


0 引言
在全球气候变化日益加剧的背景下,氧化亚氮(N2O)是仅次于二氧化碳(CO2)和甲烷(CH4)的长寿命温室气体,其浓度自工业革命以来上升超过24%,长期累积的气候反馈效应正引发全球范围的关注[1-2]。氮循环的相关研究多在农业和湿润生态系统。干旱区占全球的陆地面积40%,其对环境扰动的高度敏感性,是被严重低估的潜在热点[3-4]。土壤N2O排放受到水热等因素调控机制复杂,并且在干旱区极端气候频发的背景下,降水或灌溉引起的干湿交替会瞬间打破土壤原有水文与生物环境平衡,诱发典型的湿润脉冲效应,使N2O排放呈现特殊响应规律[5-6]。因此,探究干旱区水热因子对N2O排放规律的影响机制[7],是制定适应性减排策略的迫切需求。
生物土壤结皮(Biological soil crusts,BSCs)由蓝藻、地衣与苔藓等微生物群落与土壤颗粒共同构成,广泛分布于干旱半干旱地区,覆盖约12%的陆地表面,是中国西北干旱区的重要组成部分[8]。BSCs兼具地表结构-生物活性双重属性,不仅维系地表稳定,亦深刻影响土壤的物理、化学与生物过程[9]。BSCs在循环中发挥关键作用,通过光合作用与固氮作用,为表层土壤持续输入有机碳与活性氮,提升有机质含量、微生物生物量与养分供给,伴随BSCs演替,微环境结构愈发明显,从而为多途径氮转化发生提供条件[10]。目前多数研究聚焦于BSCs对碳、氮循环和水文过程等[11],然而对N2O排放的影响,尚存在欠缺。已有研究表明,BSCs能够通过固氮功能、增强硝化与反硝化活性来促进N2O排放[12]。在干旱区气候背景下,BSCs覆盖土壤对升温与降水脉冲尤为敏感,短期复湿常触发明显的脉冲式N2O排放峰值[13]。因此,BSCs与演替阶段对N2O通量的排放强度与调控机制仍需进一步深入研究。
综上所述,土壤温度、水分在协同调控N2O排放机理上尚不明确,并且BSCs在通气性、持水能力和微生物活性方面与沙土存在根本性差异,因此BSCs对N2O排放过程的影响值得探究。N2O主要源自硝化与反硝化反应过程,在BSCs与沙土中,两者反应过程如何随水热条件变化而发生转变仍不清楚。基于上述问题,本研究提出假设:①在干旱区,温度与水分是影响N2O排放的重要因素,且两者之间存在明显的交互作用;②BSCs通过改变微生境以影响硝化、反硝化作用,进而增强N2O排放。本研究以宁夏腾格里沙漠地区原状生物土壤结皮与裸沙土为研究对象,开展室内控制试验,设置不同梯度温度与水分处理,连续观测两类土壤的N2O通量动态与累积排放,解析BSCs对N2O排放的调控效应。通过乙炔抑制法鉴别硝化与反硝化反应过程对N2O排放的相对贡献,并测定物理、化学与生物学性质,解释环境因子和微生物过程对N2O排放的影响。该研究将有助于深化对干旱生态系统氮循环机制的理解,并为区域温室气体减排与氮管理策略提供依据。
1 材料与方法
1.1 研究区概况
土壤样品采集地点位于腾格里沙漠东南缘的沙坡头沙漠研究试验站以北约3 km范围内(37°32′—37°26′N,105°02′—104°30′E),海拔1 300~1 350 m。该区域年平均气温为9.6 ℃,冬季气温较低(1月平均温度为-6.9 ℃),夏季温暖(7月平均温度为24.3 ℃)。年降水量为186.5 mm,降水集中在5—9月。土壤类型主要为风沙土,土壤水分稳定在2%~3%。自然植被区包含多种灌木和亚灌木种类,如油蒿(Artemisia ordosica)、锦鸡儿(Caragana korshinskii)和红砂(Reaumuria soongorica)等[14]。
1.2 试验设计
本试验旨在研究不同温度和降水条件下沙土与BSCs的N2O排放通量。试验设定了3种温度条件:10 ℃(T10)作为基准温度,模拟干旱区春秋季节的土壤温度,适用于研究低温条件下土壤氮转化过程及N2O排放特征;20 ℃(T20)代表干旱区常见的土壤温度,反映温暖季节的土壤温度,适用于研究温度对N2O排放的影响,尤其是在春夏季节过渡期间;30 ℃(T30)模拟干旱区夏季高温土壤条件,用于研究温度对土壤微生物活动和N2O排放的影响。降水条件设定为0 mm和10 mm,其中,0 mm(W0)代表干旱条件下的无降水情景,模拟干旱区的极端干燥状态;10 mm(W10)代表干旱沙漠地区的有效降水量,通常触发土壤的湿润脉冲效应,用于研究其对N2O排放的影响。本试验采用乙炔抑制法(C₂H₂ inhibition method)判别N2O产生途径并量化反硝化潜力[15],试验设计为四因素全因子交叉设计,因素包括:土壤类型(沙土、BSCs)、温度(10、20、30 ℃)、降水(0、10 mm)和乙炔抑制(加、不加C₂H₂),共设24个处理,每个处理有4个重复,总计96组。
试验在研究区典型的固定沙丘区与生物结皮区内,用PVC圆筒(内径5 cm,高10 cm)自上而下垂直套取,分别采集裸沙土、生物结皮土壤(真藓,Bryum argenteum Hedw.)的原状土柱,剔除表面粗根与碎石,保持BSCs完整;装筒后以透气盖封口,低温、避光运输至实验室。所有样品于恒温室预培养48 h以消除扰动效应。培养容器为带采气口的密闭塑料盒,顶部设有丁基橡胶隔膜便于无损取气。预培养结束后,将土柱置于恒温暗箱中对应温度水平。10 mm的降水通过换算大约需要喷施20 mL超纯水,后静置30 min,在培养罐顶部采气口充入乙炔至体积分数10%(v/v);对照组(CK)不加乙炔。立即密封培养并开始定时取样,持续采集14 d。每次用无菌注射器经隔膜抽取10 mL顶空气体,注入预先抽真空的气袋,随后采用气相色谱仪(安捷伦8860,美国)进行测样。所有培养在暗条件下进行以排除光合作用干扰。
1.3 指标测定
1.3.1 气体排放通量计算
N2O排放通量计算公式:
式中:
N2O累积排放通量计算公式:
式中:M为N2O累积排放量(mg·m-2);F为瞬时排放通量(μg·m-2·h-1);dn+1-dn 为连续测量间隔时间(d)。
硝化贡献率:
1.3.2 土壤理化指标测定
在N2O气体采集结束,破坏土柱并混匀土壤,将每份土壤样品分为两部分处理:将一份土壤置于实验室阴凉干燥处自然风干,用于土壤肥力因子的测定;将另一份土壤无菌封装后保存于4 ℃冰箱中,用于微生物生物量测定。土壤含水量采用烘干法;电导率(EC)采用电导率仪测定;pH采用PB-10酸度仪测定;有机碳采用浓H2SO4-K2Cr2O7外加热法测定;采用凯氏法(全自动凯氏定氮仪SKD-1000,中国上海)测定土壤全氮(TN);速效磷(AP)采用0.5 mol·L-1 NaHCO3浸提,钼锑抗比色法测定(UV-1601,紫外可见分光光计,日本);测定土壤硝态氮(NO
1.4 数据处理与统计分析
本研究通过Origin 2024和R Studio 4.5.2等软件进行数据可视化。在统计分析部分,采用单因素方差分析(One-way ANOVA)、双因素方差分析(Two-way ANOVA)和三因素方差分析(Three-way ANOVA),用于检验不同处理组之间N2O排放通量及累积排放量的显著性差异(显著性水平α=0.05)。单因素方差分析主要用于评估单一因素(如温度或水分)对N2O排放的影响,双因素和三因素方差分析则进一步考察多个因素之间的交互作用。利用幂函数衰减模型分析不同处理下N2O排放趋势规律。偏最小二乘法结构方程模型(PLS-SEM)被用来分析土壤理化性质、微生物量与N2O排放之间的路径关系。PLS-SEM作为一种适用于多变量和复杂因果关系建模的统计方法,能够同时考虑多个独立变量和依赖变量之间的关系,并量化直接效应和间接效应。
2 结果与分析
2.1 N2O排放通量和累积排放量动态变化
随着培养时间增加,不同土壤类型下各处理N2O排放通量均表现出先快速下降后趋于平稳的变化趋势(图1)。不同处理下,N2O排放通量表现出明显的温湿度依赖性。各处理N2O排放通量排序为:W10T30>W10T20>W10T10>W0T30>W0T20>W0T10。在相同处理下,BSCs比沙土的N2O排放通量高、波动幅度大,且反硝化作用对N2O排放的贡献差异更明显。在BSCs中,W10T30处理的N2O排放峰值达到最高,其总排放量、反硝化作用和硝化作用驱动的N2O排放通量分别为40.76、32.05、9.71 μg·m²·h⁻¹;在W0T10处理下,N2O排放峰值最低,其总排放量、反硝化作用和硝化作用驱动的N2O排放通量分别为18.13、17.45、0.68 μg·m²·h⁻¹。
图1
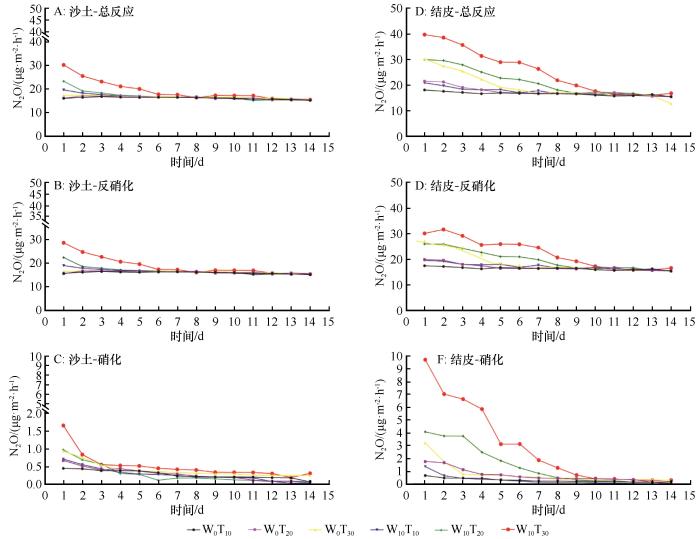
图1
N2O排放通量趋势
Fig.1
Trend of N2O emission flux
在3种反应过程下,随着培养天数的增加,两种土壤N2O累积排放量均呈上升趋势(图2)。在沙土的总反应中,随着温度增加,N2O累积排放量明显上升,W10T30处理下累积排放量最高(6.14 mg·m-2);硝化反应过程中,各处理累积排放平缓增加。在BSCs总反应过程中,各处理间排放差异明显,W10T30排放量最高(8.06 mg·m-2),其次是W10T20(6.73 mg·m-2),较W10T30降低了16.50%;硝化过程中,在1~6 d中累积排放量明显增加,各处理间差异明显。通过对比得出,W10T30处理下,BSCs累积排放量比沙土高23.94%、反硝化过程较沙土高16.94%、硝化过程较沙土高81.77%。
图2
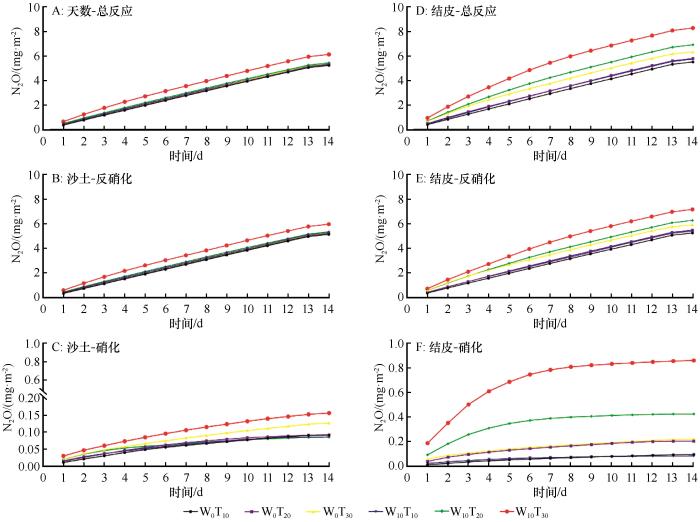
图2
N2O累积排放量趋势
Fig.2
Trend graph of cumulative N2O emissions
2.2 温度、降水和土壤类型对N2O的影响
多因素方差分析表明(表1),土壤类型、降水及温度变化均是N2O排放的关键变量,三者对各过程N2O排放通量及累积排放量均表现出极显著的独立影响(P<0.001)。在交互效应下不同反应过程中存在差异,主要关于硝化过程。其中BSCs在土壤和降水(P=0.005)及土壤和温度(P=0.004)下表现出比沙土更为敏感的硝化激发效应;降水和温度的协同作用加剧了硝化过程的排放(P=0.015);三因素交互作用(土壤类型、降水、温度)仅在硝化过程中达到显著水平(P=0.037)。
表1 温度、土壤类型、降水对N2O排放、累积排放通量的三因素方差分析
Table 1
| 影响因子 | 因变量 | N2O排放通量 | N2O累积排放通量 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 自由度 | 均方 | F值 | P值 | 自由度 | 均方 | F值 | P值 | |||
| 土壤类型 | 总反应 | 1 | 271.891 | 20.455 | 0.000 | 1 | 32 676.513 | 12.021 | 0.001 | |
| 反硝化 | 1 | 140.224 | 18.605 | 0.000 | 1 | 17 579.782 | 9.163 | 0.003 | ||
| 硝化 | 1 | 21.811 | 19.494 | 0.000 | 1 | 2 347.393 | 18.932 | 0.000 | ||
| 降水 | 总反应 | 1 | 230.853 | 17.368 | 0.000 | 1 | 26 348.350 | 9.693 | 0.002 | |
| 反硝化 | 1 | 143.733 | 19.071 | 0.000 | 1 | 16 720.353 | 8.715 | 0.004 | ||
| 硝化 | 1 | 10.418 | 9.311 | 0.003 | 1 | 1 108.008 | 8.936 | 0.003 | ||
| 温度 | 总反应 | 2 | 171.359 | 12.892 | 0.000 | 2 | 19 663.916 | 7.234 | 0.001 | |
| 反硝化 | 2 | 96.507 | 12.805 | 0.000 | 2 | 11 340.013 | 5.910 | 0.003 | ||
| 硝化 | 2 | 10.591 | 9.466 | 0.000 | 2 | 1 132.256 | 9.132 | 0.000 | ||
| 土壤类型×降水 | 总反应 | 1 | 47.931 | 3.606 | 0.050 | 1 | 7 195.752 | 2.647 | 0.106 | |
| 反硝化 | 1 | 15.065 | 1.999 | 0.159 | 1 | 2 766.099 | 1.442 | 0.232 | ||
| 硝化 | 1 | 9.115 | 8.147 | 0.005 | 1 | 1 021.559 | 8.239 | 0.005 | ||
| 土壤类型×温度 | 总反应 | 2 | 50.200 | 3.777 | 0.025 | 2 | 6 043.845 | 2.223 | 0.112 | |
| 反硝化 | 2 | 20.651 | 2.740 | 0.068 | 2 | 2 642.506 | 1.377 | 0.255 | ||
| 硝化 | 2 | 6.554 | 5.858 | 0.004 | 2 | 706.676 | 5.699 | 0.004 | ||
| 降水×温度 | 总反应 | 2 | 47.846 | 3.600 | 0.030 | 2 | 5 954.587 | 2.191 | 0.115 | |
| 反硝化 | 2 | 22.643 | 3.004 | 0.052 | 2 | 2 937.968 | 1.531 | 0.219 | ||
| 硝化 | 2 | 4.792 | 4.283 | 0.015 | 2 | 540.494 | 4.359 | 0.014 | ||
| 土壤类型×降水×温度 | 总反应 | 2 | 9.385 | 0.706 | 0.495 | 2 | 1 404.114 | 0.517 | 0.598 | |
| 反硝化 | 2 | 2.953 | 0.392 | 0.677 | 2 | 443.017 | 0.231 | 0.794 | ||
| 硝化 | 2 | 3.781 | 3.379 | 0.037 | 2 | 428.993 | 3.460 | 0.034 | ||
各处理下,N2O排放通量随着培养天数均呈初始高排放,后期非线性衰减的规律(图3)。在不同土壤类型、降水、温度条件下,N2O排放通量与培养天数的非线性曲线拟合均存在极显著正相关(P<0.001)。温度越高(T30>T20>T₁₀),初始排放通量越高,衰减速率也越快;降水显著提升了初始排放通量。随着培养天数的增加,W10T30处理下降速率比其他处理快。相比沙土,BSCs表现出更高的排放潜势及对温度变化的敏感性。随着温度和降水梯度的升高,相关系数R2从0.701提升至0.964。
图3
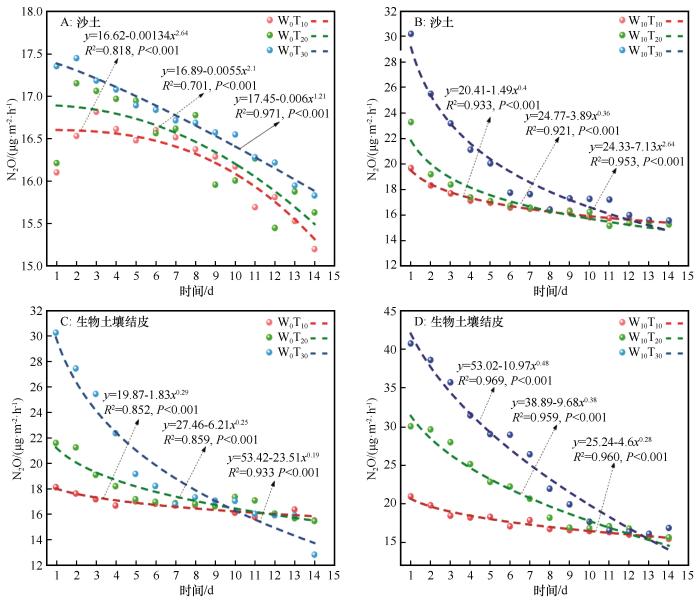
图3
不同处理下N2O排放通量与培养天数的非线性拟合
Fig.3
Nonlinear fitting between N2O emission flux and incubation days under different treatments
2.3 硝化与反硝化贡献率
乙炔抑制法结果表明,干旱区土壤N2O排放以反硝化过程为主,BSCs提升了硝化作用的贡献率(图4)。整体来看,反硝化过程是N2O产生的主要驱动力,其贡献率为79%~99%,硝化过程仅占1%~21%。不同土壤类型下,硝化与反硝化反应过程占比差异明显。相比于沙土,BSCs硝化贡献率在相同处理下高出3%~8%,在W10T30、W0T30处理尤为凸显;在沙土中,各处理反硝化占比排序为W₀T₁₀>W₀T₂₀>W₁₀T₁₀>W₀T₃₀>W₁₀T₂₀>W₁₀T₃₀,硝化占比与之相反。
图4
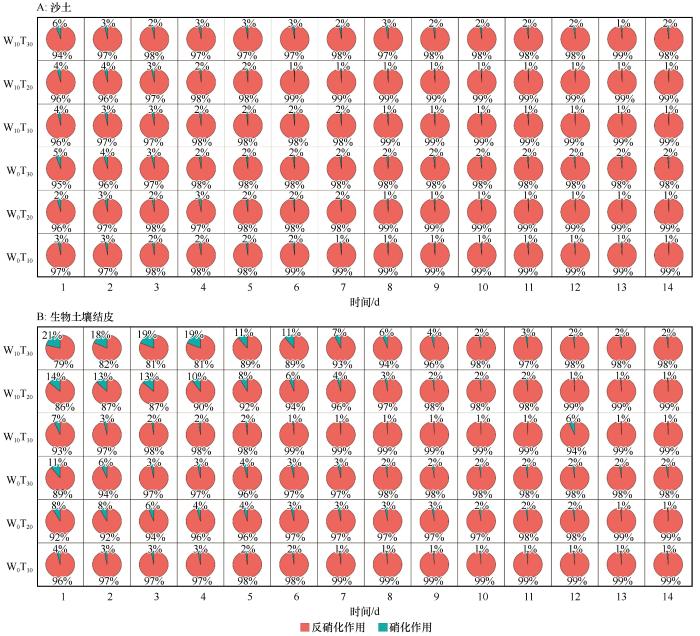
图4
硝化与反硝化贡献率
Fig.4
Analysis of contribution rates of nitrification and denitrification
2.4 土壤化学性质与微生物生物量对N2O排放通量的调控作用
不同处理间土壤化学性质与微生物生物量存在显著差异(图5)。总体来看,BSCs化学性质与微生物生物量含量均显著高于沙土(P<0.05);相较于反硝化过程,总反应过程下各处理的铵态氮(NH
图5
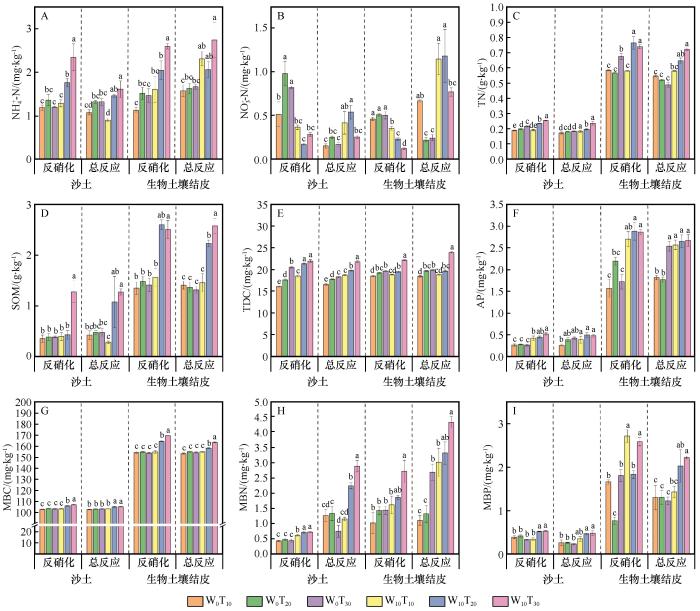
图5
不同处理土壤化学性质与生物学性质
Fig.5
Soil chemical and biological properties under different treatments
图6
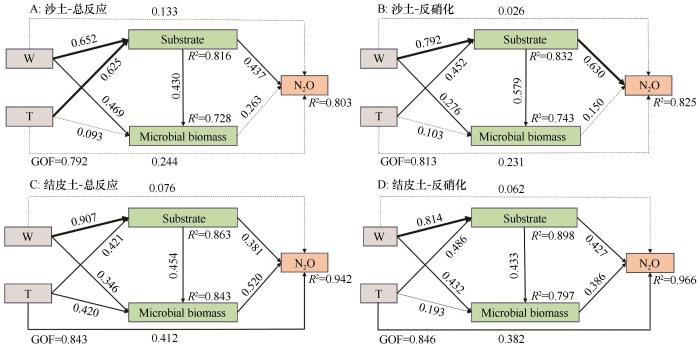
图6
土壤性质对N2O通量影响的偏最小二乘法结构方程模型
Fig.6
Partial least squares structural equation modeling (PLS-SEM) for effects of soil properties on N2O flux
3 讨论
3.1 温度和水分变化对N2O排放的影响
温度和降水是干旱区土壤N2O排放的关键水热驱动因子,直接调控土壤微生物代谢过程并影响其生理活性[16]。温度升高通过加速酶促反应、增强微生物代谢活性,进一步激活氮循环中的关键过程,从而显著促进N2O排放[17]。在实验中,W10T30处理下,沙土和BSCs的N2O排放量均达到峰值,验证了N2O排放随温度升高呈正指数响应的规律[18-19]。具体来说,高温条件下,微生物代谢活性和氮转化速率增加,导致N2O排放通量显著提升。降水事件通过改善土壤水分条件,引发典型的湿润脉冲效应,即干旱土壤在再湿润后迅速释放溶解的有机碳和无机氮,并伴随氧气扩散环境的变化和微生物的快速复苏,从而促进N2O的排放[20-21]。在试验中,降水后1 d内,N2O排放达到了峰值,且这一脉冲效应在6~7 d逐渐衰减并趋于平稳。这一现象表明,降水不仅是驱动N2O排放的重要因素,而且对N2O排放的影响具有时效性和强度的变化。
与沙土相比,BSCs对水热波动表现出更高的敏感性,这主要与其较高的有机质含量、丰富的微生物生物量及致密复杂的微生境结构有关,BSCs增强湿润脉冲条件下N2O排放的峰值和强度[22]。乙炔抑制处理结果表明,BSCs微生物群落对水分波动的适应与调节能力相对较弱。尽管较高的有机碳积累有助于维持较高的基础微生物活性,但致密的微生境结构可能限制微生物在短时间尺度内对水分变化的快速响应。相比之下,沙土有机碳含量较低、结构松散,微生物群落在水分变化后恢复更快,从而促进湿润脉冲后N2O排放的迅速增强[23]。综上所述,BSCs与沙土在湿润脉冲效应下的响应机制存在显著差异,其中微生物群落结构和有机碳供给在调控N2O排放过程中起关键作用。
3.2 沙土与BSCs对水热条件响应差异
不同土壤类型在温湿度变化下N2O排放响应呈现显著差异。BSCs因具备复杂的孔隙结构,形成大量微好氧-厌氧界面,可同时支持硝化与反硝化过程,因此微环境多样性是提升氮转化效率的结构基础[24]。温度升高时,BSCs表现尤为敏感,该响应可能与其胞外聚合物的温度敏感性密切相关[25];此类聚合物能增强土壤持水性并促进微生物生长,从而放大温度对微生物活性及氮循环的调控效应。在不同温度处理下,高含水量促使BSCs中氮转化的主导路径由硝化作用转向氨化作用[26],反映出其微生物群落能通过调整氮代谢途径适应水热变化,进而调控N2O排放。温湿度耦合变化通过调节微生物代谢活性,改变硝化与反硝化作用的相对贡献,最终影响N2O的生成与释放。
本研究表明,BSCs的N2O排放通量与累积排放量均高于沙土,表明BSCs凭借较高的微生物生物量与有机质积累,有效促进了N2O排放。BSCs中蓝细菌与真菌可通过强化氮矿化过程来提升氮循环效率,从而加剧N2O排放[27-28]。BSCs中高密度、高活性的微生物群落不仅提升了氮转化速率,也增强了系统对温湿度波动的响应;在湿润脉冲事件中,高温与适宜湿度共同作用会进一步提升N2O排放强度[29]。综上,BSCs在干旱区土壤中形成了独特的氮循环调控机制,使其在不同水热条件下均表现出较高的N2O排放潜力与环境敏感性。因此,BSCs不仅是干旱区氮转化过程的关键参与者,也可能在全球变暖背景下成为潜在的N2O排放源。
3.3 硝化与反硝化作用对N2O排放贡献的判别与机制
沙土具有低水分含量、低有机质和强透气性等特点,氮素底物稀缺且微生物活性较低,导致其整体N2O排放量较低[30]。降水能激活土壤微生物,填充土壤孔隙形成局部厌氧环境,促使反硝化作用主导N2O的排放,呈现出脉冲式的排放特征,成为沙土N2O排放的主要模式[31]。结合自然丰度同位素技术与乙炔抑制技术测定不同含水量下硝化和反硝化对N2O排放的贡献率,在一定范围内,含水量升高增强反硝化作用[15]。即使在极端干旱条件下,干燥土壤中仍能支持反硝化细菌,反硝化作用仍是沙土中N2O排放的主要途径[32]。本研究采用乙炔抑制法,通过比较抑制前后N2O排放通量的变化,评估硝化作用对N2O排放的贡献。结果显示,在沙土中,反硝化作用是N2O排放的绝对主导途径,其贡献率超过94%。虽然温度升高与水分增加会使反硝化贡献比例略有降低,硝化作用相应增强,但反硝化的主导地位并未改变。与之相比,BSCs中的N2O排放通量整体高于沙土,且硝化作用强度更高,即使施加乙炔抑制硝化过程,N2O排放以反硝化作用为主。上述结果证实乙炔可有效抑制硝化作用,并在本研究的沙漠环境中进一步表明反硝化是N2O产生的主要过程。这反映出沙土中反硝化微生物对N2O排放,尤其是在极端干旱条件下的高度适应性。
乙炔抑制法广泛应用于区分硝化与反硝化对N2O排放的贡献,其结果在复杂土壤环境或极端干旱条件下仍需审慎解读。例如,氧化还原条件等环境因素可能干扰硝化作用的真实表现。虽然乙炔抑制法在本研究中有效地区分了两者的贡献,但需要结合其他方法进行验证。例如,使用自然丰度同位素技术(如15N标记实验)或分子生物学方法(如16S rRNA基因测序)对微生物群落的结构进行分析,从而进一步确认硝化和反硝化过程的贡献。此外,还可以通过土壤氮形态分析或直接测定氮转化速率,来验证乙炔抑制法的结果。通过以上多角度的验证方法,确保乙炔抑制法结果的准确性,并提高对N2O排放机制的全面理解。
3.4 土壤理化性质与微生物群落耦合对N2O排放的影响机制
土壤理化性质与微生物群落的耦合作用是干旱区土壤N2O排放的重要驱动因素[33]。水热条件通过影响土壤基质特征和微生物代谢过程,间接调控N2O的排放。尤其是温度和水分的交互作用,能显著影响土壤中微生物群落的活动和氮转化过程[34]。在BSCs中,土壤性质与微生物群落之间耦合作用尤为显著,结构方程模型分析表明,BSCs中土壤性质与微生物群落的相互作用对N2O排放的路径系数均高于0.4,说明土壤和微生物的耦合效应在BSCs中显著增强。这一耦合效应揭示了BSCs独特的微环境特性,形成了有利于硝化和反硝化过程并行发生的条件,进而促进了氮氧化物(如N2O)的积累与释放[35]。本研究表明,在W10T30处理下,BSCs的NH
微生物群落在N2O排放过程中作用关键,既直接参与土壤氮循环,也通过改变土壤基质与底物供给间接调控N2O生成。在BSCs中,藻类、蓝藻等光合微生物通过固氮和有机碳输入显著改变土壤基质,增强了氮循环强度,从而促进N2O排放[37-38]。同时,微生物生物量与N2O排放呈正相关,表明较高的生物量可能提升N2O生成潜力[39]。但微生物生物量并不等同于代谢活性,后者还受土壤环境、群落结构和底物供应等多因素调控。因此,理解微生物对N2O排放的影响,需综合考虑群落结构、底物条件与水热环境。综上,BSCs的高微生物生物量及其独特的微环境共同提升了氮转化效率,加剧了N2O排放。水分变化既促进了氮底物的溶解与释放,也激发了微生物群落的代谢活性,进一步推升N2O排放。BSCs因其较强的保水性、有机质积累能力及生物活性,增强了环境因子对氮循环的调控作用,使得N2O排放在水分与温度变化下表现出更高的敏感性。因此,BSCs作为干旱区地表的关键调节层,不仅维持着氮循环的活跃状态,也通过其独特的微环境放大土壤对气候变化的响应,其通过强化氮转化过程、促进N2O释放,成为干旱区N2O的重要来源。BSCs的高微生物生物量、有机质积累能力以及对水热波动的敏感性使其在气候变化背景下发挥了重要作用。
4 结论
本研究通过模拟不同温度和降水,对比分析沙土与BSCs的N2O排放特征及驱动机制得出以下结论:①温度与水分是调控荒漠土壤N2O排放的关键因子,二者既独立作用又存在显著交互效应。升温加速了微生物代谢并增强排放,而降水脉冲则触发了短期N2O排放高峰。②BSCs显著增强了N2O排放。较高的有机质含量、微生物生物量为硝化和反硝化过程提供了有利条件,显著提高了N2O排放通量。③反硝化过程是荒漠土壤N2O排放的主要途径。在BSCs中,硝化作用的贡献明显增强,形成双路径调控。④土壤氮素底物和微生物生物量是N2O排放动态的核心驱动因子。温度和水分共同作用下,土壤氮素形态和微生物群落的活动对N2O排放过程起到了关键调控作用。水热因子的变化通过影响氮素底物和微生物生物量,从而提高了N2O排放通量。
参考文献
A comprehensive quantification of global nitrous oxide sources and sinks
[J].
The NOAA annual greenhouse gas index (AGGI)
[J].
A global meta-analysis of crop yield and agricultural greenhouse gas emissions under nitrogen fertilizer application
[J].
Large nitrogen oxide emission pulses from desert soils and associated microbiomes
[J].
Linking NO and N2O emission pulses with the mobilization of mineral and organic N upon rewetting dry soils
[J].
Responses of soil nitrogen and phosphorus cycling to drying and rewetting cycles:a meta-analysis
[J].
High soil temperatures alter the rates of nitrification,denitrification and associated N2O emissions
[J].
Biological Soil Crusts: Structure,Function,and Management
[M].
Biological soil crusts accelerate the nitrogen cycle through large NO and HONO emissions in drylands
[J].
Nitrogen cycling in desert biological soil crusts across biogeographic regions in the Southwestern United States
[J].
Precipitation patterns and their role in modulating nitrous oxide emissions from arid desert soil
[J].
Dynamics of short-term ecosystem carbon fluxes induced by precipitation events in a semiarid grassland
[J].
N2O accumulation from denitrification under different temperatures
[J].
Responses of soil microorganisms to simulated climate change in desert grassland in Northern China
[J].
Effects of drying-rewetting on soil CO2 emissions and the regulatory factors involved:a meta-analysis
[J].
Effects of dry-wet cycles on nitrous oxide emissions in freshwater sediments:a synthesis
[J].
Soil nutrients,enzyme activities,and microbial communities differ among biocrust types and soil layers in a degraded karst ecosystem
[J].
Moderate precipitation reduction enhances nitrogen cycling and soil nitrous oxide emissions in a semi‐arid grassland
[J].
The differential effects of sand burial on CO2,CH4,and N2O fluxes from desert biocrust-covered soils in the Tengger Desert,China
[J].
Soil nitrogen and climate drive the positive effect of biological soil crusts on soil organic carbon sequestration in drylands:a meta-analysis
[J].
Biological soil crusts increase the resistance of soil nitrogen dynamics to changes in temperatures in a semi-arid ecosystem
[J].
Soil HONO emissions at high moisture content are driven by microbial nitrate reduction to nitrite: tackling the HONO puzzle
[J].
Climate change and physical disturbance cause similar community shifts in biological soil crusts
[J].
Bacterial denitrification drives elevated N2O emissions in arid Southern California drylands
[J].
Temperature sensitivity of soil carbon decomposition and feedbacks to climate change
[J].
Water-driven microbial nitrogen transformations in biological soil crusts causing atmospheric nitrous acid and nitric oxide emissions
[J].
High rates of denitrification and nitrous oxide emission in arid biological soil crusts from the Sultanate of Oman
[J].
Effects of soil rewetting and thawing on soil gas fluxes: a review of current literature and suggestions for future research
[J].
Differences in carbon and nitrogen cycling strategies and regional variability in biological soil crust types
[J].
Bacteria and fungi differentially contribute to carbon and nitrogen cycles during biological soil crust succession in arid ecosystems
[J].
Effects of biological soil crusts on enzyme activities and microbial community in soils of an arid ecosystem
[J].

 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}