0 引言
生物结皮是细菌、藻类、真菌和孢子植物与土壤颗粒胶结而成的有机复合体[1 ] ,主要分布于沙漠或荒漠等干旱半干旱区,对干旱半干旱地区的水土保护、养分积累和生态恢复具有重要的生态意义[2 ] 。全球气候变化加剧,土地退化日益严重,而生物结皮作为土壤生态系统中的关键组成部分,在干旱半干旱区扮演关键角色,生物结皮通过固定沙漠表面的土壤,防止风蚀和水土流失,维持着土壤生态系统的稳定[3 ] 。生物结皮显著影响区域生态水文循环,直接或间接参与降水入渗、凝结水捕获和蒸发等生态水文过程[4 -5 ] 。干旱半干旱区生态水文循环中,生物结皮不仅能够根据生物特性对当地土壤特性起到一定的改善作用[6 ] ,从而维持流域生态平衡,并能够在生态水文循环的较大尺度上融入气候、土壤、高等植物作为整体考虑。研究生物结皮在生态水文循环中水分流动与转换的规律[5 ] ,有助于系统全面地揭示其在生态水文循环过程中所扮演的角色,但是目前由于缺乏生态水文循环中生物结皮的大尺度、长时序数据,生态水文研究对结皮考虑不足,因此如何发展大尺度、高频次生物结皮数据研发是干旱半干旱地区生物结皮研究的关键。
目前,生物结皮在中国北方干旱半干旱区以及南方湿润半湿润区均有大量研究,主要是通过站点观测数据分析结皮对土壤水碳固持等关键过程的影响[7 -10 ] 。生物结皮分布的提取方法主要有两种,一种是基于光谱特征的光谱分析法[11 -13 ] ,另外一种是基于遥感数据的波段代数运算构成的指数法[14 -17 ] 。在光谱分析法中,主要针对生物结皮的光谱进行分析,以光谱特征之间的差异来区分不同种类生物结皮,这种基于面向对象的提取方法在生物结皮相关提取中作为常识性应用。田园盛等[11 ] 借助光谱特征之间差异以区分藻类、藓类等不同类型生物结皮;宋丽丽等[12 ] 结合反映相片DN值信息的RGB阈值法和ML(最大似然分类)法高精度准确识别出分布于干旱半干旱退化草原区的生物结皮;Baxter等[13 ] 采用机器学习(随机森林)对生物结皮的光谱特征进行评估,以区分不同种类拥有不同结构形状的生物结皮。这些方法需要在站点尺度下的精细观测以确定阈值,但该阈值计算受人为因素或其他环境因素的影响。受野外气候、地貌、侵蚀类型等多方面环境因素的影响[18 -19 ] ,生物结皮的生物特性会发生一定变化,从而导致其光谱特性在不同条件下引起一定变化[11 ] ,生物结皮指数上下限阈值的准确性也会受一定的影响,因此采用生物结皮光谱特征提取方法的稳定性待提高。在实际应用中,由于生物结皮的空间分布复杂性和多样性,采用目视解译的方法易受到人为主观因素的影响,导致提取结果的不稳定性和不一致性。此外,生物结皮的覆盖范围广泛,包括草地、森林、湿地等不同类型的植被,这些生境特征在中低分辨率的遥感影像中难以准确区分,从而给生物结皮提取带来一定的挑战。另外,目视解译提取生物结皮需要大量的人力和时间投入,效率较低,尤其在大范围地区的生物结皮监测和研究中更为困难。
与此同时,在另外一种遥感结皮指数法中,上述方法出现的问题能够较好地被处理,少数学者通过遥感技术与站点尺度精细化观测相结合的方法对区域尺度生物结皮的空间分布及生物结皮的水文效应进行初步探讨,取得很大进展[13 -17 ,20 -21 ] 。Karnieli[20 ] 利用Landsat TM影像红色波段与蓝色波段归一化差值构建光谱生物结皮指数,在沙漠干旱环境下检测以蓝绿藻的生物结皮检测和绘制遥感图像中的不同岩性形态单元。Chen等[22 ] 通过定量实验发现生物结皮在红色与绿色波段之间的变化率比裸土、枯植或绿色植被更小,并且生物结皮在可见光与近红外波段的反射率比裸土、枯植或绿色植被要低得多,在此基础上构建了增大生物结皮与裸土、枯植、绿色植被之间差异的生物结皮指数。张元明等[14 ] 、杨伟等[16 ] 通过Landsat ETM数据中的绿光、红光、近红外波段地表反射率构建了三波段生物结皮指数,分别分析了空间上和时间上古班尔通古特沙漠以地衣为主的生物结皮分布特征与变化规律。冯秀绒等[17 ] 优化了由绿光、红光、近红外波段地表反射率构建的生物结皮提取方法,采用结皮指数、归一化植被指数、监督分类、坡度分级相结合的方法提取了毛乌素沙地以苔藓类结皮为主的生物结皮。Sun等[21 ] 基于植被辐射传输理论建立了一个生物结皮估计模型,在与其他模型进行比较后发现该模型对古尔班通古特沙漠的生物结皮具有良好的提取效果。综上所述,基于遥感方法提取生物结皮空间分布的手段主要是利用遥感数据的不同波段构建生物结皮指数,且指数主要基于红光吸收特征和近红外的反射特征的物理机制构建。然而,以上结皮指数主要在沙漠区域进行研究验证,沙漠区域地物类型以结皮和流动沙丘为主,结构单一。由于黄土高原大规模植被恢复工程的实施,致使地表地物结构发生了极大改变,植被覆盖度呈现明显增加趋势[23 ] 。因此,如何进行黄土高原植被覆盖下地表生物结皮提取,构建适用于干旱半干旱区多地物特征相互作用下生物结皮遥感提取方法是本研究的技术难点。
本研究选择干旱半干旱区黄土高原典型流域皇甫川为研究对象,利用野外观测与室内分析相结合的方法,考虑多地物混合效应,采用多遥感指数协同组合拟进行干旱半干旱区皇甫川流域生物结皮提取。主要目的是构建考虑多地物混合效应的干旱半干旱区典型流域地表生物结皮精细化提取的遥感方法,分析干旱半干旱区植被恢复后典型流域生物结皮空间分布特征及其影响因素,以期为干旱半干旱区地表生物结皮的遥感提取构建研究范式,为干旱半干旱地区生态水文过程研究提供数据支持。
1 研究区概况与数据来源
1.1 研究区概况
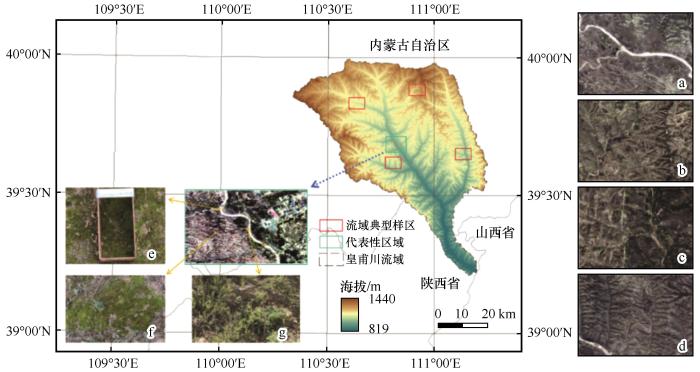
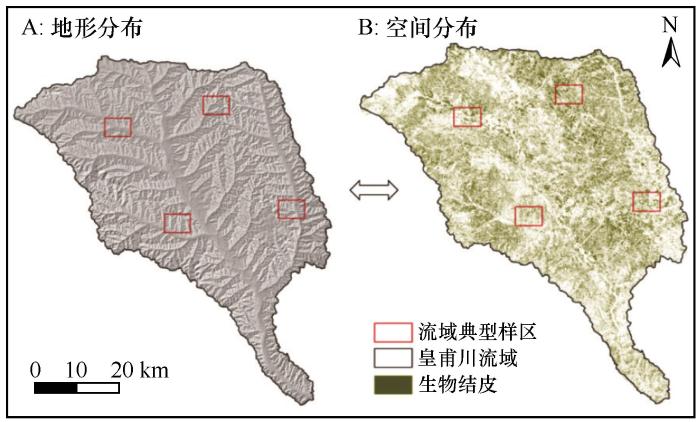
皇甫川流域面积3 246 km2 (图1 ),其中水土流失面积3 215 km2 ,占流域总面积的99.0%。流域属于干旱与半干旱气候,年降水量350~450 mm,是鄂尔多斯高原暴雨中心区,一般发生于夏季,占全年降水量的80%以上,年均暴雨5次,皇甫川沟深梁大、下游地势落差大的地形,造成了严重的流水侵蚀。皇甫川流域植被多密集分布于河流两侧,裸露河床与沙地均少有植被分布[24 ] 。而皇甫川流域生物结皮分布规律与流域内植被类型的演替密切相关,因此研究皇甫川流域生物结皮对流域内的植被恢复、生态环境保护以及生态水文循环研究具有重要意义。
图1
图1
皇甫川流域地理位置
注: a、b、c、d分别为流域典型样区;e、f、g分别为代表性区域的生物结皮解译标志
Fig.1
The geographic location of Huangfuchuan drainage basin
1.2 数据来源及数据处理
1.2.1 野外考察
于2023年8月进行皇甫川流域野外考察,受天气及道路可达性限制,采样主要沿道路选取具有代表性的800个采样点进行野外采样,样点选取标准为在一定范围内为单一植被类型,以减少影像中混合像元的干扰。收集结皮样品,采样同时利用GPS定位,记录采样点的海拔、植被种类和地貌类型等信息,使用便携式土壤水分检测仪记录土壤含水率,构建流域结皮解译标志。
1.2.2 数据来源
本研究选择Landsat8 OLI 数据为数据源;数据成像时间为2021年9月27日,空间分辨率为30 m。针对所使用的遥感特征指数,本研究选取了对地表地物类型具有较好识别效果的绿光、红光、近红外波段进行分析。此外,为探究皇甫川流域生物结皮空间分布与地形、降水、土地利用方式之间的关系,本文还选用了2021年皇甫川流域30 m分辨率地形高程数据;于Worldclim数据集中选取2021年年降水量数据,分辨率为1 km;选取了国家基础地理信息中心全球地表覆盖数据,分辨率为30 m。
1.2.3 数据处理与代表区域选取
本研究选取Landsat OLI传感器影像数据集,在ENVI 5.6上进行影像辐射定标、大气校正等预处理,并计算逐步提取过程中所使用到的各个遥感指数,随后选取影像中具有代表性的地物类型样本区进行分析研究。为了确保研究结果的准确性和精确度,选择1个代表性区域与4个典型样区(图1 )。代表性区域大小为356 m×213 m的30 m分辨率影像区域,包含各种典型地物,如建筑用地、林地、草地、水域、沙地、裸土、耕地等。在代表性区域内进行地物分类和样本点精度评估等工作;通过此区域分析,能够更好地了解该地区的地物分布情况,为后续研究和决策提供重要参考依据。
2 研究方法
2.1 生物结皮及其他目标地物光谱分割阈值确定方法
2.1.1 目标地物逐步提取过程
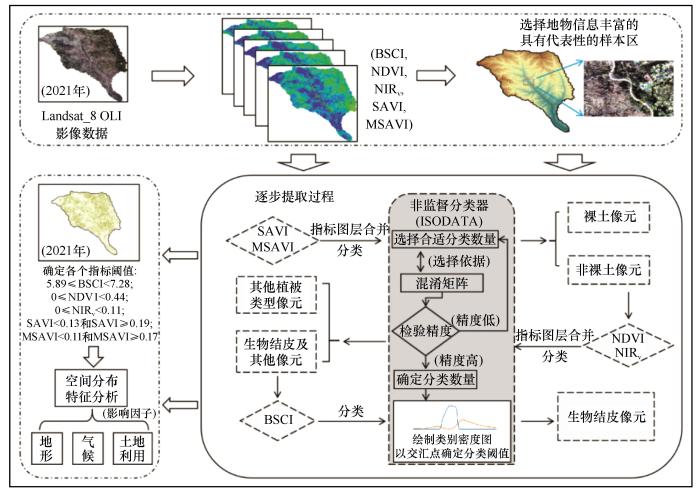
皇甫川流域地表地物类型可分为建筑用地、林地、草地、水域、沙地、裸土、耕地等[25 ] 。目视解译发现地表生物结皮多分布于草地与沙地、裸土,分类的界限难以确立,故本研究通过地物的遥感特征指数来建立目标地物提取规则以确定不同目标地物分割阈值。本研究利用两种土壤调节植被指数和两种植被指数确定土壤、植被指数值的上下限阈值。
由于裸土、植被、结皮三者光谱不同,裸土与植被的光谱差异显著,但与结皮差异较小。因此本研究遵循裸土-植被-结皮顺序,采用逐步提取-逐步验证的方法。该方法主要分3个步骤:①利用土壤调节植被指数(SAVI)与修正土壤调节植被指数(MSAVI)分离区域裸土面积,提取植被、结皮和其他地物(如:裸沙和干枯植物),并利用观测样本进行验证并调整阈值,精度大于95%时的阈值作为分割阈值;②进一步利用归一化差值植被指数(NDVI)、新型光合植被指数(NIRV )分离植被,并利用观测样本进行验证并调整阈值,精度大于95%时的阈值作为分割阈值;③最后通过生物结皮指数(BSCI)提取皇甫川流域生物结皮,实现皇甫川流域生物结皮空间分布制图。具体过程如图2 所示。
图2
图2
研究流程
注: BSCI:生物土壤结皮指数;NDVI:归一化差值植被指数;NIRV :新型植被指数(归一化差值植被指数与近红外波段反射率的乘积);SAVI:土壤调节植被指数;MSAVI:修正型土壤调节植被指数;ISODATA:迭代自组织数据分析算法
Fig.2
Research Flowchart
2.1.2 逐步提取过程中所选择的遥感指数
土壤调节植被指数(SAVI)是在NDVI指数的基础上增加了土壤调节系数L ,取值范围0~1,建立可适当描述土壤-植被系统的简单模型。而修正土壤调节植被指数(MSAVI)适当减小了SAVI中的裸土影响[26 ] 。上述指数考虑植被提取时在土壤背景下的影响,因此本研究以这两个指数结合分类时裸土分布特征,间接识别并提取裸土像元。
S A V I = R n i r - R r e d R n i r + R r e d + L 1 × 1 + L 1 (1)
M S A V I = 2 × R n i r + 1 - 2 × R n i r + 1 2 - 8 × R n i r - R r e d 2 (2)
式中:R red 、R nir 分别代表Landsat 8 OLI数据的红光和近红外波段的反射率;L 1 为常量,L 1 值随绿色植被的数量或覆盖度而变化:在植被非常高的地区L 1 =0,在没有绿色植被的地区L 1 =1。通常L 1 =0.5在大多数情况下效果很好,并且是使用的默认值,本研究取L 1 =0.5。
归一化差值植被指数(NDVI)是检测植被生长状态、植被覆盖度重要遥感指数[27 ] 。在低密度植被覆盖区域,NDVI具有非常敏感的观测效果[26 ] 。新型光合植被指数(NIRV )指的是归一化植被指数与近红外波段反射率的乘积,与NDVI相比,NIRV 可以在高生物量区域使用,有效减轻了植被-土壤混合像元的影响[28 ] 。本研究选取以上两种遥感指数对皇甫川流域的植被地物进行提取,两者在不同覆盖度植被提取上互补缺点,一定程度上减少了皇甫川流域植被的提取误差。
N D V I = R n i r - R r e d R n i r + R r e d (3)
N I R v = R n i r - R r e d R n i r + R r e d × R n i r (4)
式中:R red 、R nir 分别代表Landsat8 OLI数据的红光和近红外波段的反射率。
生物结皮指数(BSCI)是对生物结皮光谱特征的增强,使得与裸沙和干枯植物等背景地物存在较高的背景反射值。按照BSCI的定义并考虑卫星传感器的空间分辨率,一般认为BSCI值主要取决于像元内生物结皮的覆盖率。生物结皮覆盖率增加,BSCI值也增加。与之相反,背景地物的覆盖率增加,则BSCI值降低[14 ] 。本研究利用BSCI值的分布密度曲线与其他地物分布密度曲线以确定生物结皮BSCI值的上下限阈值。
B S C I = 1 - L 2 × R g r e e n - R r e d R m e a n (5)
式中:R green 、R red 分别代表Landsat8 OLI传感器的绿光和红光波段的反射率;R mean 代表绿光、红光、近红外波段反射率的平均值;L 2 为经验调整参数,用于强调红波段和绿波段差的绝对值的贡献率,本研究设定为2。
2.1.3 逐步提取方法中不同阶段目标地物选择
生物结皮与其他地物之间存在混合像元,严重影响生物结皮提取的训练样本的选择。因此,本研究采用非监督分类方式进行提取,依据地物的遥感特征指数进行叠加分类,此外,还将逐步对裸土、植被等像元进行提取分离,逐步提取目标地物样本点验证不同阶段的分类精度,逐步提取方法针对与生物结皮相似的裸土、植被像元等进行单独提取分离,在一定程度上提高了生物结皮的提取精度。
非监督分类(ISODATA)法能够根据影像数据的统计特征将像元聚类成不同的类别,从而实现地物的自动分类。因此,本研究对各个像元依靠地物光谱反射率所计算的各种特征遥感指数值进行分类。为实现影像的准确分类,本研究利用ENVI软件进行分类处理。在分类的过程中,将设定最小像元数量为20,最小迭代次数为10,其他参数将采用默认设置。通过不断调整原始期望类别数量参数,努力寻找到最佳的分类效果,以确保对目标地物的准确提取。
2.1.4 目标地物遥感指数阈值划分
生物结皮指数的阈值遥感确定是提取过程中的关键;张元明等[14 ] 通过野外测量的生物结皮的高光谱数据确定BSCI的阈值上下限,但是该值存在显著的时空差异。Guo等[29 ] 在提取黄土高原小水体时设定了一定的水体提取规则从而确定提取水体的阈值方法,该方法能够实现阈值动态调整,具有较好的时空适宜性。因此,本研究参考水体提取规则设定适用于裸土、植被、生物结皮的提取规则,即对于SAVI、MSAVI指数,裸土信号值(分布密度)大于非裸土信号值的像元判定为裸土,反之则判定为非裸土;对于NDVI、NIRV 指数,植被信号值(分布密度)大于非植被信号值的像元判定为植被,反之则判定为非植被;对于BSCI指数,生物结皮信号值(分布密度)大于非生物结皮信号值的像元判定为生物结皮,反之则判定为非生物结皮。通过以上提取规则的确定,逐步提取分离目标地物,最终得到生物结皮的上下限阈值。
2.2 基于逐步提取的目标地物数据精度验证方法
本研究采用分层抽样和整群抽样相结合的方式选取代表区域内地物样本点,即在逐步提取并分离目标地物的过程中,按照真彩色影像中各类地物分布特征选取目标地物样本。使用皇甫川流域吉林一号亚米级数据集进行地物样本点的提取,共提取了526个裸土样本点,224个非裸土样本点,269个植被样本点,159个非植被样本点,210个生物结皮样本点,119个非生物结皮样本点,其中裸土、植被、生物结皮均为目标地物样本点。基于样本点的选择,借助混淆矩阵计算分类的总体精度(OA)、生产者精度(PA)、用户精度(UA)等,在保证分类精度的前提下,最终确定分类参数,从而确定了各类目标地物的分类阈值。
O A = n M (6)
P A = n N (7)
U A = n n + k (8)
式中:n 表示正确分类样本数量;N 为该类别样本数量;M 为样本总数;k 为其他类别错分到该类别样本数量。
3 结果与分析
3.1 基于非监督分类的各目标地物阈值划分
为提取皇甫川流域的生物结皮,基于逐步解译的方法,首先确定流域裸土、植被的遥感解译阈值并将其进行组合,然后在ISODATA的基础上逐步分类提取流域裸土、植被的空间分布。不断调整期望类别数量,在进行分类处理后根据总体精度、生产者精度、用户精度等确定最佳期望类别数量参数,随后在上文所提到的提取规则基础上,根据目标地物与非目标地物在密度图中的交会点确定阈值。
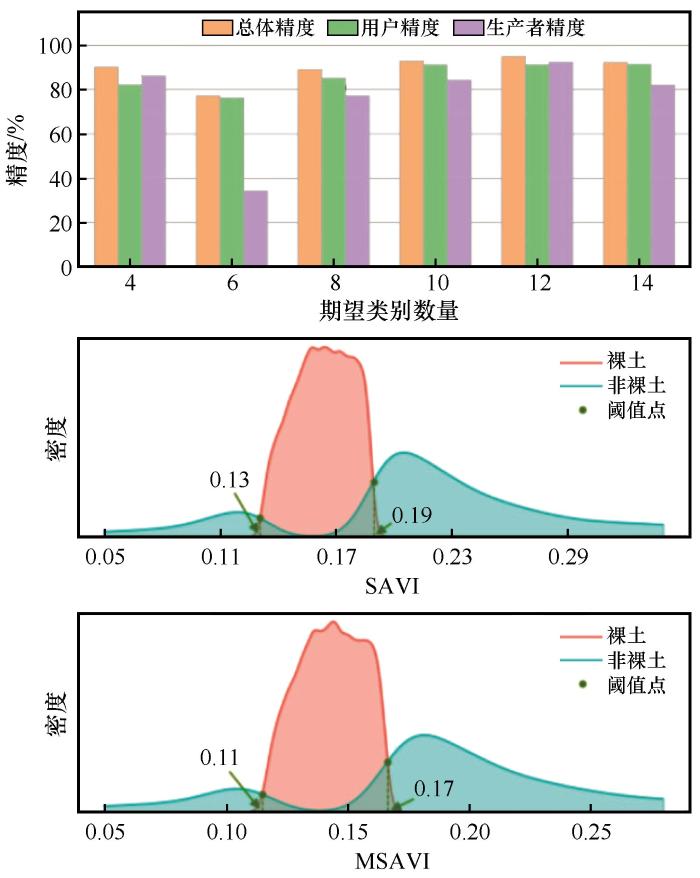
3.1.1 考虑裸土效应的阈值分割与精度评价
为探究裸土与非裸土的区分阈值,本研究借助Landsat 8 OLI数据的近红外与红波段计算了SAVI、MSAVI等指数,用以区分裸土像元与其他像元,并叠加为复合图层进行非监督分类,由于分类数量难以确定,本研究从4类地物开始依次选取复数类别数量作为期望类别数量参数输入,直到某一类别像元数量低于某个适合值时停止分类,目标地物样本点的精度检验结果如图3 所示。当采用非监督分类输入的期望类别数量为12类时,具有较高的精度,故本研究对样本区影像的裸土与非裸土的像元提取使用总体分类精度为95.07%的12类作为期望类别数量参数输入。当以SAVI指数提取时,非裸土区域像元指数阈值为SAVI≥0.19和SAVI<0.13;当以MSAVI指数提取时,非裸土区域像元的指数阈值为MSAVI≥0.17和MSAVI<0.11,此时裸土像元与非裸土像元具有明显的特征差异(图3 )。
图3
图3
提取裸土过程中各期望类别数量结果精度统计及指数阈值分割
Fig.3
Precision statistical chart and exponential threshold segmentation chart for the number of expected categories in the process of extracting bare soil
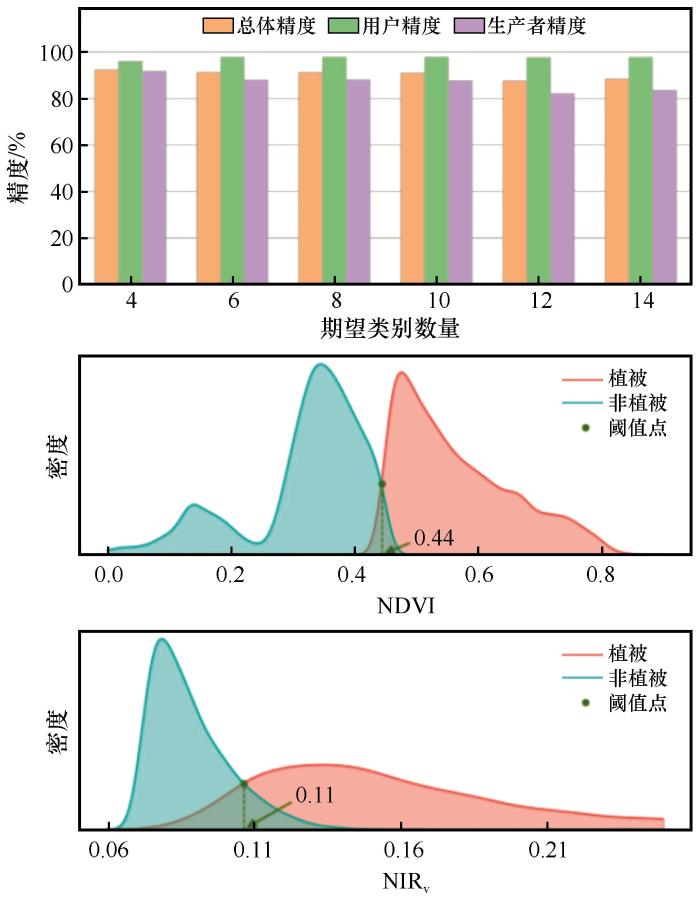
3.1.2 考虑植被效应的阈值分割与精度评价
为探究植被与非植被的区分阈值,按照提取规则在分离裸土像元后的皇甫川流域遥感影像区域进行分析,此部分以NDVI、NIRV 指数提取并分离植被像元,实验过程同裸土提取一致,提取结果如图4 所示。当期望类别数量参数较小时,分类的总体精度、生产者精度、用户精度等精度指标均在一定范围内波动;当期望类别数量参数较大时,精度有所降低。由此可知,非监督分类当参数设置于一定范围时,对于植被地物类型的像元能被很好地识别并提取;故本研究将采用期望类别数量参数为4的参数值进行。结果发现通过植被指数NDVI区分植被与非植被时,非植被区域像元的指数阈值为0≤NDVI<0.44;当以NIRV 指数区分植被与非植被时,非植被区域像元的指数阈值为0≤NIRV <0.11,此时植被像元与非植被像元具有较为明显的特征差异。
图4
图4
提取植被过程中各期望类别数量结果精度统计及阈值分割
Fig.4
Precision statistical chart and exponential threshold segmentation chart for the number of expected categories in the process of extracting vegetation
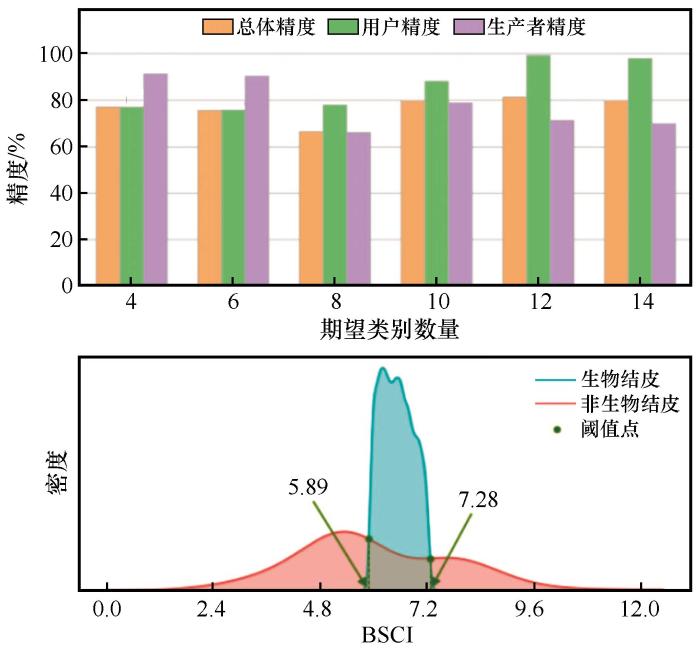
3.1.3 生物结皮提取阈值设置与精度评价
为获得生物结皮与非生物结皮(主要为不透水面区域以及部分其他地物类型混合区域)的区分阈值,按照提取规则在分离裸土、植被像元后的皇甫川流域遥感影像区域进行分析,此部分以BSCI指数提取生物结皮像元,实验过程同裸土、植被提取过程。提取总体分类效果较差于裸土、植被的提取分离精度(图5 )。最好的总体分类精度仅有12类分类时的81.46%。因此,本节以12类为期望类别参数进行分类提取生物结皮阈值。以BSCI指数提取生物结皮像元的指数阈值为5.89≤BSCI<7.28。因此,本研究获得了以Landsat 8 OLI第2、3、4、5波段数据为基础进行分析得到的皇甫川流域生物结皮遥感提取的指数阈值:SAVI<0.13和SAVI≥0.19,MSAVI<0.11和MSAVI≥0.17;0≤NDVI<0.44,0≤NIRV <0.11;5.89≤BSCI<7.28,这为干旱区大尺度生物结皮遥感制图奠定基础。
图5
图5
提取生物结皮过程中各期望类别数量结果精度统计及阈值分割
Fig.5
Precision statistical chart and exponential threshold segmentation chart for the number of expected categories in the process of extracting bio-crust
3.2 皇甫川流域生物结皮空间分布特征
利用本文计算的多植被指数阈值,通过阈值逐步提取过程实现了典型干旱半干旱区皇甫川流域生物结皮空间分布(图6 )。皇甫川流域共有生物结皮904.84 km2 ,占流域总面积的21.51%,说明生物结皮在该流域中广泛分布。进一步研究发现,在整个流域中生物结皮主要呈现破碎化分布形式。这意味着生物结皮并非连续一体地分布在土地表面,而是以分散、不规则的方式存在于不同的区域。
图6
图6
生物结皮空间分布
Fig.6
The spatial distribution of bio-crust
本研究4个典型区域如图7 所示。A区生物结皮主要分布于山区,少量分布于人类生活居住区域;B区域中生物结皮基本分布于南坡坡面或坡谷;C区域生物结皮多分布于河谷两侧植被与裸土、水域等地物类型的交汇边缘;D区域地形起伏较大,生物结皮基本分布于地势平坦开阔的区域。综上所述,生物结皮的空间分布往往需要根据其分布区域的环境因素进行分析,不同的环境因素均将影响生物结皮的空间分布,例如地形、气候、土地利用方式等等。
图7
图7
流域典型样区
注: A、B、C、D分别代表研究区内4个流域典型样区,1、2、3分别表示原始影像、生物结皮在原始影像上的分布情况、生物结皮的空间分布情况
Fig.7
The typical sample areas
3.3 影响因子
3.3.1 地形与降水交互作用对生物结皮空间分布的影响
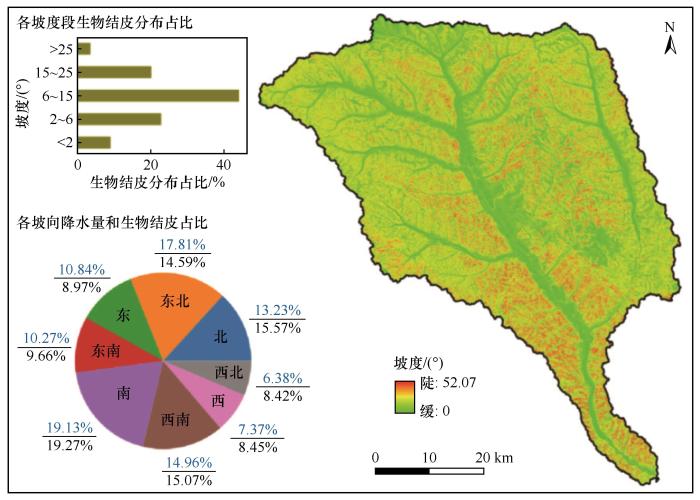
本研究使用皇甫川流域30 m分辨率的数字高程模型(DEM)计算研究区坡度分级图以及坡向图,并统计不同等级坡度以及不同坡向下生物结皮的分布特征(图8 )。不同坡度、坡向的生物结皮分布均存在一定的差异性,其中皇甫川流域的东北坡与南坡方向的生物结皮占比均在17.5%以上,是生物结皮在皇甫川流域分布的主坡向;皇甫川流域中坡度(6°~15°)生物结皮分布显著高于大坡度与小坡度地区,占比超过了40%,说明生物结皮多喜好阴凉潮湿环境。
图8
图8
生物结皮坡度坡向统计分析
注: 横线上为降水量占比,横线下为生物结皮占比
Fig.8
Statistical analysis of bio-crust slope and aspect
本研究进一步分析了不同降水量下坡度、坡向生物结皮的分布情况。在整个皇甫川流域的降水中,南坡占比达到了最高的19.27%,北坡占比达到了15.57%,南坡与北坡的降水量均显著高于其他坡向区域,而南坡与北坡的降水量之间的差异显著小于与其他坡向之间的差异。结果表明,皇甫川流域各个坡向生物结皮空间分布的趋势与降水量分布的趋势基本一致,两者呈较大相关。
3.3.2 土地利用方式对生物结皮空间分布的影响
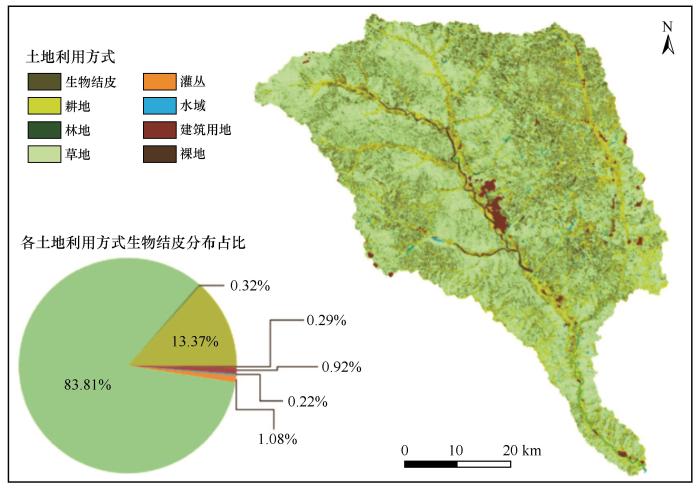
土地利用方式是生物结皮形成和分布的关键影响因素,本研究使用皇甫川流域2021年土地利用方式数据进行统计分析,主要包括不透水面(建筑用地、道路等)、裸地、水域、林地、草地、灌木丛、耕地等土地利用方式,并着重分析林地、草地、灌丛以及耕地对生物结皮分布的影响。结果如图9 所示,总体来看草地与耕地是皇甫川流域生物结皮(97.18%)分布的主要用地类型,而林地与灌丛分布的生物结皮占比(1.39%)略高于其他土地利用方式区域,而水域、不透水面、裸地等区域的生物结皮分布占比均低于1%。因此,草地、耕地是皇甫川流域生物结皮主要分布的土地利用类型。
图9
图9
生物结皮土地利用方式统计分析
Fig.9
Statistical analysis of bio-crust land use types
4 讨论
4.1 逐步提取方法的不确定性
生物结皮的光自养表层群落对土壤稳定以及区域和全球尺度上的水分、养分和微量气体循环至关重要[30 ] ,但是目前空间数据较为缺乏,仅有全球一景的粗分辨率(0.5°)的空间数据[30 ] ,缺乏区域尺度精细分辨率的遥感生物结皮提取方法。
本研究结合野外调查和遥感机制分析,开发了小流域生物结皮逐步提取方法,该方法本质上是依靠地物光谱反射率所计算的各种特征遥感指标之间聚合与分离的生物结皮遥感提取方法,该方法相比较光谱分析法[11 -13 ] 、指数法[14 -17 ] 等取得了较大进步。在考虑流域地表地物反射特性、辐射传输特性的基础上,利用逐步提取的方式针对与生物结皮相似的裸土、植被像元等进行单独提取分离,在一定程度上提高了生物结皮的提取精度。
但该方法也存在一定的局限性。皇甫川流域地形起伏较大且多山,遥感影像中存在的山体阴影限制了地表参数遥感提取的准确性[31 ] ,在逐步提取的过程中,仍然有将山体阴影误认为目标地物的情况,从而影响了目标地物提取与分离的精度与准确性。此外,在遥感影像区分绿色植被与生物结皮、裸土与生物结皮时可能有误分现象,由于不同地物的斑块边缘存在大量混合像元[32 ] ,同时,生物结皮存在厚度、类型差异,使得存在大量的异物同谱现象普遍[33 ] ,这可能导致在多指数阈值分割中本研究以提取规则建立的判别标准错分目标地物的现象。
4.2 干旱半干旱区典型流域结皮时空变化机制
生物结皮是全球干旱半干旱生态系统普遍存在的生物地被物,约占陆地总面积的40%,是该生态系统最为活跃的土壤关键层[34 ] ,生物结皮对干旱等逆境有天然的适应能力,具有重要的生态功能和环境效应[4 ] 。皇甫川流域是干旱半干旱区黄土高原典型流域,本研究基于精细化的生物结皮数据,发现该流域生物结皮基本呈现破碎化分布,空间分布显著受降水、地形、土地利用等影响。
降水、气温与地形对生物结皮存在交互影响;生物结皮生长范围广泛,主要类型包括苔藓、地衣、藻类植物等[2 ] ,生物结皮多喜好阴凉潮湿环境。因此,生物结皮对降水的变化反应极其敏感,在干旱半干旱区随着气温的升高和夏季降水频率的增多,生物结皮的生物多样性急剧下降[35 ] 。全球气候变化导致美国麦哈韦沙漠和科罗拉多州高原夏季降水增多,而夏季降水增多,促使了某些种类的生物结皮盖度下降,生理功能改变。大气温度是影响结皮形成、空间分布特征的另一个关键因子,学者通过对最近22年来的329种地衣的物种组成变化和分布变化研究,发现随全球变暖喜温生物结皮在荷兰的种类增多而耐寒地衣种减少[36 ] 。同时,不同地形因坡度和坡向的差异显著影响,降水和温度的分布格局、阴坡气温高于阳坡、不同海拔间降水存在显著差异,这些气候、地形等环境因素显著影响生物结皮的空间分布[15 ,37 ] 。此外,流域土地利用方式也影响着生物结皮的形成与分布,这种差异主要体现在林草地以及其他土地利用方式之间,草地生物结皮盖度与林地生物结皮盖度存在一定的差异性[38 ] ,而林草地生物结皮的分布与其他土地利用类型生物结皮的分布也存在着一定的差异性。
结皮的形成与降水、气温与地形等环境因素关系密切,皇甫川流域属典型干旱半干旱的风水复合侵蚀区域,生物结皮呈现出独特特征。风沙是塑造其地表形态的关键因素,风速常达3.2 m·s-1 ,在春季尤为显著,而生物结皮可有效抵御风沙侵蚀,藻类、地衣和苔藓的分泌物与假根能稳固沙粒,减少风沙对地表的破坏。降水方面,年降水约400 mm,且多在7—9月,降水强度大、历时短。生物结皮在降雨时可增加土壤入渗,减少地表径流与水土流失。当降水充沛时,有利于生物结皮中各类生物生长繁殖,加速其演替进程;但暴雨可能冲毁部分结皮。皇甫川流域昼夜温差大,利于生物结皮中微生物活动与物质分解。在温暖季节,生物结皮生长活跃,从微生物结皮向苔藓结皮逐渐演化,结构与功能愈发完善,增强对风水复合侵蚀的防控作用。
5 结论
本研究以皇甫川流域作为研究区,建立了以生物结皮为对象的基于非监督分类的逐步提取方法,方法中使用了SAVI、MSAVI、NDVI、NIRV 、BSCI等遥感指数,通过多指数阈值分割建立的提取规则提取分离各个目标地物以提取研究区的生物结皮,并结合流域内地面高程数据、土地利用方式、降水量数据进一步分析了生物结皮空间分布的影响因子。
基于土壤调节植被指数(SAVI)、修正土壤调节植被指数(MSAVI)、归一化差值植被指数(NDVI)、新型光合植被指数(NIRV )、生物结皮指数(BSCI)构建的干旱区生物结皮逐步提取模型,不同植被指数间的阈值存在显著差异(SAVI<0.13和SAVI≥0.19;MSAVI<0.11和MSAVI≥0.17;0≤NDVI<0.44;0≤NIRV <0.11;5.89≤BSCI<7.28),但该模型提取干旱区生物结皮的总体精度可靠。
在皇甫川流域中共提取生物结皮面积904.84 km2 ,占皇甫川流域总面积的21.51%。在整个流域中,从分布趋势来看,生物结皮以破碎化的形式分布为主,且主要分布于山区,少量分布于人类生活涉及区域;从地貌类型来看,山体南坡坡面或坡谷地势较为平坦地区是皇甫川流域生物结皮主要分布的区域;从分布特点来看,生物结皮在河谷两侧植被与裸土、水域等地物类型的交汇边缘区域分布较多。
皇甫川流域阴坡、阳坡均有一定占比的生物结皮分布且两者无较大差异;生物结皮基本分布于草地、耕地等土地利用方式地区;生物结皮空间分布与研究区内各区域降水量之间存在较高的相关性,降水量越大,生物结皮的分布就越多,这表明降水是影响生物结皮分布的重要因素之一,高降水量有利于生物结皮的生长和发展。总之,皇甫川流域的生物结皮分布受到多种因素的影响,包括土地利用方式、降水量、地形等。
本研究重点在于考虑地表覆盖的复杂性、时空异质性,利用多个遥感指数协同组合的方法,构建基于逐步提取的干旱区生物结皮提取的遥感模型。该方法采用逐步识别模型,以此提取裸土、植被及生物结皮像元,发展了典型干旱区皇甫川流域30 m分辨率的生物结皮空间分布数据,并探讨了地形、气候以及土地利用方式对生物结皮空间分布的影响。未来将耦合辐射传输模型,考虑裸土、植被及生物结皮的辐射传输过程,发展物理机制更加明确的干旱区生物结皮提取的遥感模型。
参考文献
View Option
[1]
吴玉环 ,高谦 ,程国栋 生物土壤结皮的生态功能
[J].生态学杂志 ,2002 ,21 (4 ):41 -45 .
[本文引用: 1]
[2]
张丙昌 ,武志芳 ,李彬 黄土高原生物土壤结皮研究进展与展望
[J].土壤学报 ,2021 ,58 (5 ):1123 -1131 .
[本文引用: 2]
[3]
张雨虹 ,张韶阳 ,张树煇 ,等 毛乌素沙地苔藓结皮对沙化土壤性质和细菌群落的影响
[J].土壤学报 ,2021 ,58 (6 ):1585 -1597 .
[本文引用: 1]
[4]
李新荣 ,张志山 ,王新平 ,等 干旱区土壤植被系统恢复的生态水文学研究进展
[J].中国沙漠 ,2009 ,29 (5 ):845 -852 .
[本文引用: 2]
[5]
杨晓晖 ,张克斌 ,赵云杰 生物土壤结皮:荒漠化地区研究的热点问题
[J].生态学报 ,2001 ,21 (3 ):474 -480 .
[本文引用: 2]
[6]
李新荣 ,张元明 ,赵允格 生物土壤结皮研究:进展、前沿与展望
[J].地球科学进展 ,2009 ,24 (1 ):11 -24 .
[本文引用: 1]
[7]
肖波 ,赵允格 ,邵明安 陕北水蚀风蚀交错区两种生物结皮对土壤理化性质的影响
[J].生态学报 ,2007 ,27 (11 ):4662 -4670 .
[本文引用: 1]
[8]
高丽倩 ,赵允格 ,秦宁强 ,等 黄土丘陵区生物结皮对土壤可蚀性的影响
[J].应用生态学报 ,2013 ,24 (1 ):105 -112 .
[9]
高丽倩 ,赵允格 ,秦宁强 ,等 黄土丘陵区生物结皮对土壤物理属性的影响
[J].自然资源学报 ,2012 ,27 (8 ):1316 -1326 .
[10]
陈瑶 ,张科利 ,罗利芳 ,等 黄土坡耕地弃耕后土壤入渗变化规律及影响因素
[J].泥沙研究 ,2005 (5 ):45 -50 .
[本文引用: 1]
[11]
田园盛 ,张玥 ,孙文义 ,等 黄土高原水蚀风蚀交错区不同类型生物土壤结皮光谱特征
[J].光谱学与光谱分析 ,2018 ,38 (7 ):2215 -2220 .
[本文引用: 4]
[12]
宋丽丽 ,白中科 ,樊翔 ,等 生物土壤结皮对照相法测量植被覆盖度结果的影响
[J].生态学报 ,2018 ,38 (4 ):1272 -1283 .
[本文引用: 1]
[13]
Baxter C Mallen-Cooper M Lyons M B et al Measuring reflectance of tiny organisms:the promise of species level biocrust remote sensing
[J].Methods in Ecology and Evolution ,2021 ,12 (11 ):2174 -2183 .
[本文引用: 4]
[14]
张元明 ,陈晋 ,王雪芹 ,等 古尔班通古特沙漠生物结皮的分布特征
[J].地理学报 ,2005 ,60 (1 ):53 -60 .
[本文引用: 5]
[15]
张元明 ,潘惠霞 ,潘伯荣 古尔班通古特沙漠不同地貌部位生物结皮的选择性分布
[J].水土保持学报 ,2004 ,18 (4 ):61 -64 .
[本文引用: 1]
[16]
杨伟 ,陈晋 ,张元明 ,等 古尔班通古特沙漠1970-2000年代生物结皮覆盖变化研究
[J].自然资源学报 ,2006 ,21 (6 ):934 -941 .
[本文引用: 1]
[17]
冯秀绒 ,卜崇峰 ,郝红科 ,等 基于光谱分析的生物结皮提取研究:以毛乌素沙地为例
[J].自然资源学报 ,2015 ,30 (6 ):1024 -1034 .
[本文引用: 4]
[18]
杨丽娜 ,赵允格 ,明姣 ,等 黄土高原不同侵蚀类型区生物结皮中蓝藻的多样性
[J].生态学报 ,2013 ,33 (14 ):4416 -4424 .
[本文引用: 1]
[19]
赵河聚 ,岳艳鹏 ,贾晓红 ,等 模拟增温对高寒沙区生物土壤结皮-土壤系统呼吸的影响
[J].植物生态学报 ,2020 ,44 (9 ):916 -925 .
[本文引用: 1]
[20]
Karnieli A Development and implementation of spectral crust index over dune sands
[J].International Journal of Remote Sensing ,1997 ,18 (6 ):1207 -1220 .
[本文引用: 2]
[21]
Sun H Ma X Liu Y et al A New multiangle method for estimating fractional biocrust coverage from sentinel-2 data in arid areas
[J].Ieee Transactions on Geoscience and Remote Sensing ,2024 ,62 :1 -15 .
[本文引用: 2]
[22]
Chen J Yuan Z M Wang L et al A new index for mapping lichen-dominated biological soil crusts in desert areas
[J].Remote Sensing of Environment ,2005 ,96 (2 ):165 -175 .
[本文引用: 1]
[23]
李双双 ,延军平 ,万佳 近10年陕甘宁黄土高原区植被覆盖时空变化特征
[J].地理学报 ,2012 ,67 (7 ):960 -970 .
[本文引用: 1]
[24]
于红博 ,杨劼 ,陈杰 皇甫川流域地表特征参数遥感反演研究
[J].人民黄河 ,2008 ,30 (4 ):5 -6 .
[本文引用: 1]
[25]
喻锋 ,李晓兵 ,王宏 生态安全条件下土地利用格局优化:以皇甫川流域为例
[J].生态学报 ,2014 ,34 (12 ):3198 -3210 .
[本文引用: 1]
[26]
田庆久 ,闵祥军 植被指数研究进展
[J].地球科学进展 ,1998 ,13 (4 ):327 -333 .
[本文引用: 2]
[27]
王正兴 ,刘闯 , Alfredo H 植被指数研究进展:从AVHRR-NDVI到MODIS-EVI
[J].生态学报 ,2003 ,23 (5 ):979 -987 .
[本文引用: 1]
[28]
王敏钰 ,罗毅 ,张正阳 ,等 植被物候参数遥感提取与验证方法研究进展
[J].遥感学报 ,2022 ,26 (3 ):431 -455 .
[本文引用: 1]
[29]
Guo J Wang X Liu B et al Remote-sensing extraction of small water bodies on the Loess Plateau
[J].Water-Sui ,2023 ,15 (5 ):866 .
[本文引用: 1]
[30]
Rodriguez-Caballero E Belnap J Büdel B et al Dryland photoautotrophic soil surface communities endangered byglobal change
[J].Nature Geoscience ,2018 ,11 (3 ):185 -189 .
[本文引用: 2]
[31]
范德芹 ,赵学胜 ,朱文泉 ,等 植物物候遥感监测精度影响因素研究综述
[J].地理科学进展 ,2016 ,35 (3 ):304 -319 .
[本文引用: 1]
[32]
李素 ,李文正 ,周建军 ,等 遥感影像混合像元分解中的端元选择方法综述
[J].地理与地理信息科学 ,2007 ,23 (5 ):35 -38 .
[本文引用: 1]
[33]
陈珂 ,彭志平 ,柯文德 遥感图像异物同谱干扰消除技术研究与仿真
[J].计算机仿真 ,2013 ,30 (1 ):423 -426 .
[本文引用: 1]
[34]
韩炳宏 ,牛得草 ,贺磊 ,等 生物土壤结皮发育及其影响因素研究进展
[J].草业科学 ,2017 ,34 (9 ):1793 -1801 .
[本文引用: 1]
[35]
房世波 ,冯凌 ,刘华杰 ,等 生物土壤结皮对全球气候变化的响应
[J].生态学报 ,2008 ,28 (7 ):3312 -3321 .
[本文引用: 1]
[36]
van Herk C M Aptroot A van Dobben H F Long-term monitoring in the Netherlands suggests that lichens respond to global warming
[J].The Lichenologist ,2002 ,34 (2 ):141 -154 .
[本文引用: 1]
[37]
焦雯珺 ,朱清科 ,张宇清 ,等 陕北黄土区退耕还林地生物结皮分布及其影响因子研究
[J].北京林业大学学报 ,2007 ,29 (1 ):102 -107 .
[本文引用: 1]
[38]
赵允格 ,吉静怡 ,张万涛 ,等 黄土高原生物土壤结皮分布时空分异特征
[J].应用生态学报 ,2024 ,35 (3 ):739 -748 .
[本文引用: 1]
生物土壤结皮的生态功能
1
2002
... 生物结皮是细菌、藻类、真菌和孢子植物与土壤颗粒胶结而成的有机复合体[1 ] ,主要分布于沙漠或荒漠等干旱半干旱区,对干旱半干旱地区的水土保护、养分积累和生态恢复具有重要的生态意义[2 ] .全球气候变化加剧,土地退化日益严重,而生物结皮作为土壤生态系统中的关键组成部分,在干旱半干旱区扮演关键角色,生物结皮通过固定沙漠表面的土壤,防止风蚀和水土流失,维持着土壤生态系统的稳定[3 ] .生物结皮显著影响区域生态水文循环,直接或间接参与降水入渗、凝结水捕获和蒸发等生态水文过程[4 -5 ] .干旱半干旱区生态水文循环中,生物结皮不仅能够根据生物特性对当地土壤特性起到一定的改善作用[6 ] ,从而维持流域生态平衡,并能够在生态水文循环的较大尺度上融入气候、土壤、高等植物作为整体考虑.研究生物结皮在生态水文循环中水分流动与转换的规律[5 ] ,有助于系统全面地揭示其在生态水文循环过程中所扮演的角色,但是目前由于缺乏生态水文循环中生物结皮的大尺度、长时序数据,生态水文研究对结皮考虑不足,因此如何发展大尺度、高频次生物结皮数据研发是干旱半干旱地区生物结皮研究的关键. ...
黄土高原生物土壤结皮研究进展与展望
2
2021
... 生物结皮是细菌、藻类、真菌和孢子植物与土壤颗粒胶结而成的有机复合体[1 ] ,主要分布于沙漠或荒漠等干旱半干旱区,对干旱半干旱地区的水土保护、养分积累和生态恢复具有重要的生态意义[2 ] .全球气候变化加剧,土地退化日益严重,而生物结皮作为土壤生态系统中的关键组成部分,在干旱半干旱区扮演关键角色,生物结皮通过固定沙漠表面的土壤,防止风蚀和水土流失,维持着土壤生态系统的稳定[3 ] .生物结皮显著影响区域生态水文循环,直接或间接参与降水入渗、凝结水捕获和蒸发等生态水文过程[4 -5 ] .干旱半干旱区生态水文循环中,生物结皮不仅能够根据生物特性对当地土壤特性起到一定的改善作用[6 ] ,从而维持流域生态平衡,并能够在生态水文循环的较大尺度上融入气候、土壤、高等植物作为整体考虑.研究生物结皮在生态水文循环中水分流动与转换的规律[5 ] ,有助于系统全面地揭示其在生态水文循环过程中所扮演的角色,但是目前由于缺乏生态水文循环中生物结皮的大尺度、长时序数据,生态水文研究对结皮考虑不足,因此如何发展大尺度、高频次生物结皮数据研发是干旱半干旱地区生物结皮研究的关键. ...
... 降水、气温与地形对生物结皮存在交互影响;生物结皮生长范围广泛,主要类型包括苔藓、地衣、藻类植物等[2 ] ,生物结皮多喜好阴凉潮湿环境.因此,生物结皮对降水的变化反应极其敏感,在干旱半干旱区随着气温的升高和夏季降水频率的增多,生物结皮的生物多样性急剧下降[35 ] .全球气候变化导致美国麦哈韦沙漠和科罗拉多州高原夏季降水增多,而夏季降水增多,促使了某些种类的生物结皮盖度下降,生理功能改变.大气温度是影响结皮形成、空间分布特征的另一个关键因子,学者通过对最近22年来的329种地衣的物种组成变化和分布变化研究,发现随全球变暖喜温生物结皮在荷兰的种类增多而耐寒地衣种减少[36 ] .同时,不同地形因坡度和坡向的差异显著影响,降水和温度的分布格局、阴坡气温高于阳坡、不同海拔间降水存在显著差异,这些气候、地形等环境因素显著影响生物结皮的空间分布[15 ,37 ] .此外,流域土地利用方式也影响着生物结皮的形成与分布,这种差异主要体现在林草地以及其他土地利用方式之间,草地生物结皮盖度与林地生物结皮盖度存在一定的差异性[38 ] ,而林草地生物结皮的分布与其他土地利用类型生物结皮的分布也存在着一定的差异性. ...
毛乌素沙地苔藓结皮对沙化土壤性质和细菌群落的影响
1
2021
... 生物结皮是细菌、藻类、真菌和孢子植物与土壤颗粒胶结而成的有机复合体[1 ] ,主要分布于沙漠或荒漠等干旱半干旱区,对干旱半干旱地区的水土保护、养分积累和生态恢复具有重要的生态意义[2 ] .全球气候变化加剧,土地退化日益严重,而生物结皮作为土壤生态系统中的关键组成部分,在干旱半干旱区扮演关键角色,生物结皮通过固定沙漠表面的土壤,防止风蚀和水土流失,维持着土壤生态系统的稳定[3 ] .生物结皮显著影响区域生态水文循环,直接或间接参与降水入渗、凝结水捕获和蒸发等生态水文过程[4 -5 ] .干旱半干旱区生态水文循环中,生物结皮不仅能够根据生物特性对当地土壤特性起到一定的改善作用[6 ] ,从而维持流域生态平衡,并能够在生态水文循环的较大尺度上融入气候、土壤、高等植物作为整体考虑.研究生物结皮在生态水文循环中水分流动与转换的规律[5 ] ,有助于系统全面地揭示其在生态水文循环过程中所扮演的角色,但是目前由于缺乏生态水文循环中生物结皮的大尺度、长时序数据,生态水文研究对结皮考虑不足,因此如何发展大尺度、高频次生物结皮数据研发是干旱半干旱地区生物结皮研究的关键. ...
干旱区土壤植被系统恢复的生态水文学研究进展
2
2009
... 生物结皮是细菌、藻类、真菌和孢子植物与土壤颗粒胶结而成的有机复合体[1 ] ,主要分布于沙漠或荒漠等干旱半干旱区,对干旱半干旱地区的水土保护、养分积累和生态恢复具有重要的生态意义[2 ] .全球气候变化加剧,土地退化日益严重,而生物结皮作为土壤生态系统中的关键组成部分,在干旱半干旱区扮演关键角色,生物结皮通过固定沙漠表面的土壤,防止风蚀和水土流失,维持着土壤生态系统的稳定[3 ] .生物结皮显著影响区域生态水文循环,直接或间接参与降水入渗、凝结水捕获和蒸发等生态水文过程[4 -5 ] .干旱半干旱区生态水文循环中,生物结皮不仅能够根据生物特性对当地土壤特性起到一定的改善作用[6 ] ,从而维持流域生态平衡,并能够在生态水文循环的较大尺度上融入气候、土壤、高等植物作为整体考虑.研究生物结皮在生态水文循环中水分流动与转换的规律[5 ] ,有助于系统全面地揭示其在生态水文循环过程中所扮演的角色,但是目前由于缺乏生态水文循环中生物结皮的大尺度、长时序数据,生态水文研究对结皮考虑不足,因此如何发展大尺度、高频次生物结皮数据研发是干旱半干旱地区生物结皮研究的关键. ...
... 生物结皮是全球干旱半干旱生态系统普遍存在的生物地被物,约占陆地总面积的40%,是该生态系统最为活跃的土壤关键层[34 ] ,生物结皮对干旱等逆境有天然的适应能力,具有重要的生态功能和环境效应[4 ] .皇甫川流域是干旱半干旱区黄土高原典型流域,本研究基于精细化的生物结皮数据,发现该流域生物结皮基本呈现破碎化分布,空间分布显著受降水、地形、土地利用等影响. ...
生物土壤结皮:荒漠化地区研究的热点问题
2
2001
... 生物结皮是细菌、藻类、真菌和孢子植物与土壤颗粒胶结而成的有机复合体[1 ] ,主要分布于沙漠或荒漠等干旱半干旱区,对干旱半干旱地区的水土保护、养分积累和生态恢复具有重要的生态意义[2 ] .全球气候变化加剧,土地退化日益严重,而生物结皮作为土壤生态系统中的关键组成部分,在干旱半干旱区扮演关键角色,生物结皮通过固定沙漠表面的土壤,防止风蚀和水土流失,维持着土壤生态系统的稳定[3 ] .生物结皮显著影响区域生态水文循环,直接或间接参与降水入渗、凝结水捕获和蒸发等生态水文过程[4 -5 ] .干旱半干旱区生态水文循环中,生物结皮不仅能够根据生物特性对当地土壤特性起到一定的改善作用[6 ] ,从而维持流域生态平衡,并能够在生态水文循环的较大尺度上融入气候、土壤、高等植物作为整体考虑.研究生物结皮在生态水文循环中水分流动与转换的规律[5 ] ,有助于系统全面地揭示其在生态水文循环过程中所扮演的角色,但是目前由于缺乏生态水文循环中生物结皮的大尺度、长时序数据,生态水文研究对结皮考虑不足,因此如何发展大尺度、高频次生物结皮数据研发是干旱半干旱地区生物结皮研究的关键. ...
... [5 ],有助于系统全面地揭示其在生态水文循环过程中所扮演的角色,但是目前由于缺乏生态水文循环中生物结皮的大尺度、长时序数据,生态水文研究对结皮考虑不足,因此如何发展大尺度、高频次生物结皮数据研发是干旱半干旱地区生物结皮研究的关键. ...
生物土壤结皮研究:进展、前沿与展望
1
2009
... 生物结皮是细菌、藻类、真菌和孢子植物与土壤颗粒胶结而成的有机复合体[1 ] ,主要分布于沙漠或荒漠等干旱半干旱区,对干旱半干旱地区的水土保护、养分积累和生态恢复具有重要的生态意义[2 ] .全球气候变化加剧,土地退化日益严重,而生物结皮作为土壤生态系统中的关键组成部分,在干旱半干旱区扮演关键角色,生物结皮通过固定沙漠表面的土壤,防止风蚀和水土流失,维持着土壤生态系统的稳定[3 ] .生物结皮显著影响区域生态水文循环,直接或间接参与降水入渗、凝结水捕获和蒸发等生态水文过程[4 -5 ] .干旱半干旱区生态水文循环中,生物结皮不仅能够根据生物特性对当地土壤特性起到一定的改善作用[6 ] ,从而维持流域生态平衡,并能够在生态水文循环的较大尺度上融入气候、土壤、高等植物作为整体考虑.研究生物结皮在生态水文循环中水分流动与转换的规律[5 ] ,有助于系统全面地揭示其在生态水文循环过程中所扮演的角色,但是目前由于缺乏生态水文循环中生物结皮的大尺度、长时序数据,生态水文研究对结皮考虑不足,因此如何发展大尺度、高频次生物结皮数据研发是干旱半干旱地区生物结皮研究的关键. ...
陕北水蚀风蚀交错区两种生物结皮对土壤理化性质的影响
1
2007
... 目前,生物结皮在中国北方干旱半干旱区以及南方湿润半湿润区均有大量研究,主要是通过站点观测数据分析结皮对土壤水碳固持等关键过程的影响[7 -10 ] .生物结皮分布的提取方法主要有两种,一种是基于光谱特征的光谱分析法[11 -13 ] ,另外一种是基于遥感数据的波段代数运算构成的指数法[14 -17 ] .在光谱分析法中,主要针对生物结皮的光谱进行分析,以光谱特征之间的差异来区分不同种类生物结皮,这种基于面向对象的提取方法在生物结皮相关提取中作为常识性应用.田园盛等[11 ] 借助光谱特征之间差异以区分藻类、藓类等不同类型生物结皮;宋丽丽等[12 ] 结合反映相片DN值信息的RGB阈值法和ML(最大似然分类)法高精度准确识别出分布于干旱半干旱退化草原区的生物结皮;Baxter等[13 ] 采用机器学习(随机森林)对生物结皮的光谱特征进行评估,以区分不同种类拥有不同结构形状的生物结皮.这些方法需要在站点尺度下的精细观测以确定阈值,但该阈值计算受人为因素或其他环境因素的影响.受野外气候、地貌、侵蚀类型等多方面环境因素的影响[18 -19 ] ,生物结皮的生物特性会发生一定变化,从而导致其光谱特性在不同条件下引起一定变化[11 ] ,生物结皮指数上下限阈值的准确性也会受一定的影响,因此采用生物结皮光谱特征提取方法的稳定性待提高.在实际应用中,由于生物结皮的空间分布复杂性和多样性,采用目视解译的方法易受到人为主观因素的影响,导致提取结果的不稳定性和不一致性.此外,生物结皮的覆盖范围广泛,包括草地、森林、湿地等不同类型的植被,这些生境特征在中低分辨率的遥感影像中难以准确区分,从而给生物结皮提取带来一定的挑战.另外,目视解译提取生物结皮需要大量的人力和时间投入,效率较低,尤其在大范围地区的生物结皮监测和研究中更为困难. ...
黄土丘陵区生物结皮对土壤可蚀性的影响
0
2013
黄土丘陵区生物结皮对土壤物理属性的影响
0
2012
黄土坡耕地弃耕后土壤入渗变化规律及影响因素
1
2005
... 目前,生物结皮在中国北方干旱半干旱区以及南方湿润半湿润区均有大量研究,主要是通过站点观测数据分析结皮对土壤水碳固持等关键过程的影响[7 -10 ] .生物结皮分布的提取方法主要有两种,一种是基于光谱特征的光谱分析法[11 -13 ] ,另外一种是基于遥感数据的波段代数运算构成的指数法[14 -17 ] .在光谱分析法中,主要针对生物结皮的光谱进行分析,以光谱特征之间的差异来区分不同种类生物结皮,这种基于面向对象的提取方法在生物结皮相关提取中作为常识性应用.田园盛等[11 ] 借助光谱特征之间差异以区分藻类、藓类等不同类型生物结皮;宋丽丽等[12 ] 结合反映相片DN值信息的RGB阈值法和ML(最大似然分类)法高精度准确识别出分布于干旱半干旱退化草原区的生物结皮;Baxter等[13 ] 采用机器学习(随机森林)对生物结皮的光谱特征进行评估,以区分不同种类拥有不同结构形状的生物结皮.这些方法需要在站点尺度下的精细观测以确定阈值,但该阈值计算受人为因素或其他环境因素的影响.受野外气候、地貌、侵蚀类型等多方面环境因素的影响[18 -19 ] ,生物结皮的生物特性会发生一定变化,从而导致其光谱特性在不同条件下引起一定变化[11 ] ,生物结皮指数上下限阈值的准确性也会受一定的影响,因此采用生物结皮光谱特征提取方法的稳定性待提高.在实际应用中,由于生物结皮的空间分布复杂性和多样性,采用目视解译的方法易受到人为主观因素的影响,导致提取结果的不稳定性和不一致性.此外,生物结皮的覆盖范围广泛,包括草地、森林、湿地等不同类型的植被,这些生境特征在中低分辨率的遥感影像中难以准确区分,从而给生物结皮提取带来一定的挑战.另外,目视解译提取生物结皮需要大量的人力和时间投入,效率较低,尤其在大范围地区的生物结皮监测和研究中更为困难. ...
黄土高原水蚀风蚀交错区不同类型生物土壤结皮光谱特征
4
2018
... 目前,生物结皮在中国北方干旱半干旱区以及南方湿润半湿润区均有大量研究,主要是通过站点观测数据分析结皮对土壤水碳固持等关键过程的影响[7 -10 ] .生物结皮分布的提取方法主要有两种,一种是基于光谱特征的光谱分析法[11 -13 ] ,另外一种是基于遥感数据的波段代数运算构成的指数法[14 -17 ] .在光谱分析法中,主要针对生物结皮的光谱进行分析,以光谱特征之间的差异来区分不同种类生物结皮,这种基于面向对象的提取方法在生物结皮相关提取中作为常识性应用.田园盛等[11 ] 借助光谱特征之间差异以区分藻类、藓类等不同类型生物结皮;宋丽丽等[12 ] 结合反映相片DN值信息的RGB阈值法和ML(最大似然分类)法高精度准确识别出分布于干旱半干旱退化草原区的生物结皮;Baxter等[13 ] 采用机器学习(随机森林)对生物结皮的光谱特征进行评估,以区分不同种类拥有不同结构形状的生物结皮.这些方法需要在站点尺度下的精细观测以确定阈值,但该阈值计算受人为因素或其他环境因素的影响.受野外气候、地貌、侵蚀类型等多方面环境因素的影响[18 -19 ] ,生物结皮的生物特性会发生一定变化,从而导致其光谱特性在不同条件下引起一定变化[11 ] ,生物结皮指数上下限阈值的准确性也会受一定的影响,因此采用生物结皮光谱特征提取方法的稳定性待提高.在实际应用中,由于生物结皮的空间分布复杂性和多样性,采用目视解译的方法易受到人为主观因素的影响,导致提取结果的不稳定性和不一致性.此外,生物结皮的覆盖范围广泛,包括草地、森林、湿地等不同类型的植被,这些生境特征在中低分辨率的遥感影像中难以准确区分,从而给生物结皮提取带来一定的挑战.另外,目视解译提取生物结皮需要大量的人力和时间投入,效率较低,尤其在大范围地区的生物结皮监测和研究中更为困难. ...
... [11 ]借助光谱特征之间差异以区分藻类、藓类等不同类型生物结皮;宋丽丽等[12 ] 结合反映相片DN值信息的RGB阈值法和ML(最大似然分类)法高精度准确识别出分布于干旱半干旱退化草原区的生物结皮;Baxter等[13 ] 采用机器学习(随机森林)对生物结皮的光谱特征进行评估,以区分不同种类拥有不同结构形状的生物结皮.这些方法需要在站点尺度下的精细观测以确定阈值,但该阈值计算受人为因素或其他环境因素的影响.受野外气候、地貌、侵蚀类型等多方面环境因素的影响[18 -19 ] ,生物结皮的生物特性会发生一定变化,从而导致其光谱特性在不同条件下引起一定变化[11 ] ,生物结皮指数上下限阈值的准确性也会受一定的影响,因此采用生物结皮光谱特征提取方法的稳定性待提高.在实际应用中,由于生物结皮的空间分布复杂性和多样性,采用目视解译的方法易受到人为主观因素的影响,导致提取结果的不稳定性和不一致性.此外,生物结皮的覆盖范围广泛,包括草地、森林、湿地等不同类型的植被,这些生境特征在中低分辨率的遥感影像中难以准确区分,从而给生物结皮提取带来一定的挑战.另外,目视解译提取生物结皮需要大量的人力和时间投入,效率较低,尤其在大范围地区的生物结皮监测和研究中更为困难. ...
... [11 ],生物结皮指数上下限阈值的准确性也会受一定的影响,因此采用生物结皮光谱特征提取方法的稳定性待提高.在实际应用中,由于生物结皮的空间分布复杂性和多样性,采用目视解译的方法易受到人为主观因素的影响,导致提取结果的不稳定性和不一致性.此外,生物结皮的覆盖范围广泛,包括草地、森林、湿地等不同类型的植被,这些生境特征在中低分辨率的遥感影像中难以准确区分,从而给生物结皮提取带来一定的挑战.另外,目视解译提取生物结皮需要大量的人力和时间投入,效率较低,尤其在大范围地区的生物结皮监测和研究中更为困难. ...
... 本研究结合野外调查和遥感机制分析,开发了小流域生物结皮逐步提取方法,该方法本质上是依靠地物光谱反射率所计算的各种特征遥感指标之间聚合与分离的生物结皮遥感提取方法,该方法相比较光谱分析法[11 -13 ] 、指数法[14 -17 ] 等取得了较大进步.在考虑流域地表地物反射特性、辐射传输特性的基础上,利用逐步提取的方式针对与生物结皮相似的裸土、植被像元等进行单独提取分离,在一定程度上提高了生物结皮的提取精度. ...
生物土壤结皮对照相法测量植被覆盖度结果的影响
1
2018
... 目前,生物结皮在中国北方干旱半干旱区以及南方湿润半湿润区均有大量研究,主要是通过站点观测数据分析结皮对土壤水碳固持等关键过程的影响[7 -10 ] .生物结皮分布的提取方法主要有两种,一种是基于光谱特征的光谱分析法[11 -13 ] ,另外一种是基于遥感数据的波段代数运算构成的指数法[14 -17 ] .在光谱分析法中,主要针对生物结皮的光谱进行分析,以光谱特征之间的差异来区分不同种类生物结皮,这种基于面向对象的提取方法在生物结皮相关提取中作为常识性应用.田园盛等[11 ] 借助光谱特征之间差异以区分藻类、藓类等不同类型生物结皮;宋丽丽等[12 ] 结合反映相片DN值信息的RGB阈值法和ML(最大似然分类)法高精度准确识别出分布于干旱半干旱退化草原区的生物结皮;Baxter等[13 ] 采用机器学习(随机森林)对生物结皮的光谱特征进行评估,以区分不同种类拥有不同结构形状的生物结皮.这些方法需要在站点尺度下的精细观测以确定阈值,但该阈值计算受人为因素或其他环境因素的影响.受野外气候、地貌、侵蚀类型等多方面环境因素的影响[18 -19 ] ,生物结皮的生物特性会发生一定变化,从而导致其光谱特性在不同条件下引起一定变化[11 ] ,生物结皮指数上下限阈值的准确性也会受一定的影响,因此采用生物结皮光谱特征提取方法的稳定性待提高.在实际应用中,由于生物结皮的空间分布复杂性和多样性,采用目视解译的方法易受到人为主观因素的影响,导致提取结果的不稳定性和不一致性.此外,生物结皮的覆盖范围广泛,包括草地、森林、湿地等不同类型的植被,这些生境特征在中低分辨率的遥感影像中难以准确区分,从而给生物结皮提取带来一定的挑战.另外,目视解译提取生物结皮需要大量的人力和时间投入,效率较低,尤其在大范围地区的生物结皮监测和研究中更为困难. ...
Measuring reflectance of tiny organisms:the promise of species level biocrust remote sensing
4
2021
... 目前,生物结皮在中国北方干旱半干旱区以及南方湿润半湿润区均有大量研究,主要是通过站点观测数据分析结皮对土壤水碳固持等关键过程的影响[7 -10 ] .生物结皮分布的提取方法主要有两种,一种是基于光谱特征的光谱分析法[11 -13 ] ,另外一种是基于遥感数据的波段代数运算构成的指数法[14 -17 ] .在光谱分析法中,主要针对生物结皮的光谱进行分析,以光谱特征之间的差异来区分不同种类生物结皮,这种基于面向对象的提取方法在生物结皮相关提取中作为常识性应用.田园盛等[11 ] 借助光谱特征之间差异以区分藻类、藓类等不同类型生物结皮;宋丽丽等[12 ] 结合反映相片DN值信息的RGB阈值法和ML(最大似然分类)法高精度准确识别出分布于干旱半干旱退化草原区的生物结皮;Baxter等[13 ] 采用机器学习(随机森林)对生物结皮的光谱特征进行评估,以区分不同种类拥有不同结构形状的生物结皮.这些方法需要在站点尺度下的精细观测以确定阈值,但该阈值计算受人为因素或其他环境因素的影响.受野外气候、地貌、侵蚀类型等多方面环境因素的影响[18 -19 ] ,生物结皮的生物特性会发生一定变化,从而导致其光谱特性在不同条件下引起一定变化[11 ] ,生物结皮指数上下限阈值的准确性也会受一定的影响,因此采用生物结皮光谱特征提取方法的稳定性待提高.在实际应用中,由于生物结皮的空间分布复杂性和多样性,采用目视解译的方法易受到人为主观因素的影响,导致提取结果的不稳定性和不一致性.此外,生物结皮的覆盖范围广泛,包括草地、森林、湿地等不同类型的植被,这些生境特征在中低分辨率的遥感影像中难以准确区分,从而给生物结皮提取带来一定的挑战.另外,目视解译提取生物结皮需要大量的人力和时间投入,效率较低,尤其在大范围地区的生物结皮监测和研究中更为困难. ...
... [13 ]采用机器学习(随机森林)对生物结皮的光谱特征进行评估,以区分不同种类拥有不同结构形状的生物结皮.这些方法需要在站点尺度下的精细观测以确定阈值,但该阈值计算受人为因素或其他环境因素的影响.受野外气候、地貌、侵蚀类型等多方面环境因素的影响[18 -19 ] ,生物结皮的生物特性会发生一定变化,从而导致其光谱特性在不同条件下引起一定变化[11 ] ,生物结皮指数上下限阈值的准确性也会受一定的影响,因此采用生物结皮光谱特征提取方法的稳定性待提高.在实际应用中,由于生物结皮的空间分布复杂性和多样性,采用目视解译的方法易受到人为主观因素的影响,导致提取结果的不稳定性和不一致性.此外,生物结皮的覆盖范围广泛,包括草地、森林、湿地等不同类型的植被,这些生境特征在中低分辨率的遥感影像中难以准确区分,从而给生物结皮提取带来一定的挑战.另外,目视解译提取生物结皮需要大量的人力和时间投入,效率较低,尤其在大范围地区的生物结皮监测和研究中更为困难. ...
... 与此同时,在另外一种遥感结皮指数法中,上述方法出现的问题能够较好地被处理,少数学者通过遥感技术与站点尺度精细化观测相结合的方法对区域尺度生物结皮的空间分布及生物结皮的水文效应进行初步探讨,取得很大进展[13 -17 ,20 -21 ] .Karnieli[20 ] 利用Landsat TM影像红色波段与蓝色波段归一化差值构建光谱生物结皮指数,在沙漠干旱环境下检测以蓝绿藻的生物结皮检测和绘制遥感图像中的不同岩性形态单元.Chen等[22 ] 通过定量实验发现生物结皮在红色与绿色波段之间的变化率比裸土、枯植或绿色植被更小,并且生物结皮在可见光与近红外波段的反射率比裸土、枯植或绿色植被要低得多,在此基础上构建了增大生物结皮与裸土、枯植、绿色植被之间差异的生物结皮指数.张元明等[14 ] 、杨伟等[16 ] 通过Landsat ETM数据中的绿光、红光、近红外波段地表反射率构建了三波段生物结皮指数,分别分析了空间上和时间上古班尔通古特沙漠以地衣为主的生物结皮分布特征与变化规律.冯秀绒等[17 ] 优化了由绿光、红光、近红外波段地表反射率构建的生物结皮提取方法,采用结皮指数、归一化植被指数、监督分类、坡度分级相结合的方法提取了毛乌素沙地以苔藓类结皮为主的生物结皮.Sun等[21 ] 基于植被辐射传输理论建立了一个生物结皮估计模型,在与其他模型进行比较后发现该模型对古尔班通古特沙漠的生物结皮具有良好的提取效果.综上所述,基于遥感方法提取生物结皮空间分布的手段主要是利用遥感数据的不同波段构建生物结皮指数,且指数主要基于红光吸收特征和近红外的反射特征的物理机制构建.然而,以上结皮指数主要在沙漠区域进行研究验证,沙漠区域地物类型以结皮和流动沙丘为主,结构单一.由于黄土高原大规模植被恢复工程的实施,致使地表地物结构发生了极大改变,植被覆盖度呈现明显增加趋势[23 ] .因此,如何进行黄土高原植被覆盖下地表生物结皮提取,构建适用于干旱半干旱区多地物特征相互作用下生物结皮遥感提取方法是本研究的技术难点. ...
... 本研究结合野外调查和遥感机制分析,开发了小流域生物结皮逐步提取方法,该方法本质上是依靠地物光谱反射率所计算的各种特征遥感指标之间聚合与分离的生物结皮遥感提取方法,该方法相比较光谱分析法[11 -13 ] 、指数法[14 -17 ] 等取得了较大进步.在考虑流域地表地物反射特性、辐射传输特性的基础上,利用逐步提取的方式针对与生物结皮相似的裸土、植被像元等进行单独提取分离,在一定程度上提高了生物结皮的提取精度. ...
古尔班通古特沙漠生物结皮的分布特征
5
2005
... 目前,生物结皮在中国北方干旱半干旱区以及南方湿润半湿润区均有大量研究,主要是通过站点观测数据分析结皮对土壤水碳固持等关键过程的影响[7 -10 ] .生物结皮分布的提取方法主要有两种,一种是基于光谱特征的光谱分析法[11 -13 ] ,另外一种是基于遥感数据的波段代数运算构成的指数法[14 -17 ] .在光谱分析法中,主要针对生物结皮的光谱进行分析,以光谱特征之间的差异来区分不同种类生物结皮,这种基于面向对象的提取方法在生物结皮相关提取中作为常识性应用.田园盛等[11 ] 借助光谱特征之间差异以区分藻类、藓类等不同类型生物结皮;宋丽丽等[12 ] 结合反映相片DN值信息的RGB阈值法和ML(最大似然分类)法高精度准确识别出分布于干旱半干旱退化草原区的生物结皮;Baxter等[13 ] 采用机器学习(随机森林)对生物结皮的光谱特征进行评估,以区分不同种类拥有不同结构形状的生物结皮.这些方法需要在站点尺度下的精细观测以确定阈值,但该阈值计算受人为因素或其他环境因素的影响.受野外气候、地貌、侵蚀类型等多方面环境因素的影响[18 -19 ] ,生物结皮的生物特性会发生一定变化,从而导致其光谱特性在不同条件下引起一定变化[11 ] ,生物结皮指数上下限阈值的准确性也会受一定的影响,因此采用生物结皮光谱特征提取方法的稳定性待提高.在实际应用中,由于生物结皮的空间分布复杂性和多样性,采用目视解译的方法易受到人为主观因素的影响,导致提取结果的不稳定性和不一致性.此外,生物结皮的覆盖范围广泛,包括草地、森林、湿地等不同类型的植被,这些生境特征在中低分辨率的遥感影像中难以准确区分,从而给生物结皮提取带来一定的挑战.另外,目视解译提取生物结皮需要大量的人力和时间投入,效率较低,尤其在大范围地区的生物结皮监测和研究中更为困难. ...
... 与此同时,在另外一种遥感结皮指数法中,上述方法出现的问题能够较好地被处理,少数学者通过遥感技术与站点尺度精细化观测相结合的方法对区域尺度生物结皮的空间分布及生物结皮的水文效应进行初步探讨,取得很大进展[13 -17 ,20 -21 ] .Karnieli[20 ] 利用Landsat TM影像红色波段与蓝色波段归一化差值构建光谱生物结皮指数,在沙漠干旱环境下检测以蓝绿藻的生物结皮检测和绘制遥感图像中的不同岩性形态单元.Chen等[22 ] 通过定量实验发现生物结皮在红色与绿色波段之间的变化率比裸土、枯植或绿色植被更小,并且生物结皮在可见光与近红外波段的反射率比裸土、枯植或绿色植被要低得多,在此基础上构建了增大生物结皮与裸土、枯植、绿色植被之间差异的生物结皮指数.张元明等[14 ] 、杨伟等[16 ] 通过Landsat ETM数据中的绿光、红光、近红外波段地表反射率构建了三波段生物结皮指数,分别分析了空间上和时间上古班尔通古特沙漠以地衣为主的生物结皮分布特征与变化规律.冯秀绒等[17 ] 优化了由绿光、红光、近红外波段地表反射率构建的生物结皮提取方法,采用结皮指数、归一化植被指数、监督分类、坡度分级相结合的方法提取了毛乌素沙地以苔藓类结皮为主的生物结皮.Sun等[21 ] 基于植被辐射传输理论建立了一个生物结皮估计模型,在与其他模型进行比较后发现该模型对古尔班通古特沙漠的生物结皮具有良好的提取效果.综上所述,基于遥感方法提取生物结皮空间分布的手段主要是利用遥感数据的不同波段构建生物结皮指数,且指数主要基于红光吸收特征和近红外的反射特征的物理机制构建.然而,以上结皮指数主要在沙漠区域进行研究验证,沙漠区域地物类型以结皮和流动沙丘为主,结构单一.由于黄土高原大规模植被恢复工程的实施,致使地表地物结构发生了极大改变,植被覆盖度呈现明显增加趋势[23 ] .因此,如何进行黄土高原植被覆盖下地表生物结皮提取,构建适用于干旱半干旱区多地物特征相互作用下生物结皮遥感提取方法是本研究的技术难点. ...
... 生物结皮指数(BSCI)是对生物结皮光谱特征的增强,使得与裸沙和干枯植物等背景地物存在较高的背景反射值.按照BSCI的定义并考虑卫星传感器的空间分辨率,一般认为BSCI值主要取决于像元内生物结皮的覆盖率.生物结皮覆盖率增加,BSCI值也增加.与之相反,背景地物的覆盖率增加,则BSCI值降低[14 ] .本研究利用BSCI值的分布密度曲线与其他地物分布密度曲线以确定生物结皮BSCI值的上下限阈值. ...
... 生物结皮指数的阈值遥感确定是提取过程中的关键;张元明等[14 ] 通过野外测量的生物结皮的高光谱数据确定BSCI的阈值上下限,但是该值存在显著的时空差异.Guo等[29 ] 在提取黄土高原小水体时设定了一定的水体提取规则从而确定提取水体的阈值方法,该方法能够实现阈值动态调整,具有较好的时空适宜性.因此,本研究参考水体提取规则设定适用于裸土、植被、生物结皮的提取规则,即对于SAVI、MSAVI指数,裸土信号值(分布密度)大于非裸土信号值的像元判定为裸土,反之则判定为非裸土;对于NDVI、NIRV 指数,植被信号值(分布密度)大于非植被信号值的像元判定为植被,反之则判定为非植被;对于BSCI指数,生物结皮信号值(分布密度)大于非生物结皮信号值的像元判定为生物结皮,反之则判定为非生物结皮.通过以上提取规则的确定,逐步提取分离目标地物,最终得到生物结皮的上下限阈值. ...
... 本研究结合野外调查和遥感机制分析,开发了小流域生物结皮逐步提取方法,该方法本质上是依靠地物光谱反射率所计算的各种特征遥感指标之间聚合与分离的生物结皮遥感提取方法,该方法相比较光谱分析法[11 -13 ] 、指数法[14 -17 ] 等取得了较大进步.在考虑流域地表地物反射特性、辐射传输特性的基础上,利用逐步提取的方式针对与生物结皮相似的裸土、植被像元等进行单独提取分离,在一定程度上提高了生物结皮的提取精度. ...
古尔班通古特沙漠不同地貌部位生物结皮的选择性分布
1
2004
... 降水、气温与地形对生物结皮存在交互影响;生物结皮生长范围广泛,主要类型包括苔藓、地衣、藻类植物等[2 ] ,生物结皮多喜好阴凉潮湿环境.因此,生物结皮对降水的变化反应极其敏感,在干旱半干旱区随着气温的升高和夏季降水频率的增多,生物结皮的生物多样性急剧下降[35 ] .全球气候变化导致美国麦哈韦沙漠和科罗拉多州高原夏季降水增多,而夏季降水增多,促使了某些种类的生物结皮盖度下降,生理功能改变.大气温度是影响结皮形成、空间分布特征的另一个关键因子,学者通过对最近22年来的329种地衣的物种组成变化和分布变化研究,发现随全球变暖喜温生物结皮在荷兰的种类增多而耐寒地衣种减少[36 ] .同时,不同地形因坡度和坡向的差异显著影响,降水和温度的分布格局、阴坡气温高于阳坡、不同海拔间降水存在显著差异,这些气候、地形等环境因素显著影响生物结皮的空间分布[15 ,37 ] .此外,流域土地利用方式也影响着生物结皮的形成与分布,这种差异主要体现在林草地以及其他土地利用方式之间,草地生物结皮盖度与林地生物结皮盖度存在一定的差异性[38 ] ,而林草地生物结皮的分布与其他土地利用类型生物结皮的分布也存在着一定的差异性. ...
古尔班通古特沙漠1970-2000年代生物结皮覆盖变化研究
1
2006
... 与此同时,在另外一种遥感结皮指数法中,上述方法出现的问题能够较好地被处理,少数学者通过遥感技术与站点尺度精细化观测相结合的方法对区域尺度生物结皮的空间分布及生物结皮的水文效应进行初步探讨,取得很大进展[13 -17 ,20 -21 ] .Karnieli[20 ] 利用Landsat TM影像红色波段与蓝色波段归一化差值构建光谱生物结皮指数,在沙漠干旱环境下检测以蓝绿藻的生物结皮检测和绘制遥感图像中的不同岩性形态单元.Chen等[22 ] 通过定量实验发现生物结皮在红色与绿色波段之间的变化率比裸土、枯植或绿色植被更小,并且生物结皮在可见光与近红外波段的反射率比裸土、枯植或绿色植被要低得多,在此基础上构建了增大生物结皮与裸土、枯植、绿色植被之间差异的生物结皮指数.张元明等[14 ] 、杨伟等[16 ] 通过Landsat ETM数据中的绿光、红光、近红外波段地表反射率构建了三波段生物结皮指数,分别分析了空间上和时间上古班尔通古特沙漠以地衣为主的生物结皮分布特征与变化规律.冯秀绒等[17 ] 优化了由绿光、红光、近红外波段地表反射率构建的生物结皮提取方法,采用结皮指数、归一化植被指数、监督分类、坡度分级相结合的方法提取了毛乌素沙地以苔藓类结皮为主的生物结皮.Sun等[21 ] 基于植被辐射传输理论建立了一个生物结皮估计模型,在与其他模型进行比较后发现该模型对古尔班通古特沙漠的生物结皮具有良好的提取效果.综上所述,基于遥感方法提取生物结皮空间分布的手段主要是利用遥感数据的不同波段构建生物结皮指数,且指数主要基于红光吸收特征和近红外的反射特征的物理机制构建.然而,以上结皮指数主要在沙漠区域进行研究验证,沙漠区域地物类型以结皮和流动沙丘为主,结构单一.由于黄土高原大规模植被恢复工程的实施,致使地表地物结构发生了极大改变,植被覆盖度呈现明显增加趋势[23 ] .因此,如何进行黄土高原植被覆盖下地表生物结皮提取,构建适用于干旱半干旱区多地物特征相互作用下生物结皮遥感提取方法是本研究的技术难点. ...
基于光谱分析的生物结皮提取研究:以毛乌素沙地为例
4
2015
... 目前,生物结皮在中国北方干旱半干旱区以及南方湿润半湿润区均有大量研究,主要是通过站点观测数据分析结皮对土壤水碳固持等关键过程的影响[7 -10 ] .生物结皮分布的提取方法主要有两种,一种是基于光谱特征的光谱分析法[11 -13 ] ,另外一种是基于遥感数据的波段代数运算构成的指数法[14 -17 ] .在光谱分析法中,主要针对生物结皮的光谱进行分析,以光谱特征之间的差异来区分不同种类生物结皮,这种基于面向对象的提取方法在生物结皮相关提取中作为常识性应用.田园盛等[11 ] 借助光谱特征之间差异以区分藻类、藓类等不同类型生物结皮;宋丽丽等[12 ] 结合反映相片DN值信息的RGB阈值法和ML(最大似然分类)法高精度准确识别出分布于干旱半干旱退化草原区的生物结皮;Baxter等[13 ] 采用机器学习(随机森林)对生物结皮的光谱特征进行评估,以区分不同种类拥有不同结构形状的生物结皮.这些方法需要在站点尺度下的精细观测以确定阈值,但该阈值计算受人为因素或其他环境因素的影响.受野外气候、地貌、侵蚀类型等多方面环境因素的影响[18 -19 ] ,生物结皮的生物特性会发生一定变化,从而导致其光谱特性在不同条件下引起一定变化[11 ] ,生物结皮指数上下限阈值的准确性也会受一定的影响,因此采用生物结皮光谱特征提取方法的稳定性待提高.在实际应用中,由于生物结皮的空间分布复杂性和多样性,采用目视解译的方法易受到人为主观因素的影响,导致提取结果的不稳定性和不一致性.此外,生物结皮的覆盖范围广泛,包括草地、森林、湿地等不同类型的植被,这些生境特征在中低分辨率的遥感影像中难以准确区分,从而给生物结皮提取带来一定的挑战.另外,目视解译提取生物结皮需要大量的人力和时间投入,效率较低,尤其在大范围地区的生物结皮监测和研究中更为困难. ...
... 与此同时,在另外一种遥感结皮指数法中,上述方法出现的问题能够较好地被处理,少数学者通过遥感技术与站点尺度精细化观测相结合的方法对区域尺度生物结皮的空间分布及生物结皮的水文效应进行初步探讨,取得很大进展[13 -17 ,20 -21 ] .Karnieli[20 ] 利用Landsat TM影像红色波段与蓝色波段归一化差值构建光谱生物结皮指数,在沙漠干旱环境下检测以蓝绿藻的生物结皮检测和绘制遥感图像中的不同岩性形态单元.Chen等[22 ] 通过定量实验发现生物结皮在红色与绿色波段之间的变化率比裸土、枯植或绿色植被更小,并且生物结皮在可见光与近红外波段的反射率比裸土、枯植或绿色植被要低得多,在此基础上构建了增大生物结皮与裸土、枯植、绿色植被之间差异的生物结皮指数.张元明等[14 ] 、杨伟等[16 ] 通过Landsat ETM数据中的绿光、红光、近红外波段地表反射率构建了三波段生物结皮指数,分别分析了空间上和时间上古班尔通古特沙漠以地衣为主的生物结皮分布特征与变化规律.冯秀绒等[17 ] 优化了由绿光、红光、近红外波段地表反射率构建的生物结皮提取方法,采用结皮指数、归一化植被指数、监督分类、坡度分级相结合的方法提取了毛乌素沙地以苔藓类结皮为主的生物结皮.Sun等[21 ] 基于植被辐射传输理论建立了一个生物结皮估计模型,在与其他模型进行比较后发现该模型对古尔班通古特沙漠的生物结皮具有良好的提取效果.综上所述,基于遥感方法提取生物结皮空间分布的手段主要是利用遥感数据的不同波段构建生物结皮指数,且指数主要基于红光吸收特征和近红外的反射特征的物理机制构建.然而,以上结皮指数主要在沙漠区域进行研究验证,沙漠区域地物类型以结皮和流动沙丘为主,结构单一.由于黄土高原大规模植被恢复工程的实施,致使地表地物结构发生了极大改变,植被覆盖度呈现明显增加趋势[23 ] .因此,如何进行黄土高原植被覆盖下地表生物结皮提取,构建适用于干旱半干旱区多地物特征相互作用下生物结皮遥感提取方法是本研究的技术难点. ...
... [17 ]优化了由绿光、红光、近红外波段地表反射率构建的生物结皮提取方法,采用结皮指数、归一化植被指数、监督分类、坡度分级相结合的方法提取了毛乌素沙地以苔藓类结皮为主的生物结皮.Sun等[21 ] 基于植被辐射传输理论建立了一个生物结皮估计模型,在与其他模型进行比较后发现该模型对古尔班通古特沙漠的生物结皮具有良好的提取效果.综上所述,基于遥感方法提取生物结皮空间分布的手段主要是利用遥感数据的不同波段构建生物结皮指数,且指数主要基于红光吸收特征和近红外的反射特征的物理机制构建.然而,以上结皮指数主要在沙漠区域进行研究验证,沙漠区域地物类型以结皮和流动沙丘为主,结构单一.由于黄土高原大规模植被恢复工程的实施,致使地表地物结构发生了极大改变,植被覆盖度呈现明显增加趋势[23 ] .因此,如何进行黄土高原植被覆盖下地表生物结皮提取,构建适用于干旱半干旱区多地物特征相互作用下生物结皮遥感提取方法是本研究的技术难点. ...
... 本研究结合野外调查和遥感机制分析,开发了小流域生物结皮逐步提取方法,该方法本质上是依靠地物光谱反射率所计算的各种特征遥感指标之间聚合与分离的生物结皮遥感提取方法,该方法相比较光谱分析法[11 -13 ] 、指数法[14 -17 ] 等取得了较大进步.在考虑流域地表地物反射特性、辐射传输特性的基础上,利用逐步提取的方式针对与生物结皮相似的裸土、植被像元等进行单独提取分离,在一定程度上提高了生物结皮的提取精度. ...
黄土高原不同侵蚀类型区生物结皮中蓝藻的多样性
1
2013
... 目前,生物结皮在中国北方干旱半干旱区以及南方湿润半湿润区均有大量研究,主要是通过站点观测数据分析结皮对土壤水碳固持等关键过程的影响[7 -10 ] .生物结皮分布的提取方法主要有两种,一种是基于光谱特征的光谱分析法[11 -13 ] ,另外一种是基于遥感数据的波段代数运算构成的指数法[14 -17 ] .在光谱分析法中,主要针对生物结皮的光谱进行分析,以光谱特征之间的差异来区分不同种类生物结皮,这种基于面向对象的提取方法在生物结皮相关提取中作为常识性应用.田园盛等[11 ] 借助光谱特征之间差异以区分藻类、藓类等不同类型生物结皮;宋丽丽等[12 ] 结合反映相片DN值信息的RGB阈值法和ML(最大似然分类)法高精度准确识别出分布于干旱半干旱退化草原区的生物结皮;Baxter等[13 ] 采用机器学习(随机森林)对生物结皮的光谱特征进行评估,以区分不同种类拥有不同结构形状的生物结皮.这些方法需要在站点尺度下的精细观测以确定阈值,但该阈值计算受人为因素或其他环境因素的影响.受野外气候、地貌、侵蚀类型等多方面环境因素的影响[18 -19 ] ,生物结皮的生物特性会发生一定变化,从而导致其光谱特性在不同条件下引起一定变化[11 ] ,生物结皮指数上下限阈值的准确性也会受一定的影响,因此采用生物结皮光谱特征提取方法的稳定性待提高.在实际应用中,由于生物结皮的空间分布复杂性和多样性,采用目视解译的方法易受到人为主观因素的影响,导致提取结果的不稳定性和不一致性.此外,生物结皮的覆盖范围广泛,包括草地、森林、湿地等不同类型的植被,这些生境特征在中低分辨率的遥感影像中难以准确区分,从而给生物结皮提取带来一定的挑战.另外,目视解译提取生物结皮需要大量的人力和时间投入,效率较低,尤其在大范围地区的生物结皮监测和研究中更为困难. ...
模拟增温对高寒沙区生物土壤结皮-土壤系统呼吸的影响
1
2020
... 目前,生物结皮在中国北方干旱半干旱区以及南方湿润半湿润区均有大量研究,主要是通过站点观测数据分析结皮对土壤水碳固持等关键过程的影响[7 -10 ] .生物结皮分布的提取方法主要有两种,一种是基于光谱特征的光谱分析法[11 -13 ] ,另外一种是基于遥感数据的波段代数运算构成的指数法[14 -17 ] .在光谱分析法中,主要针对生物结皮的光谱进行分析,以光谱特征之间的差异来区分不同种类生物结皮,这种基于面向对象的提取方法在生物结皮相关提取中作为常识性应用.田园盛等[11 ] 借助光谱特征之间差异以区分藻类、藓类等不同类型生物结皮;宋丽丽等[12 ] 结合反映相片DN值信息的RGB阈值法和ML(最大似然分类)法高精度准确识别出分布于干旱半干旱退化草原区的生物结皮;Baxter等[13 ] 采用机器学习(随机森林)对生物结皮的光谱特征进行评估,以区分不同种类拥有不同结构形状的生物结皮.这些方法需要在站点尺度下的精细观测以确定阈值,但该阈值计算受人为因素或其他环境因素的影响.受野外气候、地貌、侵蚀类型等多方面环境因素的影响[18 -19 ] ,生物结皮的生物特性会发生一定变化,从而导致其光谱特性在不同条件下引起一定变化[11 ] ,生物结皮指数上下限阈值的准确性也会受一定的影响,因此采用生物结皮光谱特征提取方法的稳定性待提高.在实际应用中,由于生物结皮的空间分布复杂性和多样性,采用目视解译的方法易受到人为主观因素的影响,导致提取结果的不稳定性和不一致性.此外,生物结皮的覆盖范围广泛,包括草地、森林、湿地等不同类型的植被,这些生境特征在中低分辨率的遥感影像中难以准确区分,从而给生物结皮提取带来一定的挑战.另外,目视解译提取生物结皮需要大量的人力和时间投入,效率较低,尤其在大范围地区的生物结皮监测和研究中更为困难. ...
Development and implementation of spectral crust index over dune sands
2
1997
... 与此同时,在另外一种遥感结皮指数法中,上述方法出现的问题能够较好地被处理,少数学者通过遥感技术与站点尺度精细化观测相结合的方法对区域尺度生物结皮的空间分布及生物结皮的水文效应进行初步探讨,取得很大进展[13 -17 ,20 -21 ] .Karnieli[20 ] 利用Landsat TM影像红色波段与蓝色波段归一化差值构建光谱生物结皮指数,在沙漠干旱环境下检测以蓝绿藻的生物结皮检测和绘制遥感图像中的不同岩性形态单元.Chen等[22 ] 通过定量实验发现生物结皮在红色与绿色波段之间的变化率比裸土、枯植或绿色植被更小,并且生物结皮在可见光与近红外波段的反射率比裸土、枯植或绿色植被要低得多,在此基础上构建了增大生物结皮与裸土、枯植、绿色植被之间差异的生物结皮指数.张元明等[14 ] 、杨伟等[16 ] 通过Landsat ETM数据中的绿光、红光、近红外波段地表反射率构建了三波段生物结皮指数,分别分析了空间上和时间上古班尔通古特沙漠以地衣为主的生物结皮分布特征与变化规律.冯秀绒等[17 ] 优化了由绿光、红光、近红外波段地表反射率构建的生物结皮提取方法,采用结皮指数、归一化植被指数、监督分类、坡度分级相结合的方法提取了毛乌素沙地以苔藓类结皮为主的生物结皮.Sun等[21 ] 基于植被辐射传输理论建立了一个生物结皮估计模型,在与其他模型进行比较后发现该模型对古尔班通古特沙漠的生物结皮具有良好的提取效果.综上所述,基于遥感方法提取生物结皮空间分布的手段主要是利用遥感数据的不同波段构建生物结皮指数,且指数主要基于红光吸收特征和近红外的反射特征的物理机制构建.然而,以上结皮指数主要在沙漠区域进行研究验证,沙漠区域地物类型以结皮和流动沙丘为主,结构单一.由于黄土高原大规模植被恢复工程的实施,致使地表地物结构发生了极大改变,植被覆盖度呈现明显增加趋势[23 ] .因此,如何进行黄土高原植被覆盖下地表生物结皮提取,构建适用于干旱半干旱区多地物特征相互作用下生物结皮遥感提取方法是本研究的技术难点. ...
... [20 ]利用Landsat TM影像红色波段与蓝色波段归一化差值构建光谱生物结皮指数,在沙漠干旱环境下检测以蓝绿藻的生物结皮检测和绘制遥感图像中的不同岩性形态单元.Chen等[22 ] 通过定量实验发现生物结皮在红色与绿色波段之间的变化率比裸土、枯植或绿色植被更小,并且生物结皮在可见光与近红外波段的反射率比裸土、枯植或绿色植被要低得多,在此基础上构建了增大生物结皮与裸土、枯植、绿色植被之间差异的生物结皮指数.张元明等[14 ] 、杨伟等[16 ] 通过Landsat ETM数据中的绿光、红光、近红外波段地表反射率构建了三波段生物结皮指数,分别分析了空间上和时间上古班尔通古特沙漠以地衣为主的生物结皮分布特征与变化规律.冯秀绒等[17 ] 优化了由绿光、红光、近红外波段地表反射率构建的生物结皮提取方法,采用结皮指数、归一化植被指数、监督分类、坡度分级相结合的方法提取了毛乌素沙地以苔藓类结皮为主的生物结皮.Sun等[21 ] 基于植被辐射传输理论建立了一个生物结皮估计模型,在与其他模型进行比较后发现该模型对古尔班通古特沙漠的生物结皮具有良好的提取效果.综上所述,基于遥感方法提取生物结皮空间分布的手段主要是利用遥感数据的不同波段构建生物结皮指数,且指数主要基于红光吸收特征和近红外的反射特征的物理机制构建.然而,以上结皮指数主要在沙漠区域进行研究验证,沙漠区域地物类型以结皮和流动沙丘为主,结构单一.由于黄土高原大规模植被恢复工程的实施,致使地表地物结构发生了极大改变,植被覆盖度呈现明显增加趋势[23 ] .因此,如何进行黄土高原植被覆盖下地表生物结皮提取,构建适用于干旱半干旱区多地物特征相互作用下生物结皮遥感提取方法是本研究的技术难点. ...
A New multiangle method for estimating fractional biocrust coverage from sentinel-2 data in arid areas
2
2024
... 与此同时,在另外一种遥感结皮指数法中,上述方法出现的问题能够较好地被处理,少数学者通过遥感技术与站点尺度精细化观测相结合的方法对区域尺度生物结皮的空间分布及生物结皮的水文效应进行初步探讨,取得很大进展[13 -17 ,20 -21 ] .Karnieli[20 ] 利用Landsat TM影像红色波段与蓝色波段归一化差值构建光谱生物结皮指数,在沙漠干旱环境下检测以蓝绿藻的生物结皮检测和绘制遥感图像中的不同岩性形态单元.Chen等[22 ] 通过定量实验发现生物结皮在红色与绿色波段之间的变化率比裸土、枯植或绿色植被更小,并且生物结皮在可见光与近红外波段的反射率比裸土、枯植或绿色植被要低得多,在此基础上构建了增大生物结皮与裸土、枯植、绿色植被之间差异的生物结皮指数.张元明等[14 ] 、杨伟等[16 ] 通过Landsat ETM数据中的绿光、红光、近红外波段地表反射率构建了三波段生物结皮指数,分别分析了空间上和时间上古班尔通古特沙漠以地衣为主的生物结皮分布特征与变化规律.冯秀绒等[17 ] 优化了由绿光、红光、近红外波段地表反射率构建的生物结皮提取方法,采用结皮指数、归一化植被指数、监督分类、坡度分级相结合的方法提取了毛乌素沙地以苔藓类结皮为主的生物结皮.Sun等[21 ] 基于植被辐射传输理论建立了一个生物结皮估计模型,在与其他模型进行比较后发现该模型对古尔班通古特沙漠的生物结皮具有良好的提取效果.综上所述,基于遥感方法提取生物结皮空间分布的手段主要是利用遥感数据的不同波段构建生物结皮指数,且指数主要基于红光吸收特征和近红外的反射特征的物理机制构建.然而,以上结皮指数主要在沙漠区域进行研究验证,沙漠区域地物类型以结皮和流动沙丘为主,结构单一.由于黄土高原大规模植被恢复工程的实施,致使地表地物结构发生了极大改变,植被覆盖度呈现明显增加趋势[23 ] .因此,如何进行黄土高原植被覆盖下地表生物结皮提取,构建适用于干旱半干旱区多地物特征相互作用下生物结皮遥感提取方法是本研究的技术难点. ...
... [21 ]基于植被辐射传输理论建立了一个生物结皮估计模型,在与其他模型进行比较后发现该模型对古尔班通古特沙漠的生物结皮具有良好的提取效果.综上所述,基于遥感方法提取生物结皮空间分布的手段主要是利用遥感数据的不同波段构建生物结皮指数,且指数主要基于红光吸收特征和近红外的反射特征的物理机制构建.然而,以上结皮指数主要在沙漠区域进行研究验证,沙漠区域地物类型以结皮和流动沙丘为主,结构单一.由于黄土高原大规模植被恢复工程的实施,致使地表地物结构发生了极大改变,植被覆盖度呈现明显增加趋势[23 ] .因此,如何进行黄土高原植被覆盖下地表生物结皮提取,构建适用于干旱半干旱区多地物特征相互作用下生物结皮遥感提取方法是本研究的技术难点. ...
A new index for mapping lichen-dominated biological soil crusts in desert areas
1
2005
... 与此同时,在另外一种遥感结皮指数法中,上述方法出现的问题能够较好地被处理,少数学者通过遥感技术与站点尺度精细化观测相结合的方法对区域尺度生物结皮的空间分布及生物结皮的水文效应进行初步探讨,取得很大进展[13 -17 ,20 -21 ] .Karnieli[20 ] 利用Landsat TM影像红色波段与蓝色波段归一化差值构建光谱生物结皮指数,在沙漠干旱环境下检测以蓝绿藻的生物结皮检测和绘制遥感图像中的不同岩性形态单元.Chen等[22 ] 通过定量实验发现生物结皮在红色与绿色波段之间的变化率比裸土、枯植或绿色植被更小,并且生物结皮在可见光与近红外波段的反射率比裸土、枯植或绿色植被要低得多,在此基础上构建了增大生物结皮与裸土、枯植、绿色植被之间差异的生物结皮指数.张元明等[14 ] 、杨伟等[16 ] 通过Landsat ETM数据中的绿光、红光、近红外波段地表反射率构建了三波段生物结皮指数,分别分析了空间上和时间上古班尔通古特沙漠以地衣为主的生物结皮分布特征与变化规律.冯秀绒等[17 ] 优化了由绿光、红光、近红外波段地表反射率构建的生物结皮提取方法,采用结皮指数、归一化植被指数、监督分类、坡度分级相结合的方法提取了毛乌素沙地以苔藓类结皮为主的生物结皮.Sun等[21 ] 基于植被辐射传输理论建立了一个生物结皮估计模型,在与其他模型进行比较后发现该模型对古尔班通古特沙漠的生物结皮具有良好的提取效果.综上所述,基于遥感方法提取生物结皮空间分布的手段主要是利用遥感数据的不同波段构建生物结皮指数,且指数主要基于红光吸收特征和近红外的反射特征的物理机制构建.然而,以上结皮指数主要在沙漠区域进行研究验证,沙漠区域地物类型以结皮和流动沙丘为主,结构单一.由于黄土高原大规模植被恢复工程的实施,致使地表地物结构发生了极大改变,植被覆盖度呈现明显增加趋势[23 ] .因此,如何进行黄土高原植被覆盖下地表生物结皮提取,构建适用于干旱半干旱区多地物特征相互作用下生物结皮遥感提取方法是本研究的技术难点. ...
近10年陕甘宁黄土高原区植被覆盖时空变化特征
1
2012
... 与此同时,在另外一种遥感结皮指数法中,上述方法出现的问题能够较好地被处理,少数学者通过遥感技术与站点尺度精细化观测相结合的方法对区域尺度生物结皮的空间分布及生物结皮的水文效应进行初步探讨,取得很大进展[13 -17 ,20 -21 ] .Karnieli[20 ] 利用Landsat TM影像红色波段与蓝色波段归一化差值构建光谱生物结皮指数,在沙漠干旱环境下检测以蓝绿藻的生物结皮检测和绘制遥感图像中的不同岩性形态单元.Chen等[22 ] 通过定量实验发现生物结皮在红色与绿色波段之间的变化率比裸土、枯植或绿色植被更小,并且生物结皮在可见光与近红外波段的反射率比裸土、枯植或绿色植被要低得多,在此基础上构建了增大生物结皮与裸土、枯植、绿色植被之间差异的生物结皮指数.张元明等[14 ] 、杨伟等[16 ] 通过Landsat ETM数据中的绿光、红光、近红外波段地表反射率构建了三波段生物结皮指数,分别分析了空间上和时间上古班尔通古特沙漠以地衣为主的生物结皮分布特征与变化规律.冯秀绒等[17 ] 优化了由绿光、红光、近红外波段地表反射率构建的生物结皮提取方法,采用结皮指数、归一化植被指数、监督分类、坡度分级相结合的方法提取了毛乌素沙地以苔藓类结皮为主的生物结皮.Sun等[21 ] 基于植被辐射传输理论建立了一个生物结皮估计模型,在与其他模型进行比较后发现该模型对古尔班通古特沙漠的生物结皮具有良好的提取效果.综上所述,基于遥感方法提取生物结皮空间分布的手段主要是利用遥感数据的不同波段构建生物结皮指数,且指数主要基于红光吸收特征和近红外的反射特征的物理机制构建.然而,以上结皮指数主要在沙漠区域进行研究验证,沙漠区域地物类型以结皮和流动沙丘为主,结构单一.由于黄土高原大规模植被恢复工程的实施,致使地表地物结构发生了极大改变,植被覆盖度呈现明显增加趋势[23 ] .因此,如何进行黄土高原植被覆盖下地表生物结皮提取,构建适用于干旱半干旱区多地物特征相互作用下生物结皮遥感提取方法是本研究的技术难点. ...
皇甫川流域地表特征参数遥感反演研究
1
2008
... 皇甫川流域面积3 246 km2 (图1 ),其中水土流失面积3 215 km2 ,占流域总面积的99.0%.流域属于干旱与半干旱气候,年降水量350~450 mm,是鄂尔多斯高原暴雨中心区,一般发生于夏季,占全年降水量的80%以上,年均暴雨5次,皇甫川沟深梁大、下游地势落差大的地形,造成了严重的流水侵蚀.皇甫川流域植被多密集分布于河流两侧,裸露河床与沙地均少有植被分布[24 ] .而皇甫川流域生物结皮分布规律与流域内植被类型的演替密切相关,因此研究皇甫川流域生物结皮对流域内的植被恢复、生态环境保护以及生态水文循环研究具有重要意义. ...
生态安全条件下土地利用格局优化:以皇甫川流域为例
1
2014
... 皇甫川流域地表地物类型可分为建筑用地、林地、草地、水域、沙地、裸土、耕地等[25 ] .目视解译发现地表生物结皮多分布于草地与沙地、裸土,分类的界限难以确立,故本研究通过地物的遥感特征指数来建立目标地物提取规则以确定不同目标地物分割阈值.本研究利用两种土壤调节植被指数和两种植被指数确定土壤、植被指数值的上下限阈值. ...
植被指数研究进展
2
1998
... 土壤调节植被指数(SAVI)是在NDVI指数的基础上增加了土壤调节系数L ,取值范围0~1,建立可适当描述土壤-植被系统的简单模型.而修正土壤调节植被指数(MSAVI)适当减小了SAVI中的裸土影响[26 ] .上述指数考虑植被提取时在土壤背景下的影响,因此本研究以这两个指数结合分类时裸土分布特征,间接识别并提取裸土像元. ...
... 归一化差值植被指数(NDVI)是检测植被生长状态、植被覆盖度重要遥感指数[27 ] .在低密度植被覆盖区域,NDVI具有非常敏感的观测效果[26 ] .新型光合植被指数(NIRV )指的是归一化植被指数与近红外波段反射率的乘积,与NDVI相比,NIRV 可以在高生物量区域使用,有效减轻了植被-土壤混合像元的影响[28 ] .本研究选取以上两种遥感指数对皇甫川流域的植被地物进行提取,两者在不同覆盖度植被提取上互补缺点,一定程度上减少了皇甫川流域植被的提取误差. ...
植被指数研究进展:从AVHRR-NDVI到MODIS-EVI
1
2003
... 归一化差值植被指数(NDVI)是检测植被生长状态、植被覆盖度重要遥感指数[27 ] .在低密度植被覆盖区域,NDVI具有非常敏感的观测效果[26 ] .新型光合植被指数(NIRV )指的是归一化植被指数与近红外波段反射率的乘积,与NDVI相比,NIRV 可以在高生物量区域使用,有效减轻了植被-土壤混合像元的影响[28 ] .本研究选取以上两种遥感指数对皇甫川流域的植被地物进行提取,两者在不同覆盖度植被提取上互补缺点,一定程度上减少了皇甫川流域植被的提取误差. ...
植被物候参数遥感提取与验证方法研究进展
1
2022
... 归一化差值植被指数(NDVI)是检测植被生长状态、植被覆盖度重要遥感指数[27 ] .在低密度植被覆盖区域,NDVI具有非常敏感的观测效果[26 ] .新型光合植被指数(NIRV )指的是归一化植被指数与近红外波段反射率的乘积,与NDVI相比,NIRV 可以在高生物量区域使用,有效减轻了植被-土壤混合像元的影响[28 ] .本研究选取以上两种遥感指数对皇甫川流域的植被地物进行提取,两者在不同覆盖度植被提取上互补缺点,一定程度上减少了皇甫川流域植被的提取误差. ...
Remote-sensing extraction of small water bodies on the Loess Plateau
1
2023
... 生物结皮指数的阈值遥感确定是提取过程中的关键;张元明等[14 ] 通过野外测量的生物结皮的高光谱数据确定BSCI的阈值上下限,但是该值存在显著的时空差异.Guo等[29 ] 在提取黄土高原小水体时设定了一定的水体提取规则从而确定提取水体的阈值方法,该方法能够实现阈值动态调整,具有较好的时空适宜性.因此,本研究参考水体提取规则设定适用于裸土、植被、生物结皮的提取规则,即对于SAVI、MSAVI指数,裸土信号值(分布密度)大于非裸土信号值的像元判定为裸土,反之则判定为非裸土;对于NDVI、NIRV 指数,植被信号值(分布密度)大于非植被信号值的像元判定为植被,反之则判定为非植被;对于BSCI指数,生物结皮信号值(分布密度)大于非生物结皮信号值的像元判定为生物结皮,反之则判定为非生物结皮.通过以上提取规则的确定,逐步提取分离目标地物,最终得到生物结皮的上下限阈值. ...
Dryland photoautotrophic soil surface communities endangered byglobal change
2
2018
... 生物结皮的光自养表层群落对土壤稳定以及区域和全球尺度上的水分、养分和微量气体循环至关重要[30 ] ,但是目前空间数据较为缺乏,仅有全球一景的粗分辨率(0.5°)的空间数据[30 ] ,缺乏区域尺度精细分辨率的遥感生物结皮提取方法. ...
... [30 ],缺乏区域尺度精细分辨率的遥感生物结皮提取方法. ...
植物物候遥感监测精度影响因素研究综述
1
2016
... 但该方法也存在一定的局限性.皇甫川流域地形起伏较大且多山,遥感影像中存在的山体阴影限制了地表参数遥感提取的准确性[31 ] ,在逐步提取的过程中,仍然有将山体阴影误认为目标地物的情况,从而影响了目标地物提取与分离的精度与准确性.此外,在遥感影像区分绿色植被与生物结皮、裸土与生物结皮时可能有误分现象,由于不同地物的斑块边缘存在大量混合像元[32 ] ,同时,生物结皮存在厚度、类型差异,使得存在大量的异物同谱现象普遍[33 ] ,这可能导致在多指数阈值分割中本研究以提取规则建立的判别标准错分目标地物的现象. ...
遥感影像混合像元分解中的端元选择方法综述
1
2007
... 但该方法也存在一定的局限性.皇甫川流域地形起伏较大且多山,遥感影像中存在的山体阴影限制了地表参数遥感提取的准确性[31 ] ,在逐步提取的过程中,仍然有将山体阴影误认为目标地物的情况,从而影响了目标地物提取与分离的精度与准确性.此外,在遥感影像区分绿色植被与生物结皮、裸土与生物结皮时可能有误分现象,由于不同地物的斑块边缘存在大量混合像元[32 ] ,同时,生物结皮存在厚度、类型差异,使得存在大量的异物同谱现象普遍[33 ] ,这可能导致在多指数阈值分割中本研究以提取规则建立的判别标准错分目标地物的现象. ...
遥感图像异物同谱干扰消除技术研究与仿真
1
2013
... 但该方法也存在一定的局限性.皇甫川流域地形起伏较大且多山,遥感影像中存在的山体阴影限制了地表参数遥感提取的准确性[31 ] ,在逐步提取的过程中,仍然有将山体阴影误认为目标地物的情况,从而影响了目标地物提取与分离的精度与准确性.此外,在遥感影像区分绿色植被与生物结皮、裸土与生物结皮时可能有误分现象,由于不同地物的斑块边缘存在大量混合像元[32 ] ,同时,生物结皮存在厚度、类型差异,使得存在大量的异物同谱现象普遍[33 ] ,这可能导致在多指数阈值分割中本研究以提取规则建立的判别标准错分目标地物的现象. ...
生物土壤结皮发育及其影响因素研究进展
1
2017
... 生物结皮是全球干旱半干旱生态系统普遍存在的生物地被物,约占陆地总面积的40%,是该生态系统最为活跃的土壤关键层[34 ] ,生物结皮对干旱等逆境有天然的适应能力,具有重要的生态功能和环境效应[4 ] .皇甫川流域是干旱半干旱区黄土高原典型流域,本研究基于精细化的生物结皮数据,发现该流域生物结皮基本呈现破碎化分布,空间分布显著受降水、地形、土地利用等影响. ...
生物土壤结皮对全球气候变化的响应
1
2008
... 降水、气温与地形对生物结皮存在交互影响;生物结皮生长范围广泛,主要类型包括苔藓、地衣、藻类植物等[2 ] ,生物结皮多喜好阴凉潮湿环境.因此,生物结皮对降水的变化反应极其敏感,在干旱半干旱区随着气温的升高和夏季降水频率的增多,生物结皮的生物多样性急剧下降[35 ] .全球气候变化导致美国麦哈韦沙漠和科罗拉多州高原夏季降水增多,而夏季降水增多,促使了某些种类的生物结皮盖度下降,生理功能改变.大气温度是影响结皮形成、空间分布特征的另一个关键因子,学者通过对最近22年来的329种地衣的物种组成变化和分布变化研究,发现随全球变暖喜温生物结皮在荷兰的种类增多而耐寒地衣种减少[36 ] .同时,不同地形因坡度和坡向的差异显著影响,降水和温度的分布格局、阴坡气温高于阳坡、不同海拔间降水存在显著差异,这些气候、地形等环境因素显著影响生物结皮的空间分布[15 ,37 ] .此外,流域土地利用方式也影响着生物结皮的形成与分布,这种差异主要体现在林草地以及其他土地利用方式之间,草地生物结皮盖度与林地生物结皮盖度存在一定的差异性[38 ] ,而林草地生物结皮的分布与其他土地利用类型生物结皮的分布也存在着一定的差异性. ...
Long-term monitoring in the Netherlands suggests that lichens respond to global warming
1
2002
... 降水、气温与地形对生物结皮存在交互影响;生物结皮生长范围广泛,主要类型包括苔藓、地衣、藻类植物等[2 ] ,生物结皮多喜好阴凉潮湿环境.因此,生物结皮对降水的变化反应极其敏感,在干旱半干旱区随着气温的升高和夏季降水频率的增多,生物结皮的生物多样性急剧下降[35 ] .全球气候变化导致美国麦哈韦沙漠和科罗拉多州高原夏季降水增多,而夏季降水增多,促使了某些种类的生物结皮盖度下降,生理功能改变.大气温度是影响结皮形成、空间分布特征的另一个关键因子,学者通过对最近22年来的329种地衣的物种组成变化和分布变化研究,发现随全球变暖喜温生物结皮在荷兰的种类增多而耐寒地衣种减少[36 ] .同时,不同地形因坡度和坡向的差异显著影响,降水和温度的分布格局、阴坡气温高于阳坡、不同海拔间降水存在显著差异,这些气候、地形等环境因素显著影响生物结皮的空间分布[15 ,37 ] .此外,流域土地利用方式也影响着生物结皮的形成与分布,这种差异主要体现在林草地以及其他土地利用方式之间,草地生物结皮盖度与林地生物结皮盖度存在一定的差异性[38 ] ,而林草地生物结皮的分布与其他土地利用类型生物结皮的分布也存在着一定的差异性. ...
陕北黄土区退耕还林地生物结皮分布及其影响因子研究
1
2007
... 降水、气温与地形对生物结皮存在交互影响;生物结皮生长范围广泛,主要类型包括苔藓、地衣、藻类植物等[2 ] ,生物结皮多喜好阴凉潮湿环境.因此,生物结皮对降水的变化反应极其敏感,在干旱半干旱区随着气温的升高和夏季降水频率的增多,生物结皮的生物多样性急剧下降[35 ] .全球气候变化导致美国麦哈韦沙漠和科罗拉多州高原夏季降水增多,而夏季降水增多,促使了某些种类的生物结皮盖度下降,生理功能改变.大气温度是影响结皮形成、空间分布特征的另一个关键因子,学者通过对最近22年来的329种地衣的物种组成变化和分布变化研究,发现随全球变暖喜温生物结皮在荷兰的种类增多而耐寒地衣种减少[36 ] .同时,不同地形因坡度和坡向的差异显著影响,降水和温度的分布格局、阴坡气温高于阳坡、不同海拔间降水存在显著差异,这些气候、地形等环境因素显著影响生物结皮的空间分布[15 ,37 ] .此外,流域土地利用方式也影响着生物结皮的形成与分布,这种差异主要体现在林草地以及其他土地利用方式之间,草地生物结皮盖度与林地生物结皮盖度存在一定的差异性[38 ] ,而林草地生物结皮的分布与其他土地利用类型生物结皮的分布也存在着一定的差异性. ...
黄土高原生物土壤结皮分布时空分异特征
1
2024
... 降水、气温与地形对生物结皮存在交互影响;生物结皮生长范围广泛,主要类型包括苔藓、地衣、藻类植物等[2 ] ,生物结皮多喜好阴凉潮湿环境.因此,生物结皮对降水的变化反应极其敏感,在干旱半干旱区随着气温的升高和夏季降水频率的增多,生物结皮的生物多样性急剧下降[35 ] .全球气候变化导致美国麦哈韦沙漠和科罗拉多州高原夏季降水增多,而夏季降水增多,促使了某些种类的生物结皮盖度下降,生理功能改变.大气温度是影响结皮形成、空间分布特征的另一个关键因子,学者通过对最近22年来的329种地衣的物种组成变化和分布变化研究,发现随全球变暖喜温生物结皮在荷兰的种类增多而耐寒地衣种减少[36 ] .同时,不同地形因坡度和坡向的差异显著影响,降水和温度的分布格局、阴坡气温高于阳坡、不同海拔间降水存在显著差异,这些气候、地形等环境因素显著影响生物结皮的空间分布[15 ,37 ] .此外,流域土地利用方式也影响着生物结皮的形成与分布,这种差异主要体现在林草地以及其他土地利用方式之间,草地生物结皮盖度与林地生物结皮盖度存在一定的差异性[38 ] ,而林草地生物结皮的分布与其他土地利用类型生物结皮的分布也存在着一定的差异性. ...





 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}