1
1999
... 生物多样性随环境梯度的变化规律是生物多样性研究的重要内容[1-2].物种多样性是生物多样性研究中最基础的部分,是群落功能复杂性与稳定性的重要度量指标[3-4].关于植物群落物种多样性的研究历来为国内外学者所重视[5-6],了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义[7].环境因素包括经纬度、海拔、气候、土壤、地形等[8],其中经纬度和海拔常用于大尺度物种多样性分布格局的研究[9];在区域尺度上,地形、气候、土壤3种环境要素的不同组合产生了复杂的群落物种多样性格局[10].探讨群落物种多样性的特征和影响因素,对维护生态系统的结构和功能的稳定性具有重要意义[11-13]. ...
生物多样性科学的热点问题
1
2016
... 生物多样性随环境梯度的变化规律是生物多样性研究的重要内容[1-2].物种多样性是生物多样性研究中最基础的部分,是群落功能复杂性与稳定性的重要度量指标[3-4].关于植物群落物种多样性的研究历来为国内外学者所重视[5-6],了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义[7].环境因素包括经纬度、海拔、气候、土壤、地形等[8],其中经纬度和海拔常用于大尺度物种多样性分布格局的研究[9];在区域尺度上,地形、气候、土壤3种环境要素的不同组合产生了复杂的群落物种多样性格局[10].探讨群落物种多样性的特征和影响因素,对维护生态系统的结构和功能的稳定性具有重要意义[11-13]. ...
北京东灵山地区植物群落多样性的研究Ⅺ.山地草甸β多样性
1
1998
... 生物多样性随环境梯度的变化规律是生物多样性研究的重要内容[1-2].物种多样性是生物多样性研究中最基础的部分,是群落功能复杂性与稳定性的重要度量指标[3-4].关于植物群落物种多样性的研究历来为国内外学者所重视[5-6],了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义[7].环境因素包括经纬度、海拔、气候、土壤、地形等[8],其中经纬度和海拔常用于大尺度物种多样性分布格局的研究[9];在区域尺度上,地形、气候、土壤3种环境要素的不同组合产生了复杂的群落物种多样性格局[10].探讨群落物种多样性的特征和影响因素,对维护生态系统的结构和功能的稳定性具有重要意义[11-13]. ...
Environmental drivers of plant distributions at global and regional scales
1
2021
... 生物多样性随环境梯度的变化规律是生物多样性研究的重要内容[1-2].物种多样性是生物多样性研究中最基础的部分,是群落功能复杂性与稳定性的重要度量指标[3-4].关于植物群落物种多样性的研究历来为国内外学者所重视[5-6],了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义[7].环境因素包括经纬度、海拔、气候、土壤、地形等[8],其中经纬度和海拔常用于大尺度物种多样性分布格局的研究[9];在区域尺度上,地形、气候、土壤3种环境要素的不同组合产生了复杂的群落物种多样性格局[10].探讨群落物种多样性的特征和影响因素,对维护生态系统的结构和功能的稳定性具有重要意义[11-13]. ...
Scale and species richness:towards a general,hierarchical theory of species diversity
1
2001
... 生物多样性随环境梯度的变化规律是生物多样性研究的重要内容[1-2].物种多样性是生物多样性研究中最基础的部分,是群落功能复杂性与稳定性的重要度量指标[3-4].关于植物群落物种多样性的研究历来为国内外学者所重视[5-6],了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义[7].环境因素包括经纬度、海拔、气候、土壤、地形等[8],其中经纬度和海拔常用于大尺度物种多样性分布格局的研究[9];在区域尺度上,地形、气候、土壤3种环境要素的不同组合产生了复杂的群落物种多样性格局[10].探讨群落物种多样性的特征和影响因素,对维护生态系统的结构和功能的稳定性具有重要意义[11-13]. ...
Toward a better understanding of the regional causes of local community richness
1
2008
... 生物多样性随环境梯度的变化规律是生物多样性研究的重要内容[1-2].物种多样性是生物多样性研究中最基础的部分,是群落功能复杂性与稳定性的重要度量指标[3-4].关于植物群落物种多样性的研究历来为国内外学者所重视[5-6],了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义[7].环境因素包括经纬度、海拔、气候、土壤、地形等[8],其中经纬度和海拔常用于大尺度物种多样性分布格局的研究[9];在区域尺度上,地形、气候、土壤3种环境要素的不同组合产生了复杂的群落物种多样性格局[10].探讨群落物种多样性的特征和影响因素,对维护生态系统的结构和功能的稳定性具有重要意义[11-13]. ...
内蒙古西部自然植被的物种多样性及其影响因素
5
2017
... 生物多样性随环境梯度的变化规律是生物多样性研究的重要内容[1-2].物种多样性是生物多样性研究中最基础的部分,是群落功能复杂性与稳定性的重要度量指标[3-4].关于植物群落物种多样性的研究历来为国内外学者所重视[5-6],了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义[7].环境因素包括经纬度、海拔、气候、土壤、地形等[8],其中经纬度和海拔常用于大尺度物种多样性分布格局的研究[9];在区域尺度上,地形、气候、土壤3种环境要素的不同组合产生了复杂的群落物种多样性格局[10].探讨群落物种多样性的特征和影响因素,对维护生态系统的结构和功能的稳定性具有重要意义[11-13]. ...
... 采用Patrick指数、Simpson指数、Shannon-Wiener指数和Pielou指数测度浑善达克沙地不同植物群落的物种多样性程度[40-41].根据样方中每个物种的高度、盖度和频度计算出重要值,作为计算多样性指数的基础[7]. ...
... 物种多样性具有丰富性和均匀性两方面内容[40],Patrick指数是最常用的丰富度指数,本文研究表明,浑善达克沙地的物种丰富度程度最高的区域在43°N、116°E、海拔1 300 m附近,是正蓝旗榆树疏林分布的核心区[45].研究区Pielou指数整体较高,较高的均匀度有利于维持群落结构的稳定性[46].在均匀度差异不显著的情况下,Shannon-Wiener指数对群落的丰富度较为敏感[40].本研究显示,物种多样性指数的变化趋势与物种丰富度基本一致,表现为物种数越多,多样性指数越高,这与在内蒙古西部的研究基本一致[7]. ...
... 温度因子是影响植物群落物种多样性的重要因子[60].一般认为物种多样性与温度因子呈正相关关系[65],而在内蒙古干旱、半干旱区的部分研究显示,物种多样性却与年均温呈显著的负相关关系[7,61,66-68].本研究结果与后者基本一致,Patrick指数、Shannon-Wiener指数、Simpson指数和Pielou指数均与生长季均温呈显著负相关关系.这说明,温度因子确实对研究区内的物种多样性存在影响.呈现负相关的原因可能有两个:一是尺度问题,浑善达克沙地东西跨度大而南北跨度小,温度因子的梯度变化不明显,气候因子的解释率反而下降[67];二是生长季温度高的地方往往蒸发量也高,而蒸发量与水分因子呈现负相关[69].温度因子对物种多样性的影响还需进一步探讨. ...
... 土壤是决定小尺度植被分布的重要环境因子,沙地土壤质地、理化因子、微生物等都对植被的分布有直接的影响[70-71].土壤和植被是土壤-植被-大气连续体(Soil-Plant-Atmosphere-Continuum,SPAC)的共同组成部分[72],是相互依存、相互影响的因子,对二者之间关系的研究具有重要意义[67].目前针对这方面的研究很多,有研究显示土壤中氮含量与物种多样性呈正相关[73],也有试验显示氮素添加会降低物种多样性[74-75];还有研究指出,在贫瘠土壤上氮含量增加可以提高物种丰富度,而养分丰富的土壤提高含氮量可能会破坏原有生态系统的共存机制而造成物种丰富度的降低[76].位于内蒙古草原上的浑善达克沙地属于氮限制地区,土壤氮含量较高的区域群落的物种多样性相应也会提高[76].本研究结果基本一致,土壤有效氮与Patrick指数、Simpson指数和Shannon-Wiener指数均呈正相关关系.对土壤磷含量与物种多样性的研究显示,土壤磷含量与物种丰富度呈现负相关关系[67,77-78],本研究结果显示出类似的趋势,Shannon-Wiener指数随土壤全磷含量的增加而降低.土壤有机碳是表征土壤肥力的重要指标,不仅代表土壤有机质的水平,还能说明氮、磷等元素的可利用状态[79].土壤有机碳的含量越高,土壤肥力越高,可支持的物种多样性水平也越高.土壤有机碳与多样性指数呈现正相关[7,61,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...
植物群落物种多样性研究综述
1
2001
... 生物多样性随环境梯度的变化规律是生物多样性研究的重要内容[1-2].物种多样性是生物多样性研究中最基础的部分,是群落功能复杂性与稳定性的重要度量指标[3-4].关于植物群落物种多样性的研究历来为国内外学者所重视[5-6],了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义[7].环境因素包括经纬度、海拔、气候、土壤、地形等[8],其中经纬度和海拔常用于大尺度物种多样性分布格局的研究[9];在区域尺度上,地形、气候、土壤3种环境要素的不同组合产生了复杂的群落物种多样性格局[10].探讨群落物种多样性的特征和影响因素,对维护生态系统的结构和功能的稳定性具有重要意义[11-13]. ...
Productivity and sustainability influenced by biodiversity in grassland ecosystems
1
1996
... 生物多样性随环境梯度的变化规律是生物多样性研究的重要内容[1-2].物种多样性是生物多样性研究中最基础的部分,是群落功能复杂性与稳定性的重要度量指标[3-4].关于植物群落物种多样性的研究历来为国内外学者所重视[5-6],了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义[7].环境因素包括经纬度、海拔、气候、土壤、地形等[8],其中经纬度和海拔常用于大尺度物种多样性分布格局的研究[9];在区域尺度上,地形、气候、土壤3种环境要素的不同组合产生了复杂的群落物种多样性格局[10].探讨群落物种多样性的特征和影响因素,对维护生态系统的结构和功能的稳定性具有重要意义[11-13]. ...
Habitat association among Amazonian tree species:a landscape-scale approach
1
2003
... 生物多样性随环境梯度的变化规律是生物多样性研究的重要内容[1-2].物种多样性是生物多样性研究中最基础的部分,是群落功能复杂性与稳定性的重要度量指标[3-4].关于植物群落物种多样性的研究历来为国内外学者所重视[5-6],了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义[7].环境因素包括经纬度、海拔、气候、土壤、地形等[8],其中经纬度和海拔常用于大尺度物种多样性分布格局的研究[9];在区域尺度上,地形、气候、土壤3种环境要素的不同组合产生了复杂的群落物种多样性格局[10].探讨群落物种多样性的特征和影响因素,对维护生态系统的结构和功能的稳定性具有重要意义[11-13]. ...
Global surface temperature change
1
2010
... 生物多样性随环境梯度的变化规律是生物多样性研究的重要内容[1-2].物种多样性是生物多样性研究中最基础的部分,是群落功能复杂性与稳定性的重要度量指标[3-4].关于植物群落物种多样性的研究历来为国内外学者所重视[5-6],了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义[7].环境因素包括经纬度、海拔、气候、土壤、地形等[8],其中经纬度和海拔常用于大尺度物种多样性分布格局的研究[9];在区域尺度上,地形、气候、土壤3种环境要素的不同组合产生了复杂的群落物种多样性格局[10].探讨群落物种多样性的特征和影响因素,对维护生态系统的结构和功能的稳定性具有重要意义[11-13]. ...
Climate science:the cause of the pause
2013
Recent trends in inner Asian forest dynamics to temperature and precipitation indicate high sensitivity to climate change
1
2013
... 生物多样性随环境梯度的变化规律是生物多样性研究的重要内容[1-2].物种多样性是生物多样性研究中最基础的部分,是群落功能复杂性与稳定性的重要度量指标[3-4].关于植物群落物种多样性的研究历来为国内外学者所重视[5-6],了解影响植物群落物种多样性的关键因素及其相互关系对预测群落多样性的动态变化有着重要意义[7].环境因素包括经纬度、海拔、气候、土壤、地形等[8],其中经纬度和海拔常用于大尺度物种多样性分布格局的研究[9];在区域尺度上,地形、气候、土壤3种环境要素的不同组合产生了复杂的群落物种多样性格局[10].探讨群落物种多样性的特征和影响因素,对维护生态系统的结构和功能的稳定性具有重要意义[11-13]. ...
群落生态学迎来新的辉煌时代
1
2009
... 生态学家对物种多样性格局的形成机制提出了中域效应假说、种库假说、物种-能量假说、地史成因假说等一系列假说[14].其中以气候因素为基础的能量假说是目前最受关注的假说之一,寒冷忍耐假说认为随着冬季温度的降低物种多样性逐渐减少[15-16],生态学代谢假说试图从机制上解释环境温度与物种多样性地理格局的关系[17-18],水热动态假说认为物种多样性的大尺度格局由水分和热量共同决定[19-20].水分和热量的作用在不同生境与尺度上并不一致[21-22],在水分充足的区域,多样性主要受温度控制,而在水分亏缺的区域,物种多样性受水分和温度的共同制约[23].此外,土壤质地与肥力等因素也被证明对植物物种多样性格局存在重要作用[24-26].这些因子对物种多样性格局的作用在前期有一定的研究支持,但对环境因子的相对重要性尚存在一定的争议[27],对不同尺度上的物种多样性格局的形成机制并未达成共识. ...
Ecology's oldest pattern?
1
2001
... 生态学家对物种多样性格局的形成机制提出了中域效应假说、种库假说、物种-能量假说、地史成因假说等一系列假说[14].其中以气候因素为基础的能量假说是目前最受关注的假说之一,寒冷忍耐假说认为随着冬季温度的降低物种多样性逐渐减少[15-16],生态学代谢假说试图从机制上解释环境温度与物种多样性地理格局的关系[17-18],水热动态假说认为物种多样性的大尺度格局由水分和热量共同决定[19-20].水分和热量的作用在不同生境与尺度上并不一致[21-22],在水分充足的区域,多样性主要受温度控制,而在水分亏缺的区域,物种多样性受水分和温度的共同制约[23].此外,土壤质地与肥力等因素也被证明对植物物种多样性格局存在重要作用[24-26].这些因子对物种多样性格局的作用在前期有一定的研究支持,但对环境因子的相对重要性尚存在一定的争议[27],对不同尺度上的物种多样性格局的形成机制并未达成共识. ...
Productivity and history as predictors of the latitudinal diversity gradient of terrestrial birds
1
2003
... 生态学家对物种多样性格局的形成机制提出了中域效应假说、种库假说、物种-能量假说、地史成因假说等一系列假说[14].其中以气候因素为基础的能量假说是目前最受关注的假说之一,寒冷忍耐假说认为随着冬季温度的降低物种多样性逐渐减少[15-16],生态学代谢假说试图从机制上解释环境温度与物种多样性地理格局的关系[17-18],水热动态假说认为物种多样性的大尺度格局由水分和热量共同决定[19-20].水分和热量的作用在不同生境与尺度上并不一致[21-22],在水分充足的区域,多样性主要受温度控制,而在水分亏缺的区域,物种多样性受水分和温度的共同制约[23].此外,土壤质地与肥力等因素也被证明对植物物种多样性格局存在重要作用[24-26].这些因子对物种多样性格局的作用在前期有一定的研究支持,但对环境因子的相对重要性尚存在一定的争议[27],对不同尺度上的物种多样性格局的形成机制并未达成共识. ...
Global biodiversity,biochemical kinetics,and the energetic-equivalence rule
1
2002
... 生态学家对物种多样性格局的形成机制提出了中域效应假说、种库假说、物种-能量假说、地史成因假说等一系列假说[14].其中以气候因素为基础的能量假说是目前最受关注的假说之一,寒冷忍耐假说认为随着冬季温度的降低物种多样性逐渐减少[15-16],生态学代谢假说试图从机制上解释环境温度与物种多样性地理格局的关系[17-18],水热动态假说认为物种多样性的大尺度格局由水分和热量共同决定[19-20].水分和热量的作用在不同生境与尺度上并不一致[21-22],在水分充足的区域,多样性主要受温度控制,而在水分亏缺的区域,物种多样性受水分和温度的共同制约[23].此外,土壤质地与肥力等因素也被证明对植物物种多样性格局存在重要作用[24-26].这些因子对物种多样性格局的作用在前期有一定的研究支持,但对环境因子的相对重要性尚存在一定的争议[27],对不同尺度上的物种多样性格局的形成机制并未达成共识. ...
Advancing the metabolic theory of biodiversity
1
2009
... 生态学家对物种多样性格局的形成机制提出了中域效应假说、种库假说、物种-能量假说、地史成因假说等一系列假说[14].其中以气候因素为基础的能量假说是目前最受关注的假说之一,寒冷忍耐假说认为随着冬季温度的降低物种多样性逐渐减少[15-16],生态学代谢假说试图从机制上解释环境温度与物种多样性地理格局的关系[17-18],水热动态假说认为物种多样性的大尺度格局由水分和热量共同决定[19-20].水分和热量的作用在不同生境与尺度上并不一致[21-22],在水分充足的区域,多样性主要受温度控制,而在水分亏缺的区域,物种多样性受水分和温度的共同制约[23].此外,土壤质地与肥力等因素也被证明对植物物种多样性格局存在重要作用[24-26].这些因子对物种多样性格局的作用在前期有一定的研究支持,但对环境因子的相对重要性尚存在一定的争议[27],对不同尺度上的物种多样性格局的形成机制并未达成共识. ...
Water-energy dynamics,climate,and prediction of woody plant species richness:an interim general model
1
1998
... 生态学家对物种多样性格局的形成机制提出了中域效应假说、种库假说、物种-能量假说、地史成因假说等一系列假说[14].其中以气候因素为基础的能量假说是目前最受关注的假说之一,寒冷忍耐假说认为随着冬季温度的降低物种多样性逐渐减少[15-16],生态学代谢假说试图从机制上解释环境温度与物种多样性地理格局的关系[17-18],水热动态假说认为物种多样性的大尺度格局由水分和热量共同决定[19-20].水分和热量的作用在不同生境与尺度上并不一致[21-22],在水分充足的区域,多样性主要受温度控制,而在水分亏缺的区域,物种多样性受水分和温度的共同制约[23].此外,土壤质地与肥力等因素也被证明对植物物种多样性格局存在重要作用[24-26].这些因子对物种多样性格局的作用在前期有一定的研究支持,但对环境因子的相对重要性尚存在一定的争议[27],对不同尺度上的物种多样性格局的形成机制并未达成共识. ...
Energy,water,and broad-scale geographic patterns of species richness
1
2003
... 生态学家对物种多样性格局的形成机制提出了中域效应假说、种库假说、物种-能量假说、地史成因假说等一系列假说[14].其中以气候因素为基础的能量假说是目前最受关注的假说之一,寒冷忍耐假说认为随着冬季温度的降低物种多样性逐渐减少[15-16],生态学代谢假说试图从机制上解释环境温度与物种多样性地理格局的关系[17-18],水热动态假说认为物种多样性的大尺度格局由水分和热量共同决定[19-20].水分和热量的作用在不同生境与尺度上并不一致[21-22],在水分充足的区域,多样性主要受温度控制,而在水分亏缺的区域,物种多样性受水分和温度的共同制约[23].此外,土壤质地与肥力等因素也被证明对植物物种多样性格局存在重要作用[24-26].这些因子对物种多样性格局的作用在前期有一定的研究支持,但对环境因子的相对重要性尚存在一定的争议[27],对不同尺度上的物种多样性格局的形成机制并未达成共识. ...
Disentangling the determinants of species richness of vascular plants and mammals from national to regional scales
1
2016
... 生态学家对物种多样性格局的形成机制提出了中域效应假说、种库假说、物种-能量假说、地史成因假说等一系列假说[14].其中以气候因素为基础的能量假说是目前最受关注的假说之一,寒冷忍耐假说认为随着冬季温度的降低物种多样性逐渐减少[15-16],生态学代谢假说试图从机制上解释环境温度与物种多样性地理格局的关系[17-18],水热动态假说认为物种多样性的大尺度格局由水分和热量共同决定[19-20].水分和热量的作用在不同生境与尺度上并不一致[21-22],在水分充足的区域,多样性主要受温度控制,而在水分亏缺的区域,物种多样性受水分和温度的共同制约[23].此外,土壤质地与肥力等因素也被证明对植物物种多样性格局存在重要作用[24-26].这些因子对物种多样性格局的作用在前期有一定的研究支持,但对环境因子的相对重要性尚存在一定的争议[27],对不同尺度上的物种多样性格局的形成机制并未达成共识. ...
Relative role of contemporary environment versus history in shaping diversity patterns of China's woody plants
1
2013
... 生态学家对物种多样性格局的形成机制提出了中域效应假说、种库假说、物种-能量假说、地史成因假说等一系列假说[14].其中以气候因素为基础的能量假说是目前最受关注的假说之一,寒冷忍耐假说认为随着冬季温度的降低物种多样性逐渐减少[15-16],生态学代谢假说试图从机制上解释环境温度与物种多样性地理格局的关系[17-18],水热动态假说认为物种多样性的大尺度格局由水分和热量共同决定[19-20].水分和热量的作用在不同生境与尺度上并不一致[21-22],在水分充足的区域,多样性主要受温度控制,而在水分亏缺的区域,物种多样性受水分和温度的共同制约[23].此外,土壤质地与肥力等因素也被证明对植物物种多样性格局存在重要作用[24-26].这些因子对物种多样性格局的作用在前期有一定的研究支持,但对环境因子的相对重要性尚存在一定的争议[27],对不同尺度上的物种多样性格局的形成机制并未达成共识. ...
物种多样性地理格局的能量假说
1
2009
... 生态学家对物种多样性格局的形成机制提出了中域效应假说、种库假说、物种-能量假说、地史成因假说等一系列假说[14].其中以气候因素为基础的能量假说是目前最受关注的假说之一,寒冷忍耐假说认为随着冬季温度的降低物种多样性逐渐减少[15-16],生态学代谢假说试图从机制上解释环境温度与物种多样性地理格局的关系[17-18],水热动态假说认为物种多样性的大尺度格局由水分和热量共同决定[19-20].水分和热量的作用在不同生境与尺度上并不一致[21-22],在水分充足的区域,多样性主要受温度控制,而在水分亏缺的区域,物种多样性受水分和温度的共同制约[23].此外,土壤质地与肥力等因素也被证明对植物物种多样性格局存在重要作用[24-26].这些因子对物种多样性格局的作用在前期有一定的研究支持,但对环境因子的相对重要性尚存在一定的争议[27],对不同尺度上的物种多样性格局的形成机制并未达成共识. ...
Disentangling the influence of climate,soil and belowground microbes on local species richness in a dryland ecosystem of Northwest China
1
2017
... 生态学家对物种多样性格局的形成机制提出了中域效应假说、种库假说、物种-能量假说、地史成因假说等一系列假说[14].其中以气候因素为基础的能量假说是目前最受关注的假说之一,寒冷忍耐假说认为随着冬季温度的降低物种多样性逐渐减少[15-16],生态学代谢假说试图从机制上解释环境温度与物种多样性地理格局的关系[17-18],水热动态假说认为物种多样性的大尺度格局由水分和热量共同决定[19-20].水分和热量的作用在不同生境与尺度上并不一致[21-22],在水分充足的区域,多样性主要受温度控制,而在水分亏缺的区域,物种多样性受水分和温度的共同制约[23].此外,土壤质地与肥力等因素也被证明对植物物种多样性格局存在重要作用[24-26].这些因子对物种多样性格局的作用在前期有一定的研究支持,但对环境因子的相对重要性尚存在一定的争议[27],对不同尺度上的物种多样性格局的形成机制并未达成共识. ...
Disentangling the effects of climate,topography,soil and vegetation on stand-scale species richness in temperate forests
2015
Species richness and diversity along edaphic and climatic gradients in Amazonia
1
2014
... 生态学家对物种多样性格局的形成机制提出了中域效应假说、种库假说、物种-能量假说、地史成因假说等一系列假说[14].其中以气候因素为基础的能量假说是目前最受关注的假说之一,寒冷忍耐假说认为随着冬季温度的降低物种多样性逐渐减少[15-16],生态学代谢假说试图从机制上解释环境温度与物种多样性地理格局的关系[17-18],水热动态假说认为物种多样性的大尺度格局由水分和热量共同决定[19-20].水分和热量的作用在不同生境与尺度上并不一致[21-22],在水分充足的区域,多样性主要受温度控制,而在水分亏缺的区域,物种多样性受水分和温度的共同制约[23].此外,土壤质地与肥力等因素也被证明对植物物种多样性格局存在重要作用[24-26].这些因子对物种多样性格局的作用在前期有一定的研究支持,但对环境因子的相对重要性尚存在一定的争议[27],对不同尺度上的物种多样性格局的形成机制并未达成共识. ...
中国裸子植物的物种多样性格局及其影响因子
1
2018
... 生态学家对物种多样性格局的形成机制提出了中域效应假说、种库假说、物种-能量假说、地史成因假说等一系列假说[14].其中以气候因素为基础的能量假说是目前最受关注的假说之一,寒冷忍耐假说认为随着冬季温度的降低物种多样性逐渐减少[15-16],生态学代谢假说试图从机制上解释环境温度与物种多样性地理格局的关系[17-18],水热动态假说认为物种多样性的大尺度格局由水分和热量共同决定[19-20].水分和热量的作用在不同生境与尺度上并不一致[21-22],在水分充足的区域,多样性主要受温度控制,而在水分亏缺的区域,物种多样性受水分和温度的共同制约[23].此外,土壤质地与肥力等因素也被证明对植物物种多样性格局存在重要作用[24-26].这些因子对物种多样性格局的作用在前期有一定的研究支持,但对环境因子的相对重要性尚存在一定的争议[27],对不同尺度上的物种多样性格局的形成机制并未达成共识. ...
浑善达克沙地植物群落主要类型与特征
2
2021
... 浑善达克沙地位于内蒙古高原中东部,跨越中国半湿润-半干旱区-干旱区,因水分条件的不同,浑善达克沙地植被从东到西具有明显的地带性特征,由东部的疏林草原、中部的典型草原向西部的荒漠草原过渡[28].近年来研究人员已从土壤、微地形、放牧等方面对影响浑善达克物种多样性的环境因素进行了一系列研究.宋创业等[29]发现浑善达克沙地固定沙丘的物种丰富度指数和多样性指数最高,均匀度指数与土壤pH值以及全氮含量显著相关,丰富度指数与有机质含量显著相关;张志永等[30]的研究表明微地形能够显著影响浑善达克沙地的群落分布和物种多样性特征;詹瑾等[31]的研究显示浑善达克沙地不同放牧程度下土壤有机碳和全氮含量存在极显著差异,而丰富度、多样性和均匀度指数差异均不显著.这些研究多在单一群落类型或浑善达克沙地的局部区域,尚未发现从生物多样性角度分析浑善达克沙地的环境对植物群落影响的理论和成果,这不利于对浑善达克沙地生物多样性格局及形成机制的认识.研究浑善达克沙地植物群落物种多样性及其与环境的关系,是深入研究沙地生态系统功能变化规律并科学解读沙地植被恢复机理的有效途径,对于促使植被恢复具有重要参考价值.本研究在野外群落调查的基础上,通过数量生态学研究方法对群落的物种多样性、空间分布格局及其影响因子进行了分析,尝试阐明该区域的主要植物群落、物种多样性特征及其环境解释.研究结果可为浑善达克沙地生物多样性保护、植被演替和多样性维持机制等生态问题的进一步研究提供数据支撑和理论依据. ...
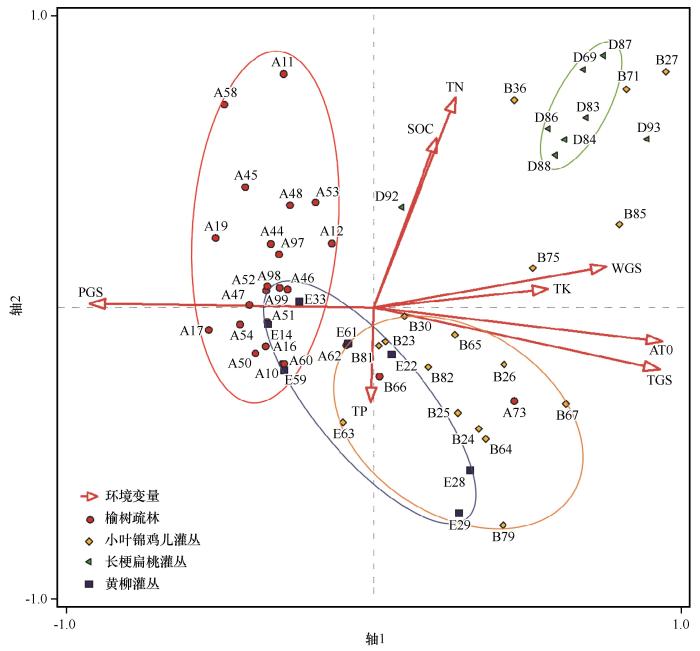
... 气候是植被大尺度分布格局的决定性因子[47],植物种群和群落的分布都是由气候(主要是水热条件)决定的.降水量被认为是影响干旱区、半干旱区荒漠植被和沙地植被的最主要因子,有研究表明,地下水位及土壤阳离子对于毛乌素沙地丘间地植物群落分布产生重要影响[48-51];浑善达克沙地镶嵌于典型草原地带之中[52],年降水量从东部的440 mm向西逐渐减少至150 mm[28].随着降水量的递减,自东向西依次分布着疏林、灌丛、荒漠等植被类型,在空间上呈现出斑块镶嵌的景观[53].其中,疏林和灌丛是最常见的两种,以榆树疏林和小叶锦鸡儿灌丛为典型代表.榆树疏林分布在沙地中东部,物种多样性较高,这与之前对榆树疏林植物多样性的结果相近[54-55].研究区的小叶锦鸡儿灌丛分布在沙地的中西部,水分条件稍差,因而多样性水平较低,这是草原灌丛化对群落多样性和生态系统功能的影响[56].研究表明,物种多样性的大尺度格局由水分和温度共同决定[57-58].在水分条件充足时,温度是影响物种丰富度的主要因子,而在降水不足时则受降水量控制[59-60].降水量是影响浑善达克沙地植被的主要气候因素[61];本研究证明,植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数与生长季降水量呈显著的正相关关系,这与前期的研究是一致的,水分条件是影响物种多样性的最主要因子[62-64]. ...
浑善达克沙地植物群落物种多样性与土壤因子的关系
1
2008
... 浑善达克沙地位于内蒙古高原中东部,跨越中国半湿润-半干旱区-干旱区,因水分条件的不同,浑善达克沙地植被从东到西具有明显的地带性特征,由东部的疏林草原、中部的典型草原向西部的荒漠草原过渡[28].近年来研究人员已从土壤、微地形、放牧等方面对影响浑善达克物种多样性的环境因素进行了一系列研究.宋创业等[29]发现浑善达克沙地固定沙丘的物种丰富度指数和多样性指数最高,均匀度指数与土壤pH值以及全氮含量显著相关,丰富度指数与有机质含量显著相关;张志永等[30]的研究表明微地形能够显著影响浑善达克沙地的群落分布和物种多样性特征;詹瑾等[31]的研究显示浑善达克沙地不同放牧程度下土壤有机碳和全氮含量存在极显著差异,而丰富度、多样性和均匀度指数差异均不显著.这些研究多在单一群落类型或浑善达克沙地的局部区域,尚未发现从生物多样性角度分析浑善达克沙地的环境对植物群落影响的理论和成果,这不利于对浑善达克沙地生物多样性格局及形成机制的认识.研究浑善达克沙地植物群落物种多样性及其与环境的关系,是深入研究沙地生态系统功能变化规律并科学解读沙地植被恢复机理的有效途径,对于促使植被恢复具有重要参考价值.本研究在野外群落调查的基础上,通过数量生态学研究方法对群落的物种多样性、空间分布格局及其影响因子进行了分析,尝试阐明该区域的主要植物群落、物种多样性特征及其环境解释.研究结果可为浑善达克沙地生物多样性保护、植被演替和多样性维持机制等生态问题的进一步研究提供数据支撑和理论依据. ...
浑善达克沙地不同微地形的土壤物理性质和草本群落分布及其相关性分析
1
2017
... 浑善达克沙地位于内蒙古高原中东部,跨越中国半湿润-半干旱区-干旱区,因水分条件的不同,浑善达克沙地植被从东到西具有明显的地带性特征,由东部的疏林草原、中部的典型草原向西部的荒漠草原过渡[28].近年来研究人员已从土壤、微地形、放牧等方面对影响浑善达克物种多样性的环境因素进行了一系列研究.宋创业等[29]发现浑善达克沙地固定沙丘的物种丰富度指数和多样性指数最高,均匀度指数与土壤pH值以及全氮含量显著相关,丰富度指数与有机质含量显著相关;张志永等[30]的研究表明微地形能够显著影响浑善达克沙地的群落分布和物种多样性特征;詹瑾等[31]的研究显示浑善达克沙地不同放牧程度下土壤有机碳和全氮含量存在极显著差异,而丰富度、多样性和均匀度指数差异均不显著.这些研究多在单一群落类型或浑善达克沙地的局部区域,尚未发现从生物多样性角度分析浑善达克沙地的环境对植物群落影响的理论和成果,这不利于对浑善达克沙地生物多样性格局及形成机制的认识.研究浑善达克沙地植物群落物种多样性及其与环境的关系,是深入研究沙地生态系统功能变化规律并科学解读沙地植被恢复机理的有效途径,对于促使植被恢复具有重要参考价值.本研究在野外群落调查的基础上,通过数量生态学研究方法对群落的物种多样性、空间分布格局及其影响因子进行了分析,尝试阐明该区域的主要植物群落、物种多样性特征及其环境解释.研究结果可为浑善达克沙地生物多样性保护、植被演替和多样性维持机制等生态问题的进一步研究提供数据支撑和理论依据. ...
放牧对浑善达克沙地丘间低地植被群落及土壤的影响
1
2019
... 浑善达克沙地位于内蒙古高原中东部,跨越中国半湿润-半干旱区-干旱区,因水分条件的不同,浑善达克沙地植被从东到西具有明显的地带性特征,由东部的疏林草原、中部的典型草原向西部的荒漠草原过渡[28].近年来研究人员已从土壤、微地形、放牧等方面对影响浑善达克物种多样性的环境因素进行了一系列研究.宋创业等[29]发现浑善达克沙地固定沙丘的物种丰富度指数和多样性指数最高,均匀度指数与土壤pH值以及全氮含量显著相关,丰富度指数与有机质含量显著相关;张志永等[30]的研究表明微地形能够显著影响浑善达克沙地的群落分布和物种多样性特征;詹瑾等[31]的研究显示浑善达克沙地不同放牧程度下土壤有机碳和全氮含量存在极显著差异,而丰富度、多样性和均匀度指数差异均不显著.这些研究多在单一群落类型或浑善达克沙地的局部区域,尚未发现从生物多样性角度分析浑善达克沙地的环境对植物群落影响的理论和成果,这不利于对浑善达克沙地生物多样性格局及形成机制的认识.研究浑善达克沙地植物群落物种多样性及其与环境的关系,是深入研究沙地生态系统功能变化规律并科学解读沙地植被恢复机理的有效途径,对于促使植被恢复具有重要参考价值.本研究在野外群落调查的基础上,通过数量生态学研究方法对群落的物种多样性、空间分布格局及其影响因子进行了分析,尝试阐明该区域的主要植物群落、物种多样性特征及其环境解释.研究结果可为浑善达克沙地生物多样性保护、植被演替和多样性维持机制等生态问题的进一步研究提供数据支撑和理论依据. ...
浑善达克沙地沙漠化过程及其植被恢复的基础研究:以正蓝旗为例
1
2003
... 浑善达克沙地位于内蒙古高原中东部,东起大兴安岭南段西麓达里诺尔,向西一直延伸到集二线铁路沿线,南部为阴山山地隆起区[32-33].本研究区域包括浑善达克沙地及其周边小面积沙地,地理坐标为41°56′—44°26′N、112°22′—117°57′E,海拔1 000—1 400 m.东部以固定半固定沙地为主,沙丘高度10—15 m,西部以半固定、流动沙地为主[34].多年平均气温2—5 ℃,年降水量东部为350—400 mm、西部为100—200 mm,降水集中在生长季(6—8月),其中7月降雨最多;年蒸发量为2 000—2 900 mm,属于温带大陆性半干旱气候区[35].土壤以风沙土和栗钙土为主,其次为棕钙土.植被具有明显的地带性,从东到西依次分布着疏林草地、灌丛和荒漠草原,建群种主要有榆树(Ulmus pumila)、小叶锦鸡儿(Caragana microphylla)、盐蒿(Artemisia halodendron)等. ...
浑善达克沙地榆树疏林中木本植物空间格局及种内和种间关系分析
1
2019
... 浑善达克沙地位于内蒙古高原中东部,东起大兴安岭南段西麓达里诺尔,向西一直延伸到集二线铁路沿线,南部为阴山山地隆起区[32-33].本研究区域包括浑善达克沙地及其周边小面积沙地,地理坐标为41°56′—44°26′N、112°22′—117°57′E,海拔1 000—1 400 m.东部以固定半固定沙地为主,沙丘高度10—15 m,西部以半固定、流动沙地为主[34].多年平均气温2—5 ℃,年降水量东部为350—400 mm、西部为100—200 mm,降水集中在生长季(6—8月),其中7月降雨最多;年蒸发量为2 000—2 900 mm,属于温带大陆性半干旱气候区[35].土壤以风沙土和栗钙土为主,其次为棕钙土.植被具有明显的地带性,从东到西依次分布着疏林草地、灌丛和荒漠草原,建群种主要有榆树(Ulmus pumila)、小叶锦鸡儿(Caragana microphylla)、盐蒿(Artemisia halodendron)等. ...
浑善达克沙地桑根达来地区榆树疏林的分布与立地因子的关系的研究
1
2008
... 浑善达克沙地位于内蒙古高原中东部,东起大兴安岭南段西麓达里诺尔,向西一直延伸到集二线铁路沿线,南部为阴山山地隆起区[32-33].本研究区域包括浑善达克沙地及其周边小面积沙地,地理坐标为41°56′—44°26′N、112°22′—117°57′E,海拔1 000—1 400 m.东部以固定半固定沙地为主,沙丘高度10—15 m,西部以半固定、流动沙地为主[34].多年平均气温2—5 ℃,年降水量东部为350—400 mm、西部为100—200 mm,降水集中在生长季(6—8月),其中7月降雨最多;年蒸发量为2 000—2 900 mm,属于温带大陆性半干旱气候区[35].土壤以风沙土和栗钙土为主,其次为棕钙土.植被具有明显的地带性,从东到西依次分布着疏林草地、灌丛和荒漠草原,建群种主要有榆树(Ulmus pumila)、小叶锦鸡儿(Caragana microphylla)、盐蒿(Artemisia halodendron)等. ...
浑善达克沙地沙漠化成因、过程及其防治对策研究:以内蒙古正蓝旗为例
1
2004
... 浑善达克沙地位于内蒙古高原中东部,东起大兴安岭南段西麓达里诺尔,向西一直延伸到集二线铁路沿线,南部为阴山山地隆起区[32-33].本研究区域包括浑善达克沙地及其周边小面积沙地,地理坐标为41°56′—44°26′N、112°22′—117°57′E,海拔1 000—1 400 m.东部以固定半固定沙地为主,沙丘高度10—15 m,西部以半固定、流动沙地为主[34].多年平均气温2—5 ℃,年降水量东部为350—400 mm、西部为100—200 mm,降水集中在生长季(6—8月),其中7月降雨最多;年蒸发量为2 000—2 900 mm,属于温带大陆性半干旱气候区[35].土壤以风沙土和栗钙土为主,其次为棕钙土.植被具有明显的地带性,从东到西依次分布着疏林草地、灌丛和荒漠草原,建群种主要有榆树(Ulmus pumila)、小叶锦鸡儿(Caragana microphylla)、盐蒿(Artemisia halodendron)等. ...
1∶100万中国植被图森林和灌丛群系类型的补充资料
1
2019
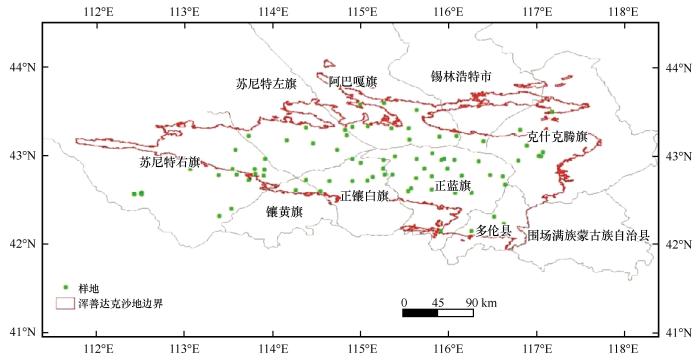
... 采用科技部科技基础资源调查专项“中国荒漠主要植物群落调查”方法,每隔10 km布设一个样地,共83个样地(图1).用GPS对每一个样地精确定位,记录地理位置、海拔、坡向、坡度、土壤类型、群落特征、干扰情况等信息.参照《中国植被》和《中国植被及其地理格局——中华人民共和国植被图集1∶100万说明书》及补充材料[36]中的植物生活型分类系统,将树高≥2 m且有明显主干的木本植物界定为乔木,没有明显主干或树高<2 m的木本植物界定为灌木.参考方精云等[37]提出的植物群落调查技术规范,设置1个100 m×100 m的大样方,对其中的乔木采用逐木调查的方法,记录高度、胸径和冠幅;在大样方中布设5个10 m×10 m的灌木样方,逐一测定灌木的高度和冠幅;在大样方中布设9个1 m×1 m的草本样方,记录样方中每种植物的种类、个体数和盖度,并选择3个标准株测量其高度以估算平均高度.共调查了83个样地,包括83个100 m×100 m的乔木样方、415个10 m×10 m的灌木样方和747个1 m×1 m的草本样方.野外调查时间为2018—2020年7—8月. ...
植物群落清查的主要内容、方法和技术规范
1
2009
... 采用科技部科技基础资源调查专项“中国荒漠主要植物群落调查”方法,每隔10 km布设一个样地,共83个样地(图1).用GPS对每一个样地精确定位,记录地理位置、海拔、坡向、坡度、土壤类型、群落特征、干扰情况等信息.参照《中国植被》和《中国植被及其地理格局——中华人民共和国植被图集1∶100万说明书》及补充材料[36]中的植物生活型分类系统,将树高≥2 m且有明显主干的木本植物界定为乔木,没有明显主干或树高<2 m的木本植物界定为灌木.参考方精云等[37]提出的植物群落调查技术规范,设置1个100 m×100 m的大样方,对其中的乔木采用逐木调查的方法,记录高度、胸径和冠幅;在大样方中布设5个10 m×10 m的灌木样方,逐一测定灌木的高度和冠幅;在大样方中布设9个1 m×1 m的草本样方,记录样方中每种植物的种类、个体数和盖度,并选择3个标准株测量其高度以估算平均高度.共调查了83个样地,包括83个100 m×100 m的乔木样方、415个10 m×10 m的灌木样方和747个1 m×1 m的草本样方.野外调查时间为2018—2020年7—8月. ...
中国区域地面气象要素驱动数据集(1979-2018)
1
2019
... 利用“中国区域地面气象要素驱动数据集(CMFD)”[38]的日尺度降水、温度和风速数据,计算每个像元的月均温度、降水和风速数据,提取每个像元1979—2018年的平均值,计算得到生长季降水量、生长季均温和生长季平均风速.≥0 ℃积温(经TEM校正)数据获取自《中国气象背景数据集》[39].环境因子见表1. ...
中国气象背景数据集
1
2017
... 利用“中国区域地面气象要素驱动数据集(CMFD)”[38]的日尺度降水、温度和风速数据,计算每个像元的月均温度、降水和风速数据,提取每个像元1979—2018年的平均值,计算得到生长季降水量、生长季均温和生长季平均风速.≥0 ℃积温(经TEM校正)数据获取自《中国气象背景数据集》[39].环境因子见表1. ...
3
2011
... 采用Patrick指数、Simpson指数、Shannon-Wiener指数和Pielou指数测度浑善达克沙地不同植物群落的物种多样性程度[40-41].根据样方中每个物种的高度、盖度和频度计算出重要值,作为计算多样性指数的基础[7]. ...
... 物种多样性具有丰富性和均匀性两方面内容[40],Patrick指数是最常用的丰富度指数,本文研究表明,浑善达克沙地的物种丰富度程度最高的区域在43°N、116°E、海拔1 300 m附近,是正蓝旗榆树疏林分布的核心区[45].研究区Pielou指数整体较高,较高的均匀度有利于维持群落结构的稳定性[46].在均匀度差异不显著的情况下,Shannon-Wiener指数对群落的丰富度较为敏感[40].本研究显示,物种多样性指数的变化趋势与物种丰富度基本一致,表现为物种数越多,多样性指数越高,这与在内蒙古西部的研究基本一致[7]. ...
... [40].本研究显示,物种多样性指数的变化趋势与物种丰富度基本一致,表现为物种数越多,多样性指数越高,这与在内蒙古西部的研究基本一致[7]. ...
北京东灵山地区植物群落多样性的研究Ⅱ丰富度、均匀度和物种多样性指数
1
1995
... 采用Patrick指数、Simpson指数、Shannon-Wiener指数和Pielou指数测度浑善达克沙地不同植物群落的物种多样性程度[40-41].根据样方中每个物种的高度、盖度和频度计算出重要值,作为计算多样性指数的基础[7]. ...
浑善达克沙地榆分布与地面因素关系的研究
1
2009
... 浑善达克沙地的植物多样性具有一定的特殊性.研究区属于温带大陆性气候区,寒冷、少雨、风沙大,广泛分布的土壤为风沙土,常与盐碱土、草甸土相间分布,形成坨甸相间的格局[42].在这样的环境条件下发育的植物群落以旱生的沙生植物最为常见,以多年生草本植物和灌木、小灌木、半灌木形成的灌丛和草地植被为主,在东部水分条件较高的区域形成榆树疏林草原.植物物种向优势科集中,表现出多数种属于少数科的特点.与毛乌素沙地[43]和科尔沁沙地[44]做对比,毛乌素沙地以菊科、蔷薇科、豆科、禾本科为主,苋科比例明显降低;科尔沁沙地以菊科、豆科、禾本科为主,蔷薇科比例较低;浑善达克沙地以菊科、禾本科、豆科、苋科、蔷薇科、唇形科为主,占到总物种数的半数以上.优势科的组成和数量体现着一个区域的区系特点,这些科表现出沙地植被的特殊性和半干旱气候影响下的植被特征. ...
毛乌素沙地天然植物多样性组成及区系特征分析
1
2018
... 浑善达克沙地的植物多样性具有一定的特殊性.研究区属于温带大陆性气候区,寒冷、少雨、风沙大,广泛分布的土壤为风沙土,常与盐碱土、草甸土相间分布,形成坨甸相间的格局[42].在这样的环境条件下发育的植物群落以旱生的沙生植物最为常见,以多年生草本植物和灌木、小灌木、半灌木形成的灌丛和草地植被为主,在东部水分条件较高的区域形成榆树疏林草原.植物物种向优势科集中,表现出多数种属于少数科的特点.与毛乌素沙地[43]和科尔沁沙地[44]做对比,毛乌素沙地以菊科、蔷薇科、豆科、禾本科为主,苋科比例明显降低;科尔沁沙地以菊科、豆科、禾本科为主,蔷薇科比例较低;浑善达克沙地以菊科、禾本科、豆科、苋科、蔷薇科、唇形科为主,占到总物种数的半数以上.优势科的组成和数量体现着一个区域的区系特点,这些科表现出沙地植被的特殊性和半干旱气候影响下的植被特征. ...
科尔沁沙地植物物种丰富度格局及其影响因素
1
2019
... 浑善达克沙地的植物多样性具有一定的特殊性.研究区属于温带大陆性气候区,寒冷、少雨、风沙大,广泛分布的土壤为风沙土,常与盐碱土、草甸土相间分布,形成坨甸相间的格局[42].在这样的环境条件下发育的植物群落以旱生的沙生植物最为常见,以多年生草本植物和灌木、小灌木、半灌木形成的灌丛和草地植被为主,在东部水分条件较高的区域形成榆树疏林草原.植物物种向优势科集中,表现出多数种属于少数科的特点.与毛乌素沙地[43]和科尔沁沙地[44]做对比,毛乌素沙地以菊科、蔷薇科、豆科、禾本科为主,苋科比例明显降低;科尔沁沙地以菊科、豆科、禾本科为主,蔷薇科比例较低;浑善达克沙地以菊科、禾本科、豆科、苋科、蔷薇科、唇形科为主,占到总物种数的半数以上.优势科的组成和数量体现着一个区域的区系特点,这些科表现出沙地植被的特殊性和半干旱气候影响下的植被特征. ...
浑善达克沙地榆树疏林自然保护区核心区设计的初步研究
1
2005
... 物种多样性具有丰富性和均匀性两方面内容[40],Patrick指数是最常用的丰富度指数,本文研究表明,浑善达克沙地的物种丰富度程度最高的区域在43°N、116°E、海拔1 300 m附近,是正蓝旗榆树疏林分布的核心区[45].研究区Pielou指数整体较高,较高的均匀度有利于维持群落结构的稳定性[46].在均匀度差异不显著的情况下,Shannon-Wiener指数对群落的丰富度较为敏感[40].本研究显示,物种多样性指数的变化趋势与物种丰富度基本一致,表现为物种数越多,多样性指数越高,这与在内蒙古西部的研究基本一致[7]. ...
Consequences of dominance:a review of evenness effects on local and regional ecosystem processes
1
2008
... 物种多样性具有丰富性和均匀性两方面内容[40],Patrick指数是最常用的丰富度指数,本文研究表明,浑善达克沙地的物种丰富度程度最高的区域在43°N、116°E、海拔1 300 m附近,是正蓝旗榆树疏林分布的核心区[45].研究区Pielou指数整体较高,较高的均匀度有利于维持群落结构的稳定性[46].在均匀度差异不显著的情况下,Shannon-Wiener指数对群落的丰富度较为敏感[40].本研究显示,物种多样性指数的变化趋势与物种丰富度基本一致,表现为物种数越多,多样性指数越高,这与在内蒙古西部的研究基本一致[7]. ...
Global patterns in biodiversity
1
2000
... 气候是植被大尺度分布格局的决定性因子[47],植物种群和群落的分布都是由气候(主要是水热条件)决定的.降水量被认为是影响干旱区、半干旱区荒漠植被和沙地植被的最主要因子,有研究表明,地下水位及土壤阳离子对于毛乌素沙地丘间地植物群落分布产生重要影响[48-51];浑善达克沙地镶嵌于典型草原地带之中[52],年降水量从东部的440 mm向西逐渐减少至150 mm[28].随着降水量的递减,自东向西依次分布着疏林、灌丛、荒漠等植被类型,在空间上呈现出斑块镶嵌的景观[53].其中,疏林和灌丛是最常见的两种,以榆树疏林和小叶锦鸡儿灌丛为典型代表.榆树疏林分布在沙地中东部,物种多样性较高,这与之前对榆树疏林植物多样性的结果相近[54-55].研究区的小叶锦鸡儿灌丛分布在沙地的中西部,水分条件稍差,因而多样性水平较低,这是草原灌丛化对群落多样性和生态系统功能的影响[56].研究表明,物种多样性的大尺度格局由水分和温度共同决定[57-58].在水分条件充足时,温度是影响物种丰富度的主要因子,而在降水不足时则受降水量控制[59-60].降水量是影响浑善达克沙地植被的主要气候因素[61];本研究证明,植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数与生长季降水量呈显著的正相关关系,这与前期的研究是一致的,水分条件是影响物种多样性的最主要因子[62-64]. ...
毛乌素沙地丘间低地植物群落分类与排序
1
2016
... 气候是植被大尺度分布格局的决定性因子[47],植物种群和群落的分布都是由气候(主要是水热条件)决定的.降水量被认为是影响干旱区、半干旱区荒漠植被和沙地植被的最主要因子,有研究表明,地下水位及土壤阳离子对于毛乌素沙地丘间地植物群落分布产生重要影响[48-51];浑善达克沙地镶嵌于典型草原地带之中[52],年降水量从东部的440 mm向西逐渐减少至150 mm[28].随着降水量的递减,自东向西依次分布着疏林、灌丛、荒漠等植被类型,在空间上呈现出斑块镶嵌的景观[53].其中,疏林和灌丛是最常见的两种,以榆树疏林和小叶锦鸡儿灌丛为典型代表.榆树疏林分布在沙地中东部,物种多样性较高,这与之前对榆树疏林植物多样性的结果相近[54-55].研究区的小叶锦鸡儿灌丛分布在沙地的中西部,水分条件稍差,因而多样性水平较低,这是草原灌丛化对群落多样性和生态系统功能的影响[56].研究表明,物种多样性的大尺度格局由水分和温度共同决定[57-58].在水分条件充足时,温度是影响物种丰富度的主要因子,而在降水不足时则受降水量控制[59-60].降水量是影响浑善达克沙地植被的主要气候因素[61];本研究证明,植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数与生长季降水量呈显著的正相关关系,这与前期的研究是一致的,水分条件是影响物种多样性的最主要因子[62-64]. ...
2000-2012年科尔沁沙地植被与气候因子间的响应关系
2016
科尔沁沙地固定沙丘植被物种多样性对降水变化的响应
1
2000
... 气候是植被大尺度分布格局的决定性因子[47],植物种群和群落的分布都是由气候(主要是水热条件)决定的.降水量被认为是影响干旱区、半干旱区荒漠植被和沙地植被的最主要因子,有研究表明,地下水位及土壤阳离子对于毛乌素沙地丘间地植物群落分布产生重要影响[48-51];浑善达克沙地镶嵌于典型草原地带之中[52],年降水量从东部的440 mm向西逐渐减少至150 mm[28].随着降水量的递减,自东向西依次分布着疏林、灌丛、荒漠等植被类型,在空间上呈现出斑块镶嵌的景观[53].其中,疏林和灌丛是最常见的两种,以榆树疏林和小叶锦鸡儿灌丛为典型代表.榆树疏林分布在沙地中东部,物种多样性较高,这与之前对榆树疏林植物多样性的结果相近[54-55].研究区的小叶锦鸡儿灌丛分布在沙地的中西部,水分条件稍差,因而多样性水平较低,这是草原灌丛化对群落多样性和生态系统功能的影响[56].研究表明,物种多样性的大尺度格局由水分和温度共同决定[57-58].在水分条件充足时,温度是影响物种丰富度的主要因子,而在降水不足时则受降水量控制[59-60].降水量是影响浑善达克沙地植被的主要气候因素[61];本研究证明,植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数与生长季降水量呈显著的正相关关系,这与前期的研究是一致的,水分条件是影响物种多样性的最主要因子[62-64]. ...
2000-2014年浑善达克沙地植被覆盖变化研究
1
2016
... 气候是植被大尺度分布格局的决定性因子[47],植物种群和群落的分布都是由气候(主要是水热条件)决定的.降水量被认为是影响干旱区、半干旱区荒漠植被和沙地植被的最主要因子,有研究表明,地下水位及土壤阳离子对于毛乌素沙地丘间地植物群落分布产生重要影响[48-51];浑善达克沙地镶嵌于典型草原地带之中[52],年降水量从东部的440 mm向西逐渐减少至150 mm[28].随着降水量的递减,自东向西依次分布着疏林、灌丛、荒漠等植被类型,在空间上呈现出斑块镶嵌的景观[53].其中,疏林和灌丛是最常见的两种,以榆树疏林和小叶锦鸡儿灌丛为典型代表.榆树疏林分布在沙地中东部,物种多样性较高,这与之前对榆树疏林植物多样性的结果相近[54-55].研究区的小叶锦鸡儿灌丛分布在沙地的中西部,水分条件稍差,因而多样性水平较低,这是草原灌丛化对群落多样性和生态系统功能的影响[56].研究表明,物种多样性的大尺度格局由水分和温度共同决定[57-58].在水分条件充足时,温度是影响物种丰富度的主要因子,而在降水不足时则受降水量控制[59-60].降水量是影响浑善达克沙地植被的主要气候因素[61];本研究证明,植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数与生长季降水量呈显著的正相关关系,这与前期的研究是一致的,水分条件是影响物种多样性的最主要因子[62-64]. ...
放牧方式对浑善达克沙地榆树疏林退化的影响
1
2020
... 气候是植被大尺度分布格局的决定性因子[47],植物种群和群落的分布都是由气候(主要是水热条件)决定的.降水量被认为是影响干旱区、半干旱区荒漠植被和沙地植被的最主要因子,有研究表明,地下水位及土壤阳离子对于毛乌素沙地丘间地植物群落分布产生重要影响[48-51];浑善达克沙地镶嵌于典型草原地带之中[52],年降水量从东部的440 mm向西逐渐减少至150 mm[28].随着降水量的递减,自东向西依次分布着疏林、灌丛、荒漠等植被类型,在空间上呈现出斑块镶嵌的景观[53].其中,疏林和灌丛是最常见的两种,以榆树疏林和小叶锦鸡儿灌丛为典型代表.榆树疏林分布在沙地中东部,物种多样性较高,这与之前对榆树疏林植物多样性的结果相近[54-55].研究区的小叶锦鸡儿灌丛分布在沙地的中西部,水分条件稍差,因而多样性水平较低,这是草原灌丛化对群落多样性和生态系统功能的影响[56].研究表明,物种多样性的大尺度格局由水分和温度共同决定[57-58].在水分条件充足时,温度是影响物种丰富度的主要因子,而在降水不足时则受降水量控制[59-60].降水量是影响浑善达克沙地植被的主要气候因素[61];本研究证明,植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数与生长季降水量呈显著的正相关关系,这与前期的研究是一致的,水分条件是影响物种多样性的最主要因子[62-64]. ...
人类活动对榆树疏林土壤环境和植物多样性的影响
1
2003
... 气候是植被大尺度分布格局的决定性因子[47],植物种群和群落的分布都是由气候(主要是水热条件)决定的.降水量被认为是影响干旱区、半干旱区荒漠植被和沙地植被的最主要因子,有研究表明,地下水位及土壤阳离子对于毛乌素沙地丘间地植物群落分布产生重要影响[48-51];浑善达克沙地镶嵌于典型草原地带之中[52],年降水量从东部的440 mm向西逐渐减少至150 mm[28].随着降水量的递减,自东向西依次分布着疏林、灌丛、荒漠等植被类型,在空间上呈现出斑块镶嵌的景观[53].其中,疏林和灌丛是最常见的两种,以榆树疏林和小叶锦鸡儿灌丛为典型代表.榆树疏林分布在沙地中东部,物种多样性较高,这与之前对榆树疏林植物多样性的结果相近[54-55].研究区的小叶锦鸡儿灌丛分布在沙地的中西部,水分条件稍差,因而多样性水平较低,这是草原灌丛化对群落多样性和生态系统功能的影响[56].研究表明,物种多样性的大尺度格局由水分和温度共同决定[57-58].在水分条件充足时,温度是影响物种丰富度的主要因子,而在降水不足时则受降水量控制[59-60].降水量是影响浑善达克沙地植被的主要气候因素[61];本研究证明,植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数与生长季降水量呈显著的正相关关系,这与前期的研究是一致的,水分条件是影响物种多样性的最主要因子[62-64]. ...
科尔沁沙地榆树疏林草地物种多样性及乔木种群空间格局
1
2005
... 气候是植被大尺度分布格局的决定性因子[47],植物种群和群落的分布都是由气候(主要是水热条件)决定的.降水量被认为是影响干旱区、半干旱区荒漠植被和沙地植被的最主要因子,有研究表明,地下水位及土壤阳离子对于毛乌素沙地丘间地植物群落分布产生重要影响[48-51];浑善达克沙地镶嵌于典型草原地带之中[52],年降水量从东部的440 mm向西逐渐减少至150 mm[28].随着降水量的递减,自东向西依次分布着疏林、灌丛、荒漠等植被类型,在空间上呈现出斑块镶嵌的景观[53].其中,疏林和灌丛是最常见的两种,以榆树疏林和小叶锦鸡儿灌丛为典型代表.榆树疏林分布在沙地中东部,物种多样性较高,这与之前对榆树疏林植物多样性的结果相近[54-55].研究区的小叶锦鸡儿灌丛分布在沙地的中西部,水分条件稍差,因而多样性水平较低,这是草原灌丛化对群落多样性和生态系统功能的影响[56].研究表明,物种多样性的大尺度格局由水分和温度共同决定[57-58].在水分条件充足时,温度是影响物种丰富度的主要因子,而在降水不足时则受降水量控制[59-60].降水量是影响浑善达克沙地植被的主要气候因素[61];本研究证明,植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数与生长季降水量呈显著的正相关关系,这与前期的研究是一致的,水分条件是影响物种多样性的最主要因子[62-64]. ...
小叶锦鸡儿灌丛化对典型草原群落结构与生态系统功能的影响
1
2020
... 气候是植被大尺度分布格局的决定性因子[47],植物种群和群落的分布都是由气候(主要是水热条件)决定的.降水量被认为是影响干旱区、半干旱区荒漠植被和沙地植被的最主要因子,有研究表明,地下水位及土壤阳离子对于毛乌素沙地丘间地植物群落分布产生重要影响[48-51];浑善达克沙地镶嵌于典型草原地带之中[52],年降水量从东部的440 mm向西逐渐减少至150 mm[28].随着降水量的递减,自东向西依次分布着疏林、灌丛、荒漠等植被类型,在空间上呈现出斑块镶嵌的景观[53].其中,疏林和灌丛是最常见的两种,以榆树疏林和小叶锦鸡儿灌丛为典型代表.榆树疏林分布在沙地中东部,物种多样性较高,这与之前对榆树疏林植物多样性的结果相近[54-55].研究区的小叶锦鸡儿灌丛分布在沙地的中西部,水分条件稍差,因而多样性水平较低,这是草原灌丛化对群落多样性和生态系统功能的影响[56].研究表明,物种多样性的大尺度格局由水分和温度共同决定[57-58].在水分条件充足时,温度是影响物种丰富度的主要因子,而在降水不足时则受降水量控制[59-60].降水量是影响浑善达克沙地植被的主要气候因素[61];本研究证明,植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数与生长季降水量呈显著的正相关关系,这与前期的研究是一致的,水分条件是影响物种多样性的最主要因子[62-64]. ...
Two decades of homage to Santa Rosalia:toward a general theory of diversity
1
1981
... 气候是植被大尺度分布格局的决定性因子[47],植物种群和群落的分布都是由气候(主要是水热条件)决定的.降水量被认为是影响干旱区、半干旱区荒漠植被和沙地植被的最主要因子,有研究表明,地下水位及土壤阳离子对于毛乌素沙地丘间地植物群落分布产生重要影响[48-51];浑善达克沙地镶嵌于典型草原地带之中[52],年降水量从东部的440 mm向西逐渐减少至150 mm[28].随着降水量的递减,自东向西依次分布着疏林、灌丛、荒漠等植被类型,在空间上呈现出斑块镶嵌的景观[53].其中,疏林和灌丛是最常见的两种,以榆树疏林和小叶锦鸡儿灌丛为典型代表.榆树疏林分布在沙地中东部,物种多样性较高,这与之前对榆树疏林植物多样性的结果相近[54-55].研究区的小叶锦鸡儿灌丛分布在沙地的中西部,水分条件稍差,因而多样性水平较低,这是草原灌丛化对群落多样性和生态系统功能的影响[56].研究表明,物种多样性的大尺度格局由水分和温度共同决定[57-58].在水分条件充足时,温度是影响物种丰富度的主要因子,而在降水不足时则受降水量控制[59-60].降水量是影响浑善达克沙地植被的主要气候因素[61];本研究证明,植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数与生长季降水量呈显著的正相关关系,这与前期的研究是一致的,水分条件是影响物种多样性的最主要因子[62-64]. ...
Species-energy theory:an extension of species-area theory
1
1983
... 气候是植被大尺度分布格局的决定性因子[47],植物种群和群落的分布都是由气候(主要是水热条件)决定的.降水量被认为是影响干旱区、半干旱区荒漠植被和沙地植被的最主要因子,有研究表明,地下水位及土壤阳离子对于毛乌素沙地丘间地植物群落分布产生重要影响[48-51];浑善达克沙地镶嵌于典型草原地带之中[52],年降水量从东部的440 mm向西逐渐减少至150 mm[28].随着降水量的递减,自东向西依次分布着疏林、灌丛、荒漠等植被类型,在空间上呈现出斑块镶嵌的景观[53].其中,疏林和灌丛是最常见的两种,以榆树疏林和小叶锦鸡儿灌丛为典型代表.榆树疏林分布在沙地中东部,物种多样性较高,这与之前对榆树疏林植物多样性的结果相近[54-55].研究区的小叶锦鸡儿灌丛分布在沙地的中西部,水分条件稍差,因而多样性水平较低,这是草原灌丛化对群落多样性和生态系统功能的影响[56].研究表明,物种多样性的大尺度格局由水分和温度共同决定[57-58].在水分条件充足时,温度是影响物种丰富度的主要因子,而在降水不足时则受降水量控制[59-60].降水量是影响浑善达克沙地植被的主要气候因素[61];本研究证明,植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数与生长季降水量呈显著的正相关关系,这与前期的研究是一致的,水分条件是影响物种多样性的最主要因子[62-64]. ...
Climatic gradients in woody plant species richness:towards an explanation based on an analysis of southern Africa’s woody flora
1
1993
... 气候是植被大尺度分布格局的决定性因子[47],植物种群和群落的分布都是由气候(主要是水热条件)决定的.降水量被认为是影响干旱区、半干旱区荒漠植被和沙地植被的最主要因子,有研究表明,地下水位及土壤阳离子对于毛乌素沙地丘间地植物群落分布产生重要影响[48-51];浑善达克沙地镶嵌于典型草原地带之中[52],年降水量从东部的440 mm向西逐渐减少至150 mm[28].随着降水量的递减,自东向西依次分布着疏林、灌丛、荒漠等植被类型,在空间上呈现出斑块镶嵌的景观[53].其中,疏林和灌丛是最常见的两种,以榆树疏林和小叶锦鸡儿灌丛为典型代表.榆树疏林分布在沙地中东部,物种多样性较高,这与之前对榆树疏林植物多样性的结果相近[54-55].研究区的小叶锦鸡儿灌丛分布在沙地的中西部,水分条件稍差,因而多样性水平较低,这是草原灌丛化对群落多样性和生态系统功能的影响[56].研究表明,物种多样性的大尺度格局由水分和温度共同决定[57-58].在水分条件充足时,温度是影响物种丰富度的主要因子,而在降水不足时则受降水量控制[59-60].降水量是影响浑善达克沙地植被的主要气候因素[61];本研究证明,植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数与生长季降水量呈显著的正相关关系,这与前期的研究是一致的,水分条件是影响物种多样性的最主要因子[62-64]. ...
Productivity and history as predictors of the latitudinal diversity gradient of terrestrial birds
2
2003
... 气候是植被大尺度分布格局的决定性因子[47],植物种群和群落的分布都是由气候(主要是水热条件)决定的.降水量被认为是影响干旱区、半干旱区荒漠植被和沙地植被的最主要因子,有研究表明,地下水位及土壤阳离子对于毛乌素沙地丘间地植物群落分布产生重要影响[48-51];浑善达克沙地镶嵌于典型草原地带之中[52],年降水量从东部的440 mm向西逐渐减少至150 mm[28].随着降水量的递减,自东向西依次分布着疏林、灌丛、荒漠等植被类型,在空间上呈现出斑块镶嵌的景观[53].其中,疏林和灌丛是最常见的两种,以榆树疏林和小叶锦鸡儿灌丛为典型代表.榆树疏林分布在沙地中东部,物种多样性较高,这与之前对榆树疏林植物多样性的结果相近[54-55].研究区的小叶锦鸡儿灌丛分布在沙地的中西部,水分条件稍差,因而多样性水平较低,这是草原灌丛化对群落多样性和生态系统功能的影响[56].研究表明,物种多样性的大尺度格局由水分和温度共同决定[57-58].在水分条件充足时,温度是影响物种丰富度的主要因子,而在降水不足时则受降水量控制[59-60].降水量是影响浑善达克沙地植被的主要气候因素[61];本研究证明,植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数与生长季降水量呈显著的正相关关系,这与前期的研究是一致的,水分条件是影响物种多样性的最主要因子[62-64]. ...
... 温度因子是影响植物群落物种多样性的重要因子[60].一般认为物种多样性与温度因子呈正相关关系[65],而在内蒙古干旱、半干旱区的部分研究显示,物种多样性却与年均温呈显著的负相关关系[7,61,66-68].本研究结果与后者基本一致,Patrick指数、Shannon-Wiener指数、Simpson指数和Pielou指数均与生长季均温呈显著负相关关系.这说明,温度因子确实对研究区内的物种多样性存在影响.呈现负相关的原因可能有两个:一是尺度问题,浑善达克沙地东西跨度大而南北跨度小,温度因子的梯度变化不明显,气候因子的解释率反而下降[67];二是生长季温度高的地方往往蒸发量也高,而蒸发量与水分因子呈现负相关[69].温度因子对物种多样性的影响还需进一步探讨. ...
荒漠草原物种多样性、生产力与气候因子和土壤养分之间关系的研究
3
2015
... 气候是植被大尺度分布格局的决定性因子[47],植物种群和群落的分布都是由气候(主要是水热条件)决定的.降水量被认为是影响干旱区、半干旱区荒漠植被和沙地植被的最主要因子,有研究表明,地下水位及土壤阳离子对于毛乌素沙地丘间地植物群落分布产生重要影响[48-51];浑善达克沙地镶嵌于典型草原地带之中[52],年降水量从东部的440 mm向西逐渐减少至150 mm[28].随着降水量的递减,自东向西依次分布着疏林、灌丛、荒漠等植被类型,在空间上呈现出斑块镶嵌的景观[53].其中,疏林和灌丛是最常见的两种,以榆树疏林和小叶锦鸡儿灌丛为典型代表.榆树疏林分布在沙地中东部,物种多样性较高,这与之前对榆树疏林植物多样性的结果相近[54-55].研究区的小叶锦鸡儿灌丛分布在沙地的中西部,水分条件稍差,因而多样性水平较低,这是草原灌丛化对群落多样性和生态系统功能的影响[56].研究表明,物种多样性的大尺度格局由水分和温度共同决定[57-58].在水分条件充足时,温度是影响物种丰富度的主要因子,而在降水不足时则受降水量控制[59-60].降水量是影响浑善达克沙地植被的主要气候因素[61];本研究证明,植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数与生长季降水量呈显著的正相关关系,这与前期的研究是一致的,水分条件是影响物种多样性的最主要因子[62-64]. ...
... 温度因子是影响植物群落物种多样性的重要因子[60].一般认为物种多样性与温度因子呈正相关关系[65],而在内蒙古干旱、半干旱区的部分研究显示,物种多样性却与年均温呈显著的负相关关系[7,61,66-68].本研究结果与后者基本一致,Patrick指数、Shannon-Wiener指数、Simpson指数和Pielou指数均与生长季均温呈显著负相关关系.这说明,温度因子确实对研究区内的物种多样性存在影响.呈现负相关的原因可能有两个:一是尺度问题,浑善达克沙地东西跨度大而南北跨度小,温度因子的梯度变化不明显,气候因子的解释率反而下降[67];二是生长季温度高的地方往往蒸发量也高,而蒸发量与水分因子呈现负相关[69].温度因子对物种多样性的影响还需进一步探讨. ...
... 土壤是决定小尺度植被分布的重要环境因子,沙地土壤质地、理化因子、微生物等都对植被的分布有直接的影响[70-71].土壤和植被是土壤-植被-大气连续体(Soil-Plant-Atmosphere-Continuum,SPAC)的共同组成部分[72],是相互依存、相互影响的因子,对二者之间关系的研究具有重要意义[67].目前针对这方面的研究很多,有研究显示土壤中氮含量与物种多样性呈正相关[73],也有试验显示氮素添加会降低物种多样性[74-75];还有研究指出,在贫瘠土壤上氮含量增加可以提高物种丰富度,而养分丰富的土壤提高含氮量可能会破坏原有生态系统的共存机制而造成物种丰富度的降低[76].位于内蒙古草原上的浑善达克沙地属于氮限制地区,土壤氮含量较高的区域群落的物种多样性相应也会提高[76].本研究结果基本一致,土壤有效氮与Patrick指数、Simpson指数和Shannon-Wiener指数均呈正相关关系.对土壤磷含量与物种多样性的研究显示,土壤磷含量与物种丰富度呈现负相关关系[67,77-78],本研究结果显示出类似的趋势,Shannon-Wiener指数随土壤全磷含量的增加而降低.土壤有机碳是表征土壤肥力的重要指标,不仅代表土壤有机质的水平,还能说明氮、磷等元素的可利用状态[79].土壤有机碳的含量越高,土壤肥力越高,可支持的物种多样性水平也越高.土壤有机碳与多样性指数呈现正相关[7,61,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...
Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau
1
2008
... 气候是植被大尺度分布格局的决定性因子[47],植物种群和群落的分布都是由气候(主要是水热条件)决定的.降水量被认为是影响干旱区、半干旱区荒漠植被和沙地植被的最主要因子,有研究表明,地下水位及土壤阳离子对于毛乌素沙地丘间地植物群落分布产生重要影响[48-51];浑善达克沙地镶嵌于典型草原地带之中[52],年降水量从东部的440 mm向西逐渐减少至150 mm[28].随着降水量的递减,自东向西依次分布着疏林、灌丛、荒漠等植被类型,在空间上呈现出斑块镶嵌的景观[53].其中,疏林和灌丛是最常见的两种,以榆树疏林和小叶锦鸡儿灌丛为典型代表.榆树疏林分布在沙地中东部,物种多样性较高,这与之前对榆树疏林植物多样性的结果相近[54-55].研究区的小叶锦鸡儿灌丛分布在沙地的中西部,水分条件稍差,因而多样性水平较低,这是草原灌丛化对群落多样性和生态系统功能的影响[56].研究表明,物种多样性的大尺度格局由水分和温度共同决定[57-58].在水分条件充足时,温度是影响物种丰富度的主要因子,而在降水不足时则受降水量控制[59-60].降水量是影响浑善达克沙地植被的主要气候因素[61];本研究证明,植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数与生长季降水量呈显著的正相关关系,这与前期的研究是一致的,水分条件是影响物种多样性的最主要因子[62-64]. ...
Stability of tallgrass prairie during a 19-year increase in growing season precipitation
2012
Climate warming and precipitation redistribution modify tree grass interactions and tree species establishment in a warm temperate savanna
1
2013
... 气候是植被大尺度分布格局的决定性因子[47],植物种群和群落的分布都是由气候(主要是水热条件)决定的.降水量被认为是影响干旱区、半干旱区荒漠植被和沙地植被的最主要因子,有研究表明,地下水位及土壤阳离子对于毛乌素沙地丘间地植物群落分布产生重要影响[48-51];浑善达克沙地镶嵌于典型草原地带之中[52],年降水量从东部的440 mm向西逐渐减少至150 mm[28].随着降水量的递减,自东向西依次分布着疏林、灌丛、荒漠等植被类型,在空间上呈现出斑块镶嵌的景观[53].其中,疏林和灌丛是最常见的两种,以榆树疏林和小叶锦鸡儿灌丛为典型代表.榆树疏林分布在沙地中东部,物种多样性较高,这与之前对榆树疏林植物多样性的结果相近[54-55].研究区的小叶锦鸡儿灌丛分布在沙地的中西部,水分条件稍差,因而多样性水平较低,这是草原灌丛化对群落多样性和生态系统功能的影响[56].研究表明,物种多样性的大尺度格局由水分和温度共同决定[57-58].在水分条件充足时,温度是影响物种丰富度的主要因子,而在降水不足时则受降水量控制[59-60].降水量是影响浑善达克沙地植被的主要气候因素[61];本研究证明,植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数与生长季降水量呈显著的正相关关系,这与前期的研究是一致的,水分条件是影响物种多样性的最主要因子[62-64]. ...
A globally consistent richness-climate relationship for angiosperms
1
2003
... 温度因子是影响植物群落物种多样性的重要因子[60].一般认为物种多样性与温度因子呈正相关关系[65],而在内蒙古干旱、半干旱区的部分研究显示,物种多样性却与年均温呈显著的负相关关系[7,61,66-68].本研究结果与后者基本一致,Patrick指数、Shannon-Wiener指数、Simpson指数和Pielou指数均与生长季均温呈显著负相关关系.这说明,温度因子确实对研究区内的物种多样性存在影响.呈现负相关的原因可能有两个:一是尺度问题,浑善达克沙地东西跨度大而南北跨度小,温度因子的梯度变化不明显,气候因子的解释率反而下降[67];二是生长季温度高的地方往往蒸发量也高,而蒸发量与水分因子呈现负相关[69].温度因子对物种多样性的影响还需进一步探讨. ...
内蒙古锡林河流域植被多样性特点及其与气候因子的关系
1
1998
... 温度因子是影响植物群落物种多样性的重要因子[60].一般认为物种多样性与温度因子呈正相关关系[65],而在内蒙古干旱、半干旱区的部分研究显示,物种多样性却与年均温呈显著的负相关关系[7,61,66-68].本研究结果与后者基本一致,Patrick指数、Shannon-Wiener指数、Simpson指数和Pielou指数均与生长季均温呈显著负相关关系.这说明,温度因子确实对研究区内的物种多样性存在影响.呈现负相关的原因可能有两个:一是尺度问题,浑善达克沙地东西跨度大而南北跨度小,温度因子的梯度变化不明显,气候因子的解释率反而下降[67];二是生长季温度高的地方往往蒸发量也高,而蒸发量与水分因子呈现负相关[69].温度因子对物种多样性的影响还需进一步探讨. ...
锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究
4
2000
... 温度因子是影响植物群落物种多样性的重要因子[60].一般认为物种多样性与温度因子呈正相关关系[65],而在内蒙古干旱、半干旱区的部分研究显示,物种多样性却与年均温呈显著的负相关关系[7,61,66-68].本研究结果与后者基本一致,Patrick指数、Shannon-Wiener指数、Simpson指数和Pielou指数均与生长季均温呈显著负相关关系.这说明,温度因子确实对研究区内的物种多样性存在影响.呈现负相关的原因可能有两个:一是尺度问题,浑善达克沙地东西跨度大而南北跨度小,温度因子的梯度变化不明显,气候因子的解释率反而下降[67];二是生长季温度高的地方往往蒸发量也高,而蒸发量与水分因子呈现负相关[69].温度因子对物种多样性的影响还需进一步探讨. ...
... 土壤是决定小尺度植被分布的重要环境因子,沙地土壤质地、理化因子、微生物等都对植被的分布有直接的影响[70-71].土壤和植被是土壤-植被-大气连续体(Soil-Plant-Atmosphere-Continuum,SPAC)的共同组成部分[72],是相互依存、相互影响的因子,对二者之间关系的研究具有重要意义[67].目前针对这方面的研究很多,有研究显示土壤中氮含量与物种多样性呈正相关[73],也有试验显示氮素添加会降低物种多样性[74-75];还有研究指出,在贫瘠土壤上氮含量增加可以提高物种丰富度,而养分丰富的土壤提高含氮量可能会破坏原有生态系统的共存机制而造成物种丰富度的降低[76].位于内蒙古草原上的浑善达克沙地属于氮限制地区,土壤氮含量较高的区域群落的物种多样性相应也会提高[76].本研究结果基本一致,土壤有效氮与Patrick指数、Simpson指数和Shannon-Wiener指数均呈正相关关系.对土壤磷含量与物种多样性的研究显示,土壤磷含量与物种丰富度呈现负相关关系[67,77-78],本研究结果显示出类似的趋势,Shannon-Wiener指数随土壤全磷含量的增加而降低.土壤有机碳是表征土壤肥力的重要指标,不仅代表土壤有机质的水平,还能说明氮、磷等元素的可利用状态[79].土壤有机碳的含量越高,土壤肥力越高,可支持的物种多样性水平也越高.土壤有机碳与多样性指数呈现正相关[7,61,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...
... [67,77-78],本研究结果显示出类似的趋势,Shannon-Wiener指数随土壤全磷含量的增加而降低.土壤有机碳是表征土壤肥力的重要指标,不仅代表土壤有机质的水平,还能说明氮、磷等元素的可利用状态[79].土壤有机碳的含量越高,土壤肥力越高,可支持的物种多样性水平也越高.土壤有机碳与多样性指数呈现正相关[7,61,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...
... ,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...
Plant species diversity is correlated with climatic factors differently at the community and the functional group levels:a case study of desert steppe in Inner Mongolia,China
1
2016
... 温度因子是影响植物群落物种多样性的重要因子[60].一般认为物种多样性与温度因子呈正相关关系[65],而在内蒙古干旱、半干旱区的部分研究显示,物种多样性却与年均温呈显著的负相关关系[7,61,66-68].本研究结果与后者基本一致,Patrick指数、Shannon-Wiener指数、Simpson指数和Pielou指数均与生长季均温呈显著负相关关系.这说明,温度因子确实对研究区内的物种多样性存在影响.呈现负相关的原因可能有两个:一是尺度问题,浑善达克沙地东西跨度大而南北跨度小,温度因子的梯度变化不明显,气候因子的解释率反而下降[67];二是生长季温度高的地方往往蒸发量也高,而蒸发量与水分因子呈现负相关[69].温度因子对物种多样性的影响还需进一步探讨. ...
Energy and large-scale patterns of animal and plant-species richness
1
1991
... 温度因子是影响植物群落物种多样性的重要因子[60].一般认为物种多样性与温度因子呈正相关关系[65],而在内蒙古干旱、半干旱区的部分研究显示,物种多样性却与年均温呈显著的负相关关系[7,61,66-68].本研究结果与后者基本一致,Patrick指数、Shannon-Wiener指数、Simpson指数和Pielou指数均与生长季均温呈显著负相关关系.这说明,温度因子确实对研究区内的物种多样性存在影响.呈现负相关的原因可能有两个:一是尺度问题,浑善达克沙地东西跨度大而南北跨度小,温度因子的梯度变化不明显,气候因子的解释率反而下降[67];二是生长季温度高的地方往往蒸发量也高,而蒸发量与水分因子呈现负相关[69].温度因子对物种多样性的影响还需进一步探讨. ...
呼伦贝尔沙地北部沙带植物群落分布格局与土壤特性的关系
1
2020
... 土壤是决定小尺度植被分布的重要环境因子,沙地土壤质地、理化因子、微生物等都对植被的分布有直接的影响[70-71].土壤和植被是土壤-植被-大气连续体(Soil-Plant-Atmosphere-Continuum,SPAC)的共同组成部分[72],是相互依存、相互影响的因子,对二者之间关系的研究具有重要意义[67].目前针对这方面的研究很多,有研究显示土壤中氮含量与物种多样性呈正相关[73],也有试验显示氮素添加会降低物种多样性[74-75];还有研究指出,在贫瘠土壤上氮含量增加可以提高物种丰富度,而养分丰富的土壤提高含氮量可能会破坏原有生态系统的共存机制而造成物种丰富度的降低[76].位于内蒙古草原上的浑善达克沙地属于氮限制地区,土壤氮含量较高的区域群落的物种多样性相应也会提高[76].本研究结果基本一致,土壤有效氮与Patrick指数、Simpson指数和Shannon-Wiener指数均呈正相关关系.对土壤磷含量与物种多样性的研究显示,土壤磷含量与物种丰富度呈现负相关关系[67,77-78],本研究结果显示出类似的趋势,Shannon-Wiener指数随土壤全磷含量的增加而降低.土壤有机碳是表征土壤肥力的重要指标,不仅代表土壤有机质的水平,还能说明氮、磷等元素的可利用状态[79].土壤有机碳的含量越高,土壤肥力越高,可支持的物种多样性水平也越高.土壤有机碳与多样性指数呈现正相关[7,61,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...
毛乌素沙地丘间低地主要植物群落土壤酶活性
1
2016
... 土壤是决定小尺度植被分布的重要环境因子,沙地土壤质地、理化因子、微生物等都对植被的分布有直接的影响[70-71].土壤和植被是土壤-植被-大气连续体(Soil-Plant-Atmosphere-Continuum,SPAC)的共同组成部分[72],是相互依存、相互影响的因子,对二者之间关系的研究具有重要意义[67].目前针对这方面的研究很多,有研究显示土壤中氮含量与物种多样性呈正相关[73],也有试验显示氮素添加会降低物种多样性[74-75];还有研究指出,在贫瘠土壤上氮含量增加可以提高物种丰富度,而养分丰富的土壤提高含氮量可能会破坏原有生态系统的共存机制而造成物种丰富度的降低[76].位于内蒙古草原上的浑善达克沙地属于氮限制地区,土壤氮含量较高的区域群落的物种多样性相应也会提高[76].本研究结果基本一致,土壤有效氮与Patrick指数、Simpson指数和Shannon-Wiener指数均呈正相关关系.对土壤磷含量与物种多样性的研究显示,土壤磷含量与物种丰富度呈现负相关关系[67,77-78],本研究结果显示出类似的趋势,Shannon-Wiener指数随土壤全磷含量的增加而降低.土壤有机碳是表征土壤肥力的重要指标,不仅代表土壤有机质的水平,还能说明氮、磷等元素的可利用状态[79].土壤有机碳的含量越高,土壤肥力越高,可支持的物种多样性水平也越高.土壤有机碳与多样性指数呈现正相关[7,61,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...
气候变暖背景下近30a北半球植被变化研究综述
1
2016
... 土壤是决定小尺度植被分布的重要环境因子,沙地土壤质地、理化因子、微生物等都对植被的分布有直接的影响[70-71].土壤和植被是土壤-植被-大气连续体(Soil-Plant-Atmosphere-Continuum,SPAC)的共同组成部分[72],是相互依存、相互影响的因子,对二者之间关系的研究具有重要意义[67].目前针对这方面的研究很多,有研究显示土壤中氮含量与物种多样性呈正相关[73],也有试验显示氮素添加会降低物种多样性[74-75];还有研究指出,在贫瘠土壤上氮含量增加可以提高物种丰富度,而养分丰富的土壤提高含氮量可能会破坏原有生态系统的共存机制而造成物种丰富度的降低[76].位于内蒙古草原上的浑善达克沙地属于氮限制地区,土壤氮含量较高的区域群落的物种多样性相应也会提高[76].本研究结果基本一致,土壤有效氮与Patrick指数、Simpson指数和Shannon-Wiener指数均呈正相关关系.对土壤磷含量与物种多样性的研究显示,土壤磷含量与物种丰富度呈现负相关关系[67,77-78],本研究结果显示出类似的趋势,Shannon-Wiener指数随土壤全磷含量的增加而降低.土壤有机碳是表征土壤肥力的重要指标,不仅代表土壤有机质的水平,还能说明氮、磷等元素的可利用状态[79].土壤有机碳的含量越高,土壤肥力越高,可支持的物种多样性水平也越高.土壤有机碳与多样性指数呈现正相关[7,61,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...
Relationship between soil nutrient availability and plant species richness in a tropical semi-arid environment
1
2006
... 土壤是决定小尺度植被分布的重要环境因子,沙地土壤质地、理化因子、微生物等都对植被的分布有直接的影响[70-71].土壤和植被是土壤-植被-大气连续体(Soil-Plant-Atmosphere-Continuum,SPAC)的共同组成部分[72],是相互依存、相互影响的因子,对二者之间关系的研究具有重要意义[67].目前针对这方面的研究很多,有研究显示土壤中氮含量与物种多样性呈正相关[73],也有试验显示氮素添加会降低物种多样性[74-75];还有研究指出,在贫瘠土壤上氮含量增加可以提高物种丰富度,而养分丰富的土壤提高含氮量可能会破坏原有生态系统的共存机制而造成物种丰富度的降低[76].位于内蒙古草原上的浑善达克沙地属于氮限制地区,土壤氮含量较高的区域群落的物种多样性相应也会提高[76].本研究结果基本一致,土壤有效氮与Patrick指数、Simpson指数和Shannon-Wiener指数均呈正相关关系.对土壤磷含量与物种多样性的研究显示,土壤磷含量与物种丰富度呈现负相关关系[67,77-78],本研究结果显示出类似的趋势,Shannon-Wiener指数随土壤全磷含量的增加而降低.土壤有机碳是表征土壤肥力的重要指标,不仅代表土壤有机质的水平,还能说明氮、磷等元素的可利用状态[79].土壤有机碳的含量越高,土壤肥力越高,可支持的物种多样性水平也越高.土壤有机碳与多样性指数呈现正相关[7,61,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...
Regional and local patterns in plant species richness with respect to resource availability
1
2003
... 土壤是决定小尺度植被分布的重要环境因子,沙地土壤质地、理化因子、微生物等都对植被的分布有直接的影响[70-71].土壤和植被是土壤-植被-大气连续体(Soil-Plant-Atmosphere-Continuum,SPAC)的共同组成部分[72],是相互依存、相互影响的因子,对二者之间关系的研究具有重要意义[67].目前针对这方面的研究很多,有研究显示土壤中氮含量与物种多样性呈正相关[73],也有试验显示氮素添加会降低物种多样性[74-75];还有研究指出,在贫瘠土壤上氮含量增加可以提高物种丰富度,而养分丰富的土壤提高含氮量可能会破坏原有生态系统的共存机制而造成物种丰富度的降低[76].位于内蒙古草原上的浑善达克沙地属于氮限制地区,土壤氮含量较高的区域群落的物种多样性相应也会提高[76].本研究结果基本一致,土壤有效氮与Patrick指数、Simpson指数和Shannon-Wiener指数均呈正相关关系.对土壤磷含量与物种多样性的研究显示,土壤磷含量与物种丰富度呈现负相关关系[67,77-78],本研究结果显示出类似的趋势,Shannon-Wiener指数随土壤全磷含量的增加而降低.土壤有机碳是表征土壤肥力的重要指标,不仅代表土壤有机质的水平,还能说明氮、磷等元素的可利用状态[79].土壤有机碳的含量越高,土壤肥力越高,可支持的物种多样性水平也越高.土壤有机碳与多样性指数呈现正相关[7,61,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...
Effects of nitrogen addition on species richness and relationship between species richness and aboveground productivity of alpine meadow of the Qinghai-Tibetan Plateau,China
1
2010
... 土壤是决定小尺度植被分布的重要环境因子,沙地土壤质地、理化因子、微生物等都对植被的分布有直接的影响[70-71].土壤和植被是土壤-植被-大气连续体(Soil-Plant-Atmosphere-Continuum,SPAC)的共同组成部分[72],是相互依存、相互影响的因子,对二者之间关系的研究具有重要意义[67].目前针对这方面的研究很多,有研究显示土壤中氮含量与物种多样性呈正相关[73],也有试验显示氮素添加会降低物种多样性[74-75];还有研究指出,在贫瘠土壤上氮含量增加可以提高物种丰富度,而养分丰富的土壤提高含氮量可能会破坏原有生态系统的共存机制而造成物种丰富度的降低[76].位于内蒙古草原上的浑善达克沙地属于氮限制地区,土壤氮含量较高的区域群落的物种多样性相应也会提高[76].本研究结果基本一致,土壤有效氮与Patrick指数、Simpson指数和Shannon-Wiener指数均呈正相关关系.对土壤磷含量与物种多样性的研究显示,土壤磷含量与物种丰富度呈现负相关关系[67,77-78],本研究结果显示出类似的趋势,Shannon-Wiener指数随土壤全磷含量的增加而降低.土壤有机碳是表征土壤肥力的重要指标,不仅代表土壤有机质的水平,还能说明氮、磷等元素的可利用状态[79].土壤有机碳的含量越高,土壤肥力越高,可支持的物种多样性水平也越高.土壤有机碳与多样性指数呈现正相关[7,61,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...
Trade offs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning:evidence from Inner Mongolia Grasslands
2
2010
... 土壤是决定小尺度植被分布的重要环境因子,沙地土壤质地、理化因子、微生物等都对植被的分布有直接的影响[70-71].土壤和植被是土壤-植被-大气连续体(Soil-Plant-Atmosphere-Continuum,SPAC)的共同组成部分[72],是相互依存、相互影响的因子,对二者之间关系的研究具有重要意义[67].目前针对这方面的研究很多,有研究显示土壤中氮含量与物种多样性呈正相关[73],也有试验显示氮素添加会降低物种多样性[74-75];还有研究指出,在贫瘠土壤上氮含量增加可以提高物种丰富度,而养分丰富的土壤提高含氮量可能会破坏原有生态系统的共存机制而造成物种丰富度的降低[76].位于内蒙古草原上的浑善达克沙地属于氮限制地区,土壤氮含量较高的区域群落的物种多样性相应也会提高[76].本研究结果基本一致,土壤有效氮与Patrick指数、Simpson指数和Shannon-Wiener指数均呈正相关关系.对土壤磷含量与物种多样性的研究显示,土壤磷含量与物种丰富度呈现负相关关系[67,77-78],本研究结果显示出类似的趋势,Shannon-Wiener指数随土壤全磷含量的增加而降低.土壤有机碳是表征土壤肥力的重要指标,不仅代表土壤有机质的水平,还能说明氮、磷等元素的可利用状态[79].土壤有机碳的含量越高,土壤肥力越高,可支持的物种多样性水平也越高.土壤有机碳与多样性指数呈现正相关[7,61,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...
... [76].本研究结果基本一致,土壤有效氮与Patrick指数、Simpson指数和Shannon-Wiener指数均呈正相关关系.对土壤磷含量与物种多样性的研究显示,土壤磷含量与物种丰富度呈现负相关关系[67,77-78],本研究结果显示出类似的趋势,Shannon-Wiener指数随土壤全磷含量的增加而降低.土壤有机碳是表征土壤肥力的重要指标,不仅代表土壤有机质的水平,还能说明氮、磷等元素的可利用状态[79].土壤有机碳的含量越高,土壤肥力越高,可支持的物种多样性水平也越高.土壤有机碳与多样性指数呈现正相关[7,61,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...
Biological phosphorus cycling in grasslands:interactions with nitrogen
1
2011
... 土壤是决定小尺度植被分布的重要环境因子,沙地土壤质地、理化因子、微生物等都对植被的分布有直接的影响[70-71].土壤和植被是土壤-植被-大气连续体(Soil-Plant-Atmosphere-Continuum,SPAC)的共同组成部分[72],是相互依存、相互影响的因子,对二者之间关系的研究具有重要意义[67].目前针对这方面的研究很多,有研究显示土壤中氮含量与物种多样性呈正相关[73],也有试验显示氮素添加会降低物种多样性[74-75];还有研究指出,在贫瘠土壤上氮含量增加可以提高物种丰富度,而养分丰富的土壤提高含氮量可能会破坏原有生态系统的共存机制而造成物种丰富度的降低[76].位于内蒙古草原上的浑善达克沙地属于氮限制地区,土壤氮含量较高的区域群落的物种多样性相应也会提高[76].本研究结果基本一致,土壤有效氮与Patrick指数、Simpson指数和Shannon-Wiener指数均呈正相关关系.对土壤磷含量与物种多样性的研究显示,土壤磷含量与物种丰富度呈现负相关关系[67,77-78],本研究结果显示出类似的趋势,Shannon-Wiener指数随土壤全磷含量的增加而降低.土壤有机碳是表征土壤肥力的重要指标,不仅代表土壤有机质的水平,还能说明氮、磷等元素的可利用状态[79].土壤有机碳的含量越高,土壤肥力越高,可支持的物种多样性水平也越高.土壤有机碳与多样性指数呈现正相关[7,61,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...
Plant species identity and soil P forms in an oligotrophic grassland-desert scrub system
1
2014
... 土壤是决定小尺度植被分布的重要环境因子,沙地土壤质地、理化因子、微生物等都对植被的分布有直接的影响[70-71].土壤和植被是土壤-植被-大气连续体(Soil-Plant-Atmosphere-Continuum,SPAC)的共同组成部分[72],是相互依存、相互影响的因子,对二者之间关系的研究具有重要意义[67].目前针对这方面的研究很多,有研究显示土壤中氮含量与物种多样性呈正相关[73],也有试验显示氮素添加会降低物种多样性[74-75];还有研究指出,在贫瘠土壤上氮含量增加可以提高物种丰富度,而养分丰富的土壤提高含氮量可能会破坏原有生态系统的共存机制而造成物种丰富度的降低[76].位于内蒙古草原上的浑善达克沙地属于氮限制地区,土壤氮含量较高的区域群落的物种多样性相应也会提高[76].本研究结果基本一致,土壤有效氮与Patrick指数、Simpson指数和Shannon-Wiener指数均呈正相关关系.对土壤磷含量与物种多样性的研究显示,土壤磷含量与物种丰富度呈现负相关关系[67,77-78],本研究结果显示出类似的趋势,Shannon-Wiener指数随土壤全磷含量的增加而降低.土壤有机碳是表征土壤肥力的重要指标,不仅代表土壤有机质的水平,还能说明氮、磷等元素的可利用状态[79].土壤有机碳的含量越高,土壤肥力越高,可支持的物种多样性水平也越高.土壤有机碳与多样性指数呈现正相关[7,61,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...
人类活动对锡林郭勒地区主要草原土壤有机碳分布的影响
1
1998
... 土壤是决定小尺度植被分布的重要环境因子,沙地土壤质地、理化因子、微生物等都对植被的分布有直接的影响[70-71].土壤和植被是土壤-植被-大气连续体(Soil-Plant-Atmosphere-Continuum,SPAC)的共同组成部分[72],是相互依存、相互影响的因子,对二者之间关系的研究具有重要意义[67].目前针对这方面的研究很多,有研究显示土壤中氮含量与物种多样性呈正相关[73],也有试验显示氮素添加会降低物种多样性[74-75];还有研究指出,在贫瘠土壤上氮含量增加可以提高物种丰富度,而养分丰富的土壤提高含氮量可能会破坏原有生态系统的共存机制而造成物种丰富度的降低[76].位于内蒙古草原上的浑善达克沙地属于氮限制地区,土壤氮含量较高的区域群落的物种多样性相应也会提高[76].本研究结果基本一致,土壤有效氮与Patrick指数、Simpson指数和Shannon-Wiener指数均呈正相关关系.对土壤磷含量与物种多样性的研究显示,土壤磷含量与物种丰富度呈现负相关关系[67,77-78],本研究结果显示出类似的趋势,Shannon-Wiener指数随土壤全磷含量的增加而降低.土壤有机碳是表征土壤肥力的重要指标,不仅代表土壤有机质的水平,还能说明氮、磷等元素的可利用状态[79].土壤有机碳的含量越高,土壤肥力越高,可支持的物种多样性水平也越高.土壤有机碳与多样性指数呈现正相关[7,61,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...
Changes of species diversity and productivity in relation to soil properties in sandy grassland in Horqin sand land
1
2007
... 土壤是决定小尺度植被分布的重要环境因子,沙地土壤质地、理化因子、微生物等都对植被的分布有直接的影响[70-71].土壤和植被是土壤-植被-大气连续体(Soil-Plant-Atmosphere-Continuum,SPAC)的共同组成部分[72],是相互依存、相互影响的因子,对二者之间关系的研究具有重要意义[67].目前针对这方面的研究很多,有研究显示土壤中氮含量与物种多样性呈正相关[73],也有试验显示氮素添加会降低物种多样性[74-75];还有研究指出,在贫瘠土壤上氮含量增加可以提高物种丰富度,而养分丰富的土壤提高含氮量可能会破坏原有生态系统的共存机制而造成物种丰富度的降低[76].位于内蒙古草原上的浑善达克沙地属于氮限制地区,土壤氮含量较高的区域群落的物种多样性相应也会提高[76].本研究结果基本一致,土壤有效氮与Patrick指数、Simpson指数和Shannon-Wiener指数均呈正相关关系.对土壤磷含量与物种多样性的研究显示,土壤磷含量与物种丰富度呈现负相关关系[67,77-78],本研究结果显示出类似的趋势,Shannon-Wiener指数随土壤全磷含量的增加而降低.土壤有机碳是表征土壤肥力的重要指标,不仅代表土壤有机质的水平,还能说明氮、磷等元素的可利用状态[79].土壤有机碳的含量越高,土壤肥力越高,可支持的物种多样性水平也越高.土壤有机碳与多样性指数呈现正相关[7,61,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...
阿尔金山自然保护区植物群落生物量和物种多样性的空间格局及其影响因素
1
2016
... 土壤是决定小尺度植被分布的重要环境因子,沙地土壤质地、理化因子、微生物等都对植被的分布有直接的影响[70-71].土壤和植被是土壤-植被-大气连续体(Soil-Plant-Atmosphere-Continuum,SPAC)的共同组成部分[72],是相互依存、相互影响的因子,对二者之间关系的研究具有重要意义[67].目前针对这方面的研究很多,有研究显示土壤中氮含量与物种多样性呈正相关[73],也有试验显示氮素添加会降低物种多样性[74-75];还有研究指出,在贫瘠土壤上氮含量增加可以提高物种丰富度,而养分丰富的土壤提高含氮量可能会破坏原有生态系统的共存机制而造成物种丰富度的降低[76].位于内蒙古草原上的浑善达克沙地属于氮限制地区,土壤氮含量较高的区域群落的物种多样性相应也会提高[76].本研究结果基本一致,土壤有效氮与Patrick指数、Simpson指数和Shannon-Wiener指数均呈正相关关系.对土壤磷含量与物种多样性的研究显示,土壤磷含量与物种丰富度呈现负相关关系[67,77-78],本研究结果显示出类似的趋势,Shannon-Wiener指数随土壤全磷含量的增加而降低.土壤有机碳是表征土壤肥力的重要指标,不仅代表土壤有机质的水平,还能说明氮、磷等元素的可利用状态[79].土壤有机碳的含量越高,土壤肥力越高,可支持的物种多样性水平也越高.土壤有机碳与多样性指数呈现正相关[7,61,67,80-81].浑善达克沙地植物群落的Patrick指数、Simpson指数和Shannon-Wiener指数均与有机碳呈显著正相关,这与前人的研究一致,说明土壤养分含量对群落的物种多样性具有一定的促进作用. ...








 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}