2
2009
... 生态位作为评价种间关系及种群在群落中地位的重要概念,在演替理论中具有极其重要的地位[1-2].生态位宽度是衡量不同物种生物对环境生态需求差异的基本测度,而生态位重叠是不同物种间生态学相似性的基本测度,开展生态位研究对于理解种间关系具有重要作用.种间关联指不同种群在空间分布上的相互关联性[3],是生态位在空间分布上的体现,能够对一定时期内组成群落和物种相互关系进行静态描述.生态系统中相邻植物个体间可能存在竞争(负相互作用)、互惠(正相互作用)和无直接联系的相对独立的中立关系.植物间的关系并非一成不变,其性质和强度随环境条件的改变而发生不同程度的变化[4-5].Bertness等[6]提出的胁迫梯度假说(stress gradient hypothesis, SGH)认为,随着环境胁迫强度的增加,物种间主要相互作用将由竞争转变为互惠,但也有个别研究指出,随着胁迫或干扰程度的增强,植物间相互作用不一定是向着互惠增强的趋势,而是呈现为单峰型曲线变化,在适度胁迫条件下,互惠效应变得更强[7-8].截止目前,对胁迫条件下种间相互作用的认识还没有形成统一认知.上述相关研究结果多基于维管束植物之间相互关系的研究,而对非维管束植物或隐花植物的种间关联有待进一步深入,这也可能是当前植物种间关系的研究还没形成统一认识的重要原因. ...
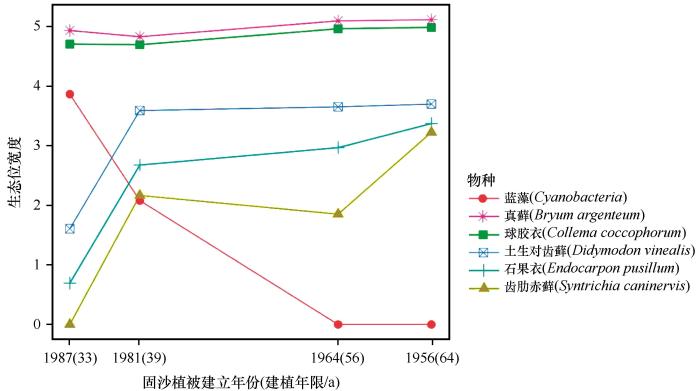
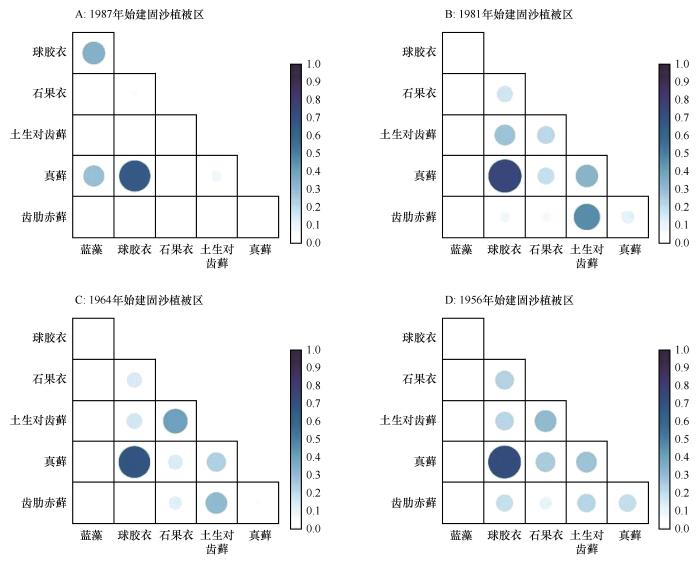
... 生态位宽度是反映物种对环境资源利用状况的尺度[21],生态位重叠则反映了种群之间对资源利用的相似程度和竞争关系[1,3].随着沙坡头地区固沙植被演替的进行,藻类生态位持续变窄(图1).根据结皮生物正向演替的一般规律模式[18],藻类为该群落的先锋类群,最先占据生境空斑,随后其代谢产物胶结沙粒等颗粒物质形成具有生物活性的团聚结构,促进演替后期物种的拓殖.根据植物CSR分类特征[22],藻类作为胁迫忍耐对策者,通常具有更宽的基础生态位,固沙植被演替后期藻类生态位宽度的降低可能源自演替后期物种对其竞争导致的实际生态位收缩.蓝藻在固沙植被演替前期与球胶衣和真藓有一定生态位重叠(图2),与其他结皮生物重叠值很小,这可能与藻类自身特性有关[22]. ...
Disentangling the drivers of β diversity along latitudinal and elevational gradients
1
2011
... 生态位作为评价种间关系及种群在群落中地位的重要概念,在演替理论中具有极其重要的地位[1-2].生态位宽度是衡量不同物种生物对环境生态需求差异的基本测度,而生态位重叠是不同物种间生态学相似性的基本测度,开展生态位研究对于理解种间关系具有重要作用.种间关联指不同种群在空间分布上的相互关联性[3],是生态位在空间分布上的体现,能够对一定时期内组成群落和物种相互关系进行静态描述.生态系统中相邻植物个体间可能存在竞争(负相互作用)、互惠(正相互作用)和无直接联系的相对独立的中立关系.植物间的关系并非一成不变,其性质和强度随环境条件的改变而发生不同程度的变化[4-5].Bertness等[6]提出的胁迫梯度假说(stress gradient hypothesis, SGH)认为,随着环境胁迫强度的增加,物种间主要相互作用将由竞争转变为互惠,但也有个别研究指出,随着胁迫或干扰程度的增强,植物间相互作用不一定是向着互惠增强的趋势,而是呈现为单峰型曲线变化,在适度胁迫条件下,互惠效应变得更强[7-8].截止目前,对胁迫条件下种间相互作用的认识还没有形成统一认知.上述相关研究结果多基于维管束植物之间相互关系的研究,而对非维管束植物或隐花植物的种间关联有待进一步深入,这也可能是当前植物种间关系的研究还没形成统一认识的重要原因. ...
4
2004
... 生态位作为评价种间关系及种群在群落中地位的重要概念,在演替理论中具有极其重要的地位[1-2].生态位宽度是衡量不同物种生物对环境生态需求差异的基本测度,而生态位重叠是不同物种间生态学相似性的基本测度,开展生态位研究对于理解种间关系具有重要作用.种间关联指不同种群在空间分布上的相互关联性[3],是生态位在空间分布上的体现,能够对一定时期内组成群落和物种相互关系进行静态描述.生态系统中相邻植物个体间可能存在竞争(负相互作用)、互惠(正相互作用)和无直接联系的相对独立的中立关系.植物间的关系并非一成不变,其性质和强度随环境条件的改变而发生不同程度的变化[4-5].Bertness等[6]提出的胁迫梯度假说(stress gradient hypothesis, SGH)认为,随着环境胁迫强度的增加,物种间主要相互作用将由竞争转变为互惠,但也有个别研究指出,随着胁迫或干扰程度的增强,植物间相互作用不一定是向着互惠增强的趋势,而是呈现为单峰型曲线变化,在适度胁迫条件下,互惠效应变得更强[7-8].截止目前,对胁迫条件下种间相互作用的认识还没有形成统一认知.上述相关研究结果多基于维管束植物之间相互关系的研究,而对非维管束植物或隐花植物的种间关联有待进一步深入,这也可能是当前植物种间关系的研究还没形成统一认识的重要原因. ...
... 生态位宽度是反映物种对环境资源利用状况的尺度[21],生态位重叠则反映了种群之间对资源利用的相似程度和竞争关系[1,3].随着沙坡头地区固沙植被演替的进行,藻类生态位持续变窄(图1).根据结皮生物正向演替的一般规律模式[18],藻类为该群落的先锋类群,最先占据生境空斑,随后其代谢产物胶结沙粒等颗粒物质形成具有生物活性的团聚结构,促进演替后期物种的拓殖.根据植物CSR分类特征[22],藻类作为胁迫忍耐对策者,通常具有更宽的基础生态位,固沙植被演替后期藻类生态位宽度的降低可能源自演替后期物种对其竞争导致的实际生态位收缩.蓝藻在固沙植被演替前期与球胶衣和真藓有一定生态位重叠(图2),与其他结皮生物重叠值很小,这可能与藻类自身特性有关[22]. ...
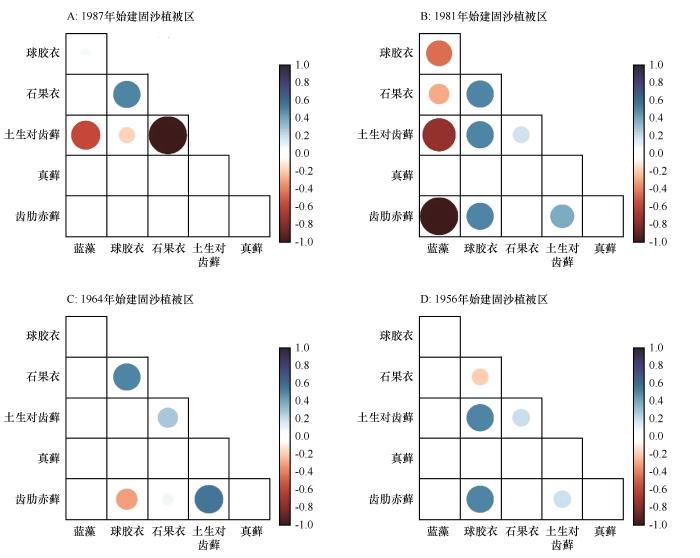
... 种间关联是不同种类在空间分布上的关联性,通常由不同物种的资源利用方式和种间关系决定[3].植物种间关联是植物群落重要的数量和结构特征,是植物群落结构形成和演化的基础[3].本研究中,不同结皮生物间的种间关联性质和强度随着固沙植被演替的进行不断发生变化(图3).蓝藻在固沙植被演替初期与球胶衣有较弱正关联,这可能得因于二者在前期资源匮乏时的相互协作,随着固沙植被演替进行,蓝藻与其他结皮生物均呈现出负关联,这可能是竞争排除作用所致.球胶衣与石果衣之间随着固沙植被演替进行,由正关联向负关联转变,这可能由于生境状况变好,二者相似的生态学特性和生物学特性导致相互之间竞争有限的环境资源和空间资源而产生竞争和排斥.石果衣与土生对齿藓的种间关联随着固沙植被演替呈现出由负转正的现象.结皮生物间的种间关联状态(图3)与生态位重叠关系基本一致(图2),因此可以推断资源利用方式是导致其种间关联的主要因素.群落的种间关联状况在一定程度上反映了群落的稳定性[25-26].我们的计算结果显示,沙坡头固沙植被区结皮生物种间总体关联性随着固沙植被演替由负关联向正关联转变,且在固沙植被演替后期产生了更广泛的正关联(表1),说明沙坡头地区不同结皮生物间随着固沙植被演替产生更强的联系,群落稳定性更强. ...
... [3].本研究中,不同结皮生物间的种间关联性质和强度随着固沙植被演替的进行不断发生变化(图3).蓝藻在固沙植被演替初期与球胶衣有较弱正关联,这可能得因于二者在前期资源匮乏时的相互协作,随着固沙植被演替进行,蓝藻与其他结皮生物均呈现出负关联,这可能是竞争排除作用所致.球胶衣与石果衣之间随着固沙植被演替进行,由正关联向负关联转变,这可能由于生境状况变好,二者相似的生态学特性和生物学特性导致相互之间竞争有限的环境资源和空间资源而产生竞争和排斥.石果衣与土生对齿藓的种间关联随着固沙植被演替呈现出由负转正的现象.结皮生物间的种间关联状态(图3)与生态位重叠关系基本一致(图2),因此可以推断资源利用方式是导致其种间关联的主要因素.群落的种间关联状况在一定程度上反映了群落的稳定性[25-26].我们的计算结果显示,沙坡头固沙植被区结皮生物种间总体关联性随着固沙植被演替由负关联向正关联转变,且在固沙植被演替后期产生了更广泛的正关联(表1),说明沙坡头地区不同结皮生物间随着固沙植被演替产生更强的联系,群落稳定性更强. ...
Refining the stress-gradient hypothesis for competition and facilitation in plant communities
1
2009
... 生态位作为评价种间关系及种群在群落中地位的重要概念,在演替理论中具有极其重要的地位[1-2].生态位宽度是衡量不同物种生物对环境生态需求差异的基本测度,而生态位重叠是不同物种间生态学相似性的基本测度,开展生态位研究对于理解种间关系具有重要作用.种间关联指不同种群在空间分布上的相互关联性[3],是生态位在空间分布上的体现,能够对一定时期内组成群落和物种相互关系进行静态描述.生态系统中相邻植物个体间可能存在竞争(负相互作用)、互惠(正相互作用)和无直接联系的相对独立的中立关系.植物间的关系并非一成不变,其性质和强度随环境条件的改变而发生不同程度的变化[4-5].Bertness等[6]提出的胁迫梯度假说(stress gradient hypothesis, SGH)认为,随着环境胁迫强度的增加,物种间主要相互作用将由竞争转变为互惠,但也有个别研究指出,随着胁迫或干扰程度的增强,植物间相互作用不一定是向着互惠增强的趋势,而是呈现为单峰型曲线变化,在适度胁迫条件下,互惠效应变得更强[7-8].截止目前,对胁迫条件下种间相互作用的认识还没有形成统一认知.上述相关研究结果多基于维管束植物之间相互关系的研究,而对非维管束植物或隐花植物的种间关联有待进一步深入,这也可能是当前植物种间关系的研究还没形成统一认识的重要原因. ...
Inclusion of biotic stress (consumer pressure) alters predictions from the stress gradient hypothesis
1
2009
... 生态位作为评价种间关系及种群在群落中地位的重要概念,在演替理论中具有极其重要的地位[1-2].生态位宽度是衡量不同物种生物对环境生态需求差异的基本测度,而生态位重叠是不同物种间生态学相似性的基本测度,开展生态位研究对于理解种间关系具有重要作用.种间关联指不同种群在空间分布上的相互关联性[3],是生态位在空间分布上的体现,能够对一定时期内组成群落和物种相互关系进行静态描述.生态系统中相邻植物个体间可能存在竞争(负相互作用)、互惠(正相互作用)和无直接联系的相对独立的中立关系.植物间的关系并非一成不变,其性质和强度随环境条件的改变而发生不同程度的变化[4-5].Bertness等[6]提出的胁迫梯度假说(stress gradient hypothesis, SGH)认为,随着环境胁迫强度的增加,物种间主要相互作用将由竞争转变为互惠,但也有个别研究指出,随着胁迫或干扰程度的增强,植物间相互作用不一定是向着互惠增强的趋势,而是呈现为单峰型曲线变化,在适度胁迫条件下,互惠效应变得更强[7-8].截止目前,对胁迫条件下种间相互作用的认识还没有形成统一认知.上述相关研究结果多基于维管束植物之间相互关系的研究,而对非维管束植物或隐花植物的种间关联有待进一步深入,这也可能是当前植物种间关系的研究还没形成统一认识的重要原因. ...
Positive interactions in communities
2
1994
... 生态位作为评价种间关系及种群在群落中地位的重要概念,在演替理论中具有极其重要的地位[1-2].生态位宽度是衡量不同物种生物对环境生态需求差异的基本测度,而生态位重叠是不同物种间生态学相似性的基本测度,开展生态位研究对于理解种间关系具有重要作用.种间关联指不同种群在空间分布上的相互关联性[3],是生态位在空间分布上的体现,能够对一定时期内组成群落和物种相互关系进行静态描述.生态系统中相邻植物个体间可能存在竞争(负相互作用)、互惠(正相互作用)和无直接联系的相对独立的中立关系.植物间的关系并非一成不变,其性质和强度随环境条件的改变而发生不同程度的变化[4-5].Bertness等[6]提出的胁迫梯度假说(stress gradient hypothesis, SGH)认为,随着环境胁迫强度的增加,物种间主要相互作用将由竞争转变为互惠,但也有个别研究指出,随着胁迫或干扰程度的增强,植物间相互作用不一定是向着互惠增强的趋势,而是呈现为单峰型曲线变化,在适度胁迫条件下,互惠效应变得更强[7-8].截止目前,对胁迫条件下种间相互作用的认识还没有形成统一认知.上述相关研究结果多基于维管束植物之间相互关系的研究,而对非维管束植物或隐花植物的种间关联有待进一步深入,这也可能是当前植物种间关系的研究还没形成统一认识的重要原因. ...
... 结皮生物的多样性、盖度和空间分布等对环境变化及干扰的响应十分敏感[27-30].沙坡头结皮生物组成和演变受到很多因素的影响,自然因素如气候、植被、土壤养分、水分等均为影响沙坡头生物多样性发展及生态系统恢复的主要因素[31-35],我们在设置样方时也有意避开或减少了植被的影响,将其设置在灌丛间.沙坡头人工固沙植被演替过程中,植被、土壤等环境的变化形成自然环境梯度[13].我们发现,当胁迫梯度增加,种间由竞争转向互惠,这与胁迫梯度假说[6]相一致.随着沙坡头固沙植被演替的进行,胁迫程度逐步降低,此时种间竞争加剧,可能有助于维持群落稳定,且土壤质地由粗变细、土壤持水量也呈现出逐步增加的趋势[36],因此可以预测,在沙坡头地区蓝藻会逐步被其他结皮生物所替代,地衣与藓类的数量会进一步增加,同时齿肋赤藓会逐步替代真藓成为该区主要的藓结皮生物.计算种间关联系数只是对种间关系的定量描述,取样尺度、种间竞争、化感作用、环境因素等都会对种间关联性产生影响[26].因此,对沙坡头人工固沙植被演替过程中结皮生物种间关联的内在机制还需进一步从植物生理、遗传学、分子生态学等多角度开展研究,得出更为合理的解释.沙区生态系统的恢复是漫长的过程,受多种自然因素与人为因素的影响,长期的监测和研究十分必要[13],沙坡头地区固沙植被演替过程中结皮生物生态位与种间关联规律还有待进一步观察研究. ...
Do positive interactions increase with abiotic stress?A test from asemi-arid steppe
1
2004
... 生态位作为评价种间关系及种群在群落中地位的重要概念,在演替理论中具有极其重要的地位[1-2].生态位宽度是衡量不同物种生物对环境生态需求差异的基本测度,而生态位重叠是不同物种间生态学相似性的基本测度,开展生态位研究对于理解种间关系具有重要作用.种间关联指不同种群在空间分布上的相互关联性[3],是生态位在空间分布上的体现,能够对一定时期内组成群落和物种相互关系进行静态描述.生态系统中相邻植物个体间可能存在竞争(负相互作用)、互惠(正相互作用)和无直接联系的相对独立的中立关系.植物间的关系并非一成不变,其性质和强度随环境条件的改变而发生不同程度的变化[4-5].Bertness等[6]提出的胁迫梯度假说(stress gradient hypothesis, SGH)认为,随着环境胁迫强度的增加,物种间主要相互作用将由竞争转变为互惠,但也有个别研究指出,随着胁迫或干扰程度的增强,植物间相互作用不一定是向着互惠增强的趋势,而是呈现为单峰型曲线变化,在适度胁迫条件下,互惠效应变得更强[7-8].截止目前,对胁迫条件下种间相互作用的认识还没有形成统一认知.上述相关研究结果多基于维管束植物之间相互关系的研究,而对非维管束植物或隐花植物的种间关联有待进一步深入,这也可能是当前植物种间关系的研究还没形成统一认识的重要原因. ...
Strong facilitation in mild environments:the stress gradient hypothesis revisited
1
2010
... 生态位作为评价种间关系及种群在群落中地位的重要概念,在演替理论中具有极其重要的地位[1-2].生态位宽度是衡量不同物种生物对环境生态需求差异的基本测度,而生态位重叠是不同物种间生态学相似性的基本测度,开展生态位研究对于理解种间关系具有重要作用.种间关联指不同种群在空间分布上的相互关联性[3],是生态位在空间分布上的体现,能够对一定时期内组成群落和物种相互关系进行静态描述.生态系统中相邻植物个体间可能存在竞争(负相互作用)、互惠(正相互作用)和无直接联系的相对独立的中立关系.植物间的关系并非一成不变,其性质和强度随环境条件的改变而发生不同程度的变化[4-5].Bertness等[6]提出的胁迫梯度假说(stress gradient hypothesis, SGH)认为,随着环境胁迫强度的增加,物种间主要相互作用将由竞争转变为互惠,但也有个别研究指出,随着胁迫或干扰程度的增强,植物间相互作用不一定是向着互惠增强的趋势,而是呈现为单峰型曲线变化,在适度胁迫条件下,互惠效应变得更强[7-8].截止目前,对胁迫条件下种间相互作用的认识还没有形成统一认知.上述相关研究结果多基于维管束植物之间相互关系的研究,而对非维管束植物或隐花植物的种间关联有待进一步深入,这也可能是当前植物种间关系的研究还没形成统一认识的重要原因. ...
Biological Soil Crusts:An Organizing Principle in Drylands
2
2016
... 生物土壤结皮(简称生物结皮)是由苔藓、地衣、藻类、真菌等土壤微生物在土壤表层与微小颗粒胶结形成的复合体[9].受水分条件的限制,干旱沙区无法支撑连片的、高密度的维管束植物群落,地表往往呈现出由维管束植物和生物结皮斑块镶嵌分布的格局[10-11].随着植被定植和流沙固定,地表结皮生物的盖度和生物量均发生不同程度的变化,表现出独特的演变规律[12].生物结皮演替通常始于稳定的表土环境[13],在风蚀[14]、水蚀[15]及沙埋[16]等严重干扰环境中很难形成生物结皮.在稳定表土环境中蓝藻首先拓殖,其代谢产物帮助形成土壤具有生物活性团聚结构,自此标志着生物结皮的初步形成[17].根据生物结皮演替过程中优势物种差异,可将其分为3个阶段,即以蓝藻为优势的演替初期阶段,以绿藻和蓝藻为优势的演替中期和以苔藓、地衣为优势的演替后期阶段[18].随着生物结皮的演替,结皮生物之间的种间关系也发生相应的变化,进而引发地表稳定性、水分、养分、土壤颗粒组成等变化,从而影响整个生态系统的稳定性[9].因此,探究结皮生物之间的关系随生境变化规律对全面理解沙区生态系统物种间关系及对生态系统结构与功能稳定性影响具有重要意义. ...
... [9].因此,探究结皮生物之间的关系随生境变化规律对全面理解沙区生态系统物种间关系及对生态系统结构与功能稳定性影响具有重要意义. ...
Long-term ecosystem effects of sand-binding vegetation in Shapotou region of Tengger Desert,Northern China
1
2004
... 生物土壤结皮(简称生物结皮)是由苔藓、地衣、藻类、真菌等土壤微生物在土壤表层与微小颗粒胶结形成的复合体[9].受水分条件的限制,干旱沙区无法支撑连片的、高密度的维管束植物群落,地表往往呈现出由维管束植物和生物结皮斑块镶嵌分布的格局[10-11].随着植被定植和流沙固定,地表结皮生物的盖度和生物量均发生不同程度的变化,表现出独特的演变规律[12].生物结皮演替通常始于稳定的表土环境[13],在风蚀[14]、水蚀[15]及沙埋[16]等严重干扰环境中很难形成生物结皮.在稳定表土环境中蓝藻首先拓殖,其代谢产物帮助形成土壤具有生物活性团聚结构,自此标志着生物结皮的初步形成[17].根据生物结皮演替过程中优势物种差异,可将其分为3个阶段,即以蓝藻为优势的演替初期阶段,以绿藻和蓝藻为优势的演替中期和以苔藓、地衣为优势的演替后期阶段[18].随着生物结皮的演替,结皮生物之间的种间关系也发生相应的变化,进而引发地表稳定性、水分、养分、土壤颗粒组成等变化,从而影响整个生态系统的稳定性[9].因此,探究结皮生物之间的关系随生境变化规律对全面理解沙区生态系统物种间关系及对生态系统结构与功能稳定性影响具有重要意义. ...
Effects of biological soil crusts on seed bank,germination and establishment of two desert annual plants
2
2005
... 生物土壤结皮(简称生物结皮)是由苔藓、地衣、藻类、真菌等土壤微生物在土壤表层与微小颗粒胶结形成的复合体[9].受水分条件的限制,干旱沙区无法支撑连片的、高密度的维管束植物群落,地表往往呈现出由维管束植物和生物结皮斑块镶嵌分布的格局[10-11].随着植被定植和流沙固定,地表结皮生物的盖度和生物量均发生不同程度的变化,表现出独特的演变规律[12].生物结皮演替通常始于稳定的表土环境[13],在风蚀[14]、水蚀[15]及沙埋[16]等严重干扰环境中很难形成生物结皮.在稳定表土环境中蓝藻首先拓殖,其代谢产物帮助形成土壤具有生物活性团聚结构,自此标志着生物结皮的初步形成[17].根据生物结皮演替过程中优势物种差异,可将其分为3个阶段,即以蓝藻为优势的演替初期阶段,以绿藻和蓝藻为优势的演替中期和以苔藓、地衣为优势的演替后期阶段[18].随着生物结皮的演替,结皮生物之间的种间关系也发生相应的变化,进而引发地表稳定性、水分、养分、土壤颗粒组成等变化,从而影响整个生态系统的稳定性[9].因此,探究结皮生物之间的关系随生境变化规律对全面理解沙区生态系统物种间关系及对生态系统结构与功能稳定性影响具有重要意义. ...
... 利用空间代替时间的方法[11],以始建于1956、1964、1981、1987年固沙植被区分别代表该区固沙植被演替的第64、56、39、33年4个阶段.沿4个演替阶段植被区设置3条样带,随后在每个样带中的各个演替阶段植被区上各选取1个典型迎风坡(生物结皮发育好、种类多且具有较好的代表性),每个坡设置3条平行样线,样线间隔1.5 m,然后,根据研究需要各样线平均间隔1 m设置一个30 cm×30 cm的样方.样方布置中为了避开或减少植被的影响,均设置在灌丛间. ...
1
2016
... 生物土壤结皮(简称生物结皮)是由苔藓、地衣、藻类、真菌等土壤微生物在土壤表层与微小颗粒胶结形成的复合体[9].受水分条件的限制,干旱沙区无法支撑连片的、高密度的维管束植物群落,地表往往呈现出由维管束植物和生物结皮斑块镶嵌分布的格局[10-11].随着植被定植和流沙固定,地表结皮生物的盖度和生物量均发生不同程度的变化,表现出独特的演变规律[12].生物结皮演替通常始于稳定的表土环境[13],在风蚀[14]、水蚀[15]及沙埋[16]等严重干扰环境中很难形成生物结皮.在稳定表土环境中蓝藻首先拓殖,其代谢产物帮助形成土壤具有生物活性团聚结构,自此标志着生物结皮的初步形成[17].根据生物结皮演替过程中优势物种差异,可将其分为3个阶段,即以蓝藻为优势的演替初期阶段,以绿藻和蓝藻为优势的演替中期和以苔藓、地衣为优势的演替后期阶段[18].随着生物结皮的演替,结皮生物之间的种间关系也发生相应的变化,进而引发地表稳定性、水分、养分、土壤颗粒组成等变化,从而影响整个生态系统的稳定性[9].因此,探究结皮生物之间的关系随生境变化规律对全面理解沙区生态系统物种间关系及对生态系统结构与功能稳定性影响具有重要意义. ...
5
2012
... 生物土壤结皮(简称生物结皮)是由苔藓、地衣、藻类、真菌等土壤微生物在土壤表层与微小颗粒胶结形成的复合体[9].受水分条件的限制,干旱沙区无法支撑连片的、高密度的维管束植物群落,地表往往呈现出由维管束植物和生物结皮斑块镶嵌分布的格局[10-11].随着植被定植和流沙固定,地表结皮生物的盖度和生物量均发生不同程度的变化,表现出独特的演变规律[12].生物结皮演替通常始于稳定的表土环境[13],在风蚀[14]、水蚀[15]及沙埋[16]等严重干扰环境中很难形成生物结皮.在稳定表土环境中蓝藻首先拓殖,其代谢产物帮助形成土壤具有生物活性团聚结构,自此标志着生物结皮的初步形成[17].根据生物结皮演替过程中优势物种差异,可将其分为3个阶段,即以蓝藻为优势的演替初期阶段,以绿藻和蓝藻为优势的演替中期和以苔藓、地衣为优势的演替后期阶段[18].随着生物结皮的演替,结皮生物之间的种间关系也发生相应的变化,进而引发地表稳定性、水分、养分、土壤颗粒组成等变化,从而影响整个生态系统的稳定性[9].因此,探究结皮生物之间的关系随生境变化规律对全面理解沙区生态系统物种间关系及对生态系统结构与功能稳定性影响具有重要意义. ...
... 腾格里沙漠人工固沙植被区拥有建植于不同年代的人工植被带,植被-土壤属性连续变化,结皮生物自身也在演变[13],其种间关系发生变化,为我们研究固沙植被演替过程中结皮生物生态位与种间关联变化的研究提供了理想场所.本文通过对腾格里沙漠人工固沙植被区演替序列中6种最主要的结皮生物分布差异调查,探讨结皮层生物物种分布规律及其种间关系变化特征,以丰富隐花植物种间关系的相关基础理论,为沙区生态系统生态修复措施中物种选择与搭配提供实践参考. ...
... 本研究选择该地区6种优势结皮生物作为研究对象,分别为蓝藻(Cyanobacteria,野外调查中具体藻种不易被鉴别,统一标记为蓝藻)、球胶衣、石果衣、真藓、土生对齿藓、齿肋赤藓.在每个小样方内采用针点样方框法调查结皮生物盖度[13].根据研究需要样框设为30 cm×30 cm,并等分为12×12格,每格2.5 cm×2.5 cm.测量时,将样方框垂直放置在待测样方上,观察每一方格是否出现隐花植物,根据隐花植物在框内出现的比率确定该结皮生物盖度,即某种结皮生物盖度(%)=隐花植物出现个数/144×100%.频度调查时,观察记录所调结皮生物出现的样方数,根据结皮生物出现的样方数占整个样方数的百分比确定该结皮生物频度. ...
... 结皮生物的多样性、盖度和空间分布等对环境变化及干扰的响应十分敏感[27-30].沙坡头结皮生物组成和演变受到很多因素的影响,自然因素如气候、植被、土壤养分、水分等均为影响沙坡头生物多样性发展及生态系统恢复的主要因素[31-35],我们在设置样方时也有意避开或减少了植被的影响,将其设置在灌丛间.沙坡头人工固沙植被演替过程中,植被、土壤等环境的变化形成自然环境梯度[13].我们发现,当胁迫梯度增加,种间由竞争转向互惠,这与胁迫梯度假说[6]相一致.随着沙坡头固沙植被演替的进行,胁迫程度逐步降低,此时种间竞争加剧,可能有助于维持群落稳定,且土壤质地由粗变细、土壤持水量也呈现出逐步增加的趋势[36],因此可以预测,在沙坡头地区蓝藻会逐步被其他结皮生物所替代,地衣与藓类的数量会进一步增加,同时齿肋赤藓会逐步替代真藓成为该区主要的藓结皮生物.计算种间关联系数只是对种间关系的定量描述,取样尺度、种间竞争、化感作用、环境因素等都会对种间关联性产生影响[26].因此,对沙坡头人工固沙植被演替过程中结皮生物种间关联的内在机制还需进一步从植物生理、遗传学、分子生态学等多角度开展研究,得出更为合理的解释.沙区生态系统的恢复是漫长的过程,受多种自然因素与人为因素的影响,长期的监测和研究十分必要[13],沙坡头地区固沙植被演替过程中结皮生物生态位与种间关联规律还有待进一步观察研究. ...
... [13],沙坡头地区固沙植被演替过程中结皮生物生态位与种间关联规律还有待进一步观察研究. ...
Differential wind tolerance of soil crust mosses explains their micro-distribution in nature
1
2012
... 生物土壤结皮(简称生物结皮)是由苔藓、地衣、藻类、真菌等土壤微生物在土壤表层与微小颗粒胶结形成的复合体[9].受水分条件的限制,干旱沙区无法支撑连片的、高密度的维管束植物群落,地表往往呈现出由维管束植物和生物结皮斑块镶嵌分布的格局[10-11].随着植被定植和流沙固定,地表结皮生物的盖度和生物量均发生不同程度的变化,表现出独特的演变规律[12].生物结皮演替通常始于稳定的表土环境[13],在风蚀[14]、水蚀[15]及沙埋[16]等严重干扰环境中很难形成生物结皮.在稳定表土环境中蓝藻首先拓殖,其代谢产物帮助形成土壤具有生物活性团聚结构,自此标志着生物结皮的初步形成[17].根据生物结皮演替过程中优势物种差异,可将其分为3个阶段,即以蓝藻为优势的演替初期阶段,以绿藻和蓝藻为优势的演替中期和以苔藓、地衣为优势的演替后期阶段[18].随着生物结皮的演替,结皮生物之间的种间关系也发生相应的变化,进而引发地表稳定性、水分、养分、土壤颗粒组成等变化,从而影响整个生态系统的稳定性[9].因此,探究结皮生物之间的关系随生境变化规律对全面理解沙区生态系统物种间关系及对生态系统结构与功能稳定性影响具有重要意义. ...
The effects of extreme rainfall events on carbon release from biological soil crusts covered soil in fixed sand dunes in the Tengger Desert,northern China
1
2013
... 生物土壤结皮(简称生物结皮)是由苔藓、地衣、藻类、真菌等土壤微生物在土壤表层与微小颗粒胶结形成的复合体[9].受水分条件的限制,干旱沙区无法支撑连片的、高密度的维管束植物群落,地表往往呈现出由维管束植物和生物结皮斑块镶嵌分布的格局[10-11].随着植被定植和流沙固定,地表结皮生物的盖度和生物量均发生不同程度的变化,表现出独特的演变规律[12].生物结皮演替通常始于稳定的表土环境[13],在风蚀[14]、水蚀[15]及沙埋[16]等严重干扰环境中很难形成生物结皮.在稳定表土环境中蓝藻首先拓殖,其代谢产物帮助形成土壤具有生物活性团聚结构,自此标志着生物结皮的初步形成[17].根据生物结皮演替过程中优势物种差异,可将其分为3个阶段,即以蓝藻为优势的演替初期阶段,以绿藻和蓝藻为优势的演替中期和以苔藓、地衣为优势的演替后期阶段[18].随着生物结皮的演替,结皮生物之间的种间关系也发生相应的变化,进而引发地表稳定性、水分、养分、土壤颗粒组成等变化,从而影响整个生态系统的稳定性[9].因此,探究结皮生物之间的关系随生境变化规律对全面理解沙区生态系统物种间关系及对生态系统结构与功能稳定性影响具有重要意义. ...
Responses of biological soil crusts to sand burial in a revegetated area of the Tengger Desert,Northern China
1
2008
... 生物土壤结皮(简称生物结皮)是由苔藓、地衣、藻类、真菌等土壤微生物在土壤表层与微小颗粒胶结形成的复合体[9].受水分条件的限制,干旱沙区无法支撑连片的、高密度的维管束植物群落,地表往往呈现出由维管束植物和生物结皮斑块镶嵌分布的格局[10-11].随着植被定植和流沙固定,地表结皮生物的盖度和生物量均发生不同程度的变化,表现出独特的演变规律[12].生物结皮演替通常始于稳定的表土环境[13],在风蚀[14]、水蚀[15]及沙埋[16]等严重干扰环境中很难形成生物结皮.在稳定表土环境中蓝藻首先拓殖,其代谢产物帮助形成土壤具有生物活性团聚结构,自此标志着生物结皮的初步形成[17].根据生物结皮演替过程中优势物种差异,可将其分为3个阶段,即以蓝藻为优势的演替初期阶段,以绿藻和蓝藻为优势的演替中期和以苔藓、地衣为优势的演替后期阶段[18].随着生物结皮的演替,结皮生物之间的种间关系也发生相应的变化,进而引发地表稳定性、水分、养分、土壤颗粒组成等变化,从而影响整个生态系统的稳定性[9].因此,探究结皮生物之间的关系随生境变化规律对全面理解沙区生态系统物种间关系及对生态系统结构与功能稳定性影响具有重要意义. ...
Response of desert biological soil crusts to alterations in precipitation frequency
1
2004
... 生物土壤结皮(简称生物结皮)是由苔藓、地衣、藻类、真菌等土壤微生物在土壤表层与微小颗粒胶结形成的复合体[9].受水分条件的限制,干旱沙区无法支撑连片的、高密度的维管束植物群落,地表往往呈现出由维管束植物和生物结皮斑块镶嵌分布的格局[10-11].随着植被定植和流沙固定,地表结皮生物的盖度和生物量均发生不同程度的变化,表现出独特的演变规律[12].生物结皮演替通常始于稳定的表土环境[13],在风蚀[14]、水蚀[15]及沙埋[16]等严重干扰环境中很难形成生物结皮.在稳定表土环境中蓝藻首先拓殖,其代谢产物帮助形成土壤具有生物活性团聚结构,自此标志着生物结皮的初步形成[17].根据生物结皮演替过程中优势物种差异,可将其分为3个阶段,即以蓝藻为优势的演替初期阶段,以绿藻和蓝藻为优势的演替中期和以苔藓、地衣为优势的演替后期阶段[18].随着生物结皮的演替,结皮生物之间的种间关系也发生相应的变化,进而引发地表稳定性、水分、养分、土壤颗粒组成等变化,从而影响整个生态系统的稳定性[9].因此,探究结皮生物之间的关系随生境变化规律对全面理解沙区生态系统物种间关系及对生态系统结构与功能稳定性影响具有重要意义. ...
Do biological soil crusts determine vegetation changes in sandy deserts?Implications for managing artificial vegetation
3
2010
... 生物土壤结皮(简称生物结皮)是由苔藓、地衣、藻类、真菌等土壤微生物在土壤表层与微小颗粒胶结形成的复合体[9].受水分条件的限制,干旱沙区无法支撑连片的、高密度的维管束植物群落,地表往往呈现出由维管束植物和生物结皮斑块镶嵌分布的格局[10-11].随着植被定植和流沙固定,地表结皮生物的盖度和生物量均发生不同程度的变化,表现出独特的演变规律[12].生物结皮演替通常始于稳定的表土环境[13],在风蚀[14]、水蚀[15]及沙埋[16]等严重干扰环境中很难形成生物结皮.在稳定表土环境中蓝藻首先拓殖,其代谢产物帮助形成土壤具有生物活性团聚结构,自此标志着生物结皮的初步形成[17].根据生物结皮演替过程中优势物种差异,可将其分为3个阶段,即以蓝藻为优势的演替初期阶段,以绿藻和蓝藻为优势的演替中期和以苔藓、地衣为优势的演替后期阶段[18].随着生物结皮的演替,结皮生物之间的种间关系也发生相应的变化,进而引发地表稳定性、水分、养分、土壤颗粒组成等变化,从而影响整个生态系统的稳定性[9].因此,探究结皮生物之间的关系随生境变化规律对全面理解沙区生态系统物种间关系及对生态系统结构与功能稳定性影响具有重要意义. ...
... 研究区位于腾格里沙漠东南缘沙坡头人工固沙植被区(37°32′N、105°02′E,海拔1 330 m).该区多年平均气温9.6 ℃,多年平均降水量186.2 mm(1956—2012年),集中在5—9月[19].为确保包兰铁路沙坡头段的安全通行,自1956年起在铁路两侧建立人工固沙植被防护体系,并于1964、1981、1987年多次扩建.不同年龄固沙区的固沙措施相同(包括采用的草方格、种植的旱生灌木种类组成、空间配置和密度与苗龄等)[20].各演替阶段固沙植被区植被以柠条锦鸡儿(Caragana korshinskii)、细枝岩黄耆(Hedysarum scoparium)、油蒿(Artemisia ordosica)、雾冰藜(Bassia dasyphylla)等为主[19].结皮生物广泛分布于该地区,类型多样,定居其间的结皮生物主要有具鞘微鞘藻(Microcoleus vaginatus)、双尖菱板藻(Hantzschia amphioxys)、隐头舟形藻(Navicula cryptocephala)等藻类和真藓(Bryum argenteum)、土生对齿藓(Didymodon vinealis)、齿肋赤藓(Syntrichia caninervis)等藓类及球胶衣(Collema coccophorum)、石果衣(Endocarpon pusillum)、坚韧胶衣(Collema tenax)等地衣[18]. ...
... 生态位宽度是反映物种对环境资源利用状况的尺度[21],生态位重叠则反映了种群之间对资源利用的相似程度和竞争关系[1,3].随着沙坡头地区固沙植被演替的进行,藻类生态位持续变窄(图1).根据结皮生物正向演替的一般规律模式[18],藻类为该群落的先锋类群,最先占据生境空斑,随后其代谢产物胶结沙粒等颗粒物质形成具有生物活性的团聚结构,促进演替后期物种的拓殖.根据植物CSR分类特征[22],藻类作为胁迫忍耐对策者,通常具有更宽的基础生态位,固沙植被演替后期藻类生态位宽度的降低可能源自演替后期物种对其竞争导致的实际生态位收缩.蓝藻在固沙植被演替前期与球胶衣和真藓有一定生态位重叠(图2),与其他结皮生物重叠值很小,这可能与藻类自身特性有关[22]. ...
我国北方风沙危害区生态重建与恢复:腾格里沙漠土壤水分与植被承载力的探讨
2
2014
... 研究区位于腾格里沙漠东南缘沙坡头人工固沙植被区(37°32′N、105°02′E,海拔1 330 m).该区多年平均气温9.6 ℃,多年平均降水量186.2 mm(1956—2012年),集中在5—9月[19].为确保包兰铁路沙坡头段的安全通行,自1956年起在铁路两侧建立人工固沙植被防护体系,并于1964、1981、1987年多次扩建.不同年龄固沙区的固沙措施相同(包括采用的草方格、种植的旱生灌木种类组成、空间配置和密度与苗龄等)[20].各演替阶段固沙植被区植被以柠条锦鸡儿(Caragana korshinskii)、细枝岩黄耆(Hedysarum scoparium)、油蒿(Artemisia ordosica)、雾冰藜(Bassia dasyphylla)等为主[19].结皮生物广泛分布于该地区,类型多样,定居其间的结皮生物主要有具鞘微鞘藻(Microcoleus vaginatus)、双尖菱板藻(Hantzschia amphioxys)、隐头舟形藻(Navicula cryptocephala)等藻类和真藓(Bryum argenteum)、土生对齿藓(Didymodon vinealis)、齿肋赤藓(Syntrichia caninervis)等藓类及球胶衣(Collema coccophorum)、石果衣(Endocarpon pusillum)、坚韧胶衣(Collema tenax)等地衣[18]. ...
... [19].结皮生物广泛分布于该地区,类型多样,定居其间的结皮生物主要有具鞘微鞘藻(Microcoleus vaginatus)、双尖菱板藻(Hantzschia amphioxys)、隐头舟形藻(Navicula cryptocephala)等藻类和真藓(Bryum argenteum)、土生对齿藓(Didymodon vinealis)、齿肋赤藓(Syntrichia caninervis)等藓类及球胶衣(Collema coccophorum)、石果衣(Endocarpon pusillum)、坚韧胶衣(Collema tenax)等地衣[18]. ...
Recovery of topsoil physicochemical properties in revegetated sites in the sand-burial ecosystems of the Tengger Desert,Northern China
1
2007
... 研究区位于腾格里沙漠东南缘沙坡头人工固沙植被区(37°32′N、105°02′E,海拔1 330 m).该区多年平均气温9.6 ℃,多年平均降水量186.2 mm(1956—2012年),集中在5—9月[19].为确保包兰铁路沙坡头段的安全通行,自1956年起在铁路两侧建立人工固沙植被防护体系,并于1964、1981、1987年多次扩建.不同年龄固沙区的固沙措施相同(包括采用的草方格、种植的旱生灌木种类组成、空间配置和密度与苗龄等)[20].各演替阶段固沙植被区植被以柠条锦鸡儿(Caragana korshinskii)、细枝岩黄耆(Hedysarum scoparium)、油蒿(Artemisia ordosica)、雾冰藜(Bassia dasyphylla)等为主[19].结皮生物广泛分布于该地区,类型多样,定居其间的结皮生物主要有具鞘微鞘藻(Microcoleus vaginatus)、双尖菱板藻(Hantzschia amphioxys)、隐头舟形藻(Navicula cryptocephala)等藻类和真藓(Bryum argenteum)、土生对齿藓(Didymodon vinealis)、齿肋赤藓(Syntrichia caninervis)等藓类及球胶衣(Collema coccophorum)、石果衣(Endocarpon pusillum)、坚韧胶衣(Collema tenax)等地衣[18]. ...
Niche breadth,rarity and ecological characteristics within a regional flora spanning large environmental gradients
1
2015
... 生态位宽度是反映物种对环境资源利用状况的尺度[21],生态位重叠则反映了种群之间对资源利用的相似程度和竞争关系[1,3].随着沙坡头地区固沙植被演替的进行,藻类生态位持续变窄(图1).根据结皮生物正向演替的一般规律模式[18],藻类为该群落的先锋类群,最先占据生境空斑,随后其代谢产物胶结沙粒等颗粒物质形成具有生物活性的团聚结构,促进演替后期物种的拓殖.根据植物CSR分类特征[22],藻类作为胁迫忍耐对策者,通常具有更宽的基础生态位,固沙植被演替后期藻类生态位宽度的降低可能源自演替后期物种对其竞争导致的实际生态位收缩.蓝藻在固沙植被演替前期与球胶衣和真藓有一定生态位重叠(图2),与其他结皮生物重叠值很小,这可能与藻类自身特性有关[22]. ...
Ecological classification (book reviews:plant strategies and vegetation processes)
2
1979
... 生态位宽度是反映物种对环境资源利用状况的尺度[21],生态位重叠则反映了种群之间对资源利用的相似程度和竞争关系[1,3].随着沙坡头地区固沙植被演替的进行,藻类生态位持续变窄(图1).根据结皮生物正向演替的一般规律模式[18],藻类为该群落的先锋类群,最先占据生境空斑,随后其代谢产物胶结沙粒等颗粒物质形成具有生物活性的团聚结构,促进演替后期物种的拓殖.根据植物CSR分类特征[22],藻类作为胁迫忍耐对策者,通常具有更宽的基础生态位,固沙植被演替后期藻类生态位宽度的降低可能源自演替后期物种对其竞争导致的实际生态位收缩.蓝藻在固沙植被演替前期与球胶衣和真藓有一定生态位重叠(图2),与其他结皮生物重叠值很小,这可能与藻类自身特性有关[22]. ...
... [22]. ...
柴达木盆地东部荒漠植物生态位特征
1
2019
... 生态位宽度越大,对环境的适应能力越强,同时伴随着较高的生态位重叠[23].沙坡头地区固沙植被演替过程中,真藓与球胶衣生态位宽度基本保持不变且高于其他种(图1).真藓与球胶衣为该区最主要的优势结皮,它们的生态位宽度在4个固沙植被演替阶段上均高于其他结皮生物类型,利用资源的能力较强,能更好地适应当地环境,为泛化种,且二者生态位重叠最高(图2),表明二者利用的资源具有很高的相似性.通常,生态位宽度较小的物种则因对资源的利用能力较弱,分布较窄而与其他物种间有较低的生态位重叠[24].在我们的研究区,土生对齿藓、齿肋赤藓和石果衣生态位宽度在固沙植被演替早期(演替33年)很小,说明它们的生长对水分和土壤条件要求较高,固沙植被演替前期较恶劣的环境不适合它们生长,对环境适应能力较差限制了它们的拓殖,从而导致与其他结皮生物有较低生态位重叠,竞争较弱;随着固沙植被演替的进行,土生对齿藓、齿肋赤藓、石果衣生态位宽度明显变宽,这可能是由于固沙植被演替引起的水分条件和土壤条件改善使它们更好地利用资源占据生长空间,且演替后期它们与优势物种产生了更多的生态位重叠(图2),对资源利用的相似性导致对资源的竞争加强. ...
安徽九华山风景区古树群落主要种群生态位的动态变化
1
2019
... 生态位宽度越大,对环境的适应能力越强,同时伴随着较高的生态位重叠[23].沙坡头地区固沙植被演替过程中,真藓与球胶衣生态位宽度基本保持不变且高于其他种(图1).真藓与球胶衣为该区最主要的优势结皮,它们的生态位宽度在4个固沙植被演替阶段上均高于其他结皮生物类型,利用资源的能力较强,能更好地适应当地环境,为泛化种,且二者生态位重叠最高(图2),表明二者利用的资源具有很高的相似性.通常,生态位宽度较小的物种则因对资源的利用能力较弱,分布较窄而与其他物种间有较低的生态位重叠[24].在我们的研究区,土生对齿藓、齿肋赤藓和石果衣生态位宽度在固沙植被演替早期(演替33年)很小,说明它们的生长对水分和土壤条件要求较高,固沙植被演替前期较恶劣的环境不适合它们生长,对环境适应能力较差限制了它们的拓殖,从而导致与其他结皮生物有较低生态位重叠,竞争较弱;随着固沙植被演替的进行,土生对齿藓、齿肋赤藓、石果衣生态位宽度明显变宽,这可能是由于固沙植被演替引起的水分条件和土壤条件改善使它们更好地利用资源占据生长空间,且演替后期它们与优势物种产生了更多的生态位重叠(图2),对资源利用的相似性导致对资源的竞争加强. ...
苏门自酒草对乡土植物群落种间联结性及稳定性的影响
1
2011
... 种间关联是不同种类在空间分布上的关联性,通常由不同物种的资源利用方式和种间关系决定[3].植物种间关联是植物群落重要的数量和结构特征,是植物群落结构形成和演化的基础[3].本研究中,不同结皮生物间的种间关联性质和强度随着固沙植被演替的进行不断发生变化(图3).蓝藻在固沙植被演替初期与球胶衣有较弱正关联,这可能得因于二者在前期资源匮乏时的相互协作,随着固沙植被演替进行,蓝藻与其他结皮生物均呈现出负关联,这可能是竞争排除作用所致.球胶衣与石果衣之间随着固沙植被演替进行,由正关联向负关联转变,这可能由于生境状况变好,二者相似的生态学特性和生物学特性导致相互之间竞争有限的环境资源和空间资源而产生竞争和排斥.石果衣与土生对齿藓的种间关联随着固沙植被演替呈现出由负转正的现象.结皮生物间的种间关联状态(图3)与生态位重叠关系基本一致(图2),因此可以推断资源利用方式是导致其种间关联的主要因素.群落的种间关联状况在一定程度上反映了群落的稳定性[25-26].我们的计算结果显示,沙坡头固沙植被区结皮生物种间总体关联性随着固沙植被演替由负关联向正关联转变,且在固沙植被演替后期产生了更广泛的正关联(表1),说明沙坡头地区不同结皮生物间随着固沙植被演替产生更强的联系,群落稳定性更强. ...
植物种间联结研究内容与方法评述
2
2016
... 种间关联是不同种类在空间分布上的关联性,通常由不同物种的资源利用方式和种间关系决定[3].植物种间关联是植物群落重要的数量和结构特征,是植物群落结构形成和演化的基础[3].本研究中,不同结皮生物间的种间关联性质和强度随着固沙植被演替的进行不断发生变化(图3).蓝藻在固沙植被演替初期与球胶衣有较弱正关联,这可能得因于二者在前期资源匮乏时的相互协作,随着固沙植被演替进行,蓝藻与其他结皮生物均呈现出负关联,这可能是竞争排除作用所致.球胶衣与石果衣之间随着固沙植被演替进行,由正关联向负关联转变,这可能由于生境状况变好,二者相似的生态学特性和生物学特性导致相互之间竞争有限的环境资源和空间资源而产生竞争和排斥.石果衣与土生对齿藓的种间关联随着固沙植被演替呈现出由负转正的现象.结皮生物间的种间关联状态(图3)与生态位重叠关系基本一致(图2),因此可以推断资源利用方式是导致其种间关联的主要因素.群落的种间关联状况在一定程度上反映了群落的稳定性[25-26].我们的计算结果显示,沙坡头固沙植被区结皮生物种间总体关联性随着固沙植被演替由负关联向正关联转变,且在固沙植被演替后期产生了更广泛的正关联(表1),说明沙坡头地区不同结皮生物间随着固沙植被演替产生更强的联系,群落稳定性更强. ...
... 结皮生物的多样性、盖度和空间分布等对环境变化及干扰的响应十分敏感[27-30].沙坡头结皮生物组成和演变受到很多因素的影响,自然因素如气候、植被、土壤养分、水分等均为影响沙坡头生物多样性发展及生态系统恢复的主要因素[31-35],我们在设置样方时也有意避开或减少了植被的影响,将其设置在灌丛间.沙坡头人工固沙植被演替过程中,植被、土壤等环境的变化形成自然环境梯度[13].我们发现,当胁迫梯度增加,种间由竞争转向互惠,这与胁迫梯度假说[6]相一致.随着沙坡头固沙植被演替的进行,胁迫程度逐步降低,此时种间竞争加剧,可能有助于维持群落稳定,且土壤质地由粗变细、土壤持水量也呈现出逐步增加的趋势[36],因此可以预测,在沙坡头地区蓝藻会逐步被其他结皮生物所替代,地衣与藓类的数量会进一步增加,同时齿肋赤藓会逐步替代真藓成为该区主要的藓结皮生物.计算种间关联系数只是对种间关系的定量描述,取样尺度、种间竞争、化感作用、环境因素等都会对种间关联性产生影响[26].因此,对沙坡头人工固沙植被演替过程中结皮生物种间关联的内在机制还需进一步从植物生理、遗传学、分子生态学等多角度开展研究,得出更为合理的解释.沙区生态系统的恢复是漫长的过程,受多种自然因素与人为因素的影响,长期的监测和研究十分必要[13],沙坡头地区固沙植被演替过程中结皮生物生态位与种间关联规律还有待进一步观察研究. ...
Drought and grazing combined:contrasting shifts in plant interactions at species pair and community level
1
2014
... 结皮生物的多样性、盖度和空间分布等对环境变化及干扰的响应十分敏感[27-30].沙坡头结皮生物组成和演变受到很多因素的影响,自然因素如气候、植被、土壤养分、水分等均为影响沙坡头生物多样性发展及生态系统恢复的主要因素[31-35],我们在设置样方时也有意避开或减少了植被的影响,将其设置在灌丛间.沙坡头人工固沙植被演替过程中,植被、土壤等环境的变化形成自然环境梯度[13].我们发现,当胁迫梯度增加,种间由竞争转向互惠,这与胁迫梯度假说[6]相一致.随着沙坡头固沙植被演替的进行,胁迫程度逐步降低,此时种间竞争加剧,可能有助于维持群落稳定,且土壤质地由粗变细、土壤持水量也呈现出逐步增加的趋势[36],因此可以预测,在沙坡头地区蓝藻会逐步被其他结皮生物所替代,地衣与藓类的数量会进一步增加,同时齿肋赤藓会逐步替代真藓成为该区主要的藓结皮生物.计算种间关联系数只是对种间关系的定量描述,取样尺度、种间竞争、化感作用、环境因素等都会对种间关联性产生影响[26].因此,对沙坡头人工固沙植被演替过程中结皮生物种间关联的内在机制还需进一步从植物生理、遗传学、分子生态学等多角度开展研究,得出更为合理的解释.沙区生态系统的恢复是漫长的过程,受多种自然因素与人为因素的影响,长期的监测和研究十分必要[13],沙坡头地区固沙植被演替过程中结皮生物生态位与种间关联规律还有待进一步观察研究. ...
Regional patterns of biological soil crust lichen species composition related tovegetation,soils,and climate in Oregon,USA
0
2012
黄土丘陵区生物结皮土壤生态化学计量学特征及其对干扰的响应
0
2019
黄土丘陵区草地根系生物量对踩踏干扰生物结皮的响应
1
2019
... 结皮生物的多样性、盖度和空间分布等对环境变化及干扰的响应十分敏感[27-30].沙坡头结皮生物组成和演变受到很多因素的影响,自然因素如气候、植被、土壤养分、水分等均为影响沙坡头生物多样性发展及生态系统恢复的主要因素[31-35],我们在设置样方时也有意避开或减少了植被的影响,将其设置在灌丛间.沙坡头人工固沙植被演替过程中,植被、土壤等环境的变化形成自然环境梯度[13].我们发现,当胁迫梯度增加,种间由竞争转向互惠,这与胁迫梯度假说[6]相一致.随着沙坡头固沙植被演替的进行,胁迫程度逐步降低,此时种间竞争加剧,可能有助于维持群落稳定,且土壤质地由粗变细、土壤持水量也呈现出逐步增加的趋势[36],因此可以预测,在沙坡头地区蓝藻会逐步被其他结皮生物所替代,地衣与藓类的数量会进一步增加,同时齿肋赤藓会逐步替代真藓成为该区主要的藓结皮生物.计算种间关联系数只是对种间关系的定量描述,取样尺度、种间竞争、化感作用、环境因素等都会对种间关联性产生影响[26].因此,对沙坡头人工固沙植被演替过程中结皮生物种间关联的内在机制还需进一步从植物生理、遗传学、分子生态学等多角度开展研究,得出更为合理的解释.沙区生态系统的恢复是漫长的过程,受多种自然因素与人为因素的影响,长期的监测和研究十分必要[13],沙坡头地区固沙植被演替过程中结皮生物生态位与种间关联规律还有待进一步观察研究. ...
Study on soil microbiotic crust and its influences on sand fixing vegetation in arid desert region
1
2000
... 结皮生物的多样性、盖度和空间分布等对环境变化及干扰的响应十分敏感[27-30].沙坡头结皮生物组成和演变受到很多因素的影响,自然因素如气候、植被、土壤养分、水分等均为影响沙坡头生物多样性发展及生态系统恢复的主要因素[31-35],我们在设置样方时也有意避开或减少了植被的影响,将其设置在灌丛间.沙坡头人工固沙植被演替过程中,植被、土壤等环境的变化形成自然环境梯度[13].我们发现,当胁迫梯度增加,种间由竞争转向互惠,这与胁迫梯度假说[6]相一致.随着沙坡头固沙植被演替的进行,胁迫程度逐步降低,此时种间竞争加剧,可能有助于维持群落稳定,且土壤质地由粗变细、土壤持水量也呈现出逐步增加的趋势[36],因此可以预测,在沙坡头地区蓝藻会逐步被其他结皮生物所替代,地衣与藓类的数量会进一步增加,同时齿肋赤藓会逐步替代真藓成为该区主要的藓结皮生物.计算种间关联系数只是对种间关系的定量描述,取样尺度、种间竞争、化感作用、环境因素等都会对种间关联性产生影响[26].因此,对沙坡头人工固沙植被演替过程中结皮生物种间关联的内在机制还需进一步从植物生理、遗传学、分子生态学等多角度开展研究,得出更为合理的解释.沙区生态系统的恢复是漫长的过程,受多种自然因素与人为因素的影响,长期的监测和研究十分必要[13],沙坡头地区固沙植被演替过程中结皮生物生态位与种间关联规律还有待进一步观察研究. ...
Ecological and environmental explanation of microbiotic crusts on sand dune scales in the Gurbantunggut Desert,Xinjiang
0
2005
Microbiotic crusts and their interrelations with environmental factors in the Gurbantunggut desert,western China
0
2007
Analysis of environmental factors determining development and succession in biological soil crusts
0
2015
Topographic differentiations of biological soil crusts and hydraulic properties in fixed sand dunes,Tengger Desert
1
2015
... 结皮生物的多样性、盖度和空间分布等对环境变化及干扰的响应十分敏感[27-30].沙坡头结皮生物组成和演变受到很多因素的影响,自然因素如气候、植被、土壤养分、水分等均为影响沙坡头生物多样性发展及生态系统恢复的主要因素[31-35],我们在设置样方时也有意避开或减少了植被的影响,将其设置在灌丛间.沙坡头人工固沙植被演替过程中,植被、土壤等环境的变化形成自然环境梯度[13].我们发现,当胁迫梯度增加,种间由竞争转向互惠,这与胁迫梯度假说[6]相一致.随着沙坡头固沙植被演替的进行,胁迫程度逐步降低,此时种间竞争加剧,可能有助于维持群落稳定,且土壤质地由粗变细、土壤持水量也呈现出逐步增加的趋势[36],因此可以预测,在沙坡头地区蓝藻会逐步被其他结皮生物所替代,地衣与藓类的数量会进一步增加,同时齿肋赤藓会逐步替代真藓成为该区主要的藓结皮生物.计算种间关联系数只是对种间关系的定量描述,取样尺度、种间竞争、化感作用、环境因素等都会对种间关联性产生影响[26].因此,对沙坡头人工固沙植被演替过程中结皮生物种间关联的内在机制还需进一步从植物生理、遗传学、分子生态学等多角度开展研究,得出更为合理的解释.沙区生态系统的恢复是漫长的过程,受多种自然因素与人为因素的影响,长期的监测和研究十分必要[13],沙坡头地区固沙植被演替过程中结皮生物生态位与种间关联规律还有待进一步观察研究. ...
Precipitation and topsoil attributes determine the species diversity and distribution patterns of crustal communities in desert ecosystems
1
2017
... 结皮生物的多样性、盖度和空间分布等对环境变化及干扰的响应十分敏感[27-30].沙坡头结皮生物组成和演变受到很多因素的影响,自然因素如气候、植被、土壤养分、水分等均为影响沙坡头生物多样性发展及生态系统恢复的主要因素[31-35],我们在设置样方时也有意避开或减少了植被的影响,将其设置在灌丛间.沙坡头人工固沙植被演替过程中,植被、土壤等环境的变化形成自然环境梯度[13].我们发现,当胁迫梯度增加,种间由竞争转向互惠,这与胁迫梯度假说[6]相一致.随着沙坡头固沙植被演替的进行,胁迫程度逐步降低,此时种间竞争加剧,可能有助于维持群落稳定,且土壤质地由粗变细、土壤持水量也呈现出逐步增加的趋势[36],因此可以预测,在沙坡头地区蓝藻会逐步被其他结皮生物所替代,地衣与藓类的数量会进一步增加,同时齿肋赤藓会逐步替代真藓成为该区主要的藓结皮生物.计算种间关联系数只是对种间关系的定量描述,取样尺度、种间竞争、化感作用、环境因素等都会对种间关联性产生影响[26].因此,对沙坡头人工固沙植被演替过程中结皮生物种间关联的内在机制还需进一步从植物生理、遗传学、分子生态学等多角度开展研究,得出更为合理的解释.沙区生态系统的恢复是漫长的过程,受多种自然因素与人为因素的影响,长期的监测和研究十分必要[13],沙坡头地区固沙植被演替过程中结皮生物生态位与种间关联规律还有待进一步观察研究. ...
生物土壤结皮研究:进展、前沿与展望
1
2009
... 生物结皮作为干旱沙区生态系统的重要组成部分,在维护沙区生态系统稳定性中的作用不容忽视[37].干旱沙区生物结皮演替过程中,深入理解结皮生物的生态位以及种间关联的变化对于沙区生态重建和恢复十分重要.随着固沙植被演替,正关联性增强是沙坡头人工固沙植被稳定的重要原因,在进行生态恢复重建中对结皮生物物种选择组合时要加以考虑,前期建议使用蓝藻进行接种,随着蓝藻定植所带来的土壤养分和水分条件的改善,再施配以藓结皮及地衣结皮.依据结皮生物种间关联随固沙植被演替过程变化特征(图3),具体结皮生物组合接种策略为:前期可以选择蓝藻、球胶衣-石果衣组合,中期选择球胶衣-石果衣、土生对齿藓-齿肋赤藓组合以及后期选择球胶衣-土生对齿藓、球胶衣-齿肋赤藓组合. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}