1
2002
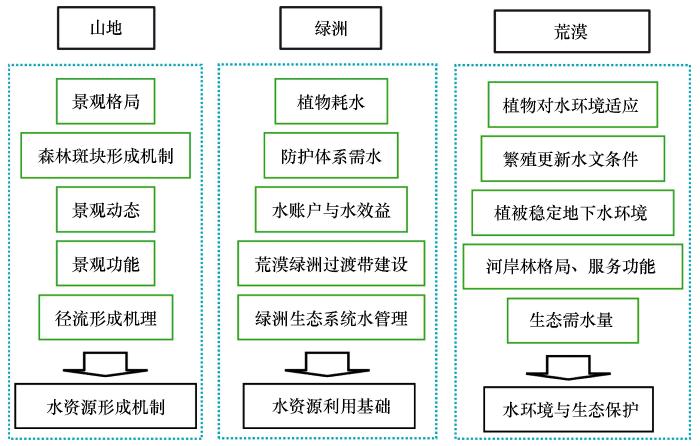
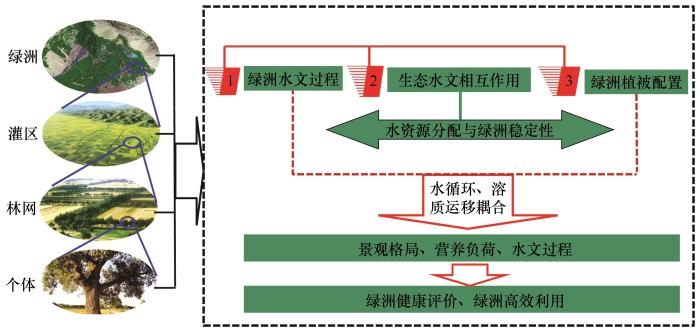
... 基于学科交叉是解决复杂环境问题的重要途径,梳理了干旱区内陆河流域管理中的瓶颈问题,尤其是水资源综合利用中的问题,认识到生态学与水文学的学科交叉有望成为推动干旱区生态建设与区域发展的抓手.翻译出版了《生态水文学——陆生环境与水生环境植物水分关系》[1],综述了干旱区生态水文研究若干问题[2],介绍了生态水文学的学科体系建设及理论与方法等[3-5],得到了国内生态学与水文学研究者的广泛关注,推动了中国生态水文学特别是寒旱区生态水文学的深入发展.针对内陆河流域是由山地、荒漠、绿洲等景观构成的复合系统,系统内要素、过程互相联系、互相作用,水是联系该系统的枢纽等问题,结合临泽站实际,逐步梳理了相关研究的科学问题与目标:①2006年开展水文水资源、水资源高效利用、水环境与生态保护相关研究(图1).②2011年聚焦绿洲生态水文研究,解析从个体、田块、灌区到绿洲尺度的蒸散发、耗水、水平衡及绿洲碳固定、生产力形成等生态-水文过程,研究绿洲系统生态水文相互作用规律及溶质运移与水循环的耦合关系,为绿洲的可持续管理提供依据(图2).③2016年梳理的生态水文研究框架,聚焦荒漠生态系统的生态-水文相互作用机制,旨在为荒漠生态系统多样性保护和适应性管理提供支撑(表1).④2020年梳理的临泽站干旱区生态水文研究框架,针对国家战略需求,开展解决生态环境问题的生态水文机制应用基础研究(表2). ...
干旱区生态水文过程研究若干问题评述
1
2001
... 基于学科交叉是解决复杂环境问题的重要途径,梳理了干旱区内陆河流域管理中的瓶颈问题,尤其是水资源综合利用中的问题,认识到生态学与水文学的学科交叉有望成为推动干旱区生态建设与区域发展的抓手.翻译出版了《生态水文学——陆生环境与水生环境植物水分关系》[1],综述了干旱区生态水文研究若干问题[2],介绍了生态水文学的学科体系建设及理论与方法等[3-5],得到了国内生态学与水文学研究者的广泛关注,推动了中国生态水文学特别是寒旱区生态水文学的深入发展.针对内陆河流域是由山地、荒漠、绿洲等景观构成的复合系统,系统内要素、过程互相联系、互相作用,水是联系该系统的枢纽等问题,结合临泽站实际,逐步梳理了相关研究的科学问题与目标:①2006年开展水文水资源、水资源高效利用、水环境与生态保护相关研究(图1).②2011年聚焦绿洲生态水文研究,解析从个体、田块、灌区到绿洲尺度的蒸散发、耗水、水平衡及绿洲碳固定、生产力形成等生态-水文过程,研究绿洲系统生态水文相互作用规律及溶质运移与水循环的耦合关系,为绿洲的可持续管理提供依据(图2).③2016年梳理的生态水文研究框架,聚焦荒漠生态系统的生态-水文相互作用机制,旨在为荒漠生态系统多样性保护和适应性管理提供支撑(表1).④2020年梳理的临泽站干旱区生态水文研究框架,针对国家战略需求,开展解决生态环境问题的生态水文机制应用基础研究(表2). ...
生态水文研究前沿问题及生态水文观测试验
1
2008
... 基于学科交叉是解决复杂环境问题的重要途径,梳理了干旱区内陆河流域管理中的瓶颈问题,尤其是水资源综合利用中的问题,认识到生态学与水文学的学科交叉有望成为推动干旱区生态建设与区域发展的抓手.翻译出版了《生态水文学——陆生环境与水生环境植物水分关系》[1],综述了干旱区生态水文研究若干问题[2],介绍了生态水文学的学科体系建设及理论与方法等[3-5],得到了国内生态学与水文学研究者的广泛关注,推动了中国生态水文学特别是寒旱区生态水文学的深入发展.针对内陆河流域是由山地、荒漠、绿洲等景观构成的复合系统,系统内要素、过程互相联系、互相作用,水是联系该系统的枢纽等问题,结合临泽站实际,逐步梳理了相关研究的科学问题与目标:①2006年开展水文水资源、水资源高效利用、水环境与生态保护相关研究(图1).②2011年聚焦绿洲生态水文研究,解析从个体、田块、灌区到绿洲尺度的蒸散发、耗水、水平衡及绿洲碳固定、生产力形成等生态-水文过程,研究绿洲系统生态水文相互作用规律及溶质运移与水循环的耦合关系,为绿洲的可持续管理提供依据(图2).③2016年梳理的生态水文研究框架,聚焦荒漠生态系统的生态-水文相互作用机制,旨在为荒漠生态系统多样性保护和适应性管理提供支撑(表1).④2020年梳理的临泽站干旱区生态水文研究框架,针对国家战略需求,开展解决生态环境问题的生态水文机制应用基础研究(表2). ...
生态水文学:揭示生态过程和生态格局水文学机制的科学
0
2001
绿水及其研究进展
1
2006
... 基于学科交叉是解决复杂环境问题的重要途径,梳理了干旱区内陆河流域管理中的瓶颈问题,尤其是水资源综合利用中的问题,认识到生态学与水文学的学科交叉有望成为推动干旱区生态建设与区域发展的抓手.翻译出版了《生态水文学——陆生环境与水生环境植物水分关系》[1],综述了干旱区生态水文研究若干问题[2],介绍了生态水文学的学科体系建设及理论与方法等[3-5],得到了国内生态学与水文学研究者的广泛关注,推动了中国生态水文学特别是寒旱区生态水文学的深入发展.针对内陆河流域是由山地、荒漠、绿洲等景观构成的复合系统,系统内要素、过程互相联系、互相作用,水是联系该系统的枢纽等问题,结合临泽站实际,逐步梳理了相关研究的科学问题与目标:①2006年开展水文水资源、水资源高效利用、水环境与生态保护相关研究(图1).②2011年聚焦绿洲生态水文研究,解析从个体、田块、灌区到绿洲尺度的蒸散发、耗水、水平衡及绿洲碳固定、生产力形成等生态-水文过程,研究绿洲系统生态水文相互作用规律及溶质运移与水循环的耦合关系,为绿洲的可持续管理提供依据(图2).③2016年梳理的生态水文研究框架,聚焦荒漠生态系统的生态-水文相互作用机制,旨在为荒漠生态系统多样性保护和适应性管理提供支撑(表1).④2020年梳理的临泽站干旱区生态水文研究框架,针对国家战略需求,开展解决生态环境问题的生态水文机制应用基础研究(表2). ...
荒漠绿洲农田防护树种二白杨生长季节树干液流的变化
1
2004
... 植物耗水量是指植物在正常生长和生存中所消耗的水分,包括蒸散发(植物蒸腾与土壤蒸发).蒸散发在不同尺度上(个体、林分等)存在尺度效应.热脉冲技术可以测定植物个体的蒸腾量,并通过林木边材面积变化参数推算林分蒸腾量.此方法适用于条带状防护林的蒸腾量估算,但未考虑土壤蒸发.常学向等[6]利用热脉冲技术研究了河西走廊绿洲中杨树、沙枣、梭梭、枣树等植株的蒸腾,并外推了林分的蒸腾量.通过典型时段的观测,建立了林木耗水量与气象因子的关系,预测了生长季的单株蒸腾量[7].结果表明:在土壤含水量为4.8%~10.7%的情况下,21年生二白杨的单株蒸腾量为77.2 kg·d-1;在土壤含水量为2.3%~6.4%的情况下,23年生沙枣的单株蒸腾量为1.5 kg·d-1;在土壤含水量为0.9%~2.0%的情况下,23年生梭梭的单株蒸腾量为3.5 kg·d-1.利用边材面积将单株林木的蒸腾量扩展到林分尺度,21年生二白杨林分蒸腾量为1.7~5.6 mm·d-1,日均蒸腾量达4.1 mm,生长季蒸腾量达到624 mm[8-9].23年生沙枣林分的蒸腾量为0.2~0.4 mm·d-1,日均蒸腾量仅为0.3 mm,生长季蒸腾量为45 mm[10].23年生梭梭林分的蒸腾量为0.2~0.5 mm·d-1,日均蒸腾量达0.3 mm,生长季蒸腾量达到47 mm[11]. ...
Radial pattern of sap flow and response to microclimate and soil moisture in Qinghai spruce (Picea crassifolia) in the upper Heihe River Basin of arid northwestern China
1
2014
... 植物耗水量是指植物在正常生长和生存中所消耗的水分,包括蒸散发(植物蒸腾与土壤蒸发).蒸散发在不同尺度上(个体、林分等)存在尺度效应.热脉冲技术可以测定植物个体的蒸腾量,并通过林木边材面积变化参数推算林分蒸腾量.此方法适用于条带状防护林的蒸腾量估算,但未考虑土壤蒸发.常学向等[6]利用热脉冲技术研究了河西走廊绿洲中杨树、沙枣、梭梭、枣树等植株的蒸腾,并外推了林分的蒸腾量.通过典型时段的观测,建立了林木耗水量与气象因子的关系,预测了生长季的单株蒸腾量[7].结果表明:在土壤含水量为4.8%~10.7%的情况下,21年生二白杨的单株蒸腾量为77.2 kg·d-1;在土壤含水量为2.3%~6.4%的情况下,23年生沙枣的单株蒸腾量为1.5 kg·d-1;在土壤含水量为0.9%~2.0%的情况下,23年生梭梭的单株蒸腾量为3.5 kg·d-1.利用边材面积将单株林木的蒸腾量扩展到林分尺度,21年生二白杨林分蒸腾量为1.7~5.6 mm·d-1,日均蒸腾量达4.1 mm,生长季蒸腾量达到624 mm[8-9].23年生沙枣林分的蒸腾量为0.2~0.4 mm·d-1,日均蒸腾量仅为0.3 mm,生长季蒸腾量为45 mm[10].23年生梭梭林分的蒸腾量为0.2~0.5 mm·d-1,日均蒸腾量达0.3 mm,生长季蒸腾量达到47 mm[11]. ...
Sap flow and tree conductance of shelter-belt in arid region of China
1
2006
... 植物耗水量是指植物在正常生长和生存中所消耗的水分,包括蒸散发(植物蒸腾与土壤蒸发).蒸散发在不同尺度上(个体、林分等)存在尺度效应.热脉冲技术可以测定植物个体的蒸腾量,并通过林木边材面积变化参数推算林分蒸腾量.此方法适用于条带状防护林的蒸腾量估算,但未考虑土壤蒸发.常学向等[6]利用热脉冲技术研究了河西走廊绿洲中杨树、沙枣、梭梭、枣树等植株的蒸腾,并外推了林分的蒸腾量.通过典型时段的观测,建立了林木耗水量与气象因子的关系,预测了生长季的单株蒸腾量[7].结果表明:在土壤含水量为4.8%~10.7%的情况下,21年生二白杨的单株蒸腾量为77.2 kg·d-1;在土壤含水量为2.3%~6.4%的情况下,23年生沙枣的单株蒸腾量为1.5 kg·d-1;在土壤含水量为0.9%~2.0%的情况下,23年生梭梭的单株蒸腾量为3.5 kg·d-1.利用边材面积将单株林木的蒸腾量扩展到林分尺度,21年生二白杨林分蒸腾量为1.7~5.6 mm·d-1,日均蒸腾量达4.1 mm,生长季蒸腾量达到624 mm[8-9].23年生沙枣林分的蒸腾量为0.2~0.4 mm·d-1,日均蒸腾量仅为0.3 mm,生长季蒸腾量为45 mm[10].23年生梭梭林分的蒸腾量为0.2~0.5 mm·d-1,日均蒸腾量达0.3 mm,生长季蒸腾量达到47 mm[11]. ...
Transpiration of a Linze jujube orchard in an arid region of China
1
2009
... 植物耗水量是指植物在正常生长和生存中所消耗的水分,包括蒸散发(植物蒸腾与土壤蒸发).蒸散发在不同尺度上(个体、林分等)存在尺度效应.热脉冲技术可以测定植物个体的蒸腾量,并通过林木边材面积变化参数推算林分蒸腾量.此方法适用于条带状防护林的蒸腾量估算,但未考虑土壤蒸发.常学向等[6]利用热脉冲技术研究了河西走廊绿洲中杨树、沙枣、梭梭、枣树等植株的蒸腾,并外推了林分的蒸腾量.通过典型时段的观测,建立了林木耗水量与气象因子的关系,预测了生长季的单株蒸腾量[7].结果表明:在土壤含水量为4.8%~10.7%的情况下,21年生二白杨的单株蒸腾量为77.2 kg·d-1;在土壤含水量为2.3%~6.4%的情况下,23年生沙枣的单株蒸腾量为1.5 kg·d-1;在土壤含水量为0.9%~2.0%的情况下,23年生梭梭的单株蒸腾量为3.5 kg·d-1.利用边材面积将单株林木的蒸腾量扩展到林分尺度,21年生二白杨林分蒸腾量为1.7~5.6 mm·d-1,日均蒸腾量达4.1 mm,生长季蒸腾量达到624 mm[8-9].23年生沙枣林分的蒸腾量为0.2~0.4 mm·d-1,日均蒸腾量仅为0.3 mm,生长季蒸腾量为45 mm[10].23年生梭梭林分的蒸腾量为0.2~0.5 mm·d-1,日均蒸腾量达0.3 mm,生长季蒸腾量达到47 mm[11]. ...
黑河中游沙枣树干液流的动态变化及其与林木个体生长的关系
1
2004
... 植物耗水量是指植物在正常生长和生存中所消耗的水分,包括蒸散发(植物蒸腾与土壤蒸发).蒸散发在不同尺度上(个体、林分等)存在尺度效应.热脉冲技术可以测定植物个体的蒸腾量,并通过林木边材面积变化参数推算林分蒸腾量.此方法适用于条带状防护林的蒸腾量估算,但未考虑土壤蒸发.常学向等[6]利用热脉冲技术研究了河西走廊绿洲中杨树、沙枣、梭梭、枣树等植株的蒸腾,并外推了林分的蒸腾量.通过典型时段的观测,建立了林木耗水量与气象因子的关系,预测了生长季的单株蒸腾量[7].结果表明:在土壤含水量为4.8%~10.7%的情况下,21年生二白杨的单株蒸腾量为77.2 kg·d-1;在土壤含水量为2.3%~6.4%的情况下,23年生沙枣的单株蒸腾量为1.5 kg·d-1;在土壤含水量为0.9%~2.0%的情况下,23年生梭梭的单株蒸腾量为3.5 kg·d-1.利用边材面积将单株林木的蒸腾量扩展到林分尺度,21年生二白杨林分蒸腾量为1.7~5.6 mm·d-1,日均蒸腾量达4.1 mm,生长季蒸腾量达到624 mm[8-9].23年生沙枣林分的蒸腾量为0.2~0.4 mm·d-1,日均蒸腾量仅为0.3 mm,生长季蒸腾量为45 mm[10].23年生梭梭林分的蒸腾量为0.2~0.5 mm·d-1,日均蒸腾量达0.3 mm,生长季蒸腾量达到47 mm[11]. ...
荒漠区固沙植物梭梭(Haloxylon ammodendron)耗水特征
1
2007
... 植物耗水量是指植物在正常生长和生存中所消耗的水分,包括蒸散发(植物蒸腾与土壤蒸发).蒸散发在不同尺度上(个体、林分等)存在尺度效应.热脉冲技术可以测定植物个体的蒸腾量,并通过林木边材面积变化参数推算林分蒸腾量.此方法适用于条带状防护林的蒸腾量估算,但未考虑土壤蒸发.常学向等[6]利用热脉冲技术研究了河西走廊绿洲中杨树、沙枣、梭梭、枣树等植株的蒸腾,并外推了林分的蒸腾量.通过典型时段的观测,建立了林木耗水量与气象因子的关系,预测了生长季的单株蒸腾量[7].结果表明:在土壤含水量为4.8%~10.7%的情况下,21年生二白杨的单株蒸腾量为77.2 kg·d-1;在土壤含水量为2.3%~6.4%的情况下,23年生沙枣的单株蒸腾量为1.5 kg·d-1;在土壤含水量为0.9%~2.0%的情况下,23年生梭梭的单株蒸腾量为3.5 kg·d-1.利用边材面积将单株林木的蒸腾量扩展到林分尺度,21年生二白杨林分蒸腾量为1.7~5.6 mm·d-1,日均蒸腾量达4.1 mm,生长季蒸腾量达到624 mm[8-9].23年生沙枣林分的蒸腾量为0.2~0.4 mm·d-1,日均蒸腾量仅为0.3 mm,生长季蒸腾量为45 mm[10].23年生梭梭林分的蒸腾量为0.2~0.5 mm·d-1,日均蒸腾量达0.3 mm,生长季蒸腾量达到47 mm[11]. ...
Seasonal variations in energy exchange and evapotranspiration of an oasis-desert ecotone in an arid region
1
2021
... 利用涡度相关法和大孔径闪烁仪可有效测定干旱区斑块状植被的耗水量.Ji等[12]利用涡度相关法,确定了绿洲外围固沙植被梭梭的耗水量.临泽北部荒漠-绿洲过渡带梭梭年蒸散发总量为203 mm,其中植物蒸腾占85%,土壤蒸发仅占15%.蒸腾量超出有效降水量,导致植被水量平衡为-114 mm,消耗了土壤水库水甚至浅层地下水.Zhao等[13]利用涡度相关、Li-6400和热脉冲同步数据,对比分析了叶片、个体和FAO-56双作物系数法获取的冠层蒸腾量,结果表明,利用上述3种方法估算的蒸腾量因作物发育阶段差异而显著不同.拔节期,双作物系数法与叶片观测的结果基本一致,但均高于个体方法观测值;灌浆期,个体和叶片观测值基本一致,但均低于双作物系数法,总体上个体观测的蒸腾量最低.整个生长季,作物蒸腾与土壤蒸发的比值大约为3∶1,田间供水充足时,作物蒸腾与土壤蒸发比值明显受作物生长过程影响,不同阶段的比值差异主要受叶面积指数影响[14]. ...
Canopy transpiration obtained from leaf transpiration,sap flow and FAO-56 dual crop coefficient method
1
2015
... 利用涡度相关法和大孔径闪烁仪可有效测定干旱区斑块状植被的耗水量.Ji等[12]利用涡度相关法,确定了绿洲外围固沙植被梭梭的耗水量.临泽北部荒漠-绿洲过渡带梭梭年蒸散发总量为203 mm,其中植物蒸腾占85%,土壤蒸发仅占15%.蒸腾量超出有效降水量,导致植被水量平衡为-114 mm,消耗了土壤水库水甚至浅层地下水.Zhao等[13]利用涡度相关、Li-6400和热脉冲同步数据,对比分析了叶片、个体和FAO-56双作物系数法获取的冠层蒸腾量,结果表明,利用上述3种方法估算的蒸腾量因作物发育阶段差异而显著不同.拔节期,双作物系数法与叶片观测的结果基本一致,但均高于个体方法观测值;灌浆期,个体和叶片观测值基本一致,但均低于双作物系数法,总体上个体观测的蒸腾量最低.整个生长季,作物蒸腾与土壤蒸发的比值大约为3∶1,田间供水充足时,作物蒸腾与土壤蒸发比值明显受作物生长过程影响,不同阶段的比值差异主要受叶面积指数影响[14]. ...
Water balance and migration for maize in an oasis farmland of Northwest China
1
2014
... 利用涡度相关法和大孔径闪烁仪可有效测定干旱区斑块状植被的耗水量.Ji等[12]利用涡度相关法,确定了绿洲外围固沙植被梭梭的耗水量.临泽北部荒漠-绿洲过渡带梭梭年蒸散发总量为203 mm,其中植物蒸腾占85%,土壤蒸发仅占15%.蒸腾量超出有效降水量,导致植被水量平衡为-114 mm,消耗了土壤水库水甚至浅层地下水.Zhao等[13]利用涡度相关、Li-6400和热脉冲同步数据,对比分析了叶片、个体和FAO-56双作物系数法获取的冠层蒸腾量,结果表明,利用上述3种方法估算的蒸腾量因作物发育阶段差异而显著不同.拔节期,双作物系数法与叶片观测的结果基本一致,但均高于个体方法观测值;灌浆期,个体和叶片观测值基本一致,但均低于双作物系数法,总体上个体观测的蒸腾量最低.整个生长季,作物蒸腾与土壤蒸发的比值大约为3∶1,田间供水充足时,作物蒸腾与土壤蒸发比值明显受作物生长过程影响,不同阶段的比值差异主要受叶面积指数影响[14]. ...
Water consumption in artificial desert oasis based on net primary productivity
2
2010
... 利用遥感方法可有效估算生产力,并通过与蒸腾系数的乘积来估算绿洲生态系统的耗水量.Zhao等[15-16]利用该方法分别估算了张掖人工绿洲和额济纳天然绿洲的耗水量,并进一步估算了额济纳荒漠河岸林的生产力及其耗水量.结果表明: 黑河中游绿洲生态系统消耗的灌溉补充水量为18.41×108~21.92×108 m3.在不考虑降水量的情况下, 绿洲生态系统内具有生产功能的农田耗水为13.97×108~16.84×108 m3, 占绿洲植被耗水的77.1%~77.8%;对绿洲农业生产起防护作用的生态系统耗水占绿洲植被总耗水的22%左右.2005年额济纳绿洲的耗水量为1.53×108 m3,若是现有的植被恢复到最高生产水平,耗水量为3.49×108 m3. ...
... 由于观测仪器和方法的限制,荒漠绿洲植物耗水量的研究结果存在差异.基于分析现有研究,得出以下关于黑河中游人工绿洲耗水与生态需水量的结论:以杨树为主的人工防护林蒸腾量约为620 mm,实际耗水量775 mm;农田的耗水量为630~675 mm,灌区耗水量为750~960 mm(绿洲边缘灌区较大),绿洲耗水量为750 mm,其中生态用水约占20%~25%.在上述水资源配置下,绿洲维持在亚稳定-稳定状态[15,17].考虑到城镇居民生活用水、河道输水损失、绿洲植被耗水、农田用水和降水补充等,额济纳绿洲维持现状所需的黑河下泄水量为1.93×108~2.23×108 m3;若是现有的植被恢复到最高生产的水平,所需的下泄水量为4.28×108~5.17×108 m3[17]. ...
Study on vegetation ecological water requirement in Ejina Oasis
1
2007
... 利用遥感方法可有效估算生产力,并通过与蒸腾系数的乘积来估算绿洲生态系统的耗水量.Zhao等[15-16]利用该方法分别估算了张掖人工绿洲和额济纳天然绿洲的耗水量,并进一步估算了额济纳荒漠河岸林的生产力及其耗水量.结果表明: 黑河中游绿洲生态系统消耗的灌溉补充水量为18.41×108~21.92×108 m3.在不考虑降水量的情况下, 绿洲生态系统内具有生产功能的农田耗水为13.97×108~16.84×108 m3, 占绿洲植被耗水的77.1%~77.8%;对绿洲农业生产起防护作用的生态系统耗水占绿洲植被总耗水的22%左右.2005年额济纳绿洲的耗水量为1.53×108 m3,若是现有的植被恢复到最高生产水平,耗水量为3.49×108 m3. ...
Water requirements and stability of oasis ecosystem in arid region,China
2
2010
... 由于观测仪器和方法的限制,荒漠绿洲植物耗水量的研究结果存在差异.基于分析现有研究,得出以下关于黑河中游人工绿洲耗水与生态需水量的结论:以杨树为主的人工防护林蒸腾量约为620 mm,实际耗水量775 mm;农田的耗水量为630~675 mm,灌区耗水量为750~960 mm(绿洲边缘灌区较大),绿洲耗水量为750 mm,其中生态用水约占20%~25%.在上述水资源配置下,绿洲维持在亚稳定-稳定状态[15,17].考虑到城镇居民生活用水、河道输水损失、绿洲植被耗水、农田用水和降水补充等,额济纳绿洲维持现状所需的黑河下泄水量为1.93×108~2.23×108 m3;若是现有的植被恢复到最高生产的水平,所需的下泄水量为4.28×108~5.17×108 m3[17]. ...
... 3[17]. ...
青海云杉(Picea crassifolia)种群格局的分形特征及其影响因素
1
2008
... 研究揭示了祁连山北坡山地森林植被格局与水文过程.张立杰等[18]量化了斑块森林的种群空间分布,揭示了采伐干扰导致斑块森林林分密度的动态变化过程,并分析了残留林木对次生林空间异质性的影响.结果表明,祁连山青海云杉种群以幼龄个体为主,处于稳定发展阶段.青海云杉种群格局的分形维数主要受海拔、坡向、坡度和土壤水分含量等生境因子的影响;祁连山青海云杉林窗干扰的线状密度为22.5个·km-1,以小规模的人为干扰为主.林窗内青海云杉幼苗的天然更新情况明显好于林下;在林窗形成初期,林窗内的个体都至少经历1次生长释放,持续时间为6~10年[19-20].林窗中心和林窗边缘的土壤含水量均显著高于正常林分内,而土壤含水量无显著差异.林冠截留率在林内最大,其次为林窗边缘和林窗中心.研究了森林斑块随海拔、坡向及坡度的分异规律;阐明了典型流域热量、土壤水分随海拔、坡度和坡向的变化,发现地形导致的辐射、蒸散潜力、土壤水分的空间分异是森林斑块形成的主要原因[21-22]. ...
祁连山青海云杉林采伐干扰与恢复过程
1
2009
... 研究揭示了祁连山北坡山地森林植被格局与水文过程.张立杰等[18]量化了斑块森林的种群空间分布,揭示了采伐干扰导致斑块森林林分密度的动态变化过程,并分析了残留林木对次生林空间异质性的影响.结果表明,祁连山青海云杉种群以幼龄个体为主,处于稳定发展阶段.青海云杉种群格局的分形维数主要受海拔、坡向、坡度和土壤水分含量等生境因子的影响;祁连山青海云杉林窗干扰的线状密度为22.5个·km-1,以小规模的人为干扰为主.林窗内青海云杉幼苗的天然更新情况明显好于林下;在林窗形成初期,林窗内的个体都至少经历1次生长释放,持续时间为6~10年[19-20].林窗中心和林窗边缘的土壤含水量均显著高于正常林分内,而土壤含水量无显著差异.林冠截留率在林内最大,其次为林窗边缘和林窗中心.研究了森林斑块随海拔、坡向及坡度的分异规律;阐明了典型流域热量、土壤水分随海拔、坡度和坡向的变化,发现地形导致的辐射、蒸散潜力、土壤水分的空间分异是森林斑块形成的主要原因[21-22]. ...
Successional process of Picea crassifolia forest after logging disturbance in semiarid mountains:a case study in the Qilian Mountains,northwestern China
1
2010
... 研究揭示了祁连山北坡山地森林植被格局与水文过程.张立杰等[18]量化了斑块森林的种群空间分布,揭示了采伐干扰导致斑块森林林分密度的动态变化过程,并分析了残留林木对次生林空间异质性的影响.结果表明,祁连山青海云杉种群以幼龄个体为主,处于稳定发展阶段.青海云杉种群格局的分形维数主要受海拔、坡向、坡度和土壤水分含量等生境因子的影响;祁连山青海云杉林窗干扰的线状密度为22.5个·km-1,以小规模的人为干扰为主.林窗内青海云杉幼苗的天然更新情况明显好于林下;在林窗形成初期,林窗内的个体都至少经历1次生长释放,持续时间为6~10年[19-20].林窗中心和林窗边缘的土壤含水量均显著高于正常林分内,而土壤含水量无显著差异.林冠截留率在林内最大,其次为林窗边缘和林窗中心.研究了森林斑块随海拔、坡向及坡度的分异规律;阐明了典型流域热量、土壤水分随海拔、坡度和坡向的变化,发现地形导致的辐射、蒸散潜力、土壤水分的空间分异是森林斑块形成的主要原因[21-22]. ...
Self-organized vegetation patterning effects on surface soil hydraulic conductivity:a case study in the Qilian Mountains,China
1
2013
... 研究揭示了祁连山北坡山地森林植被格局与水文过程.张立杰等[18]量化了斑块森林的种群空间分布,揭示了采伐干扰导致斑块森林林分密度的动态变化过程,并分析了残留林木对次生林空间异质性的影响.结果表明,祁连山青海云杉种群以幼龄个体为主,处于稳定发展阶段.青海云杉种群格局的分形维数主要受海拔、坡向、坡度和土壤水分含量等生境因子的影响;祁连山青海云杉林窗干扰的线状密度为22.5个·km-1,以小规模的人为干扰为主.林窗内青海云杉幼苗的天然更新情况明显好于林下;在林窗形成初期,林窗内的个体都至少经历1次生长释放,持续时间为6~10年[19-20].林窗中心和林窗边缘的土壤含水量均显著高于正常林分内,而土壤含水量无显著差异.林冠截留率在林内最大,其次为林窗边缘和林窗中心.研究了森林斑块随海拔、坡向及坡度的分异规律;阐明了典型流域热量、土壤水分随海拔、坡度和坡向的变化,发现地形导致的辐射、蒸散潜力、土壤水分的空间分异是森林斑块形成的主要原因[21-22]. ...
Fifty years of change in a coniferous forest in the Qilian Mountains,China:advantages of high-definition remote sensing
1
2020
... 研究揭示了祁连山北坡山地森林植被格局与水文过程.张立杰等[18]量化了斑块森林的种群空间分布,揭示了采伐干扰导致斑块森林林分密度的动态变化过程,并分析了残留林木对次生林空间异质性的影响.结果表明,祁连山青海云杉种群以幼龄个体为主,处于稳定发展阶段.青海云杉种群格局的分形维数主要受海拔、坡向、坡度和土壤水分含量等生境因子的影响;祁连山青海云杉林窗干扰的线状密度为22.5个·km-1,以小规模的人为干扰为主.林窗内青海云杉幼苗的天然更新情况明显好于林下;在林窗形成初期,林窗内的个体都至少经历1次生长释放,持续时间为6~10年[19-20].林窗中心和林窗边缘的土壤含水量均显著高于正常林分内,而土壤含水量无显著差异.林冠截留率在林内最大,其次为林窗边缘和林窗中心.研究了森林斑块随海拔、坡向及坡度的分异规律;阐明了典型流域热量、土壤水分随海拔、坡度和坡向的变化,发现地形导致的辐射、蒸散潜力、土壤水分的空间分异是森林斑块形成的主要原因[21-22]. ...
Response of tree-ring growth to climate at treeline ecotones in the Qilian Mountains,northwestern China
1
2011
... 气候变化引起山地森林生态系统变化.过去100年祁连山林线上升了近20 m,林线区幼苗成活率也呈显著增加趋势,尤其是1980年以来,树木轮宽生长速度提高了20%,表明祁连山林线动态变化对区域气候升温敏感,冬季最低温度和积雪冻土消融起到了决定性作用[23-24].近十几年来,祁连山青海云杉生长季长度平均增幅为3.7 d/10a,林木蒸腾耗水量增加超过5%[25].高山灌丛物候对气候升温更敏感,生长季平均增幅超过4 d/10a;物候变化与平均气温和最低温显著相关,而降水并非主要限制因子,干旱年份土壤含水量偏低导致植物春季物候显著推迟(平均推迟5.9 d/10a)[26-27]. ...
Response of tree recruitment to climatic variability in the alpine treeline ecotone of the Qilian Mountains,northwestern China
1
2013
... 气候变化引起山地森林生态系统变化.过去100年祁连山林线上升了近20 m,林线区幼苗成活率也呈显著增加趋势,尤其是1980年以来,树木轮宽生长速度提高了20%,表明祁连山林线动态变化对区域气候升温敏感,冬季最低温度和积雪冻土消融起到了决定性作用[23-24].近十几年来,祁连山青海云杉生长季长度平均增幅为3.7 d/10a,林木蒸腾耗水量增加超过5%[25].高山灌丛物候对气候升温更敏感,生长季平均增幅超过4 d/10a;物候变化与平均气温和最低温显著相关,而降水并非主要限制因子,干旱年份土壤含水量偏低导致植物春季物候显著推迟(平均推迟5.9 d/10a)[26-27]. ...
Detecting the effects of climate change on canopy phenology in coniferous forests in semi-arid mountain regions of China
1
2014
... 气候变化引起山地森林生态系统变化.过去100年祁连山林线上升了近20 m,林线区幼苗成活率也呈显著增加趋势,尤其是1980年以来,树木轮宽生长速度提高了20%,表明祁连山林线动态变化对区域气候升温敏感,冬季最低温度和积雪冻土消融起到了决定性作用[23-24].近十几年来,祁连山青海云杉生长季长度平均增幅为3.7 d/10a,林木蒸腾耗水量增加超过5%[25].高山灌丛物候对气候升温更敏感,生长季平均增幅超过4 d/10a;物候变化与平均气温和最低温显著相关,而降水并非主要限制因子,干旱年份土壤含水量偏低导致植物春季物候显著推迟(平均推迟5.9 d/10a)[26-27]. ...
Assessing temperature sensitivity of subalpine shrub phenology in semi-arid mountain regions of China
1
2015
... 气候变化引起山地森林生态系统变化.过去100年祁连山林线上升了近20 m,林线区幼苗成活率也呈显著增加趋势,尤其是1980年以来,树木轮宽生长速度提高了20%,表明祁连山林线动态变化对区域气候升温敏感,冬季最低温度和积雪冻土消融起到了决定性作用[23-24].近十几年来,祁连山青海云杉生长季长度平均增幅为3.7 d/10a,林木蒸腾耗水量增加超过5%[25].高山灌丛物候对气候升温更敏感,生长季平均增幅超过4 d/10a;物候变化与平均气温和最低温显著相关,而降水并非主要限制因子,干旱年份土壤含水量偏低导致植物春季物候显著推迟(平均推迟5.9 d/10a)[26-27]. ...
Impacts of recent climate extremes on spring phenology in arid-mountain ecosystems in China
1
2018
... 气候变化引起山地森林生态系统变化.过去100年祁连山林线上升了近20 m,林线区幼苗成活率也呈显著增加趋势,尤其是1980年以来,树木轮宽生长速度提高了20%,表明祁连山林线动态变化对区域气候升温敏感,冬季最低温度和积雪冻土消融起到了决定性作用[23-24].近十几年来,祁连山青海云杉生长季长度平均增幅为3.7 d/10a,林木蒸腾耗水量增加超过5%[25].高山灌丛物候对气候升温更敏感,生长季平均增幅超过4 d/10a;物候变化与平均气温和最低温显著相关,而降水并非主要限制因子,干旱年份土壤含水量偏低导致植物春季物候显著推迟(平均推迟5.9 d/10a)[26-27]. ...
Effect of forest on annual water yield in the mountains of an arid inland river basin: a case study in the Pailugou catchment on northwestern China's Qilian Mountains
1
2012
... 确定了山地森林水文过程及垂直带各景观单元的水量平衡关系.He等[28-29]建立了典型小流域生态水文数据库,定量刻画了森林生态水文特征,发现超过80%的径流主要来自高寒区(冰雪带、寒漠带、高寒草甸和灌丛带),而分布在中山带的青海云杉林斑块通过林冠截留、蒸腾和林地蒸散发消耗了超过97%的降水,森林斑块区年产水量占流域总产水量不足3%.在降水偏少的年份,森林为了维持自身生长需要消耗土壤储水,导致土壤旱化.Liu等[30]建立了祁连山典型草地系统点尺度土壤水分动态随机模型,构建了小流域生态系统生长季稳定状态的土壤湿度概率密度函数,确定了相应的变化范围;通过连续监测林线附近草地、灌丛及林地对应的土壤水分、植被截留及蒸散发过程,比较了不同植被类型间的土壤水分时间异质性,分析了导致这种差异的生态水文过程及其潜在生态水文学意义. ...
The response of soil moisture to rainfall event size in subalpine grassland and meadows in a semi-arid mountain range: a case study in northwestern China's Qilian Mountains
1
2012
... 确定了山地森林水文过程及垂直带各景观单元的水量平衡关系.He等[28-29]建立了典型小流域生态水文数据库,定量刻画了森林生态水文特征,发现超过80%的径流主要来自高寒区(冰雪带、寒漠带、高寒草甸和灌丛带),而分布在中山带的青海云杉林斑块通过林冠截留、蒸腾和林地蒸散发消耗了超过97%的降水,森林斑块区年产水量占流域总产水量不足3%.在降水偏少的年份,森林为了维持自身生长需要消耗土壤储水,导致土壤旱化.Liu等[30]建立了祁连山典型草地系统点尺度土壤水分动态随机模型,构建了小流域生态系统生长季稳定状态的土壤湿度概率密度函数,确定了相应的变化范围;通过连续监测林线附近草地、灌丛及林地对应的土壤水分、植被截留及蒸散发过程,比较了不同植被类型间的土壤水分时间异质性,分析了导致这种差异的生态水文过程及其潜在生态水文学意义. ...
Stochastic modeling of soil moisture dynamics in a grassland of Qilian Mountain at point scale
1
2017
... 确定了山地森林水文过程及垂直带各景观单元的水量平衡关系.He等[28-29]建立了典型小流域生态水文数据库,定量刻画了森林生态水文特征,发现超过80%的径流主要来自高寒区(冰雪带、寒漠带、高寒草甸和灌丛带),而分布在中山带的青海云杉林斑块通过林冠截留、蒸腾和林地蒸散发消耗了超过97%的降水,森林斑块区年产水量占流域总产水量不足3%.在降水偏少的年份,森林为了维持自身生长需要消耗土壤储水,导致土壤旱化.Liu等[30]建立了祁连山典型草地系统点尺度土壤水分动态随机模型,构建了小流域生态系统生长季稳定状态的土壤湿度概率密度函数,确定了相应的变化范围;通过连续监测林线附近草地、灌丛及林地对应的土壤水分、植被截留及蒸散发过程,比较了不同植被类型间的土壤水分时间异质性,分析了导致这种差异的生态水文过程及其潜在生态水文学意义. ...
Patterns and controls of soil organic carbon and nitrogen in alpine forests of northwestern China
1
2015
... 估算了山地森林碳储量及碳循环速率及其对气候变化的响应.结果表明,青海云杉林土壤有机碳密度随着海拔的升高而显著增加,平均值高达23.7 kg·m-2,高于其他温带森林,表现出较强的固碳能力[31-32];青海云杉林土壤呼吸速率随海拔的升高而降低,但其温度敏感性随海拔的升高而增加;增温实验表明,增温显著提高了青海云杉林土壤微生物代谢能力及多样性,随着土壤温度升高(1.3 ℃),土壤呼吸速率显著提高(30%~50%),在一定程度上削弱了干旱区山地森林的固碳能力[33]. ...
Patterns and environmental controls of soil organic carbon and total nitrogen in alpine ecosystems of northwestern China
1
2016
... 估算了山地森林碳储量及碳循环速率及其对气候变化的响应.结果表明,青海云杉林土壤有机碳密度随着海拔的升高而显著增加,平均值高达23.7 kg·m-2,高于其他温带森林,表现出较强的固碳能力[31-32];青海云杉林土壤呼吸速率随海拔的升高而降低,但其温度敏感性随海拔的升高而增加;增温实验表明,增温显著提高了青海云杉林土壤微生物代谢能力及多样性,随着土壤温度升高(1.3 ℃),土壤呼吸速率显著提高(30%~50%),在一定程度上削弱了干旱区山地森林的固碳能力[33]. ...
Empirical evidence for microbial regulation of soil respiration in alpine forests
1
2021
... 估算了山地森林碳储量及碳循环速率及其对气候变化的响应.结果表明,青海云杉林土壤有机碳密度随着海拔的升高而显著增加,平均值高达23.7 kg·m-2,高于其他温带森林,表现出较强的固碳能力[31-32];青海云杉林土壤呼吸速率随海拔的升高而降低,但其温度敏感性随海拔的升高而增加;增温实验表明,增温显著提高了青海云杉林土壤微生物代谢能力及多样性,随着土壤温度升高(1.3 ℃),土壤呼吸速率显著提高(30%~50%),在一定程度上削弱了干旱区山地森林的固碳能力[33]. ...
Changes in species diversity,aboveground biomass,and distribution characteristics along an afforestation successional gradient in semiarid Picea crassifolia plantations of Northwestern China
1
2017
... 阐明了山地人工林生态水文过程及其稳定性维持机制.Zhu等[34]系统研究了青海云杉人工林生态过程、水文过程以及生态水文相互作用,发现随着人工林的发育,林下草本层物种丰富度增加,但生物量显著下降;人工林建设显著改变了土壤理化性质,土壤有机碳、全氮含量增加,而pH值及容重降低;人工林空间分布格局从最初的大面积均匀分布逐渐演化成斑块状分布.由于郁闭度和叶面积指数较高,青海云杉人工林的林冠截留率(40.6%)高于天然林(35.1%),导致人工林地土壤水分补给受限,30~40年人工林的40~80 cm土层土壤含水量仅为天然林的60%[35-36].祁连山浅山区,青海云杉人工林应布局在坡向N、NE和NW,坡度15°~45°和海拔2 700~3 200 m的区域,人工林规模应不超过山坡坡面面积的60%[37]. ...
Spatial variability of canopy interception in a spruce forest of the semiarid mountain regions of China
1
2014
... 阐明了山地人工林生态水文过程及其稳定性维持机制.Zhu等[34]系统研究了青海云杉人工林生态过程、水文过程以及生态水文相互作用,发现随着人工林的发育,林下草本层物种丰富度增加,但生物量显著下降;人工林建设显著改变了土壤理化性质,土壤有机碳、全氮含量增加,而pH值及容重降低;人工林空间分布格局从最初的大面积均匀分布逐渐演化成斑块状分布.由于郁闭度和叶面积指数较高,青海云杉人工林的林冠截留率(40.6%)高于天然林(35.1%),导致人工林地土壤水分补给受限,30~40年人工林的40~80 cm土层土壤含水量仅为天然林的60%[35-36].祁连山浅山区,青海云杉人工林应布局在坡向N、NE和NW,坡度15°~45°和海拔2 700~3 200 m的区域,人工林规模应不超过山坡坡面面积的60%[37]. ...
Temporal variability in soil moisture after thinning in semi-arid Picea crassifolia plantations in northwestern China
1
2017
... 阐明了山地人工林生态水文过程及其稳定性维持机制.Zhu等[34]系统研究了青海云杉人工林生态过程、水文过程以及生态水文相互作用,发现随着人工林的发育,林下草本层物种丰富度增加,但生物量显著下降;人工林建设显著改变了土壤理化性质,土壤有机碳、全氮含量增加,而pH值及容重降低;人工林空间分布格局从最初的大面积均匀分布逐渐演化成斑块状分布.由于郁闭度和叶面积指数较高,青海云杉人工林的林冠截留率(40.6%)高于天然林(35.1%),导致人工林地土壤水分补给受限,30~40年人工林的40~80 cm土层土壤含水量仅为天然林的60%[35-36].祁连山浅山区,青海云杉人工林应布局在坡向N、NE和NW,坡度15°~45°和海拔2 700~3 200 m的区域,人工林规模应不超过山坡坡面面积的60%[37]. ...
Spatial patterns in natural Picea crassifolia forests of northwestern China,as basis for close-to-nature forestry
1
2018
... 阐明了山地人工林生态水文过程及其稳定性维持机制.Zhu等[34]系统研究了青海云杉人工林生态过程、水文过程以及生态水文相互作用,发现随着人工林的发育,林下草本层物种丰富度增加,但生物量显著下降;人工林建设显著改变了土壤理化性质,土壤有机碳、全氮含量增加,而pH值及容重降低;人工林空间分布格局从最初的大面积均匀分布逐渐演化成斑块状分布.由于郁闭度和叶面积指数较高,青海云杉人工林的林冠截留率(40.6%)高于天然林(35.1%),导致人工林地土壤水分补给受限,30~40年人工林的40~80 cm土层土壤含水量仅为天然林的60%[35-36].祁连山浅山区,青海云杉人工林应布局在坡向N、NE和NW,坡度15°~45°和海拔2 700~3 200 m的区域,人工林规模应不超过山坡坡面面积的60%[37]. ...
The modifiable areal unit problem of spatial heterogeneity of plant community in the transitional zone between oasis and desert using semivariance analysis
1
2007
... 荒漠植被不仅具有独特的结构、格局,而且形成了适应干旱风沙环境的机制.围绕荒漠、荒漠绿洲过渡带的植被组成、结构、格局及生态水文过程开展长期试验研究,揭示了微地形导致局地降水再分配是斑块植被格局形成的主要驱动力,而风沙活动塑造了斑块状植被格局和灌丛沙堆等典型植被景观的形成与演变[38-41].He等[42]研究表明,荒漠植被的物种组成呈空间尺度依赖性,其种-面积曲线方程S=cAz中的z值随尺度增加而增加的变化规律,与森林和草原结果相反.荒漠植被物种多样性随空间尺度增大其依赖性减弱,表现在种-面积曲线斜率z值随空间尺度增大呈幂函数降低,但调查的样方面积从1 m2增大到100 m2、1 km2时,z值从0.370降到0.213、0.035,并且在100 m2的尺度上出现拐点,表明100 m2调查样方基本可以反映荒漠植被物种多样性的信息,可以作为野外调查的最小面积单元. ...
Shielding effect of oasis-protection systems composed of various forms of wind break on sand fixation in an arid region:a case study in the Hexi Corridor,Northwest China
0
2008
Effects of sand burial depth and seed mass on seedling emergence and growth of Nitraria sphaerocarpa
0
2006
Effects of wind erosion and sand burial on growth and reproduction of a clonal shrub
1
2015
... 荒漠植被不仅具有独特的结构、格局,而且形成了适应干旱风沙环境的机制.围绕荒漠、荒漠绿洲过渡带的植被组成、结构、格局及生态水文过程开展长期试验研究,揭示了微地形导致局地降水再分配是斑块植被格局形成的主要驱动力,而风沙活动塑造了斑块状植被格局和灌丛沙堆等典型植被景观的形成与演变[38-41].He等[42]研究表明,荒漠植被的物种组成呈空间尺度依赖性,其种-面积曲线方程S=cAz中的z值随尺度增加而增加的变化规律,与森林和草原结果相反.荒漠植被物种多样性随空间尺度增大其依赖性减弱,表现在种-面积曲线斜率z值随空间尺度增大呈幂函数降低,但调查的样方面积从1 m2增大到100 m2、1 km2时,z值从0.370降到0.213、0.035,并且在100 m2的尺度上出现拐点,表明100 m2调查样方基本可以反映荒漠植被物种多样性的信息,可以作为野外调查的最小面积单元. ...
Scale dependence in desert plant diversity
2
2005
... 荒漠植被不仅具有独特的结构、格局,而且形成了适应干旱风沙环境的机制.围绕荒漠、荒漠绿洲过渡带的植被组成、结构、格局及生态水文过程开展长期试验研究,揭示了微地形导致局地降水再分配是斑块植被格局形成的主要驱动力,而风沙活动塑造了斑块状植被格局和灌丛沙堆等典型植被景观的形成与演变[38-41].He等[42]研究表明,荒漠植被的物种组成呈空间尺度依赖性,其种-面积曲线方程S=cAz中的z值随尺度增加而增加的变化规律,与森林和草原结果相反.荒漠植被物种多样性随空间尺度增大其依赖性减弱,表现在种-面积曲线斜率z值随空间尺度增大呈幂函数降低,但调查的样方面积从1 m2增大到100 m2、1 km2时,z值从0.370降到0.213、0.035,并且在100 m2的尺度上出现拐点,表明100 m2调查样方基本可以反映荒漠植被物种多样性的信息,可以作为野外调查的最小面积单元. ...
... 荒漠植被呈现明显的斑块状格局和空间异质性,不同生境灌丛斑块大小和空间格局差异显著.Luo等[43]发现,泡泡刺种群在荒漠生境中趋向于小斑块、大密度,空间自相关尺度较小(15 m),而在沙丘生境中趋向于大斑块、小密度,空间自相关尺度大(25 m).植被空间异质性越高,对空间尺度依赖性越强,尤其是在荒漠-绿洲过渡带.He等[42]对荒漠绿洲过渡带泡泡刺和红砂种群研究表明,样地面积和样方粒度对植被空间格局及异质性有显著影响.若样地面积过小,会忽略环境因子的影响,据此计算的空间异质性参数不能反映植被的真实情况;当样地面积大而样方粒度过小时,导致样方间的数据差异大,掩盖由空间位置不同带来的差异;样方太大,样方间的数据趋于同质,也难以反映植被空间异质性.因此,样地和样方的选择需根据植被格局对可塑性面积单元的敏感度来判断.如泡泡刺种群的样地应不小于200 m×200 m,红砂种群应不小于100 m×100 m,样方粒度应为20 m×20 m至30 m×30 m. ...
Spatial characteristics of two dominant shrub populations in the transition zone between oasis and desert in the Heihe River Basin,China
1
2018
... 荒漠植被呈现明显的斑块状格局和空间异质性,不同生境灌丛斑块大小和空间格局差异显著.Luo等[43]发现,泡泡刺种群在荒漠生境中趋向于小斑块、大密度,空间自相关尺度较小(15 m),而在沙丘生境中趋向于大斑块、小密度,空间自相关尺度大(25 m).植被空间异质性越高,对空间尺度依赖性越强,尤其是在荒漠-绿洲过渡带.He等[42]对荒漠绿洲过渡带泡泡刺和红砂种群研究表明,样地面积和样方粒度对植被空间格局及异质性有显著影响.若样地面积过小,会忽略环境因子的影响,据此计算的空间异质性参数不能反映植被的真实情况;当样地面积大而样方粒度过小时,导致样方间的数据差异大,掩盖由空间位置不同带来的差异;样方太大,样方间的数据趋于同质,也难以反映植被空间异质性.因此,样地和样方的选择需根据植被格局对可塑性面积单元的敏感度来判断.如泡泡刺种群的样地应不小于200 m×200 m,红砂种群应不小于100 m×100 m,样方粒度应为20 m×20 m至30 m×30 m. ...
Spatial pattern variation of artificial sand-binding vegetation based on UAV imagery and its influencing factors in an oasis-desert transitional zone
1
2022
... 荒漠区人工固沙植被格局向斑块状演变.采用空间代替时间的方法研究了40年人工梭梭固沙植被个体分异、种群分布和格局演变.20年左右,人工梭梭固沙植被种群密度减小了60%,30年左右植株冠幅最大,出现大量的天然更新幼苗,种群空间格局由均匀分布转变为斑块状[44-45].这一变化的内在驱动因素是种内竞争.5~10、10~20、20~30年的梭梭,种内竞争作用影响梭梭生长、存活以及自疏过程的发生.30~40年及更长时间的梭梭林则受到非对称竞争和非生物因素的共同作用,导致种群空间格局的演变[46].土壤水分的变化是梭梭植被格局变化的主要外部驱动因素,0~100 cm土壤含水量由2%~3%下降到1%左右.在土壤水分1%时,若前一年的6—8月累积降水量低于160 m,则梭梭地上净初级生产力明显下降,连续几年甚至导致林木枯死[47].风沙活动对天然沙丘植被演变具有重要影响.庄艳丽等[48]研究发现,风沙活动是控制沙拐枣群落演变的重要驱动因素,不仅影响植被生长和分布,也影响植被群落结构、格局和稳定性. ...
绿洲边缘人工固沙植被自组织过程
1
2018
... 荒漠区人工固沙植被格局向斑块状演变.采用空间代替时间的方法研究了40年人工梭梭固沙植被个体分异、种群分布和格局演变.20年左右,人工梭梭固沙植被种群密度减小了60%,30年左右植株冠幅最大,出现大量的天然更新幼苗,种群空间格局由均匀分布转变为斑块状[44-45].这一变化的内在驱动因素是种内竞争.5~10、10~20、20~30年的梭梭,种内竞争作用影响梭梭生长、存活以及自疏过程的发生.30~40年及更长时间的梭梭林则受到非对称竞争和非生物因素的共同作用,导致种群空间格局的演变[46].土壤水分的变化是梭梭植被格局变化的主要外部驱动因素,0~100 cm土壤含水量由2%~3%下降到1%左右.在土壤水分1%时,若前一年的6—8月累积降水量低于160 m,则梭梭地上净初级生产力明显下降,连续几年甚至导致林木枯死[47].风沙活动对天然沙丘植被演变具有重要影响.庄艳丽等[48]研究发现,风沙活动是控制沙拐枣群落演变的重要驱动因素,不仅影响植被生长和分布,也影响植被群落结构、格局和稳定性. ...
基于V_Hegyi 竞争指数的绿洲边缘人工固沙植被梭梭 (Haloxylon ammodendron)的种群竞争
1
2017
... 荒漠区人工固沙植被格局向斑块状演变.采用空间代替时间的方法研究了40年人工梭梭固沙植被个体分异、种群分布和格局演变.20年左右,人工梭梭固沙植被种群密度减小了60%,30年左右植株冠幅最大,出现大量的天然更新幼苗,种群空间格局由均匀分布转变为斑块状[44-45].这一变化的内在驱动因素是种内竞争.5~10、10~20、20~30年的梭梭,种内竞争作用影响梭梭生长、存活以及自疏过程的发生.30~40年及更长时间的梭梭林则受到非对称竞争和非生物因素的共同作用,导致种群空间格局的演变[46].土壤水分的变化是梭梭植被格局变化的主要外部驱动因素,0~100 cm土壤含水量由2%~3%下降到1%左右.在土壤水分1%时,若前一年的6—8月累积降水量低于160 m,则梭梭地上净初级生产力明显下降,连续几年甚至导致林木枯死[47].风沙活动对天然沙丘植被演变具有重要影响.庄艳丽等[48]研究发现,风沙活动是控制沙拐枣群落演变的重要驱动因素,不仅影响植被生长和分布,也影响植被群落结构、格局和稳定性. ...
Hydrologic thresholds and changes in ANPP of artificial sand-fixing vegetation in a desert-oasis ecotone in Northwest China
1
2017
... 荒漠区人工固沙植被格局向斑块状演变.采用空间代替时间的方法研究了40年人工梭梭固沙植被个体分异、种群分布和格局演变.20年左右,人工梭梭固沙植被种群密度减小了60%,30年左右植株冠幅最大,出现大量的天然更新幼苗,种群空间格局由均匀分布转变为斑块状[44-45].这一变化的内在驱动因素是种内竞争.5~10、10~20、20~30年的梭梭,种内竞争作用影响梭梭生长、存活以及自疏过程的发生.30~40年及更长时间的梭梭林则受到非对称竞争和非生物因素的共同作用,导致种群空间格局的演变[46].土壤水分的变化是梭梭植被格局变化的主要外部驱动因素,0~100 cm土壤含水量由2%~3%下降到1%左右.在土壤水分1%时,若前一年的6—8月累积降水量低于160 m,则梭梭地上净初级生产力明显下降,连续几年甚至导致林木枯死[47].风沙活动对天然沙丘植被演变具有重要影响.庄艳丽等[48]研究发现,风沙活动是控制沙拐枣群落演变的重要驱动因素,不仅影响植被生长和分布,也影响植被群落结构、格局和稳定性. ...
沙地生境沙拐枣(Calligonum mongolicum)种群特征及其扩张
1
2008
... 荒漠区人工固沙植被格局向斑块状演变.采用空间代替时间的方法研究了40年人工梭梭固沙植被个体分异、种群分布和格局演变.20年左右,人工梭梭固沙植被种群密度减小了60%,30年左右植株冠幅最大,出现大量的天然更新幼苗,种群空间格局由均匀分布转变为斑块状[44-45].这一变化的内在驱动因素是种内竞争.5~10、10~20、20~30年的梭梭,种内竞争作用影响梭梭生长、存活以及自疏过程的发生.30~40年及更长时间的梭梭林则受到非对称竞争和非生物因素的共同作用,导致种群空间格局的演变[46].土壤水分的变化是梭梭植被格局变化的主要外部驱动因素,0~100 cm土壤含水量由2%~3%下降到1%左右.在土壤水分1%时,若前一年的6—8月累积降水量低于160 m,则梭梭地上净初级生产力明显下降,连续几年甚至导致林木枯死[47].风沙活动对天然沙丘植被演变具有重要影响.庄艳丽等[48]研究发现,风沙活动是控制沙拐枣群落演变的重要驱动因素,不仅影响植被生长和分布,也影响植被群落结构、格局和稳定性. ...
1
2019
... 植物根据光合作用途径分为C3、C4和景天酸植物,甄别干旱荒漠区植物的光合途径对提高生态系统固碳能力、应对气候变化具有重要意义.基于组织结构和C同位素特征甄别了常见荒漠植物的光合途径.研究表明,梭梭和沙拐枣都具有花环结构,同化枝的δ13C值分别为-14.3‰和-14.8‰,二氧化碳补偿点分别为1.6 μmol·mol-1和4.3 μmol·mol-1,光饱和点分别为1 660 μmol·mol-1和1 755.6 μmol·mol-1.证实了这两种木本植物均具C4光合特征,其光合途径不因生长季节和水分条件的变化而改变.在雨后湿润条件下,光补偿点降低,而光饱和点升高,光能利用率显著提高.柠条、泡泡刺、花棒、红砂叶片的δ13C值分别为-25.8‰、-25.8‰、-26.4‰、-28.1‰; 沙枣、胡杨分别为-28.1‰、-29.7‰,均为C3植物[49]. ...
The response of sap flow in desert shrubs to environmental variables in an arid region of China
1
2011
... 研究发现,干旱区0~0.2 m土层土壤水分含量对降水的响应最显著,降水量与降水后土壤最大入渗深度呈指数函数关系[50].人工固沙植被梭梭根系分布有利于利用小降水事件,但对短期降水变化的适应性调节能力有限.在叶片尺度,梭梭的净光合速率、气孔导度、水势、蒸腾速率等对降水的响应显著[51];梭梭光合水分关系对降水脉动的响应主要通过对环境因子的响应来间接实现;在水分条件差的环境下,梭梭抗旱能力较强.当降水量小于5 mm时,梭梭净光合速率对降水的响应不明显;而在14 mm降水后, 净光合速率增加20%,降水前后土壤湿度的变化决定了梭梭光合水分生理对降水脉动的响应幅度[51]. ...
Physiological responses of Haloxylon ammodendron to rainfall pulses in temperate desert regions,northwestern China
4
2014
... 研究发现,干旱区0~0.2 m土层土壤水分含量对降水的响应最显著,降水量与降水后土壤最大入渗深度呈指数函数关系[50].人工固沙植被梭梭根系分布有利于利用小降水事件,但对短期降水变化的适应性调节能力有限.在叶片尺度,梭梭的净光合速率、气孔导度、水势、蒸腾速率等对降水的响应显著[51];梭梭光合水分关系对降水脉动的响应主要通过对环境因子的响应来间接实现;在水分条件差的环境下,梭梭抗旱能力较强.当降水量小于5 mm时,梭梭净光合速率对降水的响应不明显;而在14 mm降水后, 净光合速率增加20%,降水前后土壤湿度的变化决定了梭梭光合水分生理对降水脉动的响应幅度[51]. ...
... [51]. ...
... 一次降水事件后,梭梭的蒸腾速率(树干液流)随降水量的增加而显著提升,最小响应阈值为1.4 mm,滞后时间为1.4 d;最大阈值为12.3 mm,滞后时间为1 d.净光合速率(Rn)、气孔导度(Gs)也随降水量增加,但水分利用效率(WUE)对降水的响应不显著[51].Rnmax与Gs在降水事件后随时间的推移先升高后降低,但WUE呈现先降低后增加的趋势.例如,当干旱导致梭梭枝条的水势为3.09±0.05 MPa时,生长开始受到抑制,3.5~4.0 MPa时处于严重水分胁迫,导致生长停止.在这种情况下发生降水时,梭梭迅速做出响应,并在降水后第1~3天水分状况较好,但从第5天开始,水分状况显著下降.此期间的前3天梭梭胞间CO2浓度(Ci)与Rn下降,而Ls上升,表明该时期Rn下降主要是由气孔导度降低引起的;从第5天开始,Ci上升,而Ls与Rn下降,表明该过程Rn下降主要是由非气孔因素决定的[51]. ...
... [51]. ...
Modeling the physiological responses of a desert shrub to rainfall pulses in an arid environment in northwestern China
2
2020
... 在SPAC模型框架下,植入了基于降水脉动对植物生理影响的简化模型.通过同步分析光合作用、蒸腾作用和土壤水分动态,探讨降水脉动对光合作用过程的影响.模拟结果表明,在土壤干旱状态下,土壤水势和植物水势对降水脉动的响应较为显著[52];梭梭水势对降水脉动的响应较气孔导度(Gs)和光合速率(Rn)显著;水分利用效率(WUE)对降水脉动的响应不显著;初始土壤水分条件影响植物生理对降水脉动的响应,土壤水势和植物水势对降水脉动的响应比较显著.当植物处于严重干旱胁迫的情形下,较大的降水事件能使植物生理产生明显的响应,净光合速率的变化与降水脉动前的水分胁迫强度有关[52].因此,考虑初始土壤水分以及降水后土壤含水量的变化有助于理解植物的响应机制. ...
... [52].因此,考虑初始土壤水分以及降水后土壤含水量的变化有助于理解植物的响应机制. ...
The response of aboveground net primary productivity of desert vegetation to rainfall pulse in the temperate desert region of Northwest China
2
2013
... 基于阈值-延滞模型,选取了2005—2012年17次独立降水事件及降水前后的NDVI,量化了温带荒漠区砾质荒漠(砾漠)和沙质荒漠(沙漠)植被NDVI对降水脉动的响应,包括最大、最小降水阈值及响应量、响应滞后性和响应持续期等响应参数;研究表明,荒漠区植被ANPP开始响应和发生最大响应的单次降水阈值分别为5 mm和30 mm;砾漠植被响应持续期较沙漠长,响应滞后期为8~16 d,且沙漠植被响应值是砾漠的2.5倍.砾漠植被生长季出现多次生产力峰值,而沙漠植被为单次高峰[53].沿降水量80~200 mm梯度,砾质荒漠NDVI对当年3—8月的累积降水响应的敏感性逐渐增加,而对前一年9月至当年3月的累积降水量响应的敏感性逐渐减弱[54].相同降水条件下,砾漠植被对暖季降水响应较沙漠植被不明显[53]. ...
... [53]. ...
Productivity responses of desert vegetation to precipitation patterns across a rainfall gradient
1
2014
... 基于阈值-延滞模型,选取了2005—2012年17次独立降水事件及降水前后的NDVI,量化了温带荒漠区砾质荒漠(砾漠)和沙质荒漠(沙漠)植被NDVI对降水脉动的响应,包括最大、最小降水阈值及响应量、响应滞后性和响应持续期等响应参数;研究表明,荒漠区植被ANPP开始响应和发生最大响应的单次降水阈值分别为5 mm和30 mm;砾漠植被响应持续期较沙漠长,响应滞后期为8~16 d,且沙漠植被响应值是砾漠的2.5倍.砾漠植被生长季出现多次生产力峰值,而沙漠植被为单次高峰[53].沿降水量80~200 mm梯度,砾质荒漠NDVI对当年3—8月的累积降水响应的敏感性逐渐增加,而对前一年9月至当年3月的累积降水量响应的敏感性逐渐减弱[54].相同降水条件下,砾漠植被对暖季降水响应较沙漠植被不明显[53]. ...
Contrasting water-use strategies of coexisting xerophyte shrubs in a desert groundwater-dependent vegetation ecosystem
1
2025
... 运用稳定同位素技术和植物水分来源贡献率分析模型,确定了绿洲外缘固沙植被区降水、地下水以及不同深度土壤水的稳定同位素季节动态变化和对降水的响应过程[55-56],同步分析了不同年龄梭梭、沙拐枣、柠条和泡泡刺茎水的稳定同位素变化,揭示了固沙植被水分来源随季节和降水脉动的变化特征[57-59].随着梭梭的发育,主要水分来源由受降水补给的表层土壤水转变为地下水.5年和10年人工梭梭植被的水分来源季节变化较大,而20年和40年梭梭接近90%的水分来源于深层土壤水和地下水,季节变化较小[59].泡泡刺的水分来源季节动态显著,夏季降水贡献60%[60-61];沙拐枣和柠条主要依赖深层土壤水和地下水.5、10年梭梭和泡泡刺对降水响应显著,但20、40年梭梭、沙拐枣和柠条响应幅度较小[57-58]. ...
河西走廊典型荒漠区土壤水分对降水脉动响应的稳定同位素分析
1
2016
... 运用稳定同位素技术和植物水分来源贡献率分析模型,确定了绿洲外缘固沙植被区降水、地下水以及不同深度土壤水的稳定同位素季节动态变化和对降水的响应过程[55-56],同步分析了不同年龄梭梭、沙拐枣、柠条和泡泡刺茎水的稳定同位素变化,揭示了固沙植被水分来源随季节和降水脉动的变化特征[57-59].随着梭梭的发育,主要水分来源由受降水补给的表层土壤水转变为地下水.5年和10年人工梭梭植被的水分来源季节变化较大,而20年和40年梭梭接近90%的水分来源于深层土壤水和地下水,季节变化较小[59].泡泡刺的水分来源季节动态显著,夏季降水贡献60%[60-61];沙拐枣和柠条主要依赖深层土壤水和地下水.5、10年梭梭和泡泡刺对降水响应显著,但20、40年梭梭、沙拐枣和柠条响应幅度较小[57-58]. ...
Seasonal water sources and response to precipitation events of Calligonum mongolicum at the oasis margin in the Hexi River Corridor
2
2024
... 运用稳定同位素技术和植物水分来源贡献率分析模型,确定了绿洲外缘固沙植被区降水、地下水以及不同深度土壤水的稳定同位素季节动态变化和对降水的响应过程[55-56],同步分析了不同年龄梭梭、沙拐枣、柠条和泡泡刺茎水的稳定同位素变化,揭示了固沙植被水分来源随季节和降水脉动的变化特征[57-59].随着梭梭的发育,主要水分来源由受降水补给的表层土壤水转变为地下水.5年和10年人工梭梭植被的水分来源季节变化较大,而20年和40年梭梭接近90%的水分来源于深层土壤水和地下水,季节变化较小[59].泡泡刺的水分来源季节动态显著,夏季降水贡献60%[60-61];沙拐枣和柠条主要依赖深层土壤水和地下水.5、10年梭梭和泡泡刺对降水响应显著,但20、40年梭梭、沙拐枣和柠条响应幅度较小[57-58]. ...
... [57-58]. ...
Stable isotopes reveal varying water sources of Caragana microphylla in a desert-oasis ecotone near the Badain Jaran Desert
1
2018
... 运用稳定同位素技术和植物水分来源贡献率分析模型,确定了绿洲外缘固沙植被区降水、地下水以及不同深度土壤水的稳定同位素季节动态变化和对降水的响应过程[55-56],同步分析了不同年龄梭梭、沙拐枣、柠条和泡泡刺茎水的稳定同位素变化,揭示了固沙植被水分来源随季节和降水脉动的变化特征[57-59].随着梭梭的发育,主要水分来源由受降水补给的表层土壤水转变为地下水.5年和10年人工梭梭植被的水分来源季节变化较大,而20年和40年梭梭接近90%的水分来源于深层土壤水和地下水,季节变化较小[59].泡泡刺的水分来源季节动态显著,夏季降水贡献60%[60-61];沙拐枣和柠条主要依赖深层土壤水和地下水.5、10年梭梭和泡泡刺对降水响应显著,但20、40年梭梭、沙拐枣和柠条响应幅度较小[57-58]. ...
Varying water utilization of Haloxylon ammodendron plantations in a desert-oasis ecotone
2
2017
... 运用稳定同位素技术和植物水分来源贡献率分析模型,确定了绿洲外缘固沙植被区降水、地下水以及不同深度土壤水的稳定同位素季节动态变化和对降水的响应过程[55-56],同步分析了不同年龄梭梭、沙拐枣、柠条和泡泡刺茎水的稳定同位素变化,揭示了固沙植被水分来源随季节和降水脉动的变化特征[57-59].随着梭梭的发育,主要水分来源由受降水补给的表层土壤水转变为地下水.5年和10年人工梭梭植被的水分来源季节变化较大,而20年和40年梭梭接近90%的水分来源于深层土壤水和地下水,季节变化较小[59].泡泡刺的水分来源季节动态显著,夏季降水贡献60%[60-61];沙拐枣和柠条主要依赖深层土壤水和地下水.5、10年梭梭和泡泡刺对降水响应显著,但20、40年梭梭、沙拐枣和柠条响应幅度较小[57-58]. ...
... [59].泡泡刺的水分来源季节动态显著,夏季降水贡献60%[60-61];沙拐枣和柠条主要依赖深层土壤水和地下水.5、10年梭梭和泡泡刺对降水响应显著,但20、40年梭梭、沙拐枣和柠条响应幅度较小[57-58]. ...
Isotopic evidence of hydrological niche segregation and species coexisting mechanism of desert shrubs Reaumuria songarica and Nitraria sphaerocarpa
1
2025
... 运用稳定同位素技术和植物水分来源贡献率分析模型,确定了绿洲外缘固沙植被区降水、地下水以及不同深度土壤水的稳定同位素季节动态变化和对降水的响应过程[55-56],同步分析了不同年龄梭梭、沙拐枣、柠条和泡泡刺茎水的稳定同位素变化,揭示了固沙植被水分来源随季节和降水脉动的变化特征[57-59].随着梭梭的发育,主要水分来源由受降水补给的表层土壤水转变为地下水.5年和10年人工梭梭植被的水分来源季节变化较大,而20年和40年梭梭接近90%的水分来源于深层土壤水和地下水,季节变化较小[59].泡泡刺的水分来源季节动态显著,夏季降水贡献60%[60-61];沙拐枣和柠条主要依赖深层土壤水和地下水.5、10年梭梭和泡泡刺对降水响应显著,但20、40年梭梭、沙拐枣和柠条响应幅度较小[57-58]. ...
两种荒漠生境条件下泡泡刺水分来源及其对降水的响应
1
2017
... 运用稳定同位素技术和植物水分来源贡献率分析模型,确定了绿洲外缘固沙植被区降水、地下水以及不同深度土壤水的稳定同位素季节动态变化和对降水的响应过程[55-56],同步分析了不同年龄梭梭、沙拐枣、柠条和泡泡刺茎水的稳定同位素变化,揭示了固沙植被水分来源随季节和降水脉动的变化特征[57-59].随着梭梭的发育,主要水分来源由受降水补给的表层土壤水转变为地下水.5年和10年人工梭梭植被的水分来源季节变化较大,而20年和40年梭梭接近90%的水分来源于深层土壤水和地下水,季节变化较小[59].泡泡刺的水分来源季节动态显著,夏季降水贡献60%[60-61];沙拐枣和柠条主要依赖深层土壤水和地下水.5、10年梭梭和泡泡刺对降水响应显著,但20、40年梭梭、沙拐枣和柠条响应幅度较小[57-58]. ...
Characterizing the spatial structure of riparian plant communities in the lower reaches of Heihe River in China using geostatistical techniques
1
2006
... 荒漠河岸林是内陆河流域三角洲地区的独特森林景观,受控于地表水和地下水文过程.黑河下游额济纳三角洲荒漠林在垂直于河流的方向上分布着胡杨、柽柳、苦豆子等群落,呈现斑块格局,斑块内部空间异质性小,而斑块之间异质性较大.在河道至荒漠的植被样带上,空间异质性变化最大的尺度为400 m,符合荒漠河岸林种群替换尺度[62]. ...
Responses of distribution pattern of desert riparian forests to hydrologic process in Ejina Oasis
1
2004
... 1980—2000年,黑河上中游用水增加,导致下泄到额济纳的水量减少,引起了荒漠河岸林结构和格局变化.表现为植被斑块化加剧,斑块空间异质性增强,退化林地占比增加.地表水来源减少和地下水位下降是主要原因,此外河岸林中斑块状农田也是影响因素[63]. ...
人工调水对胡杨荒漠河岸林繁殖的影响
1
2005
... 荒漠河岸林优势植物胡杨主要通过种子和根蘖两种方式繁殖.1980年以前,黑河水文过程未发生明显改变,每年7—8月,祁连山冰雪融化导致额济纳三角洲形成的漫流和无数沙洲,为胡杨种子更新提供了条件.但自1980年以来,上中游用水增加,下泄到额济纳的水量减少,很少发生洪水漫流过程,导致胡杨无法进行种子更新.2005年调查显示,额济纳胡杨林自然保护区内近10年几乎没有种子繁殖的幼林,根蘖苗主要为1~3龄,4龄以上幼苗稀少.调查表明,胡杨幼苗的个体密度为213.19±112.15 株·hm-1,其中2龄幼苗最多(113.18±6.717株·hm-1).黑河分水工程自2000年开始,胡杨林保护区自2001年起全面禁牧,胡杨幼苗生长与分水工程实施和保护区围封禁牧的时间一致.在保护区内的样方调查中未出现苗龄超过4龄的胡杨幼林,表明在没有禁牧的条件下,胡杨幼苗不能定植成林.黑河分水工程有助于胡杨繁殖更新,但需消除放牧等干扰因素[64]. ...
黑河下游额济纳绿洲近20年生态输水的生态恢复效应
1
2022
... 生态分水工程实施后,额济纳绿洲生态恢复明显,表现为绿洲结构变化,绿洲规模、植被盖度和初级生产力显著增加.2000—2020年,土地利用/覆被发生明显变化,其中,湿地、林地和草地面积分别增加了79.33 km2(137%)、61.73 km2(56%)和50.26 km2(6%).景观格局逐步改善,景观多样性指数增加了14.91%.植被盖度、植被面积及NPP明显提升,其中,42.34%的区域年均NDVI显著增加,2006年以后NDVI增速明显加快;平均植被覆盖度增加了7.78%;绿洲植被面积增加了约1 100 km2;植被NPP(以C计量)增加了26.72 g·m-2a-1,植被固碳量增加了0.24 Tg(30.12%)[65]. ...
Runoff controls the development of eco-hydrological and economic conditions in an arid oasis of the downstream inland river basin
1
2022
... 用空间分析、趋势分析、回归分析和结构方程模型等方法,识别了生态输水工程实施以来(2000—2020年)额济纳绿洲地下水埋深、地表水体及NDVI的时空变化,分析了绿洲水文-生态指标对生态输水量的响应,探讨了绿洲植被恢复驱动因素的结构与强度.结果表明:2000—2020年,狼心山径流量呈显著增加趋势,年均增加2.7×107 m3,额济纳绿洲平均地下水埋深上升了0.41 m,地表水体面积增加了130.62 km2;NDVI显著增加,增加区域占42.34%,主要在西河中游和东河下游;西河地区NDVI对径流量的响应较东河地区更敏感;当狼心山年径流量大于6×108 m3、平均地下水埋深小于2.8 m时,绿洲植被面积维持在较高水平;径流量、地下水埋深及耕地面积等因素对绿洲植被具有不同程度的直接和间接影响;黑河下游径流量与地下水稳定性是额济纳绿洲恢复与可持续发展的关键[66]. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}