气候变化对陆地水循环影响研究的问题
1
2004
... 蒸散发(Evapotranspiration, ET)作为地表水循环和能量交换的重要组成部分[1],通过土壤-植被-大气连续体(Soil-plant-atmosphere-continuum, SPAC)的相互作用[2],深刻影响着生态系统的水分与能量平衡.蒸散发观测方法因空间尺度而异,植物个体尺度采用蒸渗仪法[3],站点尺度依赖涡度相关法[4]和波文比法[5],区域尺度及全球尺度则结合大孔径闪烁仪法[6]与遥感模型反演[7].随着遥感技术和蒸散发模型(如Priestley-Taylor模型)的发展,蒸散比(Evaporative Fraction, EF)作为衡量地表能量分配和水分循环的关键参数,已成为水分-能量耦合的重要指标[8-13]. ...
The future of evapotranspiration:global requirements for ecosystem functioning,carbon and climate feedbacks,agricultural management,and water resources
1
2017
... 蒸散发(Evapotranspiration, ET)作为地表水循环和能量交换的重要组成部分[1],通过土壤-植被-大气连续体(Soil-plant-atmosphere-continuum, SPAC)的相互作用[2],深刻影响着生态系统的水分与能量平衡.蒸散发观测方法因空间尺度而异,植物个体尺度采用蒸渗仪法[3],站点尺度依赖涡度相关法[4]和波文比法[5],区域尺度及全球尺度则结合大孔径闪烁仪法[6]与遥感模型反演[7].随着遥感技术和蒸散发模型(如Priestley-Taylor模型)的发展,蒸散比(Evaporative Fraction, EF)作为衡量地表能量分配和水分循环的关键参数,已成为水分-能量耦合的重要指标[8-13]. ...
蒸渗仪国内外应用现状及研究趋势
1
2018
... 蒸散发(Evapotranspiration, ET)作为地表水循环和能量交换的重要组成部分[1],通过土壤-植被-大气连续体(Soil-plant-atmosphere-continuum, SPAC)的相互作用[2],深刻影响着生态系统的水分与能量平衡.蒸散发观测方法因空间尺度而异,植物个体尺度采用蒸渗仪法[3],站点尺度依赖涡度相关法[4]和波文比法[5],区域尺度及全球尺度则结合大孔径闪烁仪法[6]与遥感模型反演[7].随着遥感技术和蒸散发模型(如Priestley-Taylor模型)的发展,蒸散比(Evaporative Fraction, EF)作为衡量地表能量分配和水分循环的关键参数,已成为水分-能量耦合的重要指标[8-13]. ...
涡度相关技术及其在陆地生态系统通量研究中的应用
1
2020
... 蒸散发(Evapotranspiration, ET)作为地表水循环和能量交换的重要组成部分[1],通过土壤-植被-大气连续体(Soil-plant-atmosphere-continuum, SPAC)的相互作用[2],深刻影响着生态系统的水分与能量平衡.蒸散发观测方法因空间尺度而异,植物个体尺度采用蒸渗仪法[3],站点尺度依赖涡度相关法[4]和波文比法[5],区域尺度及全球尺度则结合大孔径闪烁仪法[6]与遥感模型反演[7].随着遥感技术和蒸散发模型(如Priestley-Taylor模型)的发展,蒸散比(Evaporative Fraction, EF)作为衡量地表能量分配和水分循环的关键参数,已成为水分-能量耦合的重要指标[8-13]. ...
区域蒸散发的实测及模拟研究
1
2017
... 蒸散发(Evapotranspiration, ET)作为地表水循环和能量交换的重要组成部分[1],通过土壤-植被-大气连续体(Soil-plant-atmosphere-continuum, SPAC)的相互作用[2],深刻影响着生态系统的水分与能量平衡.蒸散发观测方法因空间尺度而异,植物个体尺度采用蒸渗仪法[3],站点尺度依赖涡度相关法[4]和波文比法[5],区域尺度及全球尺度则结合大孔径闪烁仪法[6]与遥感模型反演[7].随着遥感技术和蒸散发模型(如Priestley-Taylor模型)的发展,蒸散比(Evaporative Fraction, EF)作为衡量地表能量分配和水分循环的关键参数,已成为水分-能量耦合的重要指标[8-13]. ...
涡动相关系统和小孔径闪烁仪观测的森林显热通量的异同研究
1
2010
... 蒸散发(Evapotranspiration, ET)作为地表水循环和能量交换的重要组成部分[1],通过土壤-植被-大气连续体(Soil-plant-atmosphere-continuum, SPAC)的相互作用[2],深刻影响着生态系统的水分与能量平衡.蒸散发观测方法因空间尺度而异,植物个体尺度采用蒸渗仪法[3],站点尺度依赖涡度相关法[4]和波文比法[5],区域尺度及全球尺度则结合大孔径闪烁仪法[6]与遥感模型反演[7].随着遥感技术和蒸散发模型(如Priestley-Taylor模型)的发展,蒸散比(Evaporative Fraction, EF)作为衡量地表能量分配和水分循环的关键参数,已成为水分-能量耦合的重要指标[8-13]. ...
A review of remote sensing based actual evapotranspiration estimation
1
2016
... 蒸散发(Evapotranspiration, ET)作为地表水循环和能量交换的重要组成部分[1],通过土壤-植被-大气连续体(Soil-plant-atmosphere-continuum, SPAC)的相互作用[2],深刻影响着生态系统的水分与能量平衡.蒸散发观测方法因空间尺度而异,植物个体尺度采用蒸渗仪法[3],站点尺度依赖涡度相关法[4]和波文比法[5],区域尺度及全球尺度则结合大孔径闪烁仪法[6]与遥感模型反演[7].随着遥感技术和蒸散发模型(如Priestley-Taylor模型)的发展,蒸散比(Evaporative Fraction, EF)作为衡量地表能量分配和水分循环的关键参数,已成为水分-能量耦合的重要指标[8-13]. ...
Priestley-Taylor模型参数修正及在蒸散发估算中的应用
1
2013
... 蒸散发(Evapotranspiration, ET)作为地表水循环和能量交换的重要组成部分[1],通过土壤-植被-大气连续体(Soil-plant-atmosphere-continuum, SPAC)的相互作用[2],深刻影响着生态系统的水分与能量平衡.蒸散发观测方法因空间尺度而异,植物个体尺度采用蒸渗仪法[3],站点尺度依赖涡度相关法[4]和波文比法[5],区域尺度及全球尺度则结合大孔径闪烁仪法[6]与遥感模型反演[7].随着遥感技术和蒸散发模型(如Priestley-Taylor模型)的发展,蒸散比(Evaporative Fraction, EF)作为衡量地表能量分配和水分循环的关键参数,已成为水分-能量耦合的重要指标[8-13]. ...
应用Penman-Monteith模型估算稻田蒸散的误差分析
0
2015
黄河上游参考作物蒸散量变化特征及其对气候变化的响应
0
2012
松嫩平原西部生长季参考作物蒸散发的敏感性分析
0
2008
基于MODIS数据的黄河三角洲区域蒸散发量时空分布特征
0
2009
Estimation of daily evapotranspiration over Africa using MODIS/Terra and SEVIRI/MSG data
1
2012
... 蒸散发(Evapotranspiration, ET)作为地表水循环和能量交换的重要组成部分[1],通过土壤-植被-大气连续体(Soil-plant-atmosphere-continuum, SPAC)的相互作用[2],深刻影响着生态系统的水分与能量平衡.蒸散发观测方法因空间尺度而异,植物个体尺度采用蒸渗仪法[3],站点尺度依赖涡度相关法[4]和波文比法[5],区域尺度及全球尺度则结合大孔径闪烁仪法[6]与遥感模型反演[7].随着遥感技术和蒸散发模型(如Priestley-Taylor模型)的发展,蒸散比(Evaporative Fraction, EF)作为衡量地表能量分配和水分循环的关键参数,已成为水分-能量耦合的重要指标[8-13]. ...
Evaporation and surface temperature
2
1981
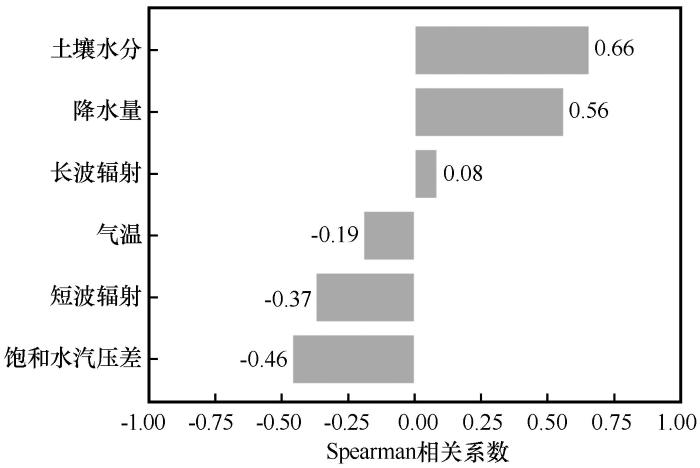
... 蒸散比为潜热通量占地表可利用能量(潜热与感热通量之和)的比例,通常用于量化地表达到恒定温度时的能量分配[14],并对大气边界层状态具有重要控制作用[15-17].蒸散比的动态受土壤水分、温度、饱和水汽压差和降水量等因子协同调控,但其主导因子在不同生态系统和气候区存在显著分异[18-20].对美国南部大平原半干旱草地的研究表明,土壤水分是蒸散比变化的主要驱动因子[21],然而同一区域其他研究却显示其影响较弱[22];半湿润草地中土壤水分对蒸散比的调控作用显著[23],而湿润草地则以叶面积指数为主导因子[24].这些争议凸显了蒸散比影响机制的复杂性,尤其在干旱半干旱区,水分胁迫条件下的蒸散比动态及其影响因素仍不明晰[25].因此,进一步深入探讨干旱半干旱区不同植被类型的蒸散比及其影响因素,有助于揭示该区域生态系统的水文和能量循环相互作用的本质. ...
... 为探讨感热通量和潜热通量的能量分配以及地表水分状况,基于能量平衡方程(Rn-G=LE+H),蒸散比定义为潜热通量(LE)占可利用能量()的比例,其计算遵循“日间稳定性假设”[14,51]. ...
温性荒漠草原生长季光伏电站地表能量交换特征
1
2024
... 蒸散比为潜热通量占地表可利用能量(潜热与感热通量之和)的比例,通常用于量化地表达到恒定温度时的能量分配[14],并对大气边界层状态具有重要控制作用[15-17].蒸散比的动态受土壤水分、温度、饱和水汽压差和降水量等因子协同调控,但其主导因子在不同生态系统和气候区存在显著分异[18-20].对美国南部大平原半干旱草地的研究表明,土壤水分是蒸散比变化的主要驱动因子[21],然而同一区域其他研究却显示其影响较弱[22];半湿润草地中土壤水分对蒸散比的调控作用显著[23],而湿润草地则以叶面积指数为主导因子[24].这些争议凸显了蒸散比影响机制的复杂性,尤其在干旱半干旱区,水分胁迫条件下的蒸散比动态及其影响因素仍不明晰[25].因此,进一步深入探讨干旱半干旱区不同植被类型的蒸散比及其影响因素,有助于揭示该区域生态系统的水文和能量循环相互作用的本质. ...
Assessment of observed and model-derived soil moisture-evaporative fraction relationships over the United States Southern Great Plains
0
2014
Probability of afternoon precipitation in eastern United States and Mexico enhanced by high evaporation
1
2011
... 蒸散比为潜热通量占地表可利用能量(潜热与感热通量之和)的比例,通常用于量化地表达到恒定温度时的能量分配[14],并对大气边界层状态具有重要控制作用[15-17].蒸散比的动态受土壤水分、温度、饱和水汽压差和降水量等因子协同调控,但其主导因子在不同生态系统和气候区存在显著分异[18-20].对美国南部大平原半干旱草地的研究表明,土壤水分是蒸散比变化的主要驱动因子[21],然而同一区域其他研究却显示其影响较弱[22];半湿润草地中土壤水分对蒸散比的调控作用显著[23],而湿润草地则以叶面积指数为主导因子[24].这些争议凸显了蒸散比影响机制的复杂性,尤其在干旱半干旱区,水分胁迫条件下的蒸散比动态及其影响因素仍不明晰[25].因此,进一步深入探讨干旱半干旱区不同植被类型的蒸散比及其影响因素,有助于揭示该区域生态系统的水文和能量循环相互作用的本质. ...
Soil and atmospheric controls on the land surface energy balance:a generalized framework for distinguishing moisture-and energy-limited evaporation regimes
1
2018
... 蒸散比为潜热通量占地表可利用能量(潜热与感热通量之和)的比例,通常用于量化地表达到恒定温度时的能量分配[14],并对大气边界层状态具有重要控制作用[15-17].蒸散比的动态受土壤水分、温度、饱和水汽压差和降水量等因子协同调控,但其主导因子在不同生态系统和气候区存在显著分异[18-20].对美国南部大平原半干旱草地的研究表明,土壤水分是蒸散比变化的主要驱动因子[21],然而同一区域其他研究却显示其影响较弱[22];半湿润草地中土壤水分对蒸散比的调控作用显著[23],而湿润草地则以叶面积指数为主导因子[24].这些争议凸显了蒸散比影响机制的复杂性,尤其在干旱半干旱区,水分胁迫条件下的蒸散比动态及其影响因素仍不明晰[25].因此,进一步深入探讨干旱半干旱区不同植被类型的蒸散比及其影响因素,有助于揭示该区域生态系统的水文和能量循环相互作用的本质. ...
The influence of land cover on surface energy partitioning and evaporative fraction regimes in the US Southern Great Plains
0
2017
Effects of soil moisture,net radiation,and atmospheric vapor pressure deficit on surface evaporation fraction at a semi-arid grass site
1
2022
... 蒸散比为潜热通量占地表可利用能量(潜热与感热通量之和)的比例,通常用于量化地表达到恒定温度时的能量分配[14],并对大气边界层状态具有重要控制作用[15-17].蒸散比的动态受土壤水分、温度、饱和水汽压差和降水量等因子协同调控,但其主导因子在不同生态系统和气候区存在显著分异[18-20].对美国南部大平原半干旱草地的研究表明,土壤水分是蒸散比变化的主要驱动因子[21],然而同一区域其他研究却显示其影响较弱[22];半湿润草地中土壤水分对蒸散比的调控作用显著[23],而湿润草地则以叶面积指数为主导因子[24].这些争议凸显了蒸散比影响机制的复杂性,尤其在干旱半干旱区,水分胁迫条件下的蒸散比动态及其影响因素仍不明晰[25].因此,进一步深入探讨干旱半干旱区不同植被类型的蒸散比及其影响因素,有助于揭示该区域生态系统的水文和能量循环相互作用的本质. ...
The sensitivity of surface fluxes to soil water content in three land surface schemes
1
2000
... 蒸散比为潜热通量占地表可利用能量(潜热与感热通量之和)的比例,通常用于量化地表达到恒定温度时的能量分配[14],并对大气边界层状态具有重要控制作用[15-17].蒸散比的动态受土壤水分、温度、饱和水汽压差和降水量等因子协同调控,但其主导因子在不同生态系统和气候区存在显著分异[18-20].对美国南部大平原半干旱草地的研究表明,土壤水分是蒸散比变化的主要驱动因子[21],然而同一区域其他研究却显示其影响较弱[22];半湿润草地中土壤水分对蒸散比的调控作用显著[23],而湿润草地则以叶面积指数为主导因子[24].这些争议凸显了蒸散比影响机制的复杂性,尤其在干旱半干旱区,水分胁迫条件下的蒸散比动态及其影响因素仍不明晰[25].因此,进一步深入探讨干旱半干旱区不同植被类型的蒸散比及其影响因素,有助于揭示该区域生态系统的水文和能量循环相互作用的本质. ...
Land-atmosphere coupling manifested in warm-season observations on the U.S.Southern Great Plains
1
2014
... 蒸散比为潜热通量占地表可利用能量(潜热与感热通量之和)的比例,通常用于量化地表达到恒定温度时的能量分配[14],并对大气边界层状态具有重要控制作用[15-17].蒸散比的动态受土壤水分、温度、饱和水汽压差和降水量等因子协同调控,但其主导因子在不同生态系统和气候区存在显著分异[18-20].对美国南部大平原半干旱草地的研究表明,土壤水分是蒸散比变化的主要驱动因子[21],然而同一区域其他研究却显示其影响较弱[22];半湿润草地中土壤水分对蒸散比的调控作用显著[23],而湿润草地则以叶面积指数为主导因子[24].这些争议凸显了蒸散比影响机制的复杂性,尤其在干旱半干旱区,水分胁迫条件下的蒸散比动态及其影响因素仍不明晰[25].因此,进一步深入探讨干旱半干旱区不同植被类型的蒸散比及其影响因素,有助于揭示该区域生态系统的水文和能量循环相互作用的本质. ...
Terrestrial and atmospheric controls on surface energy partitioning and evaporative fraction regimes over the Tibetan Plateau in the growing season
1
2021
... 蒸散比为潜热通量占地表可利用能量(潜热与感热通量之和)的比例,通常用于量化地表达到恒定温度时的能量分配[14],并对大气边界层状态具有重要控制作用[15-17].蒸散比的动态受土壤水分、温度、饱和水汽压差和降水量等因子协同调控,但其主导因子在不同生态系统和气候区存在显著分异[18-20].对美国南部大平原半干旱草地的研究表明,土壤水分是蒸散比变化的主要驱动因子[21],然而同一区域其他研究却显示其影响较弱[22];半湿润草地中土壤水分对蒸散比的调控作用显著[23],而湿润草地则以叶面积指数为主导因子[24].这些争议凸显了蒸散比影响机制的复杂性,尤其在干旱半干旱区,水分胁迫条件下的蒸散比动态及其影响因素仍不明晰[25].因此,进一步深入探讨干旱半干旱区不同植被类型的蒸散比及其影响因素,有助于揭示该区域生态系统的水文和能量循环相互作用的本质. ...
Leaf area controls on energy partitioning of a temperate mountain grassland
1
2008
... 蒸散比为潜热通量占地表可利用能量(潜热与感热通量之和)的比例,通常用于量化地表达到恒定温度时的能量分配[14],并对大气边界层状态具有重要控制作用[15-17].蒸散比的动态受土壤水分、温度、饱和水汽压差和降水量等因子协同调控,但其主导因子在不同生态系统和气候区存在显著分异[18-20].对美国南部大平原半干旱草地的研究表明,土壤水分是蒸散比变化的主要驱动因子[21],然而同一区域其他研究却显示其影响较弱[22];半湿润草地中土壤水分对蒸散比的调控作用显著[23],而湿润草地则以叶面积指数为主导因子[24].这些争议凸显了蒸散比影响机制的复杂性,尤其在干旱半干旱区,水分胁迫条件下的蒸散比动态及其影响因素仍不明晰[25].因此,进一步深入探讨干旱半干旱区不同植被类型的蒸散比及其影响因素,有助于揭示该区域生态系统的水文和能量循环相互作用的本质. ...
Investigating soil moisture-climate interactions in a changing climate:a review
1
2010
... 蒸散比为潜热通量占地表可利用能量(潜热与感热通量之和)的比例,通常用于量化地表达到恒定温度时的能量分配[14],并对大气边界层状态具有重要控制作用[15-17].蒸散比的动态受土壤水分、温度、饱和水汽压差和降水量等因子协同调控,但其主导因子在不同生态系统和气候区存在显著分异[18-20].对美国南部大平原半干旱草地的研究表明,土壤水分是蒸散比变化的主要驱动因子[21],然而同一区域其他研究却显示其影响较弱[22];半湿润草地中土壤水分对蒸散比的调控作用显著[23],而湿润草地则以叶面积指数为主导因子[24].这些争议凸显了蒸散比影响机制的复杂性,尤其在干旱半干旱区,水分胁迫条件下的蒸散比动态及其影响因素仍不明晰[25].因此,进一步深入探讨干旱半干旱区不同植被类型的蒸散比及其影响因素,有助于揭示该区域生态系统的水文和能量循环相互作用的本质. ...
Assessing vulnerability to climate change in dryland livelihood systems:conceptual challenges and interdisciplinary solutions
1
2011
... 干旱半干旱区覆盖全球陆地表面积的41%,承载约38%的人口[26],稀树草原、灌丛和草地作为该地区的典型景观类型,对气候变化具有高度敏感性[27].土壤水分是该区域蒸散发的主要限制因子[28],临界土壤水分阈值被定义为土壤水分降至某一临界值时,蒸散发开始减少并进入水分胁迫状况 [29].当土壤水分低于阈值时,蒸散发减少导致感热通量增加、地表温度升高及大气干燥度加剧 [30-31],进而通过能量再分配影响碳吸收等生态过程[32].此外,气候变化可能通过阈值效应引发生态系统功能的突变式响应,而非渐进式调整[33].因此,准确量化干旱半干旱区植物水分胁迫下的临界土壤水分阈值对于理解生态系统的水文-能量相互关系及其功能变化具有重要意义. ...
气候变化背景下干旱区碳通量的特征分析
1
2021
... 干旱半干旱区覆盖全球陆地表面积的41%,承载约38%的人口[26],稀树草原、灌丛和草地作为该地区的典型景观类型,对气候变化具有高度敏感性[27].土壤水分是该区域蒸散发的主要限制因子[28],临界土壤水分阈值被定义为土壤水分降至某一临界值时,蒸散发开始减少并进入水分胁迫状况 [29].当土壤水分低于阈值时,蒸散发减少导致感热通量增加、地表温度升高及大气干燥度加剧 [30-31],进而通过能量再分配影响碳吸收等生态过程[32].此外,气候变化可能通过阈值效应引发生态系统功能的突变式响应,而非渐进式调整[33].因此,准确量化干旱半干旱区植物水分胁迫下的临界土壤水分阈值对于理解生态系统的水文-能量相互关系及其功能变化具有重要意义. ...
土壤水分随机模型支持下的土壤水平衡研究进展
1
2008
... 干旱半干旱区覆盖全球陆地表面积的41%,承载约38%的人口[26],稀树草原、灌丛和草地作为该地区的典型景观类型,对气候变化具有高度敏感性[27].土壤水分是该区域蒸散发的主要限制因子[28],临界土壤水分阈值被定义为土壤水分降至某一临界值时,蒸散发开始减少并进入水分胁迫状况 [29].当土壤水分低于阈值时,蒸散发减少导致感热通量增加、地表温度升高及大气干燥度加剧 [30-31],进而通过能量再分配影响碳吸收等生态过程[32].此外,气候变化可能通过阈值效应引发生态系统功能的突变式响应,而非渐进式调整[33].因此,准确量化干旱半干旱区植物水分胁迫下的临界土壤水分阈值对于理解生态系统的水文-能量相互关系及其功能变化具有重要意义. ...
Satellite-based assessment of land surface energy partitioning-soil moisture relationships and effects of confounding variables
2
2019
... 干旱半干旱区覆盖全球陆地表面积的41%,承载约38%的人口[26],稀树草原、灌丛和草地作为该地区的典型景观类型,对气候变化具有高度敏感性[27].土壤水分是该区域蒸散发的主要限制因子[28],临界土壤水分阈值被定义为土壤水分降至某一临界值时,蒸散发开始减少并进入水分胁迫状况 [29].当土壤水分低于阈值时,蒸散发减少导致感热通量增加、地表温度升高及大气干燥度加剧 [30-31],进而通过能量再分配影响碳吸收等生态过程[32].此外,气候变化可能通过阈值效应引发生态系统功能的突变式响应,而非渐进式调整[33].因此,准确量化干旱半干旱区植物水分胁迫下的临界土壤水分阈值对于理解生态系统的水文-能量相互关系及其功能变化具有重要意义. ...
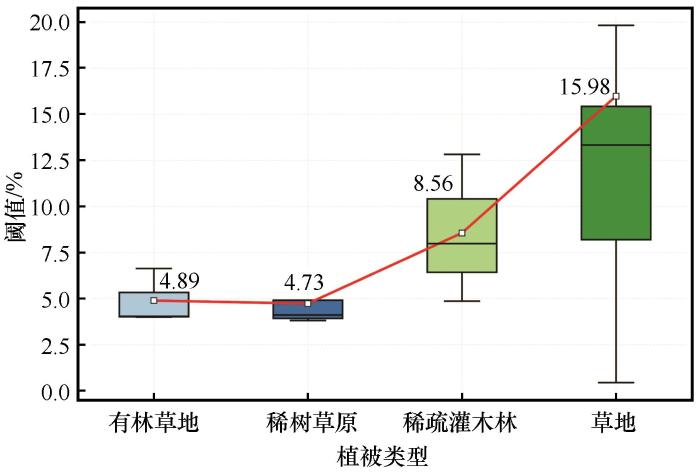
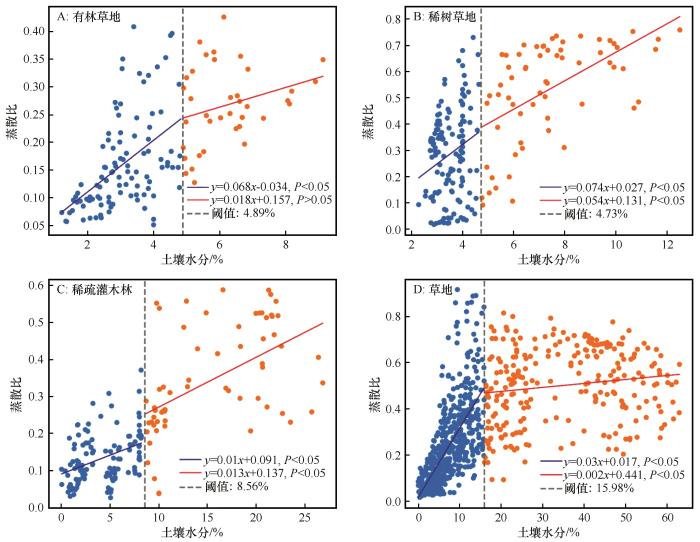
... 干旱半干旱区4种典型植被类型土壤水分阈值均值4.89%~15.98%(图5),这一结果与Fu等[40]基于涡度相关观测数据得到的12%,Buitink等[58]和Feldman等[29]基于卫星观测数据得到的16%、14%相近,但显著低于中国西北干旱区高阈值站点[59](景阳岭站、垭口站和大野口站土壤水分阈值分别为45.1%、40%和43.2%).这一差异可能由地区间土壤质地、植被类型及其环境因素差异引起[40],特别是在土壤持水能力、水分获取深度和气孔调节机制方面[60].草地的土壤水分阈值(15.98%)显著高于其他生态系统,主要原因可能在于其高植被覆盖率、高土壤有机碳含量和高叶面积指数[61-62],这些特征通过增强土壤的持水能力和蒸腾需求,提高水分利用阈值;而土壤水分阈值较低的稀树草原(4.73%)和有林草地(4.89%)则可能依赖深根系获取深层水分[63-65],并辅以快速气孔调节机制,形成低阈值-高韧性的水分利用策略[66].Fu等[41]通过计算草地和稀树草原的冠层导电率(Gc)随土壤水分降低的变化,发现稀树草原的气孔关闭反应更为迅速,以防止水分过度损失.然而,这种快速关闭虽然减少水分流失,但超过阈值后,生态系统功能可能迅速崩溃[67-68].值得注意的是,沙质土壤的低持水能力可能进一步降低稀疏灌木林的土壤水分阈值(8.56%)[61].以上分析表明不同植被类型展现出各自独特的水分利用策略,反映出它们对干旱环境存在不同的适应性,这些发现为干旱半干旱区的水资源管理和生态系统恢复提供重要依据. ...
Understanding hydrometeorology using global models
1
2004
... 干旱半干旱区覆盖全球陆地表面积的41%,承载约38%的人口[26],稀树草原、灌丛和草地作为该地区的典型景观类型,对气候变化具有高度敏感性[27].土壤水分是该区域蒸散发的主要限制因子[28],临界土壤水分阈值被定义为土壤水分降至某一临界值时,蒸散发开始减少并进入水分胁迫状况 [29].当土壤水分低于阈值时,蒸散发减少导致感热通量增加、地表温度升高及大气干燥度加剧 [30-31],进而通过能量再分配影响碳吸收等生态过程[32].此外,气候变化可能通过阈值效应引发生态系统功能的突变式响应,而非渐进式调整[33].因此,准确量化干旱半干旱区植物水分胁迫下的临界土壤水分阈值对于理解生态系统的水文-能量相互关系及其功能变化具有重要意义. ...
Coupling between the terrestrial carbon and water cycles:a review
1
2019
... 干旱半干旱区覆盖全球陆地表面积的41%,承载约38%的人口[26],稀树草原、灌丛和草地作为该地区的典型景观类型,对气候变化具有高度敏感性[27].土壤水分是该区域蒸散发的主要限制因子[28],临界土壤水分阈值被定义为土壤水分降至某一临界值时,蒸散发开始减少并进入水分胁迫状况 [29].当土壤水分低于阈值时,蒸散发减少导致感热通量增加、地表温度升高及大气干燥度加剧 [30-31],进而通过能量再分配影响碳吸收等生态过程[32].此外,气候变化可能通过阈值效应引发生态系统功能的突变式响应,而非渐进式调整[33].因此,准确量化干旱半干旱区植物水分胁迫下的临界土壤水分阈值对于理解生态系统的水文-能量相互关系及其功能变化具有重要意义. ...
Evidence for soil water control on carbon and water dynamics in European forests during the extremely dry year:2003
1
2007
... 干旱半干旱区覆盖全球陆地表面积的41%,承载约38%的人口[26],稀树草原、灌丛和草地作为该地区的典型景观类型,对气候变化具有高度敏感性[27].土壤水分是该区域蒸散发的主要限制因子[28],临界土壤水分阈值被定义为土壤水分降至某一临界值时,蒸散发开始减少并进入水分胁迫状况 [29].当土壤水分低于阈值时,蒸散发减少导致感热通量增加、地表温度升高及大气干燥度加剧 [30-31],进而通过能量再分配影响碳吸收等生态过程[32].此外,气候变化可能通过阈值效应引发生态系统功能的突变式响应,而非渐进式调整[33].因此,准确量化干旱半干旱区植物水分胁迫下的临界土壤水分阈值对于理解生态系统的水文-能量相互关系及其功能变化具有重要意义. ...
Ecological thresholds:The key to successful environmental management or an important concept with no practical application?
1
2006
... 干旱半干旱区覆盖全球陆地表面积的41%,承载约38%的人口[26],稀树草原、灌丛和草地作为该地区的典型景观类型,对气候变化具有高度敏感性[27].土壤水分是该区域蒸散发的主要限制因子[28],临界土壤水分阈值被定义为土壤水分降至某一临界值时,蒸散发开始减少并进入水分胁迫状况 [29].当土壤水分低于阈值时,蒸散发减少导致感热通量增加、地表温度升高及大气干燥度加剧 [30-31],进而通过能量再分配影响碳吸收等生态过程[32].此外,气候变化可能通过阈值效应引发生态系统功能的突变式响应,而非渐进式调整[33].因此,准确量化干旱半干旱区植物水分胁迫下的临界土壤水分阈值对于理解生态系统的水文-能量相互关系及其功能变化具有重要意义. ...
Investigating the coupling relationship between soil moisture and evaporative fraction over China's transitional climate zone
1
2023
... 蒸散比-土壤水分(EF-SM)关系是量化临界土壤水分阈值的重要依据[34],而阈值识别方法在其中发挥着至关重要的作用[35].分段回归模型(如铰链模型)因其对突变点有较好的捕捉能力,被广泛应用于EF-SM关系解析中[36-39].基于全球尺度研究证实,铰链模型可有效区分生态系统的水分限制与能量限制状态[40-41].然而,单一模型难以全面刻画不同植被类型的动态响应特征.为此,本研究集成铰链模型与其他3种分段回归模型,结合赤池信息准则(AIC)优选最优拟合模型[42-43],系统解析干旱半干旱区蒸散比与土壤水分之间的关系. ...
生态阈值确定方法综述
1
2021
... 蒸散比-土壤水分(EF-SM)关系是量化临界土壤水分阈值的重要依据[34],而阈值识别方法在其中发挥着至关重要的作用[35].分段回归模型(如铰链模型)因其对突变点有较好的捕捉能力,被广泛应用于EF-SM关系解析中[36-39].基于全球尺度研究证实,铰链模型可有效区分生态系统的水分限制与能量限制状态[40-41].然而,单一模型难以全面刻画不同植被类型的动态响应特征.为此,本研究集成铰链模型与其他3种分段回归模型,结合赤池信息准则(AIC)优选最优拟合模型[42-43],系统解析干旱半干旱区蒸散比与土壤水分之间的关系. ...
chngpt:Threshold regression model estimation and inference
2
2017
... 蒸散比-土壤水分(EF-SM)关系是量化临界土壤水分阈值的重要依据[34],而阈值识别方法在其中发挥着至关重要的作用[35].分段回归模型(如铰链模型)因其对突变点有较好的捕捉能力,被广泛应用于EF-SM关系解析中[36-39].基于全球尺度研究证实,铰链模型可有效区分生态系统的水分限制与能量限制状态[40-41].然而,单一模型难以全面刻画不同植被类型的动态响应特征.为此,本研究集成铰链模型与其他3种分段回归模型,结合赤池信息准则(AIC)优选最优拟合模型[42-43],系统解析干旱半干旱区蒸散比与土壤水分之间的关系. ...
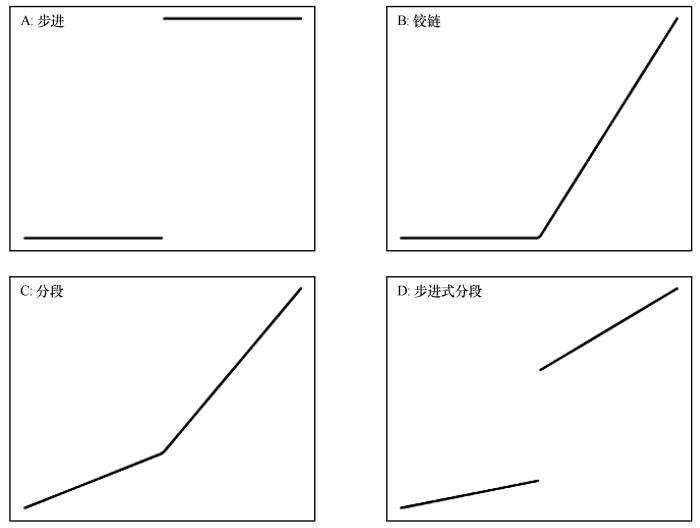
... 为量化蒸散比-土壤水分之间的阈值,采用阈值回归模型(Threshold regression models)[36]进行识别(https://cran.r-project.org/web/packages/chngpt/).该模型包括4种模式:步进模型、铰链模型、分段模型及步进式分段模型(图1).其中,铰链和分段模型可有效识别连续变化阈值,步进式分段模型结合了步进模型和分段模型的特点,适用于大量预测变量和响应变量急剧不连续的阈值识别.因此,研究采用上述4种模型捕捉蒸散比与土壤水分之间的关系. ...
Vegetation controls on surface heat flux partitioning,and land-atmosphere coupling
0
2015
基于蒸渗仪实测的参考作物蒸散发模型北京地区适用性评价
0
2014
Can surface soil moisture information identify evapotranspiration regime transitions?
1
2022
... 蒸散比-土壤水分(EF-SM)关系是量化临界土壤水分阈值的重要依据[34],而阈值识别方法在其中发挥着至关重要的作用[35].分段回归模型(如铰链模型)因其对突变点有较好的捕捉能力,被广泛应用于EF-SM关系解析中[36-39].基于全球尺度研究证实,铰链模型可有效区分生态系统的水分限制与能量限制状态[40-41].然而,单一模型难以全面刻画不同植被类型的动态响应特征.为此,本研究集成铰链模型与其他3种分段回归模型,结合赤池信息准则(AIC)优选最优拟合模型[42-43],系统解析干旱半干旱区蒸散比与土壤水分之间的关系. ...
Global critical soil moisture thresholds of plant water stress
3
2024
... 蒸散比-土壤水分(EF-SM)关系是量化临界土壤水分阈值的重要依据[34],而阈值识别方法在其中发挥着至关重要的作用[35].分段回归模型(如铰链模型)因其对突变点有较好的捕捉能力,被广泛应用于EF-SM关系解析中[36-39].基于全球尺度研究证实,铰链模型可有效区分生态系统的水分限制与能量限制状态[40-41].然而,单一模型难以全面刻画不同植被类型的动态响应特征.为此,本研究集成铰链模型与其他3种分段回归模型,结合赤池信息准则(AIC)优选最优拟合模型[42-43],系统解析干旱半干旱区蒸散比与土壤水分之间的关系. ...
... 干旱半干旱区4种典型植被类型土壤水分阈值均值4.89%~15.98%(图5),这一结果与Fu等[40]基于涡度相关观测数据得到的12%,Buitink等[58]和Feldman等[29]基于卫星观测数据得到的16%、14%相近,但显著低于中国西北干旱区高阈值站点[59](景阳岭站、垭口站和大野口站土壤水分阈值分别为45.1%、40%和43.2%).这一差异可能由地区间土壤质地、植被类型及其环境因素差异引起[40],特别是在土壤持水能力、水分获取深度和气孔调节机制方面[60].草地的土壤水分阈值(15.98%)显著高于其他生态系统,主要原因可能在于其高植被覆盖率、高土壤有机碳含量和高叶面积指数[61-62],这些特征通过增强土壤的持水能力和蒸腾需求,提高水分利用阈值;而土壤水分阈值较低的稀树草原(4.73%)和有林草地(4.89%)则可能依赖深根系获取深层水分[63-65],并辅以快速气孔调节机制,形成低阈值-高韧性的水分利用策略[66].Fu等[41]通过计算草地和稀树草原的冠层导电率(Gc)随土壤水分降低的变化,发现稀树草原的气孔关闭反应更为迅速,以防止水分过度损失.然而,这种快速关闭虽然减少水分流失,但超过阈值后,生态系统功能可能迅速崩溃[67-68].值得注意的是,沙质土壤的低持水能力可能进一步降低稀疏灌木林的土壤水分阈值(8.56%)[61].以上分析表明不同植被类型展现出各自独特的水分利用策略,反映出它们对干旱环境存在不同的适应性,这些发现为干旱半干旱区的水资源管理和生态系统恢复提供重要依据. ...
... [40],特别是在土壤持水能力、水分获取深度和气孔调节机制方面[60].草地的土壤水分阈值(15.98%)显著高于其他生态系统,主要原因可能在于其高植被覆盖率、高土壤有机碳含量和高叶面积指数[61-62],这些特征通过增强土壤的持水能力和蒸腾需求,提高水分利用阈值;而土壤水分阈值较低的稀树草原(4.73%)和有林草地(4.89%)则可能依赖深根系获取深层水分[63-65],并辅以快速气孔调节机制,形成低阈值-高韧性的水分利用策略[66].Fu等[41]通过计算草地和稀树草原的冠层导电率(Gc)随土壤水分降低的变化,发现稀树草原的气孔关闭反应更为迅速,以防止水分过度损失.然而,这种快速关闭虽然减少水分流失,但超过阈值后,生态系统功能可能迅速崩溃[67-68].值得注意的是,沙质土壤的低持水能力可能进一步降低稀疏灌木林的土壤水分阈值(8.56%)[61].以上分析表明不同植被类型展现出各自独特的水分利用策略,反映出它们对干旱环境存在不同的适应性,这些发现为干旱半干旱区的水资源管理和生态系统恢复提供重要依据. ...
Critical soil moisture thresholds of plant water stress in terrestrial ecosystems
3
2022
... 蒸散比-土壤水分(EF-SM)关系是量化临界土壤水分阈值的重要依据[34],而阈值识别方法在其中发挥着至关重要的作用[35].分段回归模型(如铰链模型)因其对突变点有较好的捕捉能力,被广泛应用于EF-SM关系解析中[36-39].基于全球尺度研究证实,铰链模型可有效区分生态系统的水分限制与能量限制状态[40-41].然而,单一模型难以全面刻画不同植被类型的动态响应特征.为此,本研究集成铰链模型与其他3种分段回归模型,结合赤池信息准则(AIC)优选最优拟合模型[42-43],系统解析干旱半干旱区蒸散比与土壤水分之间的关系. ...
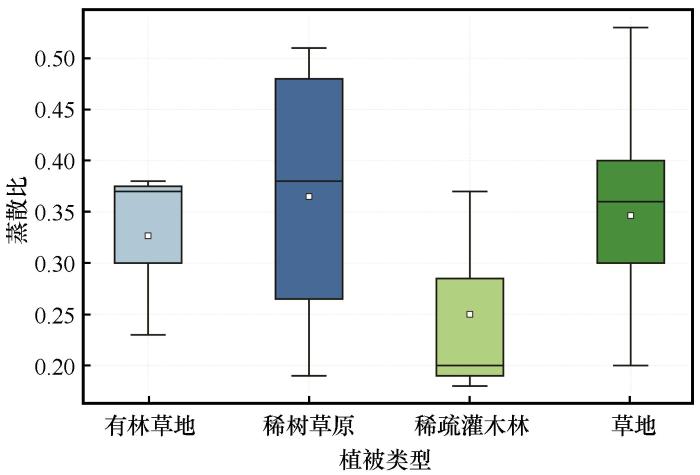
... 结果表明,干旱半干旱区月均蒸散比为0.31(表3),显著低于Chen等[53]基于涡度相关数据计算的全球多年平均蒸散比(0.52),但与该气候区多数站点观测值(<0.3)一致[53],印证了区域水分限制下植被蒸散作用普遍受限的特征[41].不同植被类型的蒸散比呈现显著分异 [53],其中,稀树草原的蒸散比季节波动最大(Cv=0.65),这可能与其独特的旱雨季交替模式密切相关[54].尽管深根系可能缓解旱季水分胁迫(通过汲取深层土壤水分)[55-56],但雨季降水的波动性仍可加剧蒸散比波动;相对而言,草地较稳定的表层水分供给使其波动性最低(Cv=0.43),凸显植被水分获取策略对蒸散动态的调控作用 [57]. ...
... 干旱半干旱区4种典型植被类型土壤水分阈值均值4.89%~15.98%(图5),这一结果与Fu等[40]基于涡度相关观测数据得到的12%,Buitink等[58]和Feldman等[29]基于卫星观测数据得到的16%、14%相近,但显著低于中国西北干旱区高阈值站点[59](景阳岭站、垭口站和大野口站土壤水分阈值分别为45.1%、40%和43.2%).这一差异可能由地区间土壤质地、植被类型及其环境因素差异引起[40],特别是在土壤持水能力、水分获取深度和气孔调节机制方面[60].草地的土壤水分阈值(15.98%)显著高于其他生态系统,主要原因可能在于其高植被覆盖率、高土壤有机碳含量和高叶面积指数[61-62],这些特征通过增强土壤的持水能力和蒸腾需求,提高水分利用阈值;而土壤水分阈值较低的稀树草原(4.73%)和有林草地(4.89%)则可能依赖深根系获取深层水分[63-65],并辅以快速气孔调节机制,形成低阈值-高韧性的水分利用策略[66].Fu等[41]通过计算草地和稀树草原的冠层导电率(Gc)随土壤水分降低的变化,发现稀树草原的气孔关闭反应更为迅速,以防止水分过度损失.然而,这种快速关闭虽然减少水分流失,但超过阈值后,生态系统功能可能迅速崩溃[67-68].值得注意的是,沙质土壤的低持水能力可能进一步降低稀疏灌木林的土壤水分阈值(8.56%)[61].以上分析表明不同植被类型展现出各自独特的水分利用策略,反映出它们对干旱环境存在不同的适应性,这些发现为干旱半干旱区的水资源管理和生态系统恢复提供重要依据. ...
Piecewise regression:a tool for identifying ecological thresholds
2
2003
... 蒸散比-土壤水分(EF-SM)关系是量化临界土壤水分阈值的重要依据[34],而阈值识别方法在其中发挥着至关重要的作用[35].分段回归模型(如铰链模型)因其对突变点有较好的捕捉能力,被广泛应用于EF-SM关系解析中[36-39].基于全球尺度研究证实,铰链模型可有效区分生态系统的水分限制与能量限制状态[40-41].然而,单一模型难以全面刻画不同植被类型的动态响应特征.为此,本研究集成铰链模型与其他3种分段回归模型,结合赤池信息准则(AIC)优选最优拟合模型[42-43],系统解析干旱半干旱区蒸散比与土壤水分之间的关系. ...
... 为确保模型的稳健性,通过马哈拉诺比斯距离法(Mahalanobis Distance)剔除异常值后,采用赤池信息准则(AIC)选择最优模型[42].AIC值越小,模型拟合效果越好. ...
A new look at the statistical model identification
1
1974
... 蒸散比-土壤水分(EF-SM)关系是量化临界土壤水分阈值的重要依据[34],而阈值识别方法在其中发挥着至关重要的作用[35].分段回归模型(如铰链模型)因其对突变点有较好的捕捉能力,被广泛应用于EF-SM关系解析中[36-39].基于全球尺度研究证实,铰链模型可有效区分生态系统的水分限制与能量限制状态[40-41].然而,单一模型难以全面刻画不同植被类型的动态响应特征.为此,本研究集成铰链模型与其他3种分段回归模型,结合赤池信息准则(AIC)优选最优拟合模型[42-43],系统解析干旱半干旱区蒸散比与土壤水分之间的关系. ...
The FLUXNET2015 dataset and the ONEFlux processing pipeline for eddy covariance data
1
2020
... 数据来源于FLUXNET 2015数据集半小时涡度相关系统长期观测数据(https://fluxnet.org/data/fluxnet2015-dataset/)[44-45],基于国际地圈-生物圈计划[46](International Geosphere-Biosphere Program,IGBP)的土地覆盖分类体系,选取草地(Grassland, GRA)、稀疏灌木林(Open shrubland, OSH)、稀树草原(Savanna, SAV)和有林草地(Woody savanna, WSA)四类典型生态系统的观测站点(表1).为探究蒸散比与土壤水分及气象因子的响应关系,揭示不同植被类型下蒸散比与土壤水分阈值的分异特征,采用严格的质量控制对全球279个通量塔站点进行筛选:首先剔除缺乏土壤水分连续观测记录的站点;其次排除湿地生态系统站点(湿地生态系统地下水位较浅,且通常情况下较少发生土壤水分胁迫);最终基于干旱指数(Arid Index,AI)>0.65的标准筛选出干旱半干旱区内的24个典型站点作为研究对象(表2). ...
FLUXNET:a new tool to study the temporal and spatial variability of ecosystem-scale carbon dioxide,water vapor,and energy flux densities
1
2001
... 数据来源于FLUXNET 2015数据集半小时涡度相关系统长期观测数据(https://fluxnet.org/data/fluxnet2015-dataset/)[44-45],基于国际地圈-生物圈计划[46](International Geosphere-Biosphere Program,IGBP)的土地覆盖分类体系,选取草地(Grassland, GRA)、稀疏灌木林(Open shrubland, OSH)、稀树草原(Savanna, SAV)和有林草地(Woody savanna, WSA)四类典型生态系统的观测站点(表1).为探究蒸散比与土壤水分及气象因子的响应关系,揭示不同植被类型下蒸散比与土壤水分阈值的分异特征,采用严格的质量控制对全球279个通量塔站点进行筛选:首先剔除缺乏土壤水分连续观测记录的站点;其次排除湿地生态系统站点(湿地生态系统地下水位较浅,且通常情况下较少发生土壤水分胁迫);最终基于干旱指数(Arid Index,AI)>0.65的标准筛选出干旱半干旱区内的24个典型站点作为研究对象(表2). ...
Development of a global land cover characteristics database and IGBP DISCover from 1 km AVHRR data
1
2000
... 数据来源于FLUXNET 2015数据集半小时涡度相关系统长期观测数据(https://fluxnet.org/data/fluxnet2015-dataset/)[44-45],基于国际地圈-生物圈计划[46](International Geosphere-Biosphere Program,IGBP)的土地覆盖分类体系,选取草地(Grassland, GRA)、稀疏灌木林(Open shrubland, OSH)、稀树草原(Savanna, SAV)和有林草地(Woody savanna, WSA)四类典型生态系统的观测站点(表1).为探究蒸散比与土壤水分及气象因子的响应关系,揭示不同植被类型下蒸散比与土壤水分阈值的分异特征,采用严格的质量控制对全球279个通量塔站点进行筛选:首先剔除缺乏土壤水分连续观测记录的站点;其次排除湿地生态系统站点(湿地生态系统地下水位较浅,且通常情况下较少发生土壤水分胁迫);最终基于干旱指数(Arid Index,AI)>0.65的标准筛选出干旱半干旱区内的24个典型站点作为研究对象(表2). ...
On the separation of net ecosystem exchange into assimilation and ecosystem respiration:review and improved algorithm
1
2005
... 研究选取的核心指标包括感热通量(W·m-2)、潜热通量(W·m-2)、土壤水分(%)、空气温度(℃)、短波辐射(W·m-2)、长波辐射(W·m-2)、饱和水汽压差(hPa)和降水量(mm).其中地表通量数据(感热通量、潜热通量)基于涡度相关系统原位测量,采用边缘分布取样法(Marginal Distribution Sampling, MDS)进行缺失值插补与能量平衡校正 [47];其他气象要素则通过融合欧洲中心再分析降尺度数据与MDS优化后的观测数据,形成具有时空连续性的组合数据集[48].所有数据均经过严格的质量控制与误差校正[49-50]. ...
Filling the gaps in meteorological continuous data measured at FLUXNET sites with ERA-Interim reanalysis
1
2015
... 研究选取的核心指标包括感热通量(W·m-2)、潜热通量(W·m-2)、土壤水分(%)、空气温度(℃)、短波辐射(W·m-2)、长波辐射(W·m-2)、饱和水汽压差(hPa)和降水量(mm).其中地表通量数据(感热通量、潜热通量)基于涡度相关系统原位测量,采用边缘分布取样法(Marginal Distribution Sampling, MDS)进行缺失值插补与能量平衡校正 [47];其他气象要素则通过融合欧洲中心再分析降尺度数据与MDS优化后的观测数据,形成具有时空连续性的组合数据集[48].所有数据均经过严格的质量控制与误差校正[49-50]. ...
Pairing FLUXNET sites to validate model representations of land-use/land-cover change
1
2018
... 研究选取的核心指标包括感热通量(W·m-2)、潜热通量(W·m-2)、土壤水分(%)、空气温度(℃)、短波辐射(W·m-2)、长波辐射(W·m-2)、饱和水汽压差(hPa)和降水量(mm).其中地表通量数据(感热通量、潜热通量)基于涡度相关系统原位测量,采用边缘分布取样法(Marginal Distribution Sampling, MDS)进行缺失值插补与能量平衡校正 [47];其他气象要素则通过融合欧洲中心再分析降尺度数据与MDS优化后的观测数据,形成具有时空连续性的组合数据集[48].所有数据均经过严格的质量控制与误差校正[49-50]. ...
Deforestation intensifies daily temperature variability in the northern extratropics
1
2022
... 研究选取的核心指标包括感热通量(W·m-2)、潜热通量(W·m-2)、土壤水分(%)、空气温度(℃)、短波辐射(W·m-2)、长波辐射(W·m-2)、饱和水汽压差(hPa)和降水量(mm).其中地表通量数据(感热通量、潜热通量)基于涡度相关系统原位测量,采用边缘分布取样法(Marginal Distribution Sampling, MDS)进行缺失值插补与能量平衡校正 [47];其他气象要素则通过融合欧洲中心再分析降尺度数据与MDS优化后的观测数据,形成具有时空连续性的组合数据集[48].所有数据均经过严格的质量控制与误差校正[49-50]. ...
Conservation and variability of the evaporative fraction during the daytime
3
1996
... 遵循蒸散比“日间稳定性假设”[51],本研究对数据处理流程进行标准化:除降水量直接采用日累计值外,其余参数均提取当地标准时间09:00—16:00的白天时段观测值计算日平均值,并通过时间序列聚合形成月尺度数据集.该处理方法有效消除了夜间蒸散发过程的不确定性,同时保证数据序列的时间一致性. ...
... 为探讨感热通量和潜热通量的能量分配以及地表水分状况,基于能量平衡方程(Rn-G=LE+H),蒸散比定义为潜热通量(LE)占可利用能量()的比例,其计算遵循“日间稳定性假设”[14,51]. ...
... 式中:EF为蒸散比(无量纲,范围为0~1);LE为潜热通量;H为感热通量;Rn为净辐射;G为土壤热通量;单位均为W·m-2.EF值表征水分状况,低值(如沙漠环境)反映感热通量主导的干旱特征,高值(如湿地环境)指示潜热通量主导的湿润状态[51]. ...
Version 3 of the global aridity index and potential evapotranspiration database
1
2022
... 注:本研究用于蒸散比的土壤水分阈值关系评估.干旱指数来源于全球干旱指数数据库(Global-AI_PET_v3,2022)[52]:0.03~0.2干旱区,0.2~0.5半干旱区,0.5~0.65半干旱半湿润区,>0.65湿润区. ...
Analysis of spatiotemporal distribution of evaporation fractions of different vegetation types based on FLUXNET site
3
2024
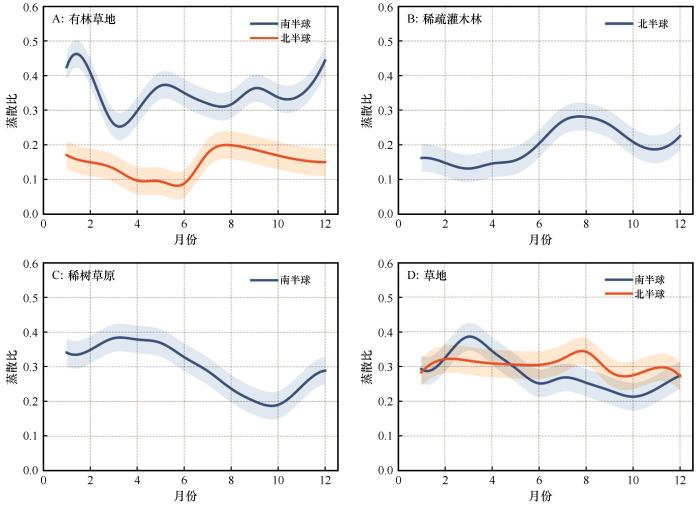
... 结果表明,干旱半干旱区月均蒸散比为0.31(表3),显著低于Chen等[53]基于涡度相关数据计算的全球多年平均蒸散比(0.52),但与该气候区多数站点观测值(<0.3)一致[53],印证了区域水分限制下植被蒸散作用普遍受限的特征[41].不同植被类型的蒸散比呈现显著分异 [53],其中,稀树草原的蒸散比季节波动最大(Cv=0.65),这可能与其独特的旱雨季交替模式密切相关[54].尽管深根系可能缓解旱季水分胁迫(通过汲取深层土壤水分)[55-56],但雨季降水的波动性仍可加剧蒸散比波动;相对而言,草地较稳定的表层水分供给使其波动性最低(Cv=0.43),凸显植被水分获取策略对蒸散动态的调控作用 [57]. ...
... [53],印证了区域水分限制下植被蒸散作用普遍受限的特征[41].不同植被类型的蒸散比呈现显著分异 [53],其中,稀树草原的蒸散比季节波动最大(Cv=0.65),这可能与其独特的旱雨季交替模式密切相关[54].尽管深根系可能缓解旱季水分胁迫(通过汲取深层土壤水分)[55-56],但雨季降水的波动性仍可加剧蒸散比波动;相对而言,草地较稳定的表层水分供给使其波动性最低(Cv=0.43),凸显植被水分获取策略对蒸散动态的调控作用 [57]. ...
... [53],其中,稀树草原的蒸散比季节波动最大(Cv=0.65),这可能与其独特的旱雨季交替模式密切相关[54].尽管深根系可能缓解旱季水分胁迫(通过汲取深层土壤水分)[55-56],但雨季降水的波动性仍可加剧蒸散比波动;相对而言,草地较稳定的表层水分供给使其波动性最低(Cv=0.43),凸显植被水分获取策略对蒸散动态的调控作用 [57]. ...
What limits trees in C4 grasslands and savannas?
1
2008
... 结果表明,干旱半干旱区月均蒸散比为0.31(表3),显著低于Chen等[53]基于涡度相关数据计算的全球多年平均蒸散比(0.52),但与该气候区多数站点观测值(<0.3)一致[53],印证了区域水分限制下植被蒸散作用普遍受限的特征[41].不同植被类型的蒸散比呈现显著分异 [53],其中,稀树草原的蒸散比季节波动最大(Cv=0.65),这可能与其独特的旱雨季交替模式密切相关[54].尽管深根系可能缓解旱季水分胁迫(通过汲取深层土壤水分)[55-56],但雨季降水的波动性仍可加剧蒸散比波动;相对而言,草地较稳定的表层水分供给使其波动性最低(Cv=0.43),凸显植被水分获取策略对蒸散动态的调控作用 [57]. ...
Hydrologic regulation of plant rooting depth
1
2017
... 结果表明,干旱半干旱区月均蒸散比为0.31(表3),显著低于Chen等[53]基于涡度相关数据计算的全球多年平均蒸散比(0.52),但与该气候区多数站点观测值(<0.3)一致[53],印证了区域水分限制下植被蒸散作用普遍受限的特征[41].不同植被类型的蒸散比呈现显著分异 [53],其中,稀树草原的蒸散比季节波动最大(Cv=0.65),这可能与其独特的旱雨季交替模式密切相关[54].尽管深根系可能缓解旱季水分胁迫(通过汲取深层土壤水分)[55-56],但雨季降水的波动性仍可加剧蒸散比波动;相对而言,草地较稳定的表层水分供给使其波动性最低(Cv=0.43),凸显植被水分获取策略对蒸散动态的调控作用 [57]. ...
Rooting depth as a key woody functional trait in savannas
1
2020
... 结果表明,干旱半干旱区月均蒸散比为0.31(表3),显著低于Chen等[53]基于涡度相关数据计算的全球多年平均蒸散比(0.52),但与该气候区多数站点观测值(<0.3)一致[53],印证了区域水分限制下植被蒸散作用普遍受限的特征[41].不同植被类型的蒸散比呈现显著分异 [53],其中,稀树草原的蒸散比季节波动最大(Cv=0.65),这可能与其独特的旱雨季交替模式密切相关[54].尽管深根系可能缓解旱季水分胁迫(通过汲取深层土壤水分)[55-56],但雨季降水的波动性仍可加剧蒸散比波动;相对而言,草地较稳定的表层水分供给使其波动性最低(Cv=0.43),凸显植被水分获取策略对蒸散动态的调控作用 [57]. ...
Root traits of grasslands rapidly respond to climate change,while community biomass mainly depends on functional composition
1
2023
... 结果表明,干旱半干旱区月均蒸散比为0.31(表3),显著低于Chen等[53]基于涡度相关数据计算的全球多年平均蒸散比(0.52),但与该气候区多数站点观测值(<0.3)一致[53],印证了区域水分限制下植被蒸散作用普遍受限的特征[41].不同植被类型的蒸散比呈现显著分异 [53],其中,稀树草原的蒸散比季节波动最大(Cv=0.65),这可能与其独特的旱雨季交替模式密切相关[54].尽管深根系可能缓解旱季水分胁迫(通过汲取深层土壤水分)[55-56],但雨季降水的波动性仍可加剧蒸散比波动;相对而言,草地较稳定的表层水分供给使其波动性最低(Cv=0.43),凸显植被水分获取策略对蒸散动态的调控作用 [57]. ...
Anatomy of the 2018 agricultural drought in the Netherlands using in situ soil moisture and satellite vegetation indices
1
2020
... 干旱半干旱区4种典型植被类型土壤水分阈值均值4.89%~15.98%(图5),这一结果与Fu等[40]基于涡度相关观测数据得到的12%,Buitink等[58]和Feldman等[29]基于卫星观测数据得到的16%、14%相近,但显著低于中国西北干旱区高阈值站点[59](景阳岭站、垭口站和大野口站土壤水分阈值分别为45.1%、40%和43.2%).这一差异可能由地区间土壤质地、植被类型及其环境因素差异引起[40],特别是在土壤持水能力、水分获取深度和气孔调节机制方面[60].草地的土壤水分阈值(15.98%)显著高于其他生态系统,主要原因可能在于其高植被覆盖率、高土壤有机碳含量和高叶面积指数[61-62],这些特征通过增强土壤的持水能力和蒸腾需求,提高水分利用阈值;而土壤水分阈值较低的稀树草原(4.73%)和有林草地(4.89%)则可能依赖深根系获取深层水分[63-65],并辅以快速气孔调节机制,形成低阈值-高韧性的水分利用策略[66].Fu等[41]通过计算草地和稀树草原的冠层导电率(Gc)随土壤水分降低的变化,发现稀树草原的气孔关闭反应更为迅速,以防止水分过度损失.然而,这种快速关闭虽然减少水分流失,但超过阈值后,生态系统功能可能迅速崩溃[67-68].值得注意的是,沙质土壤的低持水能力可能进一步降低稀疏灌木林的土壤水分阈值(8.56%)[61].以上分析表明不同植被类型展现出各自独特的水分利用策略,反映出它们对干旱环境存在不同的适应性,这些发现为干旱半干旱区的水资源管理和生态系统恢复提供重要依据. ...
干旱半干旱地区土壤水分胁迫条件下的蒸散发估算研究
1
2022
... 干旱半干旱区4种典型植被类型土壤水分阈值均值4.89%~15.98%(图5),这一结果与Fu等[40]基于涡度相关观测数据得到的12%,Buitink等[58]和Feldman等[29]基于卫星观测数据得到的16%、14%相近,但显著低于中国西北干旱区高阈值站点[59](景阳岭站、垭口站和大野口站土壤水分阈值分别为45.1%、40%和43.2%).这一差异可能由地区间土壤质地、植被类型及其环境因素差异引起[40],特别是在土壤持水能力、水分获取深度和气孔调节机制方面[60].草地的土壤水分阈值(15.98%)显著高于其他生态系统,主要原因可能在于其高植被覆盖率、高土壤有机碳含量和高叶面积指数[61-62],这些特征通过增强土壤的持水能力和蒸腾需求,提高水分利用阈值;而土壤水分阈值较低的稀树草原(4.73%)和有林草地(4.89%)则可能依赖深根系获取深层水分[63-65],并辅以快速气孔调节机制,形成低阈值-高韧性的水分利用策略[66].Fu等[41]通过计算草地和稀树草原的冠层导电率(Gc)随土壤水分降低的变化,发现稀树草原的气孔关闭反应更为迅速,以防止水分过度损失.然而,这种快速关闭虽然减少水分流失,但超过阈值后,生态系统功能可能迅速崩溃[67-68].值得注意的是,沙质土壤的低持水能力可能进一步降低稀疏灌木林的土壤水分阈值(8.56%)[61].以上分析表明不同植被类型展现出各自独特的水分利用策略,反映出它们对干旱环境存在不同的适应性,这些发现为干旱半干旱区的水资源管理和生态系统恢复提供重要依据. ...
干旱半干旱草地生态系统与土壤水分关系研究进展
1
2016
... 干旱半干旱区4种典型植被类型土壤水分阈值均值4.89%~15.98%(图5),这一结果与Fu等[40]基于涡度相关观测数据得到的12%,Buitink等[58]和Feldman等[29]基于卫星观测数据得到的16%、14%相近,但显著低于中国西北干旱区高阈值站点[59](景阳岭站、垭口站和大野口站土壤水分阈值分别为45.1%、40%和43.2%).这一差异可能由地区间土壤质地、植被类型及其环境因素差异引起[40],特别是在土壤持水能力、水分获取深度和气孔调节机制方面[60].草地的土壤水分阈值(15.98%)显著高于其他生态系统,主要原因可能在于其高植被覆盖率、高土壤有机碳含量和高叶面积指数[61-62],这些特征通过增强土壤的持水能力和蒸腾需求,提高水分利用阈值;而土壤水分阈值较低的稀树草原(4.73%)和有林草地(4.89%)则可能依赖深根系获取深层水分[63-65],并辅以快速气孔调节机制,形成低阈值-高韧性的水分利用策略[66].Fu等[41]通过计算草地和稀树草原的冠层导电率(Gc)随土壤水分降低的变化,发现稀树草原的气孔关闭反应更为迅速,以防止水分过度损失.然而,这种快速关闭虽然减少水分流失,但超过阈值后,生态系统功能可能迅速崩溃[67-68].值得注意的是,沙质土壤的低持水能力可能进一步降低稀疏灌木林的土壤水分阈值(8.56%)[61].以上分析表明不同植被类型展现出各自独特的水分利用策略,反映出它们对干旱环境存在不同的适应性,这些发现为干旱半干旱区的水资源管理和生态系统恢复提供重要依据. ...
Feedbacks of soil properties on vegetation during the Green Sahara period
2
2020
... 干旱半干旱区4种典型植被类型土壤水分阈值均值4.89%~15.98%(图5),这一结果与Fu等[40]基于涡度相关观测数据得到的12%,Buitink等[58]和Feldman等[29]基于卫星观测数据得到的16%、14%相近,但显著低于中国西北干旱区高阈值站点[59](景阳岭站、垭口站和大野口站土壤水分阈值分别为45.1%、40%和43.2%).这一差异可能由地区间土壤质地、植被类型及其环境因素差异引起[40],特别是在土壤持水能力、水分获取深度和气孔调节机制方面[60].草地的土壤水分阈值(15.98%)显著高于其他生态系统,主要原因可能在于其高植被覆盖率、高土壤有机碳含量和高叶面积指数[61-62],这些特征通过增强土壤的持水能力和蒸腾需求,提高水分利用阈值;而土壤水分阈值较低的稀树草原(4.73%)和有林草地(4.89%)则可能依赖深根系获取深层水分[63-65],并辅以快速气孔调节机制,形成低阈值-高韧性的水分利用策略[66].Fu等[41]通过计算草地和稀树草原的冠层导电率(Gc)随土壤水分降低的变化,发现稀树草原的气孔关闭反应更为迅速,以防止水分过度损失.然而,这种快速关闭虽然减少水分流失,但超过阈值后,生态系统功能可能迅速崩溃[67-68].值得注意的是,沙质土壤的低持水能力可能进一步降低稀疏灌木林的土壤水分阈值(8.56%)[61].以上分析表明不同植被类型展现出各自独特的水分利用策略,反映出它们对干旱环境存在不同的适应性,这些发现为干旱半干旱区的水资源管理和生态系统恢复提供重要依据. ...
... [61].以上分析表明不同植被类型展现出各自独特的水分利用策略,反映出它们对干旱环境存在不同的适应性,这些发现为干旱半干旱区的水资源管理和生态系统恢复提供重要依据. ...
水分对科尔沁沙地差不嘎蒿气体交换特性的影响
1
2012
... 干旱半干旱区4种典型植被类型土壤水分阈值均值4.89%~15.98%(图5),这一结果与Fu等[40]基于涡度相关观测数据得到的12%,Buitink等[58]和Feldman等[29]基于卫星观测数据得到的16%、14%相近,但显著低于中国西北干旱区高阈值站点[59](景阳岭站、垭口站和大野口站土壤水分阈值分别为45.1%、40%和43.2%).这一差异可能由地区间土壤质地、植被类型及其环境因素差异引起[40],特别是在土壤持水能力、水分获取深度和气孔调节机制方面[60].草地的土壤水分阈值(15.98%)显著高于其他生态系统,主要原因可能在于其高植被覆盖率、高土壤有机碳含量和高叶面积指数[61-62],这些特征通过增强土壤的持水能力和蒸腾需求,提高水分利用阈值;而土壤水分阈值较低的稀树草原(4.73%)和有林草地(4.89%)则可能依赖深根系获取深层水分[63-65],并辅以快速气孔调节机制,形成低阈值-高韧性的水分利用策略[66].Fu等[41]通过计算草地和稀树草原的冠层导电率(Gc)随土壤水分降低的变化,发现稀树草原的气孔关闭反应更为迅速,以防止水分过度损失.然而,这种快速关闭虽然减少水分流失,但超过阈值后,生态系统功能可能迅速崩溃[67-68].值得注意的是,沙质土壤的低持水能力可能进一步降低稀疏灌木林的土壤水分阈值(8.56%)[61].以上分析表明不同植被类型展现出各自独特的水分利用策略,反映出它们对干旱环境存在不同的适应性,这些发现为干旱半干旱区的水资源管理和生态系统恢复提供重要依据. ...
非洲和澳大利亚稀树草原树种幼苗的温室比较研究
1
2010
... 干旱半干旱区4种典型植被类型土壤水分阈值均值4.89%~15.98%(图5),这一结果与Fu等[40]基于涡度相关观测数据得到的12%,Buitink等[58]和Feldman等[29]基于卫星观测数据得到的16%、14%相近,但显著低于中国西北干旱区高阈值站点[59](景阳岭站、垭口站和大野口站土壤水分阈值分别为45.1%、40%和43.2%).这一差异可能由地区间土壤质地、植被类型及其环境因素差异引起[40],特别是在土壤持水能力、水分获取深度和气孔调节机制方面[60].草地的土壤水分阈值(15.98%)显著高于其他生态系统,主要原因可能在于其高植被覆盖率、高土壤有机碳含量和高叶面积指数[61-62],这些特征通过增强土壤的持水能力和蒸腾需求,提高水分利用阈值;而土壤水分阈值较低的稀树草原(4.73%)和有林草地(4.89%)则可能依赖深根系获取深层水分[63-65],并辅以快速气孔调节机制,形成低阈值-高韧性的水分利用策略[66].Fu等[41]通过计算草地和稀树草原的冠层导电率(Gc)随土壤水分降低的变化,发现稀树草原的气孔关闭反应更为迅速,以防止水分过度损失.然而,这种快速关闭虽然减少水分流失,但超过阈值后,生态系统功能可能迅速崩溃[67-68].值得注意的是,沙质土壤的低持水能力可能进一步降低稀疏灌木林的土壤水分阈值(8.56%)[61].以上分析表明不同植被类型展现出各自独特的水分利用策略,反映出它们对干旱环境存在不同的适应性,这些发现为干旱半干旱区的水资源管理和生态系统恢复提供重要依据. ...
A closed-form equation for predicting the hydraulic conductivity of unsaturated soils
0
1980
A new model for predicting the hydraulic conductivity of unsaturated porous media
1
1976
... 干旱半干旱区4种典型植被类型土壤水分阈值均值4.89%~15.98%(图5),这一结果与Fu等[40]基于涡度相关观测数据得到的12%,Buitink等[58]和Feldman等[29]基于卫星观测数据得到的16%、14%相近,但显著低于中国西北干旱区高阈值站点[59](景阳岭站、垭口站和大野口站土壤水分阈值分别为45.1%、40%和43.2%).这一差异可能由地区间土壤质地、植被类型及其环境因素差异引起[40],特别是在土壤持水能力、水分获取深度和气孔调节机制方面[60].草地的土壤水分阈值(15.98%)显著高于其他生态系统,主要原因可能在于其高植被覆盖率、高土壤有机碳含量和高叶面积指数[61-62],这些特征通过增强土壤的持水能力和蒸腾需求,提高水分利用阈值;而土壤水分阈值较低的稀树草原(4.73%)和有林草地(4.89%)则可能依赖深根系获取深层水分[63-65],并辅以快速气孔调节机制,形成低阈值-高韧性的水分利用策略[66].Fu等[41]通过计算草地和稀树草原的冠层导电率(Gc)随土壤水分降低的变化,发现稀树草原的气孔关闭反应更为迅速,以防止水分过度损失.然而,这种快速关闭虽然减少水分流失,但超过阈值后,生态系统功能可能迅速崩溃[67-68].值得注意的是,沙质土壤的低持水能力可能进一步降低稀疏灌木林的土壤水分阈值(8.56%)[61].以上分析表明不同植被类型展现出各自独特的水分利用策略,反映出它们对干旱环境存在不同的适应性,这些发现为干旱半干旱区的水资源管理和生态系统恢复提供重要依据. ...
Uncovering the critical soil moisture thresholds of plant water stress for European ecosystems
1
2021
... 干旱半干旱区4种典型植被类型土壤水分阈值均值4.89%~15.98%(图5),这一结果与Fu等[40]基于涡度相关观测数据得到的12%,Buitink等[58]和Feldman等[29]基于卫星观测数据得到的16%、14%相近,但显著低于中国西北干旱区高阈值站点[59](景阳岭站、垭口站和大野口站土壤水分阈值分别为45.1%、40%和43.2%).这一差异可能由地区间土壤质地、植被类型及其环境因素差异引起[40],特别是在土壤持水能力、水分获取深度和气孔调节机制方面[60].草地的土壤水分阈值(15.98%)显著高于其他生态系统,主要原因可能在于其高植被覆盖率、高土壤有机碳含量和高叶面积指数[61-62],这些特征通过增强土壤的持水能力和蒸腾需求,提高水分利用阈值;而土壤水分阈值较低的稀树草原(4.73%)和有林草地(4.89%)则可能依赖深根系获取深层水分[63-65],并辅以快速气孔调节机制,形成低阈值-高韧性的水分利用策略[66].Fu等[41]通过计算草地和稀树草原的冠层导电率(Gc)随土壤水分降低的变化,发现稀树草原的气孔关闭反应更为迅速,以防止水分过度损失.然而,这种快速关闭虽然减少水分流失,但超过阈值后,生态系统功能可能迅速崩溃[67-68].值得注意的是,沙质土壤的低持水能力可能进一步降低稀疏灌木林的土壤水分阈值(8.56%)[61].以上分析表明不同植被类型展现出各自独特的水分利用策略,反映出它们对干旱环境存在不同的适应性,这些发现为干旱半干旱区的水资源管理和生态系统恢复提供重要依据. ...
Global variations in ecosystem-scale isohydricity
1
2017
... 干旱半干旱区4种典型植被类型土壤水分阈值均值4.89%~15.98%(图5),这一结果与Fu等[40]基于涡度相关观测数据得到的12%,Buitink等[58]和Feldman等[29]基于卫星观测数据得到的16%、14%相近,但显著低于中国西北干旱区高阈值站点[59](景阳岭站、垭口站和大野口站土壤水分阈值分别为45.1%、40%和43.2%).这一差异可能由地区间土壤质地、植被类型及其环境因素差异引起[40],特别是在土壤持水能力、水分获取深度和气孔调节机制方面[60].草地的土壤水分阈值(15.98%)显著高于其他生态系统,主要原因可能在于其高植被覆盖率、高土壤有机碳含量和高叶面积指数[61-62],这些特征通过增强土壤的持水能力和蒸腾需求,提高水分利用阈值;而土壤水分阈值较低的稀树草原(4.73%)和有林草地(4.89%)则可能依赖深根系获取深层水分[63-65],并辅以快速气孔调节机制,形成低阈值-高韧性的水分利用策略[66].Fu等[41]通过计算草地和稀树草原的冠层导电率(Gc)随土壤水分降低的变化,发现稀树草原的气孔关闭反应更为迅速,以防止水分过度损失.然而,这种快速关闭虽然减少水分流失,但超过阈值后,生态系统功能可能迅速崩溃[67-68].值得注意的是,沙质土壤的低持水能力可能进一步降低稀疏灌木林的土壤水分阈值(8.56%)[61].以上分析表明不同植被类型展现出各自独特的水分利用策略,反映出它们对干旱环境存在不同的适应性,这些发现为干旱半干旱区的水资源管理和生态系统恢复提供重要依据. ...
Mechanisms of plant survival and mortality during drought:Why do some plants survive while others succumb to drought?
1
2008
... 干旱半干旱区4种典型植被类型土壤水分阈值均值4.89%~15.98%(图5),这一结果与Fu等[40]基于涡度相关观测数据得到的12%,Buitink等[58]和Feldman等[29]基于卫星观测数据得到的16%、14%相近,但显著低于中国西北干旱区高阈值站点[59](景阳岭站、垭口站和大野口站土壤水分阈值分别为45.1%、40%和43.2%).这一差异可能由地区间土壤质地、植被类型及其环境因素差异引起[40],特别是在土壤持水能力、水分获取深度和气孔调节机制方面[60].草地的土壤水分阈值(15.98%)显著高于其他生态系统,主要原因可能在于其高植被覆盖率、高土壤有机碳含量和高叶面积指数[61-62],这些特征通过增强土壤的持水能力和蒸腾需求,提高水分利用阈值;而土壤水分阈值较低的稀树草原(4.73%)和有林草地(4.89%)则可能依赖深根系获取深层水分[63-65],并辅以快速气孔调节机制,形成低阈值-高韧性的水分利用策略[66].Fu等[41]通过计算草地和稀树草原的冠层导电率(Gc)随土壤水分降低的变化,发现稀树草原的气孔关闭反应更为迅速,以防止水分过度损失.然而,这种快速关闭虽然减少水分流失,但超过阈值后,生态系统功能可能迅速崩溃[67-68].值得注意的是,沙质土壤的低持水能力可能进一步降低稀疏灌木林的土壤水分阈值(8.56%)[61].以上分析表明不同植被类型展现出各自独特的水分利用策略,反映出它们对干旱环境存在不同的适应性,这些发现为干旱半干旱区的水资源管理和生态系统恢复提供重要依据. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}