Desertification
1
2001
... 荒漠是陆地生态系统的重要组成部分,面积约占世界陆地面积的1/3[1-3].在气候变化和人类活动加剧的背景下,荒漠草地很容易退化或沙漠化[4-5],因此荒漠草地在全球碳循环中可能起着更加重要的作用.近年来,沙漠化程度不断加剧,导致荒漠草地生态系统不断退化,如植被、土壤结构的退化致使区域生态系统固碳能力下降,同时向大气中释放了大量的碳[6].全球沙漠化导致荒漠生态系统的碳损失为19—29 Pg,但这一数据还存在很大的不确定性[7].为了减缓沙漠化,促进生态系统稳定性,在荒漠地区进行植被建设已成为一种重要的管理措施.在荒漠化严重的地区进行合理的人工植被建设是提高土地生产力和生物多样性、增加荒漠生态系统碳储量和减缓全球气候变化的有效方式[8-11].但是典型荒漠草地在退化过程中究竟损失了多少碳,以及退化严重的荒漠草地的固碳潜力究竟有多大,目前还缺少强有力的证据.在区域尺度上针对这一问题开展研究是减少荒漠生态系统碳储量和固碳潜力估算的不确定性以及应对全球气候变化的关键. ...
Estimating net primary productivity from grassland biomass dynamics measurements
0
2002
Global desertification: building a science for dryland development
1
2007
... 荒漠是陆地生态系统的重要组成部分,面积约占世界陆地面积的1/3[1-3].在气候变化和人类活动加剧的背景下,荒漠草地很容易退化或沙漠化[4-5],因此荒漠草地在全球碳循环中可能起着更加重要的作用.近年来,沙漠化程度不断加剧,导致荒漠草地生态系统不断退化,如植被、土壤结构的退化致使区域生态系统固碳能力下降,同时向大气中释放了大量的碳[6].全球沙漠化导致荒漠生态系统的碳损失为19—29 Pg,但这一数据还存在很大的不确定性[7].为了减缓沙漠化,促进生态系统稳定性,在荒漠地区进行植被建设已成为一种重要的管理措施.在荒漠化严重的地区进行合理的人工植被建设是提高土地生产力和生物多样性、增加荒漠生态系统碳储量和减缓全球气候变化的有效方式[8-11].但是典型荒漠草地在退化过程中究竟损失了多少碳,以及退化严重的荒漠草地的固碳潜力究竟有多大,目前还缺少强有力的证据.在区域尺度上针对这一问题开展研究是减少荒漠生态系统碳储量和固碳潜力估算的不确定性以及应对全球气候变化的关键. ...
Biological feedbacks in global desertification
1
1990
... 荒漠是陆地生态系统的重要组成部分,面积约占世界陆地面积的1/3[1-3].在气候变化和人类活动加剧的背景下,荒漠草地很容易退化或沙漠化[4-5],因此荒漠草地在全球碳循环中可能起着更加重要的作用.近年来,沙漠化程度不断加剧,导致荒漠草地生态系统不断退化,如植被、土壤结构的退化致使区域生态系统固碳能力下降,同时向大气中释放了大量的碳[6].全球沙漠化导致荒漠生态系统的碳损失为19—29 Pg,但这一数据还存在很大的不确定性[7].为了减缓沙漠化,促进生态系统稳定性,在荒漠地区进行植被建设已成为一种重要的管理措施.在荒漠化严重的地区进行合理的人工植被建设是提高土地生产力和生物多样性、增加荒漠生态系统碳储量和减缓全球气候变化的有效方式[8-11].但是典型荒漠草地在退化过程中究竟损失了多少碳,以及退化严重的荒漠草地的固碳潜力究竟有多大,目前还缺少强有力的证据.在区域尺度上针对这一问题开展研究是减少荒漠生态系统碳储量和固碳潜力估算的不确定性以及应对全球气候变化的关键. ...
Perspectives on desertification: western Mediterranean
1
1998
... 荒漠是陆地生态系统的重要组成部分,面积约占世界陆地面积的1/3[1-3].在气候变化和人类活动加剧的背景下,荒漠草地很容易退化或沙漠化[4-5],因此荒漠草地在全球碳循环中可能起着更加重要的作用.近年来,沙漠化程度不断加剧,导致荒漠草地生态系统不断退化,如植被、土壤结构的退化致使区域生态系统固碳能力下降,同时向大气中释放了大量的碳[6].全球沙漠化导致荒漠生态系统的碳损失为19—29 Pg,但这一数据还存在很大的不确定性[7].为了减缓沙漠化,促进生态系统稳定性,在荒漠地区进行植被建设已成为一种重要的管理措施.在荒漠化严重的地区进行合理的人工植被建设是提高土地生产力和生物多样性、增加荒漠生态系统碳储量和减缓全球气候变化的有效方式[8-11].但是典型荒漠草地在退化过程中究竟损失了多少碳,以及退化严重的荒漠草地的固碳潜力究竟有多大,目前还缺少强有力的证据.在区域尺度上针对这一问题开展研究是减少荒漠生态系统碳储量和固碳潜力估算的不确定性以及应对全球气候变化的关键. ...
Regional desertification:a global synthesis
1
2008
... 荒漠是陆地生态系统的重要组成部分,面积约占世界陆地面积的1/3[1-3].在气候变化和人类活动加剧的背景下,荒漠草地很容易退化或沙漠化[4-5],因此荒漠草地在全球碳循环中可能起着更加重要的作用.近年来,沙漠化程度不断加剧,导致荒漠草地生态系统不断退化,如植被、土壤结构的退化致使区域生态系统固碳能力下降,同时向大气中释放了大量的碳[6].全球沙漠化导致荒漠生态系统的碳损失为19—29 Pg,但这一数据还存在很大的不确定性[7].为了减缓沙漠化,促进生态系统稳定性,在荒漠地区进行植被建设已成为一种重要的管理措施.在荒漠化严重的地区进行合理的人工植被建设是提高土地生产力和生物多样性、增加荒漠生态系统碳储量和减缓全球气候变化的有效方式[8-11].但是典型荒漠草地在退化过程中究竟损失了多少碳,以及退化严重的荒漠草地的固碳潜力究竟有多大,目前还缺少强有力的证据.在区域尺度上针对这一问题开展研究是减少荒漠生态系统碳储量和固碳潜力估算的不确定性以及应对全球气候变化的关键. ...
Potential of desertification control to sequester carbon and mitigate the greenhouse effect
1
2001
... 荒漠是陆地生态系统的重要组成部分,面积约占世界陆地面积的1/3[1-3].在气候变化和人类活动加剧的背景下,荒漠草地很容易退化或沙漠化[4-5],因此荒漠草地在全球碳循环中可能起着更加重要的作用.近年来,沙漠化程度不断加剧,导致荒漠草地生态系统不断退化,如植被、土壤结构的退化致使区域生态系统固碳能力下降,同时向大气中释放了大量的碳[6].全球沙漠化导致荒漠生态系统的碳损失为19—29 Pg,但这一数据还存在很大的不确定性[7].为了减缓沙漠化,促进生态系统稳定性,在荒漠地区进行植被建设已成为一种重要的管理措施.在荒漠化严重的地区进行合理的人工植被建设是提高土地生产力和生物多样性、增加荒漠生态系统碳储量和减缓全球气候变化的有效方式[8-11].但是典型荒漠草地在退化过程中究竟损失了多少碳,以及退化严重的荒漠草地的固碳潜力究竟有多大,目前还缺少强有力的证据.在区域尺度上针对这一问题开展研究是减少荒漠生态系统碳储量和固碳潜力估算的不确定性以及应对全球气候变化的关键. ...
Long-term ecosystem effects of sand-binding vegetation in the Tengger Desert,northern China
1
2004
... 荒漠是陆地生态系统的重要组成部分,面积约占世界陆地面积的1/3[1-3].在气候变化和人类活动加剧的背景下,荒漠草地很容易退化或沙漠化[4-5],因此荒漠草地在全球碳循环中可能起着更加重要的作用.近年来,沙漠化程度不断加剧,导致荒漠草地生态系统不断退化,如植被、土壤结构的退化致使区域生态系统固碳能力下降,同时向大气中释放了大量的碳[6].全球沙漠化导致荒漠生态系统的碳损失为19—29 Pg,但这一数据还存在很大的不确定性[7].为了减缓沙漠化,促进生态系统稳定性,在荒漠地区进行植被建设已成为一种重要的管理措施.在荒漠化严重的地区进行合理的人工植被建设是提高土地生产力和生物多样性、增加荒漠生态系统碳储量和减缓全球气候变化的有效方式[8-11].但是典型荒漠草地在退化过程中究竟损失了多少碳,以及退化严重的荒漠草地的固碳潜力究竟有多大,目前还缺少强有力的证据.在区域尺度上针对这一问题开展研究是减少荒漠生态系统碳储量和固碳潜力估算的不确定性以及应对全球气候变化的关键. ...
Recovery of topsoil physicochemical properties in revegetated sites in the sand-burial ecosystems of the Tengger Desert,northern China
0
2007
Changes in soil and vegetation following stabilisation of dunes in the southeastern fringe of the Tengger Desert,China
0
2007
Carbon sequestration capacity of shifting sand dune after establishing new vegetation in the Tengger Desert,northern China
2
2014
... 荒漠是陆地生态系统的重要组成部分,面积约占世界陆地面积的1/3[1-3].在气候变化和人类活动加剧的背景下,荒漠草地很容易退化或沙漠化[4-5],因此荒漠草地在全球碳循环中可能起着更加重要的作用.近年来,沙漠化程度不断加剧,导致荒漠草地生态系统不断退化,如植被、土壤结构的退化致使区域生态系统固碳能力下降,同时向大气中释放了大量的碳[6].全球沙漠化导致荒漠生态系统的碳损失为19—29 Pg,但这一数据还存在很大的不确定性[7].为了减缓沙漠化,促进生态系统稳定性,在荒漠地区进行植被建设已成为一种重要的管理措施.在荒漠化严重的地区进行合理的人工植被建设是提高土地生产力和生物多样性、增加荒漠生态系统碳储量和减缓全球气候变化的有效方式[8-11].但是典型荒漠草地在退化过程中究竟损失了多少碳,以及退化严重的荒漠草地的固碳潜力究竟有多大,目前还缺少强有力的证据.在区域尺度上针对这一问题开展研究是减少荒漠生态系统碳储量和固碳潜力估算的不确定性以及应对全球气候变化的关键. ...
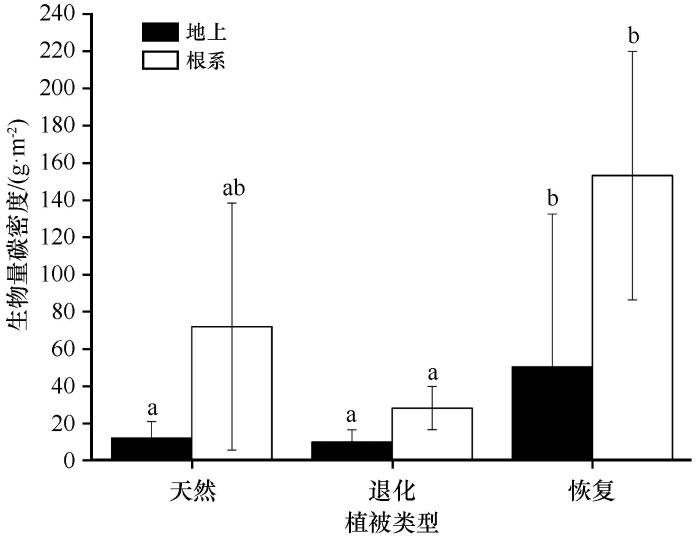
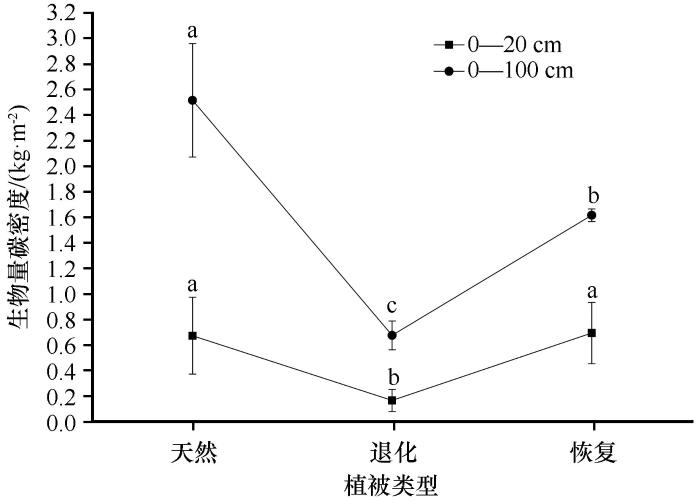
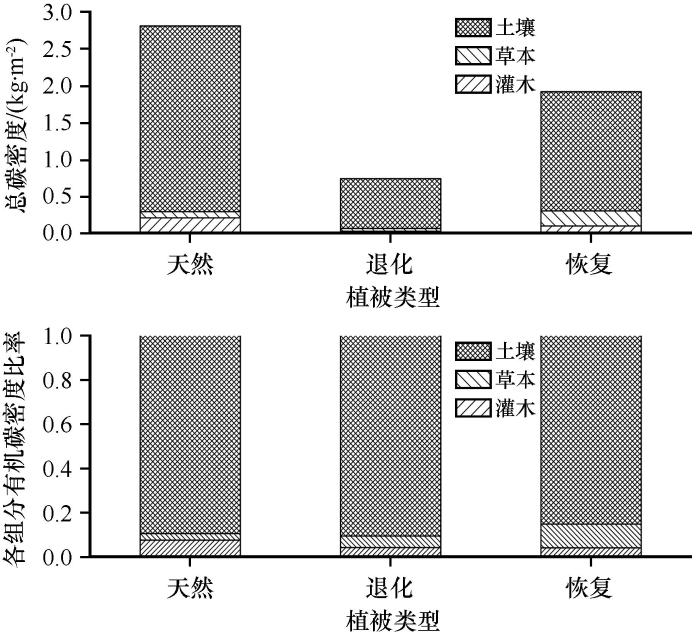
... 植被恢复还会影响土壤有机碳的分布和组成,土壤有机碳具有较强的表聚效应,本研究也证实了这一点,表层土壤(0—20 cm)理化性质受植被恢复的影响最大.主要原因为,在腾格里沙漠南缘沙漠化严重的地区进行固沙植被建设可以稳定土壤表面、增加土壤表面的粗糙度,进而增加地表对大气降尘的捕获并创造了一个稳定的物理环境[33];之后,大量的生物土壤结皮开始拓殖,草本也开始入侵,生物土壤结皮可以进行固碳和固氮活动,为表土层输入大量的有机碳[34],同时草本根系也主要分布在土壤表层,其根系周转也为表土层增加了大量的有机碳,此外灌木及草本的枯枝落叶也首先进入土壤表层.本研究显示,与退化生态系统相比,恢复生态系统各组分有机碳,包括土壤有机碳、灌木和草本生物量碳均大量增加,总有机碳密度增加了1.161 kg·m-2,但是与天然荒漠草地相比还有较大的固碳潜力(0.916 kg·m-2).由于0—20 cm土层土壤有机碳密度和草本生物量碳密度已经达到或超过天然荒漠草地,而植被地上部分容易被收获利用,吸存的碳很快又被释放出来,因此生物量碳存储的时间相对较短,尤其是草本和灌木叶片生物量碳.而土壤有机碳库和土壤养分可能会在很长时间内持续增加,即使植被的生物量已达到最大值后[11,35].考虑到旱生灌木的生物量分配特征(根冠比)[36],本研究认为,下层土壤(20—100 cm)是未来增加碳储量的主要土壤层.鉴于中国北方沙漠化土地面积巨大,干旱沙漠、半干旱沙漠以及沙漠化土地的面积分别为58.1万,10.3万km2和38.57万km2,且沙漠地区流动沙丘和半固定沙丘占比较大[37],因此中国荒漠和沙漠化地区具有巨大的固碳潜力,在未来应对全球气候变化中可能会起到更加重要的作用.我们认为在沙漠或沙漠化地区进行合理的人工植被恢复措施是增加荒漠生态系统碳储量、减缓大气中CO2浓度上升、实现中国碳达峰和碳中和以及应对全球气候变化的重要策略. ...
2
2016
... 早在20世纪50年代,在腾格里沙漠东南缘就进行了以旱生灌木为优势种的人工植被建设,有效阻止了沙漠化的进一步扩张,目前已经演替恢复为稳定的人工-自然生态系统[12].腾格里沙漠东南缘植被为典型的荒漠草原类型,是草原化荒漠与荒漠化草原的过渡带,目前分布有典型的荒漠草地生态系统、严重沙漠化的生态系统以及恢复良好的荒漠草地生态系统,是研究荒漠草地退化和重建过程中生态系统碳密度变化以及固碳潜力的理想区域.本研究选择腾格里沙漠东南缘典型的未退化的荒漠草地、退化严重的荒漠草地以及恢复良好的人工固沙植被为研究序列,研究荒漠草地退化和恢复过程中生态系统碳密度的变化特征及其固碳潜力.旨在区域尺度上准确评价荒漠草地在陆地生态系统碳循环中的作用及对人为干扰的响应,这也是未来土地利用和管理决策的重要基础. ...
... 研究区位于腾格里沙漠东南缘,属典型的温带大陆性干旱气候和季风气候[12].这一区域的年降雨量为180 mm左右,降雨量年际变率大,80.0%发生于6—9月,年均气温9.6 ℃,平均气温1月为-6.9 ℃,7月为24.3 ℃,≥10 ℃年有效积温大于3 000 ℃,年蒸发量约3 200 mm,年均风速4.2 m·s-1,年大风扬沙日数80—90 d,年沙暴日数16—25 d,多发生于4—6月,风向以西北风为主.土壤基质主要为疏松、贫瘠的流动风沙土. ...
Allometric models for estimating shrub biomass in desert grassland in northern China
1
2017
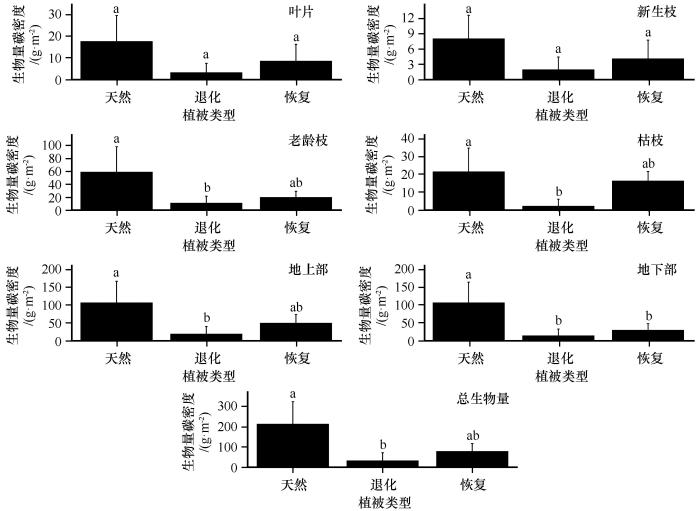
... 灌木样方面积为10 m×10 m,调查样方内所有灌木的总盖度,分物种调查所有灌木的株高及冠幅.根据调查结果,每个样地每个物种按形态选取3株具有代表性的个体,利用树剪对地上部分生物量进行取样,采用全挖开获得灌木根系生物量,并带回实验室.每一株均分解为叶片、新生枝、老龄枝、枯枝和根系,称量鲜重后置于65 ℃烘箱烘至恒重,最后获得干重.样地灌木各组分生物量估算参考Yang等[13]的方法. ...
1
2000
... 生物量碳和土壤有机碳(SOC)均采用重铬酸钾氧化-外加热法测定[14]. ...
Total carbon and nitrogen in the soils of the world
1
1996
... 土壤有机碳密度(Soil Organic Carbon Density,SOCD,kg·m-2)是指单位面积内一定深度土层中的土壤有机碳储量.土壤有机碳密度不仅是估算和评价土壤有机碳储量的重要指标,本身也是反映土壤特性的重要指标.它由土壤有机碳含量、砾石含量及容重共同确定.本文采用Batjes[15]的计算方式获得土壤有机碳密度. ...
Global soil organic carbon assessment
1
2015
... 土壤有机碳的维护和管理对生产力、土壤、水和能源安全至关重要,气候变化和土地利用变化正在对土壤产生巨大影响[16].气候变化等全球性问题需要在全球和生物圈尺度上考虑有机碳储量的变化,但人类活动的作用多发生在景观尺度上,对不同尺度有机碳的精确量化有助于制定更好的政策来保护和提高现有的有机碳,确保土壤的可持续利用[17].同时也是减缓全球气候变化,实现碳达峰和碳中和的关键.土地利用变化是全球碳循环的重要驱动力[18],多数研究表明,不合理的土地利用和生态系统退化是影响陆地生态系统土壤碳库的主要因素之一[19],如土地荒漠化直接导致封存的土壤碳释放,增高了大气中CO2的浓度,导致全球变暖[20-22].本研究显示腾格里沙漠南缘荒漠草地严重退化后生态系统有机碳密度,包括土壤有机碳密度、灌木各组分(叶片、新生枝、老龄枝、枯枝、根系)生物量碳以及草本(地上及根系)生物量碳密度均急剧降低,共减少2.068 kg·m-2,占未退化荒漠草地的73.494%,其中以土壤有机碳为主,这与科尔沁沙地沙漠化的部分研究结果相似[23].腾格里沙漠南缘荒漠草地生态系统退化从输入和输出两个方面影响生态系统碳储量,生态系统有机碳储量及其动态主要取决于有机质输入与输出之间的平衡,而在生态系统退化过程中,这一平衡遭到破坏,输入减少而输出增加.输入方面,随着生态系统的退化,灌木、草本以及地表也遭到破坏导致生产力逐渐降低,从大气中固定CO2的能力大大降低,生态系统有机碳输入的累积量减少,最终导致生态系统碳储量降低;输出方面,荒漠草地退化不仅从植物残体直接释放碳,而且导致土壤没有植被保护,在风蚀等作用下土壤结构遭到破坏,黏粉粒大量损失,一方面直接减少土壤有机碳的含量,另一方面又加速土壤有机碳的分解.总的来讲,荒漠草地的退化不仅直接影响植被生物量碳和土壤有机碳的空间分布和CO2释放,而且通过影响植被的结构和功能及土壤有机碳的形成、迁移、扩散及分解矿化等的影响因子(土壤母质和黏粒含量、土壤有机碳含量、土层深度、土壤含水量、温度)而间接影响植被生物量碳和土壤有机碳的含量和分布,最终通过改变土壤有机质的分解速率来影响有机碳密度. ...
Soil organic carbon across scales
1
2015
... 土壤有机碳的维护和管理对生产力、土壤、水和能源安全至关重要,气候变化和土地利用变化正在对土壤产生巨大影响[16].气候变化等全球性问题需要在全球和生物圈尺度上考虑有机碳储量的变化,但人类活动的作用多发生在景观尺度上,对不同尺度有机碳的精确量化有助于制定更好的政策来保护和提高现有的有机碳,确保土壤的可持续利用[17].同时也是减缓全球气候变化,实现碳达峰和碳中和的关键.土地利用变化是全球碳循环的重要驱动力[18],多数研究表明,不合理的土地利用和生态系统退化是影响陆地生态系统土壤碳库的主要因素之一[19],如土地荒漠化直接导致封存的土壤碳释放,增高了大气中CO2的浓度,导致全球变暖[20-22].本研究显示腾格里沙漠南缘荒漠草地严重退化后生态系统有机碳密度,包括土壤有机碳密度、灌木各组分(叶片、新生枝、老龄枝、枯枝、根系)生物量碳以及草本(地上及根系)生物量碳密度均急剧降低,共减少2.068 kg·m-2,占未退化荒漠草地的73.494%,其中以土壤有机碳为主,这与科尔沁沙地沙漠化的部分研究结果相似[23].腾格里沙漠南缘荒漠草地生态系统退化从输入和输出两个方面影响生态系统碳储量,生态系统有机碳储量及其动态主要取决于有机质输入与输出之间的平衡,而在生态系统退化过程中,这一平衡遭到破坏,输入减少而输出增加.输入方面,随着生态系统的退化,灌木、草本以及地表也遭到破坏导致生产力逐渐降低,从大气中固定CO2的能力大大降低,生态系统有机碳输入的累积量减少,最终导致生态系统碳储量降低;输出方面,荒漠草地退化不仅从植物残体直接释放碳,而且导致土壤没有植被保护,在风蚀等作用下土壤结构遭到破坏,黏粉粒大量损失,一方面直接减少土壤有机碳的含量,另一方面又加速土壤有机碳的分解.总的来讲,荒漠草地的退化不仅直接影响植被生物量碳和土壤有机碳的空间分布和CO2释放,而且通过影响植被的结构和功能及土壤有机碳的形成、迁移、扩散及分解矿化等的影响因子(土壤母质和黏粒含量、土壤有机碳含量、土层深度、土壤含水量、温度)而间接影响植被生物量碳和土壤有机碳的含量和分布,最终通过改变土壤有机质的分解速率来影响有机碳密度. ...
The impact of land use change on soil organic carbon and labile organic carbon stocks in the Longzhong region of Loess Plateau
1
2012
... 土壤有机碳的维护和管理对生产力、土壤、水和能源安全至关重要,气候变化和土地利用变化正在对土壤产生巨大影响[16].气候变化等全球性问题需要在全球和生物圈尺度上考虑有机碳储量的变化,但人类活动的作用多发生在景观尺度上,对不同尺度有机碳的精确量化有助于制定更好的政策来保护和提高现有的有机碳,确保土壤的可持续利用[17].同时也是减缓全球气候变化,实现碳达峰和碳中和的关键.土地利用变化是全球碳循环的重要驱动力[18],多数研究表明,不合理的土地利用和生态系统退化是影响陆地生态系统土壤碳库的主要因素之一[19],如土地荒漠化直接导致封存的土壤碳释放,增高了大气中CO2的浓度,导致全球变暖[20-22].本研究显示腾格里沙漠南缘荒漠草地严重退化后生态系统有机碳密度,包括土壤有机碳密度、灌木各组分(叶片、新生枝、老龄枝、枯枝、根系)生物量碳以及草本(地上及根系)生物量碳密度均急剧降低,共减少2.068 kg·m-2,占未退化荒漠草地的73.494%,其中以土壤有机碳为主,这与科尔沁沙地沙漠化的部分研究结果相似[23].腾格里沙漠南缘荒漠草地生态系统退化从输入和输出两个方面影响生态系统碳储量,生态系统有机碳储量及其动态主要取决于有机质输入与输出之间的平衡,而在生态系统退化过程中,这一平衡遭到破坏,输入减少而输出增加.输入方面,随着生态系统的退化,灌木、草本以及地表也遭到破坏导致生产力逐渐降低,从大气中固定CO2的能力大大降低,生态系统有机碳输入的累积量减少,最终导致生态系统碳储量降低;输出方面,荒漠草地退化不仅从植物残体直接释放碳,而且导致土壤没有植被保护,在风蚀等作用下土壤结构遭到破坏,黏粉粒大量损失,一方面直接减少土壤有机碳的含量,另一方面又加速土壤有机碳的分解.总的来讲,荒漠草地的退化不仅直接影响植被生物量碳和土壤有机碳的空间分布和CO2释放,而且通过影响植被的结构和功能及土壤有机碳的形成、迁移、扩散及分解矿化等的影响因子(土壤母质和黏粒含量、土壤有机碳含量、土层深度、土壤含水量、温度)而间接影响植被生物量碳和土壤有机碳的含量和分布,最终通过改变土壤有机质的分解速率来影响有机碳密度. ...
Land use induced changes of organic carbon storage in soils of China
1
2010
... 土壤有机碳的维护和管理对生产力、土壤、水和能源安全至关重要,气候变化和土地利用变化正在对土壤产生巨大影响[16].气候变化等全球性问题需要在全球和生物圈尺度上考虑有机碳储量的变化,但人类活动的作用多发生在景观尺度上,对不同尺度有机碳的精确量化有助于制定更好的政策来保护和提高现有的有机碳,确保土壤的可持续利用[17].同时也是减缓全球气候变化,实现碳达峰和碳中和的关键.土地利用变化是全球碳循环的重要驱动力[18],多数研究表明,不合理的土地利用和生态系统退化是影响陆地生态系统土壤碳库的主要因素之一[19],如土地荒漠化直接导致封存的土壤碳释放,增高了大气中CO2的浓度,导致全球变暖[20-22].本研究显示腾格里沙漠南缘荒漠草地严重退化后生态系统有机碳密度,包括土壤有机碳密度、灌木各组分(叶片、新生枝、老龄枝、枯枝、根系)生物量碳以及草本(地上及根系)生物量碳密度均急剧降低,共减少2.068 kg·m-2,占未退化荒漠草地的73.494%,其中以土壤有机碳为主,这与科尔沁沙地沙漠化的部分研究结果相似[23].腾格里沙漠南缘荒漠草地生态系统退化从输入和输出两个方面影响生态系统碳储量,生态系统有机碳储量及其动态主要取决于有机质输入与输出之间的平衡,而在生态系统退化过程中,这一平衡遭到破坏,输入减少而输出增加.输入方面,随着生态系统的退化,灌木、草本以及地表也遭到破坏导致生产力逐渐降低,从大气中固定CO2的能力大大降低,生态系统有机碳输入的累积量减少,最终导致生态系统碳储量降低;输出方面,荒漠草地退化不仅从植物残体直接释放碳,而且导致土壤没有植被保护,在风蚀等作用下土壤结构遭到破坏,黏粉粒大量损失,一方面直接减少土壤有机碳的含量,另一方面又加速土壤有机碳的分解.总的来讲,荒漠草地的退化不仅直接影响植被生物量碳和土壤有机碳的空间分布和CO2释放,而且通过影响植被的结构和功能及土壤有机碳的形成、迁移、扩散及分解矿化等的影响因子(土壤母质和黏粒含量、土壤有机碳含量、土层深度、土壤含水量、温度)而间接影响植被生物量碳和土壤有机碳的含量和分布,最终通过改变土壤有机质的分解速率来影响有机碳密度. ...
Pedospheric processes and the carbon cycle
1
1998
... 土壤有机碳的维护和管理对生产力、土壤、水和能源安全至关重要,气候变化和土地利用变化正在对土壤产生巨大影响[16].气候变化等全球性问题需要在全球和生物圈尺度上考虑有机碳储量的变化,但人类活动的作用多发生在景观尺度上,对不同尺度有机碳的精确量化有助于制定更好的政策来保护和提高现有的有机碳,确保土壤的可持续利用[17].同时也是减缓全球气候变化,实现碳达峰和碳中和的关键.土地利用变化是全球碳循环的重要驱动力[18],多数研究表明,不合理的土地利用和生态系统退化是影响陆地生态系统土壤碳库的主要因素之一[19],如土地荒漠化直接导致封存的土壤碳释放,增高了大气中CO2的浓度,导致全球变暖[20-22].本研究显示腾格里沙漠南缘荒漠草地严重退化后生态系统有机碳密度,包括土壤有机碳密度、灌木各组分(叶片、新生枝、老龄枝、枯枝、根系)生物量碳以及草本(地上及根系)生物量碳密度均急剧降低,共减少2.068 kg·m-2,占未退化荒漠草地的73.494%,其中以土壤有机碳为主,这与科尔沁沙地沙漠化的部分研究结果相似[23].腾格里沙漠南缘荒漠草地生态系统退化从输入和输出两个方面影响生态系统碳储量,生态系统有机碳储量及其动态主要取决于有机质输入与输出之间的平衡,而在生态系统退化过程中,这一平衡遭到破坏,输入减少而输出增加.输入方面,随着生态系统的退化,灌木、草本以及地表也遭到破坏导致生产力逐渐降低,从大气中固定CO2的能力大大降低,生态系统有机碳输入的累积量减少,最终导致生态系统碳储量降低;输出方面,荒漠草地退化不仅从植物残体直接释放碳,而且导致土壤没有植被保护,在风蚀等作用下土壤结构遭到破坏,黏粉粒大量损失,一方面直接减少土壤有机碳的含量,另一方面又加速土壤有机碳的分解.总的来讲,荒漠草地的退化不仅直接影响植被生物量碳和土壤有机碳的空间分布和CO2释放,而且通过影响植被的结构和功能及土壤有机碳的形成、迁移、扩散及分解矿化等的影响因子(土壤母质和黏粒含量、土壤有机碳含量、土层深度、土壤含水量、温度)而间接影响植被生物量碳和土壤有机碳的含量和分布,最终通过改变土壤有机质的分解速率来影响有机碳密度. ...
Carbon storage in desertified lands:a case study from North China
0
2000
Estimate of total CO2,output from desertified sandy land in China
1
2001
... 土壤有机碳的维护和管理对生产力、土壤、水和能源安全至关重要,气候变化和土地利用变化正在对土壤产生巨大影响[16].气候变化等全球性问题需要在全球和生物圈尺度上考虑有机碳储量的变化,但人类活动的作用多发生在景观尺度上,对不同尺度有机碳的精确量化有助于制定更好的政策来保护和提高现有的有机碳,确保土壤的可持续利用[17].同时也是减缓全球气候变化,实现碳达峰和碳中和的关键.土地利用变化是全球碳循环的重要驱动力[18],多数研究表明,不合理的土地利用和生态系统退化是影响陆地生态系统土壤碳库的主要因素之一[19],如土地荒漠化直接导致封存的土壤碳释放,增高了大气中CO2的浓度,导致全球变暖[20-22].本研究显示腾格里沙漠南缘荒漠草地严重退化后生态系统有机碳密度,包括土壤有机碳密度、灌木各组分(叶片、新生枝、老龄枝、枯枝、根系)生物量碳以及草本(地上及根系)生物量碳密度均急剧降低,共减少2.068 kg·m-2,占未退化荒漠草地的73.494%,其中以土壤有机碳为主,这与科尔沁沙地沙漠化的部分研究结果相似[23].腾格里沙漠南缘荒漠草地生态系统退化从输入和输出两个方面影响生态系统碳储量,生态系统有机碳储量及其动态主要取决于有机质输入与输出之间的平衡,而在生态系统退化过程中,这一平衡遭到破坏,输入减少而输出增加.输入方面,随着生态系统的退化,灌木、草本以及地表也遭到破坏导致生产力逐渐降低,从大气中固定CO2的能力大大降低,生态系统有机碳输入的累积量减少,最终导致生态系统碳储量降低;输出方面,荒漠草地退化不仅从植物残体直接释放碳,而且导致土壤没有植被保护,在风蚀等作用下土壤结构遭到破坏,黏粉粒大量损失,一方面直接减少土壤有机碳的含量,另一方面又加速土壤有机碳的分解.总的来讲,荒漠草地的退化不仅直接影响植被生物量碳和土壤有机碳的空间分布和CO2释放,而且通过影响植被的结构和功能及土壤有机碳的形成、迁移、扩散及分解矿化等的影响因子(土壤母质和黏粒含量、土壤有机碳含量、土层深度、土壤含水量、温度)而间接影响植被生物量碳和土壤有机碳的含量和分布,最终通过改变土壤有机质的分解速率来影响有机碳密度. ...
沙漠化过程中科尔沁沙地植物-土壤系统碳氮储量动态
1
2006
... 土壤有机碳的维护和管理对生产力、土壤、水和能源安全至关重要,气候变化和土地利用变化正在对土壤产生巨大影响[16].气候变化等全球性问题需要在全球和生物圈尺度上考虑有机碳储量的变化,但人类活动的作用多发生在景观尺度上,对不同尺度有机碳的精确量化有助于制定更好的政策来保护和提高现有的有机碳,确保土壤的可持续利用[17].同时也是减缓全球气候变化,实现碳达峰和碳中和的关键.土地利用变化是全球碳循环的重要驱动力[18],多数研究表明,不合理的土地利用和生态系统退化是影响陆地生态系统土壤碳库的主要因素之一[19],如土地荒漠化直接导致封存的土壤碳释放,增高了大气中CO2的浓度,导致全球变暖[20-22].本研究显示腾格里沙漠南缘荒漠草地严重退化后生态系统有机碳密度,包括土壤有机碳密度、灌木各组分(叶片、新生枝、老龄枝、枯枝、根系)生物量碳以及草本(地上及根系)生物量碳密度均急剧降低,共减少2.068 kg·m-2,占未退化荒漠草地的73.494%,其中以土壤有机碳为主,这与科尔沁沙地沙漠化的部分研究结果相似[23].腾格里沙漠南缘荒漠草地生态系统退化从输入和输出两个方面影响生态系统碳储量,生态系统有机碳储量及其动态主要取决于有机质输入与输出之间的平衡,而在生态系统退化过程中,这一平衡遭到破坏,输入减少而输出增加.输入方面,随着生态系统的退化,灌木、草本以及地表也遭到破坏导致生产力逐渐降低,从大气中固定CO2的能力大大降低,生态系统有机碳输入的累积量减少,最终导致生态系统碳储量降低;输出方面,荒漠草地退化不仅从植物残体直接释放碳,而且导致土壤没有植被保护,在风蚀等作用下土壤结构遭到破坏,黏粉粒大量损失,一方面直接减少土壤有机碳的含量,另一方面又加速土壤有机碳的分解.总的来讲,荒漠草地的退化不仅直接影响植被生物量碳和土壤有机碳的空间分布和CO2释放,而且通过影响植被的结构和功能及土壤有机碳的形成、迁移、扩散及分解矿化等的影响因子(土壤母质和黏粒含量、土壤有机碳含量、土层深度、土壤含水量、温度)而间接影响植被生物量碳和土壤有机碳的含量和分布,最终通过改变土壤有机质的分解速率来影响有机碳密度. ...
Soil management and restoration for C sequestration to mitigate the accelerated greenhouse effect
1
1999
... 有关植被恢复对生态系统碳储量影响的研究多集中在土壤有机碳方面,多数研究表明,在退化土地上进行植被恢复重建能够有效地促进土壤发育、改善土壤特性、增强土壤微生物活性、提高土壤肥力,显著增加了生态系统的固碳潜力,同时还能够有效控制土壤侵蚀减少土壤碳损失,是增加土壤碳汇的重要手段[24-26].一方面,植被恢复增加了地表覆盖,可以有效减少土壤侵蚀和养分的流失,另一方面,植被恢复增加了土壤有机碳的输入,如植物残体、根系以及根系分泌物等[27-29].此外,土壤有机碳的变化及其稳定性取决于各粒径组分[30],植被恢复显著增加了土壤黏粉粒含量,提升了土壤吸收和存储有机碳的能力,是驱动土壤碳固存和土壤恢复的关键因素[31-32]. ...
植被恢复方式对黄土丘陵区土壤理化性质及微生物特性的影响
0
2014
腾格里沙漠东南缘植被恢复过程中土壤微生物量及酶活性
1
2019
... 有关植被恢复对生态系统碳储量影响的研究多集中在土壤有机碳方面,多数研究表明,在退化土地上进行植被恢复重建能够有效地促进土壤发育、改善土壤特性、增强土壤微生物活性、提高土壤肥力,显著增加了生态系统的固碳潜力,同时还能够有效控制土壤侵蚀减少土壤碳损失,是增加土壤碳汇的重要手段[24-26].一方面,植被恢复增加了地表覆盖,可以有效减少土壤侵蚀和养分的流失,另一方面,植被恢复增加了土壤有机碳的输入,如植物残体、根系以及根系分泌物等[27-29].此外,土壤有机碳的变化及其稳定性取决于各粒径组分[30],植被恢复显著增加了土壤黏粉粒含量,提升了土壤吸收和存储有机碳的能力,是驱动土壤碳固存和土壤恢复的关键因素[31-32]. ...
Soil carbon sequestration and land-use change: processes and potential
1
2000
... 有关植被恢复对生态系统碳储量影响的研究多集中在土壤有机碳方面,多数研究表明,在退化土地上进行植被恢复重建能够有效地促进土壤发育、改善土壤特性、增强土壤微生物活性、提高土壤肥力,显著增加了生态系统的固碳潜力,同时还能够有效控制土壤侵蚀减少土壤碳损失,是增加土壤碳汇的重要手段[24-26].一方面,植被恢复增加了地表覆盖,可以有效减少土壤侵蚀和养分的流失,另一方面,植被恢复增加了土壤有机碳的输入,如植物残体、根系以及根系分泌物等[27-29].此外,土壤有机碳的变化及其稳定性取决于各粒径组分[30],植被恢复显著增加了土壤黏粉粒含量,提升了土壤吸收和存储有机碳的能力,是驱动土壤碳固存和土壤恢复的关键因素[31-32]. ...
Effect of plant cover decline on chemical and microbiological parameters under Mediterranean climate
0
2002
Different approaches to evaluating soil quality using biochemical properties
1
2005
... 有关植被恢复对生态系统碳储量影响的研究多集中在土壤有机碳方面,多数研究表明,在退化土地上进行植被恢复重建能够有效地促进土壤发育、改善土壤特性、增强土壤微生物活性、提高土壤肥力,显著增加了生态系统的固碳潜力,同时还能够有效控制土壤侵蚀减少土壤碳损失,是增加土壤碳汇的重要手段[24-26].一方面,植被恢复增加了地表覆盖,可以有效减少土壤侵蚀和养分的流失,另一方面,植被恢复增加了土壤有机碳的输入,如植物残体、根系以及根系分泌物等[27-29].此外,土壤有机碳的变化及其稳定性取决于各粒径组分[30],植被恢复显著增加了土壤黏粉粒含量,提升了土壤吸收和存储有机碳的能力,是驱动土壤碳固存和土壤恢复的关键因素[31-32]. ...
Soil organic carbon in particle-size fractions under three grassland types in Inner Mongolia,China
1
2018
... 有关植被恢复对生态系统碳储量影响的研究多集中在土壤有机碳方面,多数研究表明,在退化土地上进行植被恢复重建能够有效地促进土壤发育、改善土壤特性、增强土壤微生物活性、提高土壤肥力,显著增加了生态系统的固碳潜力,同时还能够有效控制土壤侵蚀减少土壤碳损失,是增加土壤碳汇的重要手段[24-26].一方面,植被恢复增加了地表覆盖,可以有效减少土壤侵蚀和养分的流失,另一方面,植被恢复增加了土壤有机碳的输入,如植物残体、根系以及根系分泌物等[27-29].此外,土壤有机碳的变化及其稳定性取决于各粒径组分[30],植被恢复显著增加了土壤黏粉粒含量,提升了土壤吸收和存储有机碳的能力,是驱动土壤碳固存和土壤恢复的关键因素[31-32]. ...
Restoration affect soil organic carbon and nutrients in different particle-size fractions
1
2016
... 有关植被恢复对生态系统碳储量影响的研究多集中在土壤有机碳方面,多数研究表明,在退化土地上进行植被恢复重建能够有效地促进土壤发育、改善土壤特性、增强土壤微生物活性、提高土壤肥力,显著增加了生态系统的固碳潜力,同时还能够有效控制土壤侵蚀减少土壤碳损失,是增加土壤碳汇的重要手段[24-26].一方面,植被恢复增加了地表覆盖,可以有效减少土壤侵蚀和养分的流失,另一方面,植被恢复增加了土壤有机碳的输入,如植物残体、根系以及根系分泌物等[27-29].此外,土壤有机碳的变化及其稳定性取决于各粒径组分[30],植被恢复显著增加了土壤黏粉粒含量,提升了土壤吸收和存储有机碳的能力,是驱动土壤碳固存和土壤恢复的关键因素[31-32]. ...
干旱区流动沙地建植梭梭(Haloxylon ammodendron)林后细粒物质输入对土壤碳氮积累的影响
1
2021
... 有关植被恢复对生态系统碳储量影响的研究多集中在土壤有机碳方面,多数研究表明,在退化土地上进行植被恢复重建能够有效地促进土壤发育、改善土壤特性、增强土壤微生物活性、提高土壤肥力,显著增加了生态系统的固碳潜力,同时还能够有效控制土壤侵蚀减少土壤碳损失,是增加土壤碳汇的重要手段[24-26].一方面,植被恢复增加了地表覆盖,可以有效减少土壤侵蚀和养分的流失,另一方面,植被恢复增加了土壤有机碳的输入,如植物残体、根系以及根系分泌物等[27-29].此外,土壤有机碳的变化及其稳定性取决于各粒径组分[30],植被恢复显著增加了土壤黏粉粒含量,提升了土壤吸收和存储有机碳的能力,是驱动土壤碳固存和土壤恢复的关键因素[31-32]. ...
Long-term effects of revegetation on soil water content of sand dunes in arid region of Northern China
1
2004
... 植被恢复还会影响土壤有机碳的分布和组成,土壤有机碳具有较强的表聚效应,本研究也证实了这一点,表层土壤(0—20 cm)理化性质受植被恢复的影响最大.主要原因为,在腾格里沙漠南缘沙漠化严重的地区进行固沙植被建设可以稳定土壤表面、增加土壤表面的粗糙度,进而增加地表对大气降尘的捕获并创造了一个稳定的物理环境[33];之后,大量的生物土壤结皮开始拓殖,草本也开始入侵,生物土壤结皮可以进行固碳和固氮活动,为表土层输入大量的有机碳[34],同时草本根系也主要分布在土壤表层,其根系周转也为表土层增加了大量的有机碳,此外灌木及草本的枯枝落叶也首先进入土壤表层.本研究显示,与退化生态系统相比,恢复生态系统各组分有机碳,包括土壤有机碳、灌木和草本生物量碳均大量增加,总有机碳密度增加了1.161 kg·m-2,但是与天然荒漠草地相比还有较大的固碳潜力(0.916 kg·m-2).由于0—20 cm土层土壤有机碳密度和草本生物量碳密度已经达到或超过天然荒漠草地,而植被地上部分容易被收获利用,吸存的碳很快又被释放出来,因此生物量碳存储的时间相对较短,尤其是草本和灌木叶片生物量碳.而土壤有机碳库和土壤养分可能会在很长时间内持续增加,即使植被的生物量已达到最大值后[11,35].考虑到旱生灌木的生物量分配特征(根冠比)[36],本研究认为,下层土壤(20—100 cm)是未来增加碳储量的主要土壤层.鉴于中国北方沙漠化土地面积巨大,干旱沙漠、半干旱沙漠以及沙漠化土地的面积分别为58.1万,10.3万km2和38.57万km2,且沙漠地区流动沙丘和半固定沙丘占比较大[37],因此中国荒漠和沙漠化地区具有巨大的固碳潜力,在未来应对全球气候变化中可能会起到更加重要的作用.我们认为在沙漠或沙漠化地区进行合理的人工植被恢复措施是增加荒漠生态系统碳储量、减缓大气中CO2浓度上升、实现中国碳达峰和碳中和以及应对全球气候变化的重要策略. ...
1
2012
... 植被恢复还会影响土壤有机碳的分布和组成,土壤有机碳具有较强的表聚效应,本研究也证实了这一点,表层土壤(0—20 cm)理化性质受植被恢复的影响最大.主要原因为,在腾格里沙漠南缘沙漠化严重的地区进行固沙植被建设可以稳定土壤表面、增加土壤表面的粗糙度,进而增加地表对大气降尘的捕获并创造了一个稳定的物理环境[33];之后,大量的生物土壤结皮开始拓殖,草本也开始入侵,生物土壤结皮可以进行固碳和固氮活动,为表土层输入大量的有机碳[34],同时草本根系也主要分布在土壤表层,其根系周转也为表土层增加了大量的有机碳,此外灌木及草本的枯枝落叶也首先进入土壤表层.本研究显示,与退化生态系统相比,恢复生态系统各组分有机碳,包括土壤有机碳、灌木和草本生物量碳均大量增加,总有机碳密度增加了1.161 kg·m-2,但是与天然荒漠草地相比还有较大的固碳潜力(0.916 kg·m-2).由于0—20 cm土层土壤有机碳密度和草本生物量碳密度已经达到或超过天然荒漠草地,而植被地上部分容易被收获利用,吸存的碳很快又被释放出来,因此生物量碳存储的时间相对较短,尤其是草本和灌木叶片生物量碳.而土壤有机碳库和土壤养分可能会在很长时间内持续增加,即使植被的生物量已达到最大值后[11,35].考虑到旱生灌木的生物量分配特征(根冠比)[36],本研究认为,下层土壤(20—100 cm)是未来增加碳储量的主要土壤层.鉴于中国北方沙漠化土地面积巨大,干旱沙漠、半干旱沙漠以及沙漠化土地的面积分别为58.1万,10.3万km2和38.57万km2,且沙漠地区流动沙丘和半固定沙丘占比较大[37],因此中国荒漠和沙漠化地区具有巨大的固碳潜力,在未来应对全球气候变化中可能会起到更加重要的作用.我们认为在沙漠或沙漠化地区进行合理的人工植被恢复措施是增加荒漠生态系统碳储量、减缓大气中CO2浓度上升、实现中国碳达峰和碳中和以及应对全球气候变化的重要策略. ...
Vegetation restoration drives the dynamics and distribution of nitrogen and phosphorous pools in a temperate desert soil-plant system
1
2019
... 植被恢复还会影响土壤有机碳的分布和组成,土壤有机碳具有较强的表聚效应,本研究也证实了这一点,表层土壤(0—20 cm)理化性质受植被恢复的影响最大.主要原因为,在腾格里沙漠南缘沙漠化严重的地区进行固沙植被建设可以稳定土壤表面、增加土壤表面的粗糙度,进而增加地表对大气降尘的捕获并创造了一个稳定的物理环境[33];之后,大量的生物土壤结皮开始拓殖,草本也开始入侵,生物土壤结皮可以进行固碳和固氮活动,为表土层输入大量的有机碳[34],同时草本根系也主要分布在土壤表层,其根系周转也为表土层增加了大量的有机碳,此外灌木及草本的枯枝落叶也首先进入土壤表层.本研究显示,与退化生态系统相比,恢复生态系统各组分有机碳,包括土壤有机碳、灌木和草本生物量碳均大量增加,总有机碳密度增加了1.161 kg·m-2,但是与天然荒漠草地相比还有较大的固碳潜力(0.916 kg·m-2).由于0—20 cm土层土壤有机碳密度和草本生物量碳密度已经达到或超过天然荒漠草地,而植被地上部分容易被收获利用,吸存的碳很快又被释放出来,因此生物量碳存储的时间相对较短,尤其是草本和灌木叶片生物量碳.而土壤有机碳库和土壤养分可能会在很长时间内持续增加,即使植被的生物量已达到最大值后[11,35].考虑到旱生灌木的生物量分配特征(根冠比)[36],本研究认为,下层土壤(20—100 cm)是未来增加碳储量的主要土壤层.鉴于中国北方沙漠化土地面积巨大,干旱沙漠、半干旱沙漠以及沙漠化土地的面积分别为58.1万,10.3万km2和38.57万km2,且沙漠地区流动沙丘和半固定沙丘占比较大[37],因此中国荒漠和沙漠化地区具有巨大的固碳潜力,在未来应对全球气候变化中可能会起到更加重要的作用.我们认为在沙漠或沙漠化地区进行合理的人工植被恢复措施是增加荒漠生态系统碳储量、减缓大气中CO2浓度上升、实现中国碳达峰和碳中和以及应对全球气候变化的重要策略. ...
荒漠草地4种灌木生物量分配特征
1
2013
... 植被恢复还会影响土壤有机碳的分布和组成,土壤有机碳具有较强的表聚效应,本研究也证实了这一点,表层土壤(0—20 cm)理化性质受植被恢复的影响最大.主要原因为,在腾格里沙漠南缘沙漠化严重的地区进行固沙植被建设可以稳定土壤表面、增加土壤表面的粗糙度,进而增加地表对大气降尘的捕获并创造了一个稳定的物理环境[33];之后,大量的生物土壤结皮开始拓殖,草本也开始入侵,生物土壤结皮可以进行固碳和固氮活动,为表土层输入大量的有机碳[34],同时草本根系也主要分布在土壤表层,其根系周转也为表土层增加了大量的有机碳,此外灌木及草本的枯枝落叶也首先进入土壤表层.本研究显示,与退化生态系统相比,恢复生态系统各组分有机碳,包括土壤有机碳、灌木和草本生物量碳均大量增加,总有机碳密度增加了1.161 kg·m-2,但是与天然荒漠草地相比还有较大的固碳潜力(0.916 kg·m-2).由于0—20 cm土层土壤有机碳密度和草本生物量碳密度已经达到或超过天然荒漠草地,而植被地上部分容易被收获利用,吸存的碳很快又被释放出来,因此生物量碳存储的时间相对较短,尤其是草本和灌木叶片生物量碳.而土壤有机碳库和土壤养分可能会在很长时间内持续增加,即使植被的生物量已达到最大值后[11,35].考虑到旱生灌木的生物量分配特征(根冠比)[36],本研究认为,下层土壤(20—100 cm)是未来增加碳储量的主要土壤层.鉴于中国北方沙漠化土地面积巨大,干旱沙漠、半干旱沙漠以及沙漠化土地的面积分别为58.1万,10.3万km2和38.57万km2,且沙漠地区流动沙丘和半固定沙丘占比较大[37],因此中国荒漠和沙漠化地区具有巨大的固碳潜力,在未来应对全球气候变化中可能会起到更加重要的作用.我们认为在沙漠或沙漠化地区进行合理的人工植被恢复措施是增加荒漠生态系统碳储量、减缓大气中CO2浓度上升、实现中国碳达峰和碳中和以及应对全球气候变化的重要策略. ...
腾格里沙漠土壤类型空间分布特征
1
2020
... 植被恢复还会影响土壤有机碳的分布和组成,土壤有机碳具有较强的表聚效应,本研究也证实了这一点,表层土壤(0—20 cm)理化性质受植被恢复的影响最大.主要原因为,在腾格里沙漠南缘沙漠化严重的地区进行固沙植被建设可以稳定土壤表面、增加土壤表面的粗糙度,进而增加地表对大气降尘的捕获并创造了一个稳定的物理环境[33];之后,大量的生物土壤结皮开始拓殖,草本也开始入侵,生物土壤结皮可以进行固碳和固氮活动,为表土层输入大量的有机碳[34],同时草本根系也主要分布在土壤表层,其根系周转也为表土层增加了大量的有机碳,此外灌木及草本的枯枝落叶也首先进入土壤表层.本研究显示,与退化生态系统相比,恢复生态系统各组分有机碳,包括土壤有机碳、灌木和草本生物量碳均大量增加,总有机碳密度增加了1.161 kg·m-2,但是与天然荒漠草地相比还有较大的固碳潜力(0.916 kg·m-2).由于0—20 cm土层土壤有机碳密度和草本生物量碳密度已经达到或超过天然荒漠草地,而植被地上部分容易被收获利用,吸存的碳很快又被释放出来,因此生物量碳存储的时间相对较短,尤其是草本和灌木叶片生物量碳.而土壤有机碳库和土壤养分可能会在很长时间内持续增加,即使植被的生物量已达到最大值后[11,35].考虑到旱生灌木的生物量分配特征(根冠比)[36],本研究认为,下层土壤(20—100 cm)是未来增加碳储量的主要土壤层.鉴于中国北方沙漠化土地面积巨大,干旱沙漠、半干旱沙漠以及沙漠化土地的面积分别为58.1万,10.3万km2和38.57万km2,且沙漠地区流动沙丘和半固定沙丘占比较大[37],因此中国荒漠和沙漠化地区具有巨大的固碳潜力,在未来应对全球气候变化中可能会起到更加重要的作用.我们认为在沙漠或沙漠化地区进行合理的人工植被恢复措施是增加荒漠生态系统碳储量、减缓大气中CO2浓度上升、实现中国碳达峰和碳中和以及应对全球气候变化的重要策略. ...




 甘公网安备 62010202000688号
甘公网安备 62010202000688号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}