1
2007
... 根据联合国政府间气候变化专门委员会(IPCC)第六次评估报告,近一个多世纪来,全球平均气温上升了0.85 ℃,预计到2100年升温幅度为1.8~4.0 ℃[1-2].同样地,中国的年平均气温升高了0.9~1.5 ℃,预计到2050年,中国的平均气温将进一步上升,尤其在干旱地区,气温升幅可能更加显著[3-5].随着全球气候变暖,极端气候事件和降水格局也随之改变[4],进而对陆地生态系统植物群落结构和功能产生重要影响[6]. ...
Climate Change 2021:the Physical Science Basis
1
2021
... 根据联合国政府间气候变化专门委员会(IPCC)第六次评估报告,近一个多世纪来,全球平均气温上升了0.85 ℃,预计到2100年升温幅度为1.8~4.0 ℃[1-2].同样地,中国的年平均气温升高了0.9~1.5 ℃,预计到2050年,中国的平均气温将进一步上升,尤其在干旱地区,气温升幅可能更加显著[3-5].随着全球气候变暖,极端气候事件和降水格局也随之改变[4],进而对陆地生态系统植物群落结构和功能产生重要影响[6]. ...
IPCC-AR4模式对中国21世纪气候变化的情景预估
1
2008
... 根据联合国政府间气候变化专门委员会(IPCC)第六次评估报告,近一个多世纪来,全球平均气温上升了0.85 ℃,预计到2100年升温幅度为1.8~4.0 ℃[1-2].同样地,中国的年平均气温升高了0.9~1.5 ℃,预计到2050年,中国的平均气温将进一步上升,尤其在干旱地区,气温升幅可能更加显著[3-5].随着全球气候变暖,极端气候事件和降水格局也随之改变[4],进而对陆地生态系统植物群落结构和功能产生重要影响[6]. ...
1
2015
... 根据联合国政府间气候变化专门委员会(IPCC)第六次评估报告,近一个多世纪来,全球平均气温上升了0.85 ℃,预计到2100年升温幅度为1.8~4.0 ℃[1-2].同样地,中国的年平均气温升高了0.9~1.5 ℃,预计到2050年,中国的平均气温将进一步上升,尤其在干旱地区,气温升幅可能更加显著[3-5].随着全球气候变暖,极端气候事件和降水格局也随之改变[4],进而对陆地生态系统植物群落结构和功能产生重要影响[6]. ...
Temperatures in excess of critical thresholds threaten nestling growth and survival in a rapidly-warming arid savanna:a study of common fiscals
1
2013
... 根据联合国政府间气候变化专门委员会(IPCC)第六次评估报告,近一个多世纪来,全球平均气温上升了0.85 ℃,预计到2100年升温幅度为1.8~4.0 ℃[1-2].同样地,中国的年平均气温升高了0.9~1.5 ℃,预计到2050年,中国的平均气温将进一步上升,尤其在干旱地区,气温升幅可能更加显著[3-5].随着全球气候变暖,极端气候事件和降水格局也随之改变[4],进而对陆地生态系统植物群落结构和功能产生重要影响[6]. ...
Urbanization contributes little to global warming but substantially intensifies local and regional land surface warming
1
2022
... 根据联合国政府间气候变化专门委员会(IPCC)第六次评估报告,近一个多世纪来,全球平均气温上升了0.85 ℃,预计到2100年升温幅度为1.8~4.0 ℃[1-2].同样地,中国的年平均气温升高了0.9~1.5 ℃,预计到2050年,中国的平均气温将进一步上升,尤其在干旱地区,气温升幅可能更加显著[3-5].随着全球气候变暖,极端气候事件和降水格局也随之改变[4],进而对陆地生态系统植物群落结构和功能产生重要影响[6]. ...
Plant phenology and global climate change:current progresses and challenges
1
2019
... 全球变暖对植物个体生长发育、物种组成和群落演变均有很大影响.在个体尺度上,增温对植物生长和繁殖有重要影响,例如气候变暖会使植物叶片生长和花期提前,导致植物加速成熟和衰老[7].另外,气候变暖还会增加植物的光合作用速率和蒸腾作用,导致水分流失加快.在干旱地区,这可能使植物水分胁迫加剧,限制植物生长[8].增温还对植物叶片形态、化学计量元素含量和种子质量等产生影响[9-10].而在群落尺度上,气候变暖会改变物种的分布,导致物种向极地或者高海拔地区迁移,引起植物群落结构和多样性的改变[11].同时,气候变暖会改变植物种间竞争强度来改变群落物种组成,增温可增强耐热物种竞争力,从而导致群落中不耐热物种的减少[12].增温还会导致群落中物种多样性或丰富度降低,如在北极苔原地区,短期的增温使灌木和禾草的盖度增加,但减少了物种多样性[13];增温使青藏高原区植物群落物种丰富度快速降低了26%~36%[14].在生态系统尺度上,气候变暖会改变生态系统物质和能量循环,进而影响生态系统稳定性和服务功能[15]. ...
Plant responses to rising vapor pressure deficit
1
2020
... 全球变暖对植物个体生长发育、物种组成和群落演变均有很大影响.在个体尺度上,增温对植物生长和繁殖有重要影响,例如气候变暖会使植物叶片生长和花期提前,导致植物加速成熟和衰老[7].另外,气候变暖还会增加植物的光合作用速率和蒸腾作用,导致水分流失加快.在干旱地区,这可能使植物水分胁迫加剧,限制植物生长[8].增温还对植物叶片形态、化学计量元素含量和种子质量等产生影响[9-10].而在群落尺度上,气候变暖会改变物种的分布,导致物种向极地或者高海拔地区迁移,引起植物群落结构和多样性的改变[11].同时,气候变暖会改变植物种间竞争强度来改变群落物种组成,增温可增强耐热物种竞争力,从而导致群落中不耐热物种的减少[12].增温还会导致群落中物种多样性或丰富度降低,如在北极苔原地区,短期的增温使灌木和禾草的盖度增加,但减少了物种多样性[13];增温使青藏高原区植物群落物种丰富度快速降低了26%~36%[14].在生态系统尺度上,气候变暖会改变生态系统物质和能量循环,进而影响生态系统稳定性和服务功能[15]. ...
模拟增温对川西亚高山林线交错带绵穗柳生长、叶物候和叶性状的影响
1
2009
... 全球变暖对植物个体生长发育、物种组成和群落演变均有很大影响.在个体尺度上,增温对植物生长和繁殖有重要影响,例如气候变暖会使植物叶片生长和花期提前,导致植物加速成熟和衰老[7].另外,气候变暖还会增加植物的光合作用速率和蒸腾作用,导致水分流失加快.在干旱地区,这可能使植物水分胁迫加剧,限制植物生长[8].增温还对植物叶片形态、化学计量元素含量和种子质量等产生影响[9-10].而在群落尺度上,气候变暖会改变物种的分布,导致物种向极地或者高海拔地区迁移,引起植物群落结构和多样性的改变[11].同时,气候变暖会改变植物种间竞争强度来改变群落物种组成,增温可增强耐热物种竞争力,从而导致群落中不耐热物种的减少[12].增温还会导致群落中物种多样性或丰富度降低,如在北极苔原地区,短期的增温使灌木和禾草的盖度增加,但减少了物种多样性[13];增温使青藏高原区植物群落物种丰富度快速降低了26%~36%[14].在生态系统尺度上,气候变暖会改变生态系统物质和能量循环,进而影响生态系统稳定性和服务功能[15]. ...
南北样带温带区栎属树种功能性状间的关系及其对气象因子的响应
1
2010
... 全球变暖对植物个体生长发育、物种组成和群落演变均有很大影响.在个体尺度上,增温对植物生长和繁殖有重要影响,例如气候变暖会使植物叶片生长和花期提前,导致植物加速成熟和衰老[7].另外,气候变暖还会增加植物的光合作用速率和蒸腾作用,导致水分流失加快.在干旱地区,这可能使植物水分胁迫加剧,限制植物生长[8].增温还对植物叶片形态、化学计量元素含量和种子质量等产生影响[9-10].而在群落尺度上,气候变暖会改变物种的分布,导致物种向极地或者高海拔地区迁移,引起植物群落结构和多样性的改变[11].同时,气候变暖会改变植物种间竞争强度来改变群落物种组成,增温可增强耐热物种竞争力,从而导致群落中不耐热物种的减少[12].增温还会导致群落中物种多样性或丰富度降低,如在北极苔原地区,短期的增温使灌木和禾草的盖度增加,但减少了物种多样性[13];增温使青藏高原区植物群落物种丰富度快速降低了26%~36%[14].在生态系统尺度上,气候变暖会改变生态系统物质和能量循环,进而影响生态系统稳定性和服务功能[15]. ...
Biodiversity redistribution under climate change:impacts on ecosystems and human well-being
1
2017
... 全球变暖对植物个体生长发育、物种组成和群落演变均有很大影响.在个体尺度上,增温对植物生长和繁殖有重要影响,例如气候变暖会使植物叶片生长和花期提前,导致植物加速成熟和衰老[7].另外,气候变暖还会增加植物的光合作用速率和蒸腾作用,导致水分流失加快.在干旱地区,这可能使植物水分胁迫加剧,限制植物生长[8].增温还对植物叶片形态、化学计量元素含量和种子质量等产生影响[9-10].而在群落尺度上,气候变暖会改变物种的分布,导致物种向极地或者高海拔地区迁移,引起植物群落结构和多样性的改变[11].同时,气候变暖会改变植物种间竞争强度来改变群落物种组成,增温可增强耐热物种竞争力,从而导致群落中不耐热物种的减少[12].增温还会导致群落中物种多样性或丰富度降低,如在北极苔原地区,短期的增温使灌木和禾草的盖度增加,但减少了物种多样性[13];增温使青藏高原区植物群落物种丰富度快速降低了26%~36%[14].在生态系统尺度上,气候变暖会改变生态系统物质和能量循环,进而影响生态系统稳定性和服务功能[15]. ...
Improving the forecast for biodiversity under climate change
1
2016
... 全球变暖对植物个体生长发育、物种组成和群落演变均有很大影响.在个体尺度上,增温对植物生长和繁殖有重要影响,例如气候变暖会使植物叶片生长和花期提前,导致植物加速成熟和衰老[7].另外,气候变暖还会增加植物的光合作用速率和蒸腾作用,导致水分流失加快.在干旱地区,这可能使植物水分胁迫加剧,限制植物生长[8].增温还对植物叶片形态、化学计量元素含量和种子质量等产生影响[9-10].而在群落尺度上,气候变暖会改变物种的分布,导致物种向极地或者高海拔地区迁移,引起植物群落结构和多样性的改变[11].同时,气候变暖会改变植物种间竞争强度来改变群落物种组成,增温可增强耐热物种竞争力,从而导致群落中不耐热物种的减少[12].增温还会导致群落中物种多样性或丰富度降低,如在北极苔原地区,短期的增温使灌木和禾草的盖度增加,但减少了物种多样性[13];增温使青藏高原区植物群落物种丰富度快速降低了26%~36%[14].在生态系统尺度上,气候变暖会改变生态系统物质和能量循环,进而影响生态系统稳定性和服务功能[15]. ...
Plant community responses to experimental warming across the tundra biome
1
2006
... 全球变暖对植物个体生长发育、物种组成和群落演变均有很大影响.在个体尺度上,增温对植物生长和繁殖有重要影响,例如气候变暖会使植物叶片生长和花期提前,导致植物加速成熟和衰老[7].另外,气候变暖还会增加植物的光合作用速率和蒸腾作用,导致水分流失加快.在干旱地区,这可能使植物水分胁迫加剧,限制植物生长[8].增温还对植物叶片形态、化学计量元素含量和种子质量等产生影响[9-10].而在群落尺度上,气候变暖会改变物种的分布,导致物种向极地或者高海拔地区迁移,引起植物群落结构和多样性的改变[11].同时,气候变暖会改变植物种间竞争强度来改变群落物种组成,增温可增强耐热物种竞争力,从而导致群落中不耐热物种的减少[12].增温还会导致群落中物种多样性或丰富度降低,如在北极苔原地区,短期的增温使灌木和禾草的盖度增加,但减少了物种多样性[13];增温使青藏高原区植物群落物种丰富度快速降低了26%~36%[14].在生态系统尺度上,气候变暖会改变生态系统物质和能量循环,进而影响生态系统稳定性和服务功能[15]. ...
Experimental warming causes large and rapid species loss,dampened by simulated grazing on the Tibetan Plateau
1
2004
... 全球变暖对植物个体生长发育、物种组成和群落演变均有很大影响.在个体尺度上,增温对植物生长和繁殖有重要影响,例如气候变暖会使植物叶片生长和花期提前,导致植物加速成熟和衰老[7].另外,气候变暖还会增加植物的光合作用速率和蒸腾作用,导致水分流失加快.在干旱地区,这可能使植物水分胁迫加剧,限制植物生长[8].增温还对植物叶片形态、化学计量元素含量和种子质量等产生影响[9-10].而在群落尺度上,气候变暖会改变物种的分布,导致物种向极地或者高海拔地区迁移,引起植物群落结构和多样性的改变[11].同时,气候变暖会改变植物种间竞争强度来改变群落物种组成,增温可增强耐热物种竞争力,从而导致群落中不耐热物种的减少[12].增温还会导致群落中物种多样性或丰富度降低,如在北极苔原地区,短期的增温使灌木和禾草的盖度增加,但减少了物种多样性[13];增温使青藏高原区植物群落物种丰富度快速降低了26%~36%[14].在生态系统尺度上,气候变暖会改变生态系统物质和能量循环,进而影响生态系统稳定性和服务功能[15]. ...
Multiple dimensions of climate change and their implications for biodiversity
1
2018
... 全球变暖对植物个体生长发育、物种组成和群落演变均有很大影响.在个体尺度上,增温对植物生长和繁殖有重要影响,例如气候变暖会使植物叶片生长和花期提前,导致植物加速成熟和衰老[7].另外,气候变暖还会增加植物的光合作用速率和蒸腾作用,导致水分流失加快.在干旱地区,这可能使植物水分胁迫加剧,限制植物生长[8].增温还对植物叶片形态、化学计量元素含量和种子质量等产生影响[9-10].而在群落尺度上,气候变暖会改变物种的分布,导致物种向极地或者高海拔地区迁移,引起植物群落结构和多样性的改变[11].同时,气候变暖会改变植物种间竞争强度来改变群落物种组成,增温可增强耐热物种竞争力,从而导致群落中不耐热物种的减少[12].增温还会导致群落中物种多样性或丰富度降低,如在北极苔原地区,短期的增温使灌木和禾草的盖度增加,但减少了物种多样性[13];增温使青藏高原区植物群落物种丰富度快速降低了26%~36%[14].在生态系统尺度上,气候变暖会改变生态系统物质和能量循环,进而影响生态系统稳定性和服务功能[15]. ...
Impact of climate change drivers on C4 plants:a review
2
2020
... 气候变暖对植物的影响在不同植物种和不同区域间有很大差异.温度升高可短期内提高C3植物的光合作用速率,但长期高温会导致光呼吸增强,降低净光合作用效率[16-17].C4植物由于其独特的解剖结构和CO2同化机制,叶片净光合速率在干燥和温暖的环境条件下高于C3物种,因此C4植物比C3植物更耐高温胁迫[18].在全球尺度上,全球变暖对北半球植物生长的影响明显高于南半球,北半球植被长势增加趋势高于南半球[19].对于高山草甸和高寒草甸植物群落来说,模拟增温显著降低了群落物种多样性指数[20-21];但有研究却得出相反的结论,即增温和高寒草甸草原植物群落的物种丰富度呈正相关关系[22].另外,也有研究表明增温对荒漠草原植物群落物种多样性影响不显著[23].这也表明增温对植物群落的影响和区域差异、物种类型及增温时间等密切相关. ...
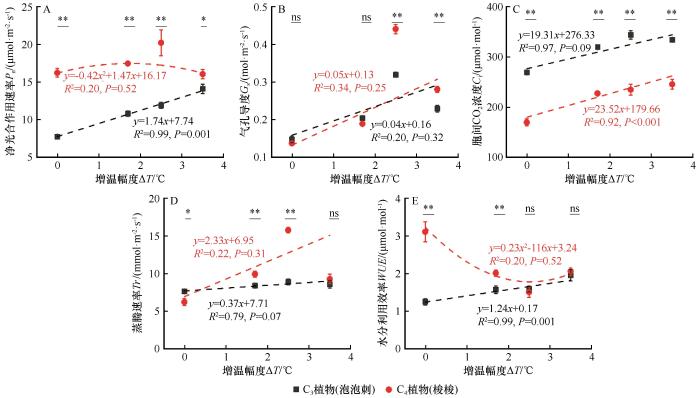
... 温度是影响植物光合作用的最重要因素,但不同植物光合作用对增温的响应却明显不同.增温会促进C4植物的光合作用,而抑制C3植物的光合作用,且C4植物净光合作用速率达到最大时的环境温度要显著高于C3植物[17,37].但也有研究提出不同结论,即一定程度的增温对C3和C4植物的光合作用都有促进作用,但是当增温幅度超过C3和C4物种叶片光合作用的最适温度后,增温会使植物气孔关闭并抑制其光合作用,但C4植物叶片光合作用的最适温度通常高于C3植物[16-17].本研究得出不同的结论,即增温会促进C3和C4植物的光合作用,且随着增温幅度的增加,C3植物泡泡刺Pn持续增大,而C4植物梭梭先增大后减小;但在相同增温幅度下,C4植物梭梭Pn高于C3植物泡泡刺.这说明本研究中模拟的3.5 ℃的增温还未达到泡泡刺和梭梭的最适温度,体现出荒漠植被对高温较好的适应性.主要原因是C4植物梭梭有独特的叶片解剖结构和CO2固定机制,这使其叶片Pn在干燥和温暖的环境条件下高于C3物种[38-39].但也有研究表明在干旱胁迫下C3植物红砂的Pn和Tr均高于C4植物珍珠(Caroxylon passerinum),体现出了两种植物对干旱荒漠生境的不同适应策略[40].有研究还表明增温会抑制梭梭的光合作用,使其Pn和Gs显著降低,主要原因是高温使梭梭叶片渗透调节物质含量和抗氧化酶活性升高,抑制了其光合作用[41].另外,虽然C4植物较高的净光合速率和较低的蒸腾速率决定了其比C3植物拥有更有效利用水分的能力,但这种能力在严重干旱条件下会丧失[42-43].这说明增温对不同植物光合作用的影响不仅与植物年龄及生长状况有关,也与其所处生境的其他环境要素紧密相关[27,44]. ...
荒漠草原植物群落特征和生态系统碳交换对增温增雨的响应机制
5
2024
... 气候变暖对植物的影响在不同植物种和不同区域间有很大差异.温度升高可短期内提高C3植物的光合作用速率,但长期高温会导致光呼吸增强,降低净光合作用效率[16-17].C4植物由于其独特的解剖结构和CO2同化机制,叶片净光合速率在干燥和温暖的环境条件下高于C3物种,因此C4植物比C3植物更耐高温胁迫[18].在全球尺度上,全球变暖对北半球植物生长的影响明显高于南半球,北半球植被长势增加趋势高于南半球[19].对于高山草甸和高寒草甸植物群落来说,模拟增温显著降低了群落物种多样性指数[20-21];但有研究却得出相反的结论,即增温和高寒草甸草原植物群落的物种丰富度呈正相关关系[22].另外,也有研究表明增温对荒漠草原植物群落物种多样性影响不显著[23].这也表明增温对植物群落的影响和区域差异、物种类型及增温时间等密切相关. ...
... 温度是影响植物光合作用的最重要因素,但不同植物光合作用对增温的响应却明显不同.增温会促进C4植物的光合作用,而抑制C3植物的光合作用,且C4植物净光合作用速率达到最大时的环境温度要显著高于C3植物[17,37].但也有研究提出不同结论,即一定程度的增温对C3和C4植物的光合作用都有促进作用,但是当增温幅度超过C3和C4物种叶片光合作用的最适温度后,增温会使植物气孔关闭并抑制其光合作用,但C4植物叶片光合作用的最适温度通常高于C3植物[16-17].本研究得出不同的结论,即增温会促进C3和C4植物的光合作用,且随着增温幅度的增加,C3植物泡泡刺Pn持续增大,而C4植物梭梭先增大后减小;但在相同增温幅度下,C4植物梭梭Pn高于C3植物泡泡刺.这说明本研究中模拟的3.5 ℃的增温还未达到泡泡刺和梭梭的最适温度,体现出荒漠植被对高温较好的适应性.主要原因是C4植物梭梭有独特的叶片解剖结构和CO2固定机制,这使其叶片Pn在干燥和温暖的环境条件下高于C3物种[38-39].但也有研究表明在干旱胁迫下C3植物红砂的Pn和Tr均高于C4植物珍珠(Caroxylon passerinum),体现出了两种植物对干旱荒漠生境的不同适应策略[40].有研究还表明增温会抑制梭梭的光合作用,使其Pn和Gs显著降低,主要原因是高温使梭梭叶片渗透调节物质含量和抗氧化酶活性升高,抑制了其光合作用[41].另外,虽然C4植物较高的净光合速率和较低的蒸腾速率决定了其比C3植物拥有更有效利用水分的能力,但这种能力在严重干旱条件下会丧失[42-43].这说明增温对不同植物光合作用的影响不仅与植物年龄及生长状况有关,也与其所处生境的其他环境要素紧密相关[27,44]. ...
... -17].本研究得出不同的结论,即增温会促进C3和C4植物的光合作用,且随着增温幅度的增加,C3植物泡泡刺Pn持续增大,而C4植物梭梭先增大后减小;但在相同增温幅度下,C4植物梭梭Pn高于C3植物泡泡刺.这说明本研究中模拟的3.5 ℃的增温还未达到泡泡刺和梭梭的最适温度,体现出荒漠植被对高温较好的适应性.主要原因是C4植物梭梭有独特的叶片解剖结构和CO2固定机制,这使其叶片Pn在干燥和温暖的环境条件下高于C3物种[38-39].但也有研究表明在干旱胁迫下C3植物红砂的Pn和Tr均高于C4植物珍珠(Caroxylon passerinum),体现出了两种植物对干旱荒漠生境的不同适应策略[40].有研究还表明增温会抑制梭梭的光合作用,使其Pn和Gs显著降低,主要原因是高温使梭梭叶片渗透调节物质含量和抗氧化酶活性升高,抑制了其光合作用[41].另外,虽然C4植物较高的净光合速率和较低的蒸腾速率决定了其比C3植物拥有更有效利用水分的能力,但这种能力在严重干旱条件下会丧失[42-43].这说明增温对不同植物光合作用的影响不仅与植物年龄及生长状况有关,也与其所处生境的其他环境要素紧密相关[27,44]. ...
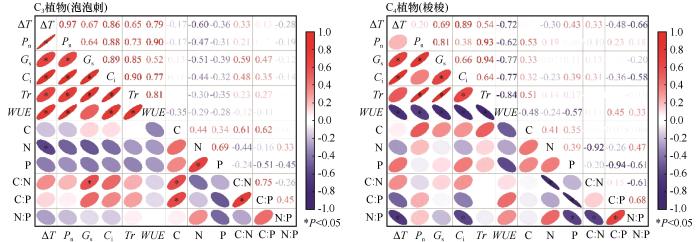
... 相关性分析表明,增温幅度和C3植物的光合作用指标均呈显著正相关关系,而和C4植物WUE呈显著负相关关系.对于C3植物,叶片N、P含量、N∶P均和光合作用指标呈负相关关系.这与前人的研究结果不尽相同,即增温会增加土壤中有效氮的含量,并对植物叶片氮含量产生积极影响,从而促进植物光合作用[17].主要原因是:一定幅度的增温促进了C3植物的光合作用,从而加强了N和P的消耗,降低了叶片N和P含量[48].另外,高温胁迫会引起土壤干旱,从而限制植物对N和P元素的吸收和利用[17].与C3植物泡泡刺不同的是,C4植物梭梭叶片C含量和光合指标间关系呈正相关关系(图5).主要原因一方面是C4植物叶肉细胞独特的解剖结构可以显著提高Rubisco酶的羧化效率,减少光呼吸碳损失,从而提高叶片C固定能力[38];另一方面,增温显著提高了C4植物的光合作用速率,加强了光合作用同化产物的合成,从而提高了叶片C含量[48].因此,植物光合指标和叶片化学计量元素间的相关关系受植物类型和环境要素共同驱动. ...
... [17].与C3植物泡泡刺不同的是,C4植物梭梭叶片C含量和光合指标间关系呈正相关关系(图5).主要原因一方面是C4植物叶肉细胞独特的解剖结构可以显著提高Rubisco酶的羧化效率,减少光呼吸碳损失,从而提高叶片C固定能力[38];另一方面,增温显著提高了C4植物的光合作用速率,加强了光合作用同化产物的合成,从而提高了叶片C含量[48].因此,植物光合指标和叶片化学计量元素间的相关关系受植物类型和环境要素共同驱动. ...
C4 photosynthesis and climate change
1
2019
... 气候变暖对植物的影响在不同植物种和不同区域间有很大差异.温度升高可短期内提高C3植物的光合作用速率,但长期高温会导致光呼吸增强,降低净光合作用效率[16-17].C4植物由于其独特的解剖结构和CO2同化机制,叶片净光合速率在干燥和温暖的环境条件下高于C3物种,因此C4植物比C3植物更耐高温胁迫[18].在全球尺度上,全球变暖对北半球植物生长的影响明显高于南半球,北半球植被长势增加趋势高于南半球[19].对于高山草甸和高寒草甸植物群落来说,模拟增温显著降低了群落物种多样性指数[20-21];但有研究却得出相反的结论,即增温和高寒草甸草原植物群落的物种丰富度呈正相关关系[22].另外,也有研究表明增温对荒漠草原植物群落物种多样性影响不显著[23].这也表明增温对植物群落的影响和区域差异、物种类型及增温时间等密切相关. ...
Global latitudinal-asymmetric vegetation growth trends and their driving mechanisms:1982-2009
1
2013
... 气候变暖对植物的影响在不同植物种和不同区域间有很大差异.温度升高可短期内提高C3植物的光合作用速率,但长期高温会导致光呼吸增强,降低净光合作用效率[16-17].C4植物由于其独特的解剖结构和CO2同化机制,叶片净光合速率在干燥和温暖的环境条件下高于C3物种,因此C4植物比C3植物更耐高温胁迫[18].在全球尺度上,全球变暖对北半球植物生长的影响明显高于南半球,北半球植被长势增加趋势高于南半球[19].对于高山草甸和高寒草甸植物群落来说,模拟增温显著降低了群落物种多样性指数[20-21];但有研究却得出相反的结论,即增温和高寒草甸草原植物群落的物种丰富度呈正相关关系[22].另外,也有研究表明增温对荒漠草原植物群落物种多样性影响不显著[23].这也表明增温对植物群落的影响和区域差异、物种类型及增温时间等密切相关. ...
模拟增温对云顶山亚高山草甸水热因子及群落结构的影响
1
2020
... 气候变暖对植物的影响在不同植物种和不同区域间有很大差异.温度升高可短期内提高C3植物的光合作用速率,但长期高温会导致光呼吸增强,降低净光合作用效率[16-17].C4植物由于其独特的解剖结构和CO2同化机制,叶片净光合速率在干燥和温暖的环境条件下高于C3物种,因此C4植物比C3植物更耐高温胁迫[18].在全球尺度上,全球变暖对北半球植物生长的影响明显高于南半球,北半球植被长势增加趋势高于南半球[19].对于高山草甸和高寒草甸植物群落来说,模拟增温显著降低了群落物种多样性指数[20-21];但有研究却得出相反的结论,即增温和高寒草甸草原植物群落的物种丰富度呈正相关关系[22].另外,也有研究表明增温对荒漠草原植物群落物种多样性影响不显著[23].这也表明增温对植物群落的影响和区域差异、物种类型及增温时间等密切相关. ...
季节性不对称模拟增温对青藏高原高寒草甸群落特征的影响
1
2022
... 气候变暖对植物的影响在不同植物种和不同区域间有很大差异.温度升高可短期内提高C3植物的光合作用速率,但长期高温会导致光呼吸增强,降低净光合作用效率[16-17].C4植物由于其独特的解剖结构和CO2同化机制,叶片净光合速率在干燥和温暖的环境条件下高于C3物种,因此C4植物比C3植物更耐高温胁迫[18].在全球尺度上,全球变暖对北半球植物生长的影响明显高于南半球,北半球植被长势增加趋势高于南半球[19].对于高山草甸和高寒草甸植物群落来说,模拟增温显著降低了群落物种多样性指数[20-21];但有研究却得出相反的结论,即增温和高寒草甸草原植物群落的物种丰富度呈正相关关系[22].另外,也有研究表明增温对荒漠草原植物群落物种多样性影响不显著[23].这也表明增温对植物群落的影响和区域差异、物种类型及增温时间等密切相关. ...
高寒草甸优势种功能多样性对增温和模拟放牧的响应
1
2021
... 气候变暖对植物的影响在不同植物种和不同区域间有很大差异.温度升高可短期内提高C3植物的光合作用速率,但长期高温会导致光呼吸增强,降低净光合作用效率[16-17].C4植物由于其独特的解剖结构和CO2同化机制,叶片净光合速率在干燥和温暖的环境条件下高于C3物种,因此C4植物比C3植物更耐高温胁迫[18].在全球尺度上,全球变暖对北半球植物生长的影响明显高于南半球,北半球植被长势增加趋势高于南半球[19].对于高山草甸和高寒草甸植物群落来说,模拟增温显著降低了群落物种多样性指数[20-21];但有研究却得出相反的结论,即增温和高寒草甸草原植物群落的物种丰富度呈正相关关系[22].另外,也有研究表明增温对荒漠草原植物群落物种多样性影响不显著[23].这也表明增温对植物群落的影响和区域差异、物种类型及增温时间等密切相关. ...
长期增温和氮素添加对荒漠草原不同植物功能群特征的影响
1
2022
... 气候变暖对植物的影响在不同植物种和不同区域间有很大差异.温度升高可短期内提高C3植物的光合作用速率,但长期高温会导致光呼吸增强,降低净光合作用效率[16-17].C4植物由于其独特的解剖结构和CO2同化机制,叶片净光合速率在干燥和温暖的环境条件下高于C3物种,因此C4植物比C3植物更耐高温胁迫[18].在全球尺度上,全球变暖对北半球植物生长的影响明显高于南半球,北半球植被长势增加趋势高于南半球[19].对于高山草甸和高寒草甸植物群落来说,模拟增温显著降低了群落物种多样性指数[20-21];但有研究却得出相反的结论,即增温和高寒草甸草原植物群落的物种丰富度呈正相关关系[22].另外,也有研究表明增温对荒漠草原植物群落物种多样性影响不显著[23].这也表明增温对植物群落的影响和区域差异、物种类型及增温时间等密切相关. ...
河西走廊绿洲生态建设和农业发展的若干思考与建议
1
2023
... 人工固沙植被建设是中国西北荒漠区荒漠化防治的最重要措施之一,其在绿洲风沙灾害防治及生物多样性维持等方面有重要意义[24].而荒漠灌木因其较强的耐干旱和高温胁迫能力,成为荒漠区人工植被建植的首选植物类型[25].为了探究荒漠植物对干旱和风沙生境的适应能力,以前的研究从生理特征、化学计量、形态特征和解剖结构等方面研究了荒漠植被对降水、风蚀和沙埋等的响应[26-29].但是,针对荒漠植被对气候变暖的响应相关的研究仍然十分匮乏,主要原因一是荒漠植被长期生长在极端高温及干旱的环境中,对气候变暖的敏感性相对较低;二是荒漠植被生长主要受降水而非温度调控.事实上,在过去的几十年间,中国西北地区气温也呈明显上升趋势,上升速率为0.34 ℃/10a,高于中国(0.25 ℃/10a)和全球平均水平(0.13 ℃/10a)[30].而目前对于荒漠植被尤其是荒漠灌木对增温响应的相关研究仍然较少,这直接限制了人们对于荒漠固沙植被适应气候变化的深入理解. ...
绿洲边缘人工固沙植被自组织过程
1
2018
... 人工固沙植被建设是中国西北荒漠区荒漠化防治的最重要措施之一,其在绿洲风沙灾害防治及生物多样性维持等方面有重要意义[24].而荒漠灌木因其较强的耐干旱和高温胁迫能力,成为荒漠区人工植被建植的首选植物类型[25].为了探究荒漠植物对干旱和风沙生境的适应能力,以前的研究从生理特征、化学计量、形态特征和解剖结构等方面研究了荒漠植被对降水、风蚀和沙埋等的响应[26-29].但是,针对荒漠植被对气候变暖的响应相关的研究仍然十分匮乏,主要原因一是荒漠植被长期生长在极端高温及干旱的环境中,对气候变暖的敏感性相对较低;二是荒漠植被生长主要受降水而非温度调控.事实上,在过去的几十年间,中国西北地区气温也呈明显上升趋势,上升速率为0.34 ℃/10a,高于中国(0.25 ℃/10a)和全球平均水平(0.13 ℃/10a)[30].而目前对于荒漠植被尤其是荒漠灌木对增温响应的相关研究仍然较少,这直接限制了人们对于荒漠固沙植被适应气候变化的深入理解. ...
梭梭(Haloxylon ammodendron)叶片气孔导度与气体交换对典型降水事件的响应
1
2014
... 人工固沙植被建设是中国西北荒漠区荒漠化防治的最重要措施之一,其在绿洲风沙灾害防治及生物多样性维持等方面有重要意义[24].而荒漠灌木因其较强的耐干旱和高温胁迫能力,成为荒漠区人工植被建植的首选植物类型[25].为了探究荒漠植物对干旱和风沙生境的适应能力,以前的研究从生理特征、化学计量、形态特征和解剖结构等方面研究了荒漠植被对降水、风蚀和沙埋等的响应[26-29].但是,针对荒漠植被对气候变暖的响应相关的研究仍然十分匮乏,主要原因一是荒漠植被长期生长在极端高温及干旱的环境中,对气候变暖的敏感性相对较低;二是荒漠植被生长主要受降水而非温度调控.事实上,在过去的几十年间,中国西北地区气温也呈明显上升趋势,上升速率为0.34 ℃/10a,高于中国(0.25 ℃/10a)和全球平均水平(0.13 ℃/10a)[30].而目前对于荒漠植被尤其是荒漠灌木对增温响应的相关研究仍然较少,这直接限制了人们对于荒漠固沙植被适应气候变化的深入理解. ...
巴丹吉林沙漠不同林龄梭梭(Haloxylon ammodendron)气体交换对增温的适应策略
2
2025
... 温度是影响植物光合作用的最重要因素,但不同植物光合作用对增温的响应却明显不同.增温会促进C4植物的光合作用,而抑制C3植物的光合作用,且C4植物净光合作用速率达到最大时的环境温度要显著高于C3植物[17,37].但也有研究提出不同结论,即一定程度的增温对C3和C4植物的光合作用都有促进作用,但是当增温幅度超过C3和C4物种叶片光合作用的最适温度后,增温会使植物气孔关闭并抑制其光合作用,但C4植物叶片光合作用的最适温度通常高于C3植物[16-17].本研究得出不同的结论,即增温会促进C3和C4植物的光合作用,且随着增温幅度的增加,C3植物泡泡刺Pn持续增大,而C4植物梭梭先增大后减小;但在相同增温幅度下,C4植物梭梭Pn高于C3植物泡泡刺.这说明本研究中模拟的3.5 ℃的增温还未达到泡泡刺和梭梭的最适温度,体现出荒漠植被对高温较好的适应性.主要原因是C4植物梭梭有独特的叶片解剖结构和CO2固定机制,这使其叶片Pn在干燥和温暖的环境条件下高于C3物种[38-39].但也有研究表明在干旱胁迫下C3植物红砂的Pn和Tr均高于C4植物珍珠(Caroxylon passerinum),体现出了两种植物对干旱荒漠生境的不同适应策略[40].有研究还表明增温会抑制梭梭的光合作用,使其Pn和Gs显著降低,主要原因是高温使梭梭叶片渗透调节物质含量和抗氧化酶活性升高,抑制了其光合作用[41].另外,虽然C4植物较高的净光合速率和较低的蒸腾速率决定了其比C3植物拥有更有效利用水分的能力,但这种能力在严重干旱条件下会丧失[42-43].这说明增温对不同植物光合作用的影响不仅与植物年龄及生长状况有关,也与其所处生境的其他环境要素紧密相关[27,44]. ...
... 荒漠C3植物泡泡刺和C4植物梭梭对模拟增温均有很好的适应性,在平均气温升高3 ℃左右的情况下,两种荒漠植物仍然能够通过提高净光合作用速率和水分利用效率来维持正常生长.因此,两种植物都可以作为荒漠区人工固沙植被建设的首选物种.但也有研究表明,不同林龄的人工梭梭林的气体交换特征对极端高温的响应有着较大差异,在极端高温下,中幼林梭梭Gs、WUE和Tr波动较小,且保持着较高的Pn,采取较为保守的水分利用策略来应对高温胁迫;而老龄梭梭则不断提高Tr致使WUE不断降低,虽然能保持较高的Pn,但也可能导致树木生长受限,面临死亡风险[27].因此,在理解荒漠人工固沙植被对全球气候变暖适应性时还应该充分考虑栽植年限对其适应性的影响,在荒漠人工固沙植被稳定性维持及其可持续性管理的过程中也应该考虑以上问题. ...
河西走廊荒漠绿洲边缘风沙生境沙拐枣的适应策略
1
2016
... 人工固沙植被建设是中国西北荒漠区荒漠化防治的最重要措施之一,其在绿洲风沙灾害防治及生物多样性维持等方面有重要意义[24].而荒漠灌木因其较强的耐干旱和高温胁迫能力,成为荒漠区人工植被建植的首选植物类型[25].为了探究荒漠植物对干旱和风沙生境的适应能力,以前的研究从生理特征、化学计量、形态特征和解剖结构等方面研究了荒漠植被对降水、风蚀和沙埋等的响应[26-29].但是,针对荒漠植被对气候变暖的响应相关的研究仍然十分匮乏,主要原因一是荒漠植被长期生长在极端高温及干旱的环境中,对气候变暖的敏感性相对较低;二是荒漠植被生长主要受降水而非温度调控.事实上,在过去的几十年间,中国西北地区气温也呈明显上升趋势,上升速率为0.34 ℃/10a,高于中国(0.25 ℃/10a)和全球平均水平(0.13 ℃/10a)[30].而目前对于荒漠植被尤其是荒漠灌木对增温响应的相关研究仍然较少,这直接限制了人们对于荒漠固沙植被适应气候变化的深入理解. ...
Why does the temperature rise faster in the arid region of northwest China?
1
2012
... 人工固沙植被建设是中国西北荒漠区荒漠化防治的最重要措施之一,其在绿洲风沙灾害防治及生物多样性维持等方面有重要意义[24].而荒漠灌木因其较强的耐干旱和高温胁迫能力,成为荒漠区人工植被建植的首选植物类型[25].为了探究荒漠植物对干旱和风沙生境的适应能力,以前的研究从生理特征、化学计量、形态特征和解剖结构等方面研究了荒漠植被对降水、风蚀和沙埋等的响应[26-29].但是,针对荒漠植被对气候变暖的响应相关的研究仍然十分匮乏,主要原因一是荒漠植被长期生长在极端高温及干旱的环境中,对气候变暖的敏感性相对较低;二是荒漠植被生长主要受降水而非温度调控.事实上,在过去的几十年间,中国西北地区气温也呈明显上升趋势,上升速率为0.34 ℃/10a,高于中国(0.25 ℃/10a)和全球平均水平(0.13 ℃/10a)[30].而目前对于荒漠植被尤其是荒漠灌木对增温响应的相关研究仍然较少,这直接限制了人们对于荒漠固沙植被适应气候变化的深入理解. ...
Physiological responses of Haloxylon ammodendron to rainfall pulses in temperate desert regions,Northwestern China
1
2014
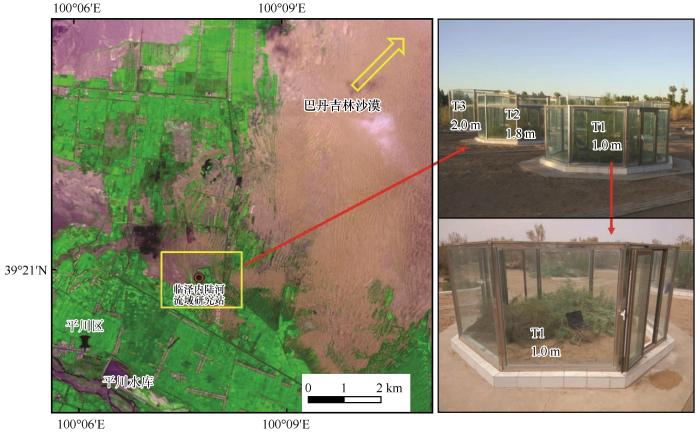
... 本研究试验场地位于甘肃省张掖市临泽县中国科学院临泽内陆河流域研究站内(图1),该区域属于典型的内陆性中温带干旱气候,多年平均降水量110 mm,降水呈显著季节变化,7—9月降水可占全年降水的50%以上,年均蒸发量为1 900~2 088 mm.年平均气温7.6 ℃,昼夜温差大,最高气温出现在7月(39.1 ℃),最低气温出现在1月(-27.3 ℃).土壤类型主要为灰棕漠土以及非地带性的风沙土.区域内地带性植被为中温带荒漠化草原植被,主要由超旱生灌木、半灌木、旱生肉质半灌木组成,典型灌木有泡泡刺(Nitraria sphaerocarpa)、梭梭(Haloxylon ammodendron)、沙拐枣(Calligonum mongolicum)和柽柳(Tamarix chinensis)等[31]. ...
内蒙古温带草原大针茅叶片光合生理特性对氮添加的响应
1
2023
... 植物光合指标测定和叶片化学计量元素采样在植物生长最旺盛的7月开展,在每种植物上选择生长相近且健康的叶片,利用Li-6400便携式光合测定系统(LI-COR, Lincoln,美国)测定两种植物的光合生理指标,自动记录净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)等值,每次读取5个测量参数[32].叶片水分利用效率根据WUE=Pn/Tr公式进行计算. ...
陆地生态系统野外增温控制实验的技术与方法
2
2020
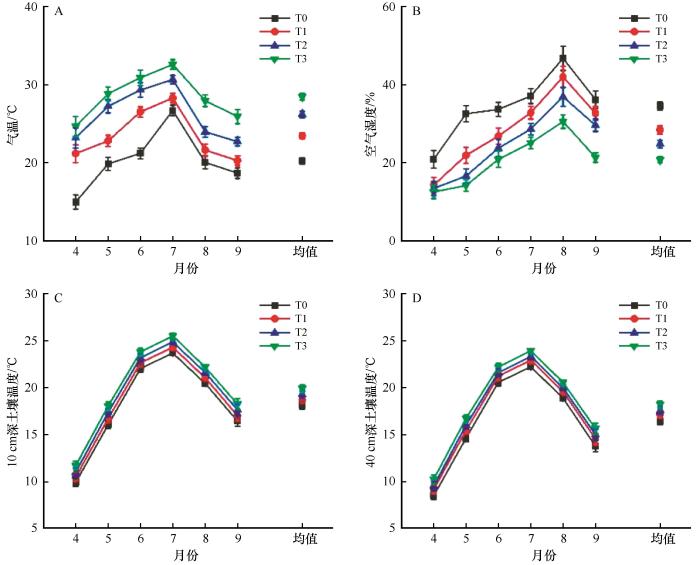
... OTC增温是陆地生态系统野外增温试验中常见的被动增温方法,其原理是利用开顶式的箱子将地面释放的长波辐射部分反射回植物和表层土壤,从而实现对生态系统的增温[33].该方法可以很好地模拟不同增温幅度对陆地生态系统过程的影响.在科尔沁沙地,1 m高的OTC增温罩可实现表层土壤10 cm平均温度增幅0.79 ℃[34].而在青藏高原的增温研究表明,高度为40、60、80、100 cm的OTC可以使空气温度分别提高2.2、2.8、3.2、3.6 ℃,使土壤温度分别提高0.9、1.9、2.1、2.1 ℃,而使土壤湿度降低3.1%、4.4%、7.2%、8.7%[35].高度为40 cm但底面积不同的OTC可使10 cm高空气温增加0.48~1.2 ℃,使10 cm深土壤温度增加0.6~1.25 ℃[36].本研究也有类似的发现,即在河西走廊中部荒漠区,1、1.8、2.3 m的OTC可使气温分别增加1.7、2.5、3.5 ℃,而整个生长季(4—9月)温度可增加3.20、5.93、8.22 ℃,空气湿度分别下降6.10%、9.69%、13.74%,10 cm深土层土壤平均温度分别增加0.61、1.19、1.83 ℃.这表明,OTC模拟增温可以不同程度地增加空气和土壤温湿度,但是对空气温湿度的影响要大于土壤温湿度.但是OTC装置在改变空气和土壤温湿度的同时也会引起风速和CO2浓度等其他气候因子的改变,因此在进行模拟增温研究时需要根据实际情况选择增温方式[33]. ...
... [33]. ...
科尔沁沙质草地植物群落对增温的响应
1
2024
... OTC增温是陆地生态系统野外增温试验中常见的被动增温方法,其原理是利用开顶式的箱子将地面释放的长波辐射部分反射回植物和表层土壤,从而实现对生态系统的增温[33].该方法可以很好地模拟不同增温幅度对陆地生态系统过程的影响.在科尔沁沙地,1 m高的OTC增温罩可实现表层土壤10 cm平均温度增幅0.79 ℃[34].而在青藏高原的增温研究表明,高度为40、60、80、100 cm的OTC可以使空气温度分别提高2.2、2.8、3.2、3.6 ℃,使土壤温度分别提高0.9、1.9、2.1、2.1 ℃,而使土壤湿度降低3.1%、4.4%、7.2%、8.7%[35].高度为40 cm但底面积不同的OTC可使10 cm高空气温增加0.48~1.2 ℃,使10 cm深土壤温度增加0.6~1.25 ℃[36].本研究也有类似的发现,即在河西走廊中部荒漠区,1、1.8、2.3 m的OTC可使气温分别增加1.7、2.5、3.5 ℃,而整个生长季(4—9月)温度可增加3.20、5.93、8.22 ℃,空气湿度分别下降6.10%、9.69%、13.74%,10 cm深土层土壤平均温度分别增加0.61、1.19、1.83 ℃.这表明,OTC模拟增温可以不同程度地增加空气和土壤温湿度,但是对空气温湿度的影响要大于土壤温湿度.但是OTC装置在改变空气和土壤温湿度的同时也会引起风速和CO2浓度等其他气候因子的改变,因此在进行模拟增温研究时需要根据实际情况选择增温方式[33]. ...
实验增温对藏北高寒草甸植物繁殖物候的影响
1
2016
... OTC增温是陆地生态系统野外增温试验中常见的被动增温方法,其原理是利用开顶式的箱子将地面释放的长波辐射部分反射回植物和表层土壤,从而实现对生态系统的增温[33].该方法可以很好地模拟不同增温幅度对陆地生态系统过程的影响.在科尔沁沙地,1 m高的OTC增温罩可实现表层土壤10 cm平均温度增幅0.79 ℃[34].而在青藏高原的增温研究表明,高度为40、60、80、100 cm的OTC可以使空气温度分别提高2.2、2.8、3.2、3.6 ℃,使土壤温度分别提高0.9、1.9、2.1、2.1 ℃,而使土壤湿度降低3.1%、4.4%、7.2%、8.7%[35].高度为40 cm但底面积不同的OTC可使10 cm高空气温增加0.48~1.2 ℃,使10 cm深土壤温度增加0.6~1.25 ℃[36].本研究也有类似的发现,即在河西走廊中部荒漠区,1、1.8、2.3 m的OTC可使气温分别增加1.7、2.5、3.5 ℃,而整个生长季(4—9月)温度可增加3.20、5.93、8.22 ℃,空气湿度分别下降6.10%、9.69%、13.74%,10 cm深土层土壤平均温度分别增加0.61、1.19、1.83 ℃.这表明,OTC模拟增温可以不同程度地增加空气和土壤温湿度,但是对空气温湿度的影响要大于土壤温湿度.但是OTC装置在改变空气和土壤温湿度的同时也会引起风速和CO2浓度等其他气候因子的改变,因此在进行模拟增温研究时需要根据实际情况选择增温方式[33]. ...
The phylogenetic structure of AMF communities shifts in response to gradient warming with and without winter grazing on the Qinghai-Tibet Plateau
1
2017
... OTC增温是陆地生态系统野外增温试验中常见的被动增温方法,其原理是利用开顶式的箱子将地面释放的长波辐射部分反射回植物和表层土壤,从而实现对生态系统的增温[33].该方法可以很好地模拟不同增温幅度对陆地生态系统过程的影响.在科尔沁沙地,1 m高的OTC增温罩可实现表层土壤10 cm平均温度增幅0.79 ℃[34].而在青藏高原的增温研究表明,高度为40、60、80、100 cm的OTC可以使空气温度分别提高2.2、2.8、3.2、3.6 ℃,使土壤温度分别提高0.9、1.9、2.1、2.1 ℃,而使土壤湿度降低3.1%、4.4%、7.2%、8.7%[35].高度为40 cm但底面积不同的OTC可使10 cm高空气温增加0.48~1.2 ℃,使10 cm深土壤温度增加0.6~1.25 ℃[36].本研究也有类似的发现,即在河西走廊中部荒漠区,1、1.8、2.3 m的OTC可使气温分别增加1.7、2.5、3.5 ℃,而整个生长季(4—9月)温度可增加3.20、5.93、8.22 ℃,空气湿度分别下降6.10%、9.69%、13.74%,10 cm深土层土壤平均温度分别增加0.61、1.19、1.83 ℃.这表明,OTC模拟增温可以不同程度地增加空气和土壤温湿度,但是对空气温湿度的影响要大于土壤温湿度.但是OTC装置在改变空气和土壤温湿度的同时也会引起风速和CO2浓度等其他气候因子的改变,因此在进行模拟增温研究时需要根据实际情况选择增温方式[33]. ...
Climatic warming changes plant photosynthesis and its temperature dependence in a temperate steppe of Northern China
1
2008
... 温度是影响植物光合作用的最重要因素,但不同植物光合作用对增温的响应却明显不同.增温会促进C4植物的光合作用,而抑制C3植物的光合作用,且C4植物净光合作用速率达到最大时的环境温度要显著高于C3植物[17,37].但也有研究提出不同结论,即一定程度的增温对C3和C4植物的光合作用都有促进作用,但是当增温幅度超过C3和C4物种叶片光合作用的最适温度后,增温会使植物气孔关闭并抑制其光合作用,但C4植物叶片光合作用的最适温度通常高于C3植物[16-17].本研究得出不同的结论,即增温会促进C3和C4植物的光合作用,且随着增温幅度的增加,C3植物泡泡刺Pn持续增大,而C4植物梭梭先增大后减小;但在相同增温幅度下,C4植物梭梭Pn高于C3植物泡泡刺.这说明本研究中模拟的3.5 ℃的增温还未达到泡泡刺和梭梭的最适温度,体现出荒漠植被对高温较好的适应性.主要原因是C4植物梭梭有独特的叶片解剖结构和CO2固定机制,这使其叶片Pn在干燥和温暖的环境条件下高于C3物种[38-39].但也有研究表明在干旱胁迫下C3植物红砂的Pn和Tr均高于C4植物珍珠(Caroxylon passerinum),体现出了两种植物对干旱荒漠生境的不同适应策略[40].有研究还表明增温会抑制梭梭的光合作用,使其Pn和Gs显著降低,主要原因是高温使梭梭叶片渗透调节物质含量和抗氧化酶活性升高,抑制了其光合作用[41].另外,虽然C4植物较高的净光合速率和较低的蒸腾速率决定了其比C3植物拥有更有效利用水分的能力,但这种能力在严重干旱条件下会丧失[42-43].这说明增温对不同植物光合作用的影响不仅与植物年龄及生长状况有关,也与其所处生境的其他环境要素紧密相关[27,44]. ...
The temperature responses of C3 and C4 photosynthesis
3
2007
... 温度是影响植物光合作用的最重要因素,但不同植物光合作用对增温的响应却明显不同.增温会促进C4植物的光合作用,而抑制C3植物的光合作用,且C4植物净光合作用速率达到最大时的环境温度要显著高于C3植物[17,37].但也有研究提出不同结论,即一定程度的增温对C3和C4植物的光合作用都有促进作用,但是当增温幅度超过C3和C4物种叶片光合作用的最适温度后,增温会使植物气孔关闭并抑制其光合作用,但C4植物叶片光合作用的最适温度通常高于C3植物[16-17].本研究得出不同的结论,即增温会促进C3和C4植物的光合作用,且随着增温幅度的增加,C3植物泡泡刺Pn持续增大,而C4植物梭梭先增大后减小;但在相同增温幅度下,C4植物梭梭Pn高于C3植物泡泡刺.这说明本研究中模拟的3.5 ℃的增温还未达到泡泡刺和梭梭的最适温度,体现出荒漠植被对高温较好的适应性.主要原因是C4植物梭梭有独特的叶片解剖结构和CO2固定机制,这使其叶片Pn在干燥和温暖的环境条件下高于C3物种[38-39].但也有研究表明在干旱胁迫下C3植物红砂的Pn和Tr均高于C4植物珍珠(Caroxylon passerinum),体现出了两种植物对干旱荒漠生境的不同适应策略[40].有研究还表明增温会抑制梭梭的光合作用,使其Pn和Gs显著降低,主要原因是高温使梭梭叶片渗透调节物质含量和抗氧化酶活性升高,抑制了其光合作用[41].另外,虽然C4植物较高的净光合速率和较低的蒸腾速率决定了其比C3植物拥有更有效利用水分的能力,但这种能力在严重干旱条件下会丧失[42-43].这说明增温对不同植物光合作用的影响不仅与植物年龄及生长状况有关,也与其所处生境的其他环境要素紧密相关[27,44]. ...
... WUE是评价植物对环境适应能力的综合指标,在同样的环境条件下,WUE值越大,表明固定单位CO2所需的水分越少,植物耐旱能力越强[44].通常来说,C4植物的WUE高于C3植物,且增温降低C3植物的WUE,却增大了C4植物的WUE.主要原因是对于C3植物来说,温度升高会促进其气孔开放,蒸腾作用加强,水分流失加快[45];另外高温还会降低Rubisco酶的羧化效率,减少CO2固定,最终导致WUE的降低[46].而C4植物独特的叶片结构使其对气温的敏感性较低,导致WUE对增温的响应更稳定[38].本研究有不同的发现,即虽然C3植物泡泡刺WUE显著低于C4植物梭梭,但是不同植物WUE和增温幅度的关系截然相反,C3植物泡泡刺WUE随着增温幅度的增加显著增大,而C4植物梭梭则随着增温幅度的增加先增大后降低.主要原因可能是OTC增温的同时也增加了CO2浓度,促进了C3植物泡泡刺光合作用,导致气孔导度的下降和WUE的增加[47].因此,在解释OTC模拟增温对不同植物光合作用影响时要综合考虑OTC导致的CO2浓度和土壤性质等其他因素的变化. ...
... 相关性分析表明,增温幅度和C3植物的光合作用指标均呈显著正相关关系,而和C4植物WUE呈显著负相关关系.对于C3植物,叶片N、P含量、N∶P均和光合作用指标呈负相关关系.这与前人的研究结果不尽相同,即增温会增加土壤中有效氮的含量,并对植物叶片氮含量产生积极影响,从而促进植物光合作用[17].主要原因是:一定幅度的增温促进了C3植物的光合作用,从而加强了N和P的消耗,降低了叶片N和P含量[48].另外,高温胁迫会引起土壤干旱,从而限制植物对N和P元素的吸收和利用[17].与C3植物泡泡刺不同的是,C4植物梭梭叶片C含量和光合指标间关系呈正相关关系(图5).主要原因一方面是C4植物叶肉细胞独特的解剖结构可以显著提高Rubisco酶的羧化效率,减少光呼吸碳损失,从而提高叶片C固定能力[38];另一方面,增温显著提高了C4植物的光合作用速率,加强了光合作用同化产物的合成,从而提高了叶片C含量[48].因此,植物光合指标和叶片化学计量元素间的相关关系受植物类型和环境要素共同驱动. ...
荒漠区植物光合器官解剖结构对水分利用效率的指示作用
1
2013
... 温度是影响植物光合作用的最重要因素,但不同植物光合作用对增温的响应却明显不同.增温会促进C4植物的光合作用,而抑制C3植物的光合作用,且C4植物净光合作用速率达到最大时的环境温度要显著高于C3植物[17,37].但也有研究提出不同结论,即一定程度的增温对C3和C4植物的光合作用都有促进作用,但是当增温幅度超过C3和C4物种叶片光合作用的最适温度后,增温会使植物气孔关闭并抑制其光合作用,但C4植物叶片光合作用的最适温度通常高于C3植物[16-17].本研究得出不同的结论,即增温会促进C3和C4植物的光合作用,且随着增温幅度的增加,C3植物泡泡刺Pn持续增大,而C4植物梭梭先增大后减小;但在相同增温幅度下,C4植物梭梭Pn高于C3植物泡泡刺.这说明本研究中模拟的3.5 ℃的增温还未达到泡泡刺和梭梭的最适温度,体现出荒漠植被对高温较好的适应性.主要原因是C4植物梭梭有独特的叶片解剖结构和CO2固定机制,这使其叶片Pn在干燥和温暖的环境条件下高于C3物种[38-39].但也有研究表明在干旱胁迫下C3植物红砂的Pn和Tr均高于C4植物珍珠(Caroxylon passerinum),体现出了两种植物对干旱荒漠生境的不同适应策略[40].有研究还表明增温会抑制梭梭的光合作用,使其Pn和Gs显著降低,主要原因是高温使梭梭叶片渗透调节物质含量和抗氧化酶活性升高,抑制了其光合作用[41].另外,虽然C4植物较高的净光合速率和较低的蒸腾速率决定了其比C3植物拥有更有效利用水分的能力,但这种能力在严重干旱条件下会丧失[42-43].这说明增温对不同植物光合作用的影响不仅与植物年龄及生长状况有关,也与其所处生境的其他环境要素紧密相关[27,44]. ...
干旱程度对C3植物红砂和C4植物珍珠光合生理参数的影响
1
2012
... 温度是影响植物光合作用的最重要因素,但不同植物光合作用对增温的响应却明显不同.增温会促进C4植物的光合作用,而抑制C3植物的光合作用,且C4植物净光合作用速率达到最大时的环境温度要显著高于C3植物[17,37].但也有研究提出不同结论,即一定程度的增温对C3和C4植物的光合作用都有促进作用,但是当增温幅度超过C3和C4物种叶片光合作用的最适温度后,增温会使植物气孔关闭并抑制其光合作用,但C4植物叶片光合作用的最适温度通常高于C3植物[16-17].本研究得出不同的结论,即增温会促进C3和C4植物的光合作用,且随着增温幅度的增加,C3植物泡泡刺Pn持续增大,而C4植物梭梭先增大后减小;但在相同增温幅度下,C4植物梭梭Pn高于C3植物泡泡刺.这说明本研究中模拟的3.5 ℃的增温还未达到泡泡刺和梭梭的最适温度,体现出荒漠植被对高温较好的适应性.主要原因是C4植物梭梭有独特的叶片解剖结构和CO2固定机制,这使其叶片Pn在干燥和温暖的环境条件下高于C3物种[38-39].但也有研究表明在干旱胁迫下C3植物红砂的Pn和Tr均高于C4植物珍珠(Caroxylon passerinum),体现出了两种植物对干旱荒漠生境的不同适应策略[40].有研究还表明增温会抑制梭梭的光合作用,使其Pn和Gs显著降低,主要原因是高温使梭梭叶片渗透调节物质含量和抗氧化酶活性升高,抑制了其光合作用[41].另外,虽然C4植物较高的净光合速率和较低的蒸腾速率决定了其比C3植物拥有更有效利用水分的能力,但这种能力在严重干旱条件下会丧失[42-43].这说明增温对不同植物光合作用的影响不仅与植物年龄及生长状况有关,也与其所处生境的其他环境要素紧密相关[27,44]. ...
模拟增温对梭梭光合生理生态特征的影响
1
2017
... 温度是影响植物光合作用的最重要因素,但不同植物光合作用对增温的响应却明显不同.增温会促进C4植物的光合作用,而抑制C3植物的光合作用,且C4植物净光合作用速率达到最大时的环境温度要显著高于C3植物[17,37].但也有研究提出不同结论,即一定程度的增温对C3和C4植物的光合作用都有促进作用,但是当增温幅度超过C3和C4物种叶片光合作用的最适温度后,增温会使植物气孔关闭并抑制其光合作用,但C4植物叶片光合作用的最适温度通常高于C3植物[16-17].本研究得出不同的结论,即增温会促进C3和C4植物的光合作用,且随着增温幅度的增加,C3植物泡泡刺Pn持续增大,而C4植物梭梭先增大后减小;但在相同增温幅度下,C4植物梭梭Pn高于C3植物泡泡刺.这说明本研究中模拟的3.5 ℃的增温还未达到泡泡刺和梭梭的最适温度,体现出荒漠植被对高温较好的适应性.主要原因是C4植物梭梭有独特的叶片解剖结构和CO2固定机制,这使其叶片Pn在干燥和温暖的环境条件下高于C3物种[38-39].但也有研究表明在干旱胁迫下C3植物红砂的Pn和Tr均高于C4植物珍珠(Caroxylon passerinum),体现出了两种植物对干旱荒漠生境的不同适应策略[40].有研究还表明增温会抑制梭梭的光合作用,使其Pn和Gs显著降低,主要原因是高温使梭梭叶片渗透调节物质含量和抗氧化酶活性升高,抑制了其光合作用[41].另外,虽然C4植物较高的净光合速率和较低的蒸腾速率决定了其比C3植物拥有更有效利用水分的能力,但这种能力在严重干旱条件下会丧失[42-43].这说明增温对不同植物光合作用的影响不仅与植物年龄及生长状况有关,也与其所处生境的其他环境要素紧密相关[27,44]. ...
C3/C4 grasslands and climate change
1
2010
... 温度是影响植物光合作用的最重要因素,但不同植物光合作用对增温的响应却明显不同.增温会促进C4植物的光合作用,而抑制C3植物的光合作用,且C4植物净光合作用速率达到最大时的环境温度要显著高于C3植物[17,37].但也有研究提出不同结论,即一定程度的增温对C3和C4植物的光合作用都有促进作用,但是当增温幅度超过C3和C4物种叶片光合作用的最适温度后,增温会使植物气孔关闭并抑制其光合作用,但C4植物叶片光合作用的最适温度通常高于C3植物[16-17].本研究得出不同的结论,即增温会促进C3和C4植物的光合作用,且随着增温幅度的增加,C3植物泡泡刺Pn持续增大,而C4植物梭梭先增大后减小;但在相同增温幅度下,C4植物梭梭Pn高于C3植物泡泡刺.这说明本研究中模拟的3.5 ℃的增温还未达到泡泡刺和梭梭的最适温度,体现出荒漠植被对高温较好的适应性.主要原因是C4植物梭梭有独特的叶片解剖结构和CO2固定机制,这使其叶片Pn在干燥和温暖的环境条件下高于C3物种[38-39].但也有研究表明在干旱胁迫下C3植物红砂的Pn和Tr均高于C4植物珍珠(Caroxylon passerinum),体现出了两种植物对干旱荒漠生境的不同适应策略[40].有研究还表明增温会抑制梭梭的光合作用,使其Pn和Gs显著降低,主要原因是高温使梭梭叶片渗透调节物质含量和抗氧化酶活性升高,抑制了其光合作用[41].另外,虽然C4植物较高的净光合速率和较低的蒸腾速率决定了其比C3植物拥有更有效利用水分的能力,但这种能力在严重干旱条件下会丧失[42-43].这说明增温对不同植物光合作用的影响不仅与植物年龄及生长状况有关,也与其所处生境的其他环境要素紧密相关[27,44]. ...
Impact of drought on photosynthesis:molecular perspective
1
2017
... 温度是影响植物光合作用的最重要因素,但不同植物光合作用对增温的响应却明显不同.增温会促进C4植物的光合作用,而抑制C3植物的光合作用,且C4植物净光合作用速率达到最大时的环境温度要显著高于C3植物[17,37].但也有研究提出不同结论,即一定程度的增温对C3和C4植物的光合作用都有促进作用,但是当增温幅度超过C3和C4物种叶片光合作用的最适温度后,增温会使植物气孔关闭并抑制其光合作用,但C4植物叶片光合作用的最适温度通常高于C3植物[16-17].本研究得出不同的结论,即增温会促进C3和C4植物的光合作用,且随着增温幅度的增加,C3植物泡泡刺Pn持续增大,而C4植物梭梭先增大后减小;但在相同增温幅度下,C4植物梭梭Pn高于C3植物泡泡刺.这说明本研究中模拟的3.5 ℃的增温还未达到泡泡刺和梭梭的最适温度,体现出荒漠植被对高温较好的适应性.主要原因是C4植物梭梭有独特的叶片解剖结构和CO2固定机制,这使其叶片Pn在干燥和温暖的环境条件下高于C3物种[38-39].但也有研究表明在干旱胁迫下C3植物红砂的Pn和Tr均高于C4植物珍珠(Caroxylon passerinum),体现出了两种植物对干旱荒漠生境的不同适应策略[40].有研究还表明增温会抑制梭梭的光合作用,使其Pn和Gs显著降低,主要原因是高温使梭梭叶片渗透调节物质含量和抗氧化酶活性升高,抑制了其光合作用[41].另外,虽然C4植物较高的净光合速率和较低的蒸腾速率决定了其比C3植物拥有更有效利用水分的能力,但这种能力在严重干旱条件下会丧失[42-43].这说明增温对不同植物光合作用的影响不仅与植物年龄及生长状况有关,也与其所处生境的其他环境要素紧密相关[27,44]. ...
C4荒漠植物猪毛菜与木本猪毛菜的叶片解剖结构及光合生理特征
2
2009
... 温度是影响植物光合作用的最重要因素,但不同植物光合作用对增温的响应却明显不同.增温会促进C4植物的光合作用,而抑制C3植物的光合作用,且C4植物净光合作用速率达到最大时的环境温度要显著高于C3植物[17,37].但也有研究提出不同结论,即一定程度的增温对C3和C4植物的光合作用都有促进作用,但是当增温幅度超过C3和C4物种叶片光合作用的最适温度后,增温会使植物气孔关闭并抑制其光合作用,但C4植物叶片光合作用的最适温度通常高于C3植物[16-17].本研究得出不同的结论,即增温会促进C3和C4植物的光合作用,且随着增温幅度的增加,C3植物泡泡刺Pn持续增大,而C4植物梭梭先增大后减小;但在相同增温幅度下,C4植物梭梭Pn高于C3植物泡泡刺.这说明本研究中模拟的3.5 ℃的增温还未达到泡泡刺和梭梭的最适温度,体现出荒漠植被对高温较好的适应性.主要原因是C4植物梭梭有独特的叶片解剖结构和CO2固定机制,这使其叶片Pn在干燥和温暖的环境条件下高于C3物种[38-39].但也有研究表明在干旱胁迫下C3植物红砂的Pn和Tr均高于C4植物珍珠(Caroxylon passerinum),体现出了两种植物对干旱荒漠生境的不同适应策略[40].有研究还表明增温会抑制梭梭的光合作用,使其Pn和Gs显著降低,主要原因是高温使梭梭叶片渗透调节物质含量和抗氧化酶活性升高,抑制了其光合作用[41].另外,虽然C4植物较高的净光合速率和较低的蒸腾速率决定了其比C3植物拥有更有效利用水分的能力,但这种能力在严重干旱条件下会丧失[42-43].这说明增温对不同植物光合作用的影响不仅与植物年龄及生长状况有关,也与其所处生境的其他环境要素紧密相关[27,44]. ...
... WUE是评价植物对环境适应能力的综合指标,在同样的环境条件下,WUE值越大,表明固定单位CO2所需的水分越少,植物耐旱能力越强[44].通常来说,C4植物的WUE高于C3植物,且增温降低C3植物的WUE,却增大了C4植物的WUE.主要原因是对于C3植物来说,温度升高会促进其气孔开放,蒸腾作用加强,水分流失加快[45];另外高温还会降低Rubisco酶的羧化效率,减少CO2固定,最终导致WUE的降低[46].而C4植物独特的叶片结构使其对气温的敏感性较低,导致WUE对增温的响应更稳定[38].本研究有不同的发现,即虽然C3植物泡泡刺WUE显著低于C4植物梭梭,但是不同植物WUE和增温幅度的关系截然相反,C3植物泡泡刺WUE随着增温幅度的增加显著增大,而C4植物梭梭则随着增温幅度的增加先增大后降低.主要原因可能是OTC增温的同时也增加了CO2浓度,促进了C3植物泡泡刺光合作用,导致气孔导度的下降和WUE的增加[47].因此,在解释OTC模拟增温对不同植物光合作用影响时要综合考虑OTC导致的CO2浓度和土壤性质等其他因素的变化. ...
Plant responses to drought and rewatering
1
2010
... WUE是评价植物对环境适应能力的综合指标,在同样的环境条件下,WUE值越大,表明固定单位CO2所需的水分越少,植物耐旱能力越强[44].通常来说,C4植物的WUE高于C3植物,且增温降低C3植物的WUE,却增大了C4植物的WUE.主要原因是对于C3植物来说,温度升高会促进其气孔开放,蒸腾作用加强,水分流失加快[45];另外高温还会降低Rubisco酶的羧化效率,减少CO2固定,最终导致WUE的降低[46].而C4植物独特的叶片结构使其对气温的敏感性较低,导致WUE对增温的响应更稳定[38].本研究有不同的发现,即虽然C3植物泡泡刺WUE显著低于C4植物梭梭,但是不同植物WUE和增温幅度的关系截然相反,C3植物泡泡刺WUE随着增温幅度的增加显著增大,而C4植物梭梭则随着增温幅度的增加先增大后降低.主要原因可能是OTC增温的同时也增加了CO2浓度,促进了C3植物泡泡刺光合作用,导致气孔导度的下降和WUE的增加[47].因此,在解释OTC模拟增温对不同植物光合作用影响时要综合考虑OTC导致的CO2浓度和土壤性质等其他因素的变化. ...
The response of photosynthesis and stomatal conductance to rising CO2:mechanisms and environmental interactions
1
2007
... WUE是评价植物对环境适应能力的综合指标,在同样的环境条件下,WUE值越大,表明固定单位CO2所需的水分越少,植物耐旱能力越强[44].通常来说,C4植物的WUE高于C3植物,且增温降低C3植物的WUE,却增大了C4植物的WUE.主要原因是对于C3植物来说,温度升高会促进其气孔开放,蒸腾作用加强,水分流失加快[45];另外高温还会降低Rubisco酶的羧化效率,减少CO2固定,最终导致WUE的降低[46].而C4植物独特的叶片结构使其对气温的敏感性较低,导致WUE对增温的响应更稳定[38].本研究有不同的发现,即虽然C3植物泡泡刺WUE显著低于C4植物梭梭,但是不同植物WUE和增温幅度的关系截然相反,C3植物泡泡刺WUE随着增温幅度的增加显著增大,而C4植物梭梭则随着增温幅度的增加先增大后降低.主要原因可能是OTC增温的同时也增加了CO2浓度,促进了C3植物泡泡刺光合作用,导致气孔导度的下降和WUE的增加[47].因此,在解释OTC模拟增温对不同植物光合作用影响时要综合考虑OTC导致的CO2浓度和土壤性质等其他因素的变化. ...
What have we learned from 15 years of free-air CO2 enrichment (FACE)?:a meta-analytic review of the responses of photosynthesis,canopy properties and plant production to rising CO2
1
2005
... WUE是评价植物对环境适应能力的综合指标,在同样的环境条件下,WUE值越大,表明固定单位CO2所需的水分越少,植物耐旱能力越强[44].通常来说,C4植物的WUE高于C3植物,且增温降低C3植物的WUE,却增大了C4植物的WUE.主要原因是对于C3植物来说,温度升高会促进其气孔开放,蒸腾作用加强,水分流失加快[45];另外高温还会降低Rubisco酶的羧化效率,减少CO2固定,最终导致WUE的降低[46].而C4植物独特的叶片结构使其对气温的敏感性较低,导致WUE对增温的响应更稳定[38].本研究有不同的发现,即虽然C3植物泡泡刺WUE显著低于C4植物梭梭,但是不同植物WUE和增温幅度的关系截然相反,C3植物泡泡刺WUE随着增温幅度的增加显著增大,而C4植物梭梭则随着增温幅度的增加先增大后降低.主要原因可能是OTC增温的同时也增加了CO2浓度,促进了C3植物泡泡刺光合作用,导致气孔导度的下降和WUE的增加[47].因此,在解释OTC模拟增温对不同植物光合作用影响时要综合考虑OTC导致的CO2浓度和土壤性质等其他因素的变化. ...
Biomass allocation to leaves,stems and roots:meta-analyses of interspecific variation and environmental control
3
2012
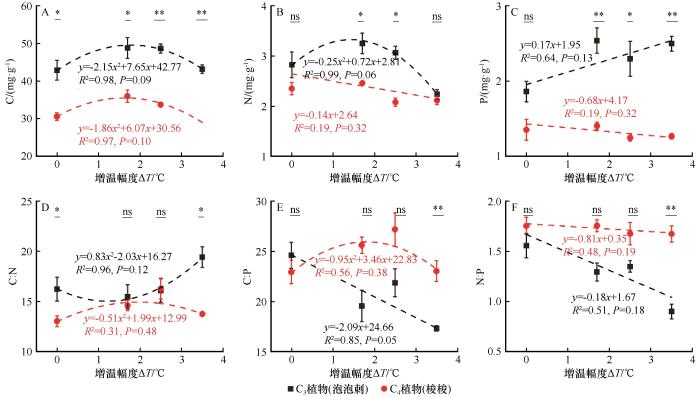
... 由于不同植被类型生理特性及养分吸收利用效率差异很大,因此不同生态系统中不同植物C、N和P含量对增温的响应也存在差异.本研究表明,C3植物泡泡刺叶片C、N和P含量显著高于C4植物梭梭.主要原因一是增温后两种植物的生物量分配策略不同,C4植物会将更多碳分配到结构性组织,而C3植物分配给非结构性碳水化合物;二是增温后两种植物光合作用系统响应不同,导致对C、N和P资源的吸收利用和分配策略不同[48].这反映出两种植物对增温的不同适应策略,C4植物在增温下叶片C、N和P含量较低,反映了其“快速生长-高效利用”的策略,而C3植物则体现出“高储存-高防御”的策略.本研究还表明泡泡刺和梭梭叶片C和N含量整体上均随着增温幅度的增加而减小,但泡泡刺P含量变化趋势相反(图4).主要原因是增温促进植物生长,增加了植物呼吸对C的消耗,从而导致C含量的下降[49-50].另外,增温也会加强植物的蒸腾作用,导致植物对营养元素的吸收减弱,进而降低植物C和N含量[51]. ...
... 相关性分析表明,增温幅度和C3植物的光合作用指标均呈显著正相关关系,而和C4植物WUE呈显著负相关关系.对于C3植物,叶片N、P含量、N∶P均和光合作用指标呈负相关关系.这与前人的研究结果不尽相同,即增温会增加土壤中有效氮的含量,并对植物叶片氮含量产生积极影响,从而促进植物光合作用[17].主要原因是:一定幅度的增温促进了C3植物的光合作用,从而加强了N和P的消耗,降低了叶片N和P含量[48].另外,高温胁迫会引起土壤干旱,从而限制植物对N和P元素的吸收和利用[17].与C3植物泡泡刺不同的是,C4植物梭梭叶片C含量和光合指标间关系呈正相关关系(图5).主要原因一方面是C4植物叶肉细胞独特的解剖结构可以显著提高Rubisco酶的羧化效率,减少光呼吸碳损失,从而提高叶片C固定能力[38];另一方面,增温显著提高了C4植物的光合作用速率,加强了光合作用同化产物的合成,从而提高了叶片C含量[48].因此,植物光合指标和叶片化学计量元素间的相关关系受植物类型和环境要素共同驱动. ...
... [48].因此,植物光合指标和叶片化学计量元素间的相关关系受植物类型和环境要素共同驱动. ...
Substantial nutrient resorption from leaves,stems and roots in a subarctic flora:What is the link with other resource economics traits?
1
2010
... 由于不同植被类型生理特性及养分吸收利用效率差异很大,因此不同生态系统中不同植物C、N和P含量对增温的响应也存在差异.本研究表明,C3植物泡泡刺叶片C、N和P含量显著高于C4植物梭梭.主要原因一是增温后两种植物的生物量分配策略不同,C4植物会将更多碳分配到结构性组织,而C3植物分配给非结构性碳水化合物;二是增温后两种植物光合作用系统响应不同,导致对C、N和P资源的吸收利用和分配策略不同[48].这反映出两种植物对增温的不同适应策略,C4植物在增温下叶片C、N和P含量较低,反映了其“快速生长-高效利用”的策略,而C3植物则体现出“高储存-高防御”的策略.本研究还表明泡泡刺和梭梭叶片C和N含量整体上均随着增温幅度的增加而减小,但泡泡刺P含量变化趋势相反(图4).主要原因是增温促进植物生长,增加了植物呼吸对C的消耗,从而导致C含量的下降[49-50].另外,增温也会加强植物的蒸腾作用,导致植物对营养元素的吸收减弱,进而降低植物C和N含量[51]. ...
增温对南亚热带常绿阔叶林4种幼树生长和碳氮磷化学计量特征的影响
1
2021
... 由于不同植被类型生理特性及养分吸收利用效率差异很大,因此不同生态系统中不同植物C、N和P含量对增温的响应也存在差异.本研究表明,C3植物泡泡刺叶片C、N和P含量显著高于C4植物梭梭.主要原因一是增温后两种植物的生物量分配策略不同,C4植物会将更多碳分配到结构性组织,而C3植物分配给非结构性碳水化合物;二是增温后两种植物光合作用系统响应不同,导致对C、N和P资源的吸收利用和分配策略不同[48].这反映出两种植物对增温的不同适应策略,C4植物在增温下叶片C、N和P含量较低,反映了其“快速生长-高效利用”的策略,而C3植物则体现出“高储存-高防御”的策略.本研究还表明泡泡刺和梭梭叶片C和N含量整体上均随着增温幅度的增加而减小,但泡泡刺P含量变化趋势相反(图4).主要原因是增温促进植物生长,增加了植物呼吸对C的消耗,从而导致C含量的下降[49-50].另外,增温也会加强植物的蒸腾作用,导致植物对营养元素的吸收减弱,进而降低植物C和N含量[51]. ...
黄渤海湿地芦苇光合特征对增温的响应
2
2023
... 由于不同植被类型生理特性及养分吸收利用效率差异很大,因此不同生态系统中不同植物C、N和P含量对增温的响应也存在差异.本研究表明,C3植物泡泡刺叶片C、N和P含量显著高于C4植物梭梭.主要原因一是增温后两种植物的生物量分配策略不同,C4植物会将更多碳分配到结构性组织,而C3植物分配给非结构性碳水化合物;二是增温后两种植物光合作用系统响应不同,导致对C、N和P资源的吸收利用和分配策略不同[48].这反映出两种植物对增温的不同适应策略,C4植物在增温下叶片C、N和P含量较低,反映了其“快速生长-高效利用”的策略,而C3植物则体现出“高储存-高防御”的策略.本研究还表明泡泡刺和梭梭叶片C和N含量整体上均随着增温幅度的增加而减小,但泡泡刺P含量变化趋势相反(图4).主要原因是增温促进植物生长,增加了植物呼吸对C的消耗,从而导致C含量的下降[49-50].另外,增温也会加强植物的蒸腾作用,导致植物对营养元素的吸收减弱,进而降低植物C和N含量[51]. ...
... 增温幅度的不同会导致植物叶片元素含量对增温的响应完全不同,例如低幅度的增温促进植物光合作用,加强CO2固定,导致C含量增加[51].相反,大幅度的增温不仅会加剧植物呼吸对C的消耗,也会引起土壤干旱,从而抑制植物对N和P元素的吸收[52-53].这也是本研究发现植物叶片C和N含量随着增温幅度的增加先增大后减小的主要原因.总之,增温对植物化学计量元素含量的影响不仅和植被类型有关,也与增温幅度和持续时间等密切相关. ...
Aridity modifies the responses of plant stoichiometry to global warming and nitrogen deposition in semi-arid steppes
1
2022
... 增温幅度的不同会导致植物叶片元素含量对增温的响应完全不同,例如低幅度的增温促进植物光合作用,加强CO2固定,导致C含量增加[51].相反,大幅度的增温不仅会加剧植物呼吸对C的消耗,也会引起土壤干旱,从而抑制植物对N和P元素的吸收[52-53].这也是本研究发现植物叶片C和N含量随着增温幅度的增加先增大后减小的主要原因.总之,增温对植物化学计量元素含量的影响不仅和植被类型有关,也与增温幅度和持续时间等密切相关. ...
陆地生态系统植物碳、氮和磷含量对增温的响应:Meta分析
1
2024
... 增温幅度的不同会导致植物叶片元素含量对增温的响应完全不同,例如低幅度的增温促进植物光合作用,加强CO2固定,导致C含量增加[51].相反,大幅度的增温不仅会加剧植物呼吸对C的消耗,也会引起土壤干旱,从而抑制植物对N和P元素的吸收[52-53].这也是本研究发现植物叶片C和N含量随着增温幅度的增加先增大后减小的主要原因.总之,增温对植物化学计量元素含量的影响不仅和植被类型有关,也与增温幅度和持续时间等密切相关. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}