


0 引言
草地是全球分布面积最大的陆地生态系统,具有重要的生态和社会功能[10]。近些年在沙化草地实施飞播补播、围栏封育和刈割等恢复措施,使得沙化草地情况有所好转[11-13]。围封作为应用面积最广的修复措施,不仅有助于恢复草地物种多样性、提升生产力,而且有利于土壤结构与功能的维持[14]。但目前关于围封对沙化草地影响的研究主要关于物种多样性以及土壤理化性质对短期围封的变化规律[15-17],而植物功能群对长期围封的响应研究较少,且在长期围封下植物功能群与土壤因子间的关系有待进一步研究。基于此,本研究以内蒙古锡林郭勒沙化草地为研究区域,通过分析围封10年后锡林郭勒沙化草地植物群落和土壤理化性质的变化特征,探讨草地植物群落与功能群对围封响应的差异,以期揭示长期围封条件下土壤因子对植物群落和功能群的影响。
1 材料与方法
1.1 研究区概况
研究区位于内蒙古自治区锡林郭勒盟苏尼特右旗(42°48′04″N、112°43′27″E),属于中温带亚干旱大陆性季风气候。平均气温4.3 ℃,年降水量215 mm。研究区土壤类型为地带性分布的淡栗钙土,地表沙化明显。植物建群种为短花针茅(Stipa breviflora),优势种为无芒隐子草(Cleistogenes songorica)和碱韭(Allium polyrhizum),偶见有狭叶锦鸡儿(Caragana stenophylla)等灌木及半灌木。
1.2 试验设计和样品采集
1.2.1 试验设计
试验始于2011年,分别设置围封和对照样地,样地内地势平坦,样地围封时植物以短花针茅、无芒隐子草等多年生禾草为主,且土地利用方式为自由放牧。围封样地外围用铁丝网及水泥桩设置围栏,面积约3.3 hm2。围封样地全年禁牧,并选取受干扰程度、土壤条件和气候条件基本一致的平行坡面作为对照样地,对照样地为自由放牧且不设置任何保护措施。根据群落植物的生活型组成,将研究区域的群落物种分为4个功能群:多年生禾草、灌木及半灌木、多年生杂类草以及一、二年生草本[18]。
1.2.2 植物和土壤样品采集
截至2021年,样区在围封处理下已禁牧10年,于2021年8月对样地进行植被调查和样品采集。在围封与对照样样地分别随机选取4个小区,每个小区面积为50 m×50 m,每个小区选用1 m×1 m的样方框随机取样方3个,记录样方内植物种类、盖度、密度、高度等信息。齐地面刈割样方内所有植株的地上部分,分种装入信封内,在65 ℃恒温下烘干至恒重,测量干重并记录生物量。在各样方内随机选取3处取样点进行土壤取样,按照0~5、5~10、10~20 cm分层,每层取3个重复土样,称重记录土壤鲜重,称重后将土样带回实验室在105 ℃烘箱中烘干至恒重,用于计算土壤含水率;同时部分鲜土自然风干后用于测定土壤理化性质。
1.3 土壤样品测定
土壤粒径占比采用激光粒度分析仪测定;土壤pH值采用水土比为5∶1的悬液用电势测定法测定;土壤有机质采用重铬酸钾氧化-外加热法测定;土壤全碳采用元素分析仪(VELP CN 802)测定;全氮测定采用微量凯氏定氮法;速效氮测定采用碱解-扩散法;全磷测定采用硫酸-高氯酸消煮法;有效磷测定采用碳酸氢钠浸提-钼锑抗分光光度法;全钾测定采用氢氧化钠熔融-火焰光度计法;速效钾测定采用醋酸铵浸提-火焰光度法测定。
1.4 植物群落基本特征及多样性指数
Shannon-Wiener物种多样性指数(H):
Simpson 物种优势度指数(D):
Pielou 物种均匀度指数(J):
Margalef物种丰富度指数(M):
1.5 数据处理与统计分析
本研究采用Microsoft Excel 2019软件对数据进行预处理,对各数据进行正态分布,对于不符合正态分布的变量进行数据转换,使其更满足正态分布。在IBM SPSS Statistics 22软件中进行单因素方差分析和多重比较。利用R4.3.2软件中的vegan包进行冗余分析(RDA),利用linkET包进行Mantel检验分析以及相关性检验。文中分析结果利用ggplot2包进行绘图。
2 结果与分析
2.1 围封对植物群落组成的影响
调查区共记录到23种植物,围封有17种,各物种重要值0.0050~0.2274,优势种为糙隐子草(Cleistogenes squarrosa)、银灰旋花(Convolvulus ammannii)和蒺藜(Tribulus terrestris),其重要值分别为0.2274、0.1314和0.1241(表1)。对照有22种,各物种重要值为0.0036~0.3390,优势种为短花针茅、糙隐子草和无芒隐子草,其重要值分别为0.3390、0.1254和0.0900。
表1 围封处理下锡林郭勒沙化草地植物群落组成的变化
Table 1
| 编号 | 物种 | 拉丁学名 | 功能群 | 物种重要值 | 功能群重要值 | |||
|---|---|---|---|---|---|---|---|---|
| 围封 | 对照 | 围封 | 对照 | |||||
| 1 | 无芒隐子草 | Cleistogenes songorica | 多年生禾草 | — | 0.0900 | 0.4475 | 0.5734 | |
| 2 | 短花针茅 | Stipa breviflora | 0.1028 | 0.3390 | ||||
| 3 | 糙隐子草 | Cleistogenes squarrosa | 0.2274 | 0.1254 | ||||
| 4 | 稗草 | Echinochloa crusgalli | 0.0192 | 0.0117 | ||||
| 5 | 寸草苔 | Carex duriuscula | 0.0981 | 0.0073 | ||||
| 6 | 狗尾草 | Setaria viridis | 一、二年生草本 | — | 0.0085 | 0.1509 | 0.1223 | |
| 7 | 栉叶蒿 | Neopallasia pectinata | — | 0.0195 | ||||
| 8 | 蒺藜 | Tribulus terrestris | 0.1241 | 0.0604 | ||||
| 9 | 猪毛菜 | Kali collinum | 0.0102 | 0.0066 | ||||
| 10 | 画眉草 | Eragrostis pilosa | 0.0166 | 0.0273 | ||||
| 11 | 草麻黄 | Ephedra sinica | 灌木及半灌木 | 0.0289 | 0.0503 | 0.1343 | 0.1022 | |
| 12 | 木地肤 | Bassia prostrata | 0.0126 | 0.0049 | ||||
| 13 | 狭叶锦鸡儿 | Caragana stenophylla | 0.0928 | 0.0470 | ||||
| 14 | 碱韭 | Allium polyrhizum | 多年生杂类草 | 0.0221 | 0.0253 | 0.2672 | 0.2019 | |
| 15 | 细叶韭 | Allium tenuissimum | 0.0050 | 0.0163 | ||||
| 16 | 蒙古韭 | Allium mongolicum | 0.0327 | 0.0269 | ||||
| 17 | 银灰旋花 | Convolvulus ammannii | 0.1314 | 0.0518 | ||||
| 18 | 刺旋花 | Convolvulus tragacanthoides | — | 0.0065 | ||||
| 19 | 鹤虱 | Lappula myosotis | — | 0.0036 | ||||
| 20 | 北芸香 | Haplophyllumdauricum | 0.0136 | 0.0172 | ||||
| 21 | 冬青叶兔唇花 | Lagochilus ilicifolius | — | 0.0066 | ||||
| 22 | 天门冬 | Asparagus cochinchinensis | 0.0394 | 0.0477 | ||||
| 23 | 支枝鸦葱 | Takhtajaniantha austriaca | 0.0230 | — | ||||
围封在一定程度改变了研究区内原有的植物优势功能群。各功能群的重要值为其所包含植物的重要值之和,围封10年后,多年生禾草重要值由0.5734降至0.4475,但依旧为围封与对照的优势功能群。其他功能群重要值均有所增加,多年生杂类草重要值由0.2019增加至0.2672,一、二年生草本重要值由0.1223增加至0.1509,灌木及半灌木重要值由0.1022增加至0.1343。
2.2 围封对植物群落和功能群数量特征的影响
植物群落和功能群的高度、盖度、密度在围封处理下均存在差异(图1)。围封显著提高植物群落高度、盖度以及密度。多年生禾草高度和一、二年生草本盖度表现为围封显著高于对照,多年生禾草盖度表现为对照显著高于围封。
图1
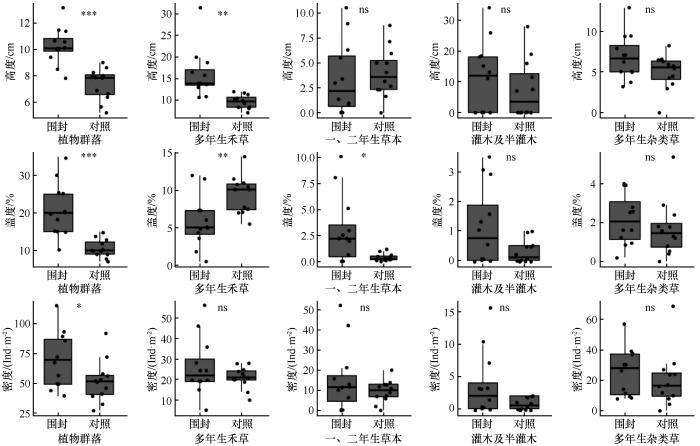
图1
围封处理下植物群落和功能群高度、盖度和密度的对比
Fig.1
Comparison of height, coverage and density of plant communities and functional groups under enclosure treatment
2.3 围封对植物群落和功能群生物量的影响
图2
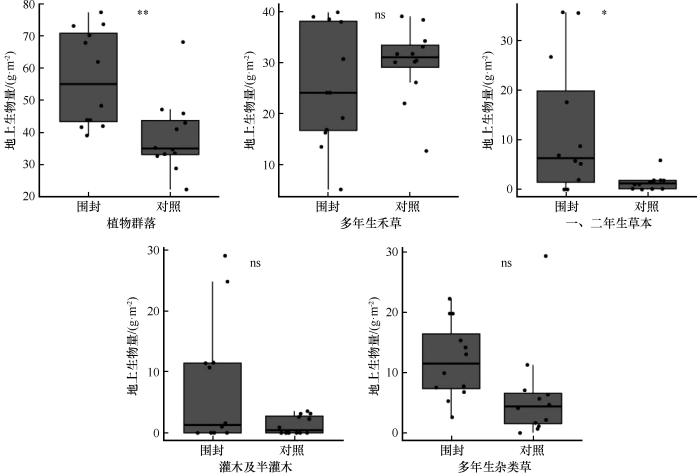
图2
围封处理下地上生物量的变化
Fig.2
Changes in aboveground biomass under enclosure treatment
图3
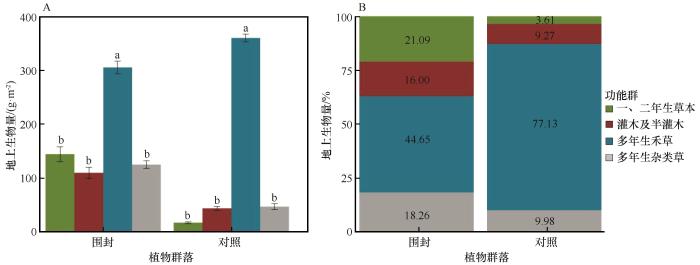
图3
围封处理下植物各功能群地上生物量(A)及其占比(B)变化
Fig.3
Changes in the aboveground biomass of plant functional groups (A) and their proportional variations (B) under enclosure treatment
2.4 围封对植物群落和功能群多样性的影响
长期围封对植物群落的物种多样性和功能群多样性表现出差异性影响(图4)。围封显著降低了植物群落的Shannon-Wiener多样性指数(P<0.05)、Simpson优势度指数(P<0.05)以及Pielou均匀度指数(P<0.05),较对照样地显著降低了14.10%、9.40%以及9.57%。而功能群的Margalef丰富度指数、Shannon-Wiener指数、Simpson指数和Pielou均匀度指数的观测值较对照均呈现上升趋势,但上升趋势不显著(P>0.05),但这一致性变化趋势表明围封可能通过优先保障关键功能群的发育,淘汰竞争能力弱的物种,以维持生态系统的功能稳定性。
图4
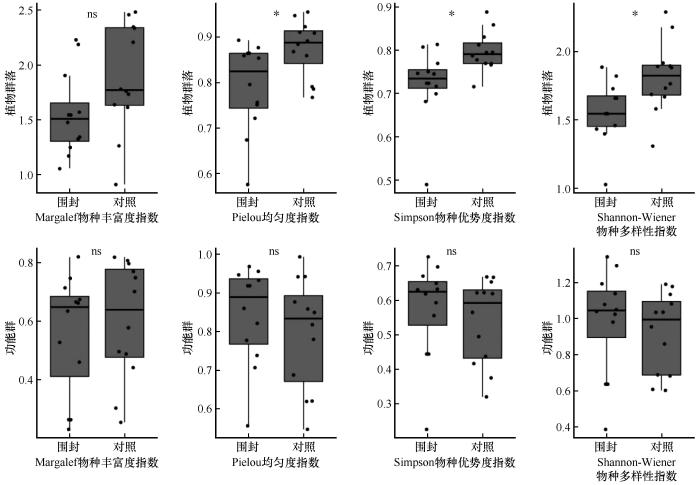
图4
围封处理下植物群落和功能群物种多样性指数
Fig.4
Species diversity index of plant communities and functional groups under enclosure treatment
2.5 围封对土壤因子的影响
围封处理下土壤因子发生不同程度的变化(表2),围封显著提高各土层全碳、有机质、全氮和水解性氮含量,总含量分别提高26.31%、22.78%、16.08%和10.17%,降低各土层土壤全磷含量,总含量降低8.08%。其中,10~20 cm土层全磷含量差异显著;在0~5 cm土层有效磷含量显著降低,而在5~10、10~20 cm土层显著升高;围封降低10~20 cm土层的粉粒含量;土壤含水率、pH、砂粒等其他土壤理化指标含量在不同土层均无显著差异。各样地0~5 cm土壤含水率均显著低于5~10 cm和10~20 cm土层,全磷含量表现为10~20 cm土层显著高于其他土层;在围封样地中,有效磷含量表现为0~5 cm显著低于其他土层;pH、全氮等其他指标含量在各样地不同土层中均无显著差异(P>0.05)。
表2 围封处理下土壤理化性质变化( N=108×2)
Table 2
| 土壤理化指标 | 围封 | 对照 | ||||
|---|---|---|---|---|---|---|
| 0~5 cm | 5~10 cm | 10~20 cm | 0~5 cm | 5~10 cm | 10~20 cm | |
| 含水率 | 2.00±1.03Ab | 2.83±0.48Aa | 2.79±0.59Aa | 1.67±0.54Ab | 2.91±0.87Aa | 3.64±1.09Aa |
| pH值 | 8.16±0.37Aa | 8.17±0.31Aa | 8.18±2.95Aa | 8.29±0.03Aa | 8.28±0.03Aa | 8.27±0.06Aa |
| 全碳/(g·kg-1) | 11.01±1.73Aa | 11.14±2.07Aa | 11.12±1.64Aa | 9.00±0.78Ba | 8.51±1.46Ba | 8.83±1.09Ba |
| 有机质/(g·kg-1) | 14.60±1.85Aa | 15.37±2.46Aa | 15.03±1.54Aa | 12.51±1.94Ba | 11.61±1.23Ba | 12.53±1.27Ba |
| 全磷/(g·kg-1) | 0.29±0.02Ab | 0.30±0.01Ab | 0.32±0.01Ba | 0.31±0.04Ab | 0.33±0.03Aab | 0.35±0.02Aa |
| 全钾/(g·kg-1) | 21.49±0.73Aa | 21.17±0.60Aa | 21.79±0.62Aa | 21.27±2.10Aa | 21.01±1.79Aa | 20.79±1.71Aa |
| 全氮/(g·kg-1) | 0.99±0.08Aa | 0.99±0.08Aa | 0.98±0.09Aa | 0.91±0.06Ba | 0.84±0.07Bab | 0.80±0.07Bb |
| 有效磷/(mg·kg-1) | 6.88±1.22Bb | 10.08±0.83Aa | 10.31±0.55Aa | 9.61±0.56Aa | 10.01±0.71Ba | 9.60±0.64Ba |
| 速效钾/(mg·kg-1) | 106.78±12.58Aa | 106.00±12.54Aa | 106.11±12.09Aa | 107.78±17.54Aa | 114.11±18.57Aa | 115.78±18.97Aa |
| 水解性氮/(mg·kg-1) | 56.33±4.06Aa | 53.33±4.16Aa | 56.33±3.07Aa | 49.78±5.80Ba | 51.67±4.27Ba | 49.22±4.44Ba |
| 砂粒(0.05~2 mm)/% | 81.28±0.98Aa | 81.59±0.65Aa | 81.51±0.69Aa | 77.52±0.61Aa | 77.69±0.70Aa | 77.77±0.68Aa |
| 粉粒(0.02~0.05mm)/% | 10.39±0.16Aa | 10.21±0.32Aa | 10.24±0.12Ba | 12.00±1.14Aa | 11.65±1.11Aa | 11.61±1.12Aa |
| 黏粒(<0.02 mm)/% | 8.33±0.87Aa | 8.38±0.79Aa | 8.26±0.69Aa | 10.48±1.58Aa | 10.67±1.67Aa | 10.61±1.71Aa |
2.6 土壤因子与植物群落的关系
2.6.1 植物群落与土壤环境因子冗余分析
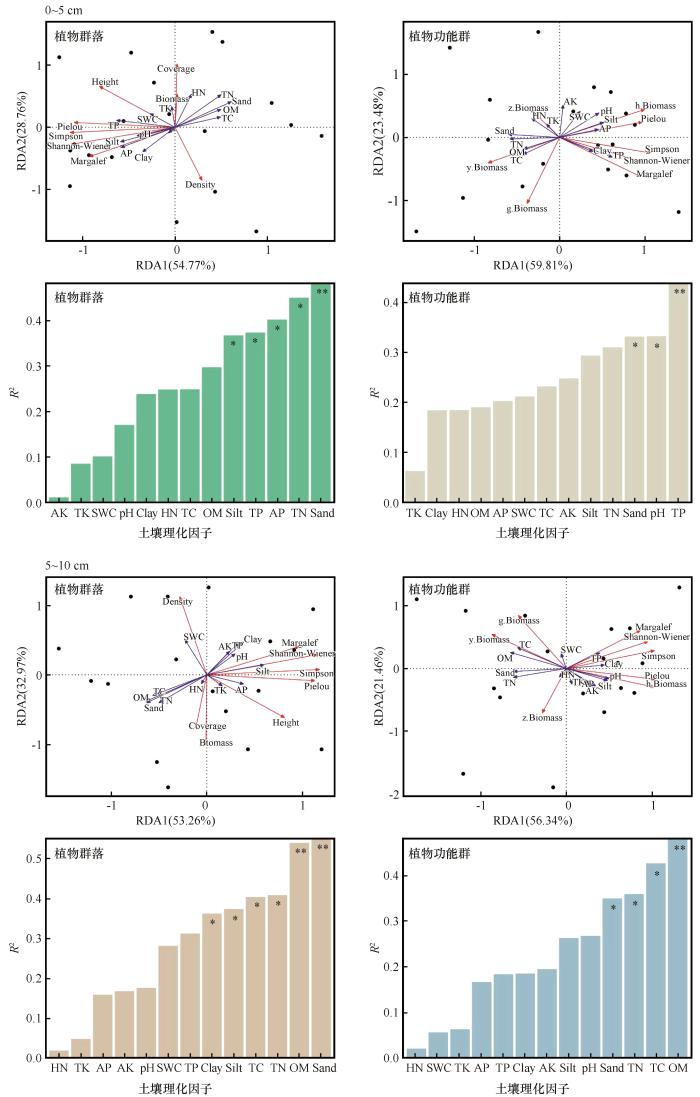
分别对各土层土壤因子与植物群落和功能群指标进行冗余分析得到每个解释变量对响应变量的解释率(图5)。0~5 cm土层前两轴对植物群落和功能群的累计解释率分别为83.53%和83.29%,5~10 cm土层前两轴对植物群落和功能群解释率分别为86.23%和77.80%,10~20 cm土层前两轴对植物群落和功能群累计解释率分别为84.77%和80.72%。对土壤因子envfit函数检验分析显示,各土层砂粒以及全氮均显著影响了植物群落与功能群特征;除此之外,有效磷和全磷含量在0~5 cm土层对植物群落和功能群特征影响显著;有机质和全碳含量在5~10 cm和10~20 cm土层显著影响植物群落和功能群特征。
图5
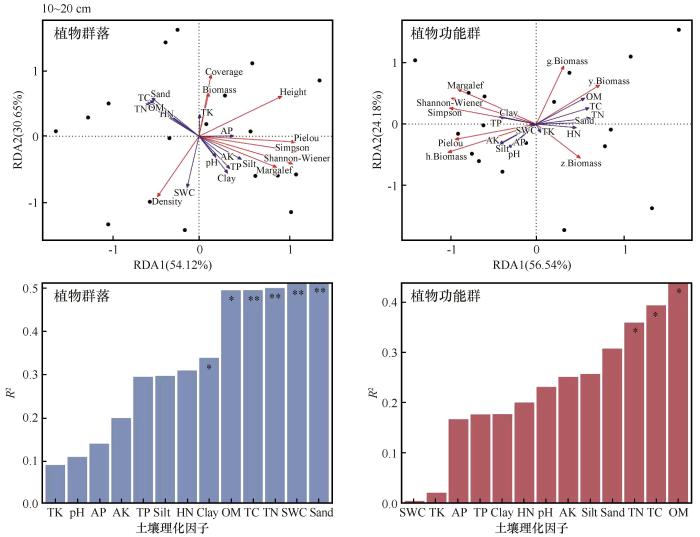
图5
围封处理下不同土层土壤因子与植物群落和功能群的冗余分析(RDA)以及各因子对植物群落和功能群的解释度
Fig.5
Redundancy analysis (RDA) of soil factors and plant communities and functional groups under enclosure treatment, and the explanatory degree of each factor on plant communities and functional groups
2.6.2 植物群落结构和功能指标与土壤理化因子的Mantel检验和组间分析
分别对各土层土壤因子与植物群落结构和功能指标进行Mantel检验和组间相关性分析结果可知(图6),植物群落基本特征与各土层砂粒、粉粒、有效磷均存在显著相关关系;植物群落多样性与各土层全氮存在显著相关关系;功能群生物量与各土层全碳、有机质以及pH均存在显著相关性;功能多样性与各土壤因子均无显著的相关关系。对植物群落指标与土壤因子进行相关性分析,所得结果在一定程度上验证了RDA和Mantel分析结果。在各土层中,砂粒与植物群落高度和盖度呈显著正相关关系,pH与功能群生物量呈负相关;在0~5 cm土层有效磷与盖度和密度呈显著负相关关系;在5~10 cm土层和10~20 cm土层中,全氮、砂粒、全碳和有机质与植物多样性指数负相关,与高度、盖度和一、二年生草本生物量显著正相关。随着土层加深,植物群落指标与土壤因子的显著相关性关系数量增多。
图6
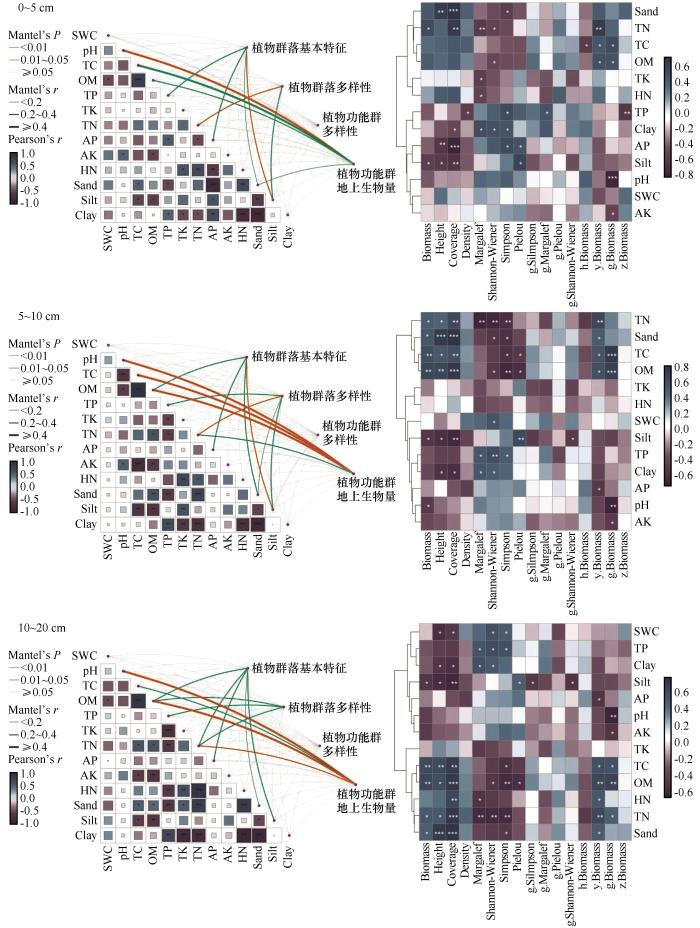
图6
围封处理下不同土层土壤因子与植物群落和功能群的Mantel检验和相关性分析
Fig.6
Mantel test and correlation analysis between soil factors and plant communities and functional groups under enclosure treatment
3 讨论
3.1 沙化草地植物群落特征对围封的响应
植物群落的变化可以较为直观地反映出样地生态系统的稳定性和可持续性[21]。本研究发现,围封10年后,植物群落整体高度、盖度、密度及生物量均呈现显著增加趋势(图1),该结果与吴婷等[22]、赵哈林等[23]研究一致。原因可能为围封消除了人类活动及家畜对地上植物的干扰,生态系统自我更新能力增强,植物种群得以快速繁殖、生长与发育,植被逐渐恢复[24-25]。但功能群层面表现出明显分异:为多年生禾草功能群的盖度和生物量显著降低,而一、二年生草本功能群的盖度和生物量则分别显著增加。一方面可能是由于一、二年生植物具有较强的入侵能力,能够利用短期降水快速生长,在一定程度上抑制了多年生禾草对光、水等资源的竞争[26];另一方面,草层高度的增加也导致竞争能力弱、低矮的禾草逐渐失去优势并消失[27]。
3.2 沙化草地不同土层土壤理化性质对围封的响应
与对照样地相比,围封样地各土层土壤全碳、有机质、全氮和水解性氮含量均显著提高(表2),该结论与Zhang等[32]、闫瑞瑞[25]研究结果一致。其原因一方面可能为围封样地枯落物积累量较多,植物枯落物是土壤中营养物质的主要来源,使得土壤中全碳和有机质含量较高;另一方面围封后植被恢复会显著增加植物根际分泌物的种类和数量,这些分泌物通过激活微生物代谢活动促进土壤中养分的释放,从而提高土壤肥力[33-34]。其中,全氮含量和水解性氮含量显著增加,也可能与狭叶锦鸡儿生物量增多有关,狭叶锦鸡儿属于豆科植物,其固氮作用使土壤中氮含量得到补充[35]。除此之外,围封处理下土壤含水率无显著变化。这可能是由于群落盖度仅为20%左右,土壤大面积裸露,表层土壤水分蒸发强烈[27]。
3.3 沙化草地土壤因子与植物群落的关系
草地植物群落和土壤因子之间存在着反馈效应,土壤因子可以影响植物种间关系,造成植物群落发生变化,从而促进沙化草地恢复[36-37]。目前研究结果表明,围封处理下引起的土壤氮磷含量的改变会进一步影响植物化学计量学特征[38]。本研究结果表明,沙化草地各土层砂粒和全氮含量与植物群落指标关系紧密并且是影响其变化的主要因子,原因可能是沙化草地地表裸露率高,土壤主要为砂粒,土壤粒径的变化会直接影响土壤理化性质,进而影响植物的生长发育[39-41],同时围封增加了枯落物和土壤养分含量,增加了土壤的持水性能[42],促进植物生长从而影响植物群落发展[43]。此外,相关研究表明,水分是环境因子中影响干旱区植物地理分布的主导因子[44],但本研究中,各土层土壤含水率对植物群落各指标的解释度均较低,这可能与研究区为沙化草地,土壤存水能力差及土壤水分快速流失有关。
不同土层土壤因子与植物群落各指标存在不同程度的关系。0~5 cm土层pH与功能群生物量表现出显著负相关关系,5~10 cm和10~20 cm土层全碳和有机质与功能群生物量表现出显著正相关关系,原因可能在于沙化草地土壤偏碱性,枯落物在分解过程中积累了大量腐殖酸,降低表层土壤碱化程度,并且枯落物输入对土壤全氮和有机碳有正效应,影响植物地上生物量[43,45]。全磷和有效磷在0~5 cm与植物群落高度和盖度呈显著负相关关系,这可能是磷元素在土壤水分的移动下迅速转移,导致表层土壤磷含量降低,围封增加了地上生物量,从而增大了地上植物对磷的需要[46]。除此之外,10~20 cm土层土壤因子与各植物指标的显著性较高。原因可能为在沙化草地由于表层土壤含水量较低,植被根系多聚集在10~20 cm土层中,因此该土层土壤因子对植物指标相关性较高,从而对植物生长发育有显著影响[47]。
4 结论
对围封10年的沙化草地生态系统恢复研究表明,尽管植物群落物种多样性下降,但地上生物量显著提升及功能群多样性增强驱动了生态恢复进程。围封显著改善土壤理化性质,全碳、有机质、总氮和水解性氮含量显著增加,其中在0~5 cm土层,pH和有效磷与植被指标存在显著相关性,全碳和有机质在5~20 cm土层对功能群的影响更为关键,各土层砂粒和全氮与植被特征均显著相关。这表明围封措施促进了植物群落结构与土壤环境的恢复,改善土壤理化环境,增强功能群多样性,以维持植物群落结构特征稳定,从而促进沙化草地自然恢复。
参考文献
Grazing exclusion promotes grasses functional group dominance via increasing of bud banks in steppe community
[J].
Benefits derived from rehabilitating a degraded semi‐arid rangeland in private enclosures in West Pokot County,Kenya
[J].
Long‐term grazing exclusion effects on vegetation characteristics,soil properties and bacterial communities in the semi‐arid grasslands of China
[J].
Grassland soil carbon sequestration: Current understanding,challenges,and solutions
[J].
围封对辽西北退化草地土壤有机碳含量的影响
[J].
Linking fungal diversity with soil water repellency through grassland enclosure in Loess Hilly Region
[J].
Reproductive responses of alpine plants to grassland degradation and artificial restoration in the Qinghai‐Tibetan Plateau
[J].
Effect of grazing on community structure and productivity of a Uruguayan grassland
[J].
Recovery of plant species composition and ecosystem function after cessation of grazing in a Mediterranean grassland
[J].
Plant diversity,soil biota and resistance to exotic invasion
[J].
Evolutionary history of grazing and resources determine herbivore exclusion effects on plant diversity
[J].
Interactions of soil bacteria and fungi with plants during long-term grazing exclusion in semiarid grasslands
[J].
Root-released organic anions in response to low phosphorus availability:recent progress,challenges and future perspectives
[J].
Grassroots ecology: plant-microbe-soil interactions as drivers of plant community structure and dynamics
[J].
乌兰布和沙漠土壤粒径与植物群落多样性特征分析
[J].

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}