蓝绿藻也称蓝细菌(Cyanobacteria),在形态上主要可分为单细胞、群落和丝状蓝藻,适应性极强,可在荒漠、热泉、极地、冰川、高盐海域等区域中生存[1],是地球上最古老的原核放氧光合生物,约35亿年前就已出现在地球上[2].蓝藻具有重要的生态功能,对地球环境演化意义重大,通过光合作用,蓝藻主导了约20亿年前发生的地球大氧化事件,并改变了地球的大气成分比例,使大气成分从还原态转变成富氧化态,为需氧型生物的进化及水生生物登陆提供了前提条件[3],同时它也是真核光合生物叶绿体的祖先[4]. ...
Responding to chemical gradients:bacterial chemotaxis
1
2012
... 蓝藻或蓝绿藻也称蓝细菌(Cyanobacteria),在形态上主要可分为单细胞、群落和丝状蓝藻,适应性极强,可在荒漠、热泉、极地、冰川、高盐海域等区域中生存[1],是地球上最古老的原核放氧光合生物,约35亿年前就已出现在地球上[2].蓝藻具有重要的生态功能,对地球环境演化意义重大,通过光合作用,蓝藻主导了约20亿年前发生的地球大氧化事件,并改变了地球的大气成分比例,使大气成分从还原态转变成富氧化态,为需氧型生物的进化及水生生物登陆提供了前提条件[3],同时它也是真核光合生物叶绿体的祖先[4]. ...
Rapid reactivation of cyanobacterial photosynthesis and migration upon rehydration of desiccated marine Microbial mats
2
2015
... 蓝藻或蓝绿藻也称蓝细菌(Cyanobacteria),在形态上主要可分为单细胞、群落和丝状蓝藻,适应性极强,可在荒漠、热泉、极地、冰川、高盐海域等区域中生存[1],是地球上最古老的原核放氧光合生物,约35亿年前就已出现在地球上[2].蓝藻具有重要的生态功能,对地球环境演化意义重大,通过光合作用,蓝藻主导了约20亿年前发生的地球大氧化事件,并改变了地球的大气成分比例,使大气成分从还原态转变成富氧化态,为需氧型生物的进化及水生生物登陆提供了前提条件[3],同时它也是真核光合生物叶绿体的祖先[4]. ...
... 微生物席(Microbial mats)也可称微生物垫或菌毯,是沉积物基质中生活的各种细菌、古菌和真核微藻的密集底栖群落与其构成的生物复合体,蓝藻是生物席中主要的初级生产者[4].由于沉积物基质中光的急剧衰减和物质传递速率的限制,微生物席的特点是拥有氧气、光强及光谱组成、pH值和硫化物浓度等的陡峭垂直梯度[31].蓝藻可以通过滑动来调整自身在微生物席内不断变化的光场中的位置,从而处于具有最佳光照强度的区域,避免光照不足或受到过强辐射的负面影响[52-54].Lichtenberg等[54]于2020年对在丹麦利姆峡湾定期干燥的沙地上收集的微生物席研究中发现,弱光下蓝藻出现在生物席表面;而在高光强下,蓝藻出现在较深层,说明蓝藻可以通过响应光照强度进行垂直滑动来调整其在生物席中的位置,并且蓝藻在高光强下是负趋光的,即具有避光性,而在低光强下则具有正趋光性.这种特性在螺旋藻(Spirulina)和颤藻(Oscillatoria)中也有发现[31].北威尔士梅奈海峡安格尔西海岸的沿岸泥滩中生活着大量微鞘藻属蓝藻(Microcoleus),泥滩表面在阴天所呈现的绿色较之于晴天时更为明显,说明泥滩中的大多数蓝藻在弱光时具有趋光性,在高光强下具有避光性,当前还尚未确定此类微鞘藻改变趋光向性的阈值光强.上述微鞘藻藻丝在黑暗时也会移动到泥滩表面,但在到达表面后,藻丝在有无光时所形成的图案有所不同:黑暗时藻丝在泥滩表面呈螺旋状,而有光照时藻丝长轴方向与光源方向平行,若此时光源方向改变 90°,藻丝则会重新定向.这说明不只是单个细胞,细胞群落也会集体响应光照条件[36,55]. ...
Establishing rates of lateral expansion of cyanobacterial biological soil crusts for optimal restoration
1
2018
... 蓝藻具有多种主动运动方式,包括泳动、蹭动和滑动等,其中陆生蓝藻的滑动运动在陆地环境中展现出独特的生理适应机制和生态功能[5].从生理层面来看,滑动运动使蓝藻能够主动迁移至适宜的微生境,有效应对陆地环境中资源分布不均和极端环境条件的挑战[6].在生态功能方面,荒漠蓝藻通过滑动过程分泌胞外聚合物(extracellular polymeric substances, EPS),这些物质不仅能够黏结固定沙砾,还能增加土壤有机质含量.特别值得注意的是,丝状蓝藻的藻丝在滑动过程中能够对沙砾进行物理性缠绕和捆绑,形成复杂的三维网状结构.这种独特的生物固定机制不仅显著提高了荒漠地表的稳定性,还促进了生物土壤结皮的形成,从而有效减少水土流失,增强土壤的持水能力[6-7].这种由蓝藻滑动驱动的生态过程对荒漠生态系统功能的维持和退化土地的生态修复具有关键作用[8].滑动行为使陆生蓝藻从“被动耐受”转变为“主动适应”环境,正是这种进化转变,使得陆生蓝藻成为陆地生态系统早期拓殖的先驱物种,并在现代生态修复实践中发挥着不可替代的重要角色. ...
Effect of desert soil algae on the stabilization of fine sands
2
2002
... 蓝藻具有多种主动运动方式,包括泳动、蹭动和滑动等,其中陆生蓝藻的滑动运动在陆地环境中展现出独特的生理适应机制和生态功能[5].从生理层面来看,滑动运动使蓝藻能够主动迁移至适宜的微生境,有效应对陆地环境中资源分布不均和极端环境条件的挑战[6].在生态功能方面,荒漠蓝藻通过滑动过程分泌胞外聚合物(extracellular polymeric substances, EPS),这些物质不仅能够黏结固定沙砾,还能增加土壤有机质含量.特别值得注意的是,丝状蓝藻的藻丝在滑动过程中能够对沙砾进行物理性缠绕和捆绑,形成复杂的三维网状结构.这种独特的生物固定机制不仅显著提高了荒漠地表的稳定性,还促进了生物土壤结皮的形成,从而有效减少水土流失,增强土壤的持水能力[6-7].这种由蓝藻滑动驱动的生态过程对荒漠生态系统功能的维持和退化土地的生态修复具有关键作用[8].滑动行为使陆生蓝藻从“被动耐受”转变为“主动适应”环境,正是这种进化转变,使得陆生蓝藻成为陆地生态系统早期拓殖的先驱物种,并在现代生态修复实践中发挥着不可替代的重要角色. ...
... [6-7].这种由蓝藻滑动驱动的生态过程对荒漠生态系统功能的维持和退化土地的生态修复具有关键作用[8].滑动行为使陆生蓝藻从“被动耐受”转变为“主动适应”环境,正是这种进化转变,使得陆生蓝藻成为陆地生态系统早期拓殖的先驱物种,并在现代生态修复实践中发挥着不可替代的重要角色. ...
Aeolian dust deposition as a driver of cyanobacterial community structure in biological soil crusts
1
2025
... 蓝藻具有多种主动运动方式,包括泳动、蹭动和滑动等,其中陆生蓝藻的滑动运动在陆地环境中展现出独特的生理适应机制和生态功能[5].从生理层面来看,滑动运动使蓝藻能够主动迁移至适宜的微生境,有效应对陆地环境中资源分布不均和极端环境条件的挑战[6].在生态功能方面,荒漠蓝藻通过滑动过程分泌胞外聚合物(extracellular polymeric substances, EPS),这些物质不仅能够黏结固定沙砾,还能增加土壤有机质含量.特别值得注意的是,丝状蓝藻的藻丝在滑动过程中能够对沙砾进行物理性缠绕和捆绑,形成复杂的三维网状结构.这种独特的生物固定机制不仅显著提高了荒漠地表的稳定性,还促进了生物土壤结皮的形成,从而有效减少水土流失,增强土壤的持水能力[6-7].这种由蓝藻滑动驱动的生态过程对荒漠生态系统功能的维持和退化土地的生态修复具有关键作用[8].滑动行为使陆生蓝藻从“被动耐受”转变为“主动适应”环境,正是这种进化转变,使得陆生蓝藻成为陆地生态系统早期拓殖的先驱物种,并在现代生态修复实践中发挥着不可替代的重要角色. ...
Chinese studies on the edible blue green alga,Nostoc flagelliforme:a review
1
1998
... 蓝藻具有多种主动运动方式,包括泳动、蹭动和滑动等,其中陆生蓝藻的滑动运动在陆地环境中展现出独特的生理适应机制和生态功能[5].从生理层面来看,滑动运动使蓝藻能够主动迁移至适宜的微生境,有效应对陆地环境中资源分布不均和极端环境条件的挑战[6].在生态功能方面,荒漠蓝藻通过滑动过程分泌胞外聚合物(extracellular polymeric substances, EPS),这些物质不仅能够黏结固定沙砾,还能增加土壤有机质含量.特别值得注意的是,丝状蓝藻的藻丝在滑动过程中能够对沙砾进行物理性缠绕和捆绑,形成复杂的三维网状结构.这种独特的生物固定机制不仅显著提高了荒漠地表的稳定性,还促进了生物土壤结皮的形成,从而有效减少水土流失,增强土壤的持水能力[6-7].这种由蓝藻滑动驱动的生态过程对荒漠生态系统功能的维持和退化土地的生态修复具有关键作用[8].滑动行为使陆生蓝藻从“被动耐受”转变为“主动适应”环境,正是这种进化转变,使得陆生蓝藻成为陆地生态系统早期拓殖的先驱物种,并在现代生态修复实践中发挥着不可替代的重要角色. ...
Gliding motility in cyanobacteria:observations and possible explanations
1
2000
... 运动是细菌中广泛存在的行为,对其在自然环境中的生存适应具有十分重要的意义.大多数细菌通过鞭毛运动,但迄今尚未在蓝藻中发现这一细胞结构.与有鞭毛细菌的游动相比,无鞭毛的蓝藻也可以进行运动,但其运动速度相对较慢,通常不会超过11 µm·s-1[9]. ...
Do cyanobacteria swim using traveling surface waves
1
1996
... 运动能力在单细胞蓝藻和丝状蓝藻中都被发现,根据蓝藻主动运动方式的不同可将其分为两类:①泳动(Swimming motility),主要指某些海洋单细胞蓝藻在水中运动的形式,如Synechococcus sp. WH8102通过细胞外膜层收缩和扩张产生波动从而使细胞在水中运动[10].②滑动(Gliding motility),主要指某些蓝藻在固体表面相对平稳的运动形式,这种运动形式在单细胞和丝状蓝藻中均存在.值得注意的是,单细胞蓝藻的这种运动有时也被称为蹭动(Twitching motility),这种运动具有不连续、速度缓慢的特点,其运动机制与丝状蓝藻存在显著差异.研究表明,某些单细胞蓝藻(如Synechocystis sp. PCC6803)主要依赖Ⅳ型菌毛(type IV pili, T4P)的周期性伸展、栓系和收缩来完成位移,其在琼脂平板上的蹭动速度通常仅为1~2 µm·min-1[11].相比之下,丝状蓝藻的滑动过程则更为高效,其移动速度可达到10 µm·s-1[12],这可能是由于其独特的运动机制和丝状结构所带来的优势.这种运动速率的显著差异反映了不同形态蓝藻在运动机制上的进化适应性.本文仅讨论蓝藻的滑动现象. ...
Photomovement of the gliding cyanobacterium Synechocystis sp.PCC 6803
1
1999
... 运动能力在单细胞蓝藻和丝状蓝藻中都被发现,根据蓝藻主动运动方式的不同可将其分为两类:①泳动(Swimming motility),主要指某些海洋单细胞蓝藻在水中运动的形式,如Synechococcus sp. WH8102通过细胞外膜层收缩和扩张产生波动从而使细胞在水中运动[10].②滑动(Gliding motility),主要指某些蓝藻在固体表面相对平稳的运动形式,这种运动形式在单细胞和丝状蓝藻中均存在.值得注意的是,单细胞蓝藻的这种运动有时也被称为蹭动(Twitching motility),这种运动具有不连续、速度缓慢的特点,其运动机制与丝状蓝藻存在显著差异.研究表明,某些单细胞蓝藻(如Synechocystis sp. PCC6803)主要依赖Ⅳ型菌毛(type IV pili, T4P)的周期性伸展、栓系和收缩来完成位移,其在琼脂平板上的蹭动速度通常仅为1~2 µm·min-1[11].相比之下,丝状蓝藻的滑动过程则更为高效,其移动速度可达到10 µm·s-1[12],这可能是由于其独特的运动机制和丝状结构所带来的优势.这种运动速率的显著差异反映了不同形态蓝藻在运动机制上的进化适应性.本文仅讨论蓝藻的滑动现象. ...
A cyanobacterium capable of swimming motility
1
1985
... 运动能力在单细胞蓝藻和丝状蓝藻中都被发现,根据蓝藻主动运动方式的不同可将其分为两类:①泳动(Swimming motility),主要指某些海洋单细胞蓝藻在水中运动的形式,如Synechococcus sp. WH8102通过细胞外膜层收缩和扩张产生波动从而使细胞在水中运动[10].②滑动(Gliding motility),主要指某些蓝藻在固体表面相对平稳的运动形式,这种运动形式在单细胞和丝状蓝藻中均存在.值得注意的是,单细胞蓝藻的这种运动有时也被称为蹭动(Twitching motility),这种运动具有不连续、速度缓慢的特点,其运动机制与丝状蓝藻存在显著差异.研究表明,某些单细胞蓝藻(如Synechocystis sp. PCC6803)主要依赖Ⅳ型菌毛(type IV pili, T4P)的周期性伸展、栓系和收缩来完成位移,其在琼脂平板上的蹭动速度通常仅为1~2 µm·min-1[11].相比之下,丝状蓝藻的滑动过程则更为高效,其移动速度可达到10 µm·s-1[12],这可能是由于其独特的运动机制和丝状结构所带来的优势.这种运动速率的显著差异反映了不同形态蓝藻在运动机制上的进化适应性.本文仅讨论蓝藻的滑动现象. ...
Envelope structure of 4 gliding filamentous cyanobacteria
1
1995
... 蓝藻滑动机制当前有3种解释模型:①表层波动模型,颤藻属、鞘丝藻属和席藻属中的某些蓝藻细胞膜上存在呈螺旋状排列的纤维丝,这些纤维丝的节律伸缩可在细胞表层产生波动力,进而使细胞沿着与固体底物的接触面滑动[13].②黏液喷出模型,一些丝状蓝藻的细胞连接处存在孔状结构,这些孔状结构按一定角度呈环形排列于细胞连接处并穿过细胞膜、肽聚糖层及外膜,位于同一侧的孔状结构可以同时喷出脱水的EPS,并在水合时膨胀产生切向力,从而推动藻丝沿着底物表层向EPS喷出侧相反方向滑动[14-15].③Khayatan等[16]认为藻殖段细胞连接处的孔状结构由Ⅳ型菌毛合成及多糖分泌蛋白复合体构成,藻殖段运动依靠此复合体中的Ⅳ型菌毛伸长、锚定与收缩提供动力,菌毛与固体基质表层的锚定借助蓝藻所分泌的EPS来实现. ...
The junctional pore complex,a prokaryotic secretion organelle,is the molecular motor underlying gliding motility in cyanobacteria
2
1998
... 蓝藻滑动机制当前有3种解释模型:①表层波动模型,颤藻属、鞘丝藻属和席藻属中的某些蓝藻细胞膜上存在呈螺旋状排列的纤维丝,这些纤维丝的节律伸缩可在细胞表层产生波动力,进而使细胞沿着与固体底物的接触面滑动[13].②黏液喷出模型,一些丝状蓝藻的细胞连接处存在孔状结构,这些孔状结构按一定角度呈环形排列于细胞连接处并穿过细胞膜、肽聚糖层及外膜,位于同一侧的孔状结构可以同时喷出脱水的EPS,并在水合时膨胀产生切向力,从而推动藻丝沿着底物表层向EPS喷出侧相反方向滑动[14-15].③Khayatan等[16]认为藻殖段细胞连接处的孔状结构由Ⅳ型菌毛合成及多糖分泌蛋白复合体构成,藻殖段运动依靠此复合体中的Ⅳ型菌毛伸长、锚定与收缩提供动力,菌毛与固体基质表层的锚定借助蓝藻所分泌的EPS来实现. ...
... EPS在蓝藻的滑动过程中具有多种功能.蓝藻滑动时,分泌的EPS通常可覆盖整个藻丝表面,但不随藻丝进行位移,以塌陷的、有时扭曲的残留物形式被遗弃在前移藻丝末端后方[14].蓝藻细胞在滑动时倾向于使用其他蓝藻细胞滑动遗留的轨迹进行运动[18],原因可能是残留EPS的存在减少或免去了跟随细胞需分泌的EPS量,使得该细胞在被跟随细胞轨迹上的滑动过程更为经济.EPS还可提供有关近期存在于附近位置的细胞浓度的信息,类似于化学群体感应,因此,随着蓝藻群体的发展,EPS 踪迹能够提供一种持久的远程间接通信机制,可以引导滞后细胞与前端群体合并[21].此外,EPS还能通过改变细胞表面和周围光的折射率来影响细胞滑动[1].当前已知影响蓝藻EPS合成及分泌的主要因素包括NaCl含量、气体流通速率、温度、光强、pH值、重金属,以及氮、磷、硫、镁、钙、钾等元素的含量等[20-24]. ...
A model of filamentous cyanobacteria leading to reticulate pattern formation
1
2014
... 蓝藻滑动机制当前有3种解释模型:①表层波动模型,颤藻属、鞘丝藻属和席藻属中的某些蓝藻细胞膜上存在呈螺旋状排列的纤维丝,这些纤维丝的节律伸缩可在细胞表层产生波动力,进而使细胞沿着与固体底物的接触面滑动[13].②黏液喷出模型,一些丝状蓝藻的细胞连接处存在孔状结构,这些孔状结构按一定角度呈环形排列于细胞连接处并穿过细胞膜、肽聚糖层及外膜,位于同一侧的孔状结构可以同时喷出脱水的EPS,并在水合时膨胀产生切向力,从而推动藻丝沿着底物表层向EPS喷出侧相反方向滑动[14-15].③Khayatan等[16]认为藻殖段细胞连接处的孔状结构由Ⅳ型菌毛合成及多糖分泌蛋白复合体构成,藻殖段运动依靠此复合体中的Ⅳ型菌毛伸长、锚定与收缩提供动力,菌毛与固体基质表层的锚定借助蓝藻所分泌的EPS来实现. ...
Evidence that a modified type IV pilus-like system powers gliding motility and polysaccharide secretion in filamentous cyanobacteria
2
2015
... 蓝藻滑动机制当前有3种解释模型:①表层波动模型,颤藻属、鞘丝藻属和席藻属中的某些蓝藻细胞膜上存在呈螺旋状排列的纤维丝,这些纤维丝的节律伸缩可在细胞表层产生波动力,进而使细胞沿着与固体底物的接触面滑动[13].②黏液喷出模型,一些丝状蓝藻的细胞连接处存在孔状结构,这些孔状结构按一定角度呈环形排列于细胞连接处并穿过细胞膜、肽聚糖层及外膜,位于同一侧的孔状结构可以同时喷出脱水的EPS,并在水合时膨胀产生切向力,从而推动藻丝沿着底物表层向EPS喷出侧相反方向滑动[14-15].③Khayatan等[16]认为藻殖段细胞连接处的孔状结构由Ⅳ型菌毛合成及多糖分泌蛋白复合体构成,藻殖段运动依靠此复合体中的Ⅳ型菌毛伸长、锚定与收缩提供动力,菌毛与固体基质表层的锚定借助蓝藻所分泌的EPS来实现. ...
... 单细胞蓝藻和丝状蓝藻感应光和响应光的方式不同.一些研究者认为,Synechocystis sp. PCC 6803能直接感知光源方向而并非光强度的空间梯度,因为其细胞能充当球形微透镜,使细胞“看到”光源并向其移动:单侧光透过细胞一侧后会汇聚于细胞背光侧表层并形成明亮光斑,触发细胞向远离聚焦点的方向移动,因此球形蓝藻可能是世界上最小、最古老的眼球和照相机镜头的原型[16,45-47].此外,在自然环境中,虽然光强度梯度很少陡峭到在几微米的单细胞尺度上产生显著差异[47],但在结构上具有一定长度的颤藻属和席藻属等丝状蓝藻展现出可感知此类光梯度并进行滑动响应的特征.如Phormidium uncinatum藻丝似乎会比较头部和尾部的入射光,如果尾部比头部接收到更多的光,藻丝很可能会改变滑动方向,且其对光非常敏感,两端接收到的光强之间的差异只要高于4%即可引发转向,而照射到藻丝中部的光线则没有这种影响;还有一些蓝藻如Anabaena variabilis可通过藻丝的弯曲和转向来响应光场的不均匀性[44]. ...
Information integration and collective motility in phototactic cyanobacteria
4
2020
... 蓝藻在滑动时通常伴随着EPS的分泌[17-18],EPS是蓝藻合成的重要物质,根据溶解度和黏性的不同可分为鞘、荚膜和黏液等3种类型,其主要成分为高黏度的糖类聚合物,其中常见的占比最高的单糖组分为葡萄糖[19].EPS单糖组分的具体构成因藻种和环境条件的不同而改变,从而表现出不同的物理化学特性,因此它也是结构和功能复杂的蓝藻胞外物质[20]. ...
... T4P 的物理连接和EPS轨迹的效应导致的群落中细胞间相互作用或可解释某些蓝藻(例如Synechocystis sp.)趋光运动的集体行为[17-21,62].蓝藻细胞的趋光性运动依赖于多个关键基因的协同作用.研究表明,参与菌毛生物合成和功能的运动相关基因对趋光性至关重要,同时调控环磷酸腺苷(Cyclic Adenosine Monophosphate,cAMP)信号通路的基因也发挥着重要作用.cAMP作为从原核生物到高等生物中保守的第二信使,由腺苷酸环化酶催化ATP生成,在细胞信号转导中具有广泛功能[63].特别值得注意的是,当缺失cya1(腺苷酸环化酶)和sycrp1(一种cAMP受体样蛋白)基因时,突变体菌落丧失趋光性能力;然而有趣的是,当这些突变体细胞与正常趋光性细胞共培养时,它们能够通过“搭便车”机制参与群体趋光运动.这种独特的群体行为源于T4P介导的细胞间相互作用:突变细胞通过Ⅳ型菌毛依附于具有光感应能力的细胞,并借助邻近细胞施加的机械力实现定向移动[18].此外,蓝藻群体趋光运动还表现出显著的协同效应.细胞滑动过程中分泌的EPS具有双重功能:一方面降低细胞运动的表面摩擦力,另一方面形成引导路径促进后续细胞的定向移动.这种正反馈机制使得群体迁移速度显著高于单个细胞,其动力学过程类似于窗户上雨滴的移动模式——后续水滴沿既有路径更快移动并与前导水滴融合,最终形成更大的水滴.这种基于EPS引导的集体运动机制,与T4P介导的细胞间力学耦合共同构成了蓝藻群体趋光性的核心基础[17]. ...
... [17]. ...
... 综上所述,一些蓝藻能根据光的波长和强度不断调整其滑动过程,在细胞层面上实现对光输入的精细整合.这一过程需要 T4P和类似光敏色素的光感受器与复杂的信号转导途径相耦合[52],从而灵敏地控制滑动的方向和速度,但细胞如何整合所有光感受器输入信号的分子机制尚不完全清楚.在宏观尺度上,菌落的集体行为模式不仅依赖于单个细胞的运动行为,还受到细胞间相互作用的影响[17,50]. ...
Phototaxis as a collective phenomenon in cyanobacterial colonies
4
2017
... 蓝藻在滑动时通常伴随着EPS的分泌[17-18],EPS是蓝藻合成的重要物质,根据溶解度和黏性的不同可分为鞘、荚膜和黏液等3种类型,其主要成分为高黏度的糖类聚合物,其中常见的占比最高的单糖组分为葡萄糖[19].EPS单糖组分的具体构成因藻种和环境条件的不同而改变,从而表现出不同的物理化学特性,因此它也是结构和功能复杂的蓝藻胞外物质[20]. ...
... EPS在蓝藻的滑动过程中具有多种功能.蓝藻滑动时,分泌的EPS通常可覆盖整个藻丝表面,但不随藻丝进行位移,以塌陷的、有时扭曲的残留物形式被遗弃在前移藻丝末端后方[14].蓝藻细胞在滑动时倾向于使用其他蓝藻细胞滑动遗留的轨迹进行运动[18],原因可能是残留EPS的存在减少或免去了跟随细胞需分泌的EPS量,使得该细胞在被跟随细胞轨迹上的滑动过程更为经济.EPS还可提供有关近期存在于附近位置的细胞浓度的信息,类似于化学群体感应,因此,随着蓝藻群体的发展,EPS 踪迹能够提供一种持久的远程间接通信机制,可以引导滞后细胞与前端群体合并[21].此外,EPS还能通过改变细胞表面和周围光的折射率来影响细胞滑动[1].当前已知影响蓝藻EPS合成及分泌的主要因素包括NaCl含量、气体流通速率、温度、光强、pH值、重金属,以及氮、磷、硫、镁、钙、钾等元素的含量等[20-24]. ...
... 趋光性是一种相对于光场方向的定向运动,即单个细胞或细胞群响应光强、光谱进行运动,从而移动到更有利于生存和生长的光条件下[42].蓝藻是光合自养生物,对光照的要求很严格,光照太弱会导致能量产生不足,在某些物种中可能会导致细胞进行异养呼吸;光照太强会降低光合作用效率,并导致光损伤,紫外线辐射对某些无直接光保护响应机制的蓝藻尤其致命,因此,光的波长、强度以及蓝藻种类都会影响蓝藻细胞对光的响应[18,43-44]. ...
... T4P 的物理连接和EPS轨迹的效应导致的群落中细胞间相互作用或可解释某些蓝藻(例如Synechocystis sp.)趋光运动的集体行为[17-21,62].蓝藻细胞的趋光性运动依赖于多个关键基因的协同作用.研究表明,参与菌毛生物合成和功能的运动相关基因对趋光性至关重要,同时调控环磷酸腺苷(Cyclic Adenosine Monophosphate,cAMP)信号通路的基因也发挥着重要作用.cAMP作为从原核生物到高等生物中保守的第二信使,由腺苷酸环化酶催化ATP生成,在细胞信号转导中具有广泛功能[63].特别值得注意的是,当缺失cya1(腺苷酸环化酶)和sycrp1(一种cAMP受体样蛋白)基因时,突变体菌落丧失趋光性能力;然而有趣的是,当这些突变体细胞与正常趋光性细胞共培养时,它们能够通过“搭便车”机制参与群体趋光运动.这种独特的群体行为源于T4P介导的细胞间相互作用:突变细胞通过Ⅳ型菌毛依附于具有光感应能力的细胞,并借助邻近细胞施加的机械力实现定向移动[18].此外,蓝藻群体趋光运动还表现出显著的协同效应.细胞滑动过程中分泌的EPS具有双重功能:一方面降低细胞运动的表面摩擦力,另一方面形成引导路径促进后续细胞的定向移动.这种正反馈机制使得群体迁移速度显著高于单个细胞,其动力学过程类似于窗户上雨滴的移动模式——后续水滴沿既有路径更快移动并与前导水滴融合,最终形成更大的水滴.这种基于EPS引导的集体运动机制,与T4P介导的细胞间力学耦合共同构成了蓝藻群体趋光性的核心基础[17]. ...
Insights into the role of external layers in cyanobacteria at varying temperatures and their regulatory mechanism
6
2022
... 蓝藻在滑动时通常伴随着EPS的分泌[17-18],EPS是蓝藻合成的重要物质,根据溶解度和黏性的不同可分为鞘、荚膜和黏液等3种类型,其主要成分为高黏度的糖类聚合物,其中常见的占比最高的单糖组分为葡萄糖[19].EPS单糖组分的具体构成因藻种和环境条件的不同而改变,从而表现出不同的物理化学特性,因此它也是结构和功能复杂的蓝藻胞外物质[20]. ...
... 蓝藻分泌的EPS具有缓冲温度变化的功能,能够应对短期内的温度波动,从而通过维持细胞活动的稳定性来提高光合作用和呼吸作用的效率 [67].鉴于蓝藻细胞外层结构(包括鞘、荚膜和黏液层)[56]的包裹,研究者提出了“球壳热传导模型”,该模型认为增加EPS的厚度可以降低细胞与环境之间的导热率,从而减缓细胞内温度的变化,最终减少环境温度波动对细胞生理活动的影响.同时EPS可增强细胞间的连接,使其形成更稳定的群体结构,有助于群体中的蓝藻细胞在环境变化时相互保护和支持[19,67-69]. ...
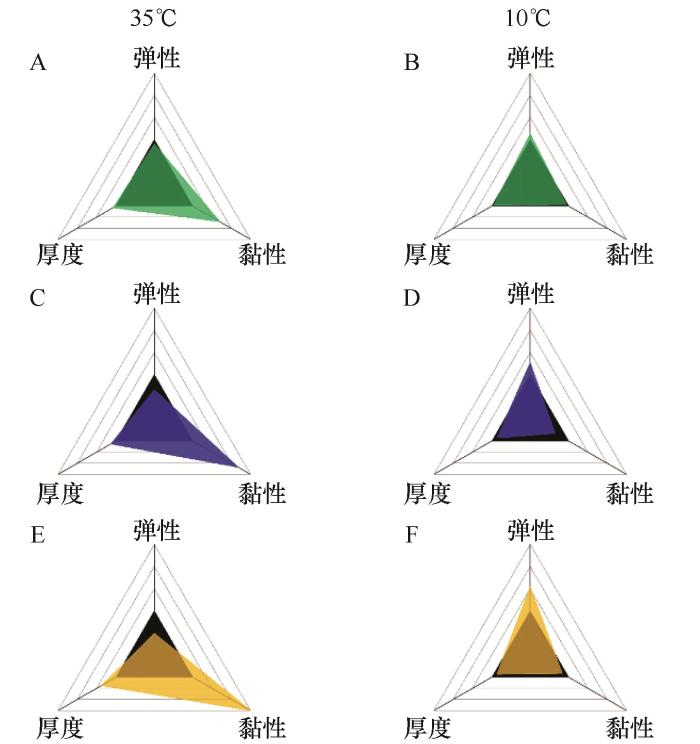
... 温度变化可显著改变蓝藻EPS的理化性质,而EPS量及其成分与特性的改变都可能对蓝藻的滑动特征产生影响[70].研究者将Nostoc sp.(FACHB-2009)、Dolichospermum sp.(FACHB-82)和Microcystis sp.(FACHB-3296)3种蓝藻菌株置于3种温度(10、25、35 ℃)下培养,发现在10 ℃和35 ℃时,单位生物量的EPS含量相对较高,表明蓝藻在相对极端温度条件下倾向于分泌更多EPS以保护细胞.在高温环境中,菌落的尺寸增大,EPS变得更厚、更松散且黏性更强(图3).此时未观察到菌落中单个细胞尺寸有显著增加,因此导致菌落尺寸增大的核心因素可能是菌落中细胞数量增多.EPS黏性的增加有助于菌落结构的形成和维持,并可能有助于蓝藻吸收营养物质,同时也会增加单个细胞滑动时的阻力[19]. ...
... [
19]
注:图A、B表示Nostoc sp.(FACHB-2009)的鞘,图C、D表示Dolichospermum sp.(FACHB-82)的荚膜,图E、F表示Microcystis sp.(FACHB-3296)的黏液.黑色实心三角形表示在25 ℃下每种EPS的生物物理特性作为比对基准,彩色三角形表示10 ℃和35 ℃下EPS物理特性的变化程度[19] ...
... 注:图A、B表示Nostoc sp.(FACHB-2009)的鞘,图C、D表示Dolichospermum sp.(FACHB-82)的荚膜,图E、F表示Microcystis sp.(FACHB-3296)的黏液.黑色实心三角形表示在25 ℃下每种EPS的生物物理特性作为比对基准,彩色三角形表示10 ℃和35 ℃下EPS物理特性的变化程度[19] ...
... [
19]
Fig.3![]() <strong>2.4</strong> <strong>CO<sub>2</sub></strong> 和<strong>O<sub>2</sub></strong>
<strong>2.4</strong> <strong>CO<sub>2</sub></strong> 和<strong>O<sub>2</sub></strong>作为产氧性光自养生物,蓝藻具有向适宜CO2浓度区域迁移的趋化行为,这对维持其光合作用效率至关重要.当环境中CO2长期匮乏时,蓝藻细胞会面临双重生理挑战:一方面,光合色素和类囊体膜会发生降解;另一方面,细胞内超氧化物离子会过量积累,导致氧化应激.同时,O2对蓝藻的生理活动同样不可或缺,它不仅是有氧呼吸的必要底物,还参与某些不饱和脂肪酸的生物合成过程[36-37]. ...
Complexity of cyanobacterial exopolysaccharides:composition,structures,inducing factors and putative genes involved in their biosynthesis and assembly
2
2009
... 蓝藻在滑动时通常伴随着EPS的分泌[17-18],EPS是蓝藻合成的重要物质,根据溶解度和黏性的不同可分为鞘、荚膜和黏液等3种类型,其主要成分为高黏度的糖类聚合物,其中常见的占比最高的单糖组分为葡萄糖[19].EPS单糖组分的具体构成因藻种和环境条件的不同而改变,从而表现出不同的物理化学特性,因此它也是结构和功能复杂的蓝藻胞外物质[20]. ...
... EPS在蓝藻的滑动过程中具有多种功能.蓝藻滑动时,分泌的EPS通常可覆盖整个藻丝表面,但不随藻丝进行位移,以塌陷的、有时扭曲的残留物形式被遗弃在前移藻丝末端后方[14].蓝藻细胞在滑动时倾向于使用其他蓝藻细胞滑动遗留的轨迹进行运动[18],原因可能是残留EPS的存在减少或免去了跟随细胞需分泌的EPS量,使得该细胞在被跟随细胞轨迹上的滑动过程更为经济.EPS还可提供有关近期存在于附近位置的细胞浓度的信息,类似于化学群体感应,因此,随着蓝藻群体的发展,EPS 踪迹能够提供一种持久的远程间接通信机制,可以引导滞后细胞与前端群体合并[21].此外,EPS还能通过改变细胞表面和周围光的折射率来影响细胞滑动[1].当前已知影响蓝藻EPS合成及分泌的主要因素包括NaCl含量、气体流通速率、温度、光强、pH值、重金属,以及氮、磷、硫、镁、钙、钾等元素的含量等[20-24]. ...
Motility enhancement through surface modification is sufficient for cyanobacterial community organization during phototaxis
2
2013
... EPS在蓝藻的滑动过程中具有多种功能.蓝藻滑动时,分泌的EPS通常可覆盖整个藻丝表面,但不随藻丝进行位移,以塌陷的、有时扭曲的残留物形式被遗弃在前移藻丝末端后方[14].蓝藻细胞在滑动时倾向于使用其他蓝藻细胞滑动遗留的轨迹进行运动[18],原因可能是残留EPS的存在减少或免去了跟随细胞需分泌的EPS量,使得该细胞在被跟随细胞轨迹上的滑动过程更为经济.EPS还可提供有关近期存在于附近位置的细胞浓度的信息,类似于化学群体感应,因此,随着蓝藻群体的发展,EPS 踪迹能够提供一种持久的远程间接通信机制,可以引导滞后细胞与前端群体合并[21].此外,EPS还能通过改变细胞表面和周围光的折射率来影响细胞滑动[1].当前已知影响蓝藻EPS合成及分泌的主要因素包括NaCl含量、气体流通速率、温度、光强、pH值、重金属,以及氮、磷、硫、镁、钙、钾等元素的含量等[20-24]. ...
... T4P 的物理连接和EPS轨迹的效应导致的群落中细胞间相互作用或可解释某些蓝藻(例如Synechocystis sp.)趋光运动的集体行为[17-21,62].蓝藻细胞的趋光性运动依赖于多个关键基因的协同作用.研究表明,参与菌毛生物合成和功能的运动相关基因对趋光性至关重要,同时调控环磷酸腺苷(Cyclic Adenosine Monophosphate,cAMP)信号通路的基因也发挥着重要作用.cAMP作为从原核生物到高等生物中保守的第二信使,由腺苷酸环化酶催化ATP生成,在细胞信号转导中具有广泛功能[63].特别值得注意的是,当缺失cya1(腺苷酸环化酶)和sycrp1(一种cAMP受体样蛋白)基因时,突变体菌落丧失趋光性能力;然而有趣的是,当这些突变体细胞与正常趋光性细胞共培养时,它们能够通过“搭便车”机制参与群体趋光运动.这种独特的群体行为源于T4P介导的细胞间相互作用:突变细胞通过Ⅳ型菌毛依附于具有光感应能力的细胞,并借助邻近细胞施加的机械力实现定向移动[18].此外,蓝藻群体趋光运动还表现出显著的协同效应.细胞滑动过程中分泌的EPS具有双重功能:一方面降低细胞运动的表面摩擦力,另一方面形成引导路径促进后续细胞的定向移动.这种正反馈机制使得群体迁移速度显著高于单个细胞,其动力学过程类似于窗户上雨滴的移动模式——后续水滴沿既有路径更快移动并与前导水滴融合,最终形成更大的水滴.这种基于EPS引导的集体运动机制,与T4P介导的细胞间力学耦合共同构成了蓝藻群体趋光性的核心基础[17]. ...
Insights into extracellular polymeric substances of cyanobacterium Microcystis aeruginosa using fractionation procedure and parallel factor analysis
0
2013
Characteristics and bacterial community dynamics during extracellular polymeric substance (EPS) degradation of cyanobacterial blooms
0
2020
The Evolution of a capacity to build supra-cellular ropes enabled filamentous cyanobacteria to colonize highly erodible substrates
1
2009
... EPS在蓝藻的滑动过程中具有多种功能.蓝藻滑动时,分泌的EPS通常可覆盖整个藻丝表面,但不随藻丝进行位移,以塌陷的、有时扭曲的残留物形式被遗弃在前移藻丝末端后方[14].蓝藻细胞在滑动时倾向于使用其他蓝藻细胞滑动遗留的轨迹进行运动[18],原因可能是残留EPS的存在减少或免去了跟随细胞需分泌的EPS量,使得该细胞在被跟随细胞轨迹上的滑动过程更为经济.EPS还可提供有关近期存在于附近位置的细胞浓度的信息,类似于化学群体感应,因此,随着蓝藻群体的发展,EPS 踪迹能够提供一种持久的远程间接通信机制,可以引导滞后细胞与前端群体合并[21].此外,EPS还能通过改变细胞表面和周围光的折射率来影响细胞滑动[1].当前已知影响蓝藻EPS合成及分泌的主要因素包括NaCl含量、气体流通速率、温度、光强、pH值、重金属,以及氮、磷、硫、镁、钙、钾等元素的含量等[20-24]. ...
Cyanobacteria track water in desert soils
3
2001
... 蓝藻可在微生物席、生物土壤结皮等生物复合体结构中通过滑动进行运动,从而对包括水分[25-29]、光照[30-32]、昼夜节律[33-35]、气体浓度[36-37]、盐度[38]和营养物质浓度[39]在内的多种因素变化进行响应. ...
... 在干旱生境,例如由蓝藻等微生物类群构成的荒漠地表生物土壤结皮中,蓝藻可通过滑动响应可利用水量的变化[25-29,40].在对西班牙东北部沙漠地区的结皮研究中发现,干燥时大量颤藻目蓝藻在距离结皮表面1.5~2.1 mm的深度形成致密层.润湿几分钟后能明显观察到结皮表面变绿,显微镜观察表明,此现象是由结皮表面出现藻丝所致;而当样本逐渐干燥时,藻丝被观察到向结皮内部移动.上述过程在有无光照时均可发生,但将样本暴露在高光强下会减缓或阻止其表面返绿.这表明在此类结皮中,光并非蓝藻响应水含量变化进行滑动的前提条件,且过强的光照可干扰或抑制藻丝向表面滑动[25].向逐渐干燥的结皮样本中添加ATP合成抑制剂或质子动力解偶联剂可抑制藻丝向下滑动,表明蓝藻向下滑动是主动响应的过程,而并非是由空气-水界面的表面张力等物理效应所致,且需要底物磷酸化以外的能量来源[26].而且从完全湿润开始逐渐干燥时,生物结皮内部会形成随深度变化的可用水梯度,即表面含水量低,较深处含水量高[40].因此,推断结皮内蓝藻的滑动是其对水环境变化的运动响应,提出了“趋水性”假说,即蓝藻细胞可以通过某种途径感知环境中可用水的短期变化,或者通过某种传感器感知环境中随含水量变化的参数,如盐度、温度或其他化学物质浓度.上述结皮样本若长时间保持湿润,表面将形成致密的主要由颤藻目蓝藻构成的生物膜,此时若进行干燥,部分蓝藻会被困在干燥的结皮表面[26].为了能够感知到可用水变化,颤藻藻丝在结皮内部的排布方向可能会有一定的垂直分量,在降雨或开始干燥时,部分蓝藻能够快速获得有关环境含水量的信息,使整根藻丝滑动到含水量较高的区域.而在足够长的水分暴露时间后,许多蓝藻藻丝已经完全到达结皮表面并在其上水平滑动,这将使得蓝藻无法有效感知藻丝与其滑动轴正交方向的含水量梯度,从而停留在结皮表面. ...
... [25].向逐渐干燥的结皮样本中添加ATP合成抑制剂或质子动力解偶联剂可抑制藻丝向下滑动,表明蓝藻向下滑动是主动响应的过程,而并非是由空气-水界面的表面张力等物理效应所致,且需要底物磷酸化以外的能量来源[26].而且从完全湿润开始逐渐干燥时,生物结皮内部会形成随深度变化的可用水梯度,即表面含水量低,较深处含水量高[40].因此,推断结皮内蓝藻的滑动是其对水环境变化的运动响应,提出了“趋水性”假说,即蓝藻细胞可以通过某种途径感知环境中可用水的短期变化,或者通过某种传感器感知环境中随含水量变化的参数,如盐度、温度或其他化学物质浓度.上述结皮样本若长时间保持湿润,表面将形成致密的主要由颤藻目蓝藻构成的生物膜,此时若进行干燥,部分蓝藻会被困在干燥的结皮表面[26].为了能够感知到可用水变化,颤藻藻丝在结皮内部的排布方向可能会有一定的垂直分量,在降雨或开始干燥时,部分蓝藻能够快速获得有关环境含水量的信息,使整根藻丝滑动到含水量较高的区域.而在足够长的水分暴露时间后,许多蓝藻藻丝已经完全到达结皮表面并在其上水平滑动,这将使得蓝藻无法有效感知藻丝与其滑动轴正交方向的含水量梯度,从而停留在结皮表面. ...
Hydrotaxis of cyanobacteria in desert crusts
3
2004
... 在干旱生境,例如由蓝藻等微生物类群构成的荒漠地表生物土壤结皮中,蓝藻可通过滑动响应可利用水量的变化[25-29,40].在对西班牙东北部沙漠地区的结皮研究中发现,干燥时大量颤藻目蓝藻在距离结皮表面1.5~2.1 mm的深度形成致密层.润湿几分钟后能明显观察到结皮表面变绿,显微镜观察表明,此现象是由结皮表面出现藻丝所致;而当样本逐渐干燥时,藻丝被观察到向结皮内部移动.上述过程在有无光照时均可发生,但将样本暴露在高光强下会减缓或阻止其表面返绿.这表明在此类结皮中,光并非蓝藻响应水含量变化进行滑动的前提条件,且过强的光照可干扰或抑制藻丝向表面滑动[25].向逐渐干燥的结皮样本中添加ATP合成抑制剂或质子动力解偶联剂可抑制藻丝向下滑动,表明蓝藻向下滑动是主动响应的过程,而并非是由空气-水界面的表面张力等物理效应所致,且需要底物磷酸化以外的能量来源[26].而且从完全湿润开始逐渐干燥时,生物结皮内部会形成随深度变化的可用水梯度,即表面含水量低,较深处含水量高[40].因此,推断结皮内蓝藻的滑动是其对水环境变化的运动响应,提出了“趋水性”假说,即蓝藻细胞可以通过某种途径感知环境中可用水的短期变化,或者通过某种传感器感知环境中随含水量变化的参数,如盐度、温度或其他化学物质浓度.上述结皮样本若长时间保持湿润,表面将形成致密的主要由颤藻目蓝藻构成的生物膜,此时若进行干燥,部分蓝藻会被困在干燥的结皮表面[26].为了能够感知到可用水变化,颤藻藻丝在结皮内部的排布方向可能会有一定的垂直分量,在降雨或开始干燥时,部分蓝藻能够快速获得有关环境含水量的信息,使整根藻丝滑动到含水量较高的区域.而在足够长的水分暴露时间后,许多蓝藻藻丝已经完全到达结皮表面并在其上水平滑动,这将使得蓝藻无法有效感知藻丝与其滑动轴正交方向的含水量梯度,从而停留在结皮表面. ...
... [26].为了能够感知到可用水变化,颤藻藻丝在结皮内部的排布方向可能会有一定的垂直分量,在降雨或开始干燥时,部分蓝藻能够快速获得有关环境含水量的信息,使整根藻丝滑动到含水量较高的区域.而在足够长的水分暴露时间后,许多蓝藻藻丝已经完全到达结皮表面并在其上水平滑动,这将使得蓝藻无法有效感知藻丝与其滑动轴正交方向的含水量梯度,从而停留在结皮表面. ...
... 紫外线,尤其是波长在280~315 nm的UV-B,对产氧型光自养生物具有负面影响[57].紫外线会抑制光养生物的生长和光合作用、导致突变和急性生理应激,甚至细胞死亡[58-59].运动是蓝藻避免受到来自较高紫外线剂量的负面影响的一种生存策略,某些微鞘藻(例如Microcoleus chthonoplastes)能通过垂直滑动来免受UV-B的照射,位于生物席表面下方300 µm处的蓝藻所受到的UV-B辐照度仅为表面的10%[30].UV-A(320~400 nm)辐射可诱导某些蓝藻合成伪枝藻素等防晒物质,伪枝藻素能有效吸收紫外辐射,从而减少紫外线对细胞的损害,这种防晒功能使得这些蓝藻能够在较高紫外线辐射的环境中,例如在荒漠生物结皮表面长期生存[60],而不具备此类保护机制的微鞘藻及颤藻则一般存在于相似环境下此类生物复合体内部,仅在伴随着光照强度降低的降水事件等发生时向生物复合体表面滑动[26,36]. ...
Effect of water potential on a Microcoleus (cyanophyceae) from a desert crust
0
1975
The involvement of type IV pili and phytochrome CphA in gliding motility,lateral motility and photophobotaxis of the cyanobacterium Phormidium lacuna
1
2021
... 部分陆生蓝藻在干燥脱水时会进入休眠状态,在有可利用水时才进行活跃的生命活动,并通过光合作用进行生长.而自然条件下,发生降雨事件时通常伴随着光照强度的降低,降雨结束后光照强度逐渐增加,过强的光照会对此时仍留在地表且缺乏光保护色素的蓝藻细胞造成光损伤[41].因此,这种追踪水的行为可能优化了蓝藻在生物结皮中的位置,从而使得缺乏光保护色素的细胞在满足光照要求的同时,避免受到高光强或紫外线等辐照的伤害;其次,蓝藻通过滑动保持细胞周围环境中有一定量的可用水,可以延长其对脱水过程的响应时间,使其更充分地对细胞进行保护以应对即将到来的干旱期,例如通过积累海藻糖和蔗糖等相容性溶质进行渗透保护,或产生EPS减缓细胞水分流失速度,因此有助于蓝藻在干旱环境中的生存[28-29]. ...
Dynamic cyanobacterial response to hydration and dehydration in a desert biological soil crust
3
2013
... 蓝藻可在微生物席、生物土壤结皮等生物复合体结构中通过滑动进行运动,从而对包括水分[25-29]、光照[30-32]、昼夜节律[33-35]、气体浓度[36-37]、盐度[38]和营养物质浓度[39]在内的多种因素变化进行响应. ...
... 在干旱生境,例如由蓝藻等微生物类群构成的荒漠地表生物土壤结皮中,蓝藻可通过滑动响应可利用水量的变化[25-29,40].在对西班牙东北部沙漠地区的结皮研究中发现,干燥时大量颤藻目蓝藻在距离结皮表面1.5~2.1 mm的深度形成致密层.润湿几分钟后能明显观察到结皮表面变绿,显微镜观察表明,此现象是由结皮表面出现藻丝所致;而当样本逐渐干燥时,藻丝被观察到向结皮内部移动.上述过程在有无光照时均可发生,但将样本暴露在高光强下会减缓或阻止其表面返绿.这表明在此类结皮中,光并非蓝藻响应水含量变化进行滑动的前提条件,且过强的光照可干扰或抑制藻丝向表面滑动[25].向逐渐干燥的结皮样本中添加ATP合成抑制剂或质子动力解偶联剂可抑制藻丝向下滑动,表明蓝藻向下滑动是主动响应的过程,而并非是由空气-水界面的表面张力等物理效应所致,且需要底物磷酸化以外的能量来源[26].而且从完全湿润开始逐渐干燥时,生物结皮内部会形成随深度变化的可用水梯度,即表面含水量低,较深处含水量高[40].因此,推断结皮内蓝藻的滑动是其对水环境变化的运动响应,提出了“趋水性”假说,即蓝藻细胞可以通过某种途径感知环境中可用水的短期变化,或者通过某种传感器感知环境中随含水量变化的参数,如盐度、温度或其他化学物质浓度.上述结皮样本若长时间保持湿润,表面将形成致密的主要由颤藻目蓝藻构成的生物膜,此时若进行干燥,部分蓝藻会被困在干燥的结皮表面[26].为了能够感知到可用水变化,颤藻藻丝在结皮内部的排布方向可能会有一定的垂直分量,在降雨或开始干燥时,部分蓝藻能够快速获得有关环境含水量的信息,使整根藻丝滑动到含水量较高的区域.而在足够长的水分暴露时间后,许多蓝藻藻丝已经完全到达结皮表面并在其上水平滑动,这将使得蓝藻无法有效感知藻丝与其滑动轴正交方向的含水量梯度,从而停留在结皮表面. ...
... 部分陆生蓝藻在干燥脱水时会进入休眠状态,在有可利用水时才进行活跃的生命活动,并通过光合作用进行生长.而自然条件下,发生降雨事件时通常伴随着光照强度的降低,降雨结束后光照强度逐渐增加,过强的光照会对此时仍留在地表且缺乏光保护色素的蓝藻细胞造成光损伤[41].因此,这种追踪水的行为可能优化了蓝藻在生物结皮中的位置,从而使得缺乏光保护色素的细胞在满足光照要求的同时,避免受到高光强或紫外线等辐照的伤害;其次,蓝藻通过滑动保持细胞周围环境中有一定量的可用水,可以延长其对脱水过程的响应时间,使其更充分地对细胞进行保护以应对即将到来的干旱期,例如通过积累海藻糖和蔗糖等相容性溶质进行渗透保护,或产生EPS减缓细胞水分流失速度,因此有助于蓝藻在干旱环境中的生存[28-29]. ...
UV B-induced vertical migrations of cyanobacteria in a microbial mat
2
1995
... 蓝藻可在微生物席、生物土壤结皮等生物复合体结构中通过滑动进行运动,从而对包括水分[25-29]、光照[30-32]、昼夜节律[33-35]、气体浓度[36-37]、盐度[38]和营养物质浓度[39]在内的多种因素变化进行响应. ...
... 紫外线,尤其是波长在280~315 nm的UV-B,对产氧型光自养生物具有负面影响[57].紫外线会抑制光养生物的生长和光合作用、导致突变和急性生理应激,甚至细胞死亡[58-59].运动是蓝藻避免受到来自较高紫外线剂量的负面影响的一种生存策略,某些微鞘藻(例如Microcoleus chthonoplastes)能通过垂直滑动来免受UV-B的照射,位于生物席表面下方300 µm处的蓝藻所受到的UV-B辐照度仅为表面的10%[30].UV-A(320~400 nm)辐射可诱导某些蓝藻合成伪枝藻素等防晒物质,伪枝藻素能有效吸收紫外辐射,从而减少紫外线对细胞的损害,这种防晒功能使得这些蓝藻能够在较高紫外线辐射的环境中,例如在荒漠生物结皮表面长期生存[60],而不具备此类保护机制的微鞘藻及颤藻则一般存在于相似环境下此类生物复合体内部,仅在伴随着光照强度降低的降水事件等发生时向生物复合体表面滑动[26,36]. ...
The effect of solar UV and visible irradiance on the vertical movements of cyanobacteria in microbial mats of hypersaline waters
2
1998
... 微生物席(Microbial mats)也可称微生物垫或菌毯,是沉积物基质中生活的各种细菌、古菌和真核微藻的密集底栖群落与其构成的生物复合体,蓝藻是生物席中主要的初级生产者[4].由于沉积物基质中光的急剧衰减和物质传递速率的限制,微生物席的特点是拥有氧气、光强及光谱组成、pH值和硫化物浓度等的陡峭垂直梯度[31].蓝藻可以通过滑动来调整自身在微生物席内不断变化的光场中的位置,从而处于具有最佳光照强度的区域,避免光照不足或受到过强辐射的负面影响[52-54].Lichtenberg等[54]于2020年对在丹麦利姆峡湾定期干燥的沙地上收集的微生物席研究中发现,弱光下蓝藻出现在生物席表面;而在高光强下,蓝藻出现在较深层,说明蓝藻可以通过响应光照强度进行垂直滑动来调整其在生物席中的位置,并且蓝藻在高光强下是负趋光的,即具有避光性,而在低光强下则具有正趋光性.这种特性在螺旋藻(Spirulina)和颤藻(Oscillatoria)中也有发现[31].北威尔士梅奈海峡安格尔西海岸的沿岸泥滩中生活着大量微鞘藻属蓝藻(Microcoleus),泥滩表面在阴天所呈现的绿色较之于晴天时更为明显,说明泥滩中的大多数蓝藻在弱光时具有趋光性,在高光强下具有避光性,当前还尚未确定此类微鞘藻改变趋光向性的阈值光强.上述微鞘藻藻丝在黑暗时也会移动到泥滩表面,但在到达表面后,藻丝在有无光时所形成的图案有所不同:黑暗时藻丝在泥滩表面呈螺旋状,而有光照时藻丝长轴方向与光源方向平行,若此时光源方向改变 90°,藻丝则会重新定向.这说明不只是单个细胞,细胞群落也会集体响应光照条件[36,55]. ...
... [31].北威尔士梅奈海峡安格尔西海岸的沿岸泥滩中生活着大量微鞘藻属蓝藻(Microcoleus),泥滩表面在阴天所呈现的绿色较之于晴天时更为明显,说明泥滩中的大多数蓝藻在弱光时具有趋光性,在高光强下具有避光性,当前还尚未确定此类微鞘藻改变趋光向性的阈值光强.上述微鞘藻藻丝在黑暗时也会移动到泥滩表面,但在到达表面后,藻丝在有无光时所形成的图案有所不同:黑暗时藻丝在泥滩表面呈螺旋状,而有光照时藻丝长轴方向与光源方向平行,若此时光源方向改变 90°,藻丝则会重新定向.这说明不只是单个细胞,细胞群落也会集体响应光照条件[36,55]. ...
Differential phototactic behavior of closely related cyanobacterial isolates from Yellowstone hot spring biofilms
4
2022
... 蓝藻可在微生物席、生物土壤结皮等生物复合体结构中通过滑动进行运动,从而对包括水分[25-29]、光照[30-32]、昼夜节律[33-35]、气体浓度[36-37]、盐度[38]和营养物质浓度[39]在内的多种因素变化进行响应. ...
... 在琼脂基质的趋光性实验中,集胞藻(Synechocystis)表现出独特的群体运动行为.单个集胞藻细胞的运动能力有限,但当细胞通过增殖和聚集达到特定群体规模阈值时,其运动能力显著增强并展现出明显的趋光性.这种群体行为表现为菌落朝向光源方向形成具有规律间隔的手指状突起(图1)[56].有趣的是,当通过改变入射光的方向将一根手指状突起重新定向到另一根“手指”的轨迹中时,细胞的运动速度会立即加快,这种现象可能与细胞运动过程中分泌的EPS的导向作用有关.在一定参数范围内,光强和细胞密度对菌落形态具有显著影响.当光强增加或细胞密度提高时,菌落前缘会形成更为均匀的冠状突起而非手指状结构[32].这一形态转变可能与光场均匀度的改变密切相关:更强的光照和更高的细胞密度都会在一定范围内提高光场的均匀性,从而影响细胞的群体运动模式.这些发现为理解蓝藻群体行为的调控机制提供了新的视角. ...
... 集胞藻属蓝藻Synechocystis sp. PCC6803表现出依赖入射光波长的趋光性,此过程通过多种光感受器的作用来实现[32,50].Synechocystis sp. PCC6803菌株偏向于朝绿光(535 nm)或红光(660 nm)光源方向移动,即细胞表现出趋光性,这种运动偏向性随着光通量的增加而增强,并逐渐产生手指状突起,但进行运动的细胞数量的比例和运动速度则与光强无关[56].当细胞暴露于多个光源时,Synechocystis sp. PCC6803会整合来自多个光源的信息,并以光场及其变化作为整体进行趋光响应,而不仅仅对单一光源做出响应.此外,由白色全光谱照明切换到黑暗条件时,蓝藻细胞的运动速度在短时间内未发生明显改变;但从红光切换到黑暗条件时,细胞运动速度显著降低;当从绿光切换到黑暗条件时,细胞运动速度明显下降[50].这可能是因为白光比单一波长的光能更有效地使细胞进行光合作用,因此细胞在白光下可以产生并储存更多的能量来支持运动过程.从美国黄石国家公园的温泉中分离出的聚球藻Synechococcus OS-A主要对红光和绿光表现出趋光性,而相同来源的Synechococcus OS-B9对UV-A、蓝光、绿光和红光都表现出趋光性[32].因此,虽然不同蓝藻细胞在趋光性相关的信号通路和分子调控机制上存在共性,但无论是种群内部还是不同种群之间,其光响应行为仍表现出显著差异.这些差异性可能源于内源性因素(如基因表达调控的变异),也可能由外部环境条件(如栖息地的异质性)所驱动. ...
... [32].因此,虽然不同蓝藻细胞在趋光性相关的信号通路和分子调控机制上存在共性,但无论是种群内部还是不同种群之间,其光响应行为仍表现出显著差异.这些差异性可能源于内源性因素(如基因表达调控的变异),也可能由外部环境条件(如栖息地的异质性)所驱动. ...
Diel vertical movements of the cyanobacterium Oscillatoria terebriformis in a sulfide-rich hot-spring microbial mat
1
1987
... 蓝藻可在微生物席、生物土壤结皮等生物复合体结构中通过滑动进行运动,从而对包括水分[25-29]、光照[30-32]、昼夜节律[33-35]、气体浓度[36-37]、盐度[38]和营养物质浓度[39]在内的多种因素变化进行响应. ...
Diel migrations of microorganisms within a benthic,hypersaline mat community
0
1994
Oscillation of type IV pili regulated by the circadian clock in cyanobacterium Synechococcus elongatus PCC7942
2
2024
... 蓝藻可在微生物席、生物土壤结皮等生物复合体结构中通过滑动进行运动,从而对包括水分[25-29]、光照[30-32]、昼夜节律[33-35]、气体浓度[36-37]、盐度[38]和营养物质浓度[39]在内的多种因素变化进行响应. ...
... 蓝藻是迄今为止已知的唯一具有内部生物钟的原核生物,这些生物钟调节细胞代谢的许多方面[64].在白天,蓝藻需要耐受强光、紫外线、氧化还原应激、高温等环境压力,夜间又要应对代谢应激、饥饿等生存压力,这种不同压力类型的规律循环使得蓝藻在长期进化过程中产生了昼夜节律来更好地适应环境[65].例如,在以12 h光照12 h黑暗来模拟自然光照的单个周期中,Synechococcus elongatus PCC7942中的T4P数量和长度发生了明显变化.对于黑暗期结束后刚进入光照期的细胞(即黎明时分),T4P通常更少且更短;而经过12 h光照培养后刚进入黑暗期时(即黄昏时分),细胞的T4P会更多且更长.T4P的活性在不同时刻也不同,黄昏时其伸展和收缩的频率几乎是黎明时的2倍.此外,经固定的光照节律培养后,将培养条件改为连续光照或与原节律相反的光照循环,细胞的T4P都能在至少12 h内保留原光照循环的节律性.而对于kaiC基因(编码核心振荡器的关键蛋白之一,控制蓝藻内部昼夜节律)被敲除的节律缺陷型突变体细胞来说,其在光暗条件变化时几乎不产生明显反应.这表明,T4P的节律性变化并非完全依赖于光照等外部条件的变化,还依赖于细胞内的生物钟机制[35]. ...
Motility of the cyanobacterium Microcoleus chthonoplastes in mud
4
1984
... 蓝藻可在微生物席、生物土壤结皮等生物复合体结构中通过滑动进行运动,从而对包括水分[25-29]、光照[30-32]、昼夜节律[33-35]、气体浓度[36-37]、盐度[38]和营养物质浓度[39]在内的多种因素变化进行响应. ...
... 微生物席(Microbial mats)也可称微生物垫或菌毯,是沉积物基质中生活的各种细菌、古菌和真核微藻的密集底栖群落与其构成的生物复合体,蓝藻是生物席中主要的初级生产者[4].由于沉积物基质中光的急剧衰减和物质传递速率的限制,微生物席的特点是拥有氧气、光强及光谱组成、pH值和硫化物浓度等的陡峭垂直梯度[31].蓝藻可以通过滑动来调整自身在微生物席内不断变化的光场中的位置,从而处于具有最佳光照强度的区域,避免光照不足或受到过强辐射的负面影响[52-54].Lichtenberg等[54]于2020年对在丹麦利姆峡湾定期干燥的沙地上收集的微生物席研究中发现,弱光下蓝藻出现在生物席表面;而在高光强下,蓝藻出现在较深层,说明蓝藻可以通过响应光照强度进行垂直滑动来调整其在生物席中的位置,并且蓝藻在高光强下是负趋光的,即具有避光性,而在低光强下则具有正趋光性.这种特性在螺旋藻(Spirulina)和颤藻(Oscillatoria)中也有发现[31].北威尔士梅奈海峡安格尔西海岸的沿岸泥滩中生活着大量微鞘藻属蓝藻(Microcoleus),泥滩表面在阴天所呈现的绿色较之于晴天时更为明显,说明泥滩中的大多数蓝藻在弱光时具有趋光性,在高光强下具有避光性,当前还尚未确定此类微鞘藻改变趋光向性的阈值光强.上述微鞘藻藻丝在黑暗时也会移动到泥滩表面,但在到达表面后,藻丝在有无光时所形成的图案有所不同:黑暗时藻丝在泥滩表面呈螺旋状,而有光照时藻丝长轴方向与光源方向平行,若此时光源方向改变 90°,藻丝则会重新定向.这说明不只是单个细胞,细胞群落也会集体响应光照条件[36,55]. ...
... 紫外线,尤其是波长在280~315 nm的UV-B,对产氧型光自养生物具有负面影响[57].紫外线会抑制光养生物的生长和光合作用、导致突变和急性生理应激,甚至细胞死亡[58-59].运动是蓝藻避免受到来自较高紫外线剂量的负面影响的一种生存策略,某些微鞘藻(例如Microcoleus chthonoplastes)能通过垂直滑动来免受UV-B的照射,位于生物席表面下方300 µm处的蓝藻所受到的UV-B辐照度仅为表面的10%[30].UV-A(320~400 nm)辐射可诱导某些蓝藻合成伪枝藻素等防晒物质,伪枝藻素能有效吸收紫外辐射,从而减少紫外线对细胞的损害,这种防晒功能使得这些蓝藻能够在较高紫外线辐射的环境中,例如在荒漠生物结皮表面长期生存[60],而不具备此类保护机制的微鞘藻及颤藻则一般存在于相似环境下此类生物复合体内部,仅在伴随着光照强度降低的降水事件等发生时向生物复合体表面滑动[26,36]. ...
... 作为产氧性光自养生物,蓝藻具有向适宜CO2浓度区域迁移的趋化行为,这对维持其光合作用效率至关重要.当环境中CO2长期匮乏时,蓝藻细胞会面临双重生理挑战:一方面,光合色素和类囊体膜会发生降解;另一方面,细胞内超氧化物离子会过量积累,导致氧化应激.同时,O2对蓝藻的生理活动同样不可或缺,它不仅是有氧呼吸的必要底物,还参与某些不饱和脂肪酸的生物合成过程[36-37]. ...
Chemotaxis of cyanobacterium on gradients of carbon-dioxide,bicarbonate and oxygen
3
1985
... 蓝藻可在微生物席、生物土壤结皮等生物复合体结构中通过滑动进行运动,从而对包括水分[25-29]、光照[30-32]、昼夜节律[33-35]、气体浓度[36-37]、盐度[38]和营养物质浓度[39]在内的多种因素变化进行响应. ...
... 作为产氧性光自养生物,蓝藻具有向适宜CO2浓度区域迁移的趋化行为,这对维持其光合作用效率至关重要.当环境中CO2长期匮乏时,蓝藻细胞会面临双重生理挑战:一方面,光合色素和类囊体膜会发生降解;另一方面,细胞内超氧化物离子会过量积累,导致氧化应激.同时,O2对蓝藻的生理活动同样不可或缺,它不仅是有氧呼吸的必要底物,还参与某些不饱和脂肪酸的生物合成过程[36-37]. ...
... 目前已发现某些颤藻属(Oscillatoria)蓝藻表现出对CO2/HCO和O2的光依赖的趋向运动,人工以及由蓝藻自身代谢活动产生的气体浓度梯度均可引发趋向性运动,且引发此类趋向运动的CO2和O2浓度都高于它们在大气中的浓度.O2和CO2会竞争核酮糖二磷酸羧化酶/加氧酶(RubisCO),过高浓度的O2会抑制蓝藻进行光合作用,从而导致蓝藻对其表出现负趋向性.蓝藻对过高浓度(在琼脂中CO2浓度高于0.9 mM)的CO2也表现出负趋向性,这可能是由pH变化导致的,高CO2会使气体出口处的琼脂pH值从8.0降至6.0,而较低浓度的CO2则不会引起该区域内pH的变化[37]. ...
Halotaxis of cyanobacteria in an intertidal hypersaline microbial mat
3
2010
... 蓝藻可在微生物席、生物土壤结皮等生物复合体结构中通过滑动进行运动,从而对包括水分[25-29]、光照[30-32]、昼夜节律[33-35]、气体浓度[36-37]、盐度[38]和营养物质浓度[39]在内的多种因素变化进行响应. ...
... 阿拉伯湾潮间带蓝藻垫的群落动态与盐度变化呈现显著相关性.该区域蓝藻垫在潮汐作用下经历6%~22%的盐度波动,并表现出明显的颜色变化特征:当表层水盐度超过15%时,富含β-胡萝卜素的Euhalothece sp.和Leptolyngbya sp.在表层富集,使藻垫呈现橙红色;盐度降低则促使微鞘藻Microcoleus chthonoplastes向表面迁移,藻垫转为深绿色;当盐度回升至20%时,橙红色特征再次显现.在恒定光强或完全黑暗且保持氧气水平恒定的情况下,藻垫中的M. chthonoplastes会响应盐度变化而滑动,这一过程受多种盐(如NaCl、Na2SO4和MgCl2)的诱导,表明此时其滑动不依赖于光和特定的离子种类.此外,垫层在缺氧状态下或去除硫化物层后藻丝的运动行为未受影响,表明藻丝滑动与氧和硫化物浓度梯度无关.若通过添加蔗糖而非无机盐来降低水活度和水势,则不会抑制盐度降低后蓝藻向表面的滑动,表明此时水活度和水势的变化不直接影响藻丝随盐度变化而产生的运动趋向性[38].这些结果共同表明,蓝藻对盐度变化的感知机制具有高度特异性,其趋向性运动主要响应离子浓度的直接变化,而非通过水活度或水势等间接因素介导[38,72]. ...
... [38,72]. ...
A symbiotic nutrient exchange within the cyanosphere microbiome of the biocrust cyanobacterium,Microcoleus vaginatus
2
2021
... 蓝藻可在微生物席、生物土壤结皮等生物复合体结构中通过滑动进行运动,从而对包括水分[25-29]、光照[30-32]、昼夜节律[33-35]、气体浓度[36-37]、盐度[38]和营养物质浓度[39]在内的多种因素变化进行响应. ...
... 环境中营养元素的丰富程度会影响蓝藻的运动性.研究表明,Microcoleus vaginatus虽不能固氮,但在陆地生态系统等缺氮环境中会聚集成束,藻丝仍会在公共鞘里面滑动,但大多不会移动到鞘外,且鞘内存在独特的细菌群落,即微观的“蓝藻圈”[74-76].结皮微鞘藻蓝藻圈内含有异养固氮细菌,其固氮潜力通常比周围的生物土壤结皮高100倍,而M. vaginatus对这些细菌而言是完成C换N互利共生所需的有机碳来源,这种互利共生关系具有高度特异性.相比之下,将成束的M. vaginatus置于富氮培养基上,其藻丝则会滑动至胶鞘外并迅速铺开,因此藻丝聚集成束可能是其对氮限制的反应[39].此外,单细胞咸水蓝藻Synechocystis sp. UNIWG在不缺氮时会在细胞表面形成刚性针状体,这种针状体似乎参与了细胞在合适环境中的锚定.由于针状体将细胞锚定在特定位置,蓝藻可以在营养富集区域停留更长时间,吸收氮、磷等溶解在水中的营养物质.当环境条件发生变化或细胞需要移动到更有利的区域时,这些针状体会被降解,使细胞恢复滑动能力,从而在环境中灵活调整其位置,寻找更优的生存条件[77]. ...
In situ X-Ray tomography imaging of soil water and cyanobacteria from biological soil crusts undergoing desiccation
2
2018
... 在干旱生境,例如由蓝藻等微生物类群构成的荒漠地表生物土壤结皮中,蓝藻可通过滑动响应可利用水量的变化[25-29,40].在对西班牙东北部沙漠地区的结皮研究中发现,干燥时大量颤藻目蓝藻在距离结皮表面1.5~2.1 mm的深度形成致密层.润湿几分钟后能明显观察到结皮表面变绿,显微镜观察表明,此现象是由结皮表面出现藻丝所致;而当样本逐渐干燥时,藻丝被观察到向结皮内部移动.上述过程在有无光照时均可发生,但将样本暴露在高光强下会减缓或阻止其表面返绿.这表明在此类结皮中,光并非蓝藻响应水含量变化进行滑动的前提条件,且过强的光照可干扰或抑制藻丝向表面滑动[25].向逐渐干燥的结皮样本中添加ATP合成抑制剂或质子动力解偶联剂可抑制藻丝向下滑动,表明蓝藻向下滑动是主动响应的过程,而并非是由空气-水界面的表面张力等物理效应所致,且需要底物磷酸化以外的能量来源[26].而且从完全湿润开始逐渐干燥时,生物结皮内部会形成随深度变化的可用水梯度,即表面含水量低,较深处含水量高[40].因此,推断结皮内蓝藻的滑动是其对水环境变化的运动响应,提出了“趋水性”假说,即蓝藻细胞可以通过某种途径感知环境中可用水的短期变化,或者通过某种传感器感知环境中随含水量变化的参数,如盐度、温度或其他化学物质浓度.上述结皮样本若长时间保持湿润,表面将形成致密的主要由颤藻目蓝藻构成的生物膜,此时若进行干燥,部分蓝藻会被困在干燥的结皮表面[26].为了能够感知到可用水变化,颤藻藻丝在结皮内部的排布方向可能会有一定的垂直分量,在降雨或开始干燥时,部分蓝藻能够快速获得有关环境含水量的信息,使整根藻丝滑动到含水量较高的区域.而在足够长的水分暴露时间后,许多蓝藻藻丝已经完全到达结皮表面并在其上水平滑动,这将使得蓝藻无法有效感知藻丝与其滑动轴正交方向的含水量梯度,从而停留在结皮表面. ...
... [40].因此,推断结皮内蓝藻的滑动是其对水环境变化的运动响应,提出了“趋水性”假说,即蓝藻细胞可以通过某种途径感知环境中可用水的短期变化,或者通过某种传感器感知环境中随含水量变化的参数,如盐度、温度或其他化学物质浓度.上述结皮样本若长时间保持湿润,表面将形成致密的主要由颤藻目蓝藻构成的生物膜,此时若进行干燥,部分蓝藻会被困在干燥的结皮表面[26].为了能够感知到可用水变化,颤藻藻丝在结皮内部的排布方向可能会有一定的垂直分量,在降雨或开始干燥时,部分蓝藻能够快速获得有关环境含水量的信息,使整根藻丝滑动到含水量较高的区域.而在足够长的水分暴露时间后,许多蓝藻藻丝已经完全到达结皮表面并在其上水平滑动,这将使得蓝藻无法有效感知藻丝与其滑动轴正交方向的含水量梯度,从而停留在结皮表面. ...
Oxidative stress and its effects during dehydration
1
2007
... 部分陆生蓝藻在干燥脱水时会进入休眠状态,在有可利用水时才进行活跃的生命活动,并通过光合作用进行生长.而自然条件下,发生降雨事件时通常伴随着光照强度的降低,降雨结束后光照强度逐渐增加,过强的光照会对此时仍留在地表且缺乏光保护色素的蓝藻细胞造成光损伤[41].因此,这种追踪水的行为可能优化了蓝藻在生物结皮中的位置,从而使得缺乏光保护色素的细胞在满足光照要求的同时,避免受到高光强或紫外线等辐照的伤害;其次,蓝藻通过滑动保持细胞周围环境中有一定量的可用水,可以延长其对脱水过程的响应时间,使其更充分地对细胞进行保护以应对即将到来的干旱期,例如通过积累海藻糖和蔗糖等相容性溶质进行渗透保护,或产生EPS减缓细胞水分流失速度,因此有助于蓝藻在干旱环境中的生存[28-29]. ...
Light matters:phototaxis and signal transduction in unicellular cyanobacteria
2
2004
... 趋光性是一种相对于光场方向的定向运动,即单个细胞或细胞群响应光强、光谱进行运动,从而移动到更有利于生存和生长的光条件下[42].蓝藻是光合自养生物,对光照的要求很严格,光照太弱会导致能量产生不足,在某些物种中可能会导致细胞进行异养呼吸;光照太强会降低光合作用效率,并导致光损伤,紫外线辐射对某些无直接光保护响应机制的蓝藻尤其致命,因此,光的波长、强度以及蓝藻种类都会影响蓝藻细胞对光的响应[18,43-44]. ...
... 对于光自养生物来说,光强并非与生长率完全正相关:当光照强度低于最佳光强时,生长率通常随光强增加而升高;然而,超过光饱和点的光照会导致其生长率下降,甚至会造成细胞光损伤并最终导致细胞死亡,某些光合生物虽可将吸收的过多能量较为有效地消散,但其生长率并不会因为光强继续增大而升高[42]. ...
中国人工蓝藻结皮研究进展
1
2023
... 趋光性是一种相对于光场方向的定向运动,即单个细胞或细胞群响应光强、光谱进行运动,从而移动到更有利于生存和生长的光条件下[42].蓝藻是光合自养生物,对光照的要求很严格,光照太弱会导致能量产生不足,在某些物种中可能会导致细胞进行异养呼吸;光照太强会降低光合作用效率,并导致光损伤,紫外线辐射对某些无直接光保护响应机制的蓝藻尤其致命,因此,光的波长、强度以及蓝藻种类都会影响蓝藻细胞对光的响应[18,43-44]. ...
Modeling filamentous cyanobacteria reveals the advantages of long and fast trichomes for optimizing light exposure
2
2011
... 趋光性是一种相对于光场方向的定向运动,即单个细胞或细胞群响应光强、光谱进行运动,从而移动到更有利于生存和生长的光条件下[42].蓝藻是光合自养生物,对光照的要求很严格,光照太弱会导致能量产生不足,在某些物种中可能会导致细胞进行异养呼吸;光照太强会降低光合作用效率,并导致光损伤,紫外线辐射对某些无直接光保护响应机制的蓝藻尤其致命,因此,光的波长、强度以及蓝藻种类都会影响蓝藻细胞对光的响应[18,43-44]. ...
... 单细胞蓝藻和丝状蓝藻感应光和响应光的方式不同.一些研究者认为,Synechocystis sp. PCC 6803能直接感知光源方向而并非光强度的空间梯度,因为其细胞能充当球形微透镜,使细胞“看到”光源并向其移动:单侧光透过细胞一侧后会汇聚于细胞背光侧表层并形成明亮光斑,触发细胞向远离聚焦点的方向移动,因此球形蓝藻可能是世界上最小、最古老的眼球和照相机镜头的原型[16,45-47].此外,在自然环境中,虽然光强度梯度很少陡峭到在几微米的单细胞尺度上产生显著差异[47],但在结构上具有一定长度的颤藻属和席藻属等丝状蓝藻展现出可感知此类光梯度并进行滑动响应的特征.如Phormidium uncinatum藻丝似乎会比较头部和尾部的入射光,如果尾部比头部接收到更多的光,藻丝很可能会改变滑动方向,且其对光非常敏感,两端接收到的光强之间的差异只要高于4%即可引发转向,而照射到藻丝中部的光线则没有这种影响;还有一些蓝藻如Anabaena variabilis可通过藻丝的弯曲和转向来响应光场的不均匀性[44]. ...
Cyanobacteria use micro-optics to sense light direction
1
2016
... 单细胞蓝藻和丝状蓝藻感应光和响应光的方式不同.一些研究者认为,Synechocystis sp. PCC 6803能直接感知光源方向而并非光强度的空间梯度,因为其细胞能充当球形微透镜,使细胞“看到”光源并向其移动:单侧光透过细胞一侧后会汇聚于细胞背光侧表层并形成明亮光斑,触发细胞向远离聚焦点的方向移动,因此球形蓝藻可能是世界上最小、最古老的眼球和照相机镜头的原型[16,45-47].此外,在自然环境中,虽然光强度梯度很少陡峭到在几微米的单细胞尺度上产生显著差异[47],但在结构上具有一定长度的颤藻属和席藻属等丝状蓝藻展现出可感知此类光梯度并进行滑动响应的特征.如Phormidium uncinatum藻丝似乎会比较头部和尾部的入射光,如果尾部比头部接收到更多的光,藻丝很可能会改变滑动方向,且其对光非常敏感,两端接收到的光强之间的差异只要高于4%即可引发转向,而照射到藻丝中部的光线则没有这种影响;还有一些蓝藻如Anabaena variabilis可通过藻丝的弯曲和转向来响应光场的不均匀性[44]. ...
Cyanobacteria in motion
0
2017
Light-controlled motility in prokaryotes and the problem of directional light perception
2
2017
... 单细胞蓝藻和丝状蓝藻感应光和响应光的方式不同.一些研究者认为,Synechocystis sp. PCC 6803能直接感知光源方向而并非光强度的空间梯度,因为其细胞能充当球形微透镜,使细胞“看到”光源并向其移动:单侧光透过细胞一侧后会汇聚于细胞背光侧表层并形成明亮光斑,触发细胞向远离聚焦点的方向移动,因此球形蓝藻可能是世界上最小、最古老的眼球和照相机镜头的原型[16,45-47].此外,在自然环境中,虽然光强度梯度很少陡峭到在几微米的单细胞尺度上产生显著差异[47],但在结构上具有一定长度的颤藻属和席藻属等丝状蓝藻展现出可感知此类光梯度并进行滑动响应的特征.如Phormidium uncinatum藻丝似乎会比较头部和尾部的入射光,如果尾部比头部接收到更多的光,藻丝很可能会改变滑动方向,且其对光非常敏感,两端接收到的光强之间的差异只要高于4%即可引发转向,而照射到藻丝中部的光线则没有这种影响;还有一些蓝藻如Anabaena variabilis可通过藻丝的弯曲和转向来响应光场的不均匀性[44]. ...
... [47],但在结构上具有一定长度的颤藻属和席藻属等丝状蓝藻展现出可感知此类光梯度并进行滑动响应的特征.如Phormidium uncinatum藻丝似乎会比较头部和尾部的入射光,如果尾部比头部接收到更多的光,藻丝很可能会改变滑动方向,且其对光非常敏感,两端接收到的光强之间的差异只要高于4%即可引发转向,而照射到藻丝中部的光线则没有这种影响;还有一些蓝藻如Anabaena variabilis可通过藻丝的弯曲和转向来响应光场的不均匀性[44]. ...
Control of light-dependent behaviour in cyanobacteria by the second messenger cyclic di-GMP
1
2023
... 同时,蓝藻细胞拥有整合多种光输入信号的能力,而不是简单地对占主导的光信号作出反应.这种信号整合可能通过信号通路与光感受器的相互作用,以及通过细胞表面菌毛之间的竞争而发生[48-51].这一过程涉及复杂的信号传导通路和分子调控机制. ...
Effects of sedimentation and light-intensity on mat-forming Oscillatoriaceae with particular reference to Microcoleus lyngbyaceus Gomont
0
1984
Emergent phototactic responses of cyanobacteria under complex light regimes
4
2017
... 一般而言,在未到达光饱和前,波长在500~1 000 nm的光(绿光到红外光)会使蓝藻细胞产生正趋光性,而在500 nm以下的蓝光或紫外线照射下,蓝藻细胞会表现出负趋光性,或由于蓝光或紫外线损害了DNA和其他细胞成分而丧失运动能力[50]. ...
... 集胞藻属蓝藻Synechocystis sp. PCC6803表现出依赖入射光波长的趋光性,此过程通过多种光感受器的作用来实现[32,50].Synechocystis sp. PCC6803菌株偏向于朝绿光(535 nm)或红光(660 nm)光源方向移动,即细胞表现出趋光性,这种运动偏向性随着光通量的增加而增强,并逐渐产生手指状突起,但进行运动的细胞数量的比例和运动速度则与光强无关[56].当细胞暴露于多个光源时,Synechocystis sp. PCC6803会整合来自多个光源的信息,并以光场及其变化作为整体进行趋光响应,而不仅仅对单一光源做出响应.此外,由白色全光谱照明切换到黑暗条件时,蓝藻细胞的运动速度在短时间内未发生明显改变;但从红光切换到黑暗条件时,细胞运动速度显著降低;当从绿光切换到黑暗条件时,细胞运动速度明显下降[50].这可能是因为白光比单一波长的光能更有效地使细胞进行光合作用,因此细胞在白光下可以产生并储存更多的能量来支持运动过程.从美国黄石国家公园的温泉中分离出的聚球藻Synechococcus OS-A主要对红光和绿光表现出趋光性,而相同来源的Synechococcus OS-B9对UV-A、蓝光、绿光和红光都表现出趋光性[32].因此,虽然不同蓝藻细胞在趋光性相关的信号通路和分子调控机制上存在共性,但无论是种群内部还是不同种群之间,其光响应行为仍表现出显著差异.这些差异性可能源于内源性因素(如基因表达调控的变异),也可能由外部环境条件(如栖息地的异质性)所驱动. ...
... [50].这可能是因为白光比单一波长的光能更有效地使细胞进行光合作用,因此细胞在白光下可以产生并储存更多的能量来支持运动过程.从美国黄石国家公园的温泉中分离出的聚球藻Synechococcus OS-A主要对红光和绿光表现出趋光性,而相同来源的Synechococcus OS-B9对UV-A、蓝光、绿光和红光都表现出趋光性[32].因此,虽然不同蓝藻细胞在趋光性相关的信号通路和分子调控机制上存在共性,但无论是种群内部还是不同种群之间,其光响应行为仍表现出显著差异.这些差异性可能源于内源性因素(如基因表达调控的变异),也可能由外部环境条件(如栖息地的异质性)所驱动. ...
... 综上所述,一些蓝藻能根据光的波长和强度不断调整其滑动过程,在细胞层面上实现对光输入的精细整合.这一过程需要 T4P和类似光敏色素的光感受器与复杂的信号转导途径相耦合[52],从而灵敏地控制滑动的方向和速度,但细胞如何整合所有光感受器输入信号的分子机制尚不完全清楚.在宏观尺度上,菌落的集体行为模式不仅依赖于单个细胞的运动行为,还受到细胞间相互作用的影响[17,50]. ...
The cyanobacterial phytochrome Cph2 inhibits phototaxis towards blue light
1
2002
... 同时,蓝藻细胞拥有整合多种光输入信号的能力,而不是简单地对占主导的光信号作出反应.这种信号整合可能通过信号通路与光感受器的相互作用,以及通过细胞表面菌毛之间的竞争而发生[48-51].这一过程涉及复杂的信号传导通路和分子调控机制. ...
Vertical migration of phototrophic bacterial populations in a hypersaline microbial mat from Salins-de-Giraud (Camargue,France)
2
2006
... 微生物席(Microbial mats)也可称微生物垫或菌毯,是沉积物基质中生活的各种细菌、古菌和真核微藻的密集底栖群落与其构成的生物复合体,蓝藻是生物席中主要的初级生产者[4].由于沉积物基质中光的急剧衰减和物质传递速率的限制,微生物席的特点是拥有氧气、光强及光谱组成、pH值和硫化物浓度等的陡峭垂直梯度[31].蓝藻可以通过滑动来调整自身在微生物席内不断变化的光场中的位置,从而处于具有最佳光照强度的区域,避免光照不足或受到过强辐射的负面影响[52-54].Lichtenberg等[54]于2020年对在丹麦利姆峡湾定期干燥的沙地上收集的微生物席研究中发现,弱光下蓝藻出现在生物席表面;而在高光强下,蓝藻出现在较深层,说明蓝藻可以通过响应光照强度进行垂直滑动来调整其在生物席中的位置,并且蓝藻在高光强下是负趋光的,即具有避光性,而在低光强下则具有正趋光性.这种特性在螺旋藻(Spirulina)和颤藻(Oscillatoria)中也有发现[31].北威尔士梅奈海峡安格尔西海岸的沿岸泥滩中生活着大量微鞘藻属蓝藻(Microcoleus),泥滩表面在阴天所呈现的绿色较之于晴天时更为明显,说明泥滩中的大多数蓝藻在弱光时具有趋光性,在高光强下具有避光性,当前还尚未确定此类微鞘藻改变趋光向性的阈值光强.上述微鞘藻藻丝在黑暗时也会移动到泥滩表面,但在到达表面后,藻丝在有无光时所形成的图案有所不同:黑暗时藻丝在泥滩表面呈螺旋状,而有光照时藻丝长轴方向与光源方向平行,若此时光源方向改变 90°,藻丝则会重新定向.这说明不只是单个细胞,细胞群落也会集体响应光照条件[36,55]. ...
... 综上所述,一些蓝藻能根据光的波长和强度不断调整其滑动过程,在细胞层面上实现对光输入的精细整合.这一过程需要 T4P和类似光敏色素的光感受器与复杂的信号转导途径相耦合[52],从而灵敏地控制滑动的方向和速度,但细胞如何整合所有光感受器输入信号的分子机制尚不完全清楚.在宏观尺度上,菌落的集体行为模式不仅依赖于单个细胞的运动行为,还受到细胞间相互作用的影响[17,50]. ...
Surface characterisation reveals substrate suitability for cyanobacterial phototaxis
0
2023
Vertical migration optimizes photosynthetic efficiency of motile cyanobacteria in a coastal microbial mat
2
2020
... 微生物席(Microbial mats)也可称微生物垫或菌毯,是沉积物基质中生活的各种细菌、古菌和真核微藻的密集底栖群落与其构成的生物复合体,蓝藻是生物席中主要的初级生产者[4].由于沉积物基质中光的急剧衰减和物质传递速率的限制,微生物席的特点是拥有氧气、光强及光谱组成、pH值和硫化物浓度等的陡峭垂直梯度[31].蓝藻可以通过滑动来调整自身在微生物席内不断变化的光场中的位置,从而处于具有最佳光照强度的区域,避免光照不足或受到过强辐射的负面影响[52-54].Lichtenberg等[54]于2020年对在丹麦利姆峡湾定期干燥的沙地上收集的微生物席研究中发现,弱光下蓝藻出现在生物席表面;而在高光强下,蓝藻出现在较深层,说明蓝藻可以通过响应光照强度进行垂直滑动来调整其在生物席中的位置,并且蓝藻在高光强下是负趋光的,即具有避光性,而在低光强下则具有正趋光性.这种特性在螺旋藻(Spirulina)和颤藻(Oscillatoria)中也有发现[31].北威尔士梅奈海峡安格尔西海岸的沿岸泥滩中生活着大量微鞘藻属蓝藻(Microcoleus),泥滩表面在阴天所呈现的绿色较之于晴天时更为明显,说明泥滩中的大多数蓝藻在弱光时具有趋光性,在高光强下具有避光性,当前还尚未确定此类微鞘藻改变趋光向性的阈值光强.上述微鞘藻藻丝在黑暗时也会移动到泥滩表面,但在到达表面后,藻丝在有无光时所形成的图案有所不同:黑暗时藻丝在泥滩表面呈螺旋状,而有光照时藻丝长轴方向与光源方向平行,若此时光源方向改变 90°,藻丝则会重新定向.这说明不只是单个细胞,细胞群落也会集体响应光照条件[36,55]. ...
... [54]于2020年对在丹麦利姆峡湾定期干燥的沙地上收集的微生物席研究中发现,弱光下蓝藻出现在生物席表面;而在高光强下,蓝藻出现在较深层,说明蓝藻可以通过响应光照强度进行垂直滑动来调整其在生物席中的位置,并且蓝藻在高光强下是负趋光的,即具有避光性,而在低光强下则具有正趋光性.这种特性在螺旋藻(Spirulina)和颤藻(Oscillatoria)中也有发现[31].北威尔士梅奈海峡安格尔西海岸的沿岸泥滩中生活着大量微鞘藻属蓝藻(Microcoleus),泥滩表面在阴天所呈现的绿色较之于晴天时更为明显,说明泥滩中的大多数蓝藻在弱光时具有趋光性,在高光强下具有避光性,当前还尚未确定此类微鞘藻改变趋光向性的阈值光强.上述微鞘藻藻丝在黑暗时也会移动到泥滩表面,但在到达表面后,藻丝在有无光时所形成的图案有所不同:黑暗时藻丝在泥滩表面呈螺旋状,而有光照时藻丝长轴方向与光源方向平行,若此时光源方向改变 90°,藻丝则会重新定向.这说明不只是单个细胞,细胞群落也会集体响应光照条件[36,55]. ...
Collective self-caging of active filaments in virtual confinement
5
2024
... 微生物席(Microbial mats)也可称微生物垫或菌毯,是沉积物基质中生活的各种细菌、古菌和真核微藻的密集底栖群落与其构成的生物复合体,蓝藻是生物席中主要的初级生产者[4].由于沉积物基质中光的急剧衰减和物质传递速率的限制,微生物席的特点是拥有氧气、光强及光谱组成、pH值和硫化物浓度等的陡峭垂直梯度[31].蓝藻可以通过滑动来调整自身在微生物席内不断变化的光场中的位置,从而处于具有最佳光照强度的区域,避免光照不足或受到过强辐射的负面影响[52-54].Lichtenberg等[54]于2020年对在丹麦利姆峡湾定期干燥的沙地上收集的微生物席研究中发现,弱光下蓝藻出现在生物席表面;而在高光强下,蓝藻出现在较深层,说明蓝藻可以通过响应光照强度进行垂直滑动来调整其在生物席中的位置,并且蓝藻在高光强下是负趋光的,即具有避光性,而在低光强下则具有正趋光性.这种特性在螺旋藻(Spirulina)和颤藻(Oscillatoria)中也有发现[31].北威尔士梅奈海峡安格尔西海岸的沿岸泥滩中生活着大量微鞘藻属蓝藻(Microcoleus),泥滩表面在阴天所呈现的绿色较之于晴天时更为明显,说明泥滩中的大多数蓝藻在弱光时具有趋光性,在高光强下具有避光性,当前还尚未确定此类微鞘藻改变趋光向性的阈值光强.上述微鞘藻藻丝在黑暗时也会移动到泥滩表面,但在到达表面后,藻丝在有无光时所形成的图案有所不同:黑暗时藻丝在泥滩表面呈螺旋状,而有光照时藻丝长轴方向与光源方向平行,若此时光源方向改变 90°,藻丝则会重新定向.这说明不只是单个细胞,细胞群落也会集体响应光照条件[36,55]. ...
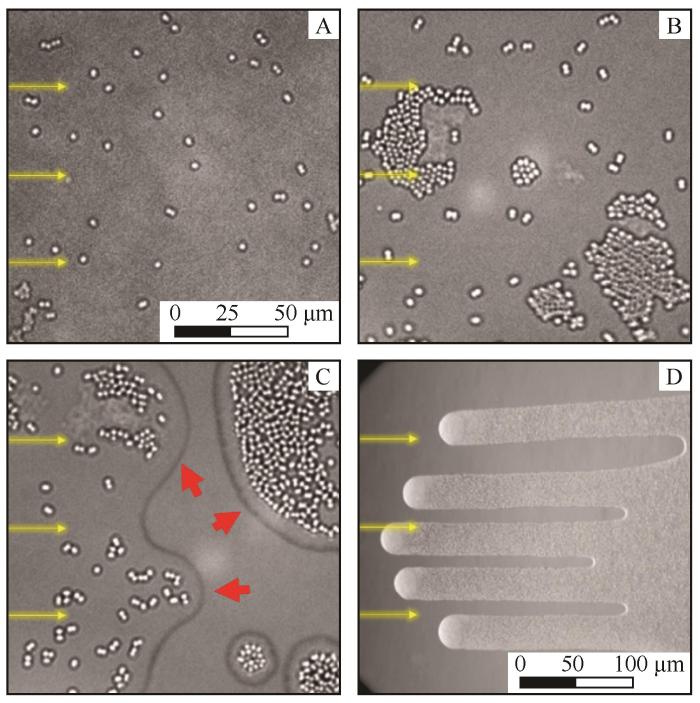
... 在无任何物理障碍的情况下,丝状蓝藻Oscillatoria lutea会向有光区域的边缘聚集(图2).研究者将Oscillatoria lutea放在装有液体培养基的培养皿中,用中心带有不同形状小孔的滤光片使培养皿底部出现不同形状的光斑.实验初期,藻丝在培养皿底部呈均匀随机分布,并沿随机方向滑动,但随着光照时间延长,藻丝会出现明显的边界聚集现象:藻丝逐渐向光斑边缘移动,并在边界处重叠形成一种3~5层藻丝厚、数十根藻丝宽的缠结结构.位于光斑边界的藻丝主要沿光斑的切向方向滑动.当藻丝的一部分进入黑暗区时,它们会开始反向滑动,这种避暗行为可能与其光合能量获取需求相关.由于藻丝具有较高的柔韧性和一定的长度,它们在光斑边界聚集过程中会产生特殊的力学效应:相邻藻丝通过相互推挤和两两对齐,最终形成有序的环形结构.这种自组织行为导致藻丝优先在光斑边缘富集,而非均匀分布于整个光照区域.在移除滤光片后,藻丝仍保持原有滑动方向,导致环形结构逐渐解体;而当光斑尺寸接近藻丝长度(0.2~2 mm)时,这种边界聚集效应就会消失.这种新发现的集体效应具有重要的生态意义:通过局部密度增加,有限数量的藻丝能更有效地构建物理屏障结构,为菌落提供一定程度的保护.尽管形成聚集结构,藻丝在聚集体内部仍保持动态位置交换,这可能是其调节光暴露和其他环境因素的适应性策略[55]. ...
... [
55]
注:上排是在约17~21 h光照后藻丝的显微镜照片;下排是数值模拟中藻丝模型的时间平均向列序取向,颜色的饱和度经过调整以反映藻丝的平均密度,颜色越亮表示藻丝密度越高,藻丝数量越少呈现的颜色越暗.图F中提供了藻丝排列长轴方向的颜色代码,每种颜色与藻丝滑动的方向相对应,每种形状对应的绕数(winding number)显示在图片左上角[55] ...
... 注:上排是在约17~21 h光照后藻丝的显微镜照片;下排是数值模拟中藻丝模型的时间平均向列序取向,颜色的饱和度经过调整以反映藻丝的平均密度,颜色越亮表示藻丝密度越高,藻丝数量越少呈现的颜色越暗.图F中提供了藻丝排列长轴方向的颜色代码,每种颜色与藻丝滑动的方向相对应,每种形状对应的绕数(winding number)显示在图片左上角[55] ...
... [
55]
Fig.2![]()
综上所述,一些蓝藻能根据光的波长和强度不断调整其滑动过程,在细胞层面上实现对光输入的精细整合.这一过程需要 T4P和类似光敏色素的光感受器与复杂的信号转导途径相耦合[52],从而灵敏地控制滑动的方向和速度,但细胞如何整合所有光感受器输入信号的分子机制尚不完全清楚.在宏观尺度上,菌落的集体行为模式不仅依赖于单个细胞的运动行为,还受到细胞间相互作用的影响[17,50]. ...
Tracking phototactic responses and modeling motility of Synechocystis sp.strain PCC6803
6
2008
... 在琼脂基质的趋光性实验中,集胞藻(Synechocystis)表现出独特的群体运动行为.单个集胞藻细胞的运动能力有限,但当细胞通过增殖和聚集达到特定群体规模阈值时,其运动能力显著增强并展现出明显的趋光性.这种群体行为表现为菌落朝向光源方向形成具有规律间隔的手指状突起(图1)[56].有趣的是,当通过改变入射光的方向将一根手指状突起重新定向到另一根“手指”的轨迹中时,细胞的运动速度会立即加快,这种现象可能与细胞运动过程中分泌的EPS的导向作用有关.在一定参数范围内,光强和细胞密度对菌落形态具有显著影响.当光强增加或细胞密度提高时,菌落前缘会形成更为均匀的冠状突起而非手指状结构[32].这一形态转变可能与光场均匀度的改变密切相关:更强的光照和更高的细胞密度都会在一定范围内提高光场的均匀性,从而影响细胞的群体运动模式.这些发现为理解蓝藻群体行为的调控机制提供了新的视角. ...
... [
56]
注:(A)第一阶段,大部分细胞独立运动,四周有少数成对细胞,没有表现出强烈的偏向光源的运动;(B)第二阶段,细胞开始聚集形成更大的群体,运动速率比第一阶段更快,但仍没有强烈偏向光源运动;(C)第三阶段,细胞群周围出现明显的冠状结构(短箭头所示),运动速率比第二阶段更快,右上方形成一个小型指状突起,在此突起中细胞挤在前面,有向光方向移动的倾向;(D)减小放大倍数,可观察到所形成的指状突起,每个“手指”长约 2 mm,前端细胞密集,可在6 h内移动1 mm,此时细胞的运动速率约为第一阶段的6~7倍[56] ...
... 注:(A)第一阶段,大部分细胞独立运动,四周有少数成对细胞,没有表现出强烈的偏向光源的运动;(B)第二阶段,细胞开始聚集形成更大的群体,运动速率比第一阶段更快,但仍没有强烈偏向光源运动;(C)第三阶段,细胞群周围出现明显的冠状结构(短箭头所示),运动速率比第二阶段更快,右上方形成一个小型指状突起,在此突起中细胞挤在前面,有向光方向移动的倾向;(D)减小放大倍数,可观察到所形成的指状突起,每个“手指”长约 2 mm,前端细胞密集,可在6 h内移动1 mm,此时细胞的运动速率约为第一阶段的6~7倍[56] ...
... [
56]
Fig.1![]() <strong>2.2.2</strong> 光谱特性
<strong>2.2.2</strong> 光谱特性一般而言,在未到达光饱和前,波长在500~1 000 nm的光(绿光到红外光)会使蓝藻细胞产生正趋光性,而在500 nm以下的蓝光或紫外线照射下,蓝藻细胞会表现出负趋光性,或由于蓝光或紫外线损害了DNA和其他细胞成分而丧失运动能力[50]. ...
... 集胞藻属蓝藻Synechocystis sp. PCC6803表现出依赖入射光波长的趋光性,此过程通过多种光感受器的作用来实现[32,50].Synechocystis sp. PCC6803菌株偏向于朝绿光(535 nm)或红光(660 nm)光源方向移动,即细胞表现出趋光性,这种运动偏向性随着光通量的增加而增强,并逐渐产生手指状突起,但进行运动的细胞数量的比例和运动速度则与光强无关[56].当细胞暴露于多个光源时,Synechocystis sp. PCC6803会整合来自多个光源的信息,并以光场及其变化作为整体进行趋光响应,而不仅仅对单一光源做出响应.此外,由白色全光谱照明切换到黑暗条件时,蓝藻细胞的运动速度在短时间内未发生明显改变;但从红光切换到黑暗条件时,细胞运动速度显著降低;当从绿光切换到黑暗条件时,细胞运动速度明显下降[50].这可能是因为白光比单一波长的光能更有效地使细胞进行光合作用,因此细胞在白光下可以产生并储存更多的能量来支持运动过程.从美国黄石国家公园的温泉中分离出的聚球藻Synechococcus OS-A主要对红光和绿光表现出趋光性,而相同来源的Synechococcus OS-B9对UV-A、蓝光、绿光和红光都表现出趋光性[32].因此,虽然不同蓝藻细胞在趋光性相关的信号通路和分子调控机制上存在共性,但无论是种群内部还是不同种群之间,其光响应行为仍表现出显著差异.这些差异性可能源于内源性因素(如基因表达调控的变异),也可能由外部环境条件(如栖息地的异质性)所驱动. ...
... 蓝藻分泌的EPS具有缓冲温度变化的功能,能够应对短期内的温度波动,从而通过维持细胞活动的稳定性来提高光合作用和呼吸作用的效率 [67].鉴于蓝藻细胞外层结构(包括鞘、荚膜和黏液层)[56]的包裹,研究者提出了“球壳热传导模型”,该模型认为增加EPS的厚度可以降低细胞与环境之间的导热率,从而减缓细胞内温度的变化,最终减少环境温度波动对细胞生理活动的影响.同时EPS可增强细胞间的连接,使其形成更稳定的群体结构,有助于群体中的蓝藻细胞在环境变化时相互保护和支持[19,67-69]. ...
Effects of solar and ultraviolet-radiation on motility,photomovement and pigmentation in filamentous,gliding cyanobacteria
1
1991
... 紫外线,尤其是波长在280~315 nm的UV-B,对产氧型光自养生物具有负面影响[57].紫外线会抑制光养生物的生长和光合作用、导致突变和急性生理应激,甚至细胞死亡[58-59].运动是蓝藻避免受到来自较高紫外线剂量的负面影响的一种生存策略,某些微鞘藻(例如Microcoleus chthonoplastes)能通过垂直滑动来免受UV-B的照射,位于生物席表面下方300 µm处的蓝藻所受到的UV-B辐照度仅为表面的10%[30].UV-A(320~400 nm)辐射可诱导某些蓝藻合成伪枝藻素等防晒物质,伪枝藻素能有效吸收紫外辐射,从而减少紫外线对细胞的损害,这种防晒功能使得这些蓝藻能够在较高紫外线辐射的环境中,例如在荒漠生物结皮表面长期生存[60],而不具备此类保护机制的微鞘藻及颤藻则一般存在于相似环境下此类生物复合体内部,仅在伴随着光照强度降低的降水事件等发生时向生物复合体表面滑动[26,36]. ...
1
1994
... 紫外线,尤其是波长在280~315 nm的UV-B,对产氧型光自养生物具有负面影响[57].紫外线会抑制光养生物的生长和光合作用、导致突变和急性生理应激,甚至细胞死亡[58-59].运动是蓝藻避免受到来自较高紫外线剂量的负面影响的一种生存策略,某些微鞘藻(例如Microcoleus chthonoplastes)能通过垂直滑动来免受UV-B的照射,位于生物席表面下方300 µm处的蓝藻所受到的UV-B辐照度仅为表面的10%[30].UV-A(320~400 nm)辐射可诱导某些蓝藻合成伪枝藻素等防晒物质,伪枝藻素能有效吸收紫外辐射,从而减少紫外线对细胞的损害,这种防晒功能使得这些蓝藻能够在较高紫外线辐射的环境中,例如在荒漠生物结皮表面长期生存[60],而不具备此类保护机制的微鞘藻及颤藻则一般存在于相似环境下此类生物复合体内部,仅在伴随着光照强度降低的降水事件等发生时向生物复合体表面滑动[26,36]. ...
Effects of solar UV and visible irradiance on photosynthesis and vertical migration of Oscillatoria sp (cyanobacteria) in an Antarctic microbial mat
1
1999
... 紫外线,尤其是波长在280~315 nm的UV-B,对产氧型光自养生物具有负面影响[57].紫外线会抑制光养生物的生长和光合作用、导致突变和急性生理应激,甚至细胞死亡[58-59].运动是蓝藻避免受到来自较高紫外线剂量的负面影响的一种生存策略,某些微鞘藻(例如Microcoleus chthonoplastes)能通过垂直滑动来免受UV-B的照射,位于生物席表面下方300 µm处的蓝藻所受到的UV-B辐照度仅为表面的10%[30].UV-A(320~400 nm)辐射可诱导某些蓝藻合成伪枝藻素等防晒物质,伪枝藻素能有效吸收紫外辐射,从而减少紫外线对细胞的损害,这种防晒功能使得这些蓝藻能够在较高紫外线辐射的环境中,例如在荒漠生物结皮表面长期生存[60],而不具备此类保护机制的微鞘藻及颤藻则一般存在于相似环境下此类生物复合体内部,仅在伴随着光照强度降低的降水事件等发生时向生物复合体表面滑动[26,36]. ...
Gene expression patterns associated with the biosynthesis of the sunscreen scytonemin in Nostoc punctiforme ATCC 29133 in response to UVA radiation
1
2009
... 紫外线,尤其是波长在280~315 nm的UV-B,对产氧型光自养生物具有负面影响[57].紫外线会抑制光养生物的生长和光合作用、导致突变和急性生理应激,甚至细胞死亡[58-59].运动是蓝藻避免受到来自较高紫外线剂量的负面影响的一种生存策略,某些微鞘藻(例如Microcoleus chthonoplastes)能通过垂直滑动来免受UV-B的照射,位于生物席表面下方300 µm处的蓝藻所受到的UV-B辐照度仅为表面的10%[30].UV-A(320~400 nm)辐射可诱导某些蓝藻合成伪枝藻素等防晒物质,伪枝藻素能有效吸收紫外辐射,从而减少紫外线对细胞的损害,这种防晒功能使得这些蓝藻能够在较高紫外线辐射的环境中,例如在荒漠生物结皮表面长期生存[60],而不具备此类保护机制的微鞘藻及颤藻则一般存在于相似环境下此类生物复合体内部,仅在伴随着光照强度降低的降水事件等发生时向生物复合体表面滑动[26,36]. ...
Phototaxis in cyanobacteria:from mutants to models of collective behavior
1
2021
... 在自然环境中,具有滑动能力的蓝藻通常以群体形式存在,它们与非生物组分共同构成密集的聚集体或分层的生物膜等复杂生物复合体.这些复合体内部存在着多种环境因子的动态相互作用,因此研究蓝藻在群体水平上的运动行为具有重要意义.值得注意的是,在趋光性运动过程中,蓝藻群体会展现出显著的形态变化,这些变化主要源于细胞间的相互作用和协同效应[61]. ...
Phototactic cyanobacteria as an active matter system
1
2022
... T4P 的物理连接和EPS轨迹的效应导致的群落中细胞间相互作用或可解释某些蓝藻(例如Synechocystis sp.)趋光运动的集体行为[17-21,62].蓝藻细胞的趋光性运动依赖于多个关键基因的协同作用.研究表明,参与菌毛生物合成和功能的运动相关基因对趋光性至关重要,同时调控环磷酸腺苷(Cyclic Adenosine Monophosphate,cAMP)信号通路的基因也发挥着重要作用.cAMP作为从原核生物到高等生物中保守的第二信使,由腺苷酸环化酶催化ATP生成,在细胞信号转导中具有广泛功能[63].特别值得注意的是,当缺失cya1(腺苷酸环化酶)和sycrp1(一种cAMP受体样蛋白)基因时,突变体菌落丧失趋光性能力;然而有趣的是,当这些突变体细胞与正常趋光性细胞共培养时,它们能够通过“搭便车”机制参与群体趋光运动.这种独特的群体行为源于T4P介导的细胞间相互作用:突变细胞通过Ⅳ型菌毛依附于具有光感应能力的细胞,并借助邻近细胞施加的机械力实现定向移动[18].此外,蓝藻群体趋光运动还表现出显著的协同效应.细胞滑动过程中分泌的EPS具有双重功能:一方面降低细胞运动的表面摩擦力,另一方面形成引导路径促进后续细胞的定向移动.这种正反馈机制使得群体迁移速度显著高于单个细胞,其动力学过程类似于窗户上雨滴的移动模式——后续水滴沿既有路径更快移动并与前导水滴融合,最终形成更大的水滴.这种基于EPS引导的集体运动机制,与T4P介导的细胞间力学耦合共同构成了蓝藻群体趋光性的核心基础[17]. ...
Blue light stimulates cyanobacterial motility via a cAMP signal transduction system
1
2004
... T4P 的物理连接和EPS轨迹的效应导致的群落中细胞间相互作用或可解释某些蓝藻(例如Synechocystis sp.)趋光运动的集体行为[17-21,62].蓝藻细胞的趋光性运动依赖于多个关键基因的协同作用.研究表明,参与菌毛生物合成和功能的运动相关基因对趋光性至关重要,同时调控环磷酸腺苷(Cyclic Adenosine Monophosphate,cAMP)信号通路的基因也发挥着重要作用.cAMP作为从原核生物到高等生物中保守的第二信使,由腺苷酸环化酶催化ATP生成,在细胞信号转导中具有广泛功能[63].特别值得注意的是,当缺失cya1(腺苷酸环化酶)和sycrp1(一种cAMP受体样蛋白)基因时,突变体菌落丧失趋光性能力;然而有趣的是,当这些突变体细胞与正常趋光性细胞共培养时,它们能够通过“搭便车”机制参与群体趋光运动.这种独特的群体行为源于T4P介导的细胞间相互作用:突变细胞通过Ⅳ型菌毛依附于具有光感应能力的细胞,并借助邻近细胞施加的机械力实现定向移动[18].此外,蓝藻群体趋光运动还表现出显著的协同效应.细胞滑动过程中分泌的EPS具有双重功能:一方面降低细胞运动的表面摩擦力,另一方面形成引导路径促进后续细胞的定向移动.这种正反馈机制使得群体迁移速度显著高于单个细胞,其动力学过程类似于窗户上雨滴的移动模式——后续水滴沿既有路径更快移动并与前导水滴融合,最终形成更大的水滴.这种基于EPS引导的集体运动机制,与T4P介导的细胞间力学耦合共同构成了蓝藻群体趋光性的核心基础[17]. ...
Circadian rhythms in cyanobacteria
1
2015
... 蓝藻是迄今为止已知的唯一具有内部生物钟的原核生物,这些生物钟调节细胞代谢的许多方面[64].在白天,蓝藻需要耐受强光、紫外线、氧化还原应激、高温等环境压力,夜间又要应对代谢应激、饥饿等生存压力,这种不同压力类型的规律循环使得蓝藻在长期进化过程中产生了昼夜节律来更好地适应环境[65].例如,在以12 h光照12 h黑暗来模拟自然光照的单个周期中,Synechococcus elongatus PCC7942中的T4P数量和长度发生了明显变化.对于黑暗期结束后刚进入光照期的细胞(即黎明时分),T4P通常更少且更短;而经过12 h光照培养后刚进入黑暗期时(即黄昏时分),细胞的T4P会更多且更长.T4P的活性在不同时刻也不同,黄昏时其伸展和收缩的频率几乎是黎明时的2倍.此外,经固定的光照节律培养后,将培养条件改为连续光照或与原节律相反的光照循环,细胞的T4P都能在至少12 h内保留原光照循环的节律性.而对于kaiC基因(编码核心振荡器的关键蛋白之一,控制蓝藻内部昼夜节律)被敲除的节律缺陷型突变体细胞来说,其在光暗条件变化时几乎不产生明显反应.这表明,T4P的节律性变化并非完全依赖于光照等外部条件的变化,还依赖于细胞内的生物钟机制[35]. ...
Bacteria can anticipate the seasons:photoperiodism in cyanobacteria
2
2024
... 蓝藻是迄今为止已知的唯一具有内部生物钟的原核生物,这些生物钟调节细胞代谢的许多方面[64].在白天,蓝藻需要耐受强光、紫外线、氧化还原应激、高温等环境压力,夜间又要应对代谢应激、饥饿等生存压力,这种不同压力类型的规律循环使得蓝藻在长期进化过程中产生了昼夜节律来更好地适应环境[65].例如,在以12 h光照12 h黑暗来模拟自然光照的单个周期中,Synechococcus elongatus PCC7942中的T4P数量和长度发生了明显变化.对于黑暗期结束后刚进入光照期的细胞(即黎明时分),T4P通常更少且更短;而经过12 h光照培养后刚进入黑暗期时(即黄昏时分),细胞的T4P会更多且更长.T4P的活性在不同时刻也不同,黄昏时其伸展和收缩的频率几乎是黎明时的2倍.此外,经固定的光照节律培养后,将培养条件改为连续光照或与原节律相反的光照循环,细胞的T4P都能在至少12 h内保留原光照循环的节律性.而对于kaiC基因(编码核心振荡器的关键蛋白之一,控制蓝藻内部昼夜节律)被敲除的节律缺陷型突变体细胞来说,其在光暗条件变化时几乎不产生明显反应.这表明,T4P的节律性变化并非完全依赖于光照等外部条件的变化,还依赖于细胞内的生物钟机制[35]. ...
... 除昼夜节律外,一些蓝藻还能通过光周期的时间长短等信息来预测关键的季节性变化,例如暴露在冬季光周期中的单细胞蓝藻Synechococcus elongatus PCC7942在低温下的存活能力比暴露在夏季光周期中的蓝藻高出2~3倍,其原因在于短日照和长日照会促进不同的转录程序,而暴露在短日照下的细胞会经历膜脂饱和度的适应性变化,类似于暴露在低温下的细胞[65].当前季节节律对蓝藻滑动特征的影响还有待研究. ...
Seeking sunlight:rapid phototactic motility of filamentous mat-forming cyanobacteria optimize photosynthesis and enhance carbon burial in Lake Huron's submerged sinkholes
2
2015
... 温度变化对蓝藻的生理活性和运动能力具有直接和间接影响[66-70].从美国休伦湖的低光、低氧和高硫环境中分离出的颤藻(Oscillatoria)能够在4~45 ℃生存.在这一范围内,随着温度升高,藻丝的滑动也相应加快,可能是因为低温时蓝藻细胞的代谢活动减缓,无法产生足够的能量来支持快速滑动.然而,当环境温度逐渐接近45 ℃时,藻丝滑动速度的增加也趋于缓慢,这主要是因为高温对蓝藻细胞的生理活动产生了抑制作用[66].高温会降低细胞内酶的活性,进而影响能量代谢和与运动相关的生化反应.此外,随着温度上升,蓝藻的EPS变得更厚、更黏稠,这增加了细胞滑动时的摩擦力,进一步限制了藻丝的滑动速度[67].高温环境还可能引发细胞的应激反应,使细胞优先分配资源用于生存保护,而不是运动,从而减缓了滑动速度的增加. ...
... [66].高温会降低细胞内酶的活性,进而影响能量代谢和与运动相关的生化反应.此外,随着温度上升,蓝藻的EPS变得更厚、更黏稠,这增加了细胞滑动时的摩擦力,进一步限制了藻丝的滑动速度[67].高温环境还可能引发细胞的应激反应,使细胞优先分配资源用于生存保护,而不是运动,从而减缓了滑动速度的增加. ...
The role of cyanobacterial external layers in mass transfer:evidence from temperature shock experiments by noninvasive microtest technology
3
2020
... 温度变化对蓝藻的生理活性和运动能力具有直接和间接影响[66-70].从美国休伦湖的低光、低氧和高硫环境中分离出的颤藻(Oscillatoria)能够在4~45 ℃生存.在这一范围内,随着温度升高,藻丝的滑动也相应加快,可能是因为低温时蓝藻细胞的代谢活动减缓,无法产生足够的能量来支持快速滑动.然而,当环境温度逐渐接近45 ℃时,藻丝滑动速度的增加也趋于缓慢,这主要是因为高温对蓝藻细胞的生理活动产生了抑制作用[66].高温会降低细胞内酶的活性,进而影响能量代谢和与运动相关的生化反应.此外,随着温度上升,蓝藻的EPS变得更厚、更黏稠,这增加了细胞滑动时的摩擦力,进一步限制了藻丝的滑动速度[67].高温环境还可能引发细胞的应激反应,使细胞优先分配资源用于生存保护,而不是运动,从而减缓了滑动速度的增加. ...
... 蓝藻分泌的EPS具有缓冲温度变化的功能,能够应对短期内的温度波动,从而通过维持细胞活动的稳定性来提高光合作用和呼吸作用的效率 [67].鉴于蓝藻细胞外层结构(包括鞘、荚膜和黏液层)[56]的包裹,研究者提出了“球壳热传导模型”,该模型认为增加EPS的厚度可以降低细胞与环境之间的导热率,从而减缓细胞内温度的变化,最终减少环境温度波动对细胞生理活动的影响.同时EPS可增强细胞间的连接,使其形成更稳定的群体结构,有助于群体中的蓝藻细胞在环境变化时相互保护和支持[19,67-69]. ...
... ,67-69]. ...
Exocellular polysaccharides from cyanobacteria and their possible applications
0
1998
Effect of temperature on photosynthesis,oxygen consumption and sulfide production in an extremely hypersaline cyanobacterial mat
1
2006
... 蓝藻分泌的EPS具有缓冲温度变化的功能,能够应对短期内的温度波动,从而通过维持细胞活动的稳定性来提高光合作用和呼吸作用的效率 [67].鉴于蓝藻细胞外层结构(包括鞘、荚膜和黏液层)[56]的包裹,研究者提出了“球壳热传导模型”,该模型认为增加EPS的厚度可以降低细胞与环境之间的导热率,从而减缓细胞内温度的变化,最终减少环境温度波动对细胞生理活动的影响.同时EPS可增强细胞间的连接,使其形成更稳定的群体结构,有助于群体中的蓝藻细胞在环境变化时相互保护和支持[19,67-69]. ...
New insights into external layers of cyanobacteria and microalgae based on multiscale analysis of AFM force-distance curves
2
2021
... 温度变化对蓝藻的生理活性和运动能力具有直接和间接影响[66-70].从美国休伦湖的低光、低氧和高硫环境中分离出的颤藻(Oscillatoria)能够在4~45 ℃生存.在这一范围内,随着温度升高,藻丝的滑动也相应加快,可能是因为低温时蓝藻细胞的代谢活动减缓,无法产生足够的能量来支持快速滑动.然而,当环境温度逐渐接近45 ℃时,藻丝滑动速度的增加也趋于缓慢,这主要是因为高温对蓝藻细胞的生理活动产生了抑制作用[66].高温会降低细胞内酶的活性,进而影响能量代谢和与运动相关的生化反应.此外,随着温度上升,蓝藻的EPS变得更厚、更黏稠,这增加了细胞滑动时的摩擦力,进一步限制了藻丝的滑动速度[67].高温环境还可能引发细胞的应激反应,使细胞优先分配资源用于生存保护,而不是运动,从而减缓了滑动速度的增加. ...
... 温度变化可显著改变蓝藻EPS的理化性质,而EPS量及其成分与特性的改变都可能对蓝藻的滑动特征产生影响[70].研究者将Nostoc sp.(FACHB-2009)、Dolichospermum sp.(FACHB-82)和Microcystis sp.(FACHB-3296)3种蓝藻菌株置于3种温度(10、25、35 ℃)下培养,发现在10 ℃和35 ℃时,单位生物量的EPS含量相对较高,表明蓝藻在相对极端温度条件下倾向于分泌更多EPS以保护细胞.在高温环境中,菌落的尺寸增大,EPS变得更厚、更松散且黏性更强(图3).此时未观察到菌落中单个细胞尺寸有显著增加,因此导致菌落尺寸增大的核心因素可能是菌落中细胞数量增多.EPS黏性的增加有助于菌落结构的形成和维持,并可能有助于蓝藻吸收营养物质,同时也会增加单个细胞滑动时的阻力[19]. ...
Salinity-dependent limitation of photosynthesis and oxygen exchange in microbial mats
1
1999
... 趋盐性(Halotaxis)是微生物沿着盐度梯度正向或反向运动的能力.盐度对微生物垫中的微生物群落多样性和功能具有重要影响.过高的盐度通常会导致细胞面临渗透胁迫、离子毒性和光合作用抑制等问题,从而抑制微生物的生长和繁殖,因此微生物多样性常随着盐浓度的增加而减少[71-73]. ...
Long-term manipulations of intact microbial mat communities in a greenhouse collaboratory:simulating earth's present and past field environments
1
2002
... 阿拉伯湾潮间带蓝藻垫的群落动态与盐度变化呈现显著相关性.该区域蓝藻垫在潮汐作用下经历6%~22%的盐度波动,并表现出明显的颜色变化特征:当表层水盐度超过15%时,富含β-胡萝卜素的Euhalothece sp.和Leptolyngbya sp.在表层富集,使藻垫呈现橙红色;盐度降低则促使微鞘藻Microcoleus chthonoplastes向表面迁移,藻垫转为深绿色;当盐度回升至20%时,橙红色特征再次显现.在恒定光强或完全黑暗且保持氧气水平恒定的情况下,藻垫中的M. chthonoplastes会响应盐度变化而滑动,这一过程受多种盐(如NaCl、Na2SO4和MgCl2)的诱导,表明此时其滑动不依赖于光和特定的离子种类.此外,垫层在缺氧状态下或去除硫化物层后藻丝的运动行为未受影响,表明藻丝滑动与氧和硫化物浓度梯度无关.若通过添加蔗糖而非无机盐来降低水活度和水势,则不会抑制盐度降低后蓝藻向表面的滑动,表明此时水活度和水势的变化不直接影响藻丝随盐度变化而产生的运动趋向性[38].这些结果共同表明,蓝藻对盐度变化的感知机制具有高度特异性,其趋向性运动主要响应离子浓度的直接变化,而非通过水活度或水势等间接因素介导[38,72]. ...
Microbial diversity of benthic mats along a tidal desiccation gradient
1
2005
... 趋盐性(Halotaxis)是微生物沿着盐度梯度正向或反向运动的能力.盐度对微生物垫中的微生物群落多样性和功能具有重要影响.过高的盐度通常会导致细胞面临渗透胁迫、离子毒性和光合作用抑制等问题,从而抑制微生物的生长和繁殖,因此微生物多样性常随着盐浓度的增加而减少[71-73]. ...
Variations in the buoyancy response of Microystis aeruginosa to nitrogen,phosphorus and light
1
2001
... 环境中营养元素的丰富程度会影响蓝藻的运动性.研究表明,Microcoleus vaginatus虽不能固氮,但在陆地生态系统等缺氮环境中会聚集成束,藻丝仍会在公共鞘里面滑动,但大多不会移动到鞘外,且鞘内存在独特的细菌群落,即微观的“蓝藻圈”[74-76].结皮微鞘藻蓝藻圈内含有异养固氮细菌,其固氮潜力通常比周围的生物土壤结皮高100倍,而M. vaginatus对这些细菌而言是完成C换N互利共生所需的有机碳来源,这种互利共生关系具有高度特异性.相比之下,将成束的M. vaginatus置于富氮培养基上,其藻丝则会滑动至胶鞘外并迅速铺开,因此藻丝聚集成束可能是其对氮限制的反应[39].此外,单细胞咸水蓝藻Synechocystis sp. UNIWG在不缺氮时会在细胞表面形成刚性针状体,这种针状体似乎参与了细胞在合适环境中的锚定.由于针状体将细胞锚定在特定位置,蓝藻可以在营养富集区域停留更长时间,吸收氮、磷等溶解在水中的营养物质.当环境条件发生变化或细胞需要移动到更有利的区域时,这些针状体会被降解,使细胞恢复滑动能力,从而在环境中灵活调整其位置,寻找更优的生存条件[77]. ...
Direct measurement of O2-depleted microzones in marine oscillatoria:relation to N2 fixation
0
1988
Role of chemotaxis in establishing a specific nitrogen-fixing cyanobacterial-bacterial association
1
1985
... 环境中营养元素的丰富程度会影响蓝藻的运动性.研究表明,Microcoleus vaginatus虽不能固氮,但在陆地生态系统等缺氮环境中会聚集成束,藻丝仍会在公共鞘里面滑动,但大多不会移动到鞘外,且鞘内存在独特的细菌群落,即微观的“蓝藻圈”[74-76].结皮微鞘藻蓝藻圈内含有异养固氮细菌,其固氮潜力通常比周围的生物土壤结皮高100倍,而M. vaginatus对这些细菌而言是完成C换N互利共生所需的有机碳来源,这种互利共生关系具有高度特异性.相比之下,将成束的M. vaginatus置于富氮培养基上,其藻丝则会滑动至胶鞘外并迅速铺开,因此藻丝聚集成束可能是其对氮限制的反应[39].此外,单细胞咸水蓝藻Synechocystis sp. UNIWG在不缺氮时会在细胞表面形成刚性针状体,这种针状体似乎参与了细胞在合适环境中的锚定.由于针状体将细胞锚定在特定位置,蓝藻可以在营养富集区域停留更长时间,吸收氮、磷等溶解在水中的营养物质.当环境条件发生变化或细胞需要移动到更有利的区域时,这些针状体会被降解,使细胞恢复滑动能力,从而在环境中灵活调整其位置,寻找更优的生存条件[77]. ...
Phototactic motility of Synechocystis sp UNIWG (cyanobacteria) from brackish environment
1
2010
... 环境中营养元素的丰富程度会影响蓝藻的运动性.研究表明,Microcoleus vaginatus虽不能固氮,但在陆地生态系统等缺氮环境中会聚集成束,藻丝仍会在公共鞘里面滑动,但大多不会移动到鞘外,且鞘内存在独特的细菌群落,即微观的“蓝藻圈”[74-76].结皮微鞘藻蓝藻圈内含有异养固氮细菌,其固氮潜力通常比周围的生物土壤结皮高100倍,而M. vaginatus对这些细菌而言是完成C换N互利共生所需的有机碳来源,这种互利共生关系具有高度特异性.相比之下,将成束的M. vaginatus置于富氮培养基上,其藻丝则会滑动至胶鞘外并迅速铺开,因此藻丝聚集成束可能是其对氮限制的反应[39].此外,单细胞咸水蓝藻Synechocystis sp. UNIWG在不缺氮时会在细胞表面形成刚性针状体,这种针状体似乎参与了细胞在合适环境中的锚定.由于针状体将细胞锚定在特定位置,蓝藻可以在营养富集区域停留更长时间,吸收氮、磷等溶解在水中的营养物质.当环境条件发生变化或细胞需要移动到更有利的区域时,这些针状体会被降解,使细胞恢复滑动能力,从而在环境中灵活调整其位置,寻找更优的生存条件[77]. ...
Cyanobacterial chemotaxis to extracts of host and nonhost plants
1
2006
... 乙烯是重要的植物激素,不仅会影响植物的生长、发育和对许多胁迫的反应,还会对一些蓝藻的趋光性运动产生影响[78].目前已知集胞藻属蓝藻Synechocystis sp. PCC6803含有功能性乙烯受体(SynEtr1),该受体可调节细胞表面结构,空气中不超过8 nL·L-1的乙烯浓度即可通过引发细胞表面结构变化来增强其生物膜形成的能力,并使单个细胞的趋光性更加显著,同时提高细胞聚集时整体向光源移动的速度[79].空气中的低浓度乙烯(8 nL·L-1)可增强Synechocystis sp. PCC6803和盖丝藻属蓝藻 Geitlerinema sp. PCC7105对白光和红光的趋光性,但似乎并不影响其对蓝光或绿光的趋光性;而较高浓度的乙烯(1 000 nL·L-1)则会使Geitlerinema sp. PCC7105对弱光产生避光性.值得注意的是,乙烯还会影响蓝藻对不同波长光的反应.与不含乙烯的空气相比,Geitlerinema sp. PCC7105在存在乙烯的情况下表现出更强的耐受多种波长光的能力.这种现象可能是由于较高浓度的乙烯抑制了蓝藻细胞内反应调节蛋白的产生,进而导致蓝藻对光的波长不再敏感[80].其他蓝藻对乙烯有何反应仍有待研究. ...
Ethylene regulates the physiology of the cyanobacterium Synechocystis sp PCC 6803 via an ethylene receptor
1
2016
... 乙烯是重要的植物激素,不仅会影响植物的生长、发育和对许多胁迫的反应,还会对一些蓝藻的趋光性运动产生影响[78].目前已知集胞藻属蓝藻Synechocystis sp. PCC6803含有功能性乙烯受体(SynEtr1),该受体可调节细胞表面结构,空气中不超过8 nL·L-1的乙烯浓度即可通过引发细胞表面结构变化来增强其生物膜形成的能力,并使单个细胞的趋光性更加显著,同时提高细胞聚集时整体向光源移动的速度[79].空气中的低浓度乙烯(8 nL·L-1)可增强Synechocystis sp. PCC6803和盖丝藻属蓝藻 Geitlerinema sp. PCC7105对白光和红光的趋光性,但似乎并不影响其对蓝光或绿光的趋光性;而较高浓度的乙烯(1 000 nL·L-1)则会使Geitlerinema sp. PCC7105对弱光产生避光性.值得注意的是,乙烯还会影响蓝藻对不同波长光的反应.与不含乙烯的空气相比,Geitlerinema sp. PCC7105在存在乙烯的情况下表现出更强的耐受多种波长光的能力.这种现象可能是由于较高浓度的乙烯抑制了蓝藻细胞内反应调节蛋白的产生,进而导致蓝藻对光的波长不再敏感[80].其他蓝藻对乙烯有何反应仍有待研究. ...
Cyanobacteria respond to low levels of ethylene
1
2019
... 乙烯是重要的植物激素,不仅会影响植物的生长、发育和对许多胁迫的反应,还会对一些蓝藻的趋光性运动产生影响[78].目前已知集胞藻属蓝藻Synechocystis sp. PCC6803含有功能性乙烯受体(SynEtr1),该受体可调节细胞表面结构,空气中不超过8 nL·L-1的乙烯浓度即可通过引发细胞表面结构变化来增强其生物膜形成的能力,并使单个细胞的趋光性更加显著,同时提高细胞聚集时整体向光源移动的速度[79].空气中的低浓度乙烯(8 nL·L-1)可增强Synechocystis sp. PCC6803和盖丝藻属蓝藻 Geitlerinema sp. PCC7105对白光和红光的趋光性,但似乎并不影响其对蓝光或绿光的趋光性;而较高浓度的乙烯(1 000 nL·L-1)则会使Geitlerinema sp. PCC7105对弱光产生避光性.值得注意的是,乙烯还会影响蓝藻对不同波长光的反应.与不含乙烯的空气相比,Geitlerinema sp. PCC7105在存在乙烯的情况下表现出更强的耐受多种波长光的能力.这种现象可能是由于较高浓度的乙烯抑制了蓝藻细胞内反应调节蛋白的产生,进而导致蓝藻对光的波长不再敏感[80].其他蓝藻对乙烯有何反应仍有待研究. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}