Global desertification:building a science for dryland development
1
2007
... 干旱半干旱区约占地球陆表面积的40%以上,是陆地生态系统中分布最广、对气候变化响应最敏感的区域之一[1].其中,荒漠生态系统作为干旱区水分限制最突出的组成部分,其植被分布与结构特征对水分变化具有高度敏感性[2].水分供给,尤其是降水的时空分布特征,是制约荒漠植被分布格局、群落结构及生态功能的首要环境因子[3].沿降水梯度,植被覆盖度、群落组成、多样性及空间格局通常呈现出系统性变化,是认识干旱区生态系统结构与功能演变的重要切入点[4].已有研究表明,降水通常通过调节土壤水分状况影响植物生长、生物量分配及物种组成,从而对群落多样性和空间格局产生重要影响.在多数情况下,降水增加能够促进物种丰富度和均匀度的提升,但这一响应关系往往受生态系统类型、土壤条件及物种组成等因素的共同调节,通常具有一定的时间滞后性[5-7].同时,在不同气候背景和土壤条件下,植被对降水梯度的响应形式及其阈值特征仍存在差异,尤其是在极端干旱环境中,现有研究多依赖遥感反演或控制试验,对基于样地调查的自然降水梯度响应研究不足[6,8-9]. ...
Desert ecosystems:environment and producers
1
1973
... 干旱半干旱区约占地球陆表面积的40%以上,是陆地生态系统中分布最广、对气候变化响应最敏感的区域之一[1].其中,荒漠生态系统作为干旱区水分限制最突出的组成部分,其植被分布与结构特征对水分变化具有高度敏感性[2].水分供给,尤其是降水的时空分布特征,是制约荒漠植被分布格局、群落结构及生态功能的首要环境因子[3].沿降水梯度,植被覆盖度、群落组成、多样性及空间格局通常呈现出系统性变化,是认识干旱区生态系统结构与功能演变的重要切入点[4].已有研究表明,降水通常通过调节土壤水分状况影响植物生长、生物量分配及物种组成,从而对群落多样性和空间格局产生重要影响.在多数情况下,降水增加能够促进物种丰富度和均匀度的提升,但这一响应关系往往受生态系统类型、土壤条件及物种组成等因素的共同调节,通常具有一定的时间滞后性[5-7].同时,在不同气候背景和土壤条件下,植被对降水梯度的响应形式及其阈值特征仍存在差异,尤其是在极端干旱环境中,现有研究多依赖遥感反演或控制试验,对基于样地调查的自然降水梯度响应研究不足[6,8-9]. ...
Precipitation pulses and carbon fluxes in semiarid and arid ecosystems
1
2004
... 干旱半干旱区约占地球陆表面积的40%以上,是陆地生态系统中分布最广、对气候变化响应最敏感的区域之一[1].其中,荒漠生态系统作为干旱区水分限制最突出的组成部分,其植被分布与结构特征对水分变化具有高度敏感性[2].水分供给,尤其是降水的时空分布特征,是制约荒漠植被分布格局、群落结构及生态功能的首要环境因子[3].沿降水梯度,植被覆盖度、群落组成、多样性及空间格局通常呈现出系统性变化,是认识干旱区生态系统结构与功能演变的重要切入点[4].已有研究表明,降水通常通过调节土壤水分状况影响植物生长、生物量分配及物种组成,从而对群落多样性和空间格局产生重要影响.在多数情况下,降水增加能够促进物种丰富度和均匀度的提升,但这一响应关系往往受生态系统类型、土壤条件及物种组成等因素的共同调节,通常具有一定的时间滞后性[5-7].同时,在不同气候背景和土壤条件下,植被对降水梯度的响应形式及其阈值特征仍存在差异,尤其是在极端干旱环境中,现有研究多依赖遥感反演或控制试验,对基于样地调查的自然降水梯度响应研究不足[6,8-9]. ...
Productivity is a poor predictor of plant species richness
1
2011
... 干旱半干旱区约占地球陆表面积的40%以上,是陆地生态系统中分布最广、对气候变化响应最敏感的区域之一[1].其中,荒漠生态系统作为干旱区水分限制最突出的组成部分,其植被分布与结构特征对水分变化具有高度敏感性[2].水分供给,尤其是降水的时空分布特征,是制约荒漠植被分布格局、群落结构及生态功能的首要环境因子[3].沿降水梯度,植被覆盖度、群落组成、多样性及空间格局通常呈现出系统性变化,是认识干旱区生态系统结构与功能演变的重要切入点[4].已有研究表明,降水通常通过调节土壤水分状况影响植物生长、生物量分配及物种组成,从而对群落多样性和空间格局产生重要影响.在多数情况下,降水增加能够促进物种丰富度和均匀度的提升,但这一响应关系往往受生态系统类型、土壤条件及物种组成等因素的共同调节,通常具有一定的时间滞后性[5-7].同时,在不同气候背景和土壤条件下,植被对降水梯度的响应形式及其阈值特征仍存在差异,尤其是在极端干旱环境中,现有研究多依赖遥感反演或控制试验,对基于样地调查的自然降水梯度响应研究不足[6,8-9]. ...
中国旱区样带尺度植物多样性和生产力与环境因子的关系
1
2023
... 干旱半干旱区约占地球陆表面积的40%以上,是陆地生态系统中分布最广、对气候变化响应最敏感的区域之一[1].其中,荒漠生态系统作为干旱区水分限制最突出的组成部分,其植被分布与结构特征对水分变化具有高度敏感性[2].水分供给,尤其是降水的时空分布特征,是制约荒漠植被分布格局、群落结构及生态功能的首要环境因子[3].沿降水梯度,植被覆盖度、群落组成、多样性及空间格局通常呈现出系统性变化,是认识干旱区生态系统结构与功能演变的重要切入点[4].已有研究表明,降水通常通过调节土壤水分状况影响植物生长、生物量分配及物种组成,从而对群落多样性和空间格局产生重要影响.在多数情况下,降水增加能够促进物种丰富度和均匀度的提升,但这一响应关系往往受生态系统类型、土壤条件及物种组成等因素的共同调节,通常具有一定的时间滞后性[5-7].同时,在不同气候背景和土壤条件下,植被对降水梯度的响应形式及其阈值特征仍存在差异,尤其是在极端干旱环境中,现有研究多依赖遥感反演或控制试验,对基于样地调查的自然降水梯度响应研究不足[6,8-9]. ...
降水控制对荒漠植物群落物种多样性和生物量的影响
1
2018
... 干旱半干旱区约占地球陆表面积的40%以上,是陆地生态系统中分布最广、对气候变化响应最敏感的区域之一[1].其中,荒漠生态系统作为干旱区水分限制最突出的组成部分,其植被分布与结构特征对水分变化具有高度敏感性[2].水分供给,尤其是降水的时空分布特征,是制约荒漠植被分布格局、群落结构及生态功能的首要环境因子[3].沿降水梯度,植被覆盖度、群落组成、多样性及空间格局通常呈现出系统性变化,是认识干旱区生态系统结构与功能演变的重要切入点[4].已有研究表明,降水通常通过调节土壤水分状况影响植物生长、生物量分配及物种组成,从而对群落多样性和空间格局产生重要影响.在多数情况下,降水增加能够促进物种丰富度和均匀度的提升,但这一响应关系往往受生态系统类型、土壤条件及物种组成等因素的共同调节,通常具有一定的时间滞后性[5-7].同时,在不同气候背景和土壤条件下,植被对降水梯度的响应形式及其阈值特征仍存在差异,尤其是在极端干旱环境中,现有研究多依赖遥感反演或控制试验,对基于样地调查的自然降水梯度响应研究不足[6,8-9]. ...
Water-mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe
1
2008
... 干旱半干旱区约占地球陆表面积的40%以上,是陆地生态系统中分布最广、对气候变化响应最敏感的区域之一[1].其中,荒漠生态系统作为干旱区水分限制最突出的组成部分,其植被分布与结构特征对水分变化具有高度敏感性[2].水分供给,尤其是降水的时空分布特征,是制约荒漠植被分布格局、群落结构及生态功能的首要环境因子[3].沿降水梯度,植被覆盖度、群落组成、多样性及空间格局通常呈现出系统性变化,是认识干旱区生态系统结构与功能演变的重要切入点[4].已有研究表明,降水通常通过调节土壤水分状况影响植物生长、生物量分配及物种组成,从而对群落多样性和空间格局产生重要影响.在多数情况下,降水增加能够促进物种丰富度和均匀度的提升,但这一响应关系往往受生态系统类型、土壤条件及物种组成等因素的共同调节,通常具有一定的时间滞后性[5-7].同时,在不同气候背景和土壤条件下,植被对降水梯度的响应形式及其阈值特征仍存在差异,尤其是在极端干旱环境中,现有研究多依赖遥感反演或控制试验,对基于样地调查的自然降水梯度响应研究不足[6,8-9]. ...
降水变化对荒漠草原植物群落生物量及其权衡关系的影响
2
2023
... 干旱半干旱区约占地球陆表面积的40%以上,是陆地生态系统中分布最广、对气候变化响应最敏感的区域之一[1].其中,荒漠生态系统作为干旱区水分限制最突出的组成部分,其植被分布与结构特征对水分变化具有高度敏感性[2].水分供给,尤其是降水的时空分布特征,是制约荒漠植被分布格局、群落结构及生态功能的首要环境因子[3].沿降水梯度,植被覆盖度、群落组成、多样性及空间格局通常呈现出系统性变化,是认识干旱区生态系统结构与功能演变的重要切入点[4].已有研究表明,降水通常通过调节土壤水分状况影响植物生长、生物量分配及物种组成,从而对群落多样性和空间格局产生重要影响.在多数情况下,降水增加能够促进物种丰富度和均匀度的提升,但这一响应关系往往受生态系统类型、土壤条件及物种组成等因素的共同调节,通常具有一定的时间滞后性[5-7].同时,在不同气候背景和土壤条件下,植被对降水梯度的响应形式及其阈值特征仍存在差异,尤其是在极端干旱环境中,现有研究多依赖遥感反演或控制试验,对基于样地调查的自然降水梯度响应研究不足[6,8-9]. ...
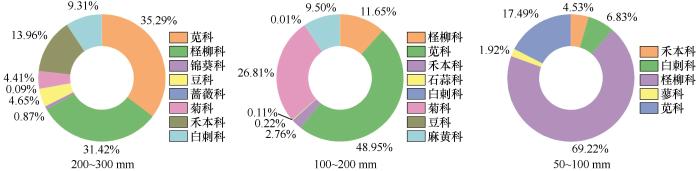
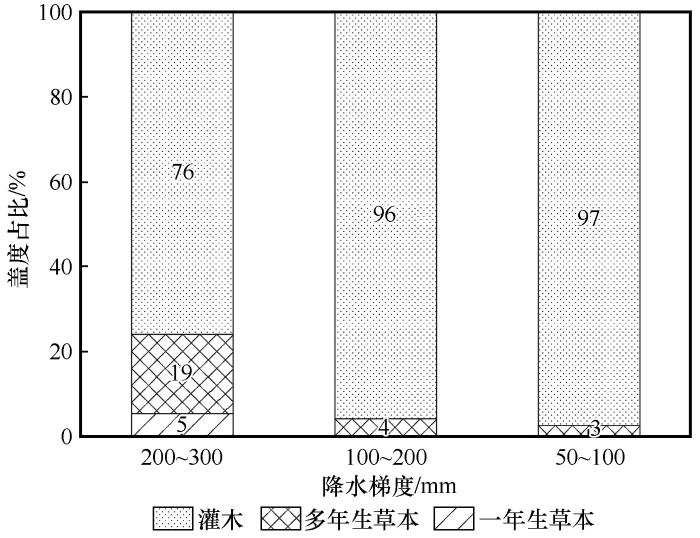
... 在3个降水梯度上,灌木均为群落的优势生活型,但群落结构对降水变化表现出明显的响应,在200~300 mm降水组禾本科与一年生草本的相对多度显著增加,与灌木共同构成群落;而在50~100 mm降水组的极端干旱区间,群落则几乎完全由梭梭、白刺、驼绒藜等耐旱灌木所主导(表1,图2和图3).这一变化趋势与低海拔干旱区的研究结果总体一致,如在宁夏荒漠草原的降水控制试验表明,随着降水减少,草本植物地上生物量显著下降,群落结构趋于简单,耐旱物种的重要性上升[32].相关研究进一步表明,降水变化主要通过直接影响豆科和禾本科植物地上生物量进而间接影响植物群落地上生物量[8].上述群落结构变化与荒漠植物的生存策略相一致,草本植物对水分变化更为敏感,在极度干旱条件下仅有少数耐旱性高的灌木能够维持稳定种群[20,33].植物多样性随降水量的减少显著降低.200~300 mm降水组的Shannon-Wiener指数和物种丰富度明显高于其余降水组,但群落均匀度指数组间差异不显著,这可能是由于沿降水梯度的多样性下降主要源于物种丰富度的减少,而非优势度结构的显著改变[34]. ...
降水变化下荒漠草原植物群落共存物种根、叶功能性状的差异与权衡
1
2026
... 干旱半干旱区约占地球陆表面积的40%以上,是陆地生态系统中分布最广、对气候变化响应最敏感的区域之一[1].其中,荒漠生态系统作为干旱区水分限制最突出的组成部分,其植被分布与结构特征对水分变化具有高度敏感性[2].水分供给,尤其是降水的时空分布特征,是制约荒漠植被分布格局、群落结构及生态功能的首要环境因子[3].沿降水梯度,植被覆盖度、群落组成、多样性及空间格局通常呈现出系统性变化,是认识干旱区生态系统结构与功能演变的重要切入点[4].已有研究表明,降水通常通过调节土壤水分状况影响植物生长、生物量分配及物种组成,从而对群落多样性和空间格局产生重要影响.在多数情况下,降水增加能够促进物种丰富度和均匀度的提升,但这一响应关系往往受生态系统类型、土壤条件及物种组成等因素的共同调节,通常具有一定的时间滞后性[5-7].同时,在不同气候背景和土壤条件下,植被对降水梯度的响应形式及其阈值特征仍存在差异,尤其是在极端干旱环境中,现有研究多依赖遥感反演或控制试验,对基于样地调查的自然降水梯度响应研究不足[6,8-9]. ...
喜马拉雅-青藏高原造山带地质演化:显生宙亚洲大陆生长
1
2001
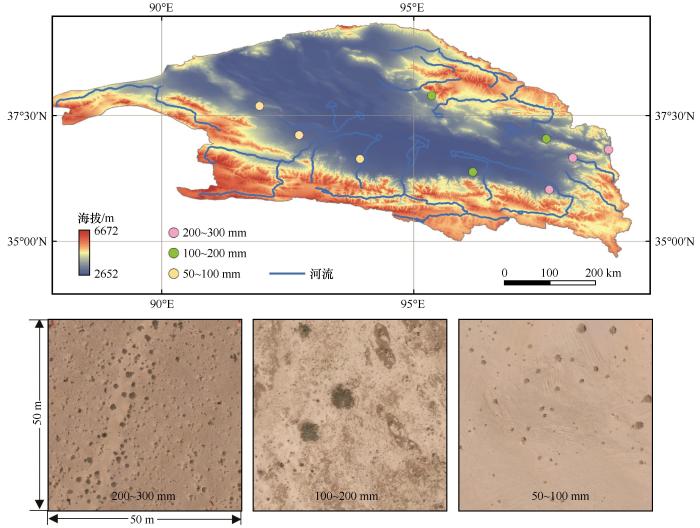
... 柴达木盆地位于青藏高原东北部的封闭内流地貌单元,四周山系环绕、地势起伏显著,是典型的内陆高原大陆性气候[10].在地形与大尺度环流共同作用下,年降水量自东南向西北急剧减少,由约250 mm降至不足15 mm;蒸发量远高于降水量,塑造出极端干旱环境[11].地表以砂质与砾质荒漠为主,盐沼与干涸湖盆呈斑块镶嵌分布,水盐时空异质性显著.植被覆盖度整体偏低,以耐旱灌木与一年生草本为主,呈现出典型的斑块镶嵌格局[12-14].在此背景下,沿降水梯度揭示植被组成、多样性与空间格局的变化特征,并阐明其对水分限制的生态响应,对于认识高原荒漠生态系统结构与功能演变、服务于干旱区生态恢复与管理具有重要意义. ...
近60年来柴达木盆地降水季节性变化分析
1
2023
... 柴达木盆地位于青藏高原东北部的封闭内流地貌单元,四周山系环绕、地势起伏显著,是典型的内陆高原大陆性气候[10].在地形与大尺度环流共同作用下,年降水量自东南向西北急剧减少,由约250 mm降至不足15 mm;蒸发量远高于降水量,塑造出极端干旱环境[11].地表以砂质与砾质荒漠为主,盐沼与干涸湖盆呈斑块镶嵌分布,水盐时空异质性显著.植被覆盖度整体偏低,以耐旱灌木与一年生草本为主,呈现出典型的斑块镶嵌格局[12-14].在此背景下,沿降水梯度揭示植被组成、多样性与空间格局的变化特征,并阐明其对水分限制的生态响应,对于认识高原荒漠生态系统结构与功能演变、服务于干旱区生态恢复与管理具有重要意义. ...
气候变化背景下柴达木盆地荒漠化时空演变及驱动力研究
1
2023
... 柴达木盆地位于青藏高原东北部的封闭内流地貌单元,四周山系环绕、地势起伏显著,是典型的内陆高原大陆性气候[10].在地形与大尺度环流共同作用下,年降水量自东南向西北急剧减少,由约250 mm降至不足15 mm;蒸发量远高于降水量,塑造出极端干旱环境[11].地表以砂质与砾质荒漠为主,盐沼与干涸湖盆呈斑块镶嵌分布,水盐时空异质性显著.植被覆盖度整体偏低,以耐旱灌木与一年生草本为主,呈现出典型的斑块镶嵌格局[12-14].在此背景下,沿降水梯度揭示植被组成、多样性与空间格局的变化特征,并阐明其对水分限制的生态响应,对于认识高原荒漠生态系统结构与功能演变、服务于干旱区生态恢复与管理具有重要意义. ...
2000-2021年柴达木盆地地表绿度变化及地形分异研究
2
2022
... 随着遥感技术的发展,基于中高分辨率卫星影像的植被指数分析已成为研究干旱区植被动态及其对降水响应的重要手段,在区域尺度揭示植被覆盖变化及其时空格局方面具有显著优势[15-16].已有研究表明,柴达木盆地植被绿度总体呈周缘升高、腹地偏低的空间格局,并在近几十年暖湿化背景下对降水变化表现出显著响应[13,17];植被物候、蒸散发及水分利用效率等生态特征亦随降水梯度和环境条件发生变化[18-20],如植被物候总体表现为生长季开始和结束时间推迟、生长期缩短,降水对物候的调控效应整体强于气温[18].然而,受空间分辨率和混合像元效应限制,传统卫星遥感难以准确刻画荒漠植被斑块的精细结构特征,尤其是在植被稀疏、斑块尺度较小的极端干旱区,其对群落尺度空间异质性和斑块格局的表征能力仍然有限[21-22].近年来,无人机遥感凭借厘米级空间分辨率、数据获取灵活等优势,在荒漠植被覆盖提取、斑块识别及空间格局量化等方面具有显著优势,为实现样方尺度与区域尺度之间的有效衔接提供了新的技术途径[23-24].而目前针对柴达木盆地自然降水梯度下的群落多样性与斑块格局研究,仍存在空间与尺度覆盖不足的问题.既往研究多局限于局地生境或单一群落类型,缺少覆盖完整降水梯度的代表性样区或样带[13,25-27].在柴达木盆地自然降水梯度背景下,将无人机获取的精细斑块格局指标与地面群落调查及土壤理化属性在相同空间尺度下进行综合分析的研究仍相对缺乏,一定程度上制约了对降水梯度下柴达木盆地荒漠植被与土壤及环境因素耦合机制的认识. ...
... [13,25-27].在柴达木盆地自然降水梯度背景下,将无人机获取的精细斑块格局指标与地面群落调查及土壤理化属性在相同空间尺度下进行综合分析的研究仍相对缺乏,一定程度上制约了对降水梯度下柴达木盆地荒漠植被与土壤及环境因素耦合机制的认识. ...
柴达木盆地典型风蚀区土壤质量评价
1
2023
... 柴达木盆地位于青藏高原东北部的封闭内流地貌单元,四周山系环绕、地势起伏显著,是典型的内陆高原大陆性气候[10].在地形与大尺度环流共同作用下,年降水量自东南向西北急剧减少,由约250 mm降至不足15 mm;蒸发量远高于降水量,塑造出极端干旱环境[11].地表以砂质与砾质荒漠为主,盐沼与干涸湖盆呈斑块镶嵌分布,水盐时空异质性显著.植被覆盖度整体偏低,以耐旱灌木与一年生草本为主,呈现出典型的斑块镶嵌格局[12-14].在此背景下,沿降水梯度揭示植被组成、多样性与空间格局的变化特征,并阐明其对水分限制的生态响应,对于认识高原荒漠生态系统结构与功能演变、服务于干旱区生态恢复与管理具有重要意义. ...
Red and photographic infrared linear combinations for monitoring vegetation
1
1979
... 随着遥感技术的发展,基于中高分辨率卫星影像的植被指数分析已成为研究干旱区植被动态及其对降水响应的重要手段,在区域尺度揭示植被覆盖变化及其时空格局方面具有显著优势[15-16].已有研究表明,柴达木盆地植被绿度总体呈周缘升高、腹地偏低的空间格局,并在近几十年暖湿化背景下对降水变化表现出显著响应[13,17];植被物候、蒸散发及水分利用效率等生态特征亦随降水梯度和环境条件发生变化[18-20],如植被物候总体表现为生长季开始和结束时间推迟、生长期缩短,降水对物候的调控效应整体强于气温[18].然而,受空间分辨率和混合像元效应限制,传统卫星遥感难以准确刻画荒漠植被斑块的精细结构特征,尤其是在植被稀疏、斑块尺度较小的极端干旱区,其对群落尺度空间异质性和斑块格局的表征能力仍然有限[21-22].近年来,无人机遥感凭借厘米级空间分辨率、数据获取灵活等优势,在荒漠植被覆盖提取、斑块识别及空间格局量化等方面具有显著优势,为实现样方尺度与区域尺度之间的有效衔接提供了新的技术途径[23-24].而目前针对柴达木盆地自然降水梯度下的群落多样性与斑块格局研究,仍存在空间与尺度覆盖不足的问题.既往研究多局限于局地生境或单一群落类型,缺少覆盖完整降水梯度的代表性样区或样带[13,25-27].在柴达木盆地自然降水梯度背景下,将无人机获取的精细斑块格局指标与地面群落调查及土壤理化属性在相同空间尺度下进行综合分析的研究仍相对缺乏,一定程度上制约了对降水梯度下柴达木盆地荒漠植被与土壤及环境因素耦合机制的认识. ...
A continuous satellite-derived measure of global terrestrial primary production
1
2004
... 随着遥感技术的发展,基于中高分辨率卫星影像的植被指数分析已成为研究干旱区植被动态及其对降水响应的重要手段,在区域尺度揭示植被覆盖变化及其时空格局方面具有显著优势[15-16].已有研究表明,柴达木盆地植被绿度总体呈周缘升高、腹地偏低的空间格局,并在近几十年暖湿化背景下对降水变化表现出显著响应[13,17];植被物候、蒸散发及水分利用效率等生态特征亦随降水梯度和环境条件发生变化[18-20],如植被物候总体表现为生长季开始和结束时间推迟、生长期缩短,降水对物候的调控效应整体强于气温[18].然而,受空间分辨率和混合像元效应限制,传统卫星遥感难以准确刻画荒漠植被斑块的精细结构特征,尤其是在植被稀疏、斑块尺度较小的极端干旱区,其对群落尺度空间异质性和斑块格局的表征能力仍然有限[21-22].近年来,无人机遥感凭借厘米级空间分辨率、数据获取灵活等优势,在荒漠植被覆盖提取、斑块识别及空间格局量化等方面具有显著优势,为实现样方尺度与区域尺度之间的有效衔接提供了新的技术途径[23-24].而目前针对柴达木盆地自然降水梯度下的群落多样性与斑块格局研究,仍存在空间与尺度覆盖不足的问题.既往研究多局限于局地生境或单一群落类型,缺少覆盖完整降水梯度的代表性样区或样带[13,25-27].在柴达木盆地自然降水梯度背景下,将无人机获取的精细斑块格局指标与地面群落调查及土壤理化属性在相同空间尺度下进行综合分析的研究仍相对缺乏,一定程度上制约了对降水梯度下柴达木盆地荒漠植被与土壤及环境因素耦合机制的认识. ...
柴达木盆地气候变化对植被的影响分析
1
2018
... 随着遥感技术的发展,基于中高分辨率卫星影像的植被指数分析已成为研究干旱区植被动态及其对降水响应的重要手段,在区域尺度揭示植被覆盖变化及其时空格局方面具有显著优势[15-16].已有研究表明,柴达木盆地植被绿度总体呈周缘升高、腹地偏低的空间格局,并在近几十年暖湿化背景下对降水变化表现出显著响应[13,17];植被物候、蒸散发及水分利用效率等生态特征亦随降水梯度和环境条件发生变化[18-20],如植被物候总体表现为生长季开始和结束时间推迟、生长期缩短,降水对物候的调控效应整体强于气温[18].然而,受空间分辨率和混合像元效应限制,传统卫星遥感难以准确刻画荒漠植被斑块的精细结构特征,尤其是在植被稀疏、斑块尺度较小的极端干旱区,其对群落尺度空间异质性和斑块格局的表征能力仍然有限[21-22].近年来,无人机遥感凭借厘米级空间分辨率、数据获取灵活等优势,在荒漠植被覆盖提取、斑块识别及空间格局量化等方面具有显著优势,为实现样方尺度与区域尺度之间的有效衔接提供了新的技术途径[23-24].而目前针对柴达木盆地自然降水梯度下的群落多样性与斑块格局研究,仍存在空间与尺度覆盖不足的问题.既往研究多局限于局地生境或单一群落类型,缺少覆盖完整降水梯度的代表性样区或样带[13,25-27].在柴达木盆地自然降水梯度背景下,将无人机获取的精细斑块格局指标与地面群落调查及土壤理化属性在相同空间尺度下进行综合分析的研究仍相对缺乏,一定程度上制约了对降水梯度下柴达木盆地荒漠植被与土壤及环境因素耦合机制的认识. ...
Spatial and temporal variation of vegetation phenology and its response to climate changes in qaidam basin from 2000 to 2015
2
2018
... 随着遥感技术的发展,基于中高分辨率卫星影像的植被指数分析已成为研究干旱区植被动态及其对降水响应的重要手段,在区域尺度揭示植被覆盖变化及其时空格局方面具有显著优势[15-16].已有研究表明,柴达木盆地植被绿度总体呈周缘升高、腹地偏低的空间格局,并在近几十年暖湿化背景下对降水变化表现出显著响应[13,17];植被物候、蒸散发及水分利用效率等生态特征亦随降水梯度和环境条件发生变化[18-20],如植被物候总体表现为生长季开始和结束时间推迟、生长期缩短,降水对物候的调控效应整体强于气温[18].然而,受空间分辨率和混合像元效应限制,传统卫星遥感难以准确刻画荒漠植被斑块的精细结构特征,尤其是在植被稀疏、斑块尺度较小的极端干旱区,其对群落尺度空间异质性和斑块格局的表征能力仍然有限[21-22].近年来,无人机遥感凭借厘米级空间分辨率、数据获取灵活等优势,在荒漠植被覆盖提取、斑块识别及空间格局量化等方面具有显著优势,为实现样方尺度与区域尺度之间的有效衔接提供了新的技术途径[23-24].而目前针对柴达木盆地自然降水梯度下的群落多样性与斑块格局研究,仍存在空间与尺度覆盖不足的问题.既往研究多局限于局地生境或单一群落类型,缺少覆盖完整降水梯度的代表性样区或样带[13,25-27].在柴达木盆地自然降水梯度背景下,将无人机获取的精细斑块格局指标与地面群落调查及土壤理化属性在相同空间尺度下进行综合分析的研究仍相对缺乏,一定程度上制约了对降水梯度下柴达木盆地荒漠植被与土壤及环境因素耦合机制的认识. ...
... [18].然而,受空间分辨率和混合像元效应限制,传统卫星遥感难以准确刻画荒漠植被斑块的精细结构特征,尤其是在植被稀疏、斑块尺度较小的极端干旱区,其对群落尺度空间异质性和斑块格局的表征能力仍然有限[21-22].近年来,无人机遥感凭借厘米级空间分辨率、数据获取灵活等优势,在荒漠植被覆盖提取、斑块识别及空间格局量化等方面具有显著优势,为实现样方尺度与区域尺度之间的有效衔接提供了新的技术途径[23-24].而目前针对柴达木盆地自然降水梯度下的群落多样性与斑块格局研究,仍存在空间与尺度覆盖不足的问题.既往研究多局限于局地生境或单一群落类型,缺少覆盖完整降水梯度的代表性样区或样带[13,25-27].在柴达木盆地自然降水梯度背景下,将无人机获取的精细斑块格局指标与地面群落调查及土壤理化属性在相同空间尺度下进行综合分析的研究仍相对缺乏,一定程度上制约了对降水梯度下柴达木盆地荒漠植被与土壤及环境因素耦合机制的认识. ...
Distribution of actual evapotranspiration over qaidam basin, an arid area in China
0
2013
柴达木盆地几种荒漠灌丛植被的生物量分配格局
2
2014
... 随着遥感技术的发展,基于中高分辨率卫星影像的植被指数分析已成为研究干旱区植被动态及其对降水响应的重要手段,在区域尺度揭示植被覆盖变化及其时空格局方面具有显著优势[15-16].已有研究表明,柴达木盆地植被绿度总体呈周缘升高、腹地偏低的空间格局,并在近几十年暖湿化背景下对降水变化表现出显著响应[13,17];植被物候、蒸散发及水分利用效率等生态特征亦随降水梯度和环境条件发生变化[18-20],如植被物候总体表现为生长季开始和结束时间推迟、生长期缩短,降水对物候的调控效应整体强于气温[18].然而,受空间分辨率和混合像元效应限制,传统卫星遥感难以准确刻画荒漠植被斑块的精细结构特征,尤其是在植被稀疏、斑块尺度较小的极端干旱区,其对群落尺度空间异质性和斑块格局的表征能力仍然有限[21-22].近年来,无人机遥感凭借厘米级空间分辨率、数据获取灵活等优势,在荒漠植被覆盖提取、斑块识别及空间格局量化等方面具有显著优势,为实现样方尺度与区域尺度之间的有效衔接提供了新的技术途径[23-24].而目前针对柴达木盆地自然降水梯度下的群落多样性与斑块格局研究,仍存在空间与尺度覆盖不足的问题.既往研究多局限于局地生境或单一群落类型,缺少覆盖完整降水梯度的代表性样区或样带[13,25-27].在柴达木盆地自然降水梯度背景下,将无人机获取的精细斑块格局指标与地面群落调查及土壤理化属性在相同空间尺度下进行综合分析的研究仍相对缺乏,一定程度上制约了对降水梯度下柴达木盆地荒漠植被与土壤及环境因素耦合机制的认识. ...
... 在3个降水梯度上,灌木均为群落的优势生活型,但群落结构对降水变化表现出明显的响应,在200~300 mm降水组禾本科与一年生草本的相对多度显著增加,与灌木共同构成群落;而在50~100 mm降水组的极端干旱区间,群落则几乎完全由梭梭、白刺、驼绒藜等耐旱灌木所主导(表1,图2和图3).这一变化趋势与低海拔干旱区的研究结果总体一致,如在宁夏荒漠草原的降水控制试验表明,随着降水减少,草本植物地上生物量显著下降,群落结构趋于简单,耐旱物种的重要性上升[32].相关研究进一步表明,降水变化主要通过直接影响豆科和禾本科植物地上生物量进而间接影响植物群落地上生物量[8].上述群落结构变化与荒漠植物的生存策略相一致,草本植物对水分变化更为敏感,在极度干旱条件下仅有少数耐旱性高的灌木能够维持稳定种群[20,33].植物多样性随降水量的减少显著降低.200~300 mm降水组的Shannon-Wiener指数和物种丰富度明显高于其余降水组,但群落均匀度指数组间差异不显著,这可能是由于沿降水梯度的多样性下降主要源于物种丰富度的减少,而非优势度结构的显著改变[34]. ...
Spatial variation in land cover and choice of spatial resolution for remote sensing
1
2004
... 随着遥感技术的发展,基于中高分辨率卫星影像的植被指数分析已成为研究干旱区植被动态及其对降水响应的重要手段,在区域尺度揭示植被覆盖变化及其时空格局方面具有显著优势[15-16].已有研究表明,柴达木盆地植被绿度总体呈周缘升高、腹地偏低的空间格局,并在近几十年暖湿化背景下对降水变化表现出显著响应[13,17];植被物候、蒸散发及水分利用效率等生态特征亦随降水梯度和环境条件发生变化[18-20],如植被物候总体表现为生长季开始和结束时间推迟、生长期缩短,降水对物候的调控效应整体强于气温[18].然而,受空间分辨率和混合像元效应限制,传统卫星遥感难以准确刻画荒漠植被斑块的精细结构特征,尤其是在植被稀疏、斑块尺度较小的极端干旱区,其对群落尺度空间异质性和斑块格局的表征能力仍然有限[21-22].近年来,无人机遥感凭借厘米级空间分辨率、数据获取灵活等优势,在荒漠植被覆盖提取、斑块识别及空间格局量化等方面具有显著优势,为实现样方尺度与区域尺度之间的有效衔接提供了新的技术途径[23-24].而目前针对柴达木盆地自然降水梯度下的群落多样性与斑块格局研究,仍存在空间与尺度覆盖不足的问题.既往研究多局限于局地生境或单一群落类型,缺少覆盖完整降水梯度的代表性样区或样带[13,25-27].在柴达木盆地自然降水梯度背景下,将无人机获取的精细斑块格局指标与地面群落调查及土壤理化属性在相同空间尺度下进行综合分析的研究仍相对缺乏,一定程度上制约了对降水梯度下柴达木盆地荒漠植被与土壤及环境因素耦合机制的认识. ...
Remote sensing for biodiversity science and conservation
1
2003
... 随着遥感技术的发展,基于中高分辨率卫星影像的植被指数分析已成为研究干旱区植被动态及其对降水响应的重要手段,在区域尺度揭示植被覆盖变化及其时空格局方面具有显著优势[15-16].已有研究表明,柴达木盆地植被绿度总体呈周缘升高、腹地偏低的空间格局,并在近几十年暖湿化背景下对降水变化表现出显著响应[13,17];植被物候、蒸散发及水分利用效率等生态特征亦随降水梯度和环境条件发生变化[18-20],如植被物候总体表现为生长季开始和结束时间推迟、生长期缩短,降水对物候的调控效应整体强于气温[18].然而,受空间分辨率和混合像元效应限制,传统卫星遥感难以准确刻画荒漠植被斑块的精细结构特征,尤其是在植被稀疏、斑块尺度较小的极端干旱区,其对群落尺度空间异质性和斑块格局的表征能力仍然有限[21-22].近年来,无人机遥感凭借厘米级空间分辨率、数据获取灵活等优势,在荒漠植被覆盖提取、斑块识别及空间格局量化等方面具有显著优势,为实现样方尺度与区域尺度之间的有效衔接提供了新的技术途径[23-24].而目前针对柴达木盆地自然降水梯度下的群落多样性与斑块格局研究,仍存在空间与尺度覆盖不足的问题.既往研究多局限于局地生境或单一群落类型,缺少覆盖完整降水梯度的代表性样区或样带[13,25-27].在柴达木盆地自然降水梯度背景下,将无人机获取的精细斑块格局指标与地面群落调查及土壤理化属性在相同空间尺度下进行综合分析的研究仍相对缺乏,一定程度上制约了对降水梯度下柴达木盆地荒漠植被与土壤及环境因素耦合机制的认识. ...
Lightweight unmanned aerial vehicles will revolutionize spatial ecology
1
2013
... 随着遥感技术的发展,基于中高分辨率卫星影像的植被指数分析已成为研究干旱区植被动态及其对降水响应的重要手段,在区域尺度揭示植被覆盖变化及其时空格局方面具有显著优势[15-16].已有研究表明,柴达木盆地植被绿度总体呈周缘升高、腹地偏低的空间格局,并在近几十年暖湿化背景下对降水变化表现出显著响应[13,17];植被物候、蒸散发及水分利用效率等生态特征亦随降水梯度和环境条件发生变化[18-20],如植被物候总体表现为生长季开始和结束时间推迟、生长期缩短,降水对物候的调控效应整体强于气温[18].然而,受空间分辨率和混合像元效应限制,传统卫星遥感难以准确刻画荒漠植被斑块的精细结构特征,尤其是在植被稀疏、斑块尺度较小的极端干旱区,其对群落尺度空间异质性和斑块格局的表征能力仍然有限[21-22].近年来,无人机遥感凭借厘米级空间分辨率、数据获取灵活等优势,在荒漠植被覆盖提取、斑块识别及空间格局量化等方面具有显著优势,为实现样方尺度与区域尺度之间的有效衔接提供了新的技术途径[23-24].而目前针对柴达木盆地自然降水梯度下的群落多样性与斑块格局研究,仍存在空间与尺度覆盖不足的问题.既往研究多局限于局地生境或单一群落类型,缺少覆盖完整降水梯度的代表性样区或样带[13,25-27].在柴达木盆地自然降水梯度背景下,将无人机获取的精细斑块格局指标与地面群落调查及土壤理化属性在相同空间尺度下进行综合分析的研究仍相对缺乏,一定程度上制约了对降水梯度下柴达木盆地荒漠植被与土壤及环境因素耦合机制的认识. ...
High spatial resolution three-dimensional mapping of vegetation spectral dynamics using computer vision
1
2013
... 随着遥感技术的发展,基于中高分辨率卫星影像的植被指数分析已成为研究干旱区植被动态及其对降水响应的重要手段,在区域尺度揭示植被覆盖变化及其时空格局方面具有显著优势[15-16].已有研究表明,柴达木盆地植被绿度总体呈周缘升高、腹地偏低的空间格局,并在近几十年暖湿化背景下对降水变化表现出显著响应[13,17];植被物候、蒸散发及水分利用效率等生态特征亦随降水梯度和环境条件发生变化[18-20],如植被物候总体表现为生长季开始和结束时间推迟、生长期缩短,降水对物候的调控效应整体强于气温[18].然而,受空间分辨率和混合像元效应限制,传统卫星遥感难以准确刻画荒漠植被斑块的精细结构特征,尤其是在植被稀疏、斑块尺度较小的极端干旱区,其对群落尺度空间异质性和斑块格局的表征能力仍然有限[21-22].近年来,无人机遥感凭借厘米级空间分辨率、数据获取灵活等优势,在荒漠植被覆盖提取、斑块识别及空间格局量化等方面具有显著优势,为实现样方尺度与区域尺度之间的有效衔接提供了新的技术途径[23-24].而目前针对柴达木盆地自然降水梯度下的群落多样性与斑块格局研究,仍存在空间与尺度覆盖不足的问题.既往研究多局限于局地生境或单一群落类型,缺少覆盖完整降水梯度的代表性样区或样带[13,25-27].在柴达木盆地自然降水梯度背景下,将无人机获取的精细斑块格局指标与地面群落调查及土壤理化属性在相同空间尺度下进行综合分析的研究仍相对缺乏,一定程度上制约了对降水梯度下柴达木盆地荒漠植被与土壤及环境因素耦合机制的认识. ...
柴达木盆地植物群落样方数据集
1
2024
... 随着遥感技术的发展,基于中高分辨率卫星影像的植被指数分析已成为研究干旱区植被动态及其对降水响应的重要手段,在区域尺度揭示植被覆盖变化及其时空格局方面具有显著优势[15-16].已有研究表明,柴达木盆地植被绿度总体呈周缘升高、腹地偏低的空间格局,并在近几十年暖湿化背景下对降水变化表现出显著响应[13,17];植被物候、蒸散发及水分利用效率等生态特征亦随降水梯度和环境条件发生变化[18-20],如植被物候总体表现为生长季开始和结束时间推迟、生长期缩短,降水对物候的调控效应整体强于气温[18].然而,受空间分辨率和混合像元效应限制,传统卫星遥感难以准确刻画荒漠植被斑块的精细结构特征,尤其是在植被稀疏、斑块尺度较小的极端干旱区,其对群落尺度空间异质性和斑块格局的表征能力仍然有限[21-22].近年来,无人机遥感凭借厘米级空间分辨率、数据获取灵活等优势,在荒漠植被覆盖提取、斑块识别及空间格局量化等方面具有显著优势,为实现样方尺度与区域尺度之间的有效衔接提供了新的技术途径[23-24].而目前针对柴达木盆地自然降水梯度下的群落多样性与斑块格局研究,仍存在空间与尺度覆盖不足的问题.既往研究多局限于局地生境或单一群落类型,缺少覆盖完整降水梯度的代表性样区或样带[13,25-27].在柴达木盆地自然降水梯度背景下,将无人机获取的精细斑块格局指标与地面群落调查及土壤理化属性在相同空间尺度下进行综合分析的研究仍相对缺乏,一定程度上制约了对降水梯度下柴达木盆地荒漠植被与土壤及环境因素耦合机制的认识. ...
柴达木盆地荒漠灌木水分利用效率及其影响因素
1
2025
... 随着遥感技术的发展,基于中高分辨率卫星影像的植被指数分析已成为研究干旱区植被动态及其对降水响应的重要手段,在区域尺度揭示植被覆盖变化及其时空格局方面具有显著优势[15-16].已有研究表明,柴达木盆地植被绿度总体呈周缘升高、腹地偏低的空间格局,并在近几十年暖湿化背景下对降水变化表现出显著响应[13,17];植被物候、蒸散发及水分利用效率等生态特征亦随降水梯度和环境条件发生变化[18-20],如植被物候总体表现为生长季开始和结束时间推迟、生长期缩短,降水对物候的调控效应整体强于气温[18].然而,受空间分辨率和混合像元效应限制,传统卫星遥感难以准确刻画荒漠植被斑块的精细结构特征,尤其是在植被稀疏、斑块尺度较小的极端干旱区,其对群落尺度空间异质性和斑块格局的表征能力仍然有限[21-22].近年来,无人机遥感凭借厘米级空间分辨率、数据获取灵活等优势,在荒漠植被覆盖提取、斑块识别及空间格局量化等方面具有显著优势,为实现样方尺度与区域尺度之间的有效衔接提供了新的技术途径[23-24].而目前针对柴达木盆地自然降水梯度下的群落多样性与斑块格局研究,仍存在空间与尺度覆盖不足的问题.既往研究多局限于局地生境或单一群落类型,缺少覆盖完整降水梯度的代表性样区或样带[13,25-27].在柴达木盆地自然降水梯度背景下,将无人机获取的精细斑块格局指标与地面群落调查及土壤理化属性在相同空间尺度下进行综合分析的研究仍相对缺乏,一定程度上制约了对降水梯度下柴达木盆地荒漠植被与土壤及环境因素耦合机制的认识. ...
Total nitrogen analysis of soil and plant tissues
1
1980
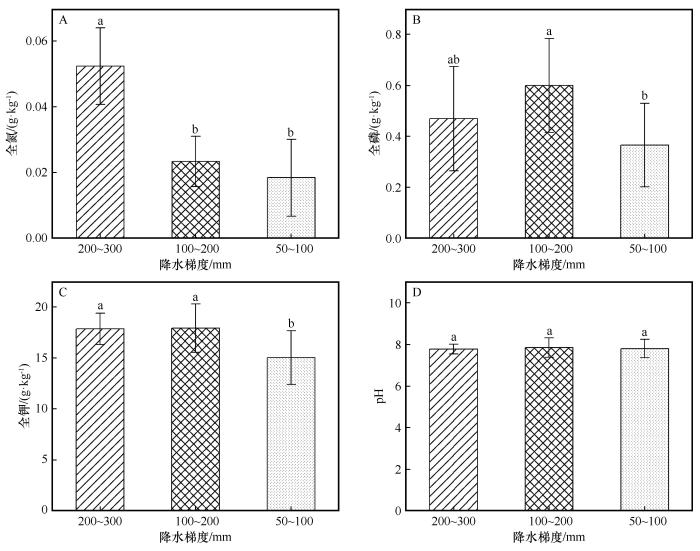
... 土壤样品与理化性质:采用五点取样法在0~20 cm土层采集混合土样.土样经风干、研磨并过2 mm筛后,分别采用凯氏定氮法测定全氮[28],酸溶-钼锑抗比色法测定全磷,酸溶-火焰光度法测定全钾,并使用玻璃电极法(1∶2.5土水比)测定pH值[29]. ...
Part 3-chemical methods
1
1982
... 土壤样品与理化性质:采用五点取样法在0~20 cm土层采集混合土样.土样经风干、研磨并过2 mm筛后,分别采用凯氏定氮法测定全氮[28],酸溶-钼锑抗比色法测定全磷,酸溶-火焰光度法测定全钾,并使用玻璃电极法(1∶2.5土水比)测定pH值[29]. ...
青藏高原气候数据(1901-2022)
1
2024
... 环境因子:气候数据,包括年降水量和年均气温,来源于国家青藏高原科学数据中心[30].地下水埋深来源于Wang等[31]的研究,并通过ArcGIS 10.2计算样地距河道的直线距离. ...
Underground well water level observation grid dataset from 2005 to 2022
1
2025
... 环境因子:气候数据,包括年降水量和年均气温,来源于国家青藏高原科学数据中心[30].地下水埋深来源于Wang等[31]的研究,并通过ArcGIS 10.2计算样地距河道的直线距离. ...
Precipitation changes alter the structure, species composition and interspecific relationships of desert steppe plant communities
2
2025
... 景观格局指标:使用大疆智图对无人机影像进行前期处理,获得研究区正射影像图.首先通过目视解释对植被和裸地进行二元划分.将结果导入R语言[32]并转为分类栅格,使用terra包进行栅格读取、类型转换,按固定编码设置植被像元值为0、非植被为1.并以像元计数法计算植被盖度(%).借助landscapemetrics包提取景观指数,包括斑块数量(NP)、最大斑块指数(LPI)和聚合度指数(AI). ...
... 在3个降水梯度上,灌木均为群落的优势生活型,但群落结构对降水变化表现出明显的响应,在200~300 mm降水组禾本科与一年生草本的相对多度显著增加,与灌木共同构成群落;而在50~100 mm降水组的极端干旱区间,群落则几乎完全由梭梭、白刺、驼绒藜等耐旱灌木所主导(表1,图2和图3).这一变化趋势与低海拔干旱区的研究结果总体一致,如在宁夏荒漠草原的降水控制试验表明,随着降水减少,草本植物地上生物量显著下降,群落结构趋于简单,耐旱物种的重要性上升[32].相关研究进一步表明,降水变化主要通过直接影响豆科和禾本科植物地上生物量进而间接影响植物群落地上生物量[8].上述群落结构变化与荒漠植物的生存策略相一致,草本植物对水分变化更为敏感,在极度干旱条件下仅有少数耐旱性高的灌木能够维持稳定种群[20,33].植物多样性随降水量的减少显著降低.200~300 mm降水组的Shannon-Wiener指数和物种丰富度明显高于其余降水组,但群落均匀度指数组间差异不显著,这可能是由于沿降水梯度的多样性下降主要源于物种丰富度的减少,而非优势度结构的显著改变[34]. ...
Soil moisture and soluble salt content dominate changes in foliar δ 13C and δ 15N of desert communities in the Qaidam Basin,Qinghai-Tibetan Plateau
1
2021
... 在3个降水梯度上,灌木均为群落的优势生活型,但群落结构对降水变化表现出明显的响应,在200~300 mm降水组禾本科与一年生草本的相对多度显著增加,与灌木共同构成群落;而在50~100 mm降水组的极端干旱区间,群落则几乎完全由梭梭、白刺、驼绒藜等耐旱灌木所主导(表1,图2和图3).这一变化趋势与低海拔干旱区的研究结果总体一致,如在宁夏荒漠草原的降水控制试验表明,随着降水减少,草本植物地上生物量显著下降,群落结构趋于简单,耐旱物种的重要性上升[32].相关研究进一步表明,降水变化主要通过直接影响豆科和禾本科植物地上生物量进而间接影响植物群落地上生物量[8].上述群落结构变化与荒漠植物的生存策略相一致,草本植物对水分变化更为敏感,在极度干旱条件下仅有少数耐旱性高的灌木能够维持稳定种群[20,33].植物多样性随降水量的减少显著降低.200~300 mm降水组的Shannon-Wiener指数和物种丰富度明显高于其余降水组,但群落均匀度指数组间差异不显著,这可能是由于沿降水梯度的多样性下降主要源于物种丰富度的减少,而非优势度结构的显著改变[34]. ...
Precipitation and temperature regulate species diversity, plant coverage and aboveground biomass through opposing mechanisms in large-scale grasslands
1
2022
... 在3个降水梯度上,灌木均为群落的优势生活型,但群落结构对降水变化表现出明显的响应,在200~300 mm降水组禾本科与一年生草本的相对多度显著增加,与灌木共同构成群落;而在50~100 mm降水组的极端干旱区间,群落则几乎完全由梭梭、白刺、驼绒藜等耐旱灌木所主导(表1,图2和图3).这一变化趋势与低海拔干旱区的研究结果总体一致,如在宁夏荒漠草原的降水控制试验表明,随着降水减少,草本植物地上生物量显著下降,群落结构趋于简单,耐旱物种的重要性上升[32].相关研究进一步表明,降水变化主要通过直接影响豆科和禾本科植物地上生物量进而间接影响植物群落地上生物量[8].上述群落结构变化与荒漠植物的生存策略相一致,草本植物对水分变化更为敏感,在极度干旱条件下仅有少数耐旱性高的灌木能够维持稳定种群[20,33].植物多样性随降水量的减少显著降低.200~300 mm降水组的Shannon-Wiener指数和物种丰富度明显高于其余降水组,但群落均匀度指数组间差异不显著,这可能是由于沿降水梯度的多样性下降主要源于物种丰富度的减少,而非优势度结构的显著改变[34]. ...
草原灌丛化对生态系统多功能性的影响
1
2023
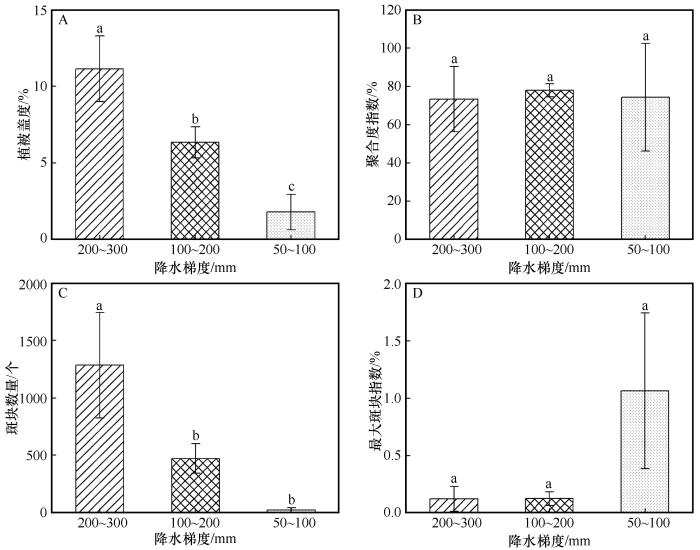
... 基于无人机影像的斑块指数分析表明,高降水区植被斑块面积较大、聚集度高,而低降水区植被斑块数量和聚集度降低,整体呈现出由连片化向破碎化转变的空间格局.这一特征符合荒漠区肥力岛效应,即在水分和养分双重限制下,植被倾向于在微地形和局地有利生境中形成聚集斑块,以集中利用有限资源;随着降水条件改善,这些斑块得以扩展并相互连接,从而提高植被盖度并维持较高的空间聚集性[35].在50~100 mm降水组观测到盖度高变异性,反映了极端干旱条件下水分供给在空间上的高度不均一性.在该降水水平下,降水多以小尺度、脉冲式事件出现,通常仅能湿润表层土壤,且入渗深度受土壤质地限制[36],导致不同样地之间实际可利用水分差异显著.在此基础上,微地形对水分再分配的影响被显著放大,局地汇水条件较好的样地能够维持相对较高的植被盖度,而水分条件不好的样地则表现为极低盖度,从而造成样地间盖度差异的显著扩大.如已有研究表明,荒漠植被高覆盖区域往往沿局地低洼沟道或短暂性汇水区分布[37]. ...
The response of soil moisture to rainfall event size in subalpine grassland and meadows in a semi-arid mountain range:a case study in Northwestern China's Qilian Mountains
1
2012
... 基于无人机影像的斑块指数分析表明,高降水区植被斑块面积较大、聚集度高,而低降水区植被斑块数量和聚集度降低,整体呈现出由连片化向破碎化转变的空间格局.这一特征符合荒漠区肥力岛效应,即在水分和养分双重限制下,植被倾向于在微地形和局地有利生境中形成聚集斑块,以集中利用有限资源;随着降水条件改善,这些斑块得以扩展并相互连接,从而提高植被盖度并维持较高的空间聚集性[35].在50~100 mm降水组观测到盖度高变异性,反映了极端干旱条件下水分供给在空间上的高度不均一性.在该降水水平下,降水多以小尺度、脉冲式事件出现,通常仅能湿润表层土壤,且入渗深度受土壤质地限制[36],导致不同样地之间实际可利用水分差异显著.在此基础上,微地形对水分再分配的影响被显著放大,局地汇水条件较好的样地能够维持相对较高的植被盖度,而水分条件不好的样地则表现为极低盖度,从而造成样地间盖度差异的显著扩大.如已有研究表明,荒漠植被高覆盖区域往往沿局地低洼沟道或短暂性汇水区分布[37]. ...
Dryland ecohydrology and climate change:critical issues and technical advances
1
2012
... 基于无人机影像的斑块指数分析表明,高降水区植被斑块面积较大、聚集度高,而低降水区植被斑块数量和聚集度降低,整体呈现出由连片化向破碎化转变的空间格局.这一特征符合荒漠区肥力岛效应,即在水分和养分双重限制下,植被倾向于在微地形和局地有利生境中形成聚集斑块,以集中利用有限资源;随着降水条件改善,这些斑块得以扩展并相互连接,从而提高植被盖度并维持较高的空间聚集性[35].在50~100 mm降水组观测到盖度高变异性,反映了极端干旱条件下水分供给在空间上的高度不均一性.在该降水水平下,降水多以小尺度、脉冲式事件出现,通常仅能湿润表层土壤,且入渗深度受土壤质地限制[36],导致不同样地之间实际可利用水分差异显著.在此基础上,微地形对水分再分配的影响被显著放大,局地汇水条件较好的样地能够维持相对较高的植被盖度,而水分条件不好的样地则表现为极低盖度,从而造成样地间盖度差异的显著扩大.如已有研究表明,荒漠植被高覆盖区域往往沿局地低洼沟道或短暂性汇水区分布[37]. ...
不同降水对荒漠灌丛土壤理化性质和地表植被分布的影响
1
2021
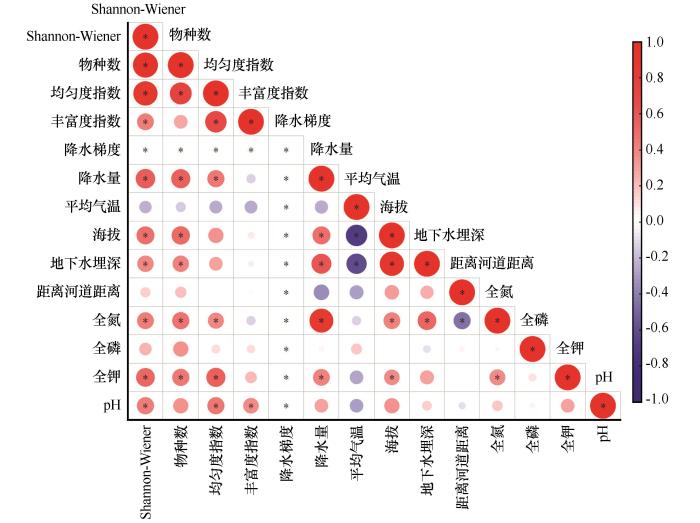
... Mantel分析表明,植被指标与土壤理化性质在整体层面相关性不显著(表4).这一结果表明,研究区所处的极端干旱背景下,土壤养分条件并非主导植被空间分布与群落结构变化的关键限制因子.研究区土壤养分含量整体处于较低水平,且不同样地间变异幅度相对有限,这在一定程度上削弱了土壤理化性质对植被格局的区分能力.与本研究结果不同,在年降水量为180、280、410 mm的较高降水梯度区研究表明,土壤理化性质与植被物种数、高度等指标呈现一致变化趋势[38].这种差异可能是由于在水分相对充足的条件下,土壤养分更容易转化为限制植被生长和群落结构的重要因子;而在本研究区的极端干旱环境中,水分胁迫强度远高于养分限制,从而掩盖了土壤理化性质对植被的潜在影响. ...
柴达木盆地生态植被的地下水阈值
2
2019
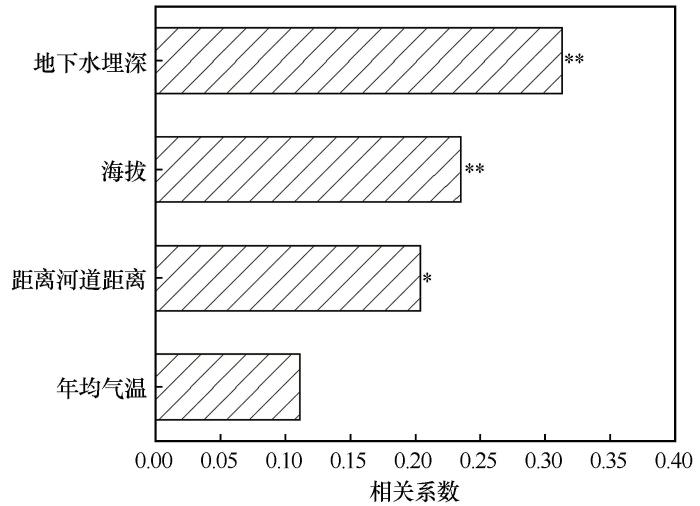
... 在各环境因子中,地下水埋深成为影响植被分布的最重要环境因子(图7).由于降水具有明显的脉冲性和年际不稳定性,地下水为浅埋区荒漠植被提供了相对稳定的水分来源.如在柴达木盆地区域尺度上的研究表明,当地下水埋深较浅、距河道较近时,NDVI明显增高;反之则迅速衰减[39-40].此外,地下水阈值研究进一步表明,柴达木盆地不同植被类型对地下水埋深具有特定的阈值范围,一旦超过这个阈值,灌丛与草本植物难以维持稳定群落结构[39].因此,在柴达木盆地这一极端干旱高原荒漠环境中,植被分布与群落结构主要受水分条件控制,其中地下水在缓冲降水变异方面发挥了关键作用,而土壤养分条件的影响则在水分限制下相对弱化. ...
... [39].因此,在柴达木盆地这一极端干旱高原荒漠环境中,植被分布与群落结构主要受水分条件控制,其中地下水在缓冲降水变异方面发挥了关键作用,而土壤养分条件的影响则在水分限制下相对弱化. ...
Vegetation response to groundwater variation in arid environments:visualization of research evolution,synthesis of response types,and estimation of groundwater threshold
1
2019
... 在各环境因子中,地下水埋深成为影响植被分布的最重要环境因子(图7).由于降水具有明显的脉冲性和年际不稳定性,地下水为浅埋区荒漠植被提供了相对稳定的水分来源.如在柴达木盆地区域尺度上的研究表明,当地下水埋深较浅、距河道较近时,NDVI明显增高;反之则迅速衰减[39-40].此外,地下水阈值研究进一步表明,柴达木盆地不同植被类型对地下水埋深具有特定的阈值范围,一旦超过这个阈值,灌丛与草本植物难以维持稳定群落结构[39].因此,在柴达木盆地这一极端干旱高原荒漠环境中,植被分布与群落结构主要受水分条件控制,其中地下水在缓冲降水变异方面发挥了关键作用,而土壤养分条件的影响则在水分限制下相对弱化. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}