半干旱沙区3 种优势固沙灌木生物量分配及其生态学意义
詹瑾 , 1 , 李玉霖 , 1 , 韩丹 1 , 2 , 杨红玲 1 , 2
1.中国科学院西北生态环境资源研究院 奈曼沙漠化研究站,甘肃 兰州 730000
2.中国科学院大学,北京 100049
Biomass allocation and its ecological significance of three dominant sand -fixing shrubs in the semi -arid desert area
Zhan Jin , 1 , Li Yulin , 1 , Han Dan 1 , 2 , Yang Hongling 1 , 2
1.Naiman Desertification Research Station,Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou 730000,China
2.University of Chinese Academy of Sciences,Beijing 100049,China
通讯作者: 李玉霖(E-mail: liyl@lzb.ac.cn )
收稿日期: 2020-03-23
修回日期: 2020-06-09
网络出版日期: 2020-09-28
基金资助:
内蒙古自治区科技计划项目 . 201601052 国家重点研发计划项目 . 2016YFC0500907
Received: 2020-03-23
Revised: 2020-06-09
Online: 2020-09-28
作者简介 About authors
詹瑾(1993—),女,甘肃兰州人,硕士研究生,主要从事干旱半干旱区生态学相关研究E-mail:18394157943@163.com
, E-mail:18394157943@163.com
摘要
为探究科尔沁沙地3种优势固沙灌木生物量的分配特征,以黄柳(Salix gordejevii )、差不嘎蒿(Artemisia halodendron )和小叶锦鸡儿(Caragana microphylla )为对象,采用全挖法在个体水平上研究了固沙灌木生物量构件及其占比、R/SP <0.05);黄柳的老枝生物量及其占比居于首位,NAB(new aboveground biomass,新枝生物量)/OBA(old branch biomass,老枝生物量)为43.90%;差不嘎蒿的地上生物量占比居于首位,地上新生生物量是老枝生物量的2.43倍;黄柳和差不嘎蒿地下生物量集中分布在浅土层,不存在深根系(根系长度>100 cm)。(2)小叶锦鸡儿地下生物量及其占比和R/S(root/stem)均居于首位;地下生物量在根系长度>30 cm处的生物量占比达到61.61%,深根系(根系长度>100 cm)层地上生物量占比达到22.33%。(3)黄柳和差不嘎蒿的R/S、NAB/AGB(aboveground biomass,地上生物量)和NAB/OBA均与冠幅呈负线性关系,在固沙后期该两种灌木随着冠幅的增大,地上新生部分锐减,生产力下降,植被出现衰退。(4)通过非线性回归拟合得到3种灌木地上(y )-地下(x )生物量异速生长模型为:黄柳y =2.928x 1.039 (R 2 =0.901,P <0.01),差不嘎蒿y =32.802x 0.685 (R 2 =0.469,P <0.01),小叶锦鸡儿y =1.337x 0.066 (R 2 =0.833,P <0.01)。
关键词:
生物量分配 灌木 科尔沁沙地
Abstract
In order to explored the biomass allocation patterns of three dominant sand-fixing shrubs in Horqin Sandy Land, we investigated the biomass components and their proportions, R/S, proportions of above-ground new biomass, aboveground new biomass and old branch biomass ratio, and their correlation with crown width and the relationship between underground and above-ground biomass of Salix gordejevii , Artemisia halodendron and Caragana microphylla on the individual level. The results showed that: (1)The biomass and proportion of three dominant sand-fixing shrubs in Horqin Sandy Land were significantly different (P <0.05), the size and proportion of old branch biomass were the first and the NAB/OBA values were 43.90% about Salix gordejevii ; the proportion of above-ground biomass were the largest and the new above-ground biomass was 2.43 times that of old branch biomass about A. halodendron ; the Underground biomass were concentrated in shallow soil and there were no deep root system (root length>100 cm) of S. gordejevii and A. halodendron . (2)The size of underground biomass and its proportion, R/S were in the first place about C. microphylla ; the underground biomass was mainly distributed in the root length >30 cm layer,the proportion of the amount reached 61.61%, and the layer of deep roots (root length>100 cm) accounted for 22.33% of the total biomass. (3)The R/S, NAB/AGB and NAB/OBA of S. gordejevii and A. halodendron showed a negative linear relationship with the crown width; In the late stage of sand fixation, the above-ground new part of S. gordejevii and A. halodendron would decrease with the increase of crown width, productivity would decrease, and vegetations would decline. (4)The allogeneic growth models of aboveground and underground biomass of three shrubs were obtained by nonlinear regression fitting: Salix gordejevii y =2.928x 1.039 (R 2 =0.901, P <0.01) A. halodendron y =32.802x 0.685 (R 2 =0.469, P <0.01), C. microphylla y =1.337x 0.066 (R 2 =0.833, P <0.01).
Keywords:
biomass allocation shrubs Horqin Sandy Land
本文引用格式
詹瑾, 李玉霖, 韩丹, 杨红玲. 半干旱沙区3 种优势固沙灌木生物量分配及其生态学意义 . 中国沙漠 [J], 2020, 40(5): 149-157 doi:10.7522/j.issn.1000-694X.2020.00039
Zhan Jin, Li Yulin, Han Dan, Yang Hongling. Biomass allocation and its ecological significance of three dominant sand -fixing shrubs in the semi -arid desert area Journal of Desert Research
0 引言
生物量作为生态系统的能量基础和营养物质来源,反映植物对有机物质和能量的积累[1 -3 ] 。生物量分配是植物生活史理论的中心。植物通过生物量分配将营养器官同化的资源用于根、茎、叶、花、果等器官的生长,植物对多样环境做出适时调整,是植物经过长期进化而形成的生存策略[4 ] 。植物生物量分配受自身特性和环境条件的影响[5 ] ,在不同生境下具有不同的生长特性与物质分配规律[6 ] 。当植物生长环境中光照或养分条件发生变化时,植物会通过调整生物量分配和形态特征来获取当下环境中的最大效率养分,从而提高存活率[7 ] 。而在不同生长环境中,植物根、茎、叶等构件间存在竞争关系,通过内部自身权衡调配,在整个生活史中获取最理想且最适合自身发展的分配格局[8 ] 。王宏信等[9 ] 发现,毛乌素沙地3种灌木整株生物量及其在不同器官分配特征与沙地生境有关,茎的木质化程度、叶构型、根系分布情况会影响各器官的异速生长,进而影响了生物量的分配特征。丁俊祥等[10 ] 发现,古尔班通古特沙漠6种荒漠草本植物的根冠比和叶片生物量分配比例的种间差异,不仅与物种自身的遗传特性有关,还受植株个体大小的影响。王艳莉等[11 ] 发现,不同生境砂蓝刺头(Echinops gmelini )形态结构及生物量特征具有明显差异,而构件生物量间相关生长关系的一致规律性及较稳定的繁殖生物量分配,有利于其适应异质生境并完成生长发育。牛存洋等[12 ] 对科尔沁沙地小叶锦鸡儿(Caragana microphylla )地上-地下生物量异速生长模型拟合结果表明其符合异速生长模型的幂函数。
在干旱半干旱风沙区,固沙灌木凭其地上多分枝茎、近地面树冠及其地下根系分布较深的特性[13 ] ,面积超过总造林面积的70%[14 ] ,已成为生态建设和退化生态系统修复中防风固沙、保持水土和维持生态平衡的关键性植物。灌木生物量已成为灌木群落及生态系统研究的重要对象,不仅反映了灌木与其他植物的竞争能力,还反映出灌木对周边生态环境的适应程度[15 ] 。科尔沁沙地由于其脆弱的生态环境及强烈的人类活动,已成为中国北方农牧交错带沙漠化最严重的地区,在过去的几十年中,约80%的土地发生了不同程度的沙漠化[16 ] ,建植防风固沙灌木林是有效遏制该地区土地沙漠化和生态治理的重要措施[17 ] 。黄柳(Salix gordejevii )、差不嘎蒿(Artemisia halodendron )和小叶锦鸡儿是科尔沁沙地固沙灌木林的主要建群种[18 ] 。因此,以该地3种优势固沙灌木为研究对象,在个体水平上探讨3种固沙灌木生物量分配特征,不仅有助于了解沙地灌木对环境变化的响应机制和适应策略,还为沙地生境变化过程中植物种群变化、植被演替及植被管理提供科学依据。
1 材料与方法
1.1 研究区概况
科尔沁沙地处于中国东北部西辽河冲积-湖积平原地区,是内蒙古高原向东北平原的过渡地带,与浑善达克沙地、毛乌素沙地和呼伦贝尔沙地被称为中国四大沙地。科尔沁沙地总面积约5.06万km2 ,地理位置42.30°—45.30°N、119°—124°E,平均海拔178.5 m,属于温带大陆性半干旱气候[19 ] 。该地降水少,年际降水波动性大,年内降水不均,年降水量364.6 mm,集中在6—8月,年蒸发量1 972.8 mm。水资源短缺,总体呈现西部多、东部少的特点。年日照时数2 900~3 100 h,年平均气温5.8~6.4 ℃,≥10 ℃年积温3 000 ℃以上,无霜期约150 d。冬季多西北风,春季多西南风,年均风速3.6~4.1 m·s-1[20 -21 ] 。自第四世纪以来科尔沁沙地沉积了深厚松散的中沙和细沙,为土地沙化提供了物质基础,土壤类型有风沙土、草甸土、沙质栗钙土和沼泽土[22 ] ,以风沙土为主,土壤的机械组成中粒径1~0.25 mm沙粒占20%~58%,粒径0.25~0.05 mm沙粒占40%~67%,粒径<0.05 mm的粉粒和黏粒占0.1%~15%,土壤有机质含量0.08%~0.49%。形成缓起伏沙地、流动沙丘、半固定沙丘、固定沙丘与丘间低地镶嵌分布的现代风沙地貌景观。植被为典型的沙地疏林草原植被,主要有盐生和沙生植物,例如小叶锦鸡儿、狗尾草(Setaria viridis )、黄柳、芦苇(Phragmites communis )、糙隐子草(Cleistogenes squarrosa )、差不嘎蒿、黄蒿(Artemisia scoparia )、达乌里胡枝子(Lespedeza davurica )和白草(Pennisetum centrasiaticum )[23 ] 。
1.2 研究方法
本研究所用数据为小叶锦鸡儿、差不嘎蒿和黄柳的生物量实测数据。2012年7—8月在科尔沁沙地典型区(翁牛特旗、奈曼旗、库伦旗、科左后旗等)选取生长环境相对一致、天然生长的3种固沙灌木纯林进行调查,首先在野外测定植株的株丛高、冠幅长轴和短轴的长度。野外数据采集结束后,人工挖取整株灌木,在获取含有根系的土块后,除去样本植物根系上的土壤及其他植被的根系,同时剔除半分解状态的死根,整个过程要保证植被地下根系的完整性。随后,将其带回室内,按叶片、新枝和老枝(依据枝条颜色进行新枝和老枝的区分)及不同长度根系(0~5、5~10、10~20、20~30、30~50、50~70、70~100 cm和大于100 cm)进行分解并称取相对应的鲜重。分别取各部分中的部分鲜样,在75 ℃下烘干称重,依据样品各部分干鲜重质量比计算相对应每株灌木的各部分干重。3种优势固沙灌木基本信息见表1 。
1.3 数据处理
用Excel 2010对数据进行初步处理,用SPSS22.0对数据进行ANOVA方差分析、多重比较(Duncans法),Origin8.0进行作图。固沙灌木生物量结构的数量指标计算公式分别为:
地上生物量(aboveground biomass,AGB)=叶片生物量(leaf biomass,LB)+新枝生物量(new branch biomass,NBA)+老枝生物量(old branch biomass,OBA)
地下生物量(underground biomass,UGB)=各层根系干物质重之和
总生物量(total biomass,TB)=AGB+UGB
地上新生生物量(new aboveground biomass,NAB)/老枝生物量(OBA)=(地上新生生物量/老枝生物量)×100%
地上新生生物量(叶与新枝生物量之和)占比=(NAB /AGB) ×100%
根冠比(root-shoot ratio,R/S)=UGB /AGB×100%
选用较为常见的异速方程来构建3种灌木地上-地下生物量模型,其通用结构形式为:
y = β 0 x 1 β 1 x 2 β 2 ⋯ x j β j + ε
式中:y 为生物量;xj 为反映灌木形态特征变量;β 为参数;ε 为误差项,采用非线性方程法。科尔沁沙地优势固沙灌木生物量模型显示幂函数回归模型较为理想[15 ] ,本研究通过地上-地下生物量进行y =ax b 异速生长模型的幂函数建立,a 、b 为模型参数,y 为地上生物量,x 为地下生物量,对原始数据拟合得到的异速生长关系经过统计学检验得到拟合评定参数R 2 ,其可以直接反映地上-地下生物量相关程度[24 ] ,并通过显著性检验和F测验对模型进行校验。
2 结果与分析
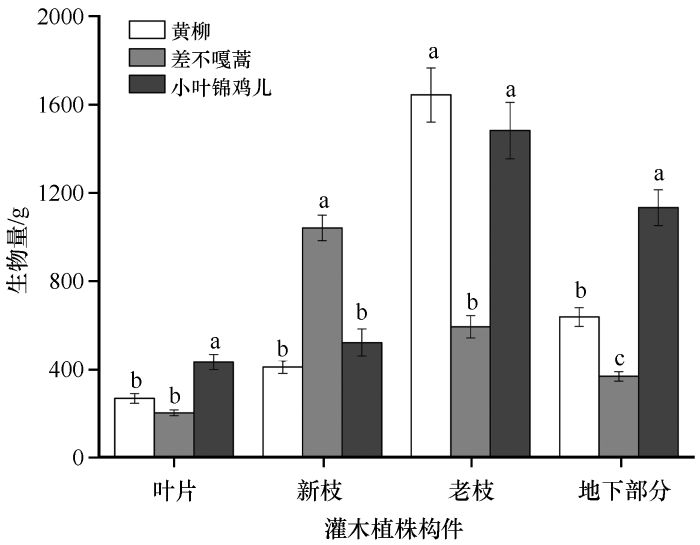
2.1 3 种优势固沙灌木各构件生物量基本特征
科尔沁沙地3种优势固沙灌木各结构生物量存在显著的种间差异(P <0.05,图1 )。其中叶片生物量中小叶锦鸡儿为432.89 g,显著高于差不嘎蒿(202.39 g)和黄柳(267.82 g);新枝生物量中差不嘎蒿为1 040.87 g,显著高于小叶锦鸡儿(521.01 g)和黄柳(410.85 g);老枝生物量中黄柳(1 643.91 g)和小叶锦鸡儿(1 483.06 g)显著高于差不嘎蒿(592.50 g);地下生物量的大小关系为小叶锦鸡儿(1 133.20 g)>黄柳(637.35 g)>差不嘎蒿(367.86 g)。
图1
图1
3种优势固沙灌木各构件生物量
不同字母表示差异显著,P <0.05
Fig.1
The biomass of each component of the three dominant sand-fixing shrubs
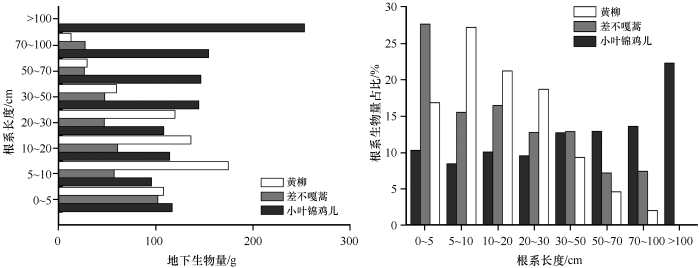
科尔沁沙地3种优势固沙灌木地下生物量均表现出明显的垂直空间分布特征(图2 ),其中黄柳和差不嘎蒿的地下生物量随着根系的不断伸长而逐渐递减,小叶锦鸡儿地下生物量随着根系的不断伸长而逐渐递增。黄柳的地下生物量集中分布在5~30 cm土层,该层生物量占比达到67.19%,差不嘎蒿的地下生物量集中分布在0~20 cm土层,该层生物量占比达到59.72%,且两者均不存在深根系(根系长度>100 cm);小叶锦鸡儿的地下生物量则集中分布在>30 cm土层,该层生物量占比达到61.61%,且深根系(根系长度>100 cm)层地上生物量占到总生物量的22.33%。
图2
图2
3种优势固沙灌木地下生物量的垂直分布及各层占比
Fig.2
Vertical distribution and each proportion of belowground biomass of the three dominant sand-fixing shrubs
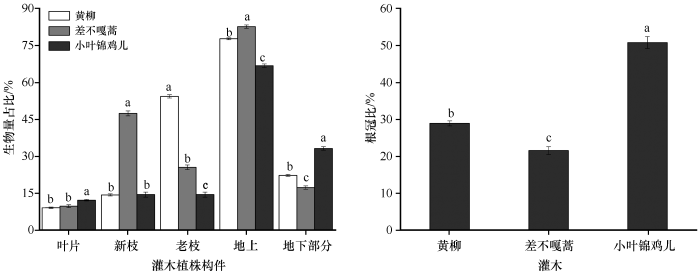
2.2 3 种优势固沙灌木各构件生物量占比、R/S 、NAB /AGB 及NAB/OBA
科尔沁沙地3种优势固沙灌木各构件生物量占比和R/S在不同物种间存在显著性差异(P <0.05,图3 )。叶片生物量占比中小叶锦鸡儿为12.16%,高于差不嘎蒿(9.76%)和黄柳(9.10%);新枝生物量占比中差不嘎蒿为47.42%,显著高于小叶锦鸡儿(14.42%)和黄柳(14.31%);老枝生物量占比的大小关系为黄柳(54.35%)>差不嘎蒿(25.53%)>小叶锦鸡儿(14.42%);地上生物量占比的大小关系为差不嘎蒿(82.71%)>黄柳(77.75%) >小叶锦鸡儿(66.80%);地下生物量占比和根冠比的大小关系均为小叶锦鸡儿>黄柳>差不嘎蒿。
图3
图3
3种优势固沙灌木各构件生物量占比和R/S
不同字母表示差异显著,P <0.05
Fig.3
The proportion of each biomass component and R/S of the three dominant sand-fixing shrubs
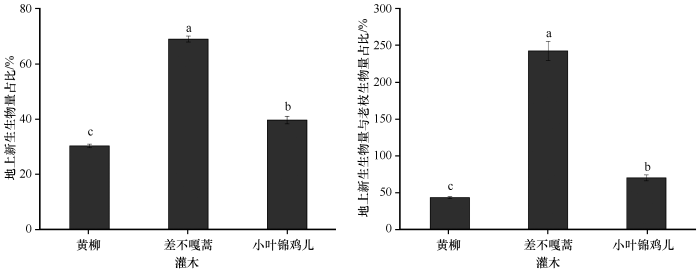
科尔沁沙地3种优势固沙灌木NAB /AGB和NAB/OBA在不同物种间存在显著性差异(P <0.05,图4 )。NAB/AGB的大小关系为差不嘎蒿(69.06%)>小叶锦鸡儿(39.57%)>黄柳(30.16%);在NAB/OBA中小叶锦鸡儿和黄柳均低于1,分别为70.69%和43.90%,而差不嘎蒿的地上新生生物量则是老枝生物量的2.43倍。
图4
图4
3种优势固沙灌木NAB /AGB和NAB/OBA
不同字母表示差异显著,P <0.05
Fig.4
The proportion of new biomass and ratio of aboveground biomass to old shoot biomass of the three dominant sand-fixing shrubs
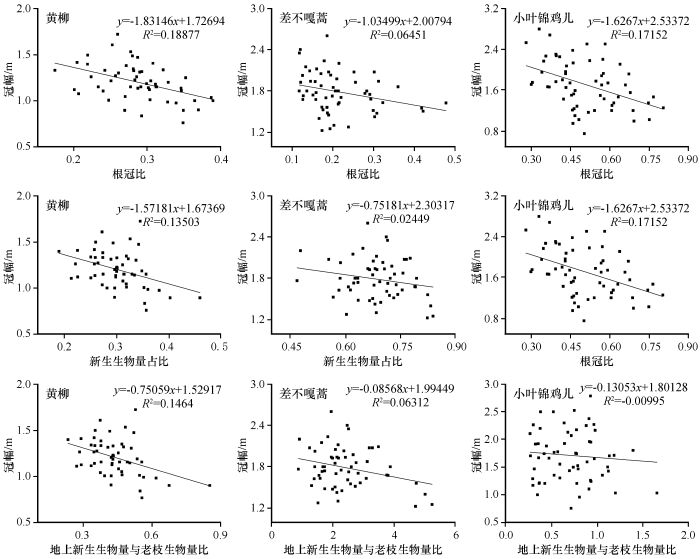
2.3 3 种优势固沙灌木R/S 、NAB /AGB 及NAB/OBA 与冠幅的关系
科尔沁沙地3种优势固沙灌木的R/S、NAB /AGB及NAB/OBA与冠幅的相关关系见表2 。黄柳的R/S、NAB /AGB、NAB/OBA均与冠幅极显著负相关(r =-0.453、r =-0.389、r =-0.403,P <0.01);差不嘎蒿的R/S、NAB/OBA均与冠幅呈显著负相关(r =-0.284、r =-0.283,P <0.05);而小叶锦鸡儿中仅R/S与冠幅呈极显著负相关(r =-0.430,P <0.01)。
科尔沁沙地3种优势固沙灌木R/S与冠幅线性拟合结果呈现负线性关系(R 黄 柳 2 R 差 不 嘎 蒿 2 R 小 叶 锦 鸡 儿 2 图5 );黄柳和差不嘎蒿NAB /AGB和NAB/OBA均与冠幅呈现负线性关系(R 黄 柳 2 R 差 不 嘎 蒿 2 R 黄 柳 2 R 差 不 嘎 蒿 2 R 小 叶 锦 鸡 儿 2 R 小 叶 锦 鸡 儿 2
图5
图5
3种优势固沙灌木R/S、NAB /AGB及NAB/OBA与冠幅的线性拟合关系
Fig.5
Linear regressions between crown and R/S,new biomass and ratio of aboveground biomass to old shoot biomass of three dominant sand-fixing shrubs
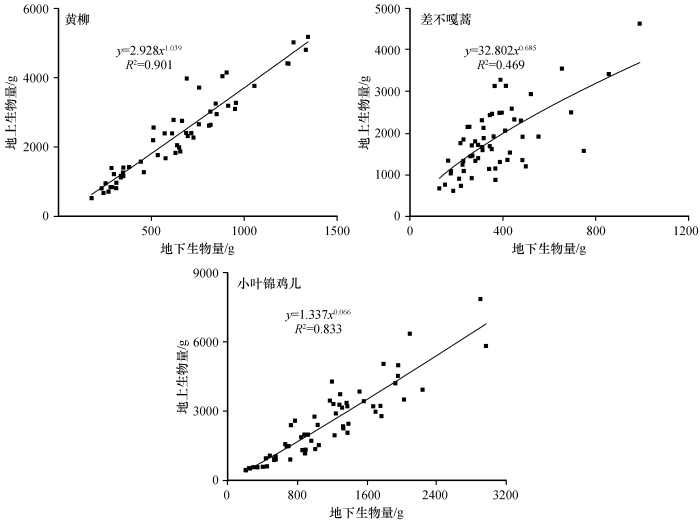
2.4 3 种优势固沙灌木地上-地下生物量异速生长模型分析
由图6 可见,科尔沁沙地3种优势固沙灌木地上-地下生物量关系符合典型的异速生长,即符合幂函数y =ax b 形式。通过非线性回归对异速生长模型的参数a 和b 进行拟合得到3种灌木地上-地下生物量异速生长模型:黄柳y =2.928x 1.039 (R 2 =0.901,P <0.01),差不嘎蒿y =32.802x 0.685 (R 2 =0.469,P <0.01),小叶锦鸡儿y =1.337x 0.066 (R 2 =0.833,P <0.01)。由回归测验可知F >F 0.01 ,P <0.01,因此地上-地下生物量存在显著的异速生长关系。
图6
图6
3种优势固沙灌木地上-地下生物量异速生长模型
Fig.6
Root-shoot biomass allometry growth model of the three dominant sand-fixing shrubs
3 讨论
生物量分配是植物对资源分配方式的反应,是通过光合和呼吸作用在不同器官间分配的结果[25 ] 。其在植物体内各构件的分配不仅受外界环境变化的调控,还与植物自身生长特性相关[26 -27 ] 。而灌木生物量作为灌木群落以及生态系统中十分重要的组成部分,是评价植被生产力的重要参数,在对灌木的生长和发育规律以及在生态系统中的地位和作用等研究方面具有十分重要意义[28 ] 。本研究中科尔沁沙地3种优势固沙灌木的各构件生物量及其占比、R/S、NAB /AGB及NAB/OBA均存在显著性差异(P <0.05),表明在同一生境中,不同的灌木物种对资源的利用能力不同。生物量分配的最优分配理论认为植物通过调节生物量在各器官的分配来适应外界环境,以获取光照、水分和养分等受限资源的最大效益,以维持其生长速率的最大化[29 ] 。本研究中科尔沁沙地3种优势固沙灌木生物量分配中黄柳的老枝生物量及其占比居于首位,NAB/OBA低于1;差不嘎蒿的地上生物量占比居于首位,地上新生生物量是老枝生物量的2.43倍;黄柳和差不嘎蒿地下生物量集中分布在浅土层,不存在深根系(根系长度>100 cm)。这表明在科尔沁沙地这种降水少、年内降水不均、水资源短缺、土壤养分较为贫瘠的环境中,黄柳和差不嘎蒿利用该生境中的资源主要用于进行地上部分的生长,地下部分受限,在固沙后期随着灌草丛(以浅根系草本为主)的出现,与黄柳和差不嘎蒿间存在对土壤浅层水分与养分资源的竞争。即植被在固沙过程中只能利用降雨后浅层土壤截留的少量地表水,同时存在对同一空间水分和养分等环境资源的竞争[30 ] 。在固沙初期植物对水分和养分的依赖较为强烈,植被倾向于把更多的资源分配于地下,且在植物生长发育过程中,与相邻植株存在光照、水分及养分竞争。而在养分和水分受限的条件下,R/S的增加有利于植物对养分和水分的吸收,可以增强植物的抗瘠和抗旱能力[31 ] 。本研究中小叶锦鸡儿地下生物量及其占比和R/S均居于首位,地下生物量主要集中分布在根系长度>30 cm处,该层生物量占比达到61.61%,且深根系(根系长度>100 cm)层地上生物量占到总生物量的22.33%。这说明随着在固沙后期小叶锦鸡儿将通过光合和呼吸作用积累的物质主要用于地下生物量的积累,随着灌木的生长,根系向更深的土层伸展,占据了更多土壤空间[1 ] ,及时从埋深较深的地下水补给根系所需水分,使其在3种优势固沙植被中独占优势,较耐旱,耐贫瘠。
冠幅作为植被进行光合作用的主要场所,不仅反映立足竞争力和生长活力,还在经营效果、生产潜力预估和养分循环等研究中存在重要的意义[32 ] 。黄柳和小叶锦鸡儿的R/S与冠幅极显著负相关(r =-0.453、r =-0.430,P <0.01),线性拟合结果呈现负线性关系(R 黄 柳 2 R 小 叶 锦 鸡 儿 2 r =-0.284,P <0.05),线性拟合结果呈现负线性关系(R 差 不 嘎 蒿 2 [33 ] 。而在科尔沁沙地3种植被固沙后期,随着植被生长,冠幅增大,地上部分增加,地下生物量并未同步增大,反而到了一定程度增加变缓,此时地下部分生长受限。其中差不嘎蒿R/S与冠幅的关系并无黄柳和小叶锦鸡儿明显,这是因为差不嘎蒿匍匐生长,生长过程中茎衍生不定根,冠幅越大,不定根越多。黄柳和差不嘎蒿NAB/AGB和NAB/OBA均与冠幅呈现负线性关系(R 黄 柳 2 R 差 不 嘎 蒿 2 R 黄 柳 2 R 差 不 嘎 蒿 2 r =-0.389、r =-0.403,P <0.01)。这说明随着植被生长冠幅的增大,新生生物量占比反而减小,地上新生部分减少,植被发生退化。在科尔沁沙地固沙后期,黄柳和差不嘎蒿随着自身生长冠幅的增大,同时受遗传和环境中水分、养分的影响,生产力在下降,植被衰退现象明显,而在小叶锦鸡儿中则不存在这种现象。
理论上获取生态系统生物量的方法有两种。其一直接测量法,即收获法所得数值虽较准确,但耗时费力,且对生态系统具有较大的破坏性;其二间接估测法,即通过异速生长关系来评估生物量,是目前较为普遍应用的方法[34 ] 。本研究参照牛存洋等[12 ] 对科尔沁沙地小叶锦鸡儿地上-地下生物量异速生长模型进行拟合的研究方法,通过地上-地下生物量关系,得出科尔沁沙地3种优势固沙灌木典型的幂函数异速生长模型,即符合幂函数y =ax b 形式,且地上-地下生物量存在显著的异速生长关系,其对黄柳和小叶锦鸡儿拟合较好。这说明3种灌木生物量分配具有不均匀性,与植物地上、地下生物量的相关生长关系与物种种类、生活型无关[35 -36 ] 的结论不一致。这可能与3种灌木在适应科尔沁沙地气候过程中,不同的资源分配策略有关,即差不嘎蒿匍匐生长,存在不定根。
4 结论
在降水少、年内降水不均、水资源短缺、土壤养分较为贫瘠的科尔沁沙地中,3种优势固沙灌木对资源的利用能力不同,其生物量分配存在显著性差异(P <0.05)。黄柳和差不嘎蒿主要用于地上部分生长,地下部分生长受限,其根系集中分布在浅土层,在固沙生长后期受遗传和环境中水分养分的影响,生产力下降,植被衰退现象明显。小叶锦鸡儿有较大的R/S, 具有深根系层,在固沙生长后期,根系向更深的土层伸展可及时从埋深较深的地下水补给水分,使其在3种优势固沙植被中独占优势,较耐旱,耐贫瘠。3种优势固沙灌木地上-地下生物量存在显著的异速生长,即符合幂函数y =axb 形式,尤其是黄柳和小叶锦鸡儿。
参考文献
View Option
[1]
杨昊天 ,李新荣 ,刘立超 ,等 荒漠草地4种灌木生物量分配特征
[J].中国沙漠 ,2013 ,33 (5 ):1340 -1348 .
[本文引用: 2]
[2]
刘长成 ,魏雅芬 ,刘玉国 ,等 贵州普定喀斯特次生林乔灌层地上生物量
[J].植物生态学报 ,2009 ,33 (4 ):698 -705 .
[3]
杨星华 毛竹单株地上部分生物量相容性模型研究
[D].北京 :北京林业大学 ,2016 .
[本文引用: 1]
[4]
金文云 猴耳环人工林地上部分生物量估测及其分配特征研究
[D].中国林业科学研究院 ,2018 .
[本文引用: 1]
[5]
Poorte A H Nagel O The role of biomass allocation in the growth response of plants to different levels of light,CO2 , nutrients and water:a quantitative review
[J].IMF Occasional Papers ,2000 ,27 (189 ):595 -607 .
[本文引用: 1]
[6]
钟泽兵 ,周国英 ,杨路存 ,等 柴达木盆地几种荒漠灌丛植被的生物量分配格局
[J].中国沙漠 ,2014 ,34 (4 ):1042 -1048 .
[本文引用: 1]
[7]
Navas M L Garnier E Plasticity of whole plant and leaf traits in Rubia peregrina in response to light, nutrient and water availability
[J].Acta Oecologica ,2002 ,23 (6 ):375 -383 .
[本文引用: 1]
[8]
安钰 ,安慧 宁夏荒漠草原优势植物生长及生物量分配对放牧干扰的响应
[J].西北植物学报 ,2015 ,35 (2 ):373 -378 .
[本文引用: 1]
[9]
王宏信 ,王伟峰 ,陈博 ,等 几种沙生灌木生物量及其在不同器官分配与土壤理化特征相关性
[J].土壤通报 ,2017 ,48 (5 ):1141 -1148 .
[本文引用: 1]
[10]
丁俊祥 ,范连连 ,李彦 ,等 古尔班通古特沙漠6种荒漠草本植物的生物量分配与相关生长关系
[J].中国沙漠 ,2016 ,36 (5 ):1323 -1330 .
[本文引用: 1]
[11]
王艳莉 ,齐欣宇 ,杨昊天 ,等 不同生境下砂蓝刺头(Echinops gmelini )形态结构及生物量分配特征
[J].中国沙漠 ,2018 ,38 (4 ):756 -764 .
[本文引用: 1]
[12]
牛存洋 ,阿拉木萨 ,宗芹 ,等 科尔沁沙地小叶锦鸡儿地上-地下生物量分配格局
[J].生态学杂志 ,2013 ,32 (8 ):1980 -1986 .
[本文引用: 2]
[13]
王蕾 ,张宏 ,哈斯 ,等 基于冠幅直径和植株高度的灌木地上生物量估测方法研究
[J].北京师范大学学报(自然科学版) ,2004 ,40 (5 ):700 -704 .
[本文引用: 1]
[14]
蒋齐 ,李生宝 ,翟明普 我国的固沙型灌木林及其研究进展
[J].干旱区资源与环境 ,1998 ,12 (2 ):88 -96 .
[本文引用: 1]
[15]
童新风 ,杨红玲 ,宁志英 ,等 科尔沁沙地优势固沙灌木的生物量预测模型
[J].中国沙漠 ,2018 ,38 (3 ):553 -559 .
[本文引用: 2]
[16]
Li Y Q Zhou X H Brandle J R et al Temporal progress in improving carbon and nitrogen storage by grazing exclosure practice in a degraded land area of China’s Horqin Sandy Grassland
[J].Agriculture Ecosystems & Environment ,2012 ,159 :55 -61 .
[本文引用: 1]
[17]
赵哈林 ,周瑞莲 ,张铜会 ,等 科尔沁沙地植被的统计学特征与土地沙漠化
[J].中国沙漠 ,2004 ,24 (3 ):20 -24 .
[本文引用: 1]
[18]
李玉霖 ,孟庆涛 ,赵学勇 ,等 科尔沁沙地流动沙丘植被恢复过程中群落组成及植物多样性演变特征
[J].草业学报 ,2007 ,16 (6 ):54 -61 .
[本文引用: 1]
[19]
赵珍珍 基于多源数据的科尔沁沙地生态环境变化研究
[D].武汉 :武汉大学 ,2017 .
[本文引用: 1]
[20]
李玉霖 ,崔建垣 ,苏永中 不同沙丘生境主要植物比叶面积和叶干物质含量的比较
[J].生态学报 ,2005 ,25 (2 ):304 -311 .
[本文引用: 1]
[21]
左小安 ,赵哈林 ,赵学勇 ,等 科尔沁沙地不同尺度上沙丘景观格局动态变化分析
[J].中国沙漠 ,2009 ,29 (5 ):785 -795 .
[本文引用: 1]
[22]
王少昆 ,赵学勇 ,张铜会 ,等 科尔沁沙地几种灌木对根际微生物的影响
[J].干旱区资源与环境 ,2012 ,26 (5 ):140 -144 .
[本文引用: 1]
[23]
宁志英 ,李玉霖 ,杨红玲 ,等 沙化草地土壤碳氮磷化学计量特征及其对植被生产力与多样性的影响
[J].生态学报 ,2019 ,39 (10 ):1 -9 .
[本文引用: 1]
[24]
李春萍 ,李刚 ,肖春旺 异速生长关系在陆地生态系统生物量估测中的应用
[J].世界科技研究与发展 ,2007 ,29 (2 ):51 -57 .
[本文引用: 1]
[25]
黄明泉 典型地段思茅松天然林生物量分配的比较分析及环境解释
[D].昆明 :西南林业大学 ,2018 .
[本文引用: 1]
[26]
Mokany K Raison R J Prokushkin A S Critical analysis of root:shoot ratios in terrestrial biomes
[J].Global Change Biology ,2005 ,12 (1 ):84 -96 .
[本文引用: 1]
[27]
Schmid I The influence of soil type and interspecific competition on the fine root system of Norway spruce and European beech
[J].Basic and Applied Ecology ,2002 ,3 (4 ):339 -346 .
[本文引用: 1]
[28]
蔡哲 ,刘琪璟 ,欧阳球林 千烟洲试验区几种灌木生物量估算模型的研究
[J].中南林学院学报 ,2006 ,26 (3 ):15 -18 ,23 .
[本文引用: 1]
[29]
Siebenkäs A Roscher C Functional composition rather than species richness determines root characteristics of experimental grasslands grown at different light and nutrient availability
[J].Plant & Soil ,2016 ,404 (1 ):1 -14 .
[本文引用: 1]
[30]
王敏 ,苏永中 ,杨荣 ,等 黑河中游荒漠草地地上和地下生物量的分配格局
[J].植物生态学报 ,2013 ,37 (3 ):209 -219 .
[本文引用: 1]
[31]
弋良朋 ,马健 ,李彦 盐胁迫对3种荒漠盐生植物苗期根系特征及活力的影响
[J].中国科学(D辑:地球科学) ,2006 ,36 ():86 -94 .
[本文引用: 1]
[32]
张冬燕 ,王冬至 ,范冬冬 ,等 不同立地类型华北落叶松人工林冠幅与胸径关系研究
[J].林业资源管理 ,2019 (4 ):69 -73 .
[本文引用: 1]
[33]
耿浩林 克氏针茅群落地上/地下生物量分配及其对水热因子响应研究
[D].北京 :中国科学院研究生院(植物研究所) ,2006 .
[本文引用: 1]
[34]
李春萍 ,李刚 ,肖春旺 异速生长关系在陆地生态系统生物量估测中的应用
[J].世界科技研究与发展 ,2007 ,29 (2 ):51 -57 .
[本文引用: 1]
[35]
陶冶 ,张元明 准噶尔荒漠6种类短命植物生物量分配与异速生长关系
[J].草业学报 ,2014 ,23 (2 ):38 -48 .
[本文引用: 1]
[36]
程栋梁 植物生物量分配模式与生长速率的相关规律研究
[D].兰州 :兰州大学 ,2007 .
[本文引用: 1]
荒漠草地4种灌木生物量分配特征
2
2013
... 生物量作为生态系统的能量基础和营养物质来源,反映植物对有机物质和能量的积累[1 -3 ] .生物量分配是植物生活史理论的中心.植物通过生物量分配将营养器官同化的资源用于根、茎、叶、花、果等器官的生长,植物对多样环境做出适时调整,是植物经过长期进化而形成的生存策略[4 ] .植物生物量分配受自身特性和环境条件的影响[5 ] ,在不同生境下具有不同的生长特性与物质分配规律[6 ] .当植物生长环境中光照或养分条件发生变化时,植物会通过调整生物量分配和形态特征来获取当下环境中的最大效率养分,从而提高存活率[7 ] .而在不同生长环境中,植物根、茎、叶等构件间存在竞争关系,通过内部自身权衡调配,在整个生活史中获取最理想且最适合自身发展的分配格局[8 ] .王宏信等[9 ] 发现,毛乌素沙地3种灌木整株生物量及其在不同器官分配特征与沙地生境有关,茎的木质化程度、叶构型、根系分布情况会影响各器官的异速生长,进而影响了生物量的分配特征.丁俊祥等[10 ] 发现,古尔班通古特沙漠6种荒漠草本植物的根冠比和叶片生物量分配比例的种间差异,不仅与物种自身的遗传特性有关,还受植株个体大小的影响.王艳莉等[11 ] 发现,不同生境砂蓝刺头(Echinops gmelini )形态结构及生物量特征具有明显差异,而构件生物量间相关生长关系的一致规律性及较稳定的繁殖生物量分配,有利于其适应异质生境并完成生长发育.牛存洋等[12 ] 对科尔沁沙地小叶锦鸡儿(Caragana microphylla )地上-地下生物量异速生长模型拟合结果表明其符合异速生长模型的幂函数. ...
... 生物量分配是植物对资源分配方式的反应,是通过光合和呼吸作用在不同器官间分配的结果[25 ] .其在植物体内各构件的分配不仅受外界环境变化的调控,还与植物自身生长特性相关[26 -27 ] .而灌木生物量作为灌木群落以及生态系统中十分重要的组成部分,是评价植被生产力的重要参数,在对灌木的生长和发育规律以及在生态系统中的地位和作用等研究方面具有十分重要意义[28 ] .本研究中科尔沁沙地3种优势固沙灌木的各构件生物量及其占比、R/S、NAB /AGB及NAB/OBA均存在显著性差异(P <0.05),表明在同一生境中,不同的灌木物种对资源的利用能力不同.生物量分配的最优分配理论认为植物通过调节生物量在各器官的分配来适应外界环境,以获取光照、水分和养分等受限资源的最大效益,以维持其生长速率的最大化[29 ] .本研究中科尔沁沙地3种优势固沙灌木生物量分配中黄柳的老枝生物量及其占比居于首位,NAB/OBA低于1;差不嘎蒿的地上生物量占比居于首位,地上新生生物量是老枝生物量的2.43倍;黄柳和差不嘎蒿地下生物量集中分布在浅土层,不存在深根系(根系长度>100 cm).这表明在科尔沁沙地这种降水少、年内降水不均、水资源短缺、土壤养分较为贫瘠的环境中,黄柳和差不嘎蒿利用该生境中的资源主要用于进行地上部分的生长,地下部分受限,在固沙后期随着灌草丛(以浅根系草本为主)的出现,与黄柳和差不嘎蒿间存在对土壤浅层水分与养分资源的竞争.即植被在固沙过程中只能利用降雨后浅层土壤截留的少量地表水,同时存在对同一空间水分和养分等环境资源的竞争[30 ] .在固沙初期植物对水分和养分的依赖较为强烈,植被倾向于把更多的资源分配于地下,且在植物生长发育过程中,与相邻植株存在光照、水分及养分竞争.而在养分和水分受限的条件下,R/S的增加有利于植物对养分和水分的吸收,可以增强植物的抗瘠和抗旱能力[31 ] .本研究中小叶锦鸡儿地下生物量及其占比和R/S均居于首位,地下生物量主要集中分布在根系长度>30 cm处,该层生物量占比达到61.61%,且深根系(根系长度>100 cm)层地上生物量占到总生物量的22.33%.这说明随着在固沙后期小叶锦鸡儿将通过光合和呼吸作用积累的物质主要用于地下生物量的积累,随着灌木的生长,根系向更深的土层伸展,占据了更多土壤空间[1 ] ,及时从埋深较深的地下水补给根系所需水分,使其在3种优势固沙植被中独占优势,较耐旱,耐贫瘠. ...
毛竹单株地上部分生物量相容性模型研究
1
2016
... 生物量作为生态系统的能量基础和营养物质来源,反映植物对有机物质和能量的积累[1 -3 ] .生物量分配是植物生活史理论的中心.植物通过生物量分配将营养器官同化的资源用于根、茎、叶、花、果等器官的生长,植物对多样环境做出适时调整,是植物经过长期进化而形成的生存策略[4 ] .植物生物量分配受自身特性和环境条件的影响[5 ] ,在不同生境下具有不同的生长特性与物质分配规律[6 ] .当植物生长环境中光照或养分条件发生变化时,植物会通过调整生物量分配和形态特征来获取当下环境中的最大效率养分,从而提高存活率[7 ] .而在不同生长环境中,植物根、茎、叶等构件间存在竞争关系,通过内部自身权衡调配,在整个生活史中获取最理想且最适合自身发展的分配格局[8 ] .王宏信等[9 ] 发现,毛乌素沙地3种灌木整株生物量及其在不同器官分配特征与沙地生境有关,茎的木质化程度、叶构型、根系分布情况会影响各器官的异速生长,进而影响了生物量的分配特征.丁俊祥等[10 ] 发现,古尔班通古特沙漠6种荒漠草本植物的根冠比和叶片生物量分配比例的种间差异,不仅与物种自身的遗传特性有关,还受植株个体大小的影响.王艳莉等[11 ] 发现,不同生境砂蓝刺头(Echinops gmelini )形态结构及生物量特征具有明显差异,而构件生物量间相关生长关系的一致规律性及较稳定的繁殖生物量分配,有利于其适应异质生境并完成生长发育.牛存洋等[12 ] 对科尔沁沙地小叶锦鸡儿(Caragana microphylla )地上-地下生物量异速生长模型拟合结果表明其符合异速生长模型的幂函数. ...
猴耳环人工林地上部分生物量估测及其分配特征研究
1
2018
... 生物量作为生态系统的能量基础和营养物质来源,反映植物对有机物质和能量的积累[1 -3 ] .生物量分配是植物生活史理论的中心.植物通过生物量分配将营养器官同化的资源用于根、茎、叶、花、果等器官的生长,植物对多样环境做出适时调整,是植物经过长期进化而形成的生存策略[4 ] .植物生物量分配受自身特性和环境条件的影响[5 ] ,在不同生境下具有不同的生长特性与物质分配规律[6 ] .当植物生长环境中光照或养分条件发生变化时,植物会通过调整生物量分配和形态特征来获取当下环境中的最大效率养分,从而提高存活率[7 ] .而在不同生长环境中,植物根、茎、叶等构件间存在竞争关系,通过内部自身权衡调配,在整个生活史中获取最理想且最适合自身发展的分配格局[8 ] .王宏信等[9 ] 发现,毛乌素沙地3种灌木整株生物量及其在不同器官分配特征与沙地生境有关,茎的木质化程度、叶构型、根系分布情况会影响各器官的异速生长,进而影响了生物量的分配特征.丁俊祥等[10 ] 发现,古尔班通古特沙漠6种荒漠草本植物的根冠比和叶片生物量分配比例的种间差异,不仅与物种自身的遗传特性有关,还受植株个体大小的影响.王艳莉等[11 ] 发现,不同生境砂蓝刺头(Echinops gmelini )形态结构及生物量特征具有明显差异,而构件生物量间相关生长关系的一致规律性及较稳定的繁殖生物量分配,有利于其适应异质生境并完成生长发育.牛存洋等[12 ] 对科尔沁沙地小叶锦鸡儿(Caragana microphylla )地上-地下生物量异速生长模型拟合结果表明其符合异速生长模型的幂函数. ...
The role of biomass allocation in the growth response of plants to different levels of light,CO2 , nutrients and water:a quantitative review
1
2000
... 生物量作为生态系统的能量基础和营养物质来源,反映植物对有机物质和能量的积累[1 -3 ] .生物量分配是植物生活史理论的中心.植物通过生物量分配将营养器官同化的资源用于根、茎、叶、花、果等器官的生长,植物对多样环境做出适时调整,是植物经过长期进化而形成的生存策略[4 ] .植物生物量分配受自身特性和环境条件的影响[5 ] ,在不同生境下具有不同的生长特性与物质分配规律[6 ] .当植物生长环境中光照或养分条件发生变化时,植物会通过调整生物量分配和形态特征来获取当下环境中的最大效率养分,从而提高存活率[7 ] .而在不同生长环境中,植物根、茎、叶等构件间存在竞争关系,通过内部自身权衡调配,在整个生活史中获取最理想且最适合自身发展的分配格局[8 ] .王宏信等[9 ] 发现,毛乌素沙地3种灌木整株生物量及其在不同器官分配特征与沙地生境有关,茎的木质化程度、叶构型、根系分布情况会影响各器官的异速生长,进而影响了生物量的分配特征.丁俊祥等[10 ] 发现,古尔班通古特沙漠6种荒漠草本植物的根冠比和叶片生物量分配比例的种间差异,不仅与物种自身的遗传特性有关,还受植株个体大小的影响.王艳莉等[11 ] 发现,不同生境砂蓝刺头(Echinops gmelini )形态结构及生物量特征具有明显差异,而构件生物量间相关生长关系的一致规律性及较稳定的繁殖生物量分配,有利于其适应异质生境并完成生长发育.牛存洋等[12 ] 对科尔沁沙地小叶锦鸡儿(Caragana microphylla )地上-地下生物量异速生长模型拟合结果表明其符合异速生长模型的幂函数. ...
柴达木盆地几种荒漠灌丛植被的生物量分配格局
1
2014
... 生物量作为生态系统的能量基础和营养物质来源,反映植物对有机物质和能量的积累[1 -3 ] .生物量分配是植物生活史理论的中心.植物通过生物量分配将营养器官同化的资源用于根、茎、叶、花、果等器官的生长,植物对多样环境做出适时调整,是植物经过长期进化而形成的生存策略[4 ] .植物生物量分配受自身特性和环境条件的影响[5 ] ,在不同生境下具有不同的生长特性与物质分配规律[6 ] .当植物生长环境中光照或养分条件发生变化时,植物会通过调整生物量分配和形态特征来获取当下环境中的最大效率养分,从而提高存活率[7 ] .而在不同生长环境中,植物根、茎、叶等构件间存在竞争关系,通过内部自身权衡调配,在整个生活史中获取最理想且最适合自身发展的分配格局[8 ] .王宏信等[9 ] 发现,毛乌素沙地3种灌木整株生物量及其在不同器官分配特征与沙地生境有关,茎的木质化程度、叶构型、根系分布情况会影响各器官的异速生长,进而影响了生物量的分配特征.丁俊祥等[10 ] 发现,古尔班通古特沙漠6种荒漠草本植物的根冠比和叶片生物量分配比例的种间差异,不仅与物种自身的遗传特性有关,还受植株个体大小的影响.王艳莉等[11 ] 发现,不同生境砂蓝刺头(Echinops gmelini )形态结构及生物量特征具有明显差异,而构件生物量间相关生长关系的一致规律性及较稳定的繁殖生物量分配,有利于其适应异质生境并完成生长发育.牛存洋等[12 ] 对科尔沁沙地小叶锦鸡儿(Caragana microphylla )地上-地下生物量异速生长模型拟合结果表明其符合异速生长模型的幂函数. ...
Plasticity of whole plant and leaf traits in Rubia peregrina in response to light, nutrient and water availability
1
2002
... 生物量作为生态系统的能量基础和营养物质来源,反映植物对有机物质和能量的积累[1 -3 ] .生物量分配是植物生活史理论的中心.植物通过生物量分配将营养器官同化的资源用于根、茎、叶、花、果等器官的生长,植物对多样环境做出适时调整,是植物经过长期进化而形成的生存策略[4 ] .植物生物量分配受自身特性和环境条件的影响[5 ] ,在不同生境下具有不同的生长特性与物质分配规律[6 ] .当植物生长环境中光照或养分条件发生变化时,植物会通过调整生物量分配和形态特征来获取当下环境中的最大效率养分,从而提高存活率[7 ] .而在不同生长环境中,植物根、茎、叶等构件间存在竞争关系,通过内部自身权衡调配,在整个生活史中获取最理想且最适合自身发展的分配格局[8 ] .王宏信等[9 ] 发现,毛乌素沙地3种灌木整株生物量及其在不同器官分配特征与沙地生境有关,茎的木质化程度、叶构型、根系分布情况会影响各器官的异速生长,进而影响了生物量的分配特征.丁俊祥等[10 ] 发现,古尔班通古特沙漠6种荒漠草本植物的根冠比和叶片生物量分配比例的种间差异,不仅与物种自身的遗传特性有关,还受植株个体大小的影响.王艳莉等[11 ] 发现,不同生境砂蓝刺头(Echinops gmelini )形态结构及生物量特征具有明显差异,而构件生物量间相关生长关系的一致规律性及较稳定的繁殖生物量分配,有利于其适应异质生境并完成生长发育.牛存洋等[12 ] 对科尔沁沙地小叶锦鸡儿(Caragana microphylla )地上-地下生物量异速生长模型拟合结果表明其符合异速生长模型的幂函数. ...
宁夏荒漠草原优势植物生长及生物量分配对放牧干扰的响应
1
2015
... 生物量作为生态系统的能量基础和营养物质来源,反映植物对有机物质和能量的积累[1 -3 ] .生物量分配是植物生活史理论的中心.植物通过生物量分配将营养器官同化的资源用于根、茎、叶、花、果等器官的生长,植物对多样环境做出适时调整,是植物经过长期进化而形成的生存策略[4 ] .植物生物量分配受自身特性和环境条件的影响[5 ] ,在不同生境下具有不同的生长特性与物质分配规律[6 ] .当植物生长环境中光照或养分条件发生变化时,植物会通过调整生物量分配和形态特征来获取当下环境中的最大效率养分,从而提高存活率[7 ] .而在不同生长环境中,植物根、茎、叶等构件间存在竞争关系,通过内部自身权衡调配,在整个生活史中获取最理想且最适合自身发展的分配格局[8 ] .王宏信等[9 ] 发现,毛乌素沙地3种灌木整株生物量及其在不同器官分配特征与沙地生境有关,茎的木质化程度、叶构型、根系分布情况会影响各器官的异速生长,进而影响了生物量的分配特征.丁俊祥等[10 ] 发现,古尔班通古特沙漠6种荒漠草本植物的根冠比和叶片生物量分配比例的种间差异,不仅与物种自身的遗传特性有关,还受植株个体大小的影响.王艳莉等[11 ] 发现,不同生境砂蓝刺头(Echinops gmelini )形态结构及生物量特征具有明显差异,而构件生物量间相关生长关系的一致规律性及较稳定的繁殖生物量分配,有利于其适应异质生境并完成生长发育.牛存洋等[12 ] 对科尔沁沙地小叶锦鸡儿(Caragana microphylla )地上-地下生物量异速生长模型拟合结果表明其符合异速生长模型的幂函数. ...
几种沙生灌木生物量及其在不同器官分配与土壤理化特征相关性
1
2017
... 生物量作为生态系统的能量基础和营养物质来源,反映植物对有机物质和能量的积累[1 -3 ] .生物量分配是植物生活史理论的中心.植物通过生物量分配将营养器官同化的资源用于根、茎、叶、花、果等器官的生长,植物对多样环境做出适时调整,是植物经过长期进化而形成的生存策略[4 ] .植物生物量分配受自身特性和环境条件的影响[5 ] ,在不同生境下具有不同的生长特性与物质分配规律[6 ] .当植物生长环境中光照或养分条件发生变化时,植物会通过调整生物量分配和形态特征来获取当下环境中的最大效率养分,从而提高存活率[7 ] .而在不同生长环境中,植物根、茎、叶等构件间存在竞争关系,通过内部自身权衡调配,在整个生活史中获取最理想且最适合自身发展的分配格局[8 ] .王宏信等[9 ] 发现,毛乌素沙地3种灌木整株生物量及其在不同器官分配特征与沙地生境有关,茎的木质化程度、叶构型、根系分布情况会影响各器官的异速生长,进而影响了生物量的分配特征.丁俊祥等[10 ] 发现,古尔班通古特沙漠6种荒漠草本植物的根冠比和叶片生物量分配比例的种间差异,不仅与物种自身的遗传特性有关,还受植株个体大小的影响.王艳莉等[11 ] 发现,不同生境砂蓝刺头(Echinops gmelini )形态结构及生物量特征具有明显差异,而构件生物量间相关生长关系的一致规律性及较稳定的繁殖生物量分配,有利于其适应异质生境并完成生长发育.牛存洋等[12 ] 对科尔沁沙地小叶锦鸡儿(Caragana microphylla )地上-地下生物量异速生长模型拟合结果表明其符合异速生长模型的幂函数. ...
古尔班通古特沙漠6种荒漠草本植物的生物量分配与相关生长关系
1
2016
... 生物量作为生态系统的能量基础和营养物质来源,反映植物对有机物质和能量的积累[1 -3 ] .生物量分配是植物生活史理论的中心.植物通过生物量分配将营养器官同化的资源用于根、茎、叶、花、果等器官的生长,植物对多样环境做出适时调整,是植物经过长期进化而形成的生存策略[4 ] .植物生物量分配受自身特性和环境条件的影响[5 ] ,在不同生境下具有不同的生长特性与物质分配规律[6 ] .当植物生长环境中光照或养分条件发生变化时,植物会通过调整生物量分配和形态特征来获取当下环境中的最大效率养分,从而提高存活率[7 ] .而在不同生长环境中,植物根、茎、叶等构件间存在竞争关系,通过内部自身权衡调配,在整个生活史中获取最理想且最适合自身发展的分配格局[8 ] .王宏信等[9 ] 发现,毛乌素沙地3种灌木整株生物量及其在不同器官分配特征与沙地生境有关,茎的木质化程度、叶构型、根系分布情况会影响各器官的异速生长,进而影响了生物量的分配特征.丁俊祥等[10 ] 发现,古尔班通古特沙漠6种荒漠草本植物的根冠比和叶片生物量分配比例的种间差异,不仅与物种自身的遗传特性有关,还受植株个体大小的影响.王艳莉等[11 ] 发现,不同生境砂蓝刺头(Echinops gmelini )形态结构及生物量特征具有明显差异,而构件生物量间相关生长关系的一致规律性及较稳定的繁殖生物量分配,有利于其适应异质生境并完成生长发育.牛存洋等[12 ] 对科尔沁沙地小叶锦鸡儿(Caragana microphylla )地上-地下生物量异速生长模型拟合结果表明其符合异速生长模型的幂函数. ...
不同生境下砂蓝刺头(Echinops gmelini )形态结构及生物量分配特征
1
2018
... 生物量作为生态系统的能量基础和营养物质来源,反映植物对有机物质和能量的积累[1 -3 ] .生物量分配是植物生活史理论的中心.植物通过生物量分配将营养器官同化的资源用于根、茎、叶、花、果等器官的生长,植物对多样环境做出适时调整,是植物经过长期进化而形成的生存策略[4 ] .植物生物量分配受自身特性和环境条件的影响[5 ] ,在不同生境下具有不同的生长特性与物质分配规律[6 ] .当植物生长环境中光照或养分条件发生变化时,植物会通过调整生物量分配和形态特征来获取当下环境中的最大效率养分,从而提高存活率[7 ] .而在不同生长环境中,植物根、茎、叶等构件间存在竞争关系,通过内部自身权衡调配,在整个生活史中获取最理想且最适合自身发展的分配格局[8 ] .王宏信等[9 ] 发现,毛乌素沙地3种灌木整株生物量及其在不同器官分配特征与沙地生境有关,茎的木质化程度、叶构型、根系分布情况会影响各器官的异速生长,进而影响了生物量的分配特征.丁俊祥等[10 ] 发现,古尔班通古特沙漠6种荒漠草本植物的根冠比和叶片生物量分配比例的种间差异,不仅与物种自身的遗传特性有关,还受植株个体大小的影响.王艳莉等[11 ] 发现,不同生境砂蓝刺头(Echinops gmelini )形态结构及生物量特征具有明显差异,而构件生物量间相关生长关系的一致规律性及较稳定的繁殖生物量分配,有利于其适应异质生境并完成生长发育.牛存洋等[12 ] 对科尔沁沙地小叶锦鸡儿(Caragana microphylla )地上-地下生物量异速生长模型拟合结果表明其符合异速生长模型的幂函数. ...
科尔沁沙地小叶锦鸡儿地上-地下生物量分配格局
2
2013
... 生物量作为生态系统的能量基础和营养物质来源,反映植物对有机物质和能量的积累[1 -3 ] .生物量分配是植物生活史理论的中心.植物通过生物量分配将营养器官同化的资源用于根、茎、叶、花、果等器官的生长,植物对多样环境做出适时调整,是植物经过长期进化而形成的生存策略[4 ] .植物生物量分配受自身特性和环境条件的影响[5 ] ,在不同生境下具有不同的生长特性与物质分配规律[6 ] .当植物生长环境中光照或养分条件发生变化时,植物会通过调整生物量分配和形态特征来获取当下环境中的最大效率养分,从而提高存活率[7 ] .而在不同生长环境中,植物根、茎、叶等构件间存在竞争关系,通过内部自身权衡调配,在整个生活史中获取最理想且最适合自身发展的分配格局[8 ] .王宏信等[9 ] 发现,毛乌素沙地3种灌木整株生物量及其在不同器官分配特征与沙地生境有关,茎的木质化程度、叶构型、根系分布情况会影响各器官的异速生长,进而影响了生物量的分配特征.丁俊祥等[10 ] 发现,古尔班通古特沙漠6种荒漠草本植物的根冠比和叶片生物量分配比例的种间差异,不仅与物种自身的遗传特性有关,还受植株个体大小的影响.王艳莉等[11 ] 发现,不同生境砂蓝刺头(Echinops gmelini )形态结构及生物量特征具有明显差异,而构件生物量间相关生长关系的一致规律性及较稳定的繁殖生物量分配,有利于其适应异质生境并完成生长发育.牛存洋等[12 ] 对科尔沁沙地小叶锦鸡儿(Caragana microphylla )地上-地下生物量异速生长模型拟合结果表明其符合异速生长模型的幂函数. ...
... 理论上获取生态系统生物量的方法有两种.其一直接测量法,即收获法所得数值虽较准确,但耗时费力,且对生态系统具有较大的破坏性;其二间接估测法,即通过异速生长关系来评估生物量,是目前较为普遍应用的方法[34 ] .本研究参照牛存洋等[12 ] 对科尔沁沙地小叶锦鸡儿地上-地下生物量异速生长模型进行拟合的研究方法,通过地上-地下生物量关系,得出科尔沁沙地3种优势固沙灌木典型的幂函数异速生长模型,即符合幂函数y =ax b 形式,且地上-地下生物量存在显著的异速生长关系,其对黄柳和小叶锦鸡儿拟合较好.这说明3种灌木生物量分配具有不均匀性,与植物地上、地下生物量的相关生长关系与物种种类、生活型无关[35 -36 ] 的结论不一致.这可能与3种灌木在适应科尔沁沙地气候过程中,不同的资源分配策略有关,即差不嘎蒿匍匐生长,存在不定根. ...
基于冠幅直径和植株高度的灌木地上生物量估测方法研究
1
2004
... 在干旱半干旱风沙区,固沙灌木凭其地上多分枝茎、近地面树冠及其地下根系分布较深的特性[13 ] ,面积超过总造林面积的70%[14 ] ,已成为生态建设和退化生态系统修复中防风固沙、保持水土和维持生态平衡的关键性植物.灌木生物量已成为灌木群落及生态系统研究的重要对象,不仅反映了灌木与其他植物的竞争能力,还反映出灌木对周边生态环境的适应程度[15 ] .科尔沁沙地由于其脆弱的生态环境及强烈的人类活动,已成为中国北方农牧交错带沙漠化最严重的地区,在过去的几十年中,约80%的土地发生了不同程度的沙漠化[16 ] ,建植防风固沙灌木林是有效遏制该地区土地沙漠化和生态治理的重要措施[17 ] .黄柳(Salix gordejevii )、差不嘎蒿(Artemisia halodendron )和小叶锦鸡儿是科尔沁沙地固沙灌木林的主要建群种[18 ] .因此,以该地3种优势固沙灌木为研究对象,在个体水平上探讨3种固沙灌木生物量分配特征,不仅有助于了解沙地灌木对环境变化的响应机制和适应策略,还为沙地生境变化过程中植物种群变化、植被演替及植被管理提供科学依据. ...
我国的固沙型灌木林及其研究进展
1
1998
... 在干旱半干旱风沙区,固沙灌木凭其地上多分枝茎、近地面树冠及其地下根系分布较深的特性[13 ] ,面积超过总造林面积的70%[14 ] ,已成为生态建设和退化生态系统修复中防风固沙、保持水土和维持生态平衡的关键性植物.灌木生物量已成为灌木群落及生态系统研究的重要对象,不仅反映了灌木与其他植物的竞争能力,还反映出灌木对周边生态环境的适应程度[15 ] .科尔沁沙地由于其脆弱的生态环境及强烈的人类活动,已成为中国北方农牧交错带沙漠化最严重的地区,在过去的几十年中,约80%的土地发生了不同程度的沙漠化[16 ] ,建植防风固沙灌木林是有效遏制该地区土地沙漠化和生态治理的重要措施[17 ] .黄柳(Salix gordejevii )、差不嘎蒿(Artemisia halodendron )和小叶锦鸡儿是科尔沁沙地固沙灌木林的主要建群种[18 ] .因此,以该地3种优势固沙灌木为研究对象,在个体水平上探讨3种固沙灌木生物量分配特征,不仅有助于了解沙地灌木对环境变化的响应机制和适应策略,还为沙地生境变化过程中植物种群变化、植被演替及植被管理提供科学依据. ...
科尔沁沙地优势固沙灌木的生物量预测模型
2
2018
... 在干旱半干旱风沙区,固沙灌木凭其地上多分枝茎、近地面树冠及其地下根系分布较深的特性[13 ] ,面积超过总造林面积的70%[14 ] ,已成为生态建设和退化生态系统修复中防风固沙、保持水土和维持生态平衡的关键性植物.灌木生物量已成为灌木群落及生态系统研究的重要对象,不仅反映了灌木与其他植物的竞争能力,还反映出灌木对周边生态环境的适应程度[15 ] .科尔沁沙地由于其脆弱的生态环境及强烈的人类活动,已成为中国北方农牧交错带沙漠化最严重的地区,在过去的几十年中,约80%的土地发生了不同程度的沙漠化[16 ] ,建植防风固沙灌木林是有效遏制该地区土地沙漠化和生态治理的重要措施[17 ] .黄柳(Salix gordejevii )、差不嘎蒿(Artemisia halodendron )和小叶锦鸡儿是科尔沁沙地固沙灌木林的主要建群种[18 ] .因此,以该地3种优势固沙灌木为研究对象,在个体水平上探讨3种固沙灌木生物量分配特征,不仅有助于了解沙地灌木对环境变化的响应机制和适应策略,还为沙地生境变化过程中植物种群变化、植被演替及植被管理提供科学依据. ...
... 式中:y 为生物量;xj 为反映灌木形态特征变量;β 为参数;ε 为误差项,采用非线性方程法.科尔沁沙地优势固沙灌木生物量模型显示幂函数回归模型较为理想[15 ] ,本研究通过地上-地下生物量进行y =ax b 异速生长模型的幂函数建立,a 、b 为模型参数,y 为地上生物量,x 为地下生物量,对原始数据拟合得到的异速生长关系经过统计学检验得到拟合评定参数R 2 ,其可以直接反映地上-地下生物量相关程度[24 ] ,并通过显著性检验和F测验对模型进行校验. ...
Temporal progress in improving carbon and nitrogen storage by grazing exclosure practice in a degraded land area of China’s Horqin Sandy Grassland
1
2012
... 在干旱半干旱风沙区,固沙灌木凭其地上多分枝茎、近地面树冠及其地下根系分布较深的特性[13 ] ,面积超过总造林面积的70%[14 ] ,已成为生态建设和退化生态系统修复中防风固沙、保持水土和维持生态平衡的关键性植物.灌木生物量已成为灌木群落及生态系统研究的重要对象,不仅反映了灌木与其他植物的竞争能力,还反映出灌木对周边生态环境的适应程度[15 ] .科尔沁沙地由于其脆弱的生态环境及强烈的人类活动,已成为中国北方农牧交错带沙漠化最严重的地区,在过去的几十年中,约80%的土地发生了不同程度的沙漠化[16 ] ,建植防风固沙灌木林是有效遏制该地区土地沙漠化和生态治理的重要措施[17 ] .黄柳(Salix gordejevii )、差不嘎蒿(Artemisia halodendron )和小叶锦鸡儿是科尔沁沙地固沙灌木林的主要建群种[18 ] .因此,以该地3种优势固沙灌木为研究对象,在个体水平上探讨3种固沙灌木生物量分配特征,不仅有助于了解沙地灌木对环境变化的响应机制和适应策略,还为沙地生境变化过程中植物种群变化、植被演替及植被管理提供科学依据. ...
科尔沁沙地植被的统计学特征与土地沙漠化
1
2004
... 在干旱半干旱风沙区,固沙灌木凭其地上多分枝茎、近地面树冠及其地下根系分布较深的特性[13 ] ,面积超过总造林面积的70%[14 ] ,已成为生态建设和退化生态系统修复中防风固沙、保持水土和维持生态平衡的关键性植物.灌木生物量已成为灌木群落及生态系统研究的重要对象,不仅反映了灌木与其他植物的竞争能力,还反映出灌木对周边生态环境的适应程度[15 ] .科尔沁沙地由于其脆弱的生态环境及强烈的人类活动,已成为中国北方农牧交错带沙漠化最严重的地区,在过去的几十年中,约80%的土地发生了不同程度的沙漠化[16 ] ,建植防风固沙灌木林是有效遏制该地区土地沙漠化和生态治理的重要措施[17 ] .黄柳(Salix gordejevii )、差不嘎蒿(Artemisia halodendron )和小叶锦鸡儿是科尔沁沙地固沙灌木林的主要建群种[18 ] .因此,以该地3种优势固沙灌木为研究对象,在个体水平上探讨3种固沙灌木生物量分配特征,不仅有助于了解沙地灌木对环境变化的响应机制和适应策略,还为沙地生境变化过程中植物种群变化、植被演替及植被管理提供科学依据. ...
科尔沁沙地流动沙丘植被恢复过程中群落组成及植物多样性演变特征
1
2007
... 在干旱半干旱风沙区,固沙灌木凭其地上多分枝茎、近地面树冠及其地下根系分布较深的特性[13 ] ,面积超过总造林面积的70%[14 ] ,已成为生态建设和退化生态系统修复中防风固沙、保持水土和维持生态平衡的关键性植物.灌木生物量已成为灌木群落及生态系统研究的重要对象,不仅反映了灌木与其他植物的竞争能力,还反映出灌木对周边生态环境的适应程度[15 ] .科尔沁沙地由于其脆弱的生态环境及强烈的人类活动,已成为中国北方农牧交错带沙漠化最严重的地区,在过去的几十年中,约80%的土地发生了不同程度的沙漠化[16 ] ,建植防风固沙灌木林是有效遏制该地区土地沙漠化和生态治理的重要措施[17 ] .黄柳(Salix gordejevii )、差不嘎蒿(Artemisia halodendron )和小叶锦鸡儿是科尔沁沙地固沙灌木林的主要建群种[18 ] .因此,以该地3种优势固沙灌木为研究对象,在个体水平上探讨3种固沙灌木生物量分配特征,不仅有助于了解沙地灌木对环境变化的响应机制和适应策略,还为沙地生境变化过程中植物种群变化、植被演替及植被管理提供科学依据. ...
基于多源数据的科尔沁沙地生态环境变化研究
1
2017
... 科尔沁沙地处于中国东北部西辽河冲积-湖积平原地区,是内蒙古高原向东北平原的过渡地带,与浑善达克沙地、毛乌素沙地和呼伦贝尔沙地被称为中国四大沙地.科尔沁沙地总面积约5.06万km2 ,地理位置42.30°—45.30°N、119°—124°E,平均海拔178.5 m,属于温带大陆性半干旱气候[19 ] .该地降水少,年际降水波动性大,年内降水不均,年降水量364.6 mm,集中在6—8月,年蒸发量1 972.8 mm.水资源短缺,总体呈现西部多、东部少的特点.年日照时数2 900~3 100 h,年平均气温5.8~6.4 ℃,≥10 ℃年积温3 000 ℃以上,无霜期约150 d.冬季多西北风,春季多西南风,年均风速3.6~4.1 m·s-1[20 -21 ] .自第四世纪以来科尔沁沙地沉积了深厚松散的中沙和细沙,为土地沙化提供了物质基础,土壤类型有风沙土、草甸土、沙质栗钙土和沼泽土[22 ] ,以风沙土为主,土壤的机械组成中粒径1~0.25 mm沙粒占20%~58%,粒径0.25~0.05 mm沙粒占40%~67%,粒径<0.05 mm的粉粒和黏粒占0.1%~15%,土壤有机质含量0.08%~0.49%.形成缓起伏沙地、流动沙丘、半固定沙丘、固定沙丘与丘间低地镶嵌分布的现代风沙地貌景观.植被为典型的沙地疏林草原植被,主要有盐生和沙生植物,例如小叶锦鸡儿、狗尾草(Setaria viridis )、黄柳、芦苇(Phragmites communis )、糙隐子草(Cleistogenes squarrosa )、差不嘎蒿、黄蒿(Artemisia scoparia )、达乌里胡枝子(Lespedeza davurica )和白草(Pennisetum centrasiaticum )[23 ] . ...
不同沙丘生境主要植物比叶面积和叶干物质含量的比较
1
2005
... 科尔沁沙地处于中国东北部西辽河冲积-湖积平原地区,是内蒙古高原向东北平原的过渡地带,与浑善达克沙地、毛乌素沙地和呼伦贝尔沙地被称为中国四大沙地.科尔沁沙地总面积约5.06万km2 ,地理位置42.30°—45.30°N、119°—124°E,平均海拔178.5 m,属于温带大陆性半干旱气候[19 ] .该地降水少,年际降水波动性大,年内降水不均,年降水量364.6 mm,集中在6—8月,年蒸发量1 972.8 mm.水资源短缺,总体呈现西部多、东部少的特点.年日照时数2 900~3 100 h,年平均气温5.8~6.4 ℃,≥10 ℃年积温3 000 ℃以上,无霜期约150 d.冬季多西北风,春季多西南风,年均风速3.6~4.1 m·s-1[20 -21 ] .自第四世纪以来科尔沁沙地沉积了深厚松散的中沙和细沙,为土地沙化提供了物质基础,土壤类型有风沙土、草甸土、沙质栗钙土和沼泽土[22 ] ,以风沙土为主,土壤的机械组成中粒径1~0.25 mm沙粒占20%~58%,粒径0.25~0.05 mm沙粒占40%~67%,粒径<0.05 mm的粉粒和黏粒占0.1%~15%,土壤有机质含量0.08%~0.49%.形成缓起伏沙地、流动沙丘、半固定沙丘、固定沙丘与丘间低地镶嵌分布的现代风沙地貌景观.植被为典型的沙地疏林草原植被,主要有盐生和沙生植物,例如小叶锦鸡儿、狗尾草(Setaria viridis )、黄柳、芦苇(Phragmites communis )、糙隐子草(Cleistogenes squarrosa )、差不嘎蒿、黄蒿(Artemisia scoparia )、达乌里胡枝子(Lespedeza davurica )和白草(Pennisetum centrasiaticum )[23 ] . ...
科尔沁沙地不同尺度上沙丘景观格局动态变化分析
1
2009
... 科尔沁沙地处于中国东北部西辽河冲积-湖积平原地区,是内蒙古高原向东北平原的过渡地带,与浑善达克沙地、毛乌素沙地和呼伦贝尔沙地被称为中国四大沙地.科尔沁沙地总面积约5.06万km2 ,地理位置42.30°—45.30°N、119°—124°E,平均海拔178.5 m,属于温带大陆性半干旱气候[19 ] .该地降水少,年际降水波动性大,年内降水不均,年降水量364.6 mm,集中在6—8月,年蒸发量1 972.8 mm.水资源短缺,总体呈现西部多、东部少的特点.年日照时数2 900~3 100 h,年平均气温5.8~6.4 ℃,≥10 ℃年积温3 000 ℃以上,无霜期约150 d.冬季多西北风,春季多西南风,年均风速3.6~4.1 m·s-1[20 -21 ] .自第四世纪以来科尔沁沙地沉积了深厚松散的中沙和细沙,为土地沙化提供了物质基础,土壤类型有风沙土、草甸土、沙质栗钙土和沼泽土[22 ] ,以风沙土为主,土壤的机械组成中粒径1~0.25 mm沙粒占20%~58%,粒径0.25~0.05 mm沙粒占40%~67%,粒径<0.05 mm的粉粒和黏粒占0.1%~15%,土壤有机质含量0.08%~0.49%.形成缓起伏沙地、流动沙丘、半固定沙丘、固定沙丘与丘间低地镶嵌分布的现代风沙地貌景观.植被为典型的沙地疏林草原植被,主要有盐生和沙生植物,例如小叶锦鸡儿、狗尾草(Setaria viridis )、黄柳、芦苇(Phragmites communis )、糙隐子草(Cleistogenes squarrosa )、差不嘎蒿、黄蒿(Artemisia scoparia )、达乌里胡枝子(Lespedeza davurica )和白草(Pennisetum centrasiaticum )[23 ] . ...
科尔沁沙地几种灌木对根际微生物的影响
1
2012
... 科尔沁沙地处于中国东北部西辽河冲积-湖积平原地区,是内蒙古高原向东北平原的过渡地带,与浑善达克沙地、毛乌素沙地和呼伦贝尔沙地被称为中国四大沙地.科尔沁沙地总面积约5.06万km2 ,地理位置42.30°—45.30°N、119°—124°E,平均海拔178.5 m,属于温带大陆性半干旱气候[19 ] .该地降水少,年际降水波动性大,年内降水不均,年降水量364.6 mm,集中在6—8月,年蒸发量1 972.8 mm.水资源短缺,总体呈现西部多、东部少的特点.年日照时数2 900~3 100 h,年平均气温5.8~6.4 ℃,≥10 ℃年积温3 000 ℃以上,无霜期约150 d.冬季多西北风,春季多西南风,年均风速3.6~4.1 m·s-1[20 -21 ] .自第四世纪以来科尔沁沙地沉积了深厚松散的中沙和细沙,为土地沙化提供了物质基础,土壤类型有风沙土、草甸土、沙质栗钙土和沼泽土[22 ] ,以风沙土为主,土壤的机械组成中粒径1~0.25 mm沙粒占20%~58%,粒径0.25~0.05 mm沙粒占40%~67%,粒径<0.05 mm的粉粒和黏粒占0.1%~15%,土壤有机质含量0.08%~0.49%.形成缓起伏沙地、流动沙丘、半固定沙丘、固定沙丘与丘间低地镶嵌分布的现代风沙地貌景观.植被为典型的沙地疏林草原植被,主要有盐生和沙生植物,例如小叶锦鸡儿、狗尾草(Setaria viridis )、黄柳、芦苇(Phragmites communis )、糙隐子草(Cleistogenes squarrosa )、差不嘎蒿、黄蒿(Artemisia scoparia )、达乌里胡枝子(Lespedeza davurica )和白草(Pennisetum centrasiaticum )[23 ] . ...
沙化草地土壤碳氮磷化学计量特征及其对植被生产力与多样性的影响
1
2019
... 科尔沁沙地处于中国东北部西辽河冲积-湖积平原地区,是内蒙古高原向东北平原的过渡地带,与浑善达克沙地、毛乌素沙地和呼伦贝尔沙地被称为中国四大沙地.科尔沁沙地总面积约5.06万km2 ,地理位置42.30°—45.30°N、119°—124°E,平均海拔178.5 m,属于温带大陆性半干旱气候[19 ] .该地降水少,年际降水波动性大,年内降水不均,年降水量364.6 mm,集中在6—8月,年蒸发量1 972.8 mm.水资源短缺,总体呈现西部多、东部少的特点.年日照时数2 900~3 100 h,年平均气温5.8~6.4 ℃,≥10 ℃年积温3 000 ℃以上,无霜期约150 d.冬季多西北风,春季多西南风,年均风速3.6~4.1 m·s-1[20 -21 ] .自第四世纪以来科尔沁沙地沉积了深厚松散的中沙和细沙,为土地沙化提供了物质基础,土壤类型有风沙土、草甸土、沙质栗钙土和沼泽土[22 ] ,以风沙土为主,土壤的机械组成中粒径1~0.25 mm沙粒占20%~58%,粒径0.25~0.05 mm沙粒占40%~67%,粒径<0.05 mm的粉粒和黏粒占0.1%~15%,土壤有机质含量0.08%~0.49%.形成缓起伏沙地、流动沙丘、半固定沙丘、固定沙丘与丘间低地镶嵌分布的现代风沙地貌景观.植被为典型的沙地疏林草原植被,主要有盐生和沙生植物,例如小叶锦鸡儿、狗尾草(Setaria viridis )、黄柳、芦苇(Phragmites communis )、糙隐子草(Cleistogenes squarrosa )、差不嘎蒿、黄蒿(Artemisia scoparia )、达乌里胡枝子(Lespedeza davurica )和白草(Pennisetum centrasiaticum )[23 ] . ...
异速生长关系在陆地生态系统生物量估测中的应用
1
2007
... 式中:y 为生物量;xj 为反映灌木形态特征变量;β 为参数;ε 为误差项,采用非线性方程法.科尔沁沙地优势固沙灌木生物量模型显示幂函数回归模型较为理想[15 ] ,本研究通过地上-地下生物量进行y =ax b 异速生长模型的幂函数建立,a 、b 为模型参数,y 为地上生物量,x 为地下生物量,对原始数据拟合得到的异速生长关系经过统计学检验得到拟合评定参数R 2 ,其可以直接反映地上-地下生物量相关程度[24 ] ,并通过显著性检验和F测验对模型进行校验. ...
典型地段思茅松天然林生物量分配的比较分析及环境解释
1
2018
... 生物量分配是植物对资源分配方式的反应,是通过光合和呼吸作用在不同器官间分配的结果[25 ] .其在植物体内各构件的分配不仅受外界环境变化的调控,还与植物自身生长特性相关[26 -27 ] .而灌木生物量作为灌木群落以及生态系统中十分重要的组成部分,是评价植被生产力的重要参数,在对灌木的生长和发育规律以及在生态系统中的地位和作用等研究方面具有十分重要意义[28 ] .本研究中科尔沁沙地3种优势固沙灌木的各构件生物量及其占比、R/S、NAB /AGB及NAB/OBA均存在显著性差异(P <0.05),表明在同一生境中,不同的灌木物种对资源的利用能力不同.生物量分配的最优分配理论认为植物通过调节生物量在各器官的分配来适应外界环境,以获取光照、水分和养分等受限资源的最大效益,以维持其生长速率的最大化[29 ] .本研究中科尔沁沙地3种优势固沙灌木生物量分配中黄柳的老枝生物量及其占比居于首位,NAB/OBA低于1;差不嘎蒿的地上生物量占比居于首位,地上新生生物量是老枝生物量的2.43倍;黄柳和差不嘎蒿地下生物量集中分布在浅土层,不存在深根系(根系长度>100 cm).这表明在科尔沁沙地这种降水少、年内降水不均、水资源短缺、土壤养分较为贫瘠的环境中,黄柳和差不嘎蒿利用该生境中的资源主要用于进行地上部分的生长,地下部分受限,在固沙后期随着灌草丛(以浅根系草本为主)的出现,与黄柳和差不嘎蒿间存在对土壤浅层水分与养分资源的竞争.即植被在固沙过程中只能利用降雨后浅层土壤截留的少量地表水,同时存在对同一空间水分和养分等环境资源的竞争[30 ] .在固沙初期植物对水分和养分的依赖较为强烈,植被倾向于把更多的资源分配于地下,且在植物生长发育过程中,与相邻植株存在光照、水分及养分竞争.而在养分和水分受限的条件下,R/S的增加有利于植物对养分和水分的吸收,可以增强植物的抗瘠和抗旱能力[31 ] .本研究中小叶锦鸡儿地下生物量及其占比和R/S均居于首位,地下生物量主要集中分布在根系长度>30 cm处,该层生物量占比达到61.61%,且深根系(根系长度>100 cm)层地上生物量占到总生物量的22.33%.这说明随着在固沙后期小叶锦鸡儿将通过光合和呼吸作用积累的物质主要用于地下生物量的积累,随着灌木的生长,根系向更深的土层伸展,占据了更多土壤空间[1 ] ,及时从埋深较深的地下水补给根系所需水分,使其在3种优势固沙植被中独占优势,较耐旱,耐贫瘠. ...
Critical analysis of root:shoot ratios in terrestrial biomes
1
2005
... 生物量分配是植物对资源分配方式的反应,是通过光合和呼吸作用在不同器官间分配的结果[25 ] .其在植物体内各构件的分配不仅受外界环境变化的调控,还与植物自身生长特性相关[26 -27 ] .而灌木生物量作为灌木群落以及生态系统中十分重要的组成部分,是评价植被生产力的重要参数,在对灌木的生长和发育规律以及在生态系统中的地位和作用等研究方面具有十分重要意义[28 ] .本研究中科尔沁沙地3种优势固沙灌木的各构件生物量及其占比、R/S、NAB /AGB及NAB/OBA均存在显著性差异(P <0.05),表明在同一生境中,不同的灌木物种对资源的利用能力不同.生物量分配的最优分配理论认为植物通过调节生物量在各器官的分配来适应外界环境,以获取光照、水分和养分等受限资源的最大效益,以维持其生长速率的最大化[29 ] .本研究中科尔沁沙地3种优势固沙灌木生物量分配中黄柳的老枝生物量及其占比居于首位,NAB/OBA低于1;差不嘎蒿的地上生物量占比居于首位,地上新生生物量是老枝生物量的2.43倍;黄柳和差不嘎蒿地下生物量集中分布在浅土层,不存在深根系(根系长度>100 cm).这表明在科尔沁沙地这种降水少、年内降水不均、水资源短缺、土壤养分较为贫瘠的环境中,黄柳和差不嘎蒿利用该生境中的资源主要用于进行地上部分的生长,地下部分受限,在固沙后期随着灌草丛(以浅根系草本为主)的出现,与黄柳和差不嘎蒿间存在对土壤浅层水分与养分资源的竞争.即植被在固沙过程中只能利用降雨后浅层土壤截留的少量地表水,同时存在对同一空间水分和养分等环境资源的竞争[30 ] .在固沙初期植物对水分和养分的依赖较为强烈,植被倾向于把更多的资源分配于地下,且在植物生长发育过程中,与相邻植株存在光照、水分及养分竞争.而在养分和水分受限的条件下,R/S的增加有利于植物对养分和水分的吸收,可以增强植物的抗瘠和抗旱能力[31 ] .本研究中小叶锦鸡儿地下生物量及其占比和R/S均居于首位,地下生物量主要集中分布在根系长度>30 cm处,该层生物量占比达到61.61%,且深根系(根系长度>100 cm)层地上生物量占到总生物量的22.33%.这说明随着在固沙后期小叶锦鸡儿将通过光合和呼吸作用积累的物质主要用于地下生物量的积累,随着灌木的生长,根系向更深的土层伸展,占据了更多土壤空间[1 ] ,及时从埋深较深的地下水补给根系所需水分,使其在3种优势固沙植被中独占优势,较耐旱,耐贫瘠. ...
The influence of soil type and interspecific competition on the fine root system of Norway spruce and European beech
1
2002
... 生物量分配是植物对资源分配方式的反应,是通过光合和呼吸作用在不同器官间分配的结果[25 ] .其在植物体内各构件的分配不仅受外界环境变化的调控,还与植物自身生长特性相关[26 -27 ] .而灌木生物量作为灌木群落以及生态系统中十分重要的组成部分,是评价植被生产力的重要参数,在对灌木的生长和发育规律以及在生态系统中的地位和作用等研究方面具有十分重要意义[28 ] .本研究中科尔沁沙地3种优势固沙灌木的各构件生物量及其占比、R/S、NAB /AGB及NAB/OBA均存在显著性差异(P <0.05),表明在同一生境中,不同的灌木物种对资源的利用能力不同.生物量分配的最优分配理论认为植物通过调节生物量在各器官的分配来适应外界环境,以获取光照、水分和养分等受限资源的最大效益,以维持其生长速率的最大化[29 ] .本研究中科尔沁沙地3种优势固沙灌木生物量分配中黄柳的老枝生物量及其占比居于首位,NAB/OBA低于1;差不嘎蒿的地上生物量占比居于首位,地上新生生物量是老枝生物量的2.43倍;黄柳和差不嘎蒿地下生物量集中分布在浅土层,不存在深根系(根系长度>100 cm).这表明在科尔沁沙地这种降水少、年内降水不均、水资源短缺、土壤养分较为贫瘠的环境中,黄柳和差不嘎蒿利用该生境中的资源主要用于进行地上部分的生长,地下部分受限,在固沙后期随着灌草丛(以浅根系草本为主)的出现,与黄柳和差不嘎蒿间存在对土壤浅层水分与养分资源的竞争.即植被在固沙过程中只能利用降雨后浅层土壤截留的少量地表水,同时存在对同一空间水分和养分等环境资源的竞争[30 ] .在固沙初期植物对水分和养分的依赖较为强烈,植被倾向于把更多的资源分配于地下,且在植物生长发育过程中,与相邻植株存在光照、水分及养分竞争.而在养分和水分受限的条件下,R/S的增加有利于植物对养分和水分的吸收,可以增强植物的抗瘠和抗旱能力[31 ] .本研究中小叶锦鸡儿地下生物量及其占比和R/S均居于首位,地下生物量主要集中分布在根系长度>30 cm处,该层生物量占比达到61.61%,且深根系(根系长度>100 cm)层地上生物量占到总生物量的22.33%.这说明随着在固沙后期小叶锦鸡儿将通过光合和呼吸作用积累的物质主要用于地下生物量的积累,随着灌木的生长,根系向更深的土层伸展,占据了更多土壤空间[1 ] ,及时从埋深较深的地下水补给根系所需水分,使其在3种优势固沙植被中独占优势,较耐旱,耐贫瘠. ...
千烟洲试验区几种灌木生物量估算模型的研究
1
2006
... 生物量分配是植物对资源分配方式的反应,是通过光合和呼吸作用在不同器官间分配的结果[25 ] .其在植物体内各构件的分配不仅受外界环境变化的调控,还与植物自身生长特性相关[26 -27 ] .而灌木生物量作为灌木群落以及生态系统中十分重要的组成部分,是评价植被生产力的重要参数,在对灌木的生长和发育规律以及在生态系统中的地位和作用等研究方面具有十分重要意义[28 ] .本研究中科尔沁沙地3种优势固沙灌木的各构件生物量及其占比、R/S、NAB /AGB及NAB/OBA均存在显著性差异(P <0.05),表明在同一生境中,不同的灌木物种对资源的利用能力不同.生物量分配的最优分配理论认为植物通过调节生物量在各器官的分配来适应外界环境,以获取光照、水分和养分等受限资源的最大效益,以维持其生长速率的最大化[29 ] .本研究中科尔沁沙地3种优势固沙灌木生物量分配中黄柳的老枝生物量及其占比居于首位,NAB/OBA低于1;差不嘎蒿的地上生物量占比居于首位,地上新生生物量是老枝生物量的2.43倍;黄柳和差不嘎蒿地下生物量集中分布在浅土层,不存在深根系(根系长度>100 cm).这表明在科尔沁沙地这种降水少、年内降水不均、水资源短缺、土壤养分较为贫瘠的环境中,黄柳和差不嘎蒿利用该生境中的资源主要用于进行地上部分的生长,地下部分受限,在固沙后期随着灌草丛(以浅根系草本为主)的出现,与黄柳和差不嘎蒿间存在对土壤浅层水分与养分资源的竞争.即植被在固沙过程中只能利用降雨后浅层土壤截留的少量地表水,同时存在对同一空间水分和养分等环境资源的竞争[30 ] .在固沙初期植物对水分和养分的依赖较为强烈,植被倾向于把更多的资源分配于地下,且在植物生长发育过程中,与相邻植株存在光照、水分及养分竞争.而在养分和水分受限的条件下,R/S的增加有利于植物对养分和水分的吸收,可以增强植物的抗瘠和抗旱能力[31 ] .本研究中小叶锦鸡儿地下生物量及其占比和R/S均居于首位,地下生物量主要集中分布在根系长度>30 cm处,该层生物量占比达到61.61%,且深根系(根系长度>100 cm)层地上生物量占到总生物量的22.33%.这说明随着在固沙后期小叶锦鸡儿将通过光合和呼吸作用积累的物质主要用于地下生物量的积累,随着灌木的生长,根系向更深的土层伸展,占据了更多土壤空间[1 ] ,及时从埋深较深的地下水补给根系所需水分,使其在3种优势固沙植被中独占优势,较耐旱,耐贫瘠. ...
Functional composition rather than species richness determines root characteristics of experimental grasslands grown at different light and nutrient availability
1
2016
... 生物量分配是植物对资源分配方式的反应,是通过光合和呼吸作用在不同器官间分配的结果[25 ] .其在植物体内各构件的分配不仅受外界环境变化的调控,还与植物自身生长特性相关[26 -27 ] .而灌木生物量作为灌木群落以及生态系统中十分重要的组成部分,是评价植被生产力的重要参数,在对灌木的生长和发育规律以及在生态系统中的地位和作用等研究方面具有十分重要意义[28 ] .本研究中科尔沁沙地3种优势固沙灌木的各构件生物量及其占比、R/S、NAB /AGB及NAB/OBA均存在显著性差异(P <0.05),表明在同一生境中,不同的灌木物种对资源的利用能力不同.生物量分配的最优分配理论认为植物通过调节生物量在各器官的分配来适应外界环境,以获取光照、水分和养分等受限资源的最大效益,以维持其生长速率的最大化[29 ] .本研究中科尔沁沙地3种优势固沙灌木生物量分配中黄柳的老枝生物量及其占比居于首位,NAB/OBA低于1;差不嘎蒿的地上生物量占比居于首位,地上新生生物量是老枝生物量的2.43倍;黄柳和差不嘎蒿地下生物量集中分布在浅土层,不存在深根系(根系长度>100 cm).这表明在科尔沁沙地这种降水少、年内降水不均、水资源短缺、土壤养分较为贫瘠的环境中,黄柳和差不嘎蒿利用该生境中的资源主要用于进行地上部分的生长,地下部分受限,在固沙后期随着灌草丛(以浅根系草本为主)的出现,与黄柳和差不嘎蒿间存在对土壤浅层水分与养分资源的竞争.即植被在固沙过程中只能利用降雨后浅层土壤截留的少量地表水,同时存在对同一空间水分和养分等环境资源的竞争[30 ] .在固沙初期植物对水分和养分的依赖较为强烈,植被倾向于把更多的资源分配于地下,且在植物生长发育过程中,与相邻植株存在光照、水分及养分竞争.而在养分和水分受限的条件下,R/S的增加有利于植物对养分和水分的吸收,可以增强植物的抗瘠和抗旱能力[31 ] .本研究中小叶锦鸡儿地下生物量及其占比和R/S均居于首位,地下生物量主要集中分布在根系长度>30 cm处,该层生物量占比达到61.61%,且深根系(根系长度>100 cm)层地上生物量占到总生物量的22.33%.这说明随着在固沙后期小叶锦鸡儿将通过光合和呼吸作用积累的物质主要用于地下生物量的积累,随着灌木的生长,根系向更深的土层伸展,占据了更多土壤空间[1 ] ,及时从埋深较深的地下水补给根系所需水分,使其在3种优势固沙植被中独占优势,较耐旱,耐贫瘠. ...
黑河中游荒漠草地地上和地下生物量的分配格局
1
2013
... 生物量分配是植物对资源分配方式的反应,是通过光合和呼吸作用在不同器官间分配的结果[25 ] .其在植物体内各构件的分配不仅受外界环境变化的调控,还与植物自身生长特性相关[26 -27 ] .而灌木生物量作为灌木群落以及生态系统中十分重要的组成部分,是评价植被生产力的重要参数,在对灌木的生长和发育规律以及在生态系统中的地位和作用等研究方面具有十分重要意义[28 ] .本研究中科尔沁沙地3种优势固沙灌木的各构件生物量及其占比、R/S、NAB /AGB及NAB/OBA均存在显著性差异(P <0.05),表明在同一生境中,不同的灌木物种对资源的利用能力不同.生物量分配的最优分配理论认为植物通过调节生物量在各器官的分配来适应外界环境,以获取光照、水分和养分等受限资源的最大效益,以维持其生长速率的最大化[29 ] .本研究中科尔沁沙地3种优势固沙灌木生物量分配中黄柳的老枝生物量及其占比居于首位,NAB/OBA低于1;差不嘎蒿的地上生物量占比居于首位,地上新生生物量是老枝生物量的2.43倍;黄柳和差不嘎蒿地下生物量集中分布在浅土层,不存在深根系(根系长度>100 cm).这表明在科尔沁沙地这种降水少、年内降水不均、水资源短缺、土壤养分较为贫瘠的环境中,黄柳和差不嘎蒿利用该生境中的资源主要用于进行地上部分的生长,地下部分受限,在固沙后期随着灌草丛(以浅根系草本为主)的出现,与黄柳和差不嘎蒿间存在对土壤浅层水分与养分资源的竞争.即植被在固沙过程中只能利用降雨后浅层土壤截留的少量地表水,同时存在对同一空间水分和养分等环境资源的竞争[30 ] .在固沙初期植物对水分和养分的依赖较为强烈,植被倾向于把更多的资源分配于地下,且在植物生长发育过程中,与相邻植株存在光照、水分及养分竞争.而在养分和水分受限的条件下,R/S的增加有利于植物对养分和水分的吸收,可以增强植物的抗瘠和抗旱能力[31 ] .本研究中小叶锦鸡儿地下生物量及其占比和R/S均居于首位,地下生物量主要集中分布在根系长度>30 cm处,该层生物量占比达到61.61%,且深根系(根系长度>100 cm)层地上生物量占到总生物量的22.33%.这说明随着在固沙后期小叶锦鸡儿将通过光合和呼吸作用积累的物质主要用于地下生物量的积累,随着灌木的生长,根系向更深的土层伸展,占据了更多土壤空间[1 ] ,及时从埋深较深的地下水补给根系所需水分,使其在3种优势固沙植被中独占优势,较耐旱,耐贫瘠. ...
盐胁迫对3种荒漠盐生植物苗期根系特征及活力的影响
1
2006
... 生物量分配是植物对资源分配方式的反应,是通过光合和呼吸作用在不同器官间分配的结果[25 ] .其在植物体内各构件的分配不仅受外界环境变化的调控,还与植物自身生长特性相关[26 -27 ] .而灌木生物量作为灌木群落以及生态系统中十分重要的组成部分,是评价植被生产力的重要参数,在对灌木的生长和发育规律以及在生态系统中的地位和作用等研究方面具有十分重要意义[28 ] .本研究中科尔沁沙地3种优势固沙灌木的各构件生物量及其占比、R/S、NAB /AGB及NAB/OBA均存在显著性差异(P <0.05),表明在同一生境中,不同的灌木物种对资源的利用能力不同.生物量分配的最优分配理论认为植物通过调节生物量在各器官的分配来适应外界环境,以获取光照、水分和养分等受限资源的最大效益,以维持其生长速率的最大化[29 ] .本研究中科尔沁沙地3种优势固沙灌木生物量分配中黄柳的老枝生物量及其占比居于首位,NAB/OBA低于1;差不嘎蒿的地上生物量占比居于首位,地上新生生物量是老枝生物量的2.43倍;黄柳和差不嘎蒿地下生物量集中分布在浅土层,不存在深根系(根系长度>100 cm).这表明在科尔沁沙地这种降水少、年内降水不均、水资源短缺、土壤养分较为贫瘠的环境中,黄柳和差不嘎蒿利用该生境中的资源主要用于进行地上部分的生长,地下部分受限,在固沙后期随着灌草丛(以浅根系草本为主)的出现,与黄柳和差不嘎蒿间存在对土壤浅层水分与养分资源的竞争.即植被在固沙过程中只能利用降雨后浅层土壤截留的少量地表水,同时存在对同一空间水分和养分等环境资源的竞争[30 ] .在固沙初期植物对水分和养分的依赖较为强烈,植被倾向于把更多的资源分配于地下,且在植物生长发育过程中,与相邻植株存在光照、水分及养分竞争.而在养分和水分受限的条件下,R/S的增加有利于植物对养分和水分的吸收,可以增强植物的抗瘠和抗旱能力[31 ] .本研究中小叶锦鸡儿地下生物量及其占比和R/S均居于首位,地下生物量主要集中分布在根系长度>30 cm处,该层生物量占比达到61.61%,且深根系(根系长度>100 cm)层地上生物量占到总生物量的22.33%.这说明随着在固沙后期小叶锦鸡儿将通过光合和呼吸作用积累的物质主要用于地下生物量的积累,随着灌木的生长,根系向更深的土层伸展,占据了更多土壤空间[1 ] ,及时从埋深较深的地下水补给根系所需水分,使其在3种优势固沙植被中独占优势,较耐旱,耐贫瘠. ...
不同立地类型华北落叶松人工林冠幅与胸径关系研究
1
2019
... 冠幅作为植被进行光合作用的主要场所,不仅反映立足竞争力和生长活力,还在经营效果、生产潜力预估和养分循环等研究中存在重要的意义[32 ] .黄柳和小叶锦鸡儿的R/S与冠幅极显著负相关(r =-0.453、r =-0.430,P <0.01),线性拟合结果呈现负线性关系(R 黄 柳 2 R 小 叶 锦 鸡 儿 2 r =-0.284,P <0.05),线性拟合结果呈现负线性关系(R 差 不 嘎 蒿 2 [33 ] .而在科尔沁沙地3种植被固沙后期,随着植被生长,冠幅增大,地上部分增加,地下生物量并未同步增大,反而到了一定程度增加变缓,此时地下部分生长受限.其中差不嘎蒿R/S与冠幅的关系并无黄柳和小叶锦鸡儿明显,这是因为差不嘎蒿匍匐生长,生长过程中茎衍生不定根,冠幅越大,不定根越多.黄柳和差不嘎蒿NAB/AGB和NAB/OBA均与冠幅呈现负线性关系(R 黄 柳 2 R 差 不 嘎 蒿 2 R 黄 柳 2 R 差 不 嘎 蒿 2 r =-0.389、r =-0.403,P <0.01).这说明随着植被生长冠幅的增大,新生生物量占比反而减小,地上新生部分减少,植被发生退化.在科尔沁沙地固沙后期,黄柳和差不嘎蒿随着自身生长冠幅的增大,同时受遗传和环境中水分、养分的影响,生产力在下降,植被衰退现象明显,而在小叶锦鸡儿中则不存在这种现象. ...
克氏针茅群落地上/地下生物量分配及其对水热因子响应研究
1
2006
... 冠幅作为植被进行光合作用的主要场所,不仅反映立足竞争力和生长活力,还在经营效果、生产潜力预估和养分循环等研究中存在重要的意义[32 ] .黄柳和小叶锦鸡儿的R/S与冠幅极显著负相关(r =-0.453、r =-0.430,P <0.01),线性拟合结果呈现负线性关系(R 黄 柳 2 R 小 叶 锦 鸡 儿 2 r =-0.284,P <0.05),线性拟合结果呈现负线性关系(R 差 不 嘎 蒿 2 [33 ] .而在科尔沁沙地3种植被固沙后期,随着植被生长,冠幅增大,地上部分增加,地下生物量并未同步增大,反而到了一定程度增加变缓,此时地下部分生长受限.其中差不嘎蒿R/S与冠幅的关系并无黄柳和小叶锦鸡儿明显,这是因为差不嘎蒿匍匐生长,生长过程中茎衍生不定根,冠幅越大,不定根越多.黄柳和差不嘎蒿NAB/AGB和NAB/OBA均与冠幅呈现负线性关系(R 黄 柳 2 R 差 不 嘎 蒿 2 R 黄 柳 2 R 差 不 嘎 蒿 2 r =-0.389、r =-0.403,P <0.01).这说明随着植被生长冠幅的增大,新生生物量占比反而减小,地上新生部分减少,植被发生退化.在科尔沁沙地固沙后期,黄柳和差不嘎蒿随着自身生长冠幅的增大,同时受遗传和环境中水分、养分的影响,生产力在下降,植被衰退现象明显,而在小叶锦鸡儿中则不存在这种现象. ...
异速生长关系在陆地生态系统生物量估测中的应用
1
2007
... 理论上获取生态系统生物量的方法有两种.其一直接测量法,即收获法所得数值虽较准确,但耗时费力,且对生态系统具有较大的破坏性;其二间接估测法,即通过异速生长关系来评估生物量,是目前较为普遍应用的方法[34 ] .本研究参照牛存洋等[12 ] 对科尔沁沙地小叶锦鸡儿地上-地下生物量异速生长模型进行拟合的研究方法,通过地上-地下生物量关系,得出科尔沁沙地3种优势固沙灌木典型的幂函数异速生长模型,即符合幂函数y =ax b 形式,且地上-地下生物量存在显著的异速生长关系,其对黄柳和小叶锦鸡儿拟合较好.这说明3种灌木生物量分配具有不均匀性,与植物地上、地下生物量的相关生长关系与物种种类、生活型无关[35 -36 ] 的结论不一致.这可能与3种灌木在适应科尔沁沙地气候过程中,不同的资源分配策略有关,即差不嘎蒿匍匐生长,存在不定根. ...
准噶尔荒漠6种类短命植物生物量分配与异速生长关系
1
2014
... 理论上获取生态系统生物量的方法有两种.其一直接测量法,即收获法所得数值虽较准确,但耗时费力,且对生态系统具有较大的破坏性;其二间接估测法,即通过异速生长关系来评估生物量,是目前较为普遍应用的方法[34 ] .本研究参照牛存洋等[12 ] 对科尔沁沙地小叶锦鸡儿地上-地下生物量异速生长模型进行拟合的研究方法,通过地上-地下生物量关系,得出科尔沁沙地3种优势固沙灌木典型的幂函数异速生长模型,即符合幂函数y =ax b 形式,且地上-地下生物量存在显著的异速生长关系,其对黄柳和小叶锦鸡儿拟合较好.这说明3种灌木生物量分配具有不均匀性,与植物地上、地下生物量的相关生长关系与物种种类、生活型无关[35 -36 ] 的结论不一致.这可能与3种灌木在适应科尔沁沙地气候过程中,不同的资源分配策略有关,即差不嘎蒿匍匐生长,存在不定根. ...
植物生物量分配模式与生长速率的相关规律研究
1
2007
... 理论上获取生态系统生物量的方法有两种.其一直接测量法,即收获法所得数值虽较准确,但耗时费力,且对生态系统具有较大的破坏性;其二间接估测法,即通过异速生长关系来评估生物量,是目前较为普遍应用的方法[34 ] .本研究参照牛存洋等[12 ] 对科尔沁沙地小叶锦鸡儿地上-地下生物量异速生长模型进行拟合的研究方法,通过地上-地下生物量关系,得出科尔沁沙地3种优势固沙灌木典型的幂函数异速生长模型,即符合幂函数y =ax b 形式,且地上-地下生物量存在显著的异速生长关系,其对黄柳和小叶锦鸡儿拟合较好.这说明3种灌木生物量分配具有不均匀性,与植物地上、地下生物量的相关生长关系与物种种类、生活型无关[35 -36 ] 的结论不一致.这可能与3种灌木在适应科尔沁沙地气候过程中,不同的资源分配策略有关,即差不嘎蒿匍匐生长,存在不定根. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}