施氮对饲用高粱/拉巴豆混播草地生产性能和氮肥贡献率的影响
1
2025
... 土壤氮循环是陆地生态系统养分循环和维持农业生产力的核心环节,其中无机氮作为作物可直接吸收利用的氮形态,是氮循环过程中最为活跃的组成部分.铵态氮(NH-N)和硝态氮(NO-N)含量及其转化速率决定了土壤氮素的有效性、氮肥利用效率以及氮素向大气和地下水的迁移风险[1].在农田生态系统中,NH-N是硝化作用的主要底物,而NO-N则是植物吸收和淋失的主要形态,两者的动态平衡直接反映了土壤氮素转化过程的方向与强度[2-3].硝化作用作为氮素矿化与气态氮(NO,N2O和N2)损失之间的关键过程,连接了无机氮的不同形态转换,对农业氮循环具有重要调控作用[4].硝化作用是氨(NH3)在氨单加氧酶(amoA)和羟胺氧化还原酶(hao)的催化下产生亚硝酸盐(NO),并进一步在亚硝酸氧化菌(NOB)作用下氧化为硝酸盐(NO)的过程[5].由氨氧化细菌(AOB)和氨氧化古菌(AOA)驱动的氨氧化过程(NH3—NO),是硝化过程的第一个限速步骤,也是农田系统氮素循环关注的焦点[6-7].在这一过程中,无机氮的含量与形态不仅影响氨氧化底物的供应,还通过调节微生物活性和群落结构影响整个硝化潜势(Potential nitrification rate,PNR)与氨氧化潜势(Potential ammonia oxidation,PAO)[8].其中,PNR反映的是土壤在非限制条件下完成NH→NO转化的最大潜在能力,而PAO仅针对氨氧化这一限速环节,其变化能够直接指示氨氧化微生物对硝化过程的调控强度,因此在解释硝化过程的驱动机制时,PNR与PAO具有互补意义[5-6]. ...
荒漠绿洲农田土壤水热动态及硝态氮淋溶特征
1
2022
... 土壤氮循环是陆地生态系统养分循环和维持农业生产力的核心环节,其中无机氮作为作物可直接吸收利用的氮形态,是氮循环过程中最为活跃的组成部分.铵态氮(NH-N)和硝态氮(NO-N)含量及其转化速率决定了土壤氮素的有效性、氮肥利用效率以及氮素向大气和地下水的迁移风险[1].在农田生态系统中,NH-N是硝化作用的主要底物,而NO-N则是植物吸收和淋失的主要形态,两者的动态平衡直接反映了土壤氮素转化过程的方向与强度[2-3].硝化作用作为氮素矿化与气态氮(NO,N2O和N2)损失之间的关键过程,连接了无机氮的不同形态转换,对农业氮循环具有重要调控作用[4].硝化作用是氨(NH3)在氨单加氧酶(amoA)和羟胺氧化还原酶(hao)的催化下产生亚硝酸盐(NO),并进一步在亚硝酸氧化菌(NOB)作用下氧化为硝酸盐(NO)的过程[5].由氨氧化细菌(AOB)和氨氧化古菌(AOA)驱动的氨氧化过程(NH3—NO),是硝化过程的第一个限速步骤,也是农田系统氮素循环关注的焦点[6-7].在这一过程中,无机氮的含量与形态不仅影响氨氧化底物的供应,还通过调节微生物活性和群落结构影响整个硝化潜势(Potential nitrification rate,PNR)与氨氧化潜势(Potential ammonia oxidation,PAO)[8].其中,PNR反映的是土壤在非限制条件下完成NH→NO转化的最大潜在能力,而PAO仅针对氨氧化这一限速环节,其变化能够直接指示氨氧化微生物对硝化过程的调控强度,因此在解释硝化过程的驱动机制时,PNR与PAO具有互补意义[5-6]. ...
Effect of vegetation degradation on Soil nitrogen components and N-cycling enzyme activities in a wet meadow on the Qinghai-Tibetan Plateau
1
2025
... 土壤氮循环是陆地生态系统养分循环和维持农业生产力的核心环节,其中无机氮作为作物可直接吸收利用的氮形态,是氮循环过程中最为活跃的组成部分.铵态氮(NH-N)和硝态氮(NO-N)含量及其转化速率决定了土壤氮素的有效性、氮肥利用效率以及氮素向大气和地下水的迁移风险[1].在农田生态系统中,NH-N是硝化作用的主要底物,而NO-N则是植物吸收和淋失的主要形态,两者的动态平衡直接反映了土壤氮素转化过程的方向与强度[2-3].硝化作用作为氮素矿化与气态氮(NO,N2O和N2)损失之间的关键过程,连接了无机氮的不同形态转换,对农业氮循环具有重要调控作用[4].硝化作用是氨(NH3)在氨单加氧酶(amoA)和羟胺氧化还原酶(hao)的催化下产生亚硝酸盐(NO),并进一步在亚硝酸氧化菌(NOB)作用下氧化为硝酸盐(NO)的过程[5].由氨氧化细菌(AOB)和氨氧化古菌(AOA)驱动的氨氧化过程(NH3—NO),是硝化过程的第一个限速步骤,也是农田系统氮素循环关注的焦点[6-7].在这一过程中,无机氮的含量与形态不仅影响氨氧化底物的供应,还通过调节微生物活性和群落结构影响整个硝化潜势(Potential nitrification rate,PNR)与氨氧化潜势(Potential ammonia oxidation,PAO)[8].其中,PNR反映的是土壤在非限制条件下完成NH→NO转化的最大潜在能力,而PAO仅针对氨氧化这一限速环节,其变化能够直接指示氨氧化微生物对硝化过程的调控强度,因此在解释硝化过程的驱动机制时,PNR与PAO具有互补意义[5-6]. ...
Effect of co-application of straw and various nitrogen fertilizers on N2O emission in acid soil
3
2023
... 土壤氮循环是陆地生态系统养分循环和维持农业生产力的核心环节,其中无机氮作为作物可直接吸收利用的氮形态,是氮循环过程中最为活跃的组成部分.铵态氮(NH-N)和硝态氮(NO-N)含量及其转化速率决定了土壤氮素的有效性、氮肥利用效率以及氮素向大气和地下水的迁移风险[1].在农田生态系统中,NH-N是硝化作用的主要底物,而NO-N则是植物吸收和淋失的主要形态,两者的动态平衡直接反映了土壤氮素转化过程的方向与强度[2-3].硝化作用作为氮素矿化与气态氮(NO,N2O和N2)损失之间的关键过程,连接了无机氮的不同形态转换,对农业氮循环具有重要调控作用[4].硝化作用是氨(NH3)在氨单加氧酶(amoA)和羟胺氧化还原酶(hao)的催化下产生亚硝酸盐(NO),并进一步在亚硝酸氧化菌(NOB)作用下氧化为硝酸盐(NO)的过程[5].由氨氧化细菌(AOB)和氨氧化古菌(AOA)驱动的氨氧化过程(NH3—NO),是硝化过程的第一个限速步骤,也是农田系统氮素循环关注的焦点[6-7].在这一过程中,无机氮的含量与形态不仅影响氨氧化底物的供应,还通过调节微生物活性和群落结构影响整个硝化潜势(Potential nitrification rate,PNR)与氨氧化潜势(Potential ammonia oxidation,PAO)[8].其中,PNR反映的是土壤在非限制条件下完成NH→NO转化的最大潜在能力,而PAO仅针对氨氧化这一限速环节,其变化能够直接指示氨氧化微生物对硝化过程的调控强度,因此在解释硝化过程的驱动机制时,PNR与PAO具有互补意义[5-6]. ...
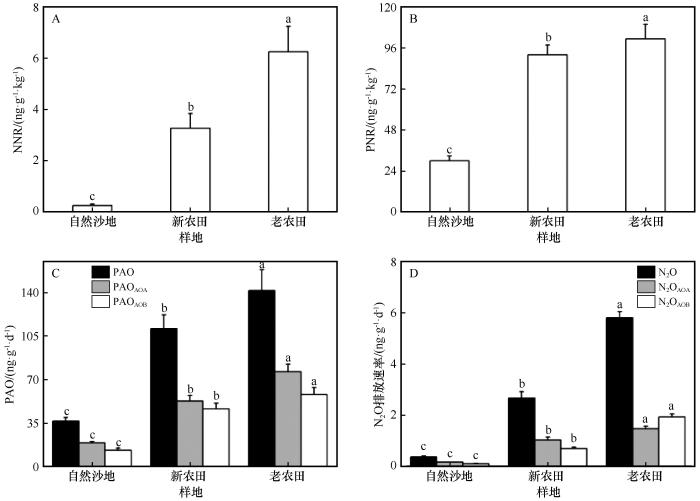
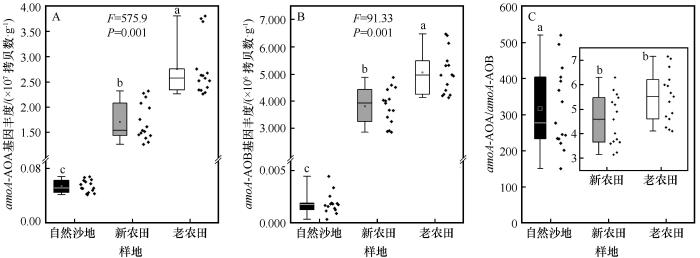
... 值得注意的是,AOA/AOB基因丰度比值随开垦年限呈降低趋势(USL>OOF>YOF).这表明在未开垦土壤中,硝化作用几乎由AOA绝对主导.而随着耕作年限延长和施肥管理的持续,AOB群落得到了更大幅度的富集,其在硝化过程中的相对重要性随之上升,反映了连续耕作过程中氨氧化群落由寡营养型向富营养型的演替趋势[35].这一转变与N2O排放来源的变化相互印证,在USL和YOF中,N2O排放主要由AOA驱动,而在OOF中则转变为AOB驱动为主.尽管AOA基因拷贝数在所有处理中均显著高于AOB,且主导了氨氧化潜势(PAO),但上述结果仍显示,微生物丰度并非决定N2O排放来源的唯一因素.研究表明,AOA多为寡营养型,对低浓度铵具有高亲和力,其代谢途径更倾向于将NH3高效转化为NO,而N2O产率较低[33];相比之下,AOB属于富营养型,在高铵环境中具有更高的最大比活性,且其氨氧化过程易于通过羟胺(NH2OH)不完全氧化等途径产生较多N2O[35].本研究中,长期耕作与高量化肥投入(OOF)塑造了持续的高铵环境,这不仅显著富集了AOB种群,更可能激发了其高代谢活性与强N2O产生能力.因此,在OOF处理中,尽管AOB的绝对基因丰度仍低于AOA,但其单位细胞或单位amoA基因的N2O产率更高,加上高底物浓度下整体硝化通量增大,共同导致AOB对N2O排放的实际贡献占据主导[4-6]. ...
... 值得注意的是,PAOAOA与AOA丰度的载荷系数均高于AOB相关变量,表明AOA在驱动PNR变化中仍占主导作用.这与AOA在干旱、碱性及低氮环境中的生态适应性相吻合[35].同时,AOA的胞外聚合物特征及对氧胁迫的耐受性有助于其在绿洲农田典型的干湿交替、弱碱性环境中占据优势[4].此外,N2O排放对PNR的负效应提示硝化与反硝化途径之间可能存在竞争性碳氮流动,当土壤含水量升高或氧气供应受限时,部分NO可能被还原为N2O而非进一步氧化为NO,从而削弱硝化潜力[39-40].这一现象在周期性灌溉条件下尤为显著,反映出绿洲农田土壤中氧化还原状态的时空变动对氮循环路径分配具有重要调节作用. ...
氮沉降和降水变化对荒漠草原土壤氨氧化细菌群落的影响
2
2024
... 土壤氮循环是陆地生态系统养分循环和维持农业生产力的核心环节,其中无机氮作为作物可直接吸收利用的氮形态,是氮循环过程中最为活跃的组成部分.铵态氮(NH-N)和硝态氮(NO-N)含量及其转化速率决定了土壤氮素的有效性、氮肥利用效率以及氮素向大气和地下水的迁移风险[1].在农田生态系统中,NH-N是硝化作用的主要底物,而NO-N则是植物吸收和淋失的主要形态,两者的动态平衡直接反映了土壤氮素转化过程的方向与强度[2-3].硝化作用作为氮素矿化与气态氮(NO,N2O和N2)损失之间的关键过程,连接了无机氮的不同形态转换,对农业氮循环具有重要调控作用[4].硝化作用是氨(NH3)在氨单加氧酶(amoA)和羟胺氧化还原酶(hao)的催化下产生亚硝酸盐(NO),并进一步在亚硝酸氧化菌(NOB)作用下氧化为硝酸盐(NO)的过程[5].由氨氧化细菌(AOB)和氨氧化古菌(AOA)驱动的氨氧化过程(NH3—NO),是硝化过程的第一个限速步骤,也是农田系统氮素循环关注的焦点[6-7].在这一过程中,无机氮的含量与形态不仅影响氨氧化底物的供应,还通过调节微生物活性和群落结构影响整个硝化潜势(Potential nitrification rate,PNR)与氨氧化潜势(Potential ammonia oxidation,PAO)[8].其中,PNR反映的是土壤在非限制条件下完成NH→NO转化的最大潜在能力,而PAO仅针对氨氧化这一限速环节,其变化能够直接指示氨氧化微生物对硝化过程的调控强度,因此在解释硝化过程的驱动机制时,PNR与PAO具有互补意义[5-6]. ...
... [5-6]. ...
紫云英还田对红壤水稻土氨氧化微生物功能基因丰度的影响
3
2025
... 土壤氮循环是陆地生态系统养分循环和维持农业生产力的核心环节,其中无机氮作为作物可直接吸收利用的氮形态,是氮循环过程中最为活跃的组成部分.铵态氮(NH-N)和硝态氮(NO-N)含量及其转化速率决定了土壤氮素的有效性、氮肥利用效率以及氮素向大气和地下水的迁移风险[1].在农田生态系统中,NH-N是硝化作用的主要底物,而NO-N则是植物吸收和淋失的主要形态,两者的动态平衡直接反映了土壤氮素转化过程的方向与强度[2-3].硝化作用作为氮素矿化与气态氮(NO,N2O和N2)损失之间的关键过程,连接了无机氮的不同形态转换,对农业氮循环具有重要调控作用[4].硝化作用是氨(NH3)在氨单加氧酶(amoA)和羟胺氧化还原酶(hao)的催化下产生亚硝酸盐(NO),并进一步在亚硝酸氧化菌(NOB)作用下氧化为硝酸盐(NO)的过程[5].由氨氧化细菌(AOB)和氨氧化古菌(AOA)驱动的氨氧化过程(NH3—NO),是硝化过程的第一个限速步骤,也是农田系统氮素循环关注的焦点[6-7].在这一过程中,无机氮的含量与形态不仅影响氨氧化底物的供应,还通过调节微生物活性和群落结构影响整个硝化潜势(Potential nitrification rate,PNR)与氨氧化潜势(Potential ammonia oxidation,PAO)[8].其中,PNR反映的是土壤在非限制条件下完成NH→NO转化的最大潜在能力,而PAO仅针对氨氧化这一限速环节,其变化能够直接指示氨氧化微生物对硝化过程的调控强度,因此在解释硝化过程的驱动机制时,PNR与PAO具有互补意义[5-6]. ...
... -6]. ...
... 值得注意的是,AOA/AOB基因丰度比值随开垦年限呈降低趋势(USL>OOF>YOF).这表明在未开垦土壤中,硝化作用几乎由AOA绝对主导.而随着耕作年限延长和施肥管理的持续,AOB群落得到了更大幅度的富集,其在硝化过程中的相对重要性随之上升,反映了连续耕作过程中氨氧化群落由寡营养型向富营养型的演替趋势[35].这一转变与N2O排放来源的变化相互印证,在USL和YOF中,N2O排放主要由AOA驱动,而在OOF中则转变为AOB驱动为主.尽管AOA基因拷贝数在所有处理中均显著高于AOB,且主导了氨氧化潜势(PAO),但上述结果仍显示,微生物丰度并非决定N2O排放来源的唯一因素.研究表明,AOA多为寡营养型,对低浓度铵具有高亲和力,其代谢途径更倾向于将NH3高效转化为NO,而N2O产率较低[33];相比之下,AOB属于富营养型,在高铵环境中具有更高的最大比活性,且其氨氧化过程易于通过羟胺(NH2OH)不完全氧化等途径产生较多N2O[35].本研究中,长期耕作与高量化肥投入(OOF)塑造了持续的高铵环境,这不仅显著富集了AOB种群,更可能激发了其高代谢活性与强N2O产生能力.因此,在OOF处理中,尽管AOB的绝对基因丰度仍低于AOA,但其单位细胞或单位amoA基因的N2O产率更高,加上高底物浓度下整体硝化通量增大,共同导致AOB对N2O排放的实际贡献占据主导[4-6]. ...
Cultivation increased soil potential denitrification rates by modifying denitrifier communities in desert-oasis ecotone
2
2023
... 土壤氮循环是陆地生态系统养分循环和维持农业生产力的核心环节,其中无机氮作为作物可直接吸收利用的氮形态,是氮循环过程中最为活跃的组成部分.铵态氮(NH-N)和硝态氮(NO-N)含量及其转化速率决定了土壤氮素的有效性、氮肥利用效率以及氮素向大气和地下水的迁移风险[1].在农田生态系统中,NH-N是硝化作用的主要底物,而NO-N则是植物吸收和淋失的主要形态,两者的动态平衡直接反映了土壤氮素转化过程的方向与强度[2-3].硝化作用作为氮素矿化与气态氮(NO,N2O和N2)损失之间的关键过程,连接了无机氮的不同形态转换,对农业氮循环具有重要调控作用[4].硝化作用是氨(NH3)在氨单加氧酶(amoA)和羟胺氧化还原酶(hao)的催化下产生亚硝酸盐(NO),并进一步在亚硝酸氧化菌(NOB)作用下氧化为硝酸盐(NO)的过程[5].由氨氧化细菌(AOB)和氨氧化古菌(AOA)驱动的氨氧化过程(NH3—NO),是硝化过程的第一个限速步骤,也是农田系统氮素循环关注的焦点[6-7].在这一过程中,无机氮的含量与形态不仅影响氨氧化底物的供应,还通过调节微生物活性和群落结构影响整个硝化潜势(Potential nitrification rate,PNR)与氨氧化潜势(Potential ammonia oxidation,PAO)[8].其中,PNR反映的是土壤在非限制条件下完成NH→NO转化的最大潜在能力,而PAO仅针对氨氧化这一限速环节,其变化能够直接指示氨氧化微生物对硝化过程的调控强度,因此在解释硝化过程的驱动机制时,PNR与PAO具有互补意义[5-6]. ...
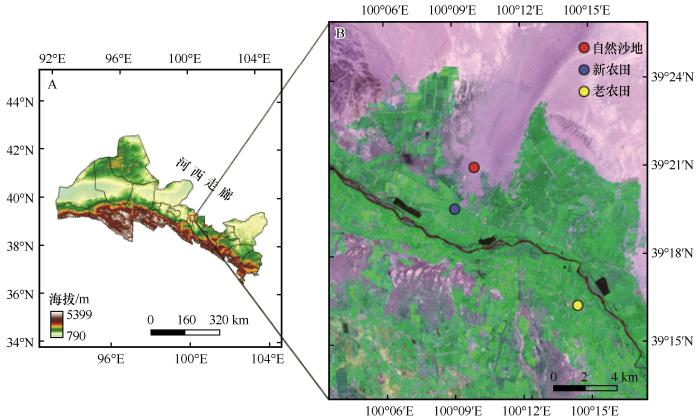
... 研究区位于甘肃河西走廊中段黑河流域的临泽县荒漠-绿洲过渡带,海拔1 350~1 400 m.该地区属典型的温带大陆性气候,年平均气温约7.6 ℃,极端最低和最高气温分别为-27 ℃和39 ℃.多年(1965—2020年)平均降水量117 mm,多年平均蒸散量高达2 390 mm,干旱特征明显.区域土壤母质为第四纪洪积-冲积物,主要土壤类型包括绿洲潮土、灌淤土、灌耕风沙土和灌耕灰棕漠土[17].由于质地偏砂,土壤普遍存在有机质偏低及保水保肥能力弱的特点.自20世纪60年代以来,为满足粮食生产需求,大量沙地被持续开垦为农田,目前近半数耕地属于新垦沙地农田,由于开垦年限不同,土壤熟化程度各异[7].该区农业以一年一熟的制种玉米种植为主(占耕地面积75%以上),普遍采用覆膜栽培和大水漫灌的传统耕作模式.由于土壤先天养分贫瘠与持水能力差,玉米生产高度依赖灌溉与化肥投入,玉米生育期内年均氮、磷、钾肥施用量分别约为200~400 kg·hm-2(以N计)、90~150 kg·hm-2(以P2O5计)和60~90 kg·hm-2(以K2O计),灌溉量在500~1 200 mm,灌溉水源主要来自黑河与地下水[15]. ...
天目山毛竹扩张对阔叶林土壤硝化过程和矿质氮含量的影响
3
2025
... 土壤氮循环是陆地生态系统养分循环和维持农业生产力的核心环节,其中无机氮作为作物可直接吸收利用的氮形态,是氮循环过程中最为活跃的组成部分.铵态氮(NH-N)和硝态氮(NO-N)含量及其转化速率决定了土壤氮素的有效性、氮肥利用效率以及氮素向大气和地下水的迁移风险[1].在农田生态系统中,NH-N是硝化作用的主要底物,而NO-N则是植物吸收和淋失的主要形态,两者的动态平衡直接反映了土壤氮素转化过程的方向与强度[2-3].硝化作用作为氮素矿化与气态氮(NO,N2O和N2)损失之间的关键过程,连接了无机氮的不同形态转换,对农业氮循环具有重要调控作用[4].硝化作用是氨(NH3)在氨单加氧酶(amoA)和羟胺氧化还原酶(hao)的催化下产生亚硝酸盐(NO),并进一步在亚硝酸氧化菌(NOB)作用下氧化为硝酸盐(NO)的过程[5].由氨氧化细菌(AOB)和氨氧化古菌(AOA)驱动的氨氧化过程(NH3—NO),是硝化过程的第一个限速步骤,也是农田系统氮素循环关注的焦点[6-7].在这一过程中,无机氮的含量与形态不仅影响氨氧化底物的供应,还通过调节微生物活性和群落结构影响整个硝化潜势(Potential nitrification rate,PNR)与氨氧化潜势(Potential ammonia oxidation,PAO)[8].其中,PNR反映的是土壤在非限制条件下完成NH→NO转化的最大潜在能力,而PAO仅针对氨氧化这一限速环节,其变化能够直接指示氨氧化微生物对硝化过程的调控强度,因此在解释硝化过程的驱动机制时,PNR与PAO具有互补意义[5-6]. ...
... 土壤潜在硝化速率(PNR)采用15N同位素标记方法测定,具体测定方法和步骤请参照王丽莎等[9].土壤净硝化速率(Net nitrification rate,NNR)采用室内培养法测定[8],称取10 g鲜土(以干重计)置于250 mL广口玻璃瓶中(同时,为测定培养期间土壤矿质氮含量的变化,称取1 kg(以干重计)土壤于2 L培养瓶中),用去离子水调节田间持水量(WHC)至50%.各处理设置3个重复,通过覆盖封口膜并在膜上刺孔,能够保持土壤微生物适宜的通气条件,既保证氨氧化和硝化微生物活性,又避免因缺氧而显著增强反硝化产生的N2O,从而使测定结果更准确反映PNR.随后将样品置于25 ℃黑暗条件下预培养3 d,以消除干湿效应并活化微生物.预培养结束后,在25 ℃条件下好氧培养28 d.培养期间,每隔2~3 d打开膜透气约30 min,并通过称重补水的方法维持WHC稳定在50%.于正式培养1、5、12、20、28 d采集烧杯中的土壤,用于测定土壤NH-N,NO-N和NO-N含量.土壤NH-N,NO-N和NO-N排放强度采用时间加权法. ...
... 添加乙炔(acetylene)和辛炔(1-octyne)可抑制AOB和AOA的活性[8].实验设置3个处理:①样本+对照;②样本+乙炔;③样本+辛炔.具体步骤如下:称取相当于10 g干土重的新鲜土壤于120 mL血清瓶中,调整WFPS至55%,密封后置于30 ℃的黑暗环境下预培养5 d.预培养结束后,向瓶内加入20 mL 1 mmol·L-1 NH4Cl 溶液,200 r·min-1震荡1 h以确保充分混合.随后使用注射器快速将1 mL乙炔或3 mL辛炔分别注入处理①和处理③中,注入前需抽出等体积空气以维持瓶内压力恒定,对照组不做处理.所有瓶中均补加15 mg·mL-1 KClO3溶液,以阻断NO向NO的进一步氧化.处理完成后,在25 ℃、200 r·min-1条件下摇床培养24 h.于培养的1 h和24 h时间点,采集1 mL土壤悬液用于测定土壤NO含量;同步采集10 mL 培养瓶内气体,采用气相色谱-质谱联用仪(7890A GC-5975C MSD,Agilent)测定其N2O浓度.PAO速率根据NO随时间积累的回归关系计算. ...
亚热带丘陵区微塑料输入对稻麦轮作农田土壤N2O排放及产生路径的影响
2
... AOB与AOA的生态位分化和生理特征决定了二者对PAO和PNR的贡献差异[9].AOA具有更高的氨亲和力和较低的能量需求,能够在低NH和干旱环境下主导氨氧化[10].而AOB虽然对氨的利用效率不及AOA,但其最大比活性更高,能够在高氮供应和中性至碱性土壤中快速响应,从而在短期内显著提高PAO和PNR[11].除微生物本身的生理差异外,土壤理化性质(如pH、含水量、可利用氮素水平)、土地利用方式及管理措施(如化肥施用、耕作制度)也会改变AOB与AOA的相对丰度和活性,进而调节PAO与PNR[12-13].在农田系统中,AOB与AOA并非简单的替代关系,而是在不同环境条件下通过功能互补共同维持氨氧化功能[14]. ...
... 土壤潜在硝化速率(PNR)采用15N同位素标记方法测定,具体测定方法和步骤请参照王丽莎等[9].土壤净硝化速率(Net nitrification rate,NNR)采用室内培养法测定[8],称取10 g鲜土(以干重计)置于250 mL广口玻璃瓶中(同时,为测定培养期间土壤矿质氮含量的变化,称取1 kg(以干重计)土壤于2 L培养瓶中),用去离子水调节田间持水量(WHC)至50%.各处理设置3个重复,通过覆盖封口膜并在膜上刺孔,能够保持土壤微生物适宜的通气条件,既保证氨氧化和硝化微生物活性,又避免因缺氧而显著增强反硝化产生的N2O,从而使测定结果更准确反映PNR.随后将样品置于25 ℃黑暗条件下预培养3 d,以消除干湿效应并活化微生物.预培养结束后,在25 ℃条件下好氧培养28 d.培养期间,每隔2~3 d打开膜透气约30 min,并通过称重补水的方法维持WHC稳定在50%.于正式培养1、5、12、20、28 d采集烧杯中的土壤,用于测定土壤NH-N,NO-N和NO-N含量.土壤NH-N,NO-N和NO-N排放强度采用时间加权法. ...
Kinetic analysis of a complete nitrifier reveals an oligotrophic lifestyle
1
2017
... AOB与AOA的生态位分化和生理特征决定了二者对PAO和PNR的贡献差异[9].AOA具有更高的氨亲和力和较低的能量需求,能够在低NH和干旱环境下主导氨氧化[10].而AOB虽然对氨的利用效率不及AOA,但其最大比活性更高,能够在高氮供应和中性至碱性土壤中快速响应,从而在短期内显著提高PAO和PNR[11].除微生物本身的生理差异外,土壤理化性质(如pH、含水量、可利用氮素水平)、土地利用方式及管理措施(如化肥施用、耕作制度)也会改变AOB与AOA的相对丰度和活性,进而调节PAO与PNR[12-13].在农田系统中,AOB与AOA并非简单的替代关系,而是在不同环境条件下通过功能互补共同维持氨氧化功能[14]. ...
Environmental determinants of spatial-temporal variability in ammonia oxidizing microorganisms across diverse Chinese agricultural soils
2
2025
... AOB与AOA的生态位分化和生理特征决定了二者对PAO和PNR的贡献差异[9].AOA具有更高的氨亲和力和较低的能量需求,能够在低NH和干旱环境下主导氨氧化[10].而AOB虽然对氨的利用效率不及AOA,但其最大比活性更高,能够在高氮供应和中性至碱性土壤中快速响应,从而在短期内显著提高PAO和PNR[11].除微生物本身的生理差异外,土壤理化性质(如pH、含水量、可利用氮素水平)、土地利用方式及管理措施(如化肥施用、耕作制度)也会改变AOB与AOA的相对丰度和活性,进而调节PAO与PNR[12-13].在农田系统中,AOB与AOA并非简单的替代关系,而是在不同环境条件下通过功能互补共同维持氨氧化功能[14]. ...
... 本研究结果表明,NNR、PNR和PAO均随开垦年限的增加显著增加.OOF处理的NNR和PNR相较于USL提高了数倍,表明农业开垦和管理(如施肥、灌溉、耕作)从根本上改变了土壤的氮素周转能力.在长期耕作农田系统中,持续施肥和翻耕通常导致NH-N含量积累及氧气供应充足,为氨氧化微生物的生长与活性提供有利条件[33].Wang等[34]的研究也表明持续耕作通过改变土壤氮素形态及通气环境,显著促进土壤硝化进程.这种环境改善直接体现在硝化微生物群落的响应上,我们的数据显示,AOA和AOB的基因丰度随开垦年限显著增加,并与PNR和PAO的变化趋势一致.以上结果表明,连续耕作通过富集土壤中的AOA和AOB群落,强化了硝化作用的微生物驱动力[11].在所有处理中,AOA的基因拷贝数始终比AOB高出1~2个数量级,并且在PAO的贡献中占主导地位.这与许多研究认为AOA在多数陆地生态系统中是氨氧化的主要执行者结论相符,尤其在低NH-N、弱碱性或干旱土壤中,AOA主导氨氧化进程[35]. ...
太湖常见3种沉水植物附着生物的生物量及潜在反硝化速率
2
2024
... AOB与AOA的生态位分化和生理特征决定了二者对PAO和PNR的贡献差异[9].AOA具有更高的氨亲和力和较低的能量需求,能够在低NH和干旱环境下主导氨氧化[10].而AOB虽然对氨的利用效率不及AOA,但其最大比活性更高,能够在高氮供应和中性至碱性土壤中快速响应,从而在短期内显著提高PAO和PNR[11].除微生物本身的生理差异外,土壤理化性质(如pH、含水量、可利用氮素水平)、土地利用方式及管理措施(如化肥施用、耕作制度)也会改变AOB与AOA的相对丰度和活性,进而调节PAO与PNR[12-13].在农田系统中,AOB与AOA并非简单的替代关系,而是在不同环境条件下通过功能互补共同维持氨氧化功能[14]. ...
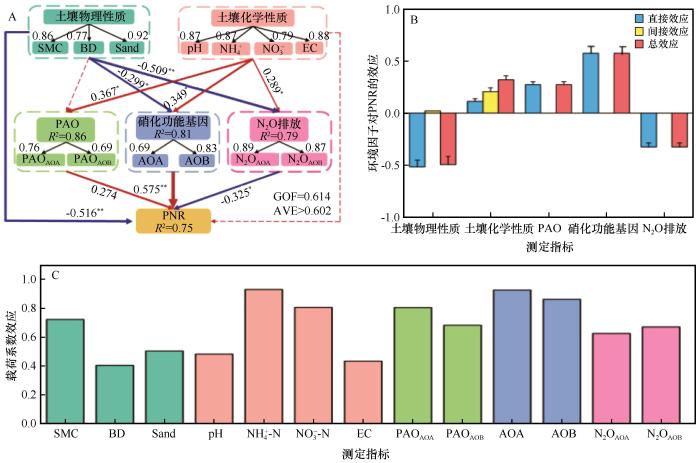
... PLS-PM结果表明硝化菌基因丰度对PNR的直接正效应最强(0.575),其次为PAO(0.274),而N2O排放对PNR的影响为负(-0.325),表明硝化潜力主要受氨氧化微生物的代谢潜能驱动,而N2O排放过程可能与硝化途径中中间产物(如NO)的积累及反硝化竞争有关.土壤物理性质对PNR表现出负效应(-0.516),反映了长期耕作后物理环境对硝化过程的复杂调控.一方面,耕作提高了土壤保水能力与团聚体稳定性,导致水分充填孔隙比例(WFPS)升高、氧气扩散受限,形成微尺度厌氧区,抑制需氧的氨氧化过程[36].另一方面,SOM与DOC积累促进异养微生物呼吸,加剧氧气竞争,进一步降低硝化微生物的代谢活性[37].此外,孔径结构由大孔向细孔转变亦可能削弱气体传输效率,使通气性改善的特征未能转化为有效氧供[37-38].土壤化学性质通过调节PAO、功能基因丰度及N2O排放间接影响PNR,交叉载荷结果表明,NH-N、NO-N、AOA丰度和AOB丰度是预测PNR的主要因子,说明氮源供给与氨氧化功能群的丰度之间存在密切耦合关系.氮素可利用性不仅直接影响氨氧化底物浓度,还会通过改变微生物群落的竞争格局,驱动AOA与AOB之间的生态分化[12]. ...
Promotion of maize straw degradation rate by altering microbial community structure through the addition of soybean straw
1
2025
... AOB与AOA的生态位分化和生理特征决定了二者对PAO和PNR的贡献差异[9].AOA具有更高的氨亲和力和较低的能量需求,能够在低NH和干旱环境下主导氨氧化[10].而AOB虽然对氨的利用效率不及AOA,但其最大比活性更高,能够在高氮供应和中性至碱性土壤中快速响应,从而在短期内显著提高PAO和PNR[11].除微生物本身的生理差异外,土壤理化性质(如pH、含水量、可利用氮素水平)、土地利用方式及管理措施(如化肥施用、耕作制度)也会改变AOB与AOA的相对丰度和活性,进而调节PAO与PNR[12-13].在农田系统中,AOB与AOA并非简单的替代关系,而是在不同环境条件下通过功能互补共同维持氨氧化功能[14]. ...
农田土壤N2O排放的关键过程及影响因素
1
2019
... AOB与AOA的生态位分化和生理特征决定了二者对PAO和PNR的贡献差异[9].AOA具有更高的氨亲和力和较低的能量需求,能够在低NH和干旱环境下主导氨氧化[10].而AOB虽然对氨的利用效率不及AOA,但其最大比活性更高,能够在高氮供应和中性至碱性土壤中快速响应,从而在短期内显著提高PAO和PNR[11].除微生物本身的生理差异外,土壤理化性质(如pH、含水量、可利用氮素水平)、土地利用方式及管理措施(如化肥施用、耕作制度)也会改变AOB与AOA的相对丰度和活性,进而调节PAO与PNR[12-13].在农田系统中,AOB与AOA并非简单的替代关系,而是在不同环境条件下通过功能互补共同维持氨氧化功能[14]. ...
长期不同施肥运筹对玉米产量,氮素转运和土壤氮积累的影响
2
2024
... 河西走廊荒漠绿洲过渡带区降水稀少,土壤疏松且肥力低.为满足粮食安全,当地农户普遍采取覆膜+大水高肥的连作制度[15].连续耕作一方面促进土壤发育和氮素供应,另一方面可能引起理化性质恶化和微生物群落重组,从而影响硝化过程及其伴随的N2O排放[16].自然沙地开垦及持续耕作引起的养分与水分变化,可能会通过调节AOA与AOB的相对贡献,影响PAO、PNR及氨氧化介导的N2O排放.然而,目前关于连续耕作背景下荒漠绿洲农田土壤AOA与AOB对PAO和N2O排放的相对贡献仍缺乏系统研究.特别是在河西走廊这一典型干旱绿洲区,土壤氮循环对长期耕作干扰的响应机制尚不明确,限制了科学施肥与可持续耕作制度的制定. ...
... 研究区位于甘肃河西走廊中段黑河流域的临泽县荒漠-绿洲过渡带,海拔1 350~1 400 m.该地区属典型的温带大陆性气候,年平均气温约7.6 ℃,极端最低和最高气温分别为-27 ℃和39 ℃.多年(1965—2020年)平均降水量117 mm,多年平均蒸散量高达2 390 mm,干旱特征明显.区域土壤母质为第四纪洪积-冲积物,主要土壤类型包括绿洲潮土、灌淤土、灌耕风沙土和灌耕灰棕漠土[17].由于质地偏砂,土壤普遍存在有机质偏低及保水保肥能力弱的特点.自20世纪60年代以来,为满足粮食生产需求,大量沙地被持续开垦为农田,目前近半数耕地属于新垦沙地农田,由于开垦年限不同,土壤熟化程度各异[7].该区农业以一年一熟的制种玉米种植为主(占耕地面积75%以上),普遍采用覆膜栽培和大水漫灌的传统耕作模式.由于土壤先天养分贫瘠与持水能力差,玉米生产高度依赖灌溉与化肥投入,玉米生育期内年均氮、磷、钾肥施用量分别约为200~400 kg·hm-2(以N计)、90~150 kg·hm-2(以P2O5计)和60~90 kg·hm-2(以K2O计),灌溉量在500~1 200 mm,灌溉水源主要来自黑河与地下水[15]. ...
荒漠土壤氧化亚氮排放及其驱动因素研究进展
1
2023
... 河西走廊荒漠绿洲过渡带区降水稀少,土壤疏松且肥力低.为满足粮食安全,当地农户普遍采取覆膜+大水高肥的连作制度[15].连续耕作一方面促进土壤发育和氮素供应,另一方面可能引起理化性质恶化和微生物群落重组,从而影响硝化过程及其伴随的N2O排放[16].自然沙地开垦及持续耕作引起的养分与水分变化,可能会通过调节AOA与AOB的相对贡献,影响PAO、PNR及氨氧化介导的N2O排放.然而,目前关于连续耕作背景下荒漠绿洲农田土壤AOA与AOB对PAO和N2O排放的相对贡献仍缺乏系统研究.特别是在河西走廊这一典型干旱绿洲区,土壤氮循环对长期耕作干扰的响应机制尚不明确,限制了科学施肥与可持续耕作制度的制定. ...
河西走廊荒漠绿洲农田氮素吸收、残留与淋失特征
1
2024
... 研究区位于甘肃河西走廊中段黑河流域的临泽县荒漠-绿洲过渡带,海拔1 350~1 400 m.该地区属典型的温带大陆性气候,年平均气温约7.6 ℃,极端最低和最高气温分别为-27 ℃和39 ℃.多年(1965—2020年)平均降水量117 mm,多年平均蒸散量高达2 390 mm,干旱特征明显.区域土壤母质为第四纪洪积-冲积物,主要土壤类型包括绿洲潮土、灌淤土、灌耕风沙土和灌耕灰棕漠土[17].由于质地偏砂,土壤普遍存在有机质偏低及保水保肥能力弱的特点.自20世纪60年代以来,为满足粮食生产需求,大量沙地被持续开垦为农田,目前近半数耕地属于新垦沙地农田,由于开垦年限不同,土壤熟化程度各异[7].该区农业以一年一熟的制种玉米种植为主(占耕地面积75%以上),普遍采用覆膜栽培和大水漫灌的传统耕作模式.由于土壤先天养分贫瘠与持水能力差,玉米生产高度依赖灌溉与化肥投入,玉米生育期内年均氮、磷、钾肥施用量分别约为200~400 kg·hm-2(以N计)、90~150 kg·hm-2(以P2O5计)和60~90 kg·hm-2(以K2O计),灌溉量在500~1 200 mm,灌溉水源主要来自黑河与地下水[15]. ...
Biochar-enriched soil mitigated N2O and NO emissions similarly as fresh biochar for wheat production
1
2019
... AOB和AOA对PAO和N2O排放的相对贡献的计算参照Wu等[18]的方法:AOA驱动的PAO(N2O)=辛炔组-乙炔组,AOB驱动PAO(N2O)=对照组-辛炔组. ...
农田土壤有机碳积累和团聚体稳定过程及其相互作用关系
1
2024
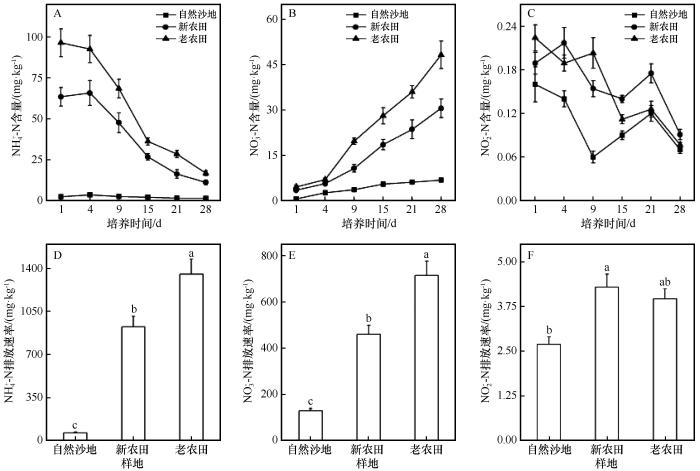
... 与USL相比,长期耕作土壤(YOF和OOF)的SMC显著提高,BD明显降低,表明耕作促进了土壤结构优化和孔隙度增加.长期耕作过程中,植物根系生长、微生物活动及有机质输入增强了团聚体形成,提高了土壤保水保肥能力[19-20].同时,沙粒含量显著下降,土壤质地由砂质向壤质转变,有利于形成更稳定的耕作层[21].尽管pH略有下降,但仍维持在碱性范围内,说明荒漠绿洲过渡带土壤碳酸盐含量高、缓冲能力强,可中和施肥或有机质分解产生的酸性物质,因此耕作对酸化的影响有限[22-23].EC变化不显著,表明盐分积累未明显发生.这可能与当地大水漫灌的传统方式有关,频繁灌溉使盐分随水下移或排出,从而抑制表层盐分聚集[24-25].此外,长期耕作显著促进了土壤养分积累,尤其是氮素和碳源相关指标.与USL相比,OOF样地NH-N和NO-N分别提高38.5倍和7.5倍,TN和SOM含量亦分别提高9倍和4.4倍,DOC显著上升至30.0 mg·L-1.这表明连续耕作通过植物残体归还、施肥管理和根系分泌等途径显著增强了氮素供应和碳输入,从而提升了土壤肥力与微生物活性[26-28]. ...
三江平原旱作黑土质量演变及其驱动因子分析
1
2025
... 与USL相比,长期耕作土壤(YOF和OOF)的SMC显著提高,BD明显降低,表明耕作促进了土壤结构优化和孔隙度增加.长期耕作过程中,植物根系生长、微生物活动及有机质输入增强了团聚体形成,提高了土壤保水保肥能力[19-20].同时,沙粒含量显著下降,土壤质地由砂质向壤质转变,有利于形成更稳定的耕作层[21].尽管pH略有下降,但仍维持在碱性范围内,说明荒漠绿洲过渡带土壤碳酸盐含量高、缓冲能力强,可中和施肥或有机质分解产生的酸性物质,因此耕作对酸化的影响有限[22-23].EC变化不显著,表明盐分积累未明显发生.这可能与当地大水漫灌的传统方式有关,频繁灌溉使盐分随水下移或排出,从而抑制表层盐分聚集[24-25].此外,长期耕作显著促进了土壤养分积累,尤其是氮素和碳源相关指标.与USL相比,OOF样地NH-N和NO-N分别提高38.5倍和7.5倍,TN和SOM含量亦分别提高9倍和4.4倍,DOC显著上升至30.0 mg·L-1.这表明连续耕作通过植物残体归还、施肥管理和根系分泌等途径显著增强了氮素供应和碳输入,从而提升了土壤肥力与微生物活性[26-28]. ...
松嫩平原不同类型土壤剖面形态描述及其性状分析
1
2024
... 与USL相比,长期耕作土壤(YOF和OOF)的SMC显著提高,BD明显降低,表明耕作促进了土壤结构优化和孔隙度增加.长期耕作过程中,植物根系生长、微生物活动及有机质输入增强了团聚体形成,提高了土壤保水保肥能力[19-20].同时,沙粒含量显著下降,土壤质地由砂质向壤质转变,有利于形成更稳定的耕作层[21].尽管pH略有下降,但仍维持在碱性范围内,说明荒漠绿洲过渡带土壤碳酸盐含量高、缓冲能力强,可中和施肥或有机质分解产生的酸性物质,因此耕作对酸化的影响有限[22-23].EC变化不显著,表明盐分积累未明显发生.这可能与当地大水漫灌的传统方式有关,频繁灌溉使盐分随水下移或排出,从而抑制表层盐分聚集[24-25].此外,长期耕作显著促进了土壤养分积累,尤其是氮素和碳源相关指标.与USL相比,OOF样地NH-N和NO-N分别提高38.5倍和7.5倍,TN和SOM含量亦分别提高9倍和4.4倍,DOC显著上升至30.0 mg·L-1.这表明连续耕作通过植物残体归还、施肥管理和根系分泌等途径显著增强了氮素供应和碳输入,从而提升了土壤肥力与微生物活性[26-28]. ...
Impact of long-term fertilization on agroecosystem multifunctionality and yield in Loess Plateau,China
1
2024
... 与USL相比,长期耕作土壤(YOF和OOF)的SMC显著提高,BD明显降低,表明耕作促进了土壤结构优化和孔隙度增加.长期耕作过程中,植物根系生长、微生物活动及有机质输入增强了团聚体形成,提高了土壤保水保肥能力[19-20].同时,沙粒含量显著下降,土壤质地由砂质向壤质转变,有利于形成更稳定的耕作层[21].尽管pH略有下降,但仍维持在碱性范围内,说明荒漠绿洲过渡带土壤碳酸盐含量高、缓冲能力强,可中和施肥或有机质分解产生的酸性物质,因此耕作对酸化的影响有限[22-23].EC变化不显著,表明盐分积累未明显发生.这可能与当地大水漫灌的传统方式有关,频繁灌溉使盐分随水下移或排出,从而抑制表层盐分聚集[24-25].此外,长期耕作显著促进了土壤养分积累,尤其是氮素和碳源相关指标.与USL相比,OOF样地NH-N和NO-N分别提高38.5倍和7.5倍,TN和SOM含量亦分别提高9倍和4.4倍,DOC显著上升至30.0 mg·L-1.这表明连续耕作通过植物残体归还、施肥管理和根系分泌等途径显著增强了氮素供应和碳输入,从而提升了土壤肥力与微生物活性[26-28]. ...
Shifting sands to sustainable soils: Spatial dynamics of soil water and salinity in a desert oasis ecotone
1
2025
... 与USL相比,长期耕作土壤(YOF和OOF)的SMC显著提高,BD明显降低,表明耕作促进了土壤结构优化和孔隙度增加.长期耕作过程中,植物根系生长、微生物活动及有机质输入增强了团聚体形成,提高了土壤保水保肥能力[19-20].同时,沙粒含量显著下降,土壤质地由砂质向壤质转变,有利于形成更稳定的耕作层[21].尽管pH略有下降,但仍维持在碱性范围内,说明荒漠绿洲过渡带土壤碳酸盐含量高、缓冲能力强,可中和施肥或有机质分解产生的酸性物质,因此耕作对酸化的影响有限[22-23].EC变化不显著,表明盐分积累未明显发生.这可能与当地大水漫灌的传统方式有关,频繁灌溉使盐分随水下移或排出,从而抑制表层盐分聚集[24-25].此外,长期耕作显著促进了土壤养分积累,尤其是氮素和碳源相关指标.与USL相比,OOF样地NH-N和NO-N分别提高38.5倍和7.5倍,TN和SOM含量亦分别提高9倍和4.4倍,DOC显著上升至30.0 mg·L-1.这表明连续耕作通过植物残体归还、施肥管理和根系分泌等途径显著增强了氮素供应和碳输入,从而提升了土壤肥力与微生物活性[26-28]. ...
Soil water and salt migration in oasis farmland during crop growing season
1
2023
... 与USL相比,长期耕作土壤(YOF和OOF)的SMC显著提高,BD明显降低,表明耕作促进了土壤结构优化和孔隙度增加.长期耕作过程中,植物根系生长、微生物活动及有机质输入增强了团聚体形成,提高了土壤保水保肥能力[19-20].同时,沙粒含量显著下降,土壤质地由砂质向壤质转变,有利于形成更稳定的耕作层[21].尽管pH略有下降,但仍维持在碱性范围内,说明荒漠绿洲过渡带土壤碳酸盐含量高、缓冲能力强,可中和施肥或有机质分解产生的酸性物质,因此耕作对酸化的影响有限[22-23].EC变化不显著,表明盐分积累未明显发生.这可能与当地大水漫灌的传统方式有关,频繁灌溉使盐分随水下移或排出,从而抑制表层盐分聚集[24-25].此外,长期耕作显著促进了土壤养分积累,尤其是氮素和碳源相关指标.与USL相比,OOF样地NH-N和NO-N分别提高38.5倍和7.5倍,TN和SOM含量亦分别提高9倍和4.4倍,DOC显著上升至30.0 mg·L-1.这表明连续耕作通过植物残体归还、施肥管理和根系分泌等途径显著增强了氮素供应和碳输入,从而提升了土壤肥力与微生物活性[26-28]. ...
Soil texture modulates microbial responses to irrigation:implications for nutrient cycling in arid agroecosystem
1
2026
... 与USL相比,长期耕作土壤(YOF和OOF)的SMC显著提高,BD明显降低,表明耕作促进了土壤结构优化和孔隙度增加.长期耕作过程中,植物根系生长、微生物活动及有机质输入增强了团聚体形成,提高了土壤保水保肥能力[19-20].同时,沙粒含量显著下降,土壤质地由砂质向壤质转变,有利于形成更稳定的耕作层[21].尽管pH略有下降,但仍维持在碱性范围内,说明荒漠绿洲过渡带土壤碳酸盐含量高、缓冲能力强,可中和施肥或有机质分解产生的酸性物质,因此耕作对酸化的影响有限[22-23].EC变化不显著,表明盐分积累未明显发生.这可能与当地大水漫灌的传统方式有关,频繁灌溉使盐分随水下移或排出,从而抑制表层盐分聚集[24-25].此外,长期耕作显著促进了土壤养分积累,尤其是氮素和碳源相关指标.与USL相比,OOF样地NH-N和NO-N分别提高38.5倍和7.5倍,TN和SOM含量亦分别提高9倍和4.4倍,DOC显著上升至30.0 mg·L-1.这表明连续耕作通过植物残体归还、施肥管理和根系分泌等途径显著增强了氮素供应和碳输入,从而提升了土壤肥力与微生物活性[26-28]. ...
减氮及有机替代对新疆棉田土壤氮素有效性和利用效率的影响
1
2024
... 与USL相比,长期耕作土壤(YOF和OOF)的SMC显著提高,BD明显降低,表明耕作促进了土壤结构优化和孔隙度增加.长期耕作过程中,植物根系生长、微生物活动及有机质输入增强了团聚体形成,提高了土壤保水保肥能力[19-20].同时,沙粒含量显著下降,土壤质地由砂质向壤质转变,有利于形成更稳定的耕作层[21].尽管pH略有下降,但仍维持在碱性范围内,说明荒漠绿洲过渡带土壤碳酸盐含量高、缓冲能力强,可中和施肥或有机质分解产生的酸性物质,因此耕作对酸化的影响有限[22-23].EC变化不显著,表明盐分积累未明显发生.这可能与当地大水漫灌的传统方式有关,频繁灌溉使盐分随水下移或排出,从而抑制表层盐分聚集[24-25].此外,长期耕作显著促进了土壤养分积累,尤其是氮素和碳源相关指标.与USL相比,OOF样地NH-N和NO-N分别提高38.5倍和7.5倍,TN和SOM含量亦分别提高9倍和4.4倍,DOC显著上升至30.0 mg·L-1.这表明连续耕作通过植物残体归还、施肥管理和根系分泌等途径显著增强了氮素供应和碳输入,从而提升了土壤肥力与微生物活性[26-28]. ...
Crop rotation and diversification in China:enhancing sustainable agriculture and resilience
0
2024
Exploring the link between soil health and crop productivity
1
2025
... 与USL相比,长期耕作土壤(YOF和OOF)的SMC显著提高,BD明显降低,表明耕作促进了土壤结构优化和孔隙度增加.长期耕作过程中,植物根系生长、微生物活动及有机质输入增强了团聚体形成,提高了土壤保水保肥能力[19-20].同时,沙粒含量显著下降,土壤质地由砂质向壤质转变,有利于形成更稳定的耕作层[21].尽管pH略有下降,但仍维持在碱性范围内,说明荒漠绿洲过渡带土壤碳酸盐含量高、缓冲能力强,可中和施肥或有机质分解产生的酸性物质,因此耕作对酸化的影响有限[22-23].EC变化不显著,表明盐分积累未明显发生.这可能与当地大水漫灌的传统方式有关,频繁灌溉使盐分随水下移或排出,从而抑制表层盐分聚集[24-25].此外,长期耕作显著促进了土壤养分积累,尤其是氮素和碳源相关指标.与USL相比,OOF样地NH-N和NO-N分别提高38.5倍和7.5倍,TN和SOM含量亦分别提高9倍和4.4倍,DOC显著上升至30.0 mg·L-1.这表明连续耕作通过植物残体归还、施肥管理和根系分泌等途径显著增强了氮素供应和碳输入,从而提升了土壤肥力与微生物活性[26-28]. ...
Natural organic matter drives nitrogen biogeochemical cycling in groundwater of the Hetao Basin: evidence from multi-isotopes and FT-ICR MS
3
2025
... 培养实验显示耕作显著影响土壤无机氮动态.NH-N在培养初期(1~4 d)短暂升高后迅速下降,反映了氨化作用先行、硝化作用随后增强的过程.OOF土壤NH-N含量和排放强度高于YOF和USL,表明有机质积累和微生物活性增强了氮素矿化与硝化潜力[29].SOM和DOC显著增加为异养微生物提供丰富碳源,促进有机氮向无机氮的转化[30].NO-N含量在培养过程中持续升高,OOF始终显著高于其他处理,说明长期耕作增强了硝化速率和无机氮累积.OOF土壤NH-N、SOM和DOC含量较高,同时土壤结构的改善提高了通气性,加快NH向NO的转化并促进无机氮积累[29,31].NO-N含量整体较低,波动小,表明其作为硝化过程的中间产物并未大量积累.然而,YOF的NO-N累积排放强度最高,可能因氨氧化菌活性较高而亚硝酸盐氧化菌活性尚未匹配,加之YOF土壤NH-N含量适中,使NO在土壤中停留时间延长[29].此外,YOF孔隙结构尚不均匀,局部缺氧,也可能短期抑制NO氧化,导致短期积累[32]. ...
... [29,31].NO-N含量整体较低,波动小,表明其作为硝化过程的中间产物并未大量积累.然而,YOF的NO-N累积排放强度最高,可能因氨氧化菌活性较高而亚硝酸盐氧化菌活性尚未匹配,加之YOF土壤NH-N含量适中,使NO在土壤中停留时间延长[29].此外,YOF孔隙结构尚不均匀,局部缺氧,也可能短期抑制NO氧化,导致短期积累[32]. ...
... [29].此外,YOF孔隙结构尚不均匀,局部缺氧,也可能短期抑制NO氧化,导致短期积累[32]. ...
有机氮替代无机氮对旱作马铃薯生长,产量及农田碳排放的影响
1
2025
... 培养实验显示耕作显著影响土壤无机氮动态.NH-N在培养初期(1~4 d)短暂升高后迅速下降,反映了氨化作用先行、硝化作用随后增强的过程.OOF土壤NH-N含量和排放强度高于YOF和USL,表明有机质积累和微生物活性增强了氮素矿化与硝化潜力[29].SOM和DOC显著增加为异养微生物提供丰富碳源,促进有机氮向无机氮的转化[30].NO-N含量在培养过程中持续升高,OOF始终显著高于其他处理,说明长期耕作增强了硝化速率和无机氮累积.OOF土壤NH-N、SOM和DOC含量较高,同时土壤结构的改善提高了通气性,加快NH向NO的转化并促进无机氮积累[29,31].NO-N含量整体较低,波动小,表明其作为硝化过程的中间产物并未大量积累.然而,YOF的NO-N累积排放强度最高,可能因氨氧化菌活性较高而亚硝酸盐氧化菌活性尚未匹配,加之YOF土壤NH-N含量适中,使NO在土壤中停留时间延长[29].此外,YOF孔隙结构尚不均匀,局部缺氧,也可能短期抑制NO氧化,导致短期积累[32]. ...
西北旱地苹果细根分布及水力特征对长期覆膜的响应
1
2021
... 培养实验显示耕作显著影响土壤无机氮动态.NH-N在培养初期(1~4 d)短暂升高后迅速下降,反映了氨化作用先行、硝化作用随后增强的过程.OOF土壤NH-N含量和排放强度高于YOF和USL,表明有机质积累和微生物活性增强了氮素矿化与硝化潜力[29].SOM和DOC显著增加为异养微生物提供丰富碳源,促进有机氮向无机氮的转化[30].NO-N含量在培养过程中持续升高,OOF始终显著高于其他处理,说明长期耕作增强了硝化速率和无机氮累积.OOF土壤NH-N、SOM和DOC含量较高,同时土壤结构的改善提高了通气性,加快NH向NO的转化并促进无机氮积累[29,31].NO-N含量整体较低,波动小,表明其作为硝化过程的中间产物并未大量积累.然而,YOF的NO-N累积排放强度最高,可能因氨氧化菌活性较高而亚硝酸盐氧化菌活性尚未匹配,加之YOF土壤NH-N含量适中,使NO在土壤中停留时间延长[29].此外,YOF孔隙结构尚不均匀,局部缺氧,也可能短期抑制NO氧化,导致短期积累[32]. ...
森林恢复对土壤氧化亚氮排放影响的生物与非生物学机制
1
2025
... 培养实验显示耕作显著影响土壤无机氮动态.NH-N在培养初期(1~4 d)短暂升高后迅速下降,反映了氨化作用先行、硝化作用随后增强的过程.OOF土壤NH-N含量和排放强度高于YOF和USL,表明有机质积累和微生物活性增强了氮素矿化与硝化潜力[29].SOM和DOC显著增加为异养微生物提供丰富碳源,促进有机氮向无机氮的转化[30].NO-N含量在培养过程中持续升高,OOF始终显著高于其他处理,说明长期耕作增强了硝化速率和无机氮累积.OOF土壤NH-N、SOM和DOC含量较高,同时土壤结构的改善提高了通气性,加快NH向NO的转化并促进无机氮积累[29,31].NO-N含量整体较低,波动小,表明其作为硝化过程的中间产物并未大量积累.然而,YOF的NO-N累积排放强度最高,可能因氨氧化菌活性较高而亚硝酸盐氧化菌活性尚未匹配,加之YOF土壤NH-N含量适中,使NO在土壤中停留时间延长[29].此外,YOF孔隙结构尚不均匀,局部缺氧,也可能短期抑制NO氧化,导致短期积累[32]. ...
草地土壤N2O排放对人为干扰的响应研究进展
2
2025
... 本研究结果表明,NNR、PNR和PAO均随开垦年限的增加显著增加.OOF处理的NNR和PNR相较于USL提高了数倍,表明农业开垦和管理(如施肥、灌溉、耕作)从根本上改变了土壤的氮素周转能力.在长期耕作农田系统中,持续施肥和翻耕通常导致NH-N含量积累及氧气供应充足,为氨氧化微生物的生长与活性提供有利条件[33].Wang等[34]的研究也表明持续耕作通过改变土壤氮素形态及通气环境,显著促进土壤硝化进程.这种环境改善直接体现在硝化微生物群落的响应上,我们的数据显示,AOA和AOB的基因丰度随开垦年限显著增加,并与PNR和PAO的变化趋势一致.以上结果表明,连续耕作通过富集土壤中的AOA和AOB群落,强化了硝化作用的微生物驱动力[11].在所有处理中,AOA的基因拷贝数始终比AOB高出1~2个数量级,并且在PAO的贡献中占主导地位.这与许多研究认为AOA在多数陆地生态系统中是氨氧化的主要执行者结论相符,尤其在低NH-N、弱碱性或干旱土壤中,AOA主导氨氧化进程[35]. ...
... 值得注意的是,AOA/AOB基因丰度比值随开垦年限呈降低趋势(USL>OOF>YOF).这表明在未开垦土壤中,硝化作用几乎由AOA绝对主导.而随着耕作年限延长和施肥管理的持续,AOB群落得到了更大幅度的富集,其在硝化过程中的相对重要性随之上升,反映了连续耕作过程中氨氧化群落由寡营养型向富营养型的演替趋势[35].这一转变与N2O排放来源的变化相互印证,在USL和YOF中,N2O排放主要由AOA驱动,而在OOF中则转变为AOB驱动为主.尽管AOA基因拷贝数在所有处理中均显著高于AOB,且主导了氨氧化潜势(PAO),但上述结果仍显示,微生物丰度并非决定N2O排放来源的唯一因素.研究表明,AOA多为寡营养型,对低浓度铵具有高亲和力,其代谢途径更倾向于将NH3高效转化为NO,而N2O产率较低[33];相比之下,AOB属于富营养型,在高铵环境中具有更高的最大比活性,且其氨氧化过程易于通过羟胺(NH2OH)不完全氧化等途径产生较多N2O[35].本研究中,长期耕作与高量化肥投入(OOF)塑造了持续的高铵环境,这不仅显著富集了AOB种群,更可能激发了其高代谢活性与强N2O产生能力.因此,在OOF处理中,尽管AOB的绝对基因丰度仍低于AOA,但其单位细胞或单位amoA基因的N2O产率更高,加上高底物浓度下整体硝化通量增大,共同导致AOB对N2O排放的实际贡献占据主导[4-6]. ...
Continuous application of conservation tillage affects in situ N2O emissions and nitrogen cycling gene abundances following nitrogen fertilization
1
2021
... 本研究结果表明,NNR、PNR和PAO均随开垦年限的增加显著增加.OOF处理的NNR和PNR相较于USL提高了数倍,表明农业开垦和管理(如施肥、灌溉、耕作)从根本上改变了土壤的氮素周转能力.在长期耕作农田系统中,持续施肥和翻耕通常导致NH-N含量积累及氧气供应充足,为氨氧化微生物的生长与活性提供有利条件[33].Wang等[34]的研究也表明持续耕作通过改变土壤氮素形态及通气环境,显著促进土壤硝化进程.这种环境改善直接体现在硝化微生物群落的响应上,我们的数据显示,AOA和AOB的基因丰度随开垦年限显著增加,并与PNR和PAO的变化趋势一致.以上结果表明,连续耕作通过富集土壤中的AOA和AOB群落,强化了硝化作用的微生物驱动力[11].在所有处理中,AOA的基因拷贝数始终比AOB高出1~2个数量级,并且在PAO的贡献中占主导地位.这与许多研究认为AOA在多数陆地生态系统中是氨氧化的主要执行者结论相符,尤其在低NH-N、弱碱性或干旱土壤中,AOA主导氨氧化进程[35]. ...
Linking N2O emissions and nosZ gene abundance:a meta-analysis of organic carbon amendments in agricultural soils
4
2025
... 本研究结果表明,NNR、PNR和PAO均随开垦年限的增加显著增加.OOF处理的NNR和PNR相较于USL提高了数倍,表明农业开垦和管理(如施肥、灌溉、耕作)从根本上改变了土壤的氮素周转能力.在长期耕作农田系统中,持续施肥和翻耕通常导致NH-N含量积累及氧气供应充足,为氨氧化微生物的生长与活性提供有利条件[33].Wang等[34]的研究也表明持续耕作通过改变土壤氮素形态及通气环境,显著促进土壤硝化进程.这种环境改善直接体现在硝化微生物群落的响应上,我们的数据显示,AOA和AOB的基因丰度随开垦年限显著增加,并与PNR和PAO的变化趋势一致.以上结果表明,连续耕作通过富集土壤中的AOA和AOB群落,强化了硝化作用的微生物驱动力[11].在所有处理中,AOA的基因拷贝数始终比AOB高出1~2个数量级,并且在PAO的贡献中占主导地位.这与许多研究认为AOA在多数陆地生态系统中是氨氧化的主要执行者结论相符,尤其在低NH-N、弱碱性或干旱土壤中,AOA主导氨氧化进程[35]. ...
... 值得注意的是,AOA/AOB基因丰度比值随开垦年限呈降低趋势(USL>OOF>YOF).这表明在未开垦土壤中,硝化作用几乎由AOA绝对主导.而随着耕作年限延长和施肥管理的持续,AOB群落得到了更大幅度的富集,其在硝化过程中的相对重要性随之上升,反映了连续耕作过程中氨氧化群落由寡营养型向富营养型的演替趋势[35].这一转变与N2O排放来源的变化相互印证,在USL和YOF中,N2O排放主要由AOA驱动,而在OOF中则转变为AOB驱动为主.尽管AOA基因拷贝数在所有处理中均显著高于AOB,且主导了氨氧化潜势(PAO),但上述结果仍显示,微生物丰度并非决定N2O排放来源的唯一因素.研究表明,AOA多为寡营养型,对低浓度铵具有高亲和力,其代谢途径更倾向于将NH3高效转化为NO,而N2O产率较低[33];相比之下,AOB属于富营养型,在高铵环境中具有更高的最大比活性,且其氨氧化过程易于通过羟胺(NH2OH)不完全氧化等途径产生较多N2O[35].本研究中,长期耕作与高量化肥投入(OOF)塑造了持续的高铵环境,这不仅显著富集了AOB种群,更可能激发了其高代谢活性与强N2O产生能力.因此,在OOF处理中,尽管AOB的绝对基因丰度仍低于AOA,但其单位细胞或单位amoA基因的N2O产率更高,加上高底物浓度下整体硝化通量增大,共同导致AOB对N2O排放的实际贡献占据主导[4-6]. ...
... [35].本研究中,长期耕作与高量化肥投入(OOF)塑造了持续的高铵环境,这不仅显著富集了AOB种群,更可能激发了其高代谢活性与强N2O产生能力.因此,在OOF处理中,尽管AOB的绝对基因丰度仍低于AOA,但其单位细胞或单位amoA基因的N2O产率更高,加上高底物浓度下整体硝化通量增大,共同导致AOB对N2O排放的实际贡献占据主导[4-6]. ...
... 值得注意的是,PAOAOA与AOA丰度的载荷系数均高于AOB相关变量,表明AOA在驱动PNR变化中仍占主导作用.这与AOA在干旱、碱性及低氮环境中的生态适应性相吻合[35].同时,AOA的胞外聚合物特征及对氧胁迫的耐受性有助于其在绿洲农田典型的干湿交替、弱碱性环境中占据优势[4].此外,N2O排放对PNR的负效应提示硝化与反硝化途径之间可能存在竞争性碳氮流动,当土壤含水量升高或氧气供应受限时,部分NO可能被还原为N2O而非进一步氧化为NO,从而削弱硝化潜力[39-40].这一现象在周期性灌溉条件下尤为显著,反映出绿洲农田土壤中氧化还原状态的时空变动对氮循环路径分配具有重要调节作用. ...
长期保护性耕作下土壤团聚体全氮与氮功能微生物关系研究
1
2024
... PLS-PM结果表明硝化菌基因丰度对PNR的直接正效应最强(0.575),其次为PAO(0.274),而N2O排放对PNR的影响为负(-0.325),表明硝化潜力主要受氨氧化微生物的代谢潜能驱动,而N2O排放过程可能与硝化途径中中间产物(如NO)的积累及反硝化竞争有关.土壤物理性质对PNR表现出负效应(-0.516),反映了长期耕作后物理环境对硝化过程的复杂调控.一方面,耕作提高了土壤保水能力与团聚体稳定性,导致水分充填孔隙比例(WFPS)升高、氧气扩散受限,形成微尺度厌氧区,抑制需氧的氨氧化过程[36].另一方面,SOM与DOC积累促进异养微生物呼吸,加剧氧气竞争,进一步降低硝化微生物的代谢活性[37].此外,孔径结构由大孔向细孔转变亦可能削弱气体传输效率,使通气性改善的特征未能转化为有效氧供[37-38].土壤化学性质通过调节PAO、功能基因丰度及N2O排放间接影响PNR,交叉载荷结果表明,NH-N、NO-N、AOA丰度和AOB丰度是预测PNR的主要因子,说明氮源供给与氨氧化功能群的丰度之间存在密切耦合关系.氮素可利用性不仅直接影响氨氧化底物浓度,还会通过改变微生物群落的竞争格局,驱动AOA与AOB之间的生态分化[12]. ...
The mechanism and optimization path of water regulation on soil nitrogen supply and release
2
2025
... PLS-PM结果表明硝化菌基因丰度对PNR的直接正效应最强(0.575),其次为PAO(0.274),而N2O排放对PNR的影响为负(-0.325),表明硝化潜力主要受氨氧化微生物的代谢潜能驱动,而N2O排放过程可能与硝化途径中中间产物(如NO)的积累及反硝化竞争有关.土壤物理性质对PNR表现出负效应(-0.516),反映了长期耕作后物理环境对硝化过程的复杂调控.一方面,耕作提高了土壤保水能力与团聚体稳定性,导致水分充填孔隙比例(WFPS)升高、氧气扩散受限,形成微尺度厌氧区,抑制需氧的氨氧化过程[36].另一方面,SOM与DOC积累促进异养微生物呼吸,加剧氧气竞争,进一步降低硝化微生物的代谢活性[37].此外,孔径结构由大孔向细孔转变亦可能削弱气体传输效率,使通气性改善的特征未能转化为有效氧供[37-38].土壤化学性质通过调节PAO、功能基因丰度及N2O排放间接影响PNR,交叉载荷结果表明,NH-N、NO-N、AOA丰度和AOB丰度是预测PNR的主要因子,说明氮源供给与氨氧化功能群的丰度之间存在密切耦合关系.氮素可利用性不仅直接影响氨氧化底物浓度,还会通过改变微生物群落的竞争格局,驱动AOA与AOB之间的生态分化[12]. ...
... [37-38].土壤化学性质通过调节PAO、功能基因丰度及N2O排放间接影响PNR,交叉载荷结果表明,NH-N、NO-N、AOA丰度和AOB丰度是预测PNR的主要因子,说明氮源供给与氨氧化功能群的丰度之间存在密切耦合关系.氮素可利用性不仅直接影响氨氧化底物浓度,还会通过改变微生物群落的竞争格局,驱动AOA与AOB之间的生态分化[12]. ...
西北荒漠区土壤氮素组分沿干旱梯度的空间变化
1
2024
... PLS-PM结果表明硝化菌基因丰度对PNR的直接正效应最强(0.575),其次为PAO(0.274),而N2O排放对PNR的影响为负(-0.325),表明硝化潜力主要受氨氧化微生物的代谢潜能驱动,而N2O排放过程可能与硝化途径中中间产物(如NO)的积累及反硝化竞争有关.土壤物理性质对PNR表现出负效应(-0.516),反映了长期耕作后物理环境对硝化过程的复杂调控.一方面,耕作提高了土壤保水能力与团聚体稳定性,导致水分充填孔隙比例(WFPS)升高、氧气扩散受限,形成微尺度厌氧区,抑制需氧的氨氧化过程[36].另一方面,SOM与DOC积累促进异养微生物呼吸,加剧氧气竞争,进一步降低硝化微生物的代谢活性[37].此外,孔径结构由大孔向细孔转变亦可能削弱气体传输效率,使通气性改善的特征未能转化为有效氧供[37-38].土壤化学性质通过调节PAO、功能基因丰度及N2O排放间接影响PNR,交叉载荷结果表明,NH-N、NO-N、AOA丰度和AOB丰度是预测PNR的主要因子,说明氮源供给与氨氧化功能群的丰度之间存在密切耦合关系.氮素可利用性不仅直接影响氨氧化底物浓度,还会通过改变微生物群落的竞争格局,驱动AOA与AOB之间的生态分化[12]. ...
秸秆还田条件下减施氮肥影响稻田土壤氧化亚氮排放的微生物机制
1
2024
... 值得注意的是,PAOAOA与AOA丰度的载荷系数均高于AOB相关变量,表明AOA在驱动PNR变化中仍占主导作用.这与AOA在干旱、碱性及低氮环境中的生态适应性相吻合[35].同时,AOA的胞外聚合物特征及对氧胁迫的耐受性有助于其在绿洲农田典型的干湿交替、弱碱性环境中占据优势[4].此外,N2O排放对PNR的负效应提示硝化与反硝化途径之间可能存在竞争性碳氮流动,当土壤含水量升高或氧气供应受限时,部分NO可能被还原为N2O而非进一步氧化为NO,从而削弱硝化潜力[39-40].这一现象在周期性灌溉条件下尤为显著,反映出绿洲农田土壤中氧化还原状态的时空变动对氮循环路径分配具有重要调节作用. ...
中国半干旱区农田土壤碳、氮、磷含量对玉米生产的影响
1
2022
... 值得注意的是,PAOAOA与AOA丰度的载荷系数均高于AOB相关变量,表明AOA在驱动PNR变化中仍占主导作用.这与AOA在干旱、碱性及低氮环境中的生态适应性相吻合[35].同时,AOA的胞外聚合物特征及对氧胁迫的耐受性有助于其在绿洲农田典型的干湿交替、弱碱性环境中占据优势[4].此外,N2O排放对PNR的负效应提示硝化与反硝化途径之间可能存在竞争性碳氮流动,当土壤含水量升高或氧气供应受限时,部分NO可能被还原为N2O而非进一步氧化为NO,从而削弱硝化潜力[39-40].这一现象在周期性灌溉条件下尤为显著,反映出绿洲农田土壤中氧化还原状态的时空变动对氮循环路径分配具有重要调节作用. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}