Biological soil crusts as an organizing principle in drylands
2
2016
... 在干旱区荒漠地表,通常发育一层相对稳定的地表生物覆盖,被称为生物土壤结皮(以下简称生物结皮).生物结皮包括真菌、细菌、蓝细菌等微生物与藻类、地衣、苔藓等孢子植物类群,分布于土壤表层之上或表层几厘米土壤范围内[1 ] (图1 ).生物结皮盖度占全球陆地面积的12%,在干旱地表(Dry land)平均盖度达24%(从半湿润区13%到干旱区31%)[2 ] ,被称为地球的“活皮肤”,也被形象地称为“生物地毯”,在维持荒漠生态系统结构和功能稳定性,以及物质和能量交换等方面都具有非常重要的意义[1 ,3 -6 ] .生物结皮能够固定沙面,调节土壤水文,影响维管植物种子萌发、生长和物种多样性.同时,生物结皮含有藻类、苔藓等大量的初级生产者以及大量的固氮细菌和蓝藻,能够固定碳和氮,其固定量可分别占陆地生态系统净生产力的1%和生物固氮量的25%[7 -8 ] ,对荒漠土壤养分积累具有重要作用.因此,生物结皮也被称为干旱区地表生物覆被层的主要构建者. ...
... [1 ,3 -6 ].生物结皮能够固定沙面,调节土壤水文,影响维管植物种子萌发、生长和物种多样性.同时,生物结皮含有藻类、苔藓等大量的初级生产者以及大量的固氮细菌和蓝藻,能够固定碳和氮,其固定量可分别占陆地生态系统净生产力的1%和生物固氮量的25%[7 -8 ] ,对荒漠土壤养分积累具有重要作用.因此,生物结皮也被称为干旱区地表生物覆被层的主要构建者. ...
Dryland photoautotrophic soil surface communities endangered by global change
1
2018
... 在干旱区荒漠地表,通常发育一层相对稳定的地表生物覆盖,被称为生物土壤结皮(以下简称生物结皮).生物结皮包括真菌、细菌、蓝细菌等微生物与藻类、地衣、苔藓等孢子植物类群,分布于土壤表层之上或表层几厘米土壤范围内[1 ] (图1 ).生物结皮盖度占全球陆地面积的12%,在干旱地表(Dry land)平均盖度达24%(从半湿润区13%到干旱区31%)[2 ] ,被称为地球的“活皮肤”,也被形象地称为“生物地毯”,在维持荒漠生态系统结构和功能稳定性,以及物质和能量交换等方面都具有非常重要的意义[1 ,3 -6 ] .生物结皮能够固定沙面,调节土壤水文,影响维管植物种子萌发、生长和物种多样性.同时,生物结皮含有藻类、苔藓等大量的初级生产者以及大量的固氮细菌和蓝藻,能够固定碳和氮,其固定量可分别占陆地生态系统净生产力的1%和生物固氮量的25%[7 -8 ] ,对荒漠土壤养分积累具有重要作用.因此,生物结皮也被称为干旱区地表生物覆被层的主要构建者. ...
Biocrusts: the living skin of the earth
1
2018
... 在干旱区荒漠地表,通常发育一层相对稳定的地表生物覆盖,被称为生物土壤结皮(以下简称生物结皮).生物结皮包括真菌、细菌、蓝细菌等微生物与藻类、地衣、苔藓等孢子植物类群,分布于土壤表层之上或表层几厘米土壤范围内[1 ] (图1 ).生物结皮盖度占全球陆地面积的12%,在干旱地表(Dry land)平均盖度达24%(从半湿润区13%到干旱区31%)[2 ] ,被称为地球的“活皮肤”,也被形象地称为“生物地毯”,在维持荒漠生态系统结构和功能稳定性,以及物质和能量交换等方面都具有非常重要的意义[1 ,3 -6 ] .生物结皮能够固定沙面,调节土壤水文,影响维管植物种子萌发、生长和物种多样性.同时,生物结皮含有藻类、苔藓等大量的初级生产者以及大量的固氮细菌和蓝藻,能够固定碳和氮,其固定量可分别占陆地生态系统净生产力的1%和生物固氮量的25%[7 -8 ] ,对荒漠土壤养分积累具有重要作用.因此,生物结皮也被称为干旱区地表生物覆被层的主要构建者. ...
沙漠生物地毯工程:干旱沙漠治理的新途径
2
2005
... 中国是沙漠化发展最快的国家之一,造成生态系统失衡、耕地萎缩,严重地影响工农业生产和人民生活[4 ] .近年来,通过一系列生态工程的实施,荒漠化和沙化面积实现双缩减[55 ] .与“三北”防护林、京津源风沙源、退耕还林等工程相比,生物结皮的恢复工程规模相对较小.在降水量相对较少的干旱区,大规模植树造林的措施并不适用[4 ] .2001—2005年,中国科学院与内蒙古自治区在库布齐沙漠建立了生物结皮与乔灌草结合的示范区,成功治理了近200 hm2 流沙,并在其他区域进行大面积推广,该项目成为了生物结皮野外成功恢复的典范[9 ] .同时,该团队采用了工程化机械接种技术克服了人工生物结皮接种耗时费工的缺点,成为生物结皮工程化大面积应用的重要措施.除此之外,在其他区域,如腾格里沙漠、古尔班通古特沙漠、毛乌素沙地、黄土高原区等均有相应的生物结皮恢复应用的报道,但均处于方法探索和前期试验阶段[15 ,56 -58 ] (表1 ).国外生物结皮恢复研究虽然开展较早,但还未见能形成规模的生物结皮恢复示范区,中国生物结皮的恢复为世界生物结皮的恢复提供了重要借鉴. ...
... [4 ].2001—2005年,中国科学院与内蒙古自治区在库布齐沙漠建立了生物结皮与乔灌草结合的示范区,成功治理了近200 hm2 流沙,并在其他区域进行大面积推广,该项目成为了生物结皮野外成功恢复的典范[9 ] .同时,该团队采用了工程化机械接种技术克服了人工生物结皮接种耗时费工的缺点,成为生物结皮工程化大面积应用的重要措施.除此之外,在其他区域,如腾格里沙漠、古尔班通古特沙漠、毛乌素沙地、黄土高原区等均有相应的生物结皮恢复应用的报道,但均处于方法探索和前期试验阶段[15 ,56 -58 ] (表1 ).国外生物结皮恢复研究虽然开展较早,但还未见能形成规模的生物结皮恢复示范区,中国生物结皮的恢复为世界生物结皮的恢复提供了重要借鉴. ...
The microstructure of microbiotic crust and its influence on wind erosion for a sandy soil surface in the Gurbantunggut Desert of Northwestern China
1
2006
... 生物结皮的藻丝对土壤颗粒的缠绕以及分泌的聚合物对阳离子的吸收,使得生物结皮具有较强的抗风蚀和抗压能力.随着生物结皮生物量的增加,抗风蚀能力增强[31 ] .研究发现,未经扰动的生物结皮,难以形成地表风蚀,随着生物结皮破损率的增加,风蚀强度显著增加[5 ,32 ] .在生物结皮形成过程中,不同物种组成和不同生物结皮类型抗蚀度不同,如不同物种中具鞘微鞘藻和席藻具有较强的抗风蚀能力,不同生物结皮类型中苔藓结皮抗风蚀能力较强[32 -33 ] .生物结皮的存在,减少土壤细沙损失的同时,使得碳氮等养分能够被固定在土壤中[34 -35 ] . ...
Cementing mechanism of algal crusts from desert area
1
2002
... 在干旱区荒漠地表,通常发育一层相对稳定的地表生物覆盖,被称为生物土壤结皮(以下简称生物结皮).生物结皮包括真菌、细菌、蓝细菌等微生物与藻类、地衣、苔藓等孢子植物类群,分布于土壤表层之上或表层几厘米土壤范围内[1 ] (图1 ).生物结皮盖度占全球陆地面积的12%,在干旱地表(Dry land)平均盖度达24%(从半湿润区13%到干旱区31%)[2 ] ,被称为地球的“活皮肤”,也被形象地称为“生物地毯”,在维持荒漠生态系统结构和功能稳定性,以及物质和能量交换等方面都具有非常重要的意义[1 ,3 -6 ] .生物结皮能够固定沙面,调节土壤水文,影响维管植物种子萌发、生长和物种多样性.同时,生物结皮含有藻类、苔藓等大量的初级生产者以及大量的固氮细菌和蓝藻,能够固定碳和氮,其固定量可分别占陆地生态系统净生产力的1%和生物固氮量的25%[7 -8 ] ,对荒漠土壤养分积累具有重要作用.因此,生物结皮也被称为干旱区地表生物覆被层的主要构建者. ...
Patterns and controls on nitrogen cycling of biological soil crusts
2
2016
... 在干旱区荒漠地表,通常发育一层相对稳定的地表生物覆盖,被称为生物土壤结皮(以下简称生物结皮).生物结皮包括真菌、细菌、蓝细菌等微生物与藻类、地衣、苔藓等孢子植物类群,分布于土壤表层之上或表层几厘米土壤范围内[1 ] (图1 ).生物结皮盖度占全球陆地面积的12%,在干旱地表(Dry land)平均盖度达24%(从半湿润区13%到干旱区31%)[2 ] ,被称为地球的“活皮肤”,也被形象地称为“生物地毯”,在维持荒漠生态系统结构和功能稳定性,以及物质和能量交换等方面都具有非常重要的意义[1 ,3 -6 ] .生物结皮能够固定沙面,调节土壤水文,影响维管植物种子萌发、生长和物种多样性.同时,生物结皮含有藻类、苔藓等大量的初级生产者以及大量的固氮细菌和蓝藻,能够固定碳和氮,其固定量可分别占陆地生态系统净生产力的1%和生物固氮量的25%[7 -8 ] ,对荒漠土壤养分积累具有重要作用.因此,生物结皮也被称为干旱区地表生物覆被层的主要构建者. ...
... 生物结皮中的蓝藻、藓类等光合类群,能够固碳,促进有机质的形成和增加.全球尺度生物结皮年总固碳量可达0.6 Pg, 相当于全球陆地生态系统净生产力的1%,在荒漠区占比可达9%[8 ,23 ] .同时,生物结皮中具有大量的固氮细菌和蓝藻,全球年固氮量可达26 Tg,约占全球孢子植物固氮量的50%,以及陆地生态系统生物总固氮量的1/4[7 -8 ] .生物结皮中丰富的微生物可矿化和溶解土壤中不溶性无机磷和有机磷复合,促进磷的吸收[24 ] .因此,生物结皮斑块是干旱区特殊的肥岛,能够促进小尺度养分的聚集[25 -27 ] .除养分外,随着生物结皮的发育,生物结皮表面粗糙度增加,增加土壤水分入渗和留存[28 ] .与裸沙相比,生物结皮可降低水分蒸发,增加对大气中凝结水、蒸汽和雾等的收集,有利于干旱区生物结皮表面的水分供给和微生物的活动[29 -30 ] .总体来讲,生物结皮对养分和水分的聚集,有助于荒漠生态系统正向演替. ...
Contribution of cryptogamic covers to the global cycles of carbon and nitrogen
3
2012
... 在干旱区荒漠地表,通常发育一层相对稳定的地表生物覆盖,被称为生物土壤结皮(以下简称生物结皮).生物结皮包括真菌、细菌、蓝细菌等微生物与藻类、地衣、苔藓等孢子植物类群,分布于土壤表层之上或表层几厘米土壤范围内[1 ] (图1 ).生物结皮盖度占全球陆地面积的12%,在干旱地表(Dry land)平均盖度达24%(从半湿润区13%到干旱区31%)[2 ] ,被称为地球的“活皮肤”,也被形象地称为“生物地毯”,在维持荒漠生态系统结构和功能稳定性,以及物质和能量交换等方面都具有非常重要的意义[1 ,3 -6 ] .生物结皮能够固定沙面,调节土壤水文,影响维管植物种子萌发、生长和物种多样性.同时,生物结皮含有藻类、苔藓等大量的初级生产者以及大量的固氮细菌和蓝藻,能够固定碳和氮,其固定量可分别占陆地生态系统净生产力的1%和生物固氮量的25%[7 -8 ] ,对荒漠土壤养分积累具有重要作用.因此,生物结皮也被称为干旱区地表生物覆被层的主要构建者. ...
... 生物结皮中的蓝藻、藓类等光合类群,能够固碳,促进有机质的形成和增加.全球尺度生物结皮年总固碳量可达0.6 Pg, 相当于全球陆地生态系统净生产力的1%,在荒漠区占比可达9%[8 ,23 ] .同时,生物结皮中具有大量的固氮细菌和蓝藻,全球年固氮量可达26 Tg,约占全球孢子植物固氮量的50%,以及陆地生态系统生物总固氮量的1/4[7 -8 ] .生物结皮中丰富的微生物可矿化和溶解土壤中不溶性无机磷和有机磷复合,促进磷的吸收[24 ] .因此,生物结皮斑块是干旱区特殊的肥岛,能够促进小尺度养分的聚集[25 -27 ] .除养分外,随着生物结皮的发育,生物结皮表面粗糙度增加,增加土壤水分入渗和留存[28 ] .与裸沙相比,生物结皮可降低水分蒸发,增加对大气中凝结水、蒸汽和雾等的收集,有利于干旱区生物结皮表面的水分供给和微生物的活动[29 -30 ] .总体来讲,生物结皮对养分和水分的聚集,有助于荒漠生态系统正向演替. ...
... -8 ].生物结皮中丰富的微生物可矿化和溶解土壤中不溶性无机磷和有机磷复合,促进磷的吸收[24 ] .因此,生物结皮斑块是干旱区特殊的肥岛,能够促进小尺度养分的聚集[25 -27 ] .除养分外,随着生物结皮的发育,生物结皮表面粗糙度增加,增加土壤水分入渗和留存[28 ] .与裸沙相比,生物结皮可降低水分蒸发,增加对大气中凝结水、蒸汽和雾等的收集,有利于干旱区生物结皮表面的水分供给和微生物的活动[29 -30 ] .总体来讲,生物结皮对养分和水分的聚集,有助于荒漠生态系统正向演替. ...
11
2013
... 生物结皮从发育初期到成熟期,经历不同的演替阶段,形成不同类型的生物结皮,结构逐步趋于稳定.发育伊始,寡营养细菌可分泌胞外多糖粘结沙粒,形成生物结皮早期结构.随着藻类、地衣和苔藓植物的不断出现,胞外多糖粘结沙粒的作用逐渐被藻丝体、菌丝体以及苔藓假根捆绑沙粒的作用取代,生物结皮的结构更加复杂和稳定,构成生物结皮固沙的重要结构基础.同时,生物结皮中很多种类具有固氮和固碳功能,在改善土壤养分和水分条件以及抗风蚀方面发挥重要生态功能,在荒漠区固沙与受损地表系统的修复领域具有广阔的应用前景.近年来,由于干旱区传统生物固沙所面临的水资源缺乏和土壤水分消耗等问题,通过人工培育生物结皮进行固沙与修复荒漠受损地表生态,逐渐成为恢复生态学者关注的焦点.通过在裸露沙面或合理密度条件下的固沙植物间(或物理沙障间)接种生物结皮,既能达到防风固沙的目的,也能减少水分消耗来维持植物长期生存[9 ] .同时,在干旱区退化生态系统恢复方面,生物结皮可以快速增加土壤稳定性,抵抗水土侵蚀,恢复受损表面[10 -12 ] .中国在生物结皮固沙和退化生态系统恢复领域具有较丰厚的研究基础,并在一定范围内取得了成功的实践,如库布齐沙漠、腾格里沙漠和黄土高原[9 ,13 -14 ] .野外人工生物结皮接种和生长有望成为干旱区受损生态系统恢复的新模式,对干旱区传统防沙治沙和退化系统恢复模式形成重要补充. ...
... [9 ,13 -14 ].野外人工生物结皮接种和生长有望成为干旱区受损生态系统恢复的新模式,对干旱区传统防沙治沙和退化系统恢复模式形成重要补充. ...
... 生物结皮不同于物理和化学结皮(雨水或外力形成),是由活的生物体及其代谢产物,通过有机物和土壤颗粒黏结而成[9 ] (图2 ).生物结皮从形成初期到成熟期,经历了物种更替和不同演替阶段[15 ] .生物结皮的演替伴随着胶结方式转变,即由胞外多糖的黏结作用转变为藻丝体、地衣菌丝体以及苔藓假根的捆绑和缠绕,使生物结皮的固沙强度发生质的改变[16 ] (图2 D-F).生物结皮主要有藻类结皮(图2 A)、地衣结皮(图2 B)与苔藓结皮(图2 C)3种,分别代表生物结皮不同的演替阶段. ...
... 物种的分离和纯化主要针对藻类开展.野外采集藻类结皮样品,在室内利用BG11、BG110 、BBM等培养基进行培养.通过稀释平板法获得纯化物种并鉴定,鉴定方法包括传统显微镜法(形态)以及分子生物学测序(16S rDNA),选定该区域优势物种[9 ] .地衣取样时,选择成熟发育良好的地衣体,利用孢子释放法分离地衣共生菌和共生藻,获得单一孢子萌发形成的菌落和藻落,利用分子生物学方法鉴定共生菌和共生藻[36 ] .藓类个体较大,一般选择单一藓类通过形态学方法鉴定,并在室内扩繁. ...
... 目前,藻类和藓类的大规模室内培养和生产技术较为成熟,但地衣培养过程十分复杂,未见大规模室内培养报道.藻类培养的温度一般20—300 ,需要一定的光照,并且通过不断搅拌等方式供给氧气.在藻种的初级扩繁阶段,通常将试管原种接种到试管固体培养基上,在培养箱内(光强约2 000 lx,温度25—28 ℃)培养10—15 d后,将固体培养基培养的藻种进行下一级培养,接种入100—500 mL三角瓶中,同样条件下静置或通气培养8—10 d,进入扩大化培养阶段[9 ] .同时,也有将藻种悬浮液直接接入三角瓶中,摇床培养(光强约600 lx,温度25—30 ℃)7—10 d后,转为下一级(约60 L)培养的报道[15 ] .中国科学院新疆生态与地理研究所团队选择古尔班通古特沙漠优势物种,分离和培养,实现了接种藻类小规模的生产(图3 ).中国科学院武汉水生生物研究所团队在规模化藻类培养方面取得突破,通过藻类纯化、试管培养、三角瓶培养、10—18 L容器培养、小跑道池(1 m×4 m)和大跑道池(6 m×40 m)培养等步骤(图4 ),能够供给大量的藻类接种物[9 ] ,每步骤时间5—15 d[38 -39 ] .除了单种培养外,中国科学院沙坡头沙漠研究试验站和新疆生态与地理研究所均尝试利用生物结皮进行混合藻类的培养和规模化生产,此法虽然产量未及单种培养,但操作相对简单高效,也成为大规模培养的潜在选择[15 ] .地衣在室内需要经过固体、液体培养基接种、转化,共生藻和共生菌在不同的条件进行培养[40 ] ,但短期内并不能形成地衣植物.藓类植物的孢子体、配子体、茎叶碎片和芽孢等均可作为人工培养的材料[41 ] .组织培养时,可将配子体部分放入knop营养液,摇床培养,20 d后取出,自然风干待用[15 ] .组织培养的缺点是需要对外殖体进行灭菌消毒等,易对藓类本身造成损害.目前使用较多的为野外采集藓类植物的碎片作为材料进行接种,可实现低成本小规模培养[42 -43 ] .藓类培养时需注意后期水分的控制,干旱和完全湿润条件均不利于藓类植物的生长[44 ] .综合各研究案例可知,同一生物结皮类型中不同物种的最适培养条件不同,各研究团队采用的培养方法也具有差异性,但流程基本一致.在培养过程中,需要注意条件控制,如温度过高将导致培养目标藻类死亡或者其他污染藻类生长[9 ] . ...
... [9 ],每步骤时间5—15 d[38 -39 ] .除了单种培养外,中国科学院沙坡头沙漠研究试验站和新疆生态与地理研究所均尝试利用生物结皮进行混合藻类的培养和规模化生产,此法虽然产量未及单种培养,但操作相对简单高效,也成为大规模培养的潜在选择[15 ] .地衣在室内需要经过固体、液体培养基接种、转化,共生藻和共生菌在不同的条件进行培养[40 ] ,但短期内并不能形成地衣植物.藓类植物的孢子体、配子体、茎叶碎片和芽孢等均可作为人工培养的材料[41 ] .组织培养时,可将配子体部分放入knop营养液,摇床培养,20 d后取出,自然风干待用[15 ] .组织培养的缺点是需要对外殖体进行灭菌消毒等,易对藓类本身造成损害.目前使用较多的为野外采集藓类植物的碎片作为材料进行接种,可实现低成本小规模培养[42 -43 ] .藓类培养时需注意后期水分的控制,干旱和完全湿润条件均不利于藓类植物的生长[44 ] .综合各研究案例可知,同一生物结皮类型中不同物种的最适培养条件不同,各研究团队采用的培养方法也具有差异性,但流程基本一致.在培养过程中,需要注意条件控制,如温度过高将导致培养目标藻类死亡或者其他污染藻类生长[9 ] . ...
... [9 ]. ...
... 生物结皮野外接种流程见图5 .在生物结皮接种前,采用草方格、立体栅栏等方法先行稳定沙面,改善生物结皮恢复初期地表条件.干藻种可与沙土混匀播撒到地表,藻悬浮液以雾化微喷的方式接种到沙面[9 ,37 ] .随着技术的进步,可利用无人机、飞机撒播等手段在更大空间尺度实现培养物的均匀喷洒.野外恢复的同时,也可辅助以微喷溉系统每天添加适量的水(20 mm·d-1 ),来保持藻类的生存活力[39 ] .虽然藓类结皮室内大规模扩繁报道较多[40 -41 ,45 ] ,但利用室内扩繁藓类成功进行野外恢复的研究鲜见报道.目前,有研究者直接采集自然藓类结皮植株,粉碎后在其他受损区域进行撒播,辅以一定的遮阳、覆膜、营养液等措施,以促进该区域藓类结皮恢复[14 ,45 -46 ] .这种藓类结皮恢复的存在较大的资源限制,如果大规模应用,将会给自然发育的藓类结皮造成巨大资源压力,并有可能造成新的退化区.为促进接种物更快生长、形成生物结皮,除添加营养液解除土壤养分限制外,有如下模式参考. ...
... 从机械和生物模式组合角度,生物结皮野外接种初期,一些稳定的措施,包括物理措施(如草方格)和生物措施(草本、灌丛和乔木)的组合,将有助于生物结皮的发育[38 ,47 ] .Chen等[39 ] 研究表明,适度的降水和降低光强的措施有助于藻类结皮的快速生长.通过藻-草-灌-乔四位一体的模式,在库布齐沙漠的大规模流沙治理中得到了应用[9 ] . ...
... 中国是沙漠化发展最快的国家之一,造成生态系统失衡、耕地萎缩,严重地影响工农业生产和人民生活[4 ] .近年来,通过一系列生态工程的实施,荒漠化和沙化面积实现双缩减[55 ] .与“三北”防护林、京津源风沙源、退耕还林等工程相比,生物结皮的恢复工程规模相对较小.在降水量相对较少的干旱区,大规模植树造林的措施并不适用[4 ] .2001—2005年,中国科学院与内蒙古自治区在库布齐沙漠建立了生物结皮与乔灌草结合的示范区,成功治理了近200 hm2 流沙,并在其他区域进行大面积推广,该项目成为了生物结皮野外成功恢复的典范[9 ] .同时,该团队采用了工程化机械接种技术克服了人工生物结皮接种耗时费工的缺点,成为生物结皮工程化大面积应用的重要措施.除此之外,在其他区域,如腾格里沙漠、古尔班通古特沙漠、毛乌素沙地、黄土高原区等均有相应的生物结皮恢复应用的报道,但均处于方法探索和前期试验阶段[15 ,56 -58 ] (表1 ).国外生物结皮恢复研究虽然开展较早,但还未见能形成规模的生物结皮恢复示范区,中国生物结皮的恢复为世界生物结皮的恢复提供了重要借鉴. ...
... Study case of biological soil crust rehabilitation in China
Table 1 结皮类型 地点 规模 结皮生长 评价 文献 藻类结皮 库布齐沙漠 约200 hm2 2—3年形成3—5 mm藻类结皮,盖度48.5% 结皮和植被得到较好恢复 [9 ,38 ,51 ] 腾格里沙漠 2 m×2 m 5个月后,0.39 mm,盖度达14.3%,生物量13.4 mg·g-1 ;盖度或先增后降,1年后13.8% 长期效应未知 [12 ,15 ] 古尔班通古特沙漠 2 m×5 m 15 d可达2.6 mg·cm-2 (叶绿素a) 未形成大规模生物结皮 未发表 地衣结皮 腾格里沙漠 3个 0.5 m2 1年后形成4—6 cm生物结皮 长期效应未知 [36 ] 藓类结皮 腾格里沙漠 10个1 m×1 m 7 d内存活,3个月内死亡 接种藓类直接采自野外 [59 ] 毛乌素沙地 18个1 m×1 m 75 d后盖度70%,密度9.8 株·cm-2 ,4年盖度达98% 接种藓类直接采自野外 [45 ,57 ] 杨凌 48个1 m×1 m 30 d,盖度90%,最大密度120株·cm-2 接种藓类直接采自野外 [14 ] 库布齐沙漠 8个1 m×1 m 3年后,盖度30%—70%,185株·cm-2 接种藓类直接采自野外 [60 ]
<strong>3.4</strong> 人工培养生物结皮的抗性锻炼 为提高野外存活率,是否对扩繁后的材料进行模拟野外环境的抗性锻炼,具有不同见解.抗性锻炼主要包括干燥和水分循环、高温和低温循环、不同光强等,以增加培养物的适合度[59 -62 ] .通过对20个蓝藻菌株进行抗性锻炼发现,13个菌株均表现出适合度的增加,尤其是结皮先锋藻类微鞘藻属的菌株,适合度均得到提升[63 ] .但通过培养混合生物结皮(藻类+地衣+藓类),进行一系列抗性锻炼(全谱光+温度变化+干旱),发现抗性锻炼无显著效果[62 ] ,藓类结皮物种作用同样不显著[64 ] .对不同种类藓类样品在不同存储温度研究发现,不同藓类种类后期生长对储存温度的敏感性不一致[65 ] .因此,生物结皮在接种前,是否有必要进行抗性锻炼,有待进一步验证. ...
Rapidly restoring biological soil crusts and ecosystem functions in a severely disturbed desert ecosystem
1
2016
... 生物结皮从发育初期到成熟期,经历不同的演替阶段,形成不同类型的生物结皮,结构逐步趋于稳定.发育伊始,寡营养细菌可分泌胞外多糖粘结沙粒,形成生物结皮早期结构.随着藻类、地衣和苔藓植物的不断出现,胞外多糖粘结沙粒的作用逐渐被藻丝体、菌丝体以及苔藓假根捆绑沙粒的作用取代,生物结皮的结构更加复杂和稳定,构成生物结皮固沙的重要结构基础.同时,生物结皮中很多种类具有固氮和固碳功能,在改善土壤养分和水分条件以及抗风蚀方面发挥重要生态功能,在荒漠区固沙与受损地表系统的修复领域具有广阔的应用前景.近年来,由于干旱区传统生物固沙所面临的水资源缺乏和土壤水分消耗等问题,通过人工培育生物结皮进行固沙与修复荒漠受损地表生态,逐渐成为恢复生态学者关注的焦点.通过在裸露沙面或合理密度条件下的固沙植物间(或物理沙障间)接种生物结皮,既能达到防风固沙的目的,也能减少水分消耗来维持植物长期生存[9 ] .同时,在干旱区退化生态系统恢复方面,生物结皮可以快速增加土壤稳定性,抵抗水土侵蚀,恢复受损表面[10 -12 ] .中国在生物结皮固沙和退化生态系统恢复领域具有较丰厚的研究基础,并在一定范围内取得了成功的实践,如库布齐沙漠、腾格里沙漠和黄土高原[9 ,13 -14 ] .野外人工生物结皮接种和生长有望成为干旱区受损生态系统恢复的新模式,对干旱区传统防沙治沙和退化系统恢复模式形成重要补充. ...
Production of a biocrust-cyanobacteria strain (Nostoc commune ) for large-scale restoration of dryland soils
2019
Towards stopping land degradation in drylands:water-saving techniques for cultivating biocrusts in situ
2
2019
... 生物结皮从发育初期到成熟期,经历不同的演替阶段,形成不同类型的生物结皮,结构逐步趋于稳定.发育伊始,寡营养细菌可分泌胞外多糖粘结沙粒,形成生物结皮早期结构.随着藻类、地衣和苔藓植物的不断出现,胞外多糖粘结沙粒的作用逐渐被藻丝体、菌丝体以及苔藓假根捆绑沙粒的作用取代,生物结皮的结构更加复杂和稳定,构成生物结皮固沙的重要结构基础.同时,生物结皮中很多种类具有固氮和固碳功能,在改善土壤养分和水分条件以及抗风蚀方面发挥重要生态功能,在荒漠区固沙与受损地表系统的修复领域具有广阔的应用前景.近年来,由于干旱区传统生物固沙所面临的水资源缺乏和土壤水分消耗等问题,通过人工培育生物结皮进行固沙与修复荒漠受损地表生态,逐渐成为恢复生态学者关注的焦点.通过在裸露沙面或合理密度条件下的固沙植物间(或物理沙障间)接种生物结皮,既能达到防风固沙的目的,也能减少水分消耗来维持植物长期生存[9 ] .同时,在干旱区退化生态系统恢复方面,生物结皮可以快速增加土壤稳定性,抵抗水土侵蚀,恢复受损表面[10 -12 ] .中国在生物结皮固沙和退化生态系统恢复领域具有较丰厚的研究基础,并在一定范围内取得了成功的实践,如库布齐沙漠、腾格里沙漠和黄土高原[9 ,13 -14 ] .野外人工生物结皮接种和生长有望成为干旱区受损生态系统恢复的新模式,对干旱区传统防沙治沙和退化系统恢复模式形成重要补充. ...
... Study case of biological soil crust rehabilitation in China
Table 1 结皮类型 地点 规模 结皮生长 评价 文献 藻类结皮 库布齐沙漠 约200 hm2 2—3年形成3—5 mm藻类结皮,盖度48.5% 结皮和植被得到较好恢复 [9 ,38 ,51 ] 腾格里沙漠 2 m×2 m 5个月后,0.39 mm,盖度达14.3%,生物量13.4 mg·g-1 ;盖度或先增后降,1年后13.8% 长期效应未知 [12 ,15 ] 古尔班通古特沙漠 2 m×5 m 15 d可达2.6 mg·cm-2 (叶绿素a) 未形成大规模生物结皮 未发表 地衣结皮 腾格里沙漠 3个 0.5 m2 1年后形成4—6 cm生物结皮 长期效应未知 [36 ] 藓类结皮 腾格里沙漠 10个1 m×1 m 7 d内存活,3个月内死亡 接种藓类直接采自野外 [59 ] 毛乌素沙地 18个1 m×1 m 75 d后盖度70%,密度9.8 株·cm-2 ,4年盖度达98% 接种藓类直接采自野外 [45 ,57 ] 杨凌 48个1 m×1 m 30 d,盖度90%,最大密度120株·cm-2 接种藓类直接采自野外 [14 ] 库布齐沙漠 8个1 m×1 m 3年后,盖度30%—70%,185株·cm-2 接种藓类直接采自野外 [60 ]
<strong>3.4</strong> 人工培养生物结皮的抗性锻炼 为提高野外存活率,是否对扩繁后的材料进行模拟野外环境的抗性锻炼,具有不同见解.抗性锻炼主要包括干燥和水分循环、高温和低温循环、不同光强等,以增加培养物的适合度[59 -62 ] .通过对20个蓝藻菌株进行抗性锻炼发现,13个菌株均表现出适合度的增加,尤其是结皮先锋藻类微鞘藻属的菌株,适合度均得到提升[63 ] .但通过培养混合生物结皮(藻类+地衣+藓类),进行一系列抗性锻炼(全谱光+温度变化+干旱),发现抗性锻炼无显著效果[62 ] ,藓类结皮物种作用同样不显著[64 ] .对不同种类藓类样品在不同存储温度研究发现,不同藓类种类后期生长对储存温度的敏感性不一致[65 ] .因此,生物结皮在接种前,是否有必要进行抗性锻炼,有待进一步验证. ...
Rapid development of cyanobacterial crust in the field for combating desertification
1
2017
... 生物结皮从发育初期到成熟期,经历不同的演替阶段,形成不同类型的生物结皮,结构逐步趋于稳定.发育伊始,寡营养细菌可分泌胞外多糖粘结沙粒,形成生物结皮早期结构.随着藻类、地衣和苔藓植物的不断出现,胞外多糖粘结沙粒的作用逐渐被藻丝体、菌丝体以及苔藓假根捆绑沙粒的作用取代,生物结皮的结构更加复杂和稳定,构成生物结皮固沙的重要结构基础.同时,生物结皮中很多种类具有固氮和固碳功能,在改善土壤养分和水分条件以及抗风蚀方面发挥重要生态功能,在荒漠区固沙与受损地表系统的修复领域具有广阔的应用前景.近年来,由于干旱区传统生物固沙所面临的水资源缺乏和土壤水分消耗等问题,通过人工培育生物结皮进行固沙与修复荒漠受损地表生态,逐渐成为恢复生态学者关注的焦点.通过在裸露沙面或合理密度条件下的固沙植物间(或物理沙障间)接种生物结皮,既能达到防风固沙的目的,也能减少水分消耗来维持植物长期生存[9 ] .同时,在干旱区退化生态系统恢复方面,生物结皮可以快速增加土壤稳定性,抵抗水土侵蚀,恢复受损表面[10 -12 ] .中国在生物结皮固沙和退化生态系统恢复领域具有较丰厚的研究基础,并在一定范围内取得了成功的实践,如库布齐沙漠、腾格里沙漠和黄土高原[9 ,13 -14 ] .野外人工生物结皮接种和生长有望成为干旱区受损生态系统恢复的新模式,对干旱区传统防沙治沙和退化系统恢复模式形成重要补充. ...
Successful field cultivation of moss biocrusts on disturbed soil surfaces in the short term
3
2018
... 生物结皮从发育初期到成熟期,经历不同的演替阶段,形成不同类型的生物结皮,结构逐步趋于稳定.发育伊始,寡营养细菌可分泌胞外多糖粘结沙粒,形成生物结皮早期结构.随着藻类、地衣和苔藓植物的不断出现,胞外多糖粘结沙粒的作用逐渐被藻丝体、菌丝体以及苔藓假根捆绑沙粒的作用取代,生物结皮的结构更加复杂和稳定,构成生物结皮固沙的重要结构基础.同时,生物结皮中很多种类具有固氮和固碳功能,在改善土壤养分和水分条件以及抗风蚀方面发挥重要生态功能,在荒漠区固沙与受损地表系统的修复领域具有广阔的应用前景.近年来,由于干旱区传统生物固沙所面临的水资源缺乏和土壤水分消耗等问题,通过人工培育生物结皮进行固沙与修复荒漠受损地表生态,逐渐成为恢复生态学者关注的焦点.通过在裸露沙面或合理密度条件下的固沙植物间(或物理沙障间)接种生物结皮,既能达到防风固沙的目的,也能减少水分消耗来维持植物长期生存[9 ] .同时,在干旱区退化生态系统恢复方面,生物结皮可以快速增加土壤稳定性,抵抗水土侵蚀,恢复受损表面[10 -12 ] .中国在生物结皮固沙和退化生态系统恢复领域具有较丰厚的研究基础,并在一定范围内取得了成功的实践,如库布齐沙漠、腾格里沙漠和黄土高原[9 ,13 -14 ] .野外人工生物结皮接种和生长有望成为干旱区受损生态系统恢复的新模式,对干旱区传统防沙治沙和退化系统恢复模式形成重要补充. ...
... 生物结皮野外接种流程见图5 .在生物结皮接种前,采用草方格、立体栅栏等方法先行稳定沙面,改善生物结皮恢复初期地表条件.干藻种可与沙土混匀播撒到地表,藻悬浮液以雾化微喷的方式接种到沙面[9 ,37 ] .随着技术的进步,可利用无人机、飞机撒播等手段在更大空间尺度实现培养物的均匀喷洒.野外恢复的同时,也可辅助以微喷溉系统每天添加适量的水(20 mm·d-1 ),来保持藻类的生存活力[39 ] .虽然藓类结皮室内大规模扩繁报道较多[40 -41 ,45 ] ,但利用室内扩繁藓类成功进行野外恢复的研究鲜见报道.目前,有研究者直接采集自然藓类结皮植株,粉碎后在其他受损区域进行撒播,辅以一定的遮阳、覆膜、营养液等措施,以促进该区域藓类结皮恢复[14 ,45 -46 ] .这种藓类结皮恢复的存在较大的资源限制,如果大规模应用,将会给自然发育的藓类结皮造成巨大资源压力,并有可能造成新的退化区.为促进接种物更快生长、形成生物结皮,除添加营养液解除土壤养分限制外,有如下模式参考. ...
... Study case of biological soil crust rehabilitation in China
Table 1 结皮类型 地点 规模 结皮生长 评价 文献 藻类结皮 库布齐沙漠 约200 hm2 2—3年形成3—5 mm藻类结皮,盖度48.5% 结皮和植被得到较好恢复 [9 ,38 ,51 ] 腾格里沙漠 2 m×2 m 5个月后,0.39 mm,盖度达14.3%,生物量13.4 mg·g-1 ;盖度或先增后降,1年后13.8% 长期效应未知 [12 ,15 ] 古尔班通古特沙漠 2 m×5 m 15 d可达2.6 mg·cm-2 (叶绿素a) 未形成大规模生物结皮 未发表 地衣结皮 腾格里沙漠 3个 0.5 m2 1年后形成4—6 cm生物结皮 长期效应未知 [36 ] 藓类结皮 腾格里沙漠 10个1 m×1 m 7 d内存活,3个月内死亡 接种藓类直接采自野外 [59 ] 毛乌素沙地 18个1 m×1 m 75 d后盖度70%,密度9.8 株·cm-2 ,4年盖度达98% 接种藓类直接采自野外 [45 ,57 ] 杨凌 48个1 m×1 m 30 d,盖度90%,最大密度120株·cm-2 接种藓类直接采自野外 [14 ] 库布齐沙漠 8个1 m×1 m 3年后,盖度30%—70%,185株·cm-2 接种藓类直接采自野外 [60 ]
<strong>3.4</strong> 人工培养生物结皮的抗性锻炼 为提高野外存活率,是否对扩繁后的材料进行模拟野外环境的抗性锻炼,具有不同见解.抗性锻炼主要包括干燥和水分循环、高温和低温循环、不同光强等,以增加培养物的适合度[59 -62 ] .通过对20个蓝藻菌株进行抗性锻炼发现,13个菌株均表现出适合度的增加,尤其是结皮先锋藻类微鞘藻属的菌株,适合度均得到提升[63 ] .但通过培养混合生物结皮(藻类+地衣+藓类),进行一系列抗性锻炼(全谱光+温度变化+干旱),发现抗性锻炼无显著效果[62 ] ,藓类结皮物种作用同样不显著[64 ] .对不同种类藓类样品在不同存储温度研究发现,不同藓类种类后期生长对储存温度的敏感性不一致[65 ] .因此,生物结皮在接种前,是否有必要进行抗性锻炼,有待进一步验证. ...
8
2016
... 生物结皮不同于物理和化学结皮(雨水或外力形成),是由活的生物体及其代谢产物,通过有机物和土壤颗粒黏结而成[9 ] (图2 ).生物结皮从形成初期到成熟期,经历了物种更替和不同演替阶段[15 ] .生物结皮的演替伴随着胶结方式转变,即由胞外多糖的黏结作用转变为藻丝体、地衣菌丝体以及苔藓假根的捆绑和缠绕,使生物结皮的固沙强度发生质的改变[16 ] (图2 D-F).生物结皮主要有藻类结皮(图2 A)、地衣结皮(图2 B)与苔藓结皮(图2 C)3种,分别代表生物结皮不同的演替阶段. ...
... 目前,藻类和藓类的大规模室内培养和生产技术较为成熟,但地衣培养过程十分复杂,未见大规模室内培养报道.藻类培养的温度一般20—300 ,需要一定的光照,并且通过不断搅拌等方式供给氧气.在藻种的初级扩繁阶段,通常将试管原种接种到试管固体培养基上,在培养箱内(光强约2 000 lx,温度25—28 ℃)培养10—15 d后,将固体培养基培养的藻种进行下一级培养,接种入100—500 mL三角瓶中,同样条件下静置或通气培养8—10 d,进入扩大化培养阶段[9 ] .同时,也有将藻种悬浮液直接接入三角瓶中,摇床培养(光强约600 lx,温度25—30 ℃)7—10 d后,转为下一级(约60 L)培养的报道[15 ] .中国科学院新疆生态与地理研究所团队选择古尔班通古特沙漠优势物种,分离和培养,实现了接种藻类小规模的生产(图3 ).中国科学院武汉水生生物研究所团队在规模化藻类培养方面取得突破,通过藻类纯化、试管培养、三角瓶培养、10—18 L容器培养、小跑道池(1 m×4 m)和大跑道池(6 m×40 m)培养等步骤(图4 ),能够供给大量的藻类接种物[9 ] ,每步骤时间5—15 d[38 -39 ] .除了单种培养外,中国科学院沙坡头沙漠研究试验站和新疆生态与地理研究所均尝试利用生物结皮进行混合藻类的培养和规模化生产,此法虽然产量未及单种培养,但操作相对简单高效,也成为大规模培养的潜在选择[15 ] .地衣在室内需要经过固体、液体培养基接种、转化,共生藻和共生菌在不同的条件进行培养[40 ] ,但短期内并不能形成地衣植物.藓类植物的孢子体、配子体、茎叶碎片和芽孢等均可作为人工培养的材料[41 ] .组织培养时,可将配子体部分放入knop营养液,摇床培养,20 d后取出,自然风干待用[15 ] .组织培养的缺点是需要对外殖体进行灭菌消毒等,易对藓类本身造成损害.目前使用较多的为野外采集藓类植物的碎片作为材料进行接种,可实现低成本小规模培养[42 -43 ] .藓类培养时需注意后期水分的控制,干旱和完全湿润条件均不利于藓类植物的生长[44 ] .综合各研究案例可知,同一生物结皮类型中不同物种的最适培养条件不同,各研究团队采用的培养方法也具有差异性,但流程基本一致.在培养过程中,需要注意条件控制,如温度过高将导致培养目标藻类死亡或者其他污染藻类生长[9 ] . ...
... [15 ].地衣在室内需要经过固体、液体培养基接种、转化,共生藻和共生菌在不同的条件进行培养[40 ] ,但短期内并不能形成地衣植物.藓类植物的孢子体、配子体、茎叶碎片和芽孢等均可作为人工培养的材料[41 ] .组织培养时,可将配子体部分放入knop营养液,摇床培养,20 d后取出,自然风干待用[15 ] .组织培养的缺点是需要对外殖体进行灭菌消毒等,易对藓类本身造成损害.目前使用较多的为野外采集藓类植物的碎片作为材料进行接种,可实现低成本小规模培养[42 -43 ] .藓类培养时需注意后期水分的控制,干旱和完全湿润条件均不利于藓类植物的生长[44 ] .综合各研究案例可知,同一生物结皮类型中不同物种的最适培养条件不同,各研究团队采用的培养方法也具有差异性,但流程基本一致.在培养过程中,需要注意条件控制,如温度过高将导致培养目标藻类死亡或者其他污染藻类生长[9 ] . ...
... [15 ].组织培养的缺点是需要对外殖体进行灭菌消毒等,易对藓类本身造成损害.目前使用较多的为野外采集藓类植物的碎片作为材料进行接种,可实现低成本小规模培养[42 -43 ] .藓类培养时需注意后期水分的控制,干旱和完全湿润条件均不利于藓类植物的生长[44 ] .综合各研究案例可知,同一生物结皮类型中不同物种的最适培养条件不同,各研究团队采用的培养方法也具有差异性,但流程基本一致.在培养过程中,需要注意条件控制,如温度过高将导致培养目标藻类死亡或者其他污染藻类生长[9 ] . ...
... 从接种藻类物种组成角度,可分成3种,选择接种区域的优势藻种:①单种培养并单种施加,如利用具鞘微鞘藻(Microcoleus vaginatus )进行培养和单独接种[39 ] ;②多种分开培养,接种前混合,如将具鞘微鞘藻和爪哇伪枝藻(S. javanicum )按干物质重10∶1进行混合接种[38 ] ,或者具鞘微鞘藻、席藻和爪哇伪枝藻按10∶5∶1混合接种;③混合培养、混合接种,如腾格里沙漠的部分实验[15 ] ,将藻类从结皮中直接分离培养,不纯化,节约了时间、技术和经济成本. ...
... 从促进藻类生长化学试剂角度,施用无毒副作用且价格低廉的化合物,是除物理和生物措施外,辅助藻类生长的重要补充.如海藻酸钠等可在沙面形成具有抗压能力的薄层,同时可促进蓝藻的克隆和生长[48 ] .有研究将藻类包裹制成海藻酸钠丸,施加到样地,取得较好的效果[49 ] .促进保水以及生物结皮生长的还有高持水性的化学材料(如SAP)、聚乙烯醇(PVA)和Tacki-Spray等化学试剂[15 ,50 ] . ...
... 中国是沙漠化发展最快的国家之一,造成生态系统失衡、耕地萎缩,严重地影响工农业生产和人民生活[4 ] .近年来,通过一系列生态工程的实施,荒漠化和沙化面积实现双缩减[55 ] .与“三北”防护林、京津源风沙源、退耕还林等工程相比,生物结皮的恢复工程规模相对较小.在降水量相对较少的干旱区,大规模植树造林的措施并不适用[4 ] .2001—2005年,中国科学院与内蒙古自治区在库布齐沙漠建立了生物结皮与乔灌草结合的示范区,成功治理了近200 hm2 流沙,并在其他区域进行大面积推广,该项目成为了生物结皮野外成功恢复的典范[9 ] .同时,该团队采用了工程化机械接种技术克服了人工生物结皮接种耗时费工的缺点,成为生物结皮工程化大面积应用的重要措施.除此之外,在其他区域,如腾格里沙漠、古尔班通古特沙漠、毛乌素沙地、黄土高原区等均有相应的生物结皮恢复应用的报道,但均处于方法探索和前期试验阶段[15 ,56 -58 ] (表1 ).国外生物结皮恢复研究虽然开展较早,但还未见能形成规模的生物结皮恢复示范区,中国生物结皮的恢复为世界生物结皮的恢复提供了重要借鉴. ...
... Study case of biological soil crust rehabilitation in China
Table 1 结皮类型 地点 规模 结皮生长 评价 文献 藻类结皮 库布齐沙漠 约200 hm2 2—3年形成3—5 mm藻类结皮,盖度48.5% 结皮和植被得到较好恢复 [9 ,38 ,51 ] 腾格里沙漠 2 m×2 m 5个月后,0.39 mm,盖度达14.3%,生物量13.4 mg·g-1 ;盖度或先增后降,1年后13.8% 长期效应未知 [12 ,15 ] 古尔班通古特沙漠 2 m×5 m 15 d可达2.6 mg·cm-2 (叶绿素a) 未形成大规模生物结皮 未发表 地衣结皮 腾格里沙漠 3个 0.5 m2 1年后形成4—6 cm生物结皮 长期效应未知 [36 ] 藓类结皮 腾格里沙漠 10个1 m×1 m 7 d内存活,3个月内死亡 接种藓类直接采自野外 [59 ] 毛乌素沙地 18个1 m×1 m 75 d后盖度70%,密度9.8 株·cm-2 ,4年盖度达98% 接种藓类直接采自野外 [45 ,57 ] 杨凌 48个1 m×1 m 30 d,盖度90%,最大密度120株·cm-2 接种藓类直接采自野外 [14 ] 库布齐沙漠 8个1 m×1 m 3年后,盖度30%—70%,185株·cm-2 接种藓类直接采自野外 [60 ]
<strong>3.4</strong> 人工培养生物结皮的抗性锻炼 为提高野外存活率,是否对扩繁后的材料进行模拟野外环境的抗性锻炼,具有不同见解.抗性锻炼主要包括干燥和水分循环、高温和低温循环、不同光强等,以增加培养物的适合度[59 -62 ] .通过对20个蓝藻菌株进行抗性锻炼发现,13个菌株均表现出适合度的增加,尤其是结皮先锋藻类微鞘藻属的菌株,适合度均得到提升[63 ] .但通过培养混合生物结皮(藻类+地衣+藓类),进行一系列抗性锻炼(全谱光+温度变化+干旱),发现抗性锻炼无显著效果[62 ] ,藓类结皮物种作用同样不显著[64 ] .对不同种类藓类样品在不同存储温度研究发现,不同藓类种类后期生长对储存温度的敏感性不一致[65 ] .因此,生物结皮在接种前,是否有必要进行抗性锻炼,有待进一步验证. ...
荒漠地表生物土壤结皮形成与演替特征概述
1
2010
... 生物结皮不同于物理和化学结皮(雨水或外力形成),是由活的生物体及其代谢产物,通过有机物和土壤颗粒黏结而成[9 ] (图2 ).生物结皮从形成初期到成熟期,经历了物种更替和不同演替阶段[15 ] .生物结皮的演替伴随着胶结方式转变,即由胞外多糖的黏结作用转变为藻丝体、地衣菌丝体以及苔藓假根的捆绑和缠绕,使生物结皮的固沙强度发生质的改变[16 ] (图2 D-F).生物结皮主要有藻类结皮(图2 A)、地衣结皮(图2 B)与苔藓结皮(图2 C)3种,分别代表生物结皮不同的演替阶段. ...
荒漠地表生物土壤结皮的微结构及其早期发育特征
1
2005
... 生物结皮发育前期,芽孢杆菌等寡营养细菌可分泌胞外多糖类黏液黏结沙粒[17 ] .随着具有光合自养能力的蓝藻类尤其是丝状蓝藻的出现,胞外多糖的分泌量和沙粒黏结能力进一步增强(图2 D).蓝藻可移动并以藻殖段拓展繁殖,在生物结皮恢复中具有较大优势[18 -19 ] .除蓝藻外,生物结皮中还具绿藻和硅藻,多以单细胞构成. ...
丝状体蓝藻藻殖段的分化及其调节机制
1
2000
... 生物结皮发育前期,芽孢杆菌等寡营养细菌可分泌胞外多糖类黏液黏结沙粒[17 ] .随着具有光合自养能力的蓝藻类尤其是丝状蓝藻的出现,胞外多糖的分泌量和沙粒黏结能力进一步增强(图2 D).蓝藻可移动并以藻殖段拓展繁殖,在生物结皮恢复中具有较大优势[18 -19 ] .除蓝藻外,生物结皮中还具绿藻和硅藻,多以单细胞构成. ...
Reproductive biology of cyanophycota
1
2001
... 生物结皮发育前期,芽孢杆菌等寡营养细菌可分泌胞外多糖类黏液黏结沙粒[17 ] .随着具有光合自养能力的蓝藻类尤其是丝状蓝藻的出现,胞外多糖的分泌量和沙粒黏结能力进一步增强(图2 D).蓝藻可移动并以藻殖段拓展繁殖,在生物结皮恢复中具有较大优势[18 -19 ] .除蓝藻外,生物结皮中还具绿藻和硅藻,多以单细胞构成. ...
荒漠藻壳的精细结构与发育
1
2000
... 地衣是由真菌和蓝藻(或绿藻)组成的互惠共生体,地衣结皮的形成是真菌侵入藻类结皮的结果,真菌通过藻类结皮边缘,伸入、生长,与藻丝或藻细胞结合,藻类结皮演变成地衣结皮[20 ] .地衣具有的菌丝体可穿透土壤,提高沙面稳定性(图2 E). ...
Enhanced recovery of Bbiological soil crusts after disturbance
2
2016
... 苔藓能够在沙漠中斑块状聚集和分布,形成肥岛,同时具有发达的假根系统,有助于沙面的稳定和沙面多样性维持(图2 F).苔藓的细胞具有全能性并在一定条件下保持长时间活力,因此能够用来进行快速的恢复[21 ] . ...
... 生物结皮人工培养物种选择的重要原则是对本地生物结皮优势物种的筛选,不仅可以保障培养体具有较强的抗逆性和适应性,也可避免外来物种生物入侵风险.同时,筛选的目标物种应具有适应高温、高pH、高盐碱、强辐射等逆境生境的能力[21 ] .多数荒漠的藻类结皮中,以微鞘藻属(Microcoleus )种类分布最广,因此该属种类是藻类结皮恢复最重要的目标种.其他优势物种也常用于人工藻类结皮恢复,如库布齐沙漠选择席藻(Phormidium tenue )和伪枝藻(Scytonema javanicum ),腾格里沙漠选择念珠藻属(Nostoc )和席藻属(Phormidium )等.中国地衣结皮的培养仅见地衣石果衣(Endocarpon pusillum )报道,包括石果衣共生真菌和石果衣共生藻的分离培养[36 ] .中国各荒漠区藓类结皮优势物种不同,常见有黄土高原极地真藓(Bryum arcticum )、毛乌素沙地黄色真藓(Bryum pallescens )、腾格里沙漠银叶真藓(Bryum argenteum )、古尔班通古特沙漠齿肋赤藓(Syntrichia caninervis )等用于藓类结皮培养与恢复研究. ...
Composition and macrostructure of biological soil crusts
1
2016
... A—C,3种生物结皮类型; D—F,胶结方式,修改自Colesie等[22 ] ; G—I,物种组成 ...
and global scales
1
2016
... 生物结皮中的蓝藻、藓类等光合类群,能够固碳,促进有机质的形成和增加.全球尺度生物结皮年总固碳量可达0.6 Pg, 相当于全球陆地生态系统净生产力的1%,在荒漠区占比可达9%[8 ,23 ] .同时,生物结皮中具有大量的固氮细菌和蓝藻,全球年固氮量可达26 Tg,约占全球孢子植物固氮量的50%,以及陆地生态系统生物总固氮量的1/4[7 -8 ] .生物结皮中丰富的微生物可矿化和溶解土壤中不溶性无机磷和有机磷复合,促进磷的吸收[24 ] .因此,生物结皮斑块是干旱区特殊的肥岛,能够促进小尺度养分的聚集[25 -27 ] .除养分外,随着生物结皮的发育,生物结皮表面粗糙度增加,增加土壤水分入渗和留存[28 ] .与裸沙相比,生物结皮可降低水分蒸发,增加对大气中凝结水、蒸汽和雾等的收集,有利于干旱区生物结皮表面的水分供给和微生物的活动[29 -30 ] .总体来讲,生物结皮对养分和水分的聚集,有助于荒漠生态系统正向演替. ...
Biological soil crusts as key player in biogeochemical P cycling during pedogenesis of sandy substrate
1
2019
... 生物结皮中的蓝藻、藓类等光合类群,能够固碳,促进有机质的形成和增加.全球尺度生物结皮年总固碳量可达0.6 Pg, 相当于全球陆地生态系统净生产力的1%,在荒漠区占比可达9%[8 ,23 ] .同时,生物结皮中具有大量的固氮细菌和蓝藻,全球年固氮量可达26 Tg,约占全球孢子植物固氮量的50%,以及陆地生态系统生物总固氮量的1/4[7 -8 ] .生物结皮中丰富的微生物可矿化和溶解土壤中不溶性无机磷和有机磷复合,促进磷的吸收[24 ] .因此,生物结皮斑块是干旱区特殊的肥岛,能够促进小尺度养分的聚集[25 -27 ] .除养分外,随着生物结皮的发育,生物结皮表面粗糙度增加,增加土壤水分入渗和留存[28 ] .与裸沙相比,生物结皮可降低水分蒸发,增加对大气中凝结水、蒸汽和雾等的收集,有利于干旱区生物结皮表面的水分供给和微生物的活动[29 -30 ] .总体来讲,生物结皮对养分和水分的聚集,有助于荒漠生态系统正向演替. ...
Linking biological soil crust attributes to the multifunctionality of vegetated patches and interspaces in a semiarid shrubland
1
2018
... 生物结皮中的蓝藻、藓类等光合类群,能够固碳,促进有机质的形成和增加.全球尺度生物结皮年总固碳量可达0.6 Pg, 相当于全球陆地生态系统净生产力的1%,在荒漠区占比可达9%[8 ,23 ] .同时,生物结皮中具有大量的固氮细菌和蓝藻,全球年固氮量可达26 Tg,约占全球孢子植物固氮量的50%,以及陆地生态系统生物总固氮量的1/4[7 -8 ] .生物结皮中丰富的微生物可矿化和溶解土壤中不溶性无机磷和有机磷复合,促进磷的吸收[24 ] .因此,生物结皮斑块是干旱区特殊的肥岛,能够促进小尺度养分的聚集[25 -27 ] .除养分外,随着生物结皮的发育,生物结皮表面粗糙度增加,增加土壤水分入渗和留存[28 ] .与裸沙相比,生物结皮可降低水分蒸发,增加对大气中凝结水、蒸汽和雾等的收集,有利于干旱区生物结皮表面的水分供给和微生物的活动[29 -30 ] .总体来讲,生物结皮对养分和水分的聚集,有助于荒漠生态系统正向演替. ...
Moss patch size and microhabitats influence stoichiometry of moss crusts in a temperate desert,Central Asia
2019
Biocrusts,inside and outside resource islands of Mimosa luisana (Leguminosae),improve soil carbon and nitrogen dynamics in a tropical semiarid ecosystem
1
2016
... 生物结皮中的蓝藻、藓类等光合类群,能够固碳,促进有机质的形成和增加.全球尺度生物结皮年总固碳量可达0.6 Pg, 相当于全球陆地生态系统净生产力的1%,在荒漠区占比可达9%[8 ,23 ] .同时,生物结皮中具有大量的固氮细菌和蓝藻,全球年固氮量可达26 Tg,约占全球孢子植物固氮量的50%,以及陆地生态系统生物总固氮量的1/4[7 -8 ] .生物结皮中丰富的微生物可矿化和溶解土壤中不溶性无机磷和有机磷复合,促进磷的吸收[24 ] .因此,生物结皮斑块是干旱区特殊的肥岛,能够促进小尺度养分的聚集[25 -27 ] .除养分外,随着生物结皮的发育,生物结皮表面粗糙度增加,增加土壤水分入渗和留存[28 ] .与裸沙相比,生物结皮可降低水分蒸发,增加对大气中凝结水、蒸汽和雾等的收集,有利于干旱区生物结皮表面的水分供给和微生物的活动[29 -30 ] .总体来讲,生物结皮对养分和水分的聚集,有助于荒漠生态系统正向演替. ...
The role of biocrusts in arid land hydrology
1
2016
... 生物结皮中的蓝藻、藓类等光合类群,能够固碳,促进有机质的形成和增加.全球尺度生物结皮年总固碳量可达0.6 Pg, 相当于全球陆地生态系统净生产力的1%,在荒漠区占比可达9%[8 ,23 ] .同时,生物结皮中具有大量的固氮细菌和蓝藻,全球年固氮量可达26 Tg,约占全球孢子植物固氮量的50%,以及陆地生态系统生物总固氮量的1/4[7 -8 ] .生物结皮中丰富的微生物可矿化和溶解土壤中不溶性无机磷和有机磷复合,促进磷的吸收[24 ] .因此,生物结皮斑块是干旱区特殊的肥岛,能够促进小尺度养分的聚集[25 -27 ] .除养分外,随着生物结皮的发育,生物结皮表面粗糙度增加,增加土壤水分入渗和留存[28 ] .与裸沙相比,生物结皮可降低水分蒸发,增加对大气中凝结水、蒸汽和雾等的收集,有利于干旱区生物结皮表面的水分供给和微生物的活动[29 -30 ] .总体来讲,生物结皮对养分和水分的聚集,有助于荒漠生态系统正向演替. ...
The influence of biological soil crusts on dew deposition in Gurbantunggut Desert,Northwestern China
1
2009
... 生物结皮中的蓝藻、藓类等光合类群,能够固碳,促进有机质的形成和增加.全球尺度生物结皮年总固碳量可达0.6 Pg, 相当于全球陆地生态系统净生产力的1%,在荒漠区占比可达9%[8 ,23 ] .同时,生物结皮中具有大量的固氮细菌和蓝藻,全球年固氮量可达26 Tg,约占全球孢子植物固氮量的50%,以及陆地生态系统生物总固氮量的1/4[7 -8 ] .生物结皮中丰富的微生物可矿化和溶解土壤中不溶性无机磷和有机磷复合,促进磷的吸收[24 ] .因此,生物结皮斑块是干旱区特殊的肥岛,能够促进小尺度养分的聚集[25 -27 ] .除养分外,随着生物结皮的发育,生物结皮表面粗糙度增加,增加土壤水分入渗和留存[28 ] .与裸沙相比,生物结皮可降低水分蒸发,增加对大气中凝结水、蒸汽和雾等的收集,有利于干旱区生物结皮表面的水分供给和微生物的活动[29 -30 ] .总体来讲,生物结皮对养分和水分的聚集,有助于荒漠生态系统正向演替. ...
Effects of microbiotic crusts on dew deposition in the restored vegetation area at Shapotou,northwest China
1
2006
... 生物结皮中的蓝藻、藓类等光合类群,能够固碳,促进有机质的形成和增加.全球尺度生物结皮年总固碳量可达0.6 Pg, 相当于全球陆地生态系统净生产力的1%,在荒漠区占比可达9%[8 ,23 ] .同时,生物结皮中具有大量的固氮细菌和蓝藻,全球年固氮量可达26 Tg,约占全球孢子植物固氮量的50%,以及陆地生态系统生物总固氮量的1/4[7 -8 ] .生物结皮中丰富的微生物可矿化和溶解土壤中不溶性无机磷和有机磷复合,促进磷的吸收[24 ] .因此,生物结皮斑块是干旱区特殊的肥岛,能够促进小尺度养分的聚集[25 -27 ] .除养分外,随着生物结皮的发育,生物结皮表面粗糙度增加,增加土壤水分入渗和留存[28 ] .与裸沙相比,生物结皮可降低水分蒸发,增加对大气中凝结水、蒸汽和雾等的收集,有利于干旱区生物结皮表面的水分供给和微生物的活动[29 -30 ] .总体来讲,生物结皮对养分和水分的聚集,有助于荒漠生态系统正向演替. ...
Relationships between the biomass of algal crusts in fields and their compressive strength
1
2007
... 生物结皮的藻丝对土壤颗粒的缠绕以及分泌的聚合物对阳离子的吸收,使得生物结皮具有较强的抗风蚀和抗压能力.随着生物结皮生物量的增加,抗风蚀能力增强[31 ] .研究发现,未经扰动的生物结皮,难以形成地表风蚀,随着生物结皮破损率的增加,风蚀强度显著增加[5 ,32 ] .在生物结皮形成过程中,不同物种组成和不同生物结皮类型抗蚀度不同,如不同物种中具鞘微鞘藻和席藻具有较强的抗风蚀能力,不同生物结皮类型中苔藓结皮抗风蚀能力较强[32 -33 ] .生物结皮的存在,减少土壤细沙损失的同时,使得碳氮等养分能够被固定在土壤中[34 -35 ] . ...
古尔班通古特沙漠生物结皮对地表风蚀作用影响的风洞实验
2
2004
... 生物结皮的藻丝对土壤颗粒的缠绕以及分泌的聚合物对阳离子的吸收,使得生物结皮具有较强的抗风蚀和抗压能力.随着生物结皮生物量的增加,抗风蚀能力增强[31 ] .研究发现,未经扰动的生物结皮,难以形成地表风蚀,随着生物结皮破损率的增加,风蚀强度显著增加[5 ,32 ] .在生物结皮形成过程中,不同物种组成和不同生物结皮类型抗蚀度不同,如不同物种中具鞘微鞘藻和席藻具有较强的抗风蚀能力,不同生物结皮类型中苔藓结皮抗风蚀能力较强[32 -33 ] .生物结皮的存在,减少土壤细沙损失的同时,使得碳氮等养分能够被固定在土壤中[34 -35 ] . ...
... [32 -33 ].生物结皮的存在,减少土壤细沙损失的同时,使得碳氮等养分能够被固定在土壤中[34 -35 ] . ...
Effect of desert soil algae on the stabilization of fine sands
1
2002
... 生物结皮的藻丝对土壤颗粒的缠绕以及分泌的聚合物对阳离子的吸收,使得生物结皮具有较强的抗风蚀和抗压能力.随着生物结皮生物量的增加,抗风蚀能力增强[31 ] .研究发现,未经扰动的生物结皮,难以形成地表风蚀,随着生物结皮破损率的增加,风蚀强度显著增加[5 ,32 ] .在生物结皮形成过程中,不同物种组成和不同生物结皮类型抗蚀度不同,如不同物种中具鞘微鞘藻和席藻具有较强的抗风蚀能力,不同生物结皮类型中苔藓结皮抗风蚀能力较强[32 -33 ] .生物结皮的存在,减少土壤细沙损失的同时,使得碳氮等养分能够被固定在土壤中[34 -35 ] . ...
Effects of wind erosion on the spatial heterogeneity of soil nutrients in two desert grassland communities
1
2008
... 生物结皮的藻丝对土壤颗粒的缠绕以及分泌的聚合物对阳离子的吸收,使得生物结皮具有较强的抗风蚀和抗压能力.随着生物结皮生物量的增加,抗风蚀能力增强[31 ] .研究发现,未经扰动的生物结皮,难以形成地表风蚀,随着生物结皮破损率的增加,风蚀强度显著增加[5 ,32 ] .在生物结皮形成过程中,不同物种组成和不同生物结皮类型抗蚀度不同,如不同物种中具鞘微鞘藻和席藻具有较强的抗风蚀能力,不同生物结皮类型中苔藓结皮抗风蚀能力较强[32 -33 ] .生物结皮的存在,减少土壤细沙损失的同时,使得碳氮等养分能够被固定在土壤中[34 -35 ] . ...
Multi-decadal impacts of grazing on soil physical and biogeochemical properties in southeast Utah
1
2005
... 生物结皮的藻丝对土壤颗粒的缠绕以及分泌的聚合物对阳离子的吸收,使得生物结皮具有较强的抗风蚀和抗压能力.随着生物结皮生物量的增加,抗风蚀能力增强[31 ] .研究发现,未经扰动的生物结皮,难以形成地表风蚀,随着生物结皮破损率的增加,风蚀强度显著增加[5 ,32 ] .在生物结皮形成过程中,不同物种组成和不同生物结皮类型抗蚀度不同,如不同物种中具鞘微鞘藻和席藻具有较强的抗风蚀能力,不同生物结皮类型中苔藓结皮抗风蚀能力较强[32 -33 ] .生物结皮的存在,减少土壤细沙损失的同时,使得碳氮等养分能够被固定在土壤中[34 -35 ] . ...
一种人工生物结皮治理荒漠的方法及专用地衣
3
... 生物结皮人工培养物种选择的重要原则是对本地生物结皮优势物种的筛选,不仅可以保障培养体具有较强的抗逆性和适应性,也可避免外来物种生物入侵风险.同时,筛选的目标物种应具有适应高温、高pH、高盐碱、强辐射等逆境生境的能力[21 ] .多数荒漠的藻类结皮中,以微鞘藻属(Microcoleus )种类分布最广,因此该属种类是藻类结皮恢复最重要的目标种.其他优势物种也常用于人工藻类结皮恢复,如库布齐沙漠选择席藻(Phormidium tenue )和伪枝藻(Scytonema javanicum ),腾格里沙漠选择念珠藻属(Nostoc )和席藻属(Phormidium )等.中国地衣结皮的培养仅见地衣石果衣(Endocarpon pusillum )报道,包括石果衣共生真菌和石果衣共生藻的分离培养[36 ] .中国各荒漠区藓类结皮优势物种不同,常见有黄土高原极地真藓(Bryum arcticum )、毛乌素沙地黄色真藓(Bryum pallescens )、腾格里沙漠银叶真藓(Bryum argenteum )、古尔班通古特沙漠齿肋赤藓(Syntrichia caninervis )等用于藓类结皮培养与恢复研究. ...
... 物种的分离和纯化主要针对藻类开展.野外采集藻类结皮样品,在室内利用BG11、BG110 、BBM等培养基进行培养.通过稀释平板法获得纯化物种并鉴定,鉴定方法包括传统显微镜法(形态)以及分子生物学测序(16S rDNA),选定该区域优势物种[9 ] .地衣取样时,选择成熟发育良好的地衣体,利用孢子释放法分离地衣共生菌和共生藻,获得单一孢子萌发形成的菌落和藻落,利用分子生物学方法鉴定共生菌和共生藻[36 ] .藓类个体较大,一般选择单一藓类通过形态学方法鉴定,并在室内扩繁. ...
... Study case of biological soil crust rehabilitation in China
Table 1 结皮类型 地点 规模 结皮生长 评价 文献 藻类结皮 库布齐沙漠 约200 hm2 2—3年形成3—5 mm藻类结皮,盖度48.5% 结皮和植被得到较好恢复 [9 ,38 ,51 ] 腾格里沙漠 2 m×2 m 5个月后,0.39 mm,盖度达14.3%,生物量13.4 mg·g-1 ;盖度或先增后降,1年后13.8% 长期效应未知 [12 ,15 ] 古尔班通古特沙漠 2 m×5 m 15 d可达2.6 mg·cm-2 (叶绿素a) 未形成大规模生物结皮 未发表 地衣结皮 腾格里沙漠 3个 0.5 m2 1年后形成4—6 cm生物结皮 长期效应未知 [36 ] 藓类结皮 腾格里沙漠 10个1 m×1 m 7 d内存活,3个月内死亡 接种藓类直接采自野外 [59 ] 毛乌素沙地 18个1 m×1 m 75 d后盖度70%,密度9.8 株·cm-2 ,4年盖度达98% 接种藓类直接采自野外 [45 ,57 ] 杨凌 48个1 m×1 m 30 d,盖度90%,最大密度120株·cm-2 接种藓类直接采自野外 [14 ] 库布齐沙漠 8个1 m×1 m 3年后,盖度30%—70%,185株·cm-2 接种藓类直接采自野外 [60 ]
<strong>3.4</strong> 人工培养生物结皮的抗性锻炼 为提高野外存活率,是否对扩繁后的材料进行模拟野外环境的抗性锻炼,具有不同见解.抗性锻炼主要包括干燥和水分循环、高温和低温循环、不同光强等,以增加培养物的适合度[59 -62 ] .通过对20个蓝藻菌株进行抗性锻炼发现,13个菌株均表现出适合度的增加,尤其是结皮先锋藻类微鞘藻属的菌株,适合度均得到提升[63 ] .但通过培养混合生物结皮(藻类+地衣+藓类),进行一系列抗性锻炼(全谱光+温度变化+干旱),发现抗性锻炼无显著效果[62 ] ,藓类结皮物种作用同样不显著[64 ] .对不同种类藓类样品在不同存储温度研究发现,不同藓类种类后期生长对储存温度的敏感性不一致[65 ] .因此,生物结皮在接种前,是否有必要进行抗性锻炼,有待进一步验证. ...
Cyanobacterial inoculation (cyanobacterisation): perspectives for the development of a standardized multifunctional technology for soil fertilization and desertification reversal
4
2017
... A—C,室内培养与扩大化生产; D,温室接种; E,野外藻类结皮接种
Mass cultivation of cyanobacterial crust and field inoculation in the Gurbantunggut Desert Fig.3 ![]()
图4 中国科学院水生生物研究所内蒙古实验基地建造的藻类培养的跑道池<sup>[<xref ref-type="bibr" rid="R37">37</xref>]</sup> The raceway pond operated by Institute of Hydrobiology, Chinese Academy of Sciences, in Inner-Mongolia<sup>[<xref ref-type="bibr" rid="R37">37</xref>]</sup> Fig.4 ![]()
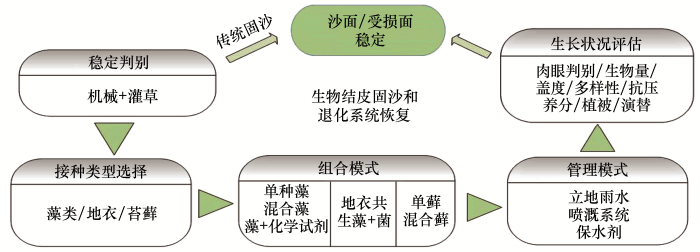
<strong>3 </strong>生物结皮培养物的野外固沙实践与生长状况评估 <strong>3.1</strong> 生物结皮的野外接种 生物结皮野外接种流程见图5 .在生物结皮接种前,采用草方格、立体栅栏等方法先行稳定沙面,改善生物结皮恢复初期地表条件.干藻种可与沙土混匀播撒到地表,藻悬浮液以雾化微喷的方式接种到沙面[9 ,37 ] .随着技术的进步,可利用无人机、飞机撒播等手段在更大空间尺度实现培养物的均匀喷洒.野外恢复的同时,也可辅助以微喷溉系统每天添加适量的水(20 mm·d-1 ),来保持藻类的生存活力[39 ] .虽然藓类结皮室内大规模扩繁报道较多[40 -41 ,45 ] ,但利用室内扩繁藓类成功进行野外恢复的研究鲜见报道.目前,有研究者直接采集自然藓类结皮植株,粉碎后在其他受损区域进行撒播,辅以一定的遮阳、覆膜、营养液等措施,以促进该区域藓类结皮恢复[14 ,45 -46 ] .这种藓类结皮恢复的存在较大的资源限制,如果大规模应用,将会给自然发育的藓类结皮造成巨大资源压力,并有可能造成新的退化区.为促进接种物更快生长、形成生物结皮,除添加营养液解除土壤养分限制外,有如下模式参考. ...
... [
37 ]
Fig.4 ![]()
<strong>3 </strong>生物结皮培养物的野外固沙实践与生长状况评估 <strong>3.1</strong> 生物结皮的野外接种 生物结皮野外接种流程见图5 .在生物结皮接种前,采用草方格、立体栅栏等方法先行稳定沙面,改善生物结皮恢复初期地表条件.干藻种可与沙土混匀播撒到地表,藻悬浮液以雾化微喷的方式接种到沙面[9 ,37 ] .随着技术的进步,可利用无人机、飞机撒播等手段在更大空间尺度实现培养物的均匀喷洒.野外恢复的同时,也可辅助以微喷溉系统每天添加适量的水(20 mm·d-1 ),来保持藻类的生存活力[39 ] .虽然藓类结皮室内大规模扩繁报道较多[40 -41 ,45 ] ,但利用室内扩繁藓类成功进行野外恢复的研究鲜见报道.目前,有研究者直接采集自然藓类结皮植株,粉碎后在其他受损区域进行撒播,辅以一定的遮阳、覆膜、营养液等措施,以促进该区域藓类结皮恢复[14 ,45 -46 ] .这种藓类结皮恢复的存在较大的资源限制,如果大规模应用,将会给自然发育的藓类结皮造成巨大资源压力,并有可能造成新的退化区.为促进接种物更快生长、形成生物结皮,除添加营养液解除土壤养分限制外,有如下模式参考. ...
... 生物结皮野外接种流程见图5 .在生物结皮接种前,采用草方格、立体栅栏等方法先行稳定沙面,改善生物结皮恢复初期地表条件.干藻种可与沙土混匀播撒到地表,藻悬浮液以雾化微喷的方式接种到沙面[9 ,37 ] .随着技术的进步,可利用无人机、飞机撒播等手段在更大空间尺度实现培养物的均匀喷洒.野外恢复的同时,也可辅助以微喷溉系统每天添加适量的水(20 mm·d-1 ),来保持藻类的生存活力[39 ] .虽然藓类结皮室内大规模扩繁报道较多[40 -41 ,45 ] ,但利用室内扩繁藓类成功进行野外恢复的研究鲜见报道.目前,有研究者直接采集自然藓类结皮植株,粉碎后在其他受损区域进行撒播,辅以一定的遮阳、覆膜、营养液等措施,以促进该区域藓类结皮恢复[14 ,45 -46 ] .这种藓类结皮恢复的存在较大的资源限制,如果大规模应用,将会给自然发育的藓类结皮造成巨大资源压力,并有可能造成新的退化区.为促进接种物更快生长、形成生物结皮,除添加营养液解除土壤养分限制外,有如下模式参考. ...
... 许多指标可用来指示人工生物结皮生长、恢复的状况,能够实现自身持续的可支持系统,是其恢复成功的重要标志[37 ] .从生物结皮本身的角度来看,形成肉眼可见的藻类结皮是一个重要的标志[38 ] .以生物结皮生物量表征恢复实验的效果,可直接反映生物结皮的生长状况,如通过对藻类结皮的2—3年人工培养,叶绿素含量可达35 μg·cm-2[39 ,51 ] .人工生物结皮的抗压强度和生物结皮厚度能够指示其固沙能力的强弱,随着培育时间的推移,其抗压强度得到显著增加,3年后可达3.83 kg·cm-2[38 ] .对于地衣结皮,通过扫描电镜观察生物结皮菌藻复合物,可发现石果衣菌丝向土壤基质下部延伸6—8 mm[40 ] .对于藓类结皮,盖度和密度常用来指示一定面积的生物结皮恢复,如培养65 d后,盖度可达83%,密度可达122株·cm-2[42 ] .从环境改善的角度,人工生物结皮的发育可促进土壤有机碳、多糖、总氮等养分和土壤酶活性的增加[40 ,52 -53 ] .人工生物结皮区沙生植物得到拓殖和生长,植物群落演替加速,同时土壤微生物数量也大量增加,固沙能力增强[38 ,51 ,54 ] . ...
Feasibility of cyanobacterial inoculation for biological soil crusts formation in desert area
7
2009
... 目前,藻类和藓类的大规模室内培养和生产技术较为成熟,但地衣培养过程十分复杂,未见大规模室内培养报道.藻类培养的温度一般20—300 ,需要一定的光照,并且通过不断搅拌等方式供给氧气.在藻种的初级扩繁阶段,通常将试管原种接种到试管固体培养基上,在培养箱内(光强约2 000 lx,温度25—28 ℃)培养10—15 d后,将固体培养基培养的藻种进行下一级培养,接种入100—500 mL三角瓶中,同样条件下静置或通气培养8—10 d,进入扩大化培养阶段[9 ] .同时,也有将藻种悬浮液直接接入三角瓶中,摇床培养(光强约600 lx,温度25—30 ℃)7—10 d后,转为下一级(约60 L)培养的报道[15 ] .中国科学院新疆生态与地理研究所团队选择古尔班通古特沙漠优势物种,分离和培养,实现了接种藻类小规模的生产(图3 ).中国科学院武汉水生生物研究所团队在规模化藻类培养方面取得突破,通过藻类纯化、试管培养、三角瓶培养、10—18 L容器培养、小跑道池(1 m×4 m)和大跑道池(6 m×40 m)培养等步骤(图4 ),能够供给大量的藻类接种物[9 ] ,每步骤时间5—15 d[38 -39 ] .除了单种培养外,中国科学院沙坡头沙漠研究试验站和新疆生态与地理研究所均尝试利用生物结皮进行混合藻类的培养和规模化生产,此法虽然产量未及单种培养,但操作相对简单高效,也成为大规模培养的潜在选择[15 ] .地衣在室内需要经过固体、液体培养基接种、转化,共生藻和共生菌在不同的条件进行培养[40 ] ,但短期内并不能形成地衣植物.藓类植物的孢子体、配子体、茎叶碎片和芽孢等均可作为人工培养的材料[41 ] .组织培养时,可将配子体部分放入knop营养液,摇床培养,20 d后取出,自然风干待用[15 ] .组织培养的缺点是需要对外殖体进行灭菌消毒等,易对藓类本身造成损害.目前使用较多的为野外采集藓类植物的碎片作为材料进行接种,可实现低成本小规模培养[42 -43 ] .藓类培养时需注意后期水分的控制,干旱和完全湿润条件均不利于藓类植物的生长[44 ] .综合各研究案例可知,同一生物结皮类型中不同物种的最适培养条件不同,各研究团队采用的培养方法也具有差异性,但流程基本一致.在培养过程中,需要注意条件控制,如温度过高将导致培养目标藻类死亡或者其他污染藻类生长[9 ] . ...
... 从接种藻类物种组成角度,可分成3种,选择接种区域的优势藻种:①单种培养并单种施加,如利用具鞘微鞘藻(Microcoleus vaginatus )进行培养和单独接种[39 ] ;②多种分开培养,接种前混合,如将具鞘微鞘藻和爪哇伪枝藻(S. javanicum )按干物质重10∶1进行混合接种[38 ] ,或者具鞘微鞘藻、席藻和爪哇伪枝藻按10∶5∶1混合接种;③混合培养、混合接种,如腾格里沙漠的部分实验[15 ] ,将藻类从结皮中直接分离培养,不纯化,节约了时间、技术和经济成本. ...
... 从机械和生物模式组合角度,生物结皮野外接种初期,一些稳定的措施,包括物理措施(如草方格)和生物措施(草本、灌丛和乔木)的组合,将有助于生物结皮的发育[38 ,47 ] .Chen等[39 ] 研究表明,适度的降水和降低光强的措施有助于藻类结皮的快速生长.通过藻-草-灌-乔四位一体的模式,在库布齐沙漠的大规模流沙治理中得到了应用[9 ] . ...
... 许多指标可用来指示人工生物结皮生长、恢复的状况,能够实现自身持续的可支持系统,是其恢复成功的重要标志[37 ] .从生物结皮本身的角度来看,形成肉眼可见的藻类结皮是一个重要的标志[38 ] .以生物结皮生物量表征恢复实验的效果,可直接反映生物结皮的生长状况,如通过对藻类结皮的2—3年人工培养,叶绿素含量可达35 μg·cm-2[39 ,51 ] .人工生物结皮的抗压强度和生物结皮厚度能够指示其固沙能力的强弱,随着培育时间的推移,其抗压强度得到显著增加,3年后可达3.83 kg·cm-2[38 ] .对于地衣结皮,通过扫描电镜观察生物结皮菌藻复合物,可发现石果衣菌丝向土壤基质下部延伸6—8 mm[40 ] .对于藓类结皮,盖度和密度常用来指示一定面积的生物结皮恢复,如培养65 d后,盖度可达83%,密度可达122株·cm-2[42 ] .从环境改善的角度,人工生物结皮的发育可促进土壤有机碳、多糖、总氮等养分和土壤酶活性的增加[40 ,52 -53 ] .人工生物结皮区沙生植物得到拓殖和生长,植物群落演替加速,同时土壤微生物数量也大量增加,固沙能力增强[38 ,51 ,54 ] . ...
... -2[38 ].对于地衣结皮,通过扫描电镜观察生物结皮菌藻复合物,可发现石果衣菌丝向土壤基质下部延伸6—8 mm[40 ] .对于藓类结皮,盖度和密度常用来指示一定面积的生物结皮恢复,如培养65 d后,盖度可达83%,密度可达122株·cm-2[42 ] .从环境改善的角度,人工生物结皮的发育可促进土壤有机碳、多糖、总氮等养分和土壤酶活性的增加[40 ,52 -53 ] .人工生物结皮区沙生植物得到拓殖和生长,植物群落演替加速,同时土壤微生物数量也大量增加,固沙能力增强[38 ,51 ,54 ] . ...
... [38 ,51 ,54 ]. ...
... Study case of biological soil crust rehabilitation in China
Table 1 结皮类型 地点 规模 结皮生长 评价 文献 藻类结皮 库布齐沙漠 约200 hm2 2—3年形成3—5 mm藻类结皮,盖度48.5% 结皮和植被得到较好恢复 [9 ,38 ,51 ] 腾格里沙漠 2 m×2 m 5个月后,0.39 mm,盖度达14.3%,生物量13.4 mg·g-1 ;盖度或先增后降,1年后13.8% 长期效应未知 [12 ,15 ] 古尔班通古特沙漠 2 m×5 m 15 d可达2.6 mg·cm-2 (叶绿素a) 未形成大规模生物结皮 未发表 地衣结皮 腾格里沙漠 3个 0.5 m2 1年后形成4—6 cm生物结皮 长期效应未知 [36 ] 藓类结皮 腾格里沙漠 10个1 m×1 m 7 d内存活,3个月内死亡 接种藓类直接采自野外 [59 ] 毛乌素沙地 18个1 m×1 m 75 d后盖度70%,密度9.8 株·cm-2 ,4年盖度达98% 接种藓类直接采自野外 [45 ,57 ] 杨凌 48个1 m×1 m 30 d,盖度90%,最大密度120株·cm-2 接种藓类直接采自野外 [14 ] 库布齐沙漠 8个1 m×1 m 3年后,盖度30%—70%,185株·cm-2 接种藓类直接采自野外 [60 ]
<strong>3.4</strong> 人工培养生物结皮的抗性锻炼 为提高野外存活率,是否对扩繁后的材料进行模拟野外环境的抗性锻炼,具有不同见解.抗性锻炼主要包括干燥和水分循环、高温和低温循环、不同光强等,以增加培养物的适合度[59 -62 ] .通过对20个蓝藻菌株进行抗性锻炼发现,13个菌株均表现出适合度的增加,尤其是结皮先锋藻类微鞘藻属的菌株,适合度均得到提升[63 ] .但通过培养混合生物结皮(藻类+地衣+藓类),进行一系列抗性锻炼(全谱光+温度变化+干旱),发现抗性锻炼无显著效果[62 ] ,藓类结皮物种作用同样不显著[64 ] .对不同种类藓类样品在不同存储温度研究发现,不同藓类种类后期生长对储存温度的敏感性不一致[65 ] .因此,生物结皮在接种前,是否有必要进行抗性锻炼,有待进一步验证. ...
Man-made desert algal crusts as affected by environmental factors in Inner Mongolia,China
5
2006
... 目前,藻类和藓类的大规模室内培养和生产技术较为成熟,但地衣培养过程十分复杂,未见大规模室内培养报道.藻类培养的温度一般20—300 ,需要一定的光照,并且通过不断搅拌等方式供给氧气.在藻种的初级扩繁阶段,通常将试管原种接种到试管固体培养基上,在培养箱内(光强约2 000 lx,温度25—28 ℃)培养10—15 d后,将固体培养基培养的藻种进行下一级培养,接种入100—500 mL三角瓶中,同样条件下静置或通气培养8—10 d,进入扩大化培养阶段[9 ] .同时,也有将藻种悬浮液直接接入三角瓶中,摇床培养(光强约600 lx,温度25—30 ℃)7—10 d后,转为下一级(约60 L)培养的报道[15 ] .中国科学院新疆生态与地理研究所团队选择古尔班通古特沙漠优势物种,分离和培养,实现了接种藻类小规模的生产(图3 ).中国科学院武汉水生生物研究所团队在规模化藻类培养方面取得突破,通过藻类纯化、试管培养、三角瓶培养、10—18 L容器培养、小跑道池(1 m×4 m)和大跑道池(6 m×40 m)培养等步骤(图4 ),能够供给大量的藻类接种物[9 ] ,每步骤时间5—15 d[38 -39 ] .除了单种培养外,中国科学院沙坡头沙漠研究试验站和新疆生态与地理研究所均尝试利用生物结皮进行混合藻类的培养和规模化生产,此法虽然产量未及单种培养,但操作相对简单高效,也成为大规模培养的潜在选择[15 ] .地衣在室内需要经过固体、液体培养基接种、转化,共生藻和共生菌在不同的条件进行培养[40 ] ,但短期内并不能形成地衣植物.藓类植物的孢子体、配子体、茎叶碎片和芽孢等均可作为人工培养的材料[41 ] .组织培养时,可将配子体部分放入knop营养液,摇床培养,20 d后取出,自然风干待用[15 ] .组织培养的缺点是需要对外殖体进行灭菌消毒等,易对藓类本身造成损害.目前使用较多的为野外采集藓类植物的碎片作为材料进行接种,可实现低成本小规模培养[42 -43 ] .藓类培养时需注意后期水分的控制,干旱和完全湿润条件均不利于藓类植物的生长[44 ] .综合各研究案例可知,同一生物结皮类型中不同物种的最适培养条件不同,各研究团队采用的培养方法也具有差异性,但流程基本一致.在培养过程中,需要注意条件控制,如温度过高将导致培养目标藻类死亡或者其他污染藻类生长[9 ] . ...
... 生物结皮野外接种流程见图5 .在生物结皮接种前,采用草方格、立体栅栏等方法先行稳定沙面,改善生物结皮恢复初期地表条件.干藻种可与沙土混匀播撒到地表,藻悬浮液以雾化微喷的方式接种到沙面[9 ,37 ] .随着技术的进步,可利用无人机、飞机撒播等手段在更大空间尺度实现培养物的均匀喷洒.野外恢复的同时,也可辅助以微喷溉系统每天添加适量的水(20 mm·d-1 ),来保持藻类的生存活力[39 ] .虽然藓类结皮室内大规模扩繁报道较多[40 -41 ,45 ] ,但利用室内扩繁藓类成功进行野外恢复的研究鲜见报道.目前,有研究者直接采集自然藓类结皮植株,粉碎后在其他受损区域进行撒播,辅以一定的遮阳、覆膜、营养液等措施,以促进该区域藓类结皮恢复[14 ,45 -46 ] .这种藓类结皮恢复的存在较大的资源限制,如果大规模应用,将会给自然发育的藓类结皮造成巨大资源压力,并有可能造成新的退化区.为促进接种物更快生长、形成生物结皮,除添加营养液解除土壤养分限制外,有如下模式参考. ...
... 从接种藻类物种组成角度,可分成3种,选择接种区域的优势藻种:①单种培养并单种施加,如利用具鞘微鞘藻(Microcoleus vaginatus )进行培养和单独接种[39 ] ;②多种分开培养,接种前混合,如将具鞘微鞘藻和爪哇伪枝藻(S. javanicum )按干物质重10∶1进行混合接种[38 ] ,或者具鞘微鞘藻、席藻和爪哇伪枝藻按10∶5∶1混合接种;③混合培养、混合接种,如腾格里沙漠的部分实验[15 ] ,将藻类从结皮中直接分离培养,不纯化,节约了时间、技术和经济成本. ...
... 从机械和生物模式组合角度,生物结皮野外接种初期,一些稳定的措施,包括物理措施(如草方格)和生物措施(草本、灌丛和乔木)的组合,将有助于生物结皮的发育[38 ,47 ] .Chen等[39 ] 研究表明,适度的降水和降低光强的措施有助于藻类结皮的快速生长.通过藻-草-灌-乔四位一体的模式,在库布齐沙漠的大规模流沙治理中得到了应用[9 ] . ...
... 许多指标可用来指示人工生物结皮生长、恢复的状况,能够实现自身持续的可支持系统,是其恢复成功的重要标志[37 ] .从生物结皮本身的角度来看,形成肉眼可见的藻类结皮是一个重要的标志[38 ] .以生物结皮生物量表征恢复实验的效果,可直接反映生物结皮的生长状况,如通过对藻类结皮的2—3年人工培养,叶绿素含量可达35 μg·cm-2[39 ,51 ] .人工生物结皮的抗压强度和生物结皮厚度能够指示其固沙能力的强弱,随着培育时间的推移,其抗压强度得到显著增加,3年后可达3.83 kg·cm-2[38 ] .对于地衣结皮,通过扫描电镜观察生物结皮菌藻复合物,可发现石果衣菌丝向土壤基质下部延伸6—8 mm[40 ] .对于藓类结皮,盖度和密度常用来指示一定面积的生物结皮恢复,如培养65 d后,盖度可达83%,密度可达122株·cm-2[42 ] .从环境改善的角度,人工生物结皮的发育可促进土壤有机碳、多糖、总氮等养分和土壤酶活性的增加[40 ,52 -53 ] .人工生物结皮区沙生植物得到拓殖和生长,植物群落演替加速,同时土壤微生物数量也大量增加,固沙能力增强[38 ,51 ,54 ] . ...
沙漠地衣结皮快速繁殖技术探索.多彩菌物 美丽中国
4
2019
... 目前,藻类和藓类的大规模室内培养和生产技术较为成熟,但地衣培养过程十分复杂,未见大规模室内培养报道.藻类培养的温度一般20—300 ,需要一定的光照,并且通过不断搅拌等方式供给氧气.在藻种的初级扩繁阶段,通常将试管原种接种到试管固体培养基上,在培养箱内(光强约2 000 lx,温度25—28 ℃)培养10—15 d后,将固体培养基培养的藻种进行下一级培养,接种入100—500 mL三角瓶中,同样条件下静置或通气培养8—10 d,进入扩大化培养阶段[9 ] .同时,也有将藻种悬浮液直接接入三角瓶中,摇床培养(光强约600 lx,温度25—30 ℃)7—10 d后,转为下一级(约60 L)培养的报道[15 ] .中国科学院新疆生态与地理研究所团队选择古尔班通古特沙漠优势物种,分离和培养,实现了接种藻类小规模的生产(图3 ).中国科学院武汉水生生物研究所团队在规模化藻类培养方面取得突破,通过藻类纯化、试管培养、三角瓶培养、10—18 L容器培养、小跑道池(1 m×4 m)和大跑道池(6 m×40 m)培养等步骤(图4 ),能够供给大量的藻类接种物[9 ] ,每步骤时间5—15 d[38 -39 ] .除了单种培养外,中国科学院沙坡头沙漠研究试验站和新疆生态与地理研究所均尝试利用生物结皮进行混合藻类的培养和规模化生产,此法虽然产量未及单种培养,但操作相对简单高效,也成为大规模培养的潜在选择[15 ] .地衣在室内需要经过固体、液体培养基接种、转化,共生藻和共生菌在不同的条件进行培养[40 ] ,但短期内并不能形成地衣植物.藓类植物的孢子体、配子体、茎叶碎片和芽孢等均可作为人工培养的材料[41 ] .组织培养时,可将配子体部分放入knop营养液,摇床培养,20 d后取出,自然风干待用[15 ] .组织培养的缺点是需要对外殖体进行灭菌消毒等,易对藓类本身造成损害.目前使用较多的为野外采集藓类植物的碎片作为材料进行接种,可实现低成本小规模培养[42 -43 ] .藓类培养时需注意后期水分的控制,干旱和完全湿润条件均不利于藓类植物的生长[44 ] .综合各研究案例可知,同一生物结皮类型中不同物种的最适培养条件不同,各研究团队采用的培养方法也具有差异性,但流程基本一致.在培养过程中,需要注意条件控制,如温度过高将导致培养目标藻类死亡或者其他污染藻类生长[9 ] . ...
... 生物结皮野外接种流程见图5 .在生物结皮接种前,采用草方格、立体栅栏等方法先行稳定沙面,改善生物结皮恢复初期地表条件.干藻种可与沙土混匀播撒到地表,藻悬浮液以雾化微喷的方式接种到沙面[9 ,37 ] .随着技术的进步,可利用无人机、飞机撒播等手段在更大空间尺度实现培养物的均匀喷洒.野外恢复的同时,也可辅助以微喷溉系统每天添加适量的水(20 mm·d-1 ),来保持藻类的生存活力[39 ] .虽然藓类结皮室内大规模扩繁报道较多[40 -41 ,45 ] ,但利用室内扩繁藓类成功进行野外恢复的研究鲜见报道.目前,有研究者直接采集自然藓类结皮植株,粉碎后在其他受损区域进行撒播,辅以一定的遮阳、覆膜、营养液等措施,以促进该区域藓类结皮恢复[14 ,45 -46 ] .这种藓类结皮恢复的存在较大的资源限制,如果大规模应用,将会给自然发育的藓类结皮造成巨大资源压力,并有可能造成新的退化区.为促进接种物更快生长、形成生物结皮,除添加营养液解除土壤养分限制外,有如下模式参考. ...
... 许多指标可用来指示人工生物结皮生长、恢复的状况,能够实现自身持续的可支持系统,是其恢复成功的重要标志[37 ] .从生物结皮本身的角度来看,形成肉眼可见的藻类结皮是一个重要的标志[38 ] .以生物结皮生物量表征恢复实验的效果,可直接反映生物结皮的生长状况,如通过对藻类结皮的2—3年人工培养,叶绿素含量可达35 μg·cm-2[39 ,51 ] .人工生物结皮的抗压强度和生物结皮厚度能够指示其固沙能力的强弱,随着培育时间的推移,其抗压强度得到显著增加,3年后可达3.83 kg·cm-2[38 ] .对于地衣结皮,通过扫描电镜观察生物结皮菌藻复合物,可发现石果衣菌丝向土壤基质下部延伸6—8 mm[40 ] .对于藓类结皮,盖度和密度常用来指示一定面积的生物结皮恢复,如培养65 d后,盖度可达83%,密度可达122株·cm-2[42 ] .从环境改善的角度,人工生物结皮的发育可促进土壤有机碳、多糖、总氮等养分和土壤酶活性的增加[40 ,52 -53 ] .人工生物结皮区沙生植物得到拓殖和生长,植物群落演替加速,同时土壤微生物数量也大量增加,固沙能力增强[38 ,51 ,54 ] . ...
... [40 ,52 -53 ].人工生物结皮区沙生植物得到拓殖和生长,植物群落演替加速,同时土壤微生物数量也大量增加,固沙能力增强[38 ,51 ,54 ] . ...
苔藓植物组织培养繁殖技术及其应用前景
2
2005
... 目前,藻类和藓类的大规模室内培养和生产技术较为成熟,但地衣培养过程十分复杂,未见大规模室内培养报道.藻类培养的温度一般20—300 ,需要一定的光照,并且通过不断搅拌等方式供给氧气.在藻种的初级扩繁阶段,通常将试管原种接种到试管固体培养基上,在培养箱内(光强约2 000 lx,温度25—28 ℃)培养10—15 d后,将固体培养基培养的藻种进行下一级培养,接种入100—500 mL三角瓶中,同样条件下静置或通气培养8—10 d,进入扩大化培养阶段[9 ] .同时,也有将藻种悬浮液直接接入三角瓶中,摇床培养(光强约600 lx,温度25—30 ℃)7—10 d后,转为下一级(约60 L)培养的报道[15 ] .中国科学院新疆生态与地理研究所团队选择古尔班通古特沙漠优势物种,分离和培养,实现了接种藻类小规模的生产(图3 ).中国科学院武汉水生生物研究所团队在规模化藻类培养方面取得突破,通过藻类纯化、试管培养、三角瓶培养、10—18 L容器培养、小跑道池(1 m×4 m)和大跑道池(6 m×40 m)培养等步骤(图4 ),能够供给大量的藻类接种物[9 ] ,每步骤时间5—15 d[38 -39 ] .除了单种培养外,中国科学院沙坡头沙漠研究试验站和新疆生态与地理研究所均尝试利用生物结皮进行混合藻类的培养和规模化生产,此法虽然产量未及单种培养,但操作相对简单高效,也成为大规模培养的潜在选择[15 ] .地衣在室内需要经过固体、液体培养基接种、转化,共生藻和共生菌在不同的条件进行培养[40 ] ,但短期内并不能形成地衣植物.藓类植物的孢子体、配子体、茎叶碎片和芽孢等均可作为人工培养的材料[41 ] .组织培养时,可将配子体部分放入knop营养液,摇床培养,20 d后取出,自然风干待用[15 ] .组织培养的缺点是需要对外殖体进行灭菌消毒等,易对藓类本身造成损害.目前使用较多的为野外采集藓类植物的碎片作为材料进行接种,可实现低成本小规模培养[42 -43 ] .藓类培养时需注意后期水分的控制,干旱和完全湿润条件均不利于藓类植物的生长[44 ] .综合各研究案例可知,同一生物结皮类型中不同物种的最适培养条件不同,各研究团队采用的培养方法也具有差异性,但流程基本一致.在培养过程中,需要注意条件控制,如温度过高将导致培养目标藻类死亡或者其他污染藻类生长[9 ] . ...
... 生物结皮野外接种流程见图5 .在生物结皮接种前,采用草方格、立体栅栏等方法先行稳定沙面,改善生物结皮恢复初期地表条件.干藻种可与沙土混匀播撒到地表,藻悬浮液以雾化微喷的方式接种到沙面[9 ,37 ] .随着技术的进步,可利用无人机、飞机撒播等手段在更大空间尺度实现培养物的均匀喷洒.野外恢复的同时,也可辅助以微喷溉系统每天添加适量的水(20 mm·d-1 ),来保持藻类的生存活力[39 ] .虽然藓类结皮室内大规模扩繁报道较多[40 -41 ,45 ] ,但利用室内扩繁藓类成功进行野外恢复的研究鲜见报道.目前,有研究者直接采集自然藓类结皮植株,粉碎后在其他受损区域进行撒播,辅以一定的遮阳、覆膜、营养液等措施,以促进该区域藓类结皮恢复[14 ,45 -46 ] .这种藓类结皮恢复的存在较大的资源限制,如果大规模应用,将会给自然发育的藓类结皮造成巨大资源压力,并有可能造成新的退化区.为促进接种物更快生长、形成生物结皮,除添加营养液解除土壤养分限制外,有如下模式参考. ...
黄土高原苔藓结皮的快速培育及其对逆境的生理响应研究
2
2015
... 目前,藻类和藓类的大规模室内培养和生产技术较为成熟,但地衣培养过程十分复杂,未见大规模室内培养报道.藻类培养的温度一般20—300 ,需要一定的光照,并且通过不断搅拌等方式供给氧气.在藻种的初级扩繁阶段,通常将试管原种接种到试管固体培养基上,在培养箱内(光强约2 000 lx,温度25—28 ℃)培养10—15 d后,将固体培养基培养的藻种进行下一级培养,接种入100—500 mL三角瓶中,同样条件下静置或通气培养8—10 d,进入扩大化培养阶段[9 ] .同时,也有将藻种悬浮液直接接入三角瓶中,摇床培养(光强约600 lx,温度25—30 ℃)7—10 d后,转为下一级(约60 L)培养的报道[15 ] .中国科学院新疆生态与地理研究所团队选择古尔班通古特沙漠优势物种,分离和培养,实现了接种藻类小规模的生产(图3 ).中国科学院武汉水生生物研究所团队在规模化藻类培养方面取得突破,通过藻类纯化、试管培养、三角瓶培养、10—18 L容器培养、小跑道池(1 m×4 m)和大跑道池(6 m×40 m)培养等步骤(图4 ),能够供给大量的藻类接种物[9 ] ,每步骤时间5—15 d[38 -39 ] .除了单种培养外,中国科学院沙坡头沙漠研究试验站和新疆生态与地理研究所均尝试利用生物结皮进行混合藻类的培养和规模化生产,此法虽然产量未及单种培养,但操作相对简单高效,也成为大规模培养的潜在选择[15 ] .地衣在室内需要经过固体、液体培养基接种、转化,共生藻和共生菌在不同的条件进行培养[40 ] ,但短期内并不能形成地衣植物.藓类植物的孢子体、配子体、茎叶碎片和芽孢等均可作为人工培养的材料[41 ] .组织培养时,可将配子体部分放入knop营养液,摇床培养,20 d后取出,自然风干待用[15 ] .组织培养的缺点是需要对外殖体进行灭菌消毒等,易对藓类本身造成损害.目前使用较多的为野外采集藓类植物的碎片作为材料进行接种,可实现低成本小规模培养[42 -43 ] .藓类培养时需注意后期水分的控制,干旱和完全湿润条件均不利于藓类植物的生长[44 ] .综合各研究案例可知,同一生物结皮类型中不同物种的最适培养条件不同,各研究团队采用的培养方法也具有差异性,但流程基本一致.在培养过程中,需要注意条件控制,如温度过高将导致培养目标藻类死亡或者其他污染藻类生长[9 ] . ...
... 许多指标可用来指示人工生物结皮生长、恢复的状况,能够实现自身持续的可支持系统,是其恢复成功的重要标志[37 ] .从生物结皮本身的角度来看,形成肉眼可见的藻类结皮是一个重要的标志[38 ] .以生物结皮生物量表征恢复实验的效果,可直接反映生物结皮的生长状况,如通过对藻类结皮的2—3年人工培养,叶绿素含量可达35 μg·cm-2[39 ,51 ] .人工生物结皮的抗压强度和生物结皮厚度能够指示其固沙能力的强弱,随着培育时间的推移,其抗压强度得到显著增加,3年后可达3.83 kg·cm-2[38 ] .对于地衣结皮,通过扫描电镜观察生物结皮菌藻复合物,可发现石果衣菌丝向土壤基质下部延伸6—8 mm[40 ] .对于藓类结皮,盖度和密度常用来指示一定面积的生物结皮恢复,如培养65 d后,盖度可达83%,密度可达122株·cm-2[42 ] .从环境改善的角度,人工生物结皮的发育可促进土壤有机碳、多糖、总氮等养分和土壤酶活性的增加[40 ,52 -53 ] .人工生物结皮区沙生植物得到拓殖和生长,植物群落演替加速,同时土壤微生物数量也大量增加,固沙能力增强[38 ,51 ,54 ] . ...
黄土丘陵区藓结皮人工培养方法试验研究
1
2009
... 目前,藻类和藓类的大规模室内培养和生产技术较为成熟,但地衣培养过程十分复杂,未见大规模室内培养报道.藻类培养的温度一般20—300 ,需要一定的光照,并且通过不断搅拌等方式供给氧气.在藻种的初级扩繁阶段,通常将试管原种接种到试管固体培养基上,在培养箱内(光强约2 000 lx,温度25—28 ℃)培养10—15 d后,将固体培养基培养的藻种进行下一级培养,接种入100—500 mL三角瓶中,同样条件下静置或通气培养8—10 d,进入扩大化培养阶段[9 ] .同时,也有将藻种悬浮液直接接入三角瓶中,摇床培养(光强约600 lx,温度25—30 ℃)7—10 d后,转为下一级(约60 L)培养的报道[15 ] .中国科学院新疆生态与地理研究所团队选择古尔班通古特沙漠优势物种,分离和培养,实现了接种藻类小规模的生产(图3 ).中国科学院武汉水生生物研究所团队在规模化藻类培养方面取得突破,通过藻类纯化、试管培养、三角瓶培养、10—18 L容器培养、小跑道池(1 m×4 m)和大跑道池(6 m×40 m)培养等步骤(图4 ),能够供给大量的藻类接种物[9 ] ,每步骤时间5—15 d[38 -39 ] .除了单种培养外,中国科学院沙坡头沙漠研究试验站和新疆生态与地理研究所均尝试利用生物结皮进行混合藻类的培养和规模化生产,此法虽然产量未及单种培养,但操作相对简单高效,也成为大规模培养的潜在选择[15 ] .地衣在室内需要经过固体、液体培养基接种、转化,共生藻和共生菌在不同的条件进行培养[40 ] ,但短期内并不能形成地衣植物.藓类植物的孢子体、配子体、茎叶碎片和芽孢等均可作为人工培养的材料[41 ] .组织培养时,可将配子体部分放入knop营养液,摇床培养,20 d后取出,自然风干待用[15 ] .组织培养的缺点是需要对外殖体进行灭菌消毒等,易对藓类本身造成损害.目前使用较多的为野外采集藓类植物的碎片作为材料进行接种,可实现低成本小规模培养[42 -43 ] .藓类培养时需注意后期水分的控制,干旱和完全湿润条件均不利于藓类植物的生长[44 ] .综合各研究案例可知,同一生物结皮类型中不同物种的最适培养条件不同,各研究团队采用的培养方法也具有差异性,但流程基本一致.在培养过程中,需要注意条件控制,如温度过高将导致培养目标藻类死亡或者其他污染藻类生长[9 ] . ...
水分条件对人工培养齿肋赤藓光化学效率及生理特性的影响
1
2017
... 目前,藻类和藓类的大规模室内培养和生产技术较为成熟,但地衣培养过程十分复杂,未见大规模室内培养报道.藻类培养的温度一般20—300 ,需要一定的光照,并且通过不断搅拌等方式供给氧气.在藻种的初级扩繁阶段,通常将试管原种接种到试管固体培养基上,在培养箱内(光强约2 000 lx,温度25—28 ℃)培养10—15 d后,将固体培养基培养的藻种进行下一级培养,接种入100—500 mL三角瓶中,同样条件下静置或通气培养8—10 d,进入扩大化培养阶段[9 ] .同时,也有将藻种悬浮液直接接入三角瓶中,摇床培养(光强约600 lx,温度25—30 ℃)7—10 d后,转为下一级(约60 L)培养的报道[15 ] .中国科学院新疆生态与地理研究所团队选择古尔班通古特沙漠优势物种,分离和培养,实现了接种藻类小规模的生产(图3 ).中国科学院武汉水生生物研究所团队在规模化藻类培养方面取得突破,通过藻类纯化、试管培养、三角瓶培养、10—18 L容器培养、小跑道池(1 m×4 m)和大跑道池(6 m×40 m)培养等步骤(图4 ),能够供给大量的藻类接种物[9 ] ,每步骤时间5—15 d[38 -39 ] .除了单种培养外,中国科学院沙坡头沙漠研究试验站和新疆生态与地理研究所均尝试利用生物结皮进行混合藻类的培养和规模化生产,此法虽然产量未及单种培养,但操作相对简单高效,也成为大规模培养的潜在选择[15 ] .地衣在室内需要经过固体、液体培养基接种、转化,共生藻和共生菌在不同的条件进行培养[40 ] ,但短期内并不能形成地衣植物.藓类植物的孢子体、配子体、茎叶碎片和芽孢等均可作为人工培养的材料[41 ] .组织培养时,可将配子体部分放入knop营养液,摇床培养,20 d后取出,自然风干待用[15 ] .组织培养的缺点是需要对外殖体进行灭菌消毒等,易对藓类本身造成损害.目前使用较多的为野外采集藓类植物的碎片作为材料进行接种,可实现低成本小规模培养[42 -43 ] .藓类培养时需注意后期水分的控制,干旱和完全湿润条件均不利于藓类植物的生长[44 ] .综合各研究案例可知,同一生物结皮类型中不同物种的最适培养条件不同,各研究团队采用的培养方法也具有差异性,但流程基本一致.在培养过程中,需要注意条件控制,如温度过高将导致培养目标藻类死亡或者其他污染藻类生长[9 ] . ...
Development of artificial moss-dominated biological soil crusts and their effects on runoff and soil water content in a semi-arid environment
3
2015
... 生物结皮野外接种流程见图5 .在生物结皮接种前,采用草方格、立体栅栏等方法先行稳定沙面,改善生物结皮恢复初期地表条件.干藻种可与沙土混匀播撒到地表,藻悬浮液以雾化微喷的方式接种到沙面[9 ,37 ] .随着技术的进步,可利用无人机、飞机撒播等手段在更大空间尺度实现培养物的均匀喷洒.野外恢复的同时,也可辅助以微喷溉系统每天添加适量的水(20 mm·d-1 ),来保持藻类的生存活力[39 ] .虽然藓类结皮室内大规模扩繁报道较多[40 -41 ,45 ] ,但利用室内扩繁藓类成功进行野外恢复的研究鲜见报道.目前,有研究者直接采集自然藓类结皮植株,粉碎后在其他受损区域进行撒播,辅以一定的遮阳、覆膜、营养液等措施,以促进该区域藓类结皮恢复[14 ,45 -46 ] .这种藓类结皮恢复的存在较大的资源限制,如果大规模应用,将会给自然发育的藓类结皮造成巨大资源压力,并有可能造成新的退化区.为促进接种物更快生长、形成生物结皮,除添加营养液解除土壤养分限制外,有如下模式参考. ...
... ,45 -46 ].这种藓类结皮恢复的存在较大的资源限制,如果大规模应用,将会给自然发育的藓类结皮造成巨大资源压力,并有可能造成新的退化区.为促进接种物更快生长、形成生物结皮,除添加营养液解除土壤养分限制外,有如下模式参考. ...
... Study case of biological soil crust rehabilitation in China
Table 1 结皮类型 地点 规模 结皮生长 评价 文献 藻类结皮 库布齐沙漠 约200 hm2 2—3年形成3—5 mm藻类结皮,盖度48.5% 结皮和植被得到较好恢复 [9 ,38 ,51 ] 腾格里沙漠 2 m×2 m 5个月后,0.39 mm,盖度达14.3%,生物量13.4 mg·g-1 ;盖度或先增后降,1年后13.8% 长期效应未知 [12 ,15 ] 古尔班通古特沙漠 2 m×5 m 15 d可达2.6 mg·cm-2 (叶绿素a) 未形成大规模生物结皮 未发表 地衣结皮 腾格里沙漠 3个 0.5 m2 1年后形成4—6 cm生物结皮 长期效应未知 [36 ] 藓类结皮 腾格里沙漠 10个1 m×1 m 7 d内存活,3个月内死亡 接种藓类直接采自野外 [59 ] 毛乌素沙地 18个1 m×1 m 75 d后盖度70%,密度9.8 株·cm-2 ,4年盖度达98% 接种藓类直接采自野外 [45 ,57 ] 杨凌 48个1 m×1 m 30 d,盖度90%,最大密度120株·cm-2 接种藓类直接采自野外 [14 ] 库布齐沙漠 8个1 m×1 m 3年后,盖度30%—70%,185株·cm-2 接种藓类直接采自野外 [60 ]
<strong>3.4</strong> 人工培养生物结皮的抗性锻炼 为提高野外存活率,是否对扩繁后的材料进行模拟野外环境的抗性锻炼,具有不同见解.抗性锻炼主要包括干燥和水分循环、高温和低温循环、不同光强等,以增加培养物的适合度[59 -62 ] .通过对20个蓝藻菌株进行抗性锻炼发现,13个菌株均表现出适合度的增加,尤其是结皮先锋藻类微鞘藻属的菌株,适合度均得到提升[63 ] .但通过培养混合生物结皮(藻类+地衣+藓类),进行一系列抗性锻炼(全谱光+温度变化+干旱),发现抗性锻炼无显著效果[62 ] ,藓类结皮物种作用同样不显著[64 ] .对不同种类藓类样品在不同存储温度研究发现,不同藓类种类后期生长对储存温度的敏感性不一致[65 ] .因此,生物结皮在接种前,是否有必要进行抗性锻炼,有待进一步验证. ...
毛乌素沙地苔藓结皮的人工培育技术
1
2012
... 生物结皮野外接种流程见图5 .在生物结皮接种前,采用草方格、立体栅栏等方法先行稳定沙面,改善生物结皮恢复初期地表条件.干藻种可与沙土混匀播撒到地表,藻悬浮液以雾化微喷的方式接种到沙面[9 ,37 ] .随着技术的进步,可利用无人机、飞机撒播等手段在更大空间尺度实现培养物的均匀喷洒.野外恢复的同时,也可辅助以微喷溉系统每天添加适量的水(20 mm·d-1 ),来保持藻类的生存活力[39 ] .虽然藓类结皮室内大规模扩繁报道较多[40 -41 ,45 ] ,但利用室内扩繁藓类成功进行野外恢复的研究鲜见报道.目前,有研究者直接采集自然藓类结皮植株,粉碎后在其他受损区域进行撒播,辅以一定的遮阳、覆膜、营养液等措施,以促进该区域藓类结皮恢复[14 ,45 -46 ] .这种藓类结皮恢复的存在较大的资源限制,如果大规模应用,将会给自然发育的藓类结皮造成巨大资源压力,并有可能造成新的退化区.为促进接种物更快生长、形成生物结皮,除添加营养液解除土壤养分限制外,有如下模式参考. ...
Practices of biological soil crust rehabilitation in China:experiences and challenges
2
2020
... 从机械和生物模式组合角度,生物结皮野外接种初期,一些稳定的措施,包括物理措施(如草方格)和生物措施(草本、灌丛和乔木)的组合,将有助于生物结皮的发育[38 ,47 ] .Chen等[39 ] 研究表明,适度的降水和降低光强的措施有助于藻类结皮的快速生长.通过藻-草-灌-乔四位一体的模式,在库布齐沙漠的大规模流沙治理中得到了应用[9 ] . ...
... 近几十年来,应用人工生物结皮技术开展固沙实践得到了国内外的广泛关注,通过生物结皮人工扩繁等手段,已经在局部区域实现生物结皮的成功野外恢复.随着研究的深入和技术进步,生物结皮人工恢复具有极大的应用潜力,尤其是在降水稀少、不宜进行林草恢复的干旱荒漠区.结合当地条件,因地制宜地开展生物结皮的恢复,将有利于防沙固沙的综合防治和技术开发利用,也有利于荒漠化防治和生态修复工程的节约成本.根据粗略对比分析,每公顷人工藻类结皮的成本0.14—1.4万元,草方格成本1.9—2.9万元,防风固沙人工林0.2—0.8万元(降水量100—200 mm地区)或4.2—5.8万元(降水量<50 mm地区)[47 ] .因此,干旱荒漠区生物结皮人工恢复在受损荒漠生态系统地表稳定与修复,以及荒漠化防治等领域具有重要的应用前景. ...
Application of sodium alginate in induced biological soil crusts: enhancing the sand stabilization in the early stage
1
2017
... 从促进藻类生长化学试剂角度,施用无毒副作用且价格低廉的化合物,是除物理和生物措施外,辅助藻类生长的重要补充.如海藻酸钠等可在沙面形成具有抗压能力的薄层,同时可促进蓝藻的克隆和生长[48 ] .有研究将藻类包裹制成海藻酸钠丸,施加到样地,取得较好的效果[49 ] .促进保水以及生物结皮生长的还有高持水性的化学材料(如SAP)、聚乙烯醇(PVA)和Tacki-Spray等化学试剂[15 ,50 ] . ...
Pelletized cyanobacterial soil amendments: laboratory testing for survival,escapability,and nitrogen fixation
1
1998
... 从促进藻类生长化学试剂角度,施用无毒副作用且价格低廉的化合物,是除物理和生物措施外,辅助藻类生长的重要补充.如海藻酸钠等可在沙面形成具有抗压能力的薄层,同时可促进蓝藻的克隆和生长[48 ] .有研究将藻类包裹制成海藻酸钠丸,施加到样地,取得较好的效果[49 ] .促进保水以及生物结皮生长的还有高持水性的化学材料(如SAP)、聚乙烯醇(PVA)和Tacki-Spray等化学试剂[15 ,50 ] . ...
Combined application of cyanobacteria with soil fixing chemicals for rapid induction of biological soil crust formation
1
2017
... 从促进藻类生长化学试剂角度,施用无毒副作用且价格低廉的化合物,是除物理和生物措施外,辅助藻类生长的重要补充.如海藻酸钠等可在沙面形成具有抗压能力的薄层,同时可促进蓝藻的克隆和生长[48 ] .有研究将藻类包裹制成海藻酸钠丸,施加到样地,取得较好的效果[49 ] .促进保水以及生物结皮生长的还有高持水性的化学材料(如SAP)、聚乙烯醇(PVA)和Tacki-Spray等化学试剂[15 ,50 ] . ...
库布齐沙地三年生人工藻结皮发育特征及微生物分布
3
2009
... 许多指标可用来指示人工生物结皮生长、恢复的状况,能够实现自身持续的可支持系统,是其恢复成功的重要标志[37 ] .从生物结皮本身的角度来看,形成肉眼可见的藻类结皮是一个重要的标志[38 ] .以生物结皮生物量表征恢复实验的效果,可直接反映生物结皮的生长状况,如通过对藻类结皮的2—3年人工培养,叶绿素含量可达35 μg·cm-2[39 ,51 ] .人工生物结皮的抗压强度和生物结皮厚度能够指示其固沙能力的强弱,随着培育时间的推移,其抗压强度得到显著增加,3年后可达3.83 kg·cm-2[38 ] .对于地衣结皮,通过扫描电镜观察生物结皮菌藻复合物,可发现石果衣菌丝向土壤基质下部延伸6—8 mm[40 ] .对于藓类结皮,盖度和密度常用来指示一定面积的生物结皮恢复,如培养65 d后,盖度可达83%,密度可达122株·cm-2[42 ] .从环境改善的角度,人工生物结皮的发育可促进土壤有机碳、多糖、总氮等养分和土壤酶活性的增加[40 ,52 -53 ] .人工生物结皮区沙生植物得到拓殖和生长,植物群落演替加速,同时土壤微生物数量也大量增加,固沙能力增强[38 ,51 ,54 ] . ...
... ,51 ,54 ]. ...
... Study case of biological soil crust rehabilitation in China
Table 1 结皮类型 地点 规模 结皮生长 评价 文献 藻类结皮 库布齐沙漠 约200 hm2 2—3年形成3—5 mm藻类结皮,盖度48.5% 结皮和植被得到较好恢复 [9 ,38 ,51 ] 腾格里沙漠 2 m×2 m 5个月后,0.39 mm,盖度达14.3%,生物量13.4 mg·g-1 ;盖度或先增后降,1年后13.8% 长期效应未知 [12 ,15 ] 古尔班通古特沙漠 2 m×5 m 15 d可达2.6 mg·cm-2 (叶绿素a) 未形成大规模生物结皮 未发表 地衣结皮 腾格里沙漠 3个 0.5 m2 1年后形成4—6 cm生物结皮 长期效应未知 [36 ] 藓类结皮 腾格里沙漠 10个1 m×1 m 7 d内存活,3个月内死亡 接种藓类直接采自野外 [59 ] 毛乌素沙地 18个1 m×1 m 75 d后盖度70%,密度9.8 株·cm-2 ,4年盖度达98% 接种藓类直接采自野外 [45 ,57 ] 杨凌 48个1 m×1 m 30 d,盖度90%,最大密度120株·cm-2 接种藓类直接采自野外 [14 ] 库布齐沙漠 8个1 m×1 m 3年后,盖度30%—70%,185株·cm-2 接种藓类直接采自野外 [60 ]
<strong>3.4</strong> 人工培养生物结皮的抗性锻炼 为提高野外存活率,是否对扩繁后的材料进行模拟野外环境的抗性锻炼,具有不同见解.抗性锻炼主要包括干燥和水分循环、高温和低温循环、不同光强等,以增加培养物的适合度[59 -62 ] .通过对20个蓝藻菌株进行抗性锻炼发现,13个菌株均表现出适合度的增加,尤其是结皮先锋藻类微鞘藻属的菌株,适合度均得到提升[63 ] .但通过培养混合生物结皮(藻类+地衣+藓类),进行一系列抗性锻炼(全谱光+温度变化+干旱),发现抗性锻炼无显著效果[62 ] ,藓类结皮物种作用同样不显著[64 ] .对不同种类藓类样品在不同存储温度研究发现,不同藓类种类后期生长对储存温度的敏感性不一致[65 ] .因此,生物结皮在接种前,是否有必要进行抗性锻炼,有待进一步验证. ...
人工藻结皮对库布齐沙地土壤酶活性的影响
1
2007
... 许多指标可用来指示人工生物结皮生长、恢复的状况,能够实现自身持续的可支持系统,是其恢复成功的重要标志[37 ] .从生物结皮本身的角度来看,形成肉眼可见的藻类结皮是一个重要的标志[38 ] .以生物结皮生物量表征恢复实验的效果,可直接反映生物结皮的生长状况,如通过对藻类结皮的2—3年人工培养,叶绿素含量可达35 μg·cm-2[39 ,51 ] .人工生物结皮的抗压强度和生物结皮厚度能够指示其固沙能力的强弱,随着培育时间的推移,其抗压强度得到显著增加,3年后可达3.83 kg·cm-2[38 ] .对于地衣结皮,通过扫描电镜观察生物结皮菌藻复合物,可发现石果衣菌丝向土壤基质下部延伸6—8 mm[40 ] .对于藓类结皮,盖度和密度常用来指示一定面积的生物结皮恢复,如培养65 d后,盖度可达83%,密度可达122株·cm-2[42 ] .从环境改善的角度,人工生物结皮的发育可促进土壤有机碳、多糖、总氮等养分和土壤酶活性的增加[40 ,52 -53 ] .人工生物结皮区沙生植物得到拓殖和生长,植物群落演替加速,同时土壤微生物数量也大量增加,固沙能力增强[38 ,51 ,54 ] . ...
人工生物结皮的发育演替及表土持水特性研究
1
2014
... 许多指标可用来指示人工生物结皮生长、恢复的状况,能够实现自身持续的可支持系统,是其恢复成功的重要标志[37 ] .从生物结皮本身的角度来看,形成肉眼可见的藻类结皮是一个重要的标志[38 ] .以生物结皮生物量表征恢复实验的效果,可直接反映生物结皮的生长状况,如通过对藻类结皮的2—3年人工培养,叶绿素含量可达35 μg·cm-2[39 ,51 ] .人工生物结皮的抗压强度和生物结皮厚度能够指示其固沙能力的强弱,随着培育时间的推移,其抗压强度得到显著增加,3年后可达3.83 kg·cm-2[38 ] .对于地衣结皮,通过扫描电镜观察生物结皮菌藻复合物,可发现石果衣菌丝向土壤基质下部延伸6—8 mm[40 ] .对于藓类结皮,盖度和密度常用来指示一定面积的生物结皮恢复,如培养65 d后,盖度可达83%,密度可达122株·cm-2[42 ] .从环境改善的角度,人工生物结皮的发育可促进土壤有机碳、多糖、总氮等养分和土壤酶活性的增加[40 ,52 -53 ] .人工生物结皮区沙生植物得到拓殖和生长,植物群落演替加速,同时土壤微生物数量也大量增加,固沙能力增强[38 ,51 ,54 ] . ...
Artificially accelerating the reversal of desertification: cyanobacterial inoculation facilitates the succession of vegetation communities
1
2014
... 许多指标可用来指示人工生物结皮生长、恢复的状况,能够实现自身持续的可支持系统,是其恢复成功的重要标志[37 ] .从生物结皮本身的角度来看,形成肉眼可见的藻类结皮是一个重要的标志[38 ] .以生物结皮生物量表征恢复实验的效果,可直接反映生物结皮的生长状况,如通过对藻类结皮的2—3年人工培养,叶绿素含量可达35 μg·cm-2[39 ,51 ] .人工生物结皮的抗压强度和生物结皮厚度能够指示其固沙能力的强弱,随着培育时间的推移,其抗压强度得到显著增加,3年后可达3.83 kg·cm-2[38 ] .对于地衣结皮,通过扫描电镜观察生物结皮菌藻复合物,可发现石果衣菌丝向土壤基质下部延伸6—8 mm[40 ] .对于藓类结皮,盖度和密度常用来指示一定面积的生物结皮恢复,如培养65 d后,盖度可达83%,密度可达122株·cm-2[42 ] .从环境改善的角度,人工生物结皮的发育可促进土壤有机碳、多糖、总氮等养分和土壤酶活性的增加[40 ,52 -53 ] .人工生物结皮区沙生植物得到拓殖和生长,植物群落演替加速,同时土壤微生物数量也大量增加,固沙能力增强[38 ,51 ,54 ] . ...
我国荒漠化和沙化面积连续15年“双缩减”
1
2020
... 中国是沙漠化发展最快的国家之一,造成生态系统失衡、耕地萎缩,严重地影响工农业生产和人民生活[4 ] .近年来,通过一系列生态工程的实施,荒漠化和沙化面积实现双缩减[55 ] .与“三北”防护林、京津源风沙源、退耕还林等工程相比,生物结皮的恢复工程规模相对较小.在降水量相对较少的干旱区,大规模植树造林的措施并不适用[4 ] .2001—2005年,中国科学院与内蒙古自治区在库布齐沙漠建立了生物结皮与乔灌草结合的示范区,成功治理了近200 hm2 流沙,并在其他区域进行大面积推广,该项目成为了生物结皮野外成功恢复的典范[9 ] .同时,该团队采用了工程化机械接种技术克服了人工生物结皮接种耗时费工的缺点,成为生物结皮工程化大面积应用的重要措施.除此之外,在其他区域,如腾格里沙漠、古尔班通古特沙漠、毛乌素沙地、黄土高原区等均有相应的生物结皮恢复应用的报道,但均处于方法探索和前期试验阶段[15 ,56 -58 ] (表1 ).国外生物结皮恢复研究虽然开展较早,但还未见能形成规模的生物结皮恢复示范区,中国生物结皮的恢复为世界生物结皮的恢复提供了重要借鉴. ...
人工培育生物结皮在毛乌素沙地光伏电站施工迹地的风蚀防治研究
1
2016
... 中国是沙漠化发展最快的国家之一,造成生态系统失衡、耕地萎缩,严重地影响工农业生产和人民生活[4 ] .近年来,通过一系列生态工程的实施,荒漠化和沙化面积实现双缩减[55 ] .与“三北”防护林、京津源风沙源、退耕还林等工程相比,生物结皮的恢复工程规模相对较小.在降水量相对较少的干旱区,大规模植树造林的措施并不适用[4 ] .2001—2005年,中国科学院与内蒙古自治区在库布齐沙漠建立了生物结皮与乔灌草结合的示范区,成功治理了近200 hm2 流沙,并在其他区域进行大面积推广,该项目成为了生物结皮野外成功恢复的典范[9 ] .同时,该团队采用了工程化机械接种技术克服了人工生物结皮接种耗时费工的缺点,成为生物结皮工程化大面积应用的重要措施.除此之外,在其他区域,如腾格里沙漠、古尔班通古特沙漠、毛乌素沙地、黄土高原区等均有相应的生物结皮恢复应用的报道,但均处于方法探索和前期试验阶段[15 ,56 -58 ] (表1 ).国外生物结皮恢复研究虽然开展较早,但还未见能形成规模的生物结皮恢复示范区,中国生物结皮的恢复为世界生物结皮的恢复提供了重要借鉴. ...
陕北毛乌素沙地生物结皮的土壤水分效应及其人工培育技术初探
1
2010
... Study case of biological soil crust rehabilitation in China
Table 1 结皮类型 地点 规模 结皮生长 评价 文献 藻类结皮 库布齐沙漠 约200 hm2 2—3年形成3—5 mm藻类结皮,盖度48.5% 结皮和植被得到较好恢复 [9 ,38 ,51 ] 腾格里沙漠 2 m×2 m 5个月后,0.39 mm,盖度达14.3%,生物量13.4 mg·g-1 ;盖度或先增后降,1年后13.8% 长期效应未知 [12 ,15 ] 古尔班通古特沙漠 2 m×5 m 15 d可达2.6 mg·cm-2 (叶绿素a) 未形成大规模生物结皮 未发表 地衣结皮 腾格里沙漠 3个 0.5 m2 1年后形成4—6 cm生物结皮 长期效应未知 [36 ] 藓类结皮 腾格里沙漠 10个1 m×1 m 7 d内存活,3个月内死亡 接种藓类直接采自野外 [59 ] 毛乌素沙地 18个1 m×1 m 75 d后盖度70%,密度9.8 株·cm-2 ,4年盖度达98% 接种藓类直接采自野外 [45 ,57 ] 杨凌 48个1 m×1 m 30 d,盖度90%,最大密度120株·cm-2 接种藓类直接采自野外 [14 ] 库布齐沙漠 8个1 m×1 m 3年后,盖度30%—70%,185株·cm-2 接种藓类直接采自野外 [60 ]
<strong>3.4</strong> 人工培养生物结皮的抗性锻炼 为提高野外存活率,是否对扩繁后的材料进行模拟野外环境的抗性锻炼,具有不同见解.抗性锻炼主要包括干燥和水分循环、高温和低温循环、不同光强等,以增加培养物的适合度[59 -62 ] .通过对20个蓝藻菌株进行抗性锻炼发现,13个菌株均表现出适合度的增加,尤其是结皮先锋藻类微鞘藻属的菌株,适合度均得到提升[63 ] .但通过培养混合生物结皮(藻类+地衣+藓类),进行一系列抗性锻炼(全谱光+温度变化+干旱),发现抗性锻炼无显著效果[62 ] ,藓类结皮物种作用同样不显著[64 ] .对不同种类藓类样品在不同存储温度研究发现,不同藓类种类后期生长对储存温度的敏感性不一致[65 ] .因此,生物结皮在接种前,是否有必要进行抗性锻炼,有待进一步验证. ...
水分对具鞘微鞘藻构建人工藻结皮的作用
1
2013
... 中国是沙漠化发展最快的国家之一,造成生态系统失衡、耕地萎缩,严重地影响工农业生产和人民生活[4 ] .近年来,通过一系列生态工程的实施,荒漠化和沙化面积实现双缩减[55 ] .与“三北”防护林、京津源风沙源、退耕还林等工程相比,生物结皮的恢复工程规模相对较小.在降水量相对较少的干旱区,大规模植树造林的措施并不适用[4 ] .2001—2005年,中国科学院与内蒙古自治区在库布齐沙漠建立了生物结皮与乔灌草结合的示范区,成功治理了近200 hm2 流沙,并在其他区域进行大面积推广,该项目成为了生物结皮野外成功恢复的典范[9 ] .同时,该团队采用了工程化机械接种技术克服了人工生物结皮接种耗时费工的缺点,成为生物结皮工程化大面积应用的重要措施.除此之外,在其他区域,如腾格里沙漠、古尔班通古特沙漠、毛乌素沙地、黄土高原区等均有相应的生物结皮恢复应用的报道,但均处于方法探索和前期试验阶段[15 ,56 -58 ] (表1 ).国外生物结皮恢复研究虽然开展较早,但还未见能形成规模的生物结皮恢复示范区,中国生物结皮的恢复为世界生物结皮的恢复提供了重要借鉴. ...
腾格里沙漠固定沙丘藓类植物结皮层的自然恢复及人工培养试验研究
2
2005
... Study case of biological soil crust rehabilitation in China
Table 1 结皮类型 地点 规模 结皮生长 评价 文献 藻类结皮 库布齐沙漠 约200 hm2 2—3年形成3—5 mm藻类结皮,盖度48.5% 结皮和植被得到较好恢复 [9 ,38 ,51 ] 腾格里沙漠 2 m×2 m 5个月后,0.39 mm,盖度达14.3%,生物量13.4 mg·g-1 ;盖度或先增后降,1年后13.8% 长期效应未知 [12 ,15 ] 古尔班通古特沙漠 2 m×5 m 15 d可达2.6 mg·cm-2 (叶绿素a) 未形成大规模生物结皮 未发表 地衣结皮 腾格里沙漠 3个 0.5 m2 1年后形成4—6 cm生物结皮 长期效应未知 [36 ] 藓类结皮 腾格里沙漠 10个1 m×1 m 7 d内存活,3个月内死亡 接种藓类直接采自野外 [59 ] 毛乌素沙地 18个1 m×1 m 75 d后盖度70%,密度9.8 株·cm-2 ,4年盖度达98% 接种藓类直接采自野外 [45 ,57 ] 杨凌 48个1 m×1 m 30 d,盖度90%,最大密度120株·cm-2 接种藓类直接采自野外 [14 ] 库布齐沙漠 8个1 m×1 m 3年后,盖度30%—70%,185株·cm-2 接种藓类直接采自野外 [60 ]
<strong>3.4</strong> 人工培养生物结皮的抗性锻炼 为提高野外存活率,是否对扩繁后的材料进行模拟野外环境的抗性锻炼,具有不同见解.抗性锻炼主要包括干燥和水分循环、高温和低温循环、不同光强等,以增加培养物的适合度[59 -62 ] .通过对20个蓝藻菌株进行抗性锻炼发现,13个菌株均表现出适合度的增加,尤其是结皮先锋藻类微鞘藻属的菌株,适合度均得到提升[63 ] .但通过培养混合生物结皮(藻类+地衣+藓类),进行一系列抗性锻炼(全谱光+温度变化+干旱),发现抗性锻炼无显著效果[62 ] ,藓类结皮物种作用同样不显著[64 ] .对不同种类藓类样品在不同存储温度研究发现,不同藓类种类后期生长对储存温度的敏感性不一致[65 ] .因此,生物结皮在接种前,是否有必要进行抗性锻炼,有待进一步验证. ...
... 为提高野外存活率,是否对扩繁后的材料进行模拟野外环境的抗性锻炼,具有不同见解.抗性锻炼主要包括干燥和水分循环、高温和低温循环、不同光强等,以增加培养物的适合度[59 -62 ] .通过对20个蓝藻菌株进行抗性锻炼发现,13个菌株均表现出适合度的增加,尤其是结皮先锋藻类微鞘藻属的菌株,适合度均得到提升[63 ] .但通过培养混合生物结皮(藻类+地衣+藓类),进行一系列抗性锻炼(全谱光+温度变化+干旱),发现抗性锻炼无显著效果[62 ] ,藓类结皮物种作用同样不显著[64 ] .对不同种类藓类样品在不同存储温度研究发现,不同藓类种类后期生长对储存温度的敏感性不一致[65 ] .因此,生物结皮在接种前,是否有必要进行抗性锻炼,有待进一步验证. ...
藓类结皮层人工培养试验和维持机制研究
1
2012
... Study case of biological soil crust rehabilitation in China
Table 1 结皮类型 地点 规模 结皮生长 评价 文献 藻类结皮 库布齐沙漠 约200 hm2 2—3年形成3—5 mm藻类结皮,盖度48.5% 结皮和植被得到较好恢复 [9 ,38 ,51 ] 腾格里沙漠 2 m×2 m 5个月后,0.39 mm,盖度达14.3%,生物量13.4 mg·g-1 ;盖度或先增后降,1年后13.8% 长期效应未知 [12 ,15 ] 古尔班通古特沙漠 2 m×5 m 15 d可达2.6 mg·cm-2 (叶绿素a) 未形成大规模生物结皮 未发表 地衣结皮 腾格里沙漠 3个 0.5 m2 1年后形成4—6 cm生物结皮 长期效应未知 [36 ] 藓类结皮 腾格里沙漠 10个1 m×1 m 7 d内存活,3个月内死亡 接种藓类直接采自野外 [59 ] 毛乌素沙地 18个1 m×1 m 75 d后盖度70%,密度9.8 株·cm-2 ,4年盖度达98% 接种藓类直接采自野外 [45 ,57 ] 杨凌 48个1 m×1 m 30 d,盖度90%,最大密度120株·cm-2 接种藓类直接采自野外 [14 ] 库布齐沙漠 8个1 m×1 m 3年后,盖度30%—70%,185株·cm-2 接种藓类直接采自野外 [60 ]
<strong>3.4</strong> 人工培养生物结皮的抗性锻炼 为提高野外存活率,是否对扩繁后的材料进行模拟野外环境的抗性锻炼,具有不同见解.抗性锻炼主要包括干燥和水分循环、高温和低温循环、不同光强等,以增加培养物的适合度[59 -62 ] .通过对20个蓝藻菌株进行抗性锻炼发现,13个菌株均表现出适合度的增加,尤其是结皮先锋藻类微鞘藻属的菌株,适合度均得到提升[63 ] .但通过培养混合生物结皮(藻类+地衣+藓类),进行一系列抗性锻炼(全谱光+温度变化+干旱),发现抗性锻炼无显著效果[62 ] ,藓类结皮物种作用同样不显著[64 ] .对不同种类藓类样品在不同存储温度研究发现,不同藓类种类后期生长对储存温度的敏感性不一致[65 ] .因此,生物结皮在接种前,是否有必要进行抗性锻炼,有待进一步验证. ...
Nursing biocrusts: isolation,cultivation,and fitness test of indigenous cyanobacteria
2019
Improving field success of biocrust rehabilitation materials: hardening the organisms or softening the environment?
2
2019
... 为提高野外存活率,是否对扩繁后的材料进行模拟野外环境的抗性锻炼,具有不同见解.抗性锻炼主要包括干燥和水分循环、高温和低温循环、不同光强等,以增加培养物的适合度[59 -62 ] .通过对20个蓝藻菌株进行抗性锻炼发现,13个菌株均表现出适合度的增加,尤其是结皮先锋藻类微鞘藻属的菌株,适合度均得到提升[63 ] .但通过培养混合生物结皮(藻类+地衣+藓类),进行一系列抗性锻炼(全谱光+温度变化+干旱),发现抗性锻炼无显著效果[62 ] ,藓类结皮物种作用同样不显著[64 ] .对不同种类藓类样品在不同存储温度研究发现,不同藓类种类后期生长对储存温度的敏感性不一致[65 ] .因此,生物结皮在接种前,是否有必要进行抗性锻炼,有待进一步验证. ...
... [62 ],藓类结皮物种作用同样不显著[64 ] .对不同种类藓类样品在不同存储温度研究发现,不同藓类种类后期生长对储存温度的敏感性不一致[65 ] .因此,生物结皮在接种前,是否有必要进行抗性锻炼,有待进一步验证. ...
Effect of preconditioning to the soil environment on the performance of 20 cyanobacterial strains used as inoculum for biocrust restoration
1
2019
... 为提高野外存活率,是否对扩繁后的材料进行模拟野外环境的抗性锻炼,具有不同见解.抗性锻炼主要包括干燥和水分循环、高温和低温循环、不同光强等,以增加培养物的适合度[59 -62 ] .通过对20个蓝藻菌株进行抗性锻炼发现,13个菌株均表现出适合度的增加,尤其是结皮先锋藻类微鞘藻属的菌株,适合度均得到提升[63 ] .但通过培养混合生物结皮(藻类+地衣+藓类),进行一系列抗性锻炼(全谱光+温度变化+干旱),发现抗性锻炼无显著效果[62 ] ,藓类结皮物种作用同样不显著[64 ] .对不同种类藓类样品在不同存储温度研究发现,不同藓类种类后期生长对储存温度的敏感性不一致[65 ] .因此,生物结皮在接种前,是否有必要进行抗性锻炼,有待进一步验证. ...
Maximizing establishment and survivorship of field-collected and greenhouse-cultivated biocrusts in a semi-cold desert
1
2018
... 为提高野外存活率,是否对扩繁后的材料进行模拟野外环境的抗性锻炼,具有不同见解.抗性锻炼主要包括干燥和水分循环、高温和低温循环、不同光强等,以增加培养物的适合度[59 -62 ] .通过对20个蓝藻菌株进行抗性锻炼发现,13个菌株均表现出适合度的增加,尤其是结皮先锋藻类微鞘藻属的菌株,适合度均得到提升[63 ] .但通过培养混合生物结皮(藻类+地衣+藓类),进行一系列抗性锻炼(全谱光+温度变化+干旱),发现抗性锻炼无显著效果[62 ] ,藓类结皮物种作用同样不显著[64 ] .对不同种类藓类样品在不同存储温度研究发现,不同藓类种类后期生长对储存温度的敏感性不一致[65 ] .因此,生物结皮在接种前,是否有必要进行抗性锻炼,有待进一步验证. ...
Effects of storage temperature on the physiological characteristics and vegetative propagation of desiccation-tolerant mosses
1
2018
... 为提高野外存活率,是否对扩繁后的材料进行模拟野外环境的抗性锻炼,具有不同见解.抗性锻炼主要包括干燥和水分循环、高温和低温循环、不同光强等,以增加培养物的适合度[59 -62 ] .通过对20个蓝藻菌株进行抗性锻炼发现,13个菌株均表现出适合度的增加,尤其是结皮先锋藻类微鞘藻属的菌株,适合度均得到提升[63 ] .但通过培养混合生物结皮(藻类+地衣+藓类),进行一系列抗性锻炼(全谱光+温度变化+干旱),发现抗性锻炼无显著效果[62 ] ,藓类结皮物种作用同样不显著[64 ] .对不同种类藓类样品在不同存储温度研究发现,不同藓类种类后期生长对储存温度的敏感性不一致[65 ] .因此,生物结皮在接种前,是否有必要进行抗性锻炼,有待进一步验证. ...








 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}