腾格里沙漠典型固沙灌木生存的生态水文学机理综述
3
2025
... 水分是干旱半干旱区植被建设和生态系统稳定的最关键限制因子[1].在该地区,植物的生存、生长及分布很大程度上取决于其应对水分胁迫的能力与策略[2-3].植物水力系统连接土壤-植物-大气连续体,其功能性状直接影响个体水分利用效率及整体适应性,决定种群动态和群落结构[4].近年来,随着全球气候变化加剧,极端干旱事件频率增加,导致植被退化及林木死亡现象频发[5-7].因此,深入研究植物水力功能对干旱环境的响应与适应机制,对于预测植被动态及指导生态恢复具有重要理论与实践意义. ...
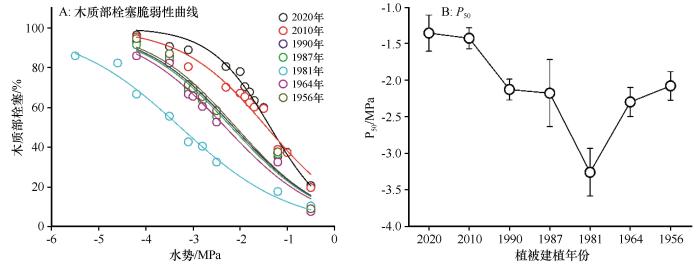
... 研究发现柠条的栓塞脆弱性(P₅₀)随建植年份呈现出非线性的“U”型变化模式.这一模式揭示了柠条在不同生活史阶段,为了适应腾格里沙漠极端干旱环境而进行的策略性调整 [1].在植被建植初期,柠条需要维持较高的水力导度和光合速率,以支持快速的生物量积累.随着柠条进入演替阶段中期,P₅₀值达到最低点,表现出最强的栓塞抗性.此时,植株已经建立了稳定的生态位,生长速率放缓,生存和维持功能稳定成为其主要策略.在腾格里沙漠极端干旱、降水年际波动巨大的环境中,高栓塞抗性意味着植株能够在遭遇极端干旱事件时,依然维持木质部导水系统的完整性,从而保证关键生理活动(如光合作用)的最低水分需求,避免在干旱中死亡[1,23].本研究中观测到-3.25 MPa的P₅₀值,在全球木本植物中属于中高抗性水平,证明了柠条具有强烈的适应腾格里沙漠极端干旱环境的能力.这种抗性的获得可能涉及多种复杂的生理和解剖学机制,例如纹孔膜结构优化、导管壁增厚以增强其抗气穴化能力,以及导管网络的冗余设计等,这些机制共同作用,构建了一个高效且安全的水分传输系统[30].当柠条进入演替阶段后期,其栓塞脆弱性有增加的趋势,P₅₀值增大.这一变化反映了植物在生理衰老过程中,维持高栓塞抗性的能力开始下降.维持高栓塞抗性需要持续的能量和资源投入,包括合成渗透调节物质、维护细胞壁结构、运行抗氧化系统等[30].后期植株的代谢活性普遍下降,可能无法支撑这些高成本的防御机制,其预示着植株抵抗极端干旱事件的能力减弱,是其功能衰退和最终死亡的一个重要信号[38]. ...
... [1,23].本研究中观测到-3.25 MPa的P₅₀值,在全球木本植物中属于中高抗性水平,证明了柠条具有强烈的适应腾格里沙漠极端干旱环境的能力.这种抗性的获得可能涉及多种复杂的生理和解剖学机制,例如纹孔膜结构优化、导管壁增厚以增强其抗气穴化能力,以及导管网络的冗余设计等,这些机制共同作用,构建了一个高效且安全的水分传输系统[30].当柠条进入演替阶段后期,其栓塞脆弱性有增加的趋势,P₅₀值增大.这一变化反映了植物在生理衰老过程中,维持高栓塞抗性的能力开始下降.维持高栓塞抗性需要持续的能量和资源投入,包括合成渗透调节物质、维护细胞壁结构、运行抗氧化系统等[30].后期植株的代谢活性普遍下降,可能无法支撑这些高成本的防御机制,其预示着植株抵抗极端干旱事件的能力减弱,是其功能衰退和最终死亡的一个重要信号[38]. ...
Triggers of tree mortality under drought
1
2018
... 水分是干旱半干旱区植被建设和生态系统稳定的最关键限制因子[1].在该地区,植物的生存、生长及分布很大程度上取决于其应对水分胁迫的能力与策略[2-3].植物水力系统连接土壤-植物-大气连续体,其功能性状直接影响个体水分利用效率及整体适应性,决定种群动态和群落结构[4].近年来,随着全球气候变化加剧,极端干旱事件频率增加,导致植被退化及林木死亡现象频发[5-7].因此,深入研究植物水力功能对干旱环境的响应与适应机制,对于预测植被动态及指导生态恢复具有重要理论与实践意义. ...
Mechanisms of woody-plant mortality under rising drought, CO2 and vapour pressure deficit
1
2022
... 水分是干旱半干旱区植被建设和生态系统稳定的最关键限制因子[1].在该地区,植物的生存、生长及分布很大程度上取决于其应对水分胁迫的能力与策略[2-3].植物水力系统连接土壤-植物-大气连续体,其功能性状直接影响个体水分利用效率及整体适应性,决定种群动态和群落结构[4].近年来,随着全球气候变化加剧,极端干旱事件频率增加,导致植被退化及林木死亡现象频发[5-7].因此,深入研究植物水力功能对干旱环境的响应与适应机制,对于预测植被动态及指导生态恢复具有重要理论与实践意义. ...
Hydraulic diversity of forests regulates ecosystem resilience during drought
1
2018
... 水分是干旱半干旱区植被建设和生态系统稳定的最关键限制因子[1].在该地区,植物的生存、生长及分布很大程度上取决于其应对水分胁迫的能力与策略[2-3].植物水力系统连接土壤-植物-大气连续体,其功能性状直接影响个体水分利用效率及整体适应性,决定种群动态和群落结构[4].近年来,随着全球气候变化加剧,极端干旱事件频率增加,导致植被退化及林木死亡现象频发[5-7].因此,深入研究植物水力功能对干旱环境的响应与适应机制,对于预测植被动态及指导生态恢复具有重要理论与实践意义. ...
Research frontiers for improving our understanding of drought-induced tree and forest mortality
1
2018
... 水分是干旱半干旱区植被建设和生态系统稳定的最关键限制因子[1].在该地区,植物的生存、生长及分布很大程度上取决于其应对水分胁迫的能力与策略[2-3].植物水力系统连接土壤-植物-大气连续体,其功能性状直接影响个体水分利用效率及整体适应性,决定种群动态和群落结构[4].近年来,随着全球气候变化加剧,极端干旱事件频率增加,导致植被退化及林木死亡现象频发[5-7].因此,深入研究植物水力功能对干旱环境的响应与适应机制,对于预测植被动态及指导生态恢复具有重要理论与实践意义. ...
木本植物水力系统对干旱胁迫的响应机制
1
2021
... 水分是干旱半干旱区植被建设和生态系统稳定的最关键限制因子[1].在该地区,植物的生存、生长及分布很大程度上取决于其应对水分胁迫的能力与策略[2-3].植物水力系统连接土壤-植物-大气连续体,其功能性状直接影响个体水分利用效率及整体适应性,决定种群动态和群落结构[4].近年来,随着全球气候变化加剧,极端干旱事件频率增加,导致植被退化及林木死亡现象频发[5-7].因此,深入研究植物水力功能对干旱环境的响应与适应机制,对于预测植被动态及指导生态恢复具有重要理论与实践意义. ...
The hydraulic architecture of trees and other woody plants
1
1991
... 植物的水力结构通常采用导水率(PLC)、边材比导率(Ks)、叶比导率(Kl)和胡伯尔值(Hv)来表征,这些参数分别从不同角度反映了植物的水分运输能力、输水效率以及叶片供水情况[8-9].利用木质部在不同水势下导水率变化得出木质部的脆弱性曲线[10].水力结构参数直接或间接地反映树木水分状况的指标,通过对这些指标的变化规律及其相互关系的研究认识树木的水分生理生态特点,从而阐明树木的耐旱特性及其机理[11].不同演替阶段,植物的水力结构参数呈现出明显的变化趋势[9].例如,贵州喀斯特地区的石漠化先锋植物中,常绿树种占据优势,其木质部导管直径相对较小,以牺牲部分水分传输效率为代价,换取更强的抗栓塞能力,从而在干旱、贫瘠的环境中生存[12].随着演替进入中期阶段,植物种类逐渐丰富,生境条件有所改善但依然存在一定的胁迫因素.此时,植物的水力结构呈现出多样化的适应策略.一些植物通过增加木质部导管直径来提高水分传输效率,以满足快速生长的需求;另一些植物则侧重于增强木质部的抗栓塞能力,以应对可能出现的干旱或冻融等胁迫事件[13].对长白山阔叶红松林的研究发现,不同功能类群的树种在这一阶段展现出明显的水力结构差异.具有木质部正压力(如根压或茎压)的树种能够有效修复冬季冻融循环诱导的气穴化栓塞,保持较高的水分传输效率;而不具备这种修复机制的树种则通过调整导管的形态和密度,寻求水分传输效率与安全性的平衡[14-15].顶级群落阶段植物的水力结构趋于稳定和优化,该阶段的植物通常具有较高的生物量和复杂的生态系统结构,水力结构的演化更加注重整体的协调性和对环境的精细适应.植物木质部导管在直径、长度、数量等方面达到一种动态平衡,既能保证高效的水分传输以支持庞大的生物量和生理活动,又能有效抵御各种环境胁迫[9,16]. ...
浙江天童常绿阔叶林不同演替阶段木本植物的水力结构特征
3
2016
... 植物的水力结构通常采用导水率(PLC)、边材比导率(Ks)、叶比导率(Kl)和胡伯尔值(Hv)来表征,这些参数分别从不同角度反映了植物的水分运输能力、输水效率以及叶片供水情况[8-9].利用木质部在不同水势下导水率变化得出木质部的脆弱性曲线[10].水力结构参数直接或间接地反映树木水分状况的指标,通过对这些指标的变化规律及其相互关系的研究认识树木的水分生理生态特点,从而阐明树木的耐旱特性及其机理[11].不同演替阶段,植物的水力结构参数呈现出明显的变化趋势[9].例如,贵州喀斯特地区的石漠化先锋植物中,常绿树种占据优势,其木质部导管直径相对较小,以牺牲部分水分传输效率为代价,换取更强的抗栓塞能力,从而在干旱、贫瘠的环境中生存[12].随着演替进入中期阶段,植物种类逐渐丰富,生境条件有所改善但依然存在一定的胁迫因素.此时,植物的水力结构呈现出多样化的适应策略.一些植物通过增加木质部导管直径来提高水分传输效率,以满足快速生长的需求;另一些植物则侧重于增强木质部的抗栓塞能力,以应对可能出现的干旱或冻融等胁迫事件[13].对长白山阔叶红松林的研究发现,不同功能类群的树种在这一阶段展现出明显的水力结构差异.具有木质部正压力(如根压或茎压)的树种能够有效修复冬季冻融循环诱导的气穴化栓塞,保持较高的水分传输效率;而不具备这种修复机制的树种则通过调整导管的形态和密度,寻求水分传输效率与安全性的平衡[14-15].顶级群落阶段植物的水力结构趋于稳定和优化,该阶段的植物通常具有较高的生物量和复杂的生态系统结构,水力结构的演化更加注重整体的协调性和对环境的精细适应.植物木质部导管在直径、长度、数量等方面达到一种动态平衡,既能保证高效的水分传输以支持庞大的生物量和生理活动,又能有效抵御各种环境胁迫[9,16]. ...
... [9].例如,贵州喀斯特地区的石漠化先锋植物中,常绿树种占据优势,其木质部导管直径相对较小,以牺牲部分水分传输效率为代价,换取更强的抗栓塞能力,从而在干旱、贫瘠的环境中生存[12].随着演替进入中期阶段,植物种类逐渐丰富,生境条件有所改善但依然存在一定的胁迫因素.此时,植物的水力结构呈现出多样化的适应策略.一些植物通过增加木质部导管直径来提高水分传输效率,以满足快速生长的需求;另一些植物则侧重于增强木质部的抗栓塞能力,以应对可能出现的干旱或冻融等胁迫事件[13].对长白山阔叶红松林的研究发现,不同功能类群的树种在这一阶段展现出明显的水力结构差异.具有木质部正压力(如根压或茎压)的树种能够有效修复冬季冻融循环诱导的气穴化栓塞,保持较高的水分传输效率;而不具备这种修复机制的树种则通过调整导管的形态和密度,寻求水分传输效率与安全性的平衡[14-15].顶级群落阶段植物的水力结构趋于稳定和优化,该阶段的植物通常具有较高的生物量和复杂的生态系统结构,水力结构的演化更加注重整体的协调性和对环境的精细适应.植物木质部导管在直径、长度、数量等方面达到一种动态平衡,既能保证高效的水分传输以支持庞大的生物量和生理活动,又能有效抵御各种环境胁迫[9,16]. ...
... [9,16]. ...
6个耐旱树种木质部结构与栓塞脆弱性的关系
1
2016
... 植物的水力结构通常采用导水率(PLC)、边材比导率(Ks)、叶比导率(Kl)和胡伯尔值(Hv)来表征,这些参数分别从不同角度反映了植物的水分运输能力、输水效率以及叶片供水情况[8-9].利用木质部在不同水势下导水率变化得出木质部的脆弱性曲线[10].水力结构参数直接或间接地反映树木水分状况的指标,通过对这些指标的变化规律及其相互关系的研究认识树木的水分生理生态特点,从而阐明树木的耐旱特性及其机理[11].不同演替阶段,植物的水力结构参数呈现出明显的变化趋势[9].例如,贵州喀斯特地区的石漠化先锋植物中,常绿树种占据优势,其木质部导管直径相对较小,以牺牲部分水分传输效率为代价,换取更强的抗栓塞能力,从而在干旱、贫瘠的环境中生存[12].随着演替进入中期阶段,植物种类逐渐丰富,生境条件有所改善但依然存在一定的胁迫因素.此时,植物的水力结构呈现出多样化的适应策略.一些植物通过增加木质部导管直径来提高水分传输效率,以满足快速生长的需求;另一些植物则侧重于增强木质部的抗栓塞能力,以应对可能出现的干旱或冻融等胁迫事件[13].对长白山阔叶红松林的研究发现,不同功能类群的树种在这一阶段展现出明显的水力结构差异.具有木质部正压力(如根压或茎压)的树种能够有效修复冬季冻融循环诱导的气穴化栓塞,保持较高的水分传输效率;而不具备这种修复机制的树种则通过调整导管的形态和密度,寻求水分传输效率与安全性的平衡[14-15].顶级群落阶段植物的水力结构趋于稳定和优化,该阶段的植物通常具有较高的生物量和复杂的生态系统结构,水力结构的演化更加注重整体的协调性和对环境的精细适应.植物木质部导管在直径、长度、数量等方面达到一种动态平衡,既能保证高效的水分传输以支持庞大的生物量和生理活动,又能有效抵御各种环境胁迫[9,16]. ...
植物木质部水力学研究进展
1
2020
... 植物的水力结构通常采用导水率(PLC)、边材比导率(Ks)、叶比导率(Kl)和胡伯尔值(Hv)来表征,这些参数分别从不同角度反映了植物的水分运输能力、输水效率以及叶片供水情况[8-9].利用木质部在不同水势下导水率变化得出木质部的脆弱性曲线[10].水力结构参数直接或间接地反映树木水分状况的指标,通过对这些指标的变化规律及其相互关系的研究认识树木的水分生理生态特点,从而阐明树木的耐旱特性及其机理[11].不同演替阶段,植物的水力结构参数呈现出明显的变化趋势[9].例如,贵州喀斯特地区的石漠化先锋植物中,常绿树种占据优势,其木质部导管直径相对较小,以牺牲部分水分传输效率为代价,换取更强的抗栓塞能力,从而在干旱、贫瘠的环境中生存[12].随着演替进入中期阶段,植物种类逐渐丰富,生境条件有所改善但依然存在一定的胁迫因素.此时,植物的水力结构呈现出多样化的适应策略.一些植物通过增加木质部导管直径来提高水分传输效率,以满足快速生长的需求;另一些植物则侧重于增强木质部的抗栓塞能力,以应对可能出现的干旱或冻融等胁迫事件[13].对长白山阔叶红松林的研究发现,不同功能类群的树种在这一阶段展现出明显的水力结构差异.具有木质部正压力(如根压或茎压)的树种能够有效修复冬季冻融循环诱导的气穴化栓塞,保持较高的水分传输效率;而不具备这种修复机制的树种则通过调整导管的形态和密度,寻求水分传输效率与安全性的平衡[14-15].顶级群落阶段植物的水力结构趋于稳定和优化,该阶段的植物通常具有较高的生物量和复杂的生态系统结构,水力结构的演化更加注重整体的协调性和对环境的精细适应.植物木质部导管在直径、长度、数量等方面达到一种动态平衡,既能保证高效的水分传输以支持庞大的生物量和生理活动,又能有效抵御各种环境胁迫[9,16]. ...
中亚热带喀斯特常绿落叶阔叶混交林典型树种的木质部解剖与功能特征分析
1
2021
... 植物的水力结构通常采用导水率(PLC)、边材比导率(Ks)、叶比导率(Kl)和胡伯尔值(Hv)来表征,这些参数分别从不同角度反映了植物的水分运输能力、输水效率以及叶片供水情况[8-9].利用木质部在不同水势下导水率变化得出木质部的脆弱性曲线[10].水力结构参数直接或间接地反映树木水分状况的指标,通过对这些指标的变化规律及其相互关系的研究认识树木的水分生理生态特点,从而阐明树木的耐旱特性及其机理[11].不同演替阶段,植物的水力结构参数呈现出明显的变化趋势[9].例如,贵州喀斯特地区的石漠化先锋植物中,常绿树种占据优势,其木质部导管直径相对较小,以牺牲部分水分传输效率为代价,换取更强的抗栓塞能力,从而在干旱、贫瘠的环境中生存[12].随着演替进入中期阶段,植物种类逐渐丰富,生境条件有所改善但依然存在一定的胁迫因素.此时,植物的水力结构呈现出多样化的适应策略.一些植物通过增加木质部导管直径来提高水分传输效率,以满足快速生长的需求;另一些植物则侧重于增强木质部的抗栓塞能力,以应对可能出现的干旱或冻融等胁迫事件[13].对长白山阔叶红松林的研究发现,不同功能类群的树种在这一阶段展现出明显的水力结构差异.具有木质部正压力(如根压或茎压)的树种能够有效修复冬季冻融循环诱导的气穴化栓塞,保持较高的水分传输效率;而不具备这种修复机制的树种则通过调整导管的形态和密度,寻求水分传输效率与安全性的平衡[14-15].顶级群落阶段植物的水力结构趋于稳定和优化,该阶段的植物通常具有较高的生物量和复杂的生态系统结构,水力结构的演化更加注重整体的协调性和对环境的精细适应.植物木质部导管在直径、长度、数量等方面达到一种动态平衡,既能保证高效的水分传输以支持庞大的生物量和生理活动,又能有效抵御各种环境胁迫[9,16]. ...
Divergent hydraulic strategies to cope with freezing in co‐occurring temperate tree species with special reference to root and stem pressure generation
1
2018
... 植物的水力结构通常采用导水率(PLC)、边材比导率(Ks)、叶比导率(Kl)和胡伯尔值(Hv)来表征,这些参数分别从不同角度反映了植物的水分运输能力、输水效率以及叶片供水情况[8-9].利用木质部在不同水势下导水率变化得出木质部的脆弱性曲线[10].水力结构参数直接或间接地反映树木水分状况的指标,通过对这些指标的变化规律及其相互关系的研究认识树木的水分生理生态特点,从而阐明树木的耐旱特性及其机理[11].不同演替阶段,植物的水力结构参数呈现出明显的变化趋势[9].例如,贵州喀斯特地区的石漠化先锋植物中,常绿树种占据优势,其木质部导管直径相对较小,以牺牲部分水分传输效率为代价,换取更强的抗栓塞能力,从而在干旱、贫瘠的环境中生存[12].随着演替进入中期阶段,植物种类逐渐丰富,生境条件有所改善但依然存在一定的胁迫因素.此时,植物的水力结构呈现出多样化的适应策略.一些植物通过增加木质部导管直径来提高水分传输效率,以满足快速生长的需求;另一些植物则侧重于增强木质部的抗栓塞能力,以应对可能出现的干旱或冻融等胁迫事件[13].对长白山阔叶红松林的研究发现,不同功能类群的树种在这一阶段展现出明显的水力结构差异.具有木质部正压力(如根压或茎压)的树种能够有效修复冬季冻融循环诱导的气穴化栓塞,保持较高的水分传输效率;而不具备这种修复机制的树种则通过调整导管的形态和密度,寻求水分传输效率与安全性的平衡[14-15].顶级群落阶段植物的水力结构趋于稳定和优化,该阶段的植物通常具有较高的生物量和复杂的生态系统结构,水力结构的演化更加注重整体的协调性和对环境的精细适应.植物木质部导管在直径、长度、数量等方面达到一种动态平衡,既能保证高效的水分传输以支持庞大的生物量和生理活动,又能有效抵御各种环境胁迫[9,16]. ...
长白山阔叶树种木质部环孔和散孔结构特征的分化导致其水力学性状的显著差异
3
2018
... 植物的水力结构通常采用导水率(PLC)、边材比导率(Ks)、叶比导率(Kl)和胡伯尔值(Hv)来表征,这些参数分别从不同角度反映了植物的水分运输能力、输水效率以及叶片供水情况[8-9].利用木质部在不同水势下导水率变化得出木质部的脆弱性曲线[10].水力结构参数直接或间接地反映树木水分状况的指标,通过对这些指标的变化规律及其相互关系的研究认识树木的水分生理生态特点,从而阐明树木的耐旱特性及其机理[11].不同演替阶段,植物的水力结构参数呈现出明显的变化趋势[9].例如,贵州喀斯特地区的石漠化先锋植物中,常绿树种占据优势,其木质部导管直径相对较小,以牺牲部分水分传输效率为代价,换取更强的抗栓塞能力,从而在干旱、贫瘠的环境中生存[12].随着演替进入中期阶段,植物种类逐渐丰富,生境条件有所改善但依然存在一定的胁迫因素.此时,植物的水力结构呈现出多样化的适应策略.一些植物通过增加木质部导管直径来提高水分传输效率,以满足快速生长的需求;另一些植物则侧重于增强木质部的抗栓塞能力,以应对可能出现的干旱或冻融等胁迫事件[13].对长白山阔叶红松林的研究发现,不同功能类群的树种在这一阶段展现出明显的水力结构差异.具有木质部正压力(如根压或茎压)的树种能够有效修复冬季冻融循环诱导的气穴化栓塞,保持较高的水分传输效率;而不具备这种修复机制的树种则通过调整导管的形态和密度,寻求水分传输效率与安全性的平衡[14-15].顶级群落阶段植物的水力结构趋于稳定和优化,该阶段的植物通常具有较高的生物量和复杂的生态系统结构,水力结构的演化更加注重整体的协调性和对环境的精细适应.植物木质部导管在直径、长度、数量等方面达到一种动态平衡,既能保证高效的水分传输以支持庞大的生物量和生理活动,又能有效抵御各种环境胁迫[9,16]. ...
... 研究区位于腾格里沙漠以南中卫市沙坡头区包兰铁路以北的人工固沙植被区(37°32′N,105°02′E),该地区平均海拔1 339 m,最高气温可达38.1 ℃,最低气温-24.5 ℃,年平均气温为9.6 ℃,多年平均降水量为186.2 mm,降水量较为集中(5—9月),平均风速为2.9 m·s-1,最大风速可达19 m·s-1,年潜在蒸发量为2 900 mm,植被类型为荒漠化草原向草原化荒漠的过渡地带[14-15].包兰铁路穿过沙坡头地段的流动沙区,为了确保铁路正常运行,1956年起科研人员在流动沙区上设置了1 m×1 m的麦草方格,并且分别在1956、1964、1981、1987、1990、2010、2020年种植了以柠条、油蒿(Artemisia ordosica)、花棒(Hedysarum scoparium)为主的旱生灌木,目前已经在铁路两侧形成了长16 km,宽700 m的非灌溉人工固沙防护体系[14].在长期的演替过程中,铁路两侧的人工固沙植被体系内形成了丰富的生物土壤结皮,主要有藻类、藓类、地衣类等[16],草本植物以雾冰藜(Bassia dasyphlla)、小画眉草(Eragorostis minoe)、狗尾草(Setara viridis)等为主[26]. ...
... [14].在长期的演替过程中,铁路两侧的人工固沙植被体系内形成了丰富的生物土壤结皮,主要有藻类、藓类、地衣类等[16],草本植物以雾冰藜(Bassia dasyphlla)、小画眉草(Eragorostis minoe)、狗尾草(Setara viridis)等为主[26]. ...
A trade‐off between growth and hydraulic resilience against freezing leads to divergent adaptations among temperate tree species
2
2022
... 植物的水力结构通常采用导水率(PLC)、边材比导率(Ks)、叶比导率(Kl)和胡伯尔值(Hv)来表征,这些参数分别从不同角度反映了植物的水分运输能力、输水效率以及叶片供水情况[8-9].利用木质部在不同水势下导水率变化得出木质部的脆弱性曲线[10].水力结构参数直接或间接地反映树木水分状况的指标,通过对这些指标的变化规律及其相互关系的研究认识树木的水分生理生态特点,从而阐明树木的耐旱特性及其机理[11].不同演替阶段,植物的水力结构参数呈现出明显的变化趋势[9].例如,贵州喀斯特地区的石漠化先锋植物中,常绿树种占据优势,其木质部导管直径相对较小,以牺牲部分水分传输效率为代价,换取更强的抗栓塞能力,从而在干旱、贫瘠的环境中生存[12].随着演替进入中期阶段,植物种类逐渐丰富,生境条件有所改善但依然存在一定的胁迫因素.此时,植物的水力结构呈现出多样化的适应策略.一些植物通过增加木质部导管直径来提高水分传输效率,以满足快速生长的需求;另一些植物则侧重于增强木质部的抗栓塞能力,以应对可能出现的干旱或冻融等胁迫事件[13].对长白山阔叶红松林的研究发现,不同功能类群的树种在这一阶段展现出明显的水力结构差异.具有木质部正压力(如根压或茎压)的树种能够有效修复冬季冻融循环诱导的气穴化栓塞,保持较高的水分传输效率;而不具备这种修复机制的树种则通过调整导管的形态和密度,寻求水分传输效率与安全性的平衡[14-15].顶级群落阶段植物的水力结构趋于稳定和优化,该阶段的植物通常具有较高的生物量和复杂的生态系统结构,水力结构的演化更加注重整体的协调性和对环境的精细适应.植物木质部导管在直径、长度、数量等方面达到一种动态平衡,既能保证高效的水分传输以支持庞大的生物量和生理活动,又能有效抵御各种环境胁迫[9,16]. ...
... 研究区位于腾格里沙漠以南中卫市沙坡头区包兰铁路以北的人工固沙植被区(37°32′N,105°02′E),该地区平均海拔1 339 m,最高气温可达38.1 ℃,最低气温-24.5 ℃,年平均气温为9.6 ℃,多年平均降水量为186.2 mm,降水量较为集中(5—9月),平均风速为2.9 m·s-1,最大风速可达19 m·s-1,年潜在蒸发量为2 900 mm,植被类型为荒漠化草原向草原化荒漠的过渡地带[14-15].包兰铁路穿过沙坡头地段的流动沙区,为了确保铁路正常运行,1956年起科研人员在流动沙区上设置了1 m×1 m的麦草方格,并且分别在1956、1964、1981、1987、1990、2010、2020年种植了以柠条、油蒿(Artemisia ordosica)、花棒(Hedysarum scoparium)为主的旱生灌木,目前已经在铁路两侧形成了长16 km,宽700 m的非灌溉人工固沙防护体系[14].在长期的演替过程中,铁路两侧的人工固沙植被体系内形成了丰富的生物土壤结皮,主要有藻类、藓类、地衣类等[16],草本植物以雾冰藜(Bassia dasyphlla)、小画眉草(Eragorostis minoe)、狗尾草(Setara viridis)等为主[26]. ...
Hydraulic prediction of drought‐induced plant dieback and top‐kill depends on leaf habit and growth form
2
2021
... 植物的水力结构通常采用导水率(PLC)、边材比导率(Ks)、叶比导率(Kl)和胡伯尔值(Hv)来表征,这些参数分别从不同角度反映了植物的水分运输能力、输水效率以及叶片供水情况[8-9].利用木质部在不同水势下导水率变化得出木质部的脆弱性曲线[10].水力结构参数直接或间接地反映树木水分状况的指标,通过对这些指标的变化规律及其相互关系的研究认识树木的水分生理生态特点,从而阐明树木的耐旱特性及其机理[11].不同演替阶段,植物的水力结构参数呈现出明显的变化趋势[9].例如,贵州喀斯特地区的石漠化先锋植物中,常绿树种占据优势,其木质部导管直径相对较小,以牺牲部分水分传输效率为代价,换取更强的抗栓塞能力,从而在干旱、贫瘠的环境中生存[12].随着演替进入中期阶段,植物种类逐渐丰富,生境条件有所改善但依然存在一定的胁迫因素.此时,植物的水力结构呈现出多样化的适应策略.一些植物通过增加木质部导管直径来提高水分传输效率,以满足快速生长的需求;另一些植物则侧重于增强木质部的抗栓塞能力,以应对可能出现的干旱或冻融等胁迫事件[13].对长白山阔叶红松林的研究发现,不同功能类群的树种在这一阶段展现出明显的水力结构差异.具有木质部正压力(如根压或茎压)的树种能够有效修复冬季冻融循环诱导的气穴化栓塞,保持较高的水分传输效率;而不具备这种修复机制的树种则通过调整导管的形态和密度,寻求水分传输效率与安全性的平衡[14-15].顶级群落阶段植物的水力结构趋于稳定和优化,该阶段的植物通常具有较高的生物量和复杂的生态系统结构,水力结构的演化更加注重整体的协调性和对环境的精细适应.植物木质部导管在直径、长度、数量等方面达到一种动态平衡,既能保证高效的水分传输以支持庞大的生物量和生理活动,又能有效抵御各种环境胁迫[9,16]. ...
... 研究区位于腾格里沙漠以南中卫市沙坡头区包兰铁路以北的人工固沙植被区(37°32′N,105°02′E),该地区平均海拔1 339 m,最高气温可达38.1 ℃,最低气温-24.5 ℃,年平均气温为9.6 ℃,多年平均降水量为186.2 mm,降水量较为集中(5—9月),平均风速为2.9 m·s-1,最大风速可达19 m·s-1,年潜在蒸发量为2 900 mm,植被类型为荒漠化草原向草原化荒漠的过渡地带[14-15].包兰铁路穿过沙坡头地段的流动沙区,为了确保铁路正常运行,1956年起科研人员在流动沙区上设置了1 m×1 m的麦草方格,并且分别在1956、1964、1981、1987、1990、2010、2020年种植了以柠条、油蒿(Artemisia ordosica)、花棒(Hedysarum scoparium)为主的旱生灌木,目前已经在铁路两侧形成了长16 km,宽700 m的非灌溉人工固沙防护体系[14].在长期的演替过程中,铁路两侧的人工固沙植被体系内形成了丰富的生物土壤结皮,主要有藻类、藓类、地衣类等[16],草本植物以雾冰藜(Bassia dasyphlla)、小画眉草(Eragorostis minoe)、狗尾草(Setara viridis)等为主[26]. ...
Weak tradeoff between xylem hydraulic efficiency and safety:climatic seasonality matters
2
2021
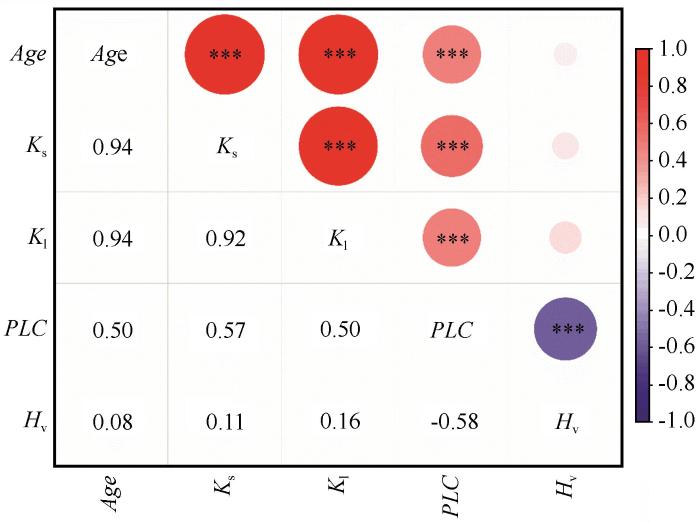
... 植物水力系统存在效率与安全性之间的内在权衡,即高水分输导效率往往以牺牲栓塞抗性为代价,反之亦然[17].这一权衡关系由木质部解剖结构所约束——大直径导管提升导水效率,但易引发栓塞;小直径导管增强安全性,但限制传输速率[18].该权衡广泛存在于物种间及种内不同发育阶段,是植物生态策略形成的基础[19].然而,其在极端干旱环境下的表达强度及调控机制仍不明确,尤其是在人工植被中,植被演替如何影响植物水力系统存在效率与安全性之间的权衡尚缺乏系统研究.植物水力功能随年龄增长发生显著变化.多数研究表明,林木水力导度往往随年龄增加而下降,其原因包括形成层活动衰退、导管结构变化及栓塞累积等[20].与之相比,栓塞抗性的年龄动态则较为复杂,常呈非线性变化,部分物种在中龄阶段表现出最优栓塞抗性[21].这动态反映了植物在不同生活史阶段对生长与生存的权衡,但相关研究多集中于天然林和乔木物种,对干旱区灌木林关注较少. ...
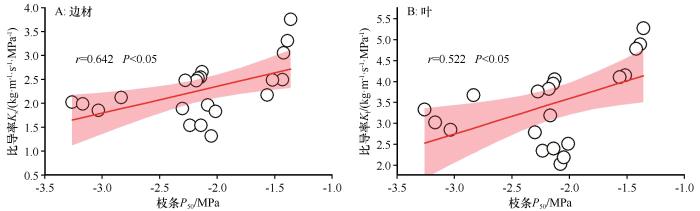
... 在植物水力系统中,水分传输效率与安全性之间普遍存在着一种权衡关系[17].本研究发现,柠条在不同的演替阶段会动态地调整其在效率与安全之间的策略侧重,表现出高适应性和可塑性.柠条的茎水力导度(Kₛ)与栓塞脆弱性(P₅₀)之间存在强烈的相关关系,其相关系数(r=0.642)高于全球平均水平(r=0.5)[25].在植被建设早期,柠条的主要目标是快速生长,以便在激烈的竞争中占据有利的生态位.因此,早期柠条具有较高的水力导度(Kₛ)和光合能力,但栓塞抗性(P₅₀)相对较弱,其策略明显偏向于效率优先.在应对干旱胁迫时,早期柠条主要采取快速关闭气孔的方式来减少水分散失,这是一种相对被动的、以牺牲光合作用为代价的短期应对策略[39].这种策略带来的快速生长收益弥补了栓塞损失,从而确保个体的成功定居.进入演替阶段中期后,柠条已经成功建立了稳定的生态位,其生态策略从效率转向安全.这一阶段,柠条表现出最强的栓塞抗性(P₅₀最低),同时通过提高胡伯尔值(Hᵥ)进行结构补偿,以应对水力导度的自然衰退.在应对干旱胁迫时,柠条不再仅仅依赖于快速关闭气孔,而是通过调整其水分利用策略来更好地利用水分[40].当进入演替阶段后期,柠条生理机能开始全面衰退,效率与安全之间的平衡被打破,呈现出双重下降的趋势.一方面,水力导度(Kₛ)持续降低,光合作用能力也显著下降,导致生长停滞.另一方面,栓塞抗性(P₅₀)的降低使其抵抗干旱的能力减弱.尽管后期柠条会进一步提高其水分利用效率,但这种效率的提高,是以牺牲生长和生物量积累为代价,无法阻止植株整体的衰退[41].最终,由于水力传输效率低下,无法有效供应叶片,同时水力安全性也得不到保障,使其在遭遇极端干旱事件时变得异常脆弱,容易发生大面积死亡,导致整个人工植被生态系统的崩溃[42].除去不同演替过程中柠条的年龄因素,区域环境条件的变化,不同水分、养分、生物结皮等土壤环境的改变也可能是重要的影响因素,其与年龄形成协同作用而影响水力性状与效率-安全性的权衡关系[43]有待更加深入地研究. ...
An ecophysiological and developmental perspective on variation in vessel diameter
1
2017
... 植物水力系统存在效率与安全性之间的内在权衡,即高水分输导效率往往以牺牲栓塞抗性为代价,反之亦然[17].这一权衡关系由木质部解剖结构所约束——大直径导管提升导水效率,但易引发栓塞;小直径导管增强安全性,但限制传输速率[18].该权衡广泛存在于物种间及种内不同发育阶段,是植物生态策略形成的基础[19].然而,其在极端干旱环境下的表达强度及调控机制仍不明确,尤其是在人工植被中,植被演替如何影响植物水力系统存在效率与安全性之间的权衡尚缺乏系统研究.植物水力功能随年龄增长发生显著变化.多数研究表明,林木水力导度往往随年龄增加而下降,其原因包括形成层活动衰退、导管结构变化及栓塞累积等[20].与之相比,栓塞抗性的年龄动态则较为复杂,常呈非线性变化,部分物种在中龄阶段表现出最优栓塞抗性[21].这动态反映了植物在不同生活史阶段对生长与生存的权衡,但相关研究多集中于天然林和乔木物种,对干旱区灌木林关注较少. ...
The world‐wide ‘fast-slow’plant economics spectrum:a traits manifesto
1
2014
... 植物水力系统存在效率与安全性之间的内在权衡,即高水分输导效率往往以牺牲栓塞抗性为代价,反之亦然[17].这一权衡关系由木质部解剖结构所约束——大直径导管提升导水效率,但易引发栓塞;小直径导管增强安全性,但限制传输速率[18].该权衡广泛存在于物种间及种内不同发育阶段,是植物生态策略形成的基础[19].然而,其在极端干旱环境下的表达强度及调控机制仍不明确,尤其是在人工植被中,植被演替如何影响植物水力系统存在效率与安全性之间的权衡尚缺乏系统研究.植物水力功能随年龄增长发生显著变化.多数研究表明,林木水力导度往往随年龄增加而下降,其原因包括形成层活动衰退、导管结构变化及栓塞累积等[20].与之相比,栓塞抗性的年龄动态则较为复杂,常呈非线性变化,部分物种在中龄阶段表现出最优栓塞抗性[21].这动态反映了植物在不同生活史阶段对生长与生存的权衡,但相关研究多集中于天然林和乔木物种,对干旱区灌木林关注较少. ...
Wood structure and function change with maturity:age of the vascular cambium is associated with xylem changes in current‐year growth
1
2019
... 植物水力系统存在效率与安全性之间的内在权衡,即高水分输导效率往往以牺牲栓塞抗性为代价,反之亦然[17].这一权衡关系由木质部解剖结构所约束——大直径导管提升导水效率,但易引发栓塞;小直径导管增强安全性,但限制传输速率[18].该权衡广泛存在于物种间及种内不同发育阶段,是植物生态策略形成的基础[19].然而,其在极端干旱环境下的表达强度及调控机制仍不明确,尤其是在人工植被中,植被演替如何影响植物水力系统存在效率与安全性之间的权衡尚缺乏系统研究.植物水力功能随年龄增长发生显著变化.多数研究表明,林木水力导度往往随年龄增加而下降,其原因包括形成层活动衰退、导管结构变化及栓塞累积等[20].与之相比,栓塞抗性的年龄动态则较为复杂,常呈非线性变化,部分物种在中龄阶段表现出最优栓塞抗性[21].这动态反映了植物在不同生活史阶段对生长与生存的权衡,但相关研究多集中于天然林和乔木物种,对干旱区灌木林关注较少. ...
Tree mortality across biomes is promoted by drought intensity,lower wood density and higher specific leaf area
1
2017
... 植物水力系统存在效率与安全性之间的内在权衡,即高水分输导效率往往以牺牲栓塞抗性为代价,反之亦然[17].这一权衡关系由木质部解剖结构所约束——大直径导管提升导水效率,但易引发栓塞;小直径导管增强安全性,但限制传输速率[18].该权衡广泛存在于物种间及种内不同发育阶段,是植物生态策略形成的基础[19].然而,其在极端干旱环境下的表达强度及调控机制仍不明确,尤其是在人工植被中,植被演替如何影响植物水力系统存在效率与安全性之间的权衡尚缺乏系统研究.植物水力功能随年龄增长发生显著变化.多数研究表明,林木水力导度往往随年龄增加而下降,其原因包括形成层活动衰退、导管结构变化及栓塞累积等[20].与之相比,栓塞抗性的年龄动态则较为复杂,常呈非线性变化,部分物种在中龄阶段表现出最优栓塞抗性[21].这动态反映了植物在不同生活史阶段对生长与生存的权衡,但相关研究多集中于天然林和乔木物种,对干旱区灌木林关注较少. ...
固沙灌木柠条锦鸡儿和中间锦鸡儿木质部导水与叶片光合能力对土壤水分的响应
2
2023
... 柠条(Caragana korshinskii)是中国北方干旱半干旱地区生态修复的常用树种,在防风固沙和生态恢复中起到关键作用[22].位于宁夏中卫沙坡头的人工植被防护体系从1956年开始人工柠条林的栽植,其盖度经过40年的演替最终稳定在10%左右[23],许多研究者从水量平衡、土壤水分植被承载力以及物种多样性等多种外部因素来解释此现象[24],而植物自身的生物学特征如何变化,是否存在安全效率权衡,至今未有定论.事实上,植物自身的水力结构特征有可能会随着植被土壤系统的演替进行调整和进化,从而保证植物的生存[25].基于此,本研究以腾格里沙漠不同建植年份(1956—2020年)的柠条人工植被为对象,通过系统测定分析其关键水力性状指标,旨在阐明人工植被演替过程中柠条水力功能随年龄的变化规律,揭示其水力效率与安全性之间的权衡关系.研究结果将深化对干旱区植物水分适应机制的理解,为人工植被的可持续管理及未来全球变化背景下的生态修复策略制定提供科学依据. ...
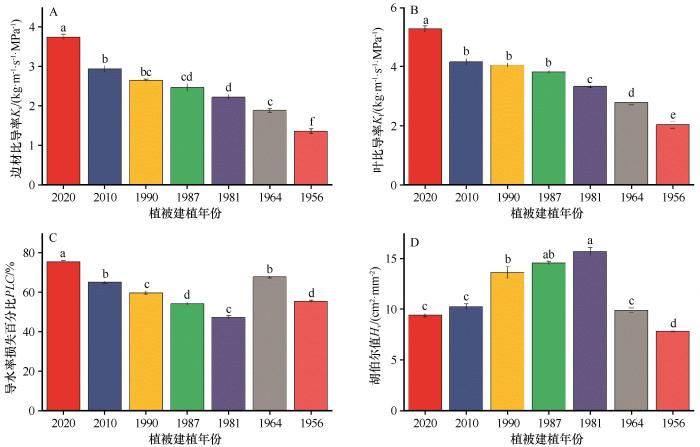
... 在腾格里沙漠人工植被生态系统中,柠条的水力结构特征随建植年份的变化,不仅反映了植物个体对环境的适应策略,也影响着整个植被系统的稳定性与可持续性[22].本研究通过对不同建植年份柠条关键水力参数的测定,揭示了其水力功能随植被演替的动态变化规律,并深入探讨了这些变化背后的生理生态学机制及其在干旱适应中的意义.研究发现,柠条的茎水力导度(Kₛ)和叶水力导度(Kₗ)均随植被演替呈现出显著的下降趋势.水力导度下降直接限制了植物从土壤中吸收水分并供应叶片的能力.这与全球范围内对多种木本植物的研究结果一致,被认为是植物衰老过程中的一个标志性特征[27-29].在人工植被建植早期,柠条为了快速生长和占据生态位,可能维持较高的水力导度,以支持旺盛的光合作用和生物量积累.但随着年龄增长,尤其是在土壤水分被持续消耗、环境胁迫加剧的情况下,维持高水力导度的成本过高,进而采取更为保守的水分利用策略,从而导致水力导度的主动或被动下降[30].随着植被的演替,柠条木质部结构也有可能出现衰退.如导管直径减小、纹孔密度降低、纹孔膜厚度增加等均会显著增加水分在导管间传输的阻力,从而进一步降低整株植物的水力导度[31].此外,演替阶段后期,柠条木质部中次生代谢产物(如酚类、树脂等)的积累也可能对水分传输构成物理性阻碍[32]. ...
我国沙区人工植被系统生态-水文过程和互馈机理研究评述
3
2013
... 柠条(Caragana korshinskii)是中国北方干旱半干旱地区生态修复的常用树种,在防风固沙和生态恢复中起到关键作用[22].位于宁夏中卫沙坡头的人工植被防护体系从1956年开始人工柠条林的栽植,其盖度经过40年的演替最终稳定在10%左右[23],许多研究者从水量平衡、土壤水分植被承载力以及物种多样性等多种外部因素来解释此现象[24],而植物自身的生物学特征如何变化,是否存在安全效率权衡,至今未有定论.事实上,植物自身的水力结构特征有可能会随着植被土壤系统的演替进行调整和进化,从而保证植物的生存[25].基于此,本研究以腾格里沙漠不同建植年份(1956—2020年)的柠条人工植被为对象,通过系统测定分析其关键水力性状指标,旨在阐明人工植被演替过程中柠条水力功能随年龄的变化规律,揭示其水力效率与安全性之间的权衡关系.研究结果将深化对干旱区植物水分适应机制的理解,为人工植被的可持续管理及未来全球变化背景下的生态修复策略制定提供科学依据. ...
... 胡伯尔值(Hᵥ)的单峰分布(图1D)揭示了柠条通过调整水力架构来优化水分利用的策略.这种结构调整具有多重效应:首先,能够补偿导管效率的下降,维持叶片水分供应;其次,提供水分储存缓冲,增强应对短期水分亏缺的能力;第三,增加水力安全边际,降低失控栓塞风险[37].在水分充足条件下,高Hᵥ可能造成过度投资,降低生长效率;而在干旱条件下其提供了关键的生存优势.本研究区年降水量仅186 mm,极端干旱事件频发,在这种环境下,结构补偿的收益可能大于成本.此外,补偿能力随演替阶段变化:前期植株资源有限,难以承担高补偿成本;后期植株生理功能衰退,补偿效果有限;只有中期植株在资源可用性和生理活力之间达到最佳平衡,能够实现有效补偿.演替后期Hᵥ的急剧下降表明补偿策略的不可持续性,这就使得植株既失去了高效的水分传输能力,又无法通过结构调整缓解水力限制,导致水力功能的全面崩溃.这也解释了许多人工灌木林在达到一定年龄后出现大面积衰退和死亡的现象[23]. ...
... 研究发现柠条的栓塞脆弱性(P₅₀)随建植年份呈现出非线性的“U”型变化模式.这一模式揭示了柠条在不同生活史阶段,为了适应腾格里沙漠极端干旱环境而进行的策略性调整 [1].在植被建植初期,柠条需要维持较高的水力导度和光合速率,以支持快速的生物量积累.随着柠条进入演替阶段中期,P₅₀值达到最低点,表现出最强的栓塞抗性.此时,植株已经建立了稳定的生态位,生长速率放缓,生存和维持功能稳定成为其主要策略.在腾格里沙漠极端干旱、降水年际波动巨大的环境中,高栓塞抗性意味着植株能够在遭遇极端干旱事件时,依然维持木质部导水系统的完整性,从而保证关键生理活动(如光合作用)的最低水分需求,避免在干旱中死亡[1,23].本研究中观测到-3.25 MPa的P₅₀值,在全球木本植物中属于中高抗性水平,证明了柠条具有强烈的适应腾格里沙漠极端干旱环境的能力.这种抗性的获得可能涉及多种复杂的生理和解剖学机制,例如纹孔膜结构优化、导管壁增厚以增强其抗气穴化能力,以及导管网络的冗余设计等,这些机制共同作用,构建了一个高效且安全的水分传输系统[30].当柠条进入演替阶段后期,其栓塞脆弱性有增加的趋势,P₅₀值增大.这一变化反映了植物在生理衰老过程中,维持高栓塞抗性的能力开始下降.维持高栓塞抗性需要持续的能量和资源投入,包括合成渗透调节物质、维护细胞壁结构、运行抗氧化系统等[30].后期植株的代谢活性普遍下降,可能无法支撑这些高成本的防御机制,其预示着植株抵抗极端干旱事件的能力减弱,是其功能衰退和最终死亡的一个重要信号[38]. ...
中国干旱沙区的生态重建与恢复:沙坡头站60年重要研究进展综述
1
2016
... 柠条(Caragana korshinskii)是中国北方干旱半干旱地区生态修复的常用树种,在防风固沙和生态恢复中起到关键作用[22].位于宁夏中卫沙坡头的人工植被防护体系从1956年开始人工柠条林的栽植,其盖度经过40年的演替最终稳定在10%左右[23],许多研究者从水量平衡、土壤水分植被承载力以及物种多样性等多种外部因素来解释此现象[24],而植物自身的生物学特征如何变化,是否存在安全效率权衡,至今未有定论.事实上,植物自身的水力结构特征有可能会随着植被土壤系统的演替进行调整和进化,从而保证植物的生存[25].基于此,本研究以腾格里沙漠不同建植年份(1956—2020年)的柠条人工植被为对象,通过系统测定分析其关键水力性状指标,旨在阐明人工植被演替过程中柠条水力功能随年龄的变化规律,揭示其水力效率与安全性之间的权衡关系.研究结果将深化对干旱区植物水分适应机制的理解,为人工植被的可持续管理及未来全球变化背景下的生态修复策略制定提供科学依据. ...
木本植物应对干旱胁迫的响应机制:基于水力学性状视角
2
2024
... 柠条(Caragana korshinskii)是中国北方干旱半干旱地区生态修复的常用树种,在防风固沙和生态恢复中起到关键作用[22].位于宁夏中卫沙坡头的人工植被防护体系从1956年开始人工柠条林的栽植,其盖度经过40年的演替最终稳定在10%左右[23],许多研究者从水量平衡、土壤水分植被承载力以及物种多样性等多种外部因素来解释此现象[24],而植物自身的生物学特征如何变化,是否存在安全效率权衡,至今未有定论.事实上,植物自身的水力结构特征有可能会随着植被土壤系统的演替进行调整和进化,从而保证植物的生存[25].基于此,本研究以腾格里沙漠不同建植年份(1956—2020年)的柠条人工植被为对象,通过系统测定分析其关键水力性状指标,旨在阐明人工植被演替过程中柠条水力功能随年龄的变化规律,揭示其水力效率与安全性之间的权衡关系.研究结果将深化对干旱区植物水分适应机制的理解,为人工植被的可持续管理及未来全球变化背景下的生态修复策略制定提供科学依据. ...
... 在植物水力系统中,水分传输效率与安全性之间普遍存在着一种权衡关系[17].本研究发现,柠条在不同的演替阶段会动态地调整其在效率与安全之间的策略侧重,表现出高适应性和可塑性.柠条的茎水力导度(Kₛ)与栓塞脆弱性(P₅₀)之间存在强烈的相关关系,其相关系数(r=0.642)高于全球平均水平(r=0.5)[25].在植被建设早期,柠条的主要目标是快速生长,以便在激烈的竞争中占据有利的生态位.因此,早期柠条具有较高的水力导度(Kₛ)和光合能力,但栓塞抗性(P₅₀)相对较弱,其策略明显偏向于效率优先.在应对干旱胁迫时,早期柠条主要采取快速关闭气孔的方式来减少水分散失,这是一种相对被动的、以牺牲光合作用为代价的短期应对策略[39].这种策略带来的快速生长收益弥补了栓塞损失,从而确保个体的成功定居.进入演替阶段中期后,柠条已经成功建立了稳定的生态位,其生态策略从效率转向安全.这一阶段,柠条表现出最强的栓塞抗性(P₅₀最低),同时通过提高胡伯尔值(Hᵥ)进行结构补偿,以应对水力导度的自然衰退.在应对干旱胁迫时,柠条不再仅仅依赖于快速关闭气孔,而是通过调整其水分利用策略来更好地利用水分[40].当进入演替阶段后期,柠条生理机能开始全面衰退,效率与安全之间的平衡被打破,呈现出双重下降的趋势.一方面,水力导度(Kₛ)持续降低,光合作用能力也显著下降,导致生长停滞.另一方面,栓塞抗性(P₅₀)的降低使其抵抗干旱的能力减弱.尽管后期柠条会进一步提高其水分利用效率,但这种效率的提高,是以牺牲生长和生物量积累为代价,无法阻止植株整体的衰退[41].最终,由于水力传输效率低下,无法有效供应叶片,同时水力安全性也得不到保障,使其在遭遇极端干旱事件时变得异常脆弱,容易发生大面积死亡,导致整个人工植被生态系统的崩溃[42].除去不同演替过程中柠条的年龄因素,区域环境条件的变化,不同水分、养分、生物结皮等土壤环境的改变也可能是重要的影响因素,其与年龄形成协同作用而影响水力性状与效率-安全性的权衡关系[43]有待更加深入地研究. ...
The stability of revegetated ecosystems in sandy areas:an assessment and prediction index
1
2015
... 研究区位于腾格里沙漠以南中卫市沙坡头区包兰铁路以北的人工固沙植被区(37°32′N,105°02′E),该地区平均海拔1 339 m,最高气温可达38.1 ℃,最低气温-24.5 ℃,年平均气温为9.6 ℃,多年平均降水量为186.2 mm,降水量较为集中(5—9月),平均风速为2.9 m·s-1,最大风速可达19 m·s-1,年潜在蒸发量为2 900 mm,植被类型为荒漠化草原向草原化荒漠的过渡地带[14-15].包兰铁路穿过沙坡头地段的流动沙区,为了确保铁路正常运行,1956年起科研人员在流动沙区上设置了1 m×1 m的麦草方格,并且分别在1956、1964、1981、1987、1990、2010、2020年种植了以柠条、油蒿(Artemisia ordosica)、花棒(Hedysarum scoparium)为主的旱生灌木,目前已经在铁路两侧形成了长16 km,宽700 m的非灌溉人工固沙防护体系[14].在长期的演替过程中,铁路两侧的人工固沙植被体系内形成了丰富的生物土壤结皮,主要有藻类、藓类、地衣类等[16],草本植物以雾冰藜(Bassia dasyphlla)、小画眉草(Eragorostis minoe)、狗尾草(Setara viridis)等为主[26]. ...
Limits to the height growth of Caragana korshinskii resprouts
1
2013
... 在腾格里沙漠人工植被生态系统中,柠条的水力结构特征随建植年份的变化,不仅反映了植物个体对环境的适应策略,也影响着整个植被系统的稳定性与可持续性[22].本研究通过对不同建植年份柠条关键水力参数的测定,揭示了其水力功能随植被演替的动态变化规律,并深入探讨了这些变化背后的生理生态学机制及其在干旱适应中的意义.研究发现,柠条的茎水力导度(Kₛ)和叶水力导度(Kₗ)均随植被演替呈现出显著的下降趋势.水力导度下降直接限制了植物从土壤中吸收水分并供应叶片的能力.这与全球范围内对多种木本植物的研究结果一致,被认为是植物衰老过程中的一个标志性特征[27-29].在人工植被建植早期,柠条为了快速生长和占据生态位,可能维持较高的水力导度,以支持旺盛的光合作用和生物量积累.但随着年龄增长,尤其是在土壤水分被持续消耗、环境胁迫加剧的情况下,维持高水力导度的成本过高,进而采取更为保守的水分利用策略,从而导致水力导度的主动或被动下降[30].随着植被的演替,柠条木质部结构也有可能出现衰退.如导管直径减小、纹孔密度降低、纹孔膜厚度增加等均会显著增加水分在导管间传输的阻力,从而进一步降低整株植物的水力导度[31].此外,演替阶段后期,柠条木质部中次生代谢产物(如酚类、树脂等)的积累也可能对水分传输构成物理性阻碍[32]. ...
Interactive effect between tree ageing and trunk-boring pest reduces hydraulics and carbon metabolism in Hippophae rhamnoides
1
2022
... 在腾格里沙漠人工植被生态系统中,柠条的水力结构特征随建植年份的变化,不仅反映了植物个体对环境的适应策略,也影响着整个植被系统的稳定性与可持续性[22].本研究通过对不同建植年份柠条关键水力参数的测定,揭示了其水力功能随植被演替的动态变化规律,并深入探讨了这些变化背后的生理生态学机制及其在干旱适应中的意义.研究发现,柠条的茎水力导度(Kₛ)和叶水力导度(Kₗ)均随植被演替呈现出显著的下降趋势.水力导度下降直接限制了植物从土壤中吸收水分并供应叶片的能力.这与全球范围内对多种木本植物的研究结果一致,被认为是植物衰老过程中的一个标志性特征[27-29].在人工植被建植早期,柠条为了快速生长和占据生态位,可能维持较高的水力导度,以支持旺盛的光合作用和生物量积累.但随着年龄增长,尤其是在土壤水分被持续消耗、环境胁迫加剧的情况下,维持高水力导度的成本过高,进而采取更为保守的水分利用策略,从而导致水力导度的主动或被动下降[30].随着植被的演替,柠条木质部结构也有可能出现衰退.如导管直径减小、纹孔密度降低、纹孔膜厚度增加等均会显著增加水分在导管间传输的阻力,从而进一步降低整株植物的水力导度[31].此外,演替阶段后期,柠条木质部中次生代谢产物(如酚类、树脂等)的积累也可能对水分传输构成物理性阻碍[32]. ...
维管植物木质部水分传输过程的影响因素及研究进展
3
2017
... 在腾格里沙漠人工植被生态系统中,柠条的水力结构特征随建植年份的变化,不仅反映了植物个体对环境的适应策略,也影响着整个植被系统的稳定性与可持续性[22].本研究通过对不同建植年份柠条关键水力参数的测定,揭示了其水力功能随植被演替的动态变化规律,并深入探讨了这些变化背后的生理生态学机制及其在干旱适应中的意义.研究发现,柠条的茎水力导度(Kₛ)和叶水力导度(Kₗ)均随植被演替呈现出显著的下降趋势.水力导度下降直接限制了植物从土壤中吸收水分并供应叶片的能力.这与全球范围内对多种木本植物的研究结果一致,被认为是植物衰老过程中的一个标志性特征[27-29].在人工植被建植早期,柠条为了快速生长和占据生态位,可能维持较高的水力导度,以支持旺盛的光合作用和生物量积累.但随着年龄增长,尤其是在土壤水分被持续消耗、环境胁迫加剧的情况下,维持高水力导度的成本过高,进而采取更为保守的水分利用策略,从而导致水力导度的主动或被动下降[30].随着植被的演替,柠条木质部结构也有可能出现衰退.如导管直径减小、纹孔密度降低、纹孔膜厚度增加等均会显著增加水分在导管间传输的阻力,从而进一步降低整株植物的水力导度[31].此外,演替阶段后期,柠条木质部中次生代谢产物(如酚类、树脂等)的积累也可能对水分传输构成物理性阻碍[32]. ...
... 研究发现柠条的栓塞脆弱性(P₅₀)随建植年份呈现出非线性的“U”型变化模式.这一模式揭示了柠条在不同生活史阶段,为了适应腾格里沙漠极端干旱环境而进行的策略性调整 [1].在植被建植初期,柠条需要维持较高的水力导度和光合速率,以支持快速的生物量积累.随着柠条进入演替阶段中期,P₅₀值达到最低点,表现出最强的栓塞抗性.此时,植株已经建立了稳定的生态位,生长速率放缓,生存和维持功能稳定成为其主要策略.在腾格里沙漠极端干旱、降水年际波动巨大的环境中,高栓塞抗性意味着植株能够在遭遇极端干旱事件时,依然维持木质部导水系统的完整性,从而保证关键生理活动(如光合作用)的最低水分需求,避免在干旱中死亡[1,23].本研究中观测到-3.25 MPa的P₅₀值,在全球木本植物中属于中高抗性水平,证明了柠条具有强烈的适应腾格里沙漠极端干旱环境的能力.这种抗性的获得可能涉及多种复杂的生理和解剖学机制,例如纹孔膜结构优化、导管壁增厚以增强其抗气穴化能力,以及导管网络的冗余设计等,这些机制共同作用,构建了一个高效且安全的水分传输系统[30].当柠条进入演替阶段后期,其栓塞脆弱性有增加的趋势,P₅₀值增大.这一变化反映了植物在生理衰老过程中,维持高栓塞抗性的能力开始下降.维持高栓塞抗性需要持续的能量和资源投入,包括合成渗透调节物质、维护细胞壁结构、运行抗氧化系统等[30].后期植株的代谢活性普遍下降,可能无法支撑这些高成本的防御机制,其预示着植株抵抗极端干旱事件的能力减弱,是其功能衰退和最终死亡的一个重要信号[38]. ...
... [30].后期植株的代谢活性普遍下降,可能无法支撑这些高成本的防御机制,其预示着植株抵抗极端干旱事件的能力减弱,是其功能衰退和最终死亡的一个重要信号[38]. ...
Effects of limited water availability on xylem transport in liana Humulus lupulus L
1
2016
... 在腾格里沙漠人工植被生态系统中,柠条的水力结构特征随建植年份的变化,不仅反映了植物个体对环境的适应策略,也影响着整个植被系统的稳定性与可持续性[22].本研究通过对不同建植年份柠条关键水力参数的测定,揭示了其水力功能随植被演替的动态变化规律,并深入探讨了这些变化背后的生理生态学机制及其在干旱适应中的意义.研究发现,柠条的茎水力导度(Kₛ)和叶水力导度(Kₗ)均随植被演替呈现出显著的下降趋势.水力导度下降直接限制了植物从土壤中吸收水分并供应叶片的能力.这与全球范围内对多种木本植物的研究结果一致,被认为是植物衰老过程中的一个标志性特征[27-29].在人工植被建植早期,柠条为了快速生长和占据生态位,可能维持较高的水力导度,以支持旺盛的光合作用和生物量积累.但随着年龄增长,尤其是在土壤水分被持续消耗、环境胁迫加剧的情况下,维持高水力导度的成本过高,进而采取更为保守的水分利用策略,从而导致水力导度的主动或被动下降[30].随着植被的演替,柠条木质部结构也有可能出现衰退.如导管直径减小、纹孔密度降低、纹孔膜厚度增加等均会显著增加水分在导管间传输的阻力,从而进一步降低整株植物的水力导度[31].此外,演替阶段后期,柠条木质部中次生代谢产物(如酚类、树脂等)的积累也可能对水分传输构成物理性阻碍[32]. ...
Plant phenolics: neglected secondary metabolites in plant stress tolerance
1
2024
... 在腾格里沙漠人工植被生态系统中,柠条的水力结构特征随建植年份的变化,不仅反映了植物个体对环境的适应策略,也影响着整个植被系统的稳定性与可持续性[22].本研究通过对不同建植年份柠条关键水力参数的测定,揭示了其水力功能随植被演替的动态变化规律,并深入探讨了这些变化背后的生理生态学机制及其在干旱适应中的意义.研究发现,柠条的茎水力导度(Kₛ)和叶水力导度(Kₗ)均随植被演替呈现出显著的下降趋势.水力导度下降直接限制了植物从土壤中吸收水分并供应叶片的能力.这与全球范围内对多种木本植物的研究结果一致,被认为是植物衰老过程中的一个标志性特征[27-29].在人工植被建植早期,柠条为了快速生长和占据生态位,可能维持较高的水力导度,以支持旺盛的光合作用和生物量积累.但随着年龄增长,尤其是在土壤水分被持续消耗、环境胁迫加剧的情况下,维持高水力导度的成本过高,进而采取更为保守的水分利用策略,从而导致水力导度的主动或被动下降[30].随着植被的演替,柠条木质部结构也有可能出现衰退.如导管直径减小、纹孔密度降低、纹孔膜厚度增加等均会显著增加水分在导管间传输的阻力,从而进一步降低整株植物的水力导度[31].此外,演替阶段后期,柠条木质部中次生代谢产物(如酚类、树脂等)的积累也可能对水分传输构成物理性阻碍[32]. ...
木本植物木质部栓塞修复机制: 研究进展与问题
2
2016
... 除了木质部结构的衰老,栓塞累积与修复能力下降也是导致水力功能衰退的重要因素[33].植物经历多次环境胁迫事件(干旱、冻融、机械损伤等)会导致木质部栓塞的逐渐累积.虽然植物拥有一定的栓塞修复机制(如根压驱动再充水、新生木质部补偿等),但这种修复能力随年龄增长而下降[34-35].本研究中,演替后期柠条栓塞水平(PLC)显著高于早期植株,支持累积损伤假说.后期植株能量代谢水平降低、碳水化合物储备减少以及细胞膜修复能力下降,都可能影响其栓塞修复效率.长期累积栓塞不仅直接降低了水力导度,还可能触发一系列负反馈过程,例如导致栓塞导管周围薄壁细胞死亡、为病原体入侵提供通道,甚至引发局部木质部功能的永久性丧失,最终导致整株植物水力功能的全面崩溃[33,36]. ...
... [33,36]. ...
6个树种1年生枝木质部的水力特征及与栓塞修复能力的关系
1
2017
... 除了木质部结构的衰老,栓塞累积与修复能力下降也是导致水力功能衰退的重要因素[33].植物经历多次环境胁迫事件(干旱、冻融、机械损伤等)会导致木质部栓塞的逐渐累积.虽然植物拥有一定的栓塞修复机制(如根压驱动再充水、新生木质部补偿等),但这种修复能力随年龄增长而下降[34-35].本研究中,演替后期柠条栓塞水平(PLC)显著高于早期植株,支持累积损伤假说.后期植株能量代谢水平降低、碳水化合物储备减少以及细胞膜修复能力下降,都可能影响其栓塞修复效率.长期累积栓塞不仅直接降低了水力导度,还可能触发一系列负反馈过程,例如导致栓塞导管周围薄壁细胞死亡、为病原体入侵提供通道,甚至引发局部木质部功能的永久性丧失,最终导致整株植物水力功能的全面崩溃[33,36]. ...
Divergent stem hydraulic strategies of Caragana korshinskii resprouts following a disturbance
1
2022
... 除了木质部结构的衰老,栓塞累积与修复能力下降也是导致水力功能衰退的重要因素[33].植物经历多次环境胁迫事件(干旱、冻融、机械损伤等)会导致木质部栓塞的逐渐累积.虽然植物拥有一定的栓塞修复机制(如根压驱动再充水、新生木质部补偿等),但这种修复能力随年龄增长而下降[34-35].本研究中,演替后期柠条栓塞水平(PLC)显著高于早期植株,支持累积损伤假说.后期植株能量代谢水平降低、碳水化合物储备减少以及细胞膜修复能力下降,都可能影响其栓塞修复效率.长期累积栓塞不仅直接降低了水力导度,还可能触发一系列负反馈过程,例如导致栓塞导管周围薄壁细胞死亡、为病原体入侵提供通道,甚至引发局部木质部功能的永久性丧失,最终导致整株植物水力功能的全面崩溃[33,36]. ...
干旱与冻害胁迫对植物水分运输的影响机制:木质部栓塞和栓塞疲劳
1
2024
... 除了木质部结构的衰老,栓塞累积与修复能力下降也是导致水力功能衰退的重要因素[33].植物经历多次环境胁迫事件(干旱、冻融、机械损伤等)会导致木质部栓塞的逐渐累积.虽然植物拥有一定的栓塞修复机制(如根压驱动再充水、新生木质部补偿等),但这种修复能力随年龄增长而下降[34-35].本研究中,演替后期柠条栓塞水平(PLC)显著高于早期植株,支持累积损伤假说.后期植株能量代谢水平降低、碳水化合物储备减少以及细胞膜修复能力下降,都可能影响其栓塞修复效率.长期累积栓塞不仅直接降低了水力导度,还可能触发一系列负反馈过程,例如导致栓塞导管周围薄壁细胞死亡、为病原体入侵提供通道,甚至引发局部木质部功能的永久性丧失,最终导致整株植物水力功能的全面崩溃[33,36]. ...
内蒙古高原西部荒漠区四种锦鸡儿属植物水力结构变化的比较研究
1
2015
... 胡伯尔值(Hᵥ)的单峰分布(图1D)揭示了柠条通过调整水力架构来优化水分利用的策略.这种结构调整具有多重效应:首先,能够补偿导管效率的下降,维持叶片水分供应;其次,提供水分储存缓冲,增强应对短期水分亏缺的能力;第三,增加水力安全边际,降低失控栓塞风险[37].在水分充足条件下,高Hᵥ可能造成过度投资,降低生长效率;而在干旱条件下其提供了关键的生存优势.本研究区年降水量仅186 mm,极端干旱事件频发,在这种环境下,结构补偿的收益可能大于成本.此外,补偿能力随演替阶段变化:前期植株资源有限,难以承担高补偿成本;后期植株生理功能衰退,补偿效果有限;只有中期植株在资源可用性和生理活力之间达到最佳平衡,能够实现有效补偿.演替后期Hᵥ的急剧下降表明补偿策略的不可持续性,这就使得植株既失去了高效的水分传输能力,又无法通过结构调整缓解水力限制,导致水力功能的全面崩溃.这也解释了许多人工灌木林在达到一定年龄后出现大面积衰退和死亡的现象[23]. ...
荒漠植物逆境生理生态学研究进展
1
2025
... 研究发现柠条的栓塞脆弱性(P₅₀)随建植年份呈现出非线性的“U”型变化模式.这一模式揭示了柠条在不同生活史阶段,为了适应腾格里沙漠极端干旱环境而进行的策略性调整 [1].在植被建植初期,柠条需要维持较高的水力导度和光合速率,以支持快速的生物量积累.随着柠条进入演替阶段中期,P₅₀值达到最低点,表现出最强的栓塞抗性.此时,植株已经建立了稳定的生态位,生长速率放缓,生存和维持功能稳定成为其主要策略.在腾格里沙漠极端干旱、降水年际波动巨大的环境中,高栓塞抗性意味着植株能够在遭遇极端干旱事件时,依然维持木质部导水系统的完整性,从而保证关键生理活动(如光合作用)的最低水分需求,避免在干旱中死亡[1,23].本研究中观测到-3.25 MPa的P₅₀值,在全球木本植物中属于中高抗性水平,证明了柠条具有强烈的适应腾格里沙漠极端干旱环境的能力.这种抗性的获得可能涉及多种复杂的生理和解剖学机制,例如纹孔膜结构优化、导管壁增厚以增强其抗气穴化能力,以及导管网络的冗余设计等,这些机制共同作用,构建了一个高效且安全的水分传输系统[30].当柠条进入演替阶段后期,其栓塞脆弱性有增加的趋势,P₅₀值增大.这一变化反映了植物在生理衰老过程中,维持高栓塞抗性的能力开始下降.维持高栓塞抗性需要持续的能量和资源投入,包括合成渗透调节物质、维护细胞壁结构、运行抗氧化系统等[30].后期植株的代谢活性普遍下降,可能无法支撑这些高成本的防御机制,其预示着植株抵抗极端干旱事件的能力减弱,是其功能衰退和最终死亡的一个重要信号[38]. ...
Weak tradeoff between xylem hydraulic efficiency and safety:climatic seasonality matters
1
2021
... 在植物水力系统中,水分传输效率与安全性之间普遍存在着一种权衡关系[17].本研究发现,柠条在不同的演替阶段会动态地调整其在效率与安全之间的策略侧重,表现出高适应性和可塑性.柠条的茎水力导度(Kₛ)与栓塞脆弱性(P₅₀)之间存在强烈的相关关系,其相关系数(r=0.642)高于全球平均水平(r=0.5)[25].在植被建设早期,柠条的主要目标是快速生长,以便在激烈的竞争中占据有利的生态位.因此,早期柠条具有较高的水力导度(Kₛ)和光合能力,但栓塞抗性(P₅₀)相对较弱,其策略明显偏向于效率优先.在应对干旱胁迫时,早期柠条主要采取快速关闭气孔的方式来减少水分散失,这是一种相对被动的、以牺牲光合作用为代价的短期应对策略[39].这种策略带来的快速生长收益弥补了栓塞损失,从而确保个体的成功定居.进入演替阶段中期后,柠条已经成功建立了稳定的生态位,其生态策略从效率转向安全.这一阶段,柠条表现出最强的栓塞抗性(P₅₀最低),同时通过提高胡伯尔值(Hᵥ)进行结构补偿,以应对水力导度的自然衰退.在应对干旱胁迫时,柠条不再仅仅依赖于快速关闭气孔,而是通过调整其水分利用策略来更好地利用水分[40].当进入演替阶段后期,柠条生理机能开始全面衰退,效率与安全之间的平衡被打破,呈现出双重下降的趋势.一方面,水力导度(Kₛ)持续降低,光合作用能力也显著下降,导致生长停滞.另一方面,栓塞抗性(P₅₀)的降低使其抵抗干旱的能力减弱.尽管后期柠条会进一步提高其水分利用效率,但这种效率的提高,是以牺牲生长和生物量积累为代价,无法阻止植株整体的衰退[41].最终,由于水力传输效率低下,无法有效供应叶片,同时水力安全性也得不到保障,使其在遭遇极端干旱事件时变得异常脆弱,容易发生大面积死亡,导致整个人工植被生态系统的崩溃[42].除去不同演替过程中柠条的年龄因素,区域环境条件的变化,不同水分、养分、生物结皮等土壤环境的改变也可能是重要的影响因素,其与年龄形成协同作用而影响水力性状与效率-安全性的权衡关系[43]有待更加深入地研究. ...
植物水分调节对策:等水与非等水行为
1
2017
... 在植物水力系统中,水分传输效率与安全性之间普遍存在着一种权衡关系[17].本研究发现,柠条在不同的演替阶段会动态地调整其在效率与安全之间的策略侧重,表现出高适应性和可塑性.柠条的茎水力导度(Kₛ)与栓塞脆弱性(P₅₀)之间存在强烈的相关关系,其相关系数(r=0.642)高于全球平均水平(r=0.5)[25].在植被建设早期,柠条的主要目标是快速生长,以便在激烈的竞争中占据有利的生态位.因此,早期柠条具有较高的水力导度(Kₛ)和光合能力,但栓塞抗性(P₅₀)相对较弱,其策略明显偏向于效率优先.在应对干旱胁迫时,早期柠条主要采取快速关闭气孔的方式来减少水分散失,这是一种相对被动的、以牺牲光合作用为代价的短期应对策略[39].这种策略带来的快速生长收益弥补了栓塞损失,从而确保个体的成功定居.进入演替阶段中期后,柠条已经成功建立了稳定的生态位,其生态策略从效率转向安全.这一阶段,柠条表现出最强的栓塞抗性(P₅₀最低),同时通过提高胡伯尔值(Hᵥ)进行结构补偿,以应对水力导度的自然衰退.在应对干旱胁迫时,柠条不再仅仅依赖于快速关闭气孔,而是通过调整其水分利用策略来更好地利用水分[40].当进入演替阶段后期,柠条生理机能开始全面衰退,效率与安全之间的平衡被打破,呈现出双重下降的趋势.一方面,水力导度(Kₛ)持续降低,光合作用能力也显著下降,导致生长停滞.另一方面,栓塞抗性(P₅₀)的降低使其抵抗干旱的能力减弱.尽管后期柠条会进一步提高其水分利用效率,但这种效率的提高,是以牺牲生长和生物量积累为代价,无法阻止植株整体的衰退[41].最终,由于水力传输效率低下,无法有效供应叶片,同时水力安全性也得不到保障,使其在遭遇极端干旱事件时变得异常脆弱,容易发生大面积死亡,导致整个人工植被生态系统的崩溃[42].除去不同演替过程中柠条的年龄因素,区域环境条件的变化,不同水分、养分、生物结皮等土壤环境的改变也可能是重要的影响因素,其与年龄形成协同作用而影响水力性状与效率-安全性的权衡关系[43]有待更加深入地研究. ...
Stable isotopic analysis on water utilization of two xerophytic shrubs in a revegetated desert area:Tengger Desert,China
1
2015
... 在植物水力系统中,水分传输效率与安全性之间普遍存在着一种权衡关系[17].本研究发现,柠条在不同的演替阶段会动态地调整其在效率与安全之间的策略侧重,表现出高适应性和可塑性.柠条的茎水力导度(Kₛ)与栓塞脆弱性(P₅₀)之间存在强烈的相关关系,其相关系数(r=0.642)高于全球平均水平(r=0.5)[25].在植被建设早期,柠条的主要目标是快速生长,以便在激烈的竞争中占据有利的生态位.因此,早期柠条具有较高的水力导度(Kₛ)和光合能力,但栓塞抗性(P₅₀)相对较弱,其策略明显偏向于效率优先.在应对干旱胁迫时,早期柠条主要采取快速关闭气孔的方式来减少水分散失,这是一种相对被动的、以牺牲光合作用为代价的短期应对策略[39].这种策略带来的快速生长收益弥补了栓塞损失,从而确保个体的成功定居.进入演替阶段中期后,柠条已经成功建立了稳定的生态位,其生态策略从效率转向安全.这一阶段,柠条表现出最强的栓塞抗性(P₅₀最低),同时通过提高胡伯尔值(Hᵥ)进行结构补偿,以应对水力导度的自然衰退.在应对干旱胁迫时,柠条不再仅仅依赖于快速关闭气孔,而是通过调整其水分利用策略来更好地利用水分[40].当进入演替阶段后期,柠条生理机能开始全面衰退,效率与安全之间的平衡被打破,呈现出双重下降的趋势.一方面,水力导度(Kₛ)持续降低,光合作用能力也显著下降,导致生长停滞.另一方面,栓塞抗性(P₅₀)的降低使其抵抗干旱的能力减弱.尽管后期柠条会进一步提高其水分利用效率,但这种效率的提高,是以牺牲生长和生物量积累为代价,无法阻止植株整体的衰退[41].最终,由于水力传输效率低下,无法有效供应叶片,同时水力安全性也得不到保障,使其在遭遇极端干旱事件时变得异常脆弱,容易发生大面积死亡,导致整个人工植被生态系统的崩溃[42].除去不同演替过程中柠条的年龄因素,区域环境条件的变化,不同水分、养分、生物结皮等土壤环境的改变也可能是重要的影响因素,其与年龄形成协同作用而影响水力性状与效率-安全性的权衡关系[43]有待更加深入地研究. ...
Hydraulic safety and growth rather than climate of origin influence survival in desert shrubs and trees
1
2023
... 在植物水力系统中,水分传输效率与安全性之间普遍存在着一种权衡关系[17].本研究发现,柠条在不同的演替阶段会动态地调整其在效率与安全之间的策略侧重,表现出高适应性和可塑性.柠条的茎水力导度(Kₛ)与栓塞脆弱性(P₅₀)之间存在强烈的相关关系,其相关系数(r=0.642)高于全球平均水平(r=0.5)[25].在植被建设早期,柠条的主要目标是快速生长,以便在激烈的竞争中占据有利的生态位.因此,早期柠条具有较高的水力导度(Kₛ)和光合能力,但栓塞抗性(P₅₀)相对较弱,其策略明显偏向于效率优先.在应对干旱胁迫时,早期柠条主要采取快速关闭气孔的方式来减少水分散失,这是一种相对被动的、以牺牲光合作用为代价的短期应对策略[39].这种策略带来的快速生长收益弥补了栓塞损失,从而确保个体的成功定居.进入演替阶段中期后,柠条已经成功建立了稳定的生态位,其生态策略从效率转向安全.这一阶段,柠条表现出最强的栓塞抗性(P₅₀最低),同时通过提高胡伯尔值(Hᵥ)进行结构补偿,以应对水力导度的自然衰退.在应对干旱胁迫时,柠条不再仅仅依赖于快速关闭气孔,而是通过调整其水分利用策略来更好地利用水分[40].当进入演替阶段后期,柠条生理机能开始全面衰退,效率与安全之间的平衡被打破,呈现出双重下降的趋势.一方面,水力导度(Kₛ)持续降低,光合作用能力也显著下降,导致生长停滞.另一方面,栓塞抗性(P₅₀)的降低使其抵抗干旱的能力减弱.尽管后期柠条会进一步提高其水分利用效率,但这种效率的提高,是以牺牲生长和生物量积累为代价,无法阻止植株整体的衰退[41].最终,由于水力传输效率低下,无法有效供应叶片,同时水力安全性也得不到保障,使其在遭遇极端干旱事件时变得异常脆弱,容易发生大面积死亡,导致整个人工植被生态系统的崩溃[42].除去不同演替过程中柠条的年龄因素,区域环境条件的变化,不同水分、养分、生物结皮等土壤环境的改变也可能是重要的影响因素,其与年龄形成协同作用而影响水力性状与效率-安全性的权衡关系[43]有待更加深入地研究. ...
Size-related decline in dryland shrubs is related to reductions in hydraulic efficiency and carbon assimilation and not nonstructural carbohydrate depletion
1
2025
... 在植物水力系统中,水分传输效率与安全性之间普遍存在着一种权衡关系[17].本研究发现,柠条在不同的演替阶段会动态地调整其在效率与安全之间的策略侧重,表现出高适应性和可塑性.柠条的茎水力导度(Kₛ)与栓塞脆弱性(P₅₀)之间存在强烈的相关关系,其相关系数(r=0.642)高于全球平均水平(r=0.5)[25].在植被建设早期,柠条的主要目标是快速生长,以便在激烈的竞争中占据有利的生态位.因此,早期柠条具有较高的水力导度(Kₛ)和光合能力,但栓塞抗性(P₅₀)相对较弱,其策略明显偏向于效率优先.在应对干旱胁迫时,早期柠条主要采取快速关闭气孔的方式来减少水分散失,这是一种相对被动的、以牺牲光合作用为代价的短期应对策略[39].这种策略带来的快速生长收益弥补了栓塞损失,从而确保个体的成功定居.进入演替阶段中期后,柠条已经成功建立了稳定的生态位,其生态策略从效率转向安全.这一阶段,柠条表现出最强的栓塞抗性(P₅₀最低),同时通过提高胡伯尔值(Hᵥ)进行结构补偿,以应对水力导度的自然衰退.在应对干旱胁迫时,柠条不再仅仅依赖于快速关闭气孔,而是通过调整其水分利用策略来更好地利用水分[40].当进入演替阶段后期,柠条生理机能开始全面衰退,效率与安全之间的平衡被打破,呈现出双重下降的趋势.一方面,水力导度(Kₛ)持续降低,光合作用能力也显著下降,导致生长停滞.另一方面,栓塞抗性(P₅₀)的降低使其抵抗干旱的能力减弱.尽管后期柠条会进一步提高其水分利用效率,但这种效率的提高,是以牺牲生长和生物量积累为代价,无法阻止植株整体的衰退[41].最终,由于水力传输效率低下,无法有效供应叶片,同时水力安全性也得不到保障,使其在遭遇极端干旱事件时变得异常脆弱,容易发生大面积死亡,导致整个人工植被生态系统的崩溃[42].除去不同演替过程中柠条的年龄因素,区域环境条件的变化,不同水分、养分、生物结皮等土壤环境的改变也可能是重要的影响因素,其与年龄形成协同作用而影响水力性状与效率-安全性的权衡关系[43]有待更加深入地研究. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}